อ่าน 35 นาที
ไดโนแฟลเจลเลต
ไดโนแฟลเจลเลต (จากภาษากรีกโบราณδῖνος ( dînos ) ' หมุนวน'และภาษาละตินflagellum ' แส้, แส้' ) หรือที่เรียกว่าไดโนไฟต์เป็น กลุ่ม โมโนฟิเลติก ของ...
ไดโนแฟลเจลเลต
| ไดโนแฟลเจลเลต ช่วงเวลา: ยุคไทรแอสสิกหรือก่อนหน้านั้น – ปัจจุบัน | |
|---|---|
 | |
| เซราเทียมสปีชีส์ | |
| การจำแนกทางวิทยาศาสตร์ | |
| โดเมน: | ยูคาริโอตา |
| กลุ่มสายพันธุ์ : | ซาร์ |
| กลุ่มสายพันธุ์ : | อัลวีโอลาตา |
| กลุ่มสายพันธุ์ : | ไดโนซัว |
| แผนก: | ไดโนแฟลเจลลาตาบุตชลี 1885 [ 2 ] |
| ชั้นเรียน | |
| คำพ้องความหมาย | |
| |
ไดโนแฟลเจลเลต (จากภาษากรีกโบราณδῖνος ( dînos ) ' หมุนวน'และภาษาละตินflagellum ' แส้, แส้' ) หรือที่เรียกว่าไดโนไฟต์เป็น กลุ่ม โมโนฟิเลติก ของ ยูคาริโอตเซลล์เดียวที่ประกอบเป็นไฟลัมไดโนแฟลเจลเล ตา [ 3 ]และโดยทั่วไปถือว่าเป็นโปรติสต์ ไดโนแฟลเจลเลตส่วนใหญ่เป็นแพลงก์ตอนในทะเลแต่ก็พบได้ทั่วไปในแหล่งน้ำจืดประชากรของพวกมันแตกต่างกันไปตามอุณหภูมิผิวน้ำทะเลความเค็มและความลึก ไดโนแฟลเจลเลตหลายชนิดสังเคราะห์แสงได้แต่ส่วนใหญ่เป็นแบบมิกโซ โทรฟิก คือผสมผสานการสังเคราะห์ แสงกับการกินเหยื่อ ( ฟาโกโทรฟีและไมโซไซโตซิส ) [ 4 ] [ 5 ]
ในแง่ของจำนวนชนิด ไดโนแฟลเจลเลตเป็นหนึ่งในกลุ่มยูคาริโอตในทะเลที่ใหญ่ที่สุด แม้ว่าจะมีขนาดเล็กกว่าไดอะตอม มาก ก็ตาม[ 6 ]บางชนิดเป็นเอนโดซิมไบออนต์ของสัตว์ทะเลและมีบทบาทสำคัญในชีววิทยาของแนวปะการัง ไดโนแฟลเจลเลตบางชนิดเป็นผู้ล่าที่ไม่มีเม็ดสีบนโปรโตซัว ชนิดอื่น และบางชนิดเป็นปรสิต (เช่นOodiniumและPfiesteria ) ไดโนแฟลเจลเลตบางชนิดสร้างระยะพักตัวที่เรียกว่าซีสต์ไดโนแฟลเจลเลตหรือไดโนซิสต์ซึ่งเป็นส่วนหนึ่งของวงจรชีวิต โดยพบใน 84 จาก 350 ชนิดในน้ำจืดที่ได้รับการอธิบาย และมากกว่า 10% เล็กน้อยของชนิดในทะเลที่รู้จัก[ 7 ] [ 8 ]ไดโนแฟลเจลเลตเป็นอัลวีโอเลต ที่มีแฟล เจลลาสอง เส้น ซึ่งเป็นลักษณะดั้งเดิมของไบคอนต์
ปัจจุบันมีการระบุชนิดของไดโนแฟลเจลเลตในทะเลที่ดำรงชีวิตอิสระไว้ประมาณ 1,555 ชนิด[ 9 ]การประมาณการอีกครั้งหนึ่งชี้ให้เห็นว่ามีสิ่งมีชีวิตประมาณ 2,000 ชนิด ซึ่งมากกว่า 1,700 ชนิดอยู่ในทะเล (ทั้งที่ดำรงชีวิตอิสระและที่อาศัยอยู่บนพื้นทะเล) และประมาณ 220 ชนิดอยู่ในน้ำจืด[ 10 ]การประมาณการล่าสุดชี้ให้เห็นว่ามีสิ่งมีชีวิตไดโนแฟลเจลเลตทั้งหมด 2,294 ชนิด ซึ่งรวมถึงไดโนแฟลเจลเลตในทะเล น้ำจืด และปรสิต[ 11 ]
การสะสมตัวอย่างรวดเร็วของไดโนแฟลเจลเลตบางชนิดอาจทำให้เกิดการเปลี่ยนสีของน้ำอย่างเห็นได้ชัด ซึ่งเรียกกันทั่วไปว่าปรากฏการณ์น้ำแดง ( การแพร่กระจายของสาหร่ายที่เป็นอันตราย ) และอาจก่อให้เกิดพิษจากหอยหากมนุษย์รับประทานหอยที่ปนเปื้อนเข้าไป ไดโนแฟลเจลเลตบางชนิดยังแสดงคุณสมบัติเรืองแสงโดยส่วนใหญ่จะปล่อยแสงสีฟ้าอมเขียว ซึ่งอาจมองเห็นได้ในบริเวณมหาสมุทรภายใต้เงื่อนไขบางประการ
นิรุกติศาสตร์
คำว่า "ไดโนแฟลเจลเลต" มาจากการรวมกันของคำว่าdinos ในภาษากรีก และflagellum ในภาษา ละตินDinosหมายถึง "การหมุนวน" และหมายถึงลักษณะเฉพาะของการว่ายน้ำของไดโนแฟลเจลเลตที่ถูกสังเกตFlagellumหมายถึง "แส้" และหมายถึงแฟลเจลลาของ พวกมัน [ 12 ]
ประวัติศาสตร์
ในปี ค.ศ. 1753 ไดโนแฟลเจลเลตสมัยใหม่ชนิดแรก ( Noctilucaแม้ว่าจะยังไม่ได้รับการตั้งชื่ออย่างเป็นทางการจนกระทั่งปี ค.ศ. 1816) ได้รับการอธิบายโดยเฮนรี เบเกอร์ว่าเป็น "สิ่งมีชีวิตขนาดเล็กที่ทำให้เกิดแสงระยิบระยับในน้ำทะเล" [ 13 ]ออตโต ฟรีดริช มุลเลอร์ได้บันทึกBursaria (ปัจจุบันคือCeratium ) hirundinellaและVorticella (ปัจจุบันคือ Peridinium ) cinctumในปี ค.ศ. 1773 [ 14 ]
ชื่อสามัญที่เก่าแก่ที่สุดของไดโนแฟลเจลเลตคือCeratiumซึ่งเสนอโดย Schrank (1793)
ในช่วงทศวรรษ 1830 นักจุลทรรศน์ชาวเยอรมันChristian Gottfried Ehrenberg ได้ตรวจสอบตัวอย่างน้ำและแพลงก์ตอนจำนวนมาก และ เสนอสกุลไดโนแฟลเจลเลตหลายสกุลที่ยังคงใช้กันอยู่ในปัจจุบัน ได้แก่Peridinium, ProrocentrumและDinophysis [ 15 ]
ไดโนแฟลเจลเลตเหล่านี้ได้รับการกำหนดครั้งแรกโดยOtto Bütschliในปี 1885 ว่าเป็นอันดับไดโน แฟลเจลเลต (Dinoflagellida) [ 16 ]นักพฤกษศาสตร์ถือว่าพวกมันเป็นส่วนหนึ่งของสาหร่าย โดยตั้งชื่อว่าPyrrophytaหรือPyrrhophyta ("สาหร่ายไฟ"; ภาษากรีกpyrr(h)osแปลว่าไฟ) ตามรูปแบบที่เรืองแสงได้ หรือDinophytaในช่วงเวลาต่างๆคริปโตโมนาดอีบริอิดและเอลโลไบโอป ซิด ถูกรวมอยู่ในกลุ่มนี้ แต่ปัจจุบันมีเพียงกลุ่มหลังเท่านั้นที่ถือว่าเป็นญาติใกล้ชิด ไดโนแฟลเจลเลตมีความสามารถในการเปลี่ยนจากกลยุทธ์ที่ไม่สร้างซีสต์ไปเป็นกลยุทธ์ที่สร้างซีสต์ ซึ่งทำให้การสร้างประวัติวิวัฒนาการของพวกมันขึ้นมาใหม่เป็นเรื่องยากมาก
สัณฐานวิทยา

- เยื่อหุ้มพลา สติด (3, สีแดงทุติยภูมิ)
- ไทลาคอยด์คือบริเวณที่เกิดปฏิกิริยาที่ต้องอาศัยแสงในกระบวนการสังเคราะห์แสง
- ไพรีนอยด์ศูนย์กลางการตรึงคาร์บอน
- ไตรโคซิสต์
- ถุงลม, โพรงหรือหลุมที่ผิว
- แผ่นเทคาล
- ถุงหนอง
- สุญญากาศ
- เครื่องมือก็อลจิ (Golgi apparatus ) ทำหน้าที่ดัดแปลงโปรตีนและส่งโปรตีนเหล่านั้นออกนอกเซลล์
- เอนโดพลาสมิกเรติคูลัมคือเครือข่ายการขนส่งโมเลกุลไปยังส่วนต่างๆ ของเซลล์
- แฟลเจลลัมตามขวาง
- เส้นใยลายริ้ว
- การเก็บหนอง
- ไมโตคอนเดรียสร้างATP (พลังงาน) ให้กับเซลล์
- นิวเคลียส
- นิวคลีโอลัส
- โครโมโซมที่ควบแน่น
- เม็ดแป้ง
- ไลโซโซมเป็นที่เก็บเอนไซม์
- ฟาโกโซมคือถุงที่ก่อตัวขึ้นรอบอนุภาค
- มาสติโกนีมคือ "ขน" ที่ยึดติดกับแฟลเจลลัม
- แฟลเจลลัมตามยาว
ไดโนแฟลเจลเลตเป็นสิ่งมีชีวิตเซลล์เดียวและมีแฟลเจลลาสองแบบที่ไม่เหมือนกันซึ่งเกิดขึ้นจากด้านท้องของเซลล์ (การสร้างแฟลเจลลาแบบไดโนคอนต์) พวกมันมีแฟลเจลลัมตามขวางที่มีลักษณะคล้ายริบบิ้นที่มีหลายคลื่นซึ่งโบกไปทางซ้ายของเซลล์ และแฟลเจลลัมตามยาวแบบทั่วไปซึ่งโบกไปทางด้านหลัง[ 17 ] [ 18 ] [ 19 ]แฟลเจลลัมตามขวางเป็นริบบิ้นหยักซึ่งมีเพียงขอบด้านนอกเท่านั้นที่กระเพื่อมจากฐานถึงปลายเนื่องจากการทำงานของแอ็กโซนีมที่วิ่งไปตามนั้น ขอบของแอ็กโซนีมมีขนแบบง่ายๆ ซึ่งอาจมีความยาวแตกต่างกัน การเคลื่อนที่ของแฟลเจลลัมทำให้เกิดแรงผลักดันไปข้างหน้าและแรงหมุน แฟลเจลลัมตามยาวมีลักษณะค่อนข้างทั่วไป มีขนน้อยหรือไม่มีเลย มันโบกด้วยคลื่นเพียงหนึ่งหรือสองรอบเท่านั้น แฟลเจลลาจะอยู่ภายในร่องบนพื้นผิวเซลล์ โดยแฟลเจลลาตามขวางจะอยู่ภายในซิงกูลัม และแฟลเจลลาตามยาวจะอยู่ภายในซัลคัส แม้ว่าส่วนปลายสุดของมันจะยื่นออกมาอย่างอิสระด้านหลังเซลล์ก็ตาม ในไดโนแฟลเจลเลตชนิดที่มีแฟลเจลลาแบบเดสโมคอนต์ (เช่นProrocentrum ) แฟลเจลลาทั้งสองจะแยกออกจากกันเช่นเดียวกับในไดโนคอนต์ แต่ไม่ได้เกี่ยวข้องกับร่องบนพื้นผิวเซลล์
ไดโนแฟลเจลเลตมีเปลือกเซลล์ที่ซับซ้อนเรียกว่าแอมฟีสมาหรือคอร์เทกซ์ ซึ่งประกอบด้วยเยื่อหุ้มหลายชั้นถุง แบนๆ ที่เรียกว่าอัลวีโอลิ (= ถุงแอมฟีสมา) และโครงสร้างที่เกี่ยวข้อง[ 20 ] [ 21 ]ในไดโนแฟลเจลเลตแบบมีเกราะ (thecate) เปลือกเหล่านี้จะรองรับ แผ่น เซลลูโลส ที่ซ้อนทับกัน เพื่อสร้างเกราะชนิดหนึ่งที่เรียกว่าเทกาหรือโลริกาซึ่งแตกต่างจากไดโนแฟลเจลเลตแบบไม่มีเกราะ (athecate) เปลือกเหล่านี้มีรูปร่างและการจัดเรียงที่หลากหลาย ขึ้นอยู่กับชนิดและบางครั้งก็ขึ้นอยู่กับระยะของไดโนแฟลเจลเลต โดยทั่วไป คำว่า tabulation ถูกใช้เพื่ออ้างถึงการจัดเรียงแผ่นเทกา แบบนี้ การกำหนดค่าแผ่นสามารถแสดงได้ด้วยสูตรแผ่นหรือสูตร tabulation นอกจากนี้ยังพบเอ็กซ์ทรูโซม ที่เป็นเส้นใยในหลายรูปแบบ [ 22 ] [ 23 ]
ร่องขวางที่เรียกว่า cingulum (หรือ cigulum) วิ่งรอบเซลล์ ทำให้แบ่งเซลล์ออกเป็นส่วนหน้า (episoma) และส่วนหลัง (hyposoma) หากมี theca อยู่ ส่วนต่างๆ จะเรียกว่า epitheca และ hypotheca ตามลำดับ ทางด้านหลัง เริ่มจากร่องขวาง จะมีร่องตามยาวที่เรียกว่า sulcus แฟลเจลลัมตามขวางจะกระทบกับ cingulum แฟลเจลลัมตามยาวจะกระทบกับ sulcus [ 24 ] [ 23 ]
เมื่อรวมกับรายละเอียดโครงสร้างและพันธุกรรมอื่นๆ องค์กรนี้แสดงให้เห็นถึงความสัมพันธ์ที่ใกล้ชิดระหว่างไดโนแฟลเจลเลต อะพิคอมเพล็กซาและซิลิเอตซึ่งเรียกรวมกันว่า อัลวี โอเลต[ 22 ]
ตารางไดโนแฟลเจลเลตสามารถจัดกลุ่มได้เป็น 6 "ประเภทตาราง": จิมโนดินอยด์ซูเอสซอยด์กอนยาอูลาคอยด์ - เพอริดิ นิออยด์ แน นโนเซรา ทอปซิออยด์ไดโนฟิซิออยด์และโปรโรเซนทรอยด์[ 25 ]
ไดโนแฟลเจลเลตส่วนใหญ่มีพลาสติดที่ได้มาจากเอนโดซิมไบโอซิสทุติยภูมิของสาหร่ายแดง อย่างไรก็ตาม ไดโนแฟลเจลเลตที่มีพลาสติดที่ได้มาจากสาหร่ายสีเขียวและเอนโดซิมไบโอซิสตติยภูมิของไดอะตอมก็ถูกค้นพบเช่นกัน[ 26 ] เช่นเดียวกับสิ่งมีชีวิตสังเคราะห์แสงอื่นๆ ไดโนแฟลเจลเลตมีคลอโรฟิลล์aและ c2 และแคโรทีนอยด์เบต้าแคโรทีน ไดโนแฟลเจลเลตยังผลิตแซนโทฟิลล์ได้แก่เพอริ ดินิน ไดนอกแซนทินและ ไดอะได นอกแซ นทิน เม็ดสี เหล่านี้ทำให้ไดโนแฟลเจลเลตหลายชนิดมีสีน้ำตาลทองที่เป็นเอกลักษณ์ อย่างไรก็ตาม ไดโนแฟลเจลเลตKarenia brevis , Karenia mikimotoiและKarlodinium micrumได้รับเม็ดสีอื่นๆ ผ่านเอนโดซิมไบโอซิส รวมถึงฟูโคแซนทิน[ 27 ] สิ่งนี้ชี้ให้เห็นว่าคลอโรพลาสต์ของพวกมันถูกรวมเข้าด้วยกันโดยเหตุการณ์เอนโดซิมไบโอซิส หลายครั้ง ที่เกี่ยวข้องกับรูปแบบที่มีสีอยู่แล้วหรือรูปแบบที่ไม่มีสีในภายหลัง การค้นพบพลาสติดในApicomplexaทำให้บางคนเสนอว่าพวกมันได้รับการสืบทอดมาจากบรรพบุรุษร่วมกันของทั้งสองกลุ่ม แต่ไม่มีสายพันธุ์พื้นฐานใดที่มีพลาสติด อย่างไรก็ตาม เซลล์ไดโนแฟลเจลเลตประกอบด้วยออร์แกเนลล์ทั่วไป เช่น เอนโด พลาสมิกเรติคูลัม แบบหยาบและแบบเรียบ อุปกรณ์กอลจิไมโตคอนเดรีย เม็ด ไขมันและแป้งและแวคิวโอล อาหาร บางชนิดยังพบว่ามีออร์แกเนลล์ที่ไวต่อแสง เช่นจุดตาหรือสติ๊กมาหรือนิวเคลียสขนาดใหญ่ที่มีนิวคลีโอลัส ที่เด่นชัด ไดโนแฟลเจลเลตErythropsidiniumมีตาที่เล็กที่สุดเท่าที่รู้จัก[ 28 ]
บางชนิดที่ไม่มีเปลือกหุ้มจะมีโครงกระดูกภายในที่ประกอบด้วย องค์ประกอบ ซิลิกาที่ มีรูปร่างคล้ายดาวสองอัน ซึ่งมีหน้าที่ที่ไม่ทราบแน่ชัด และสามารถพบได้ในรูปของไมโครฟอสซิล Tappan [ 29 ]ได้ทำการสำรวจไดโนแฟลเจลเลตที่มีโครงกระดูกภายในซึ่งรวมถึงคำอธิบายโดยละเอียดครั้งแรกของเพนตาสเตอร์ในActiniscus pentasteriasโดยอาศัยกล้องจุลทรรศน์อิเล็กตรอนแบบสแกนพวกมันถูกจัดอยู่ในอันดับGymnodinialesอันดับย่อยActiniscineae [ 3 ]
โครงสร้างและการก่อตัวของธีคา
การก่อตัวของแผ่นเทคาลได้รับการศึกษาอย่างละเอียดผ่านการศึกษาโครงสร้างระดับอัลตรา[ 21 ]
นิวเคลียสไดโนแฟลเจลเลต: ไดโนคาริโอน
'ไดโนแฟลเจลเลตแกนกลาง' ( ไดโนคาริโอต ) มี นิวเคลียสรูปแบบพิเศษที่เรียก ว่า ไดโนคาริออนซึ่งโครโมโซมจะยึดติดกับเยื่อหุ้มนิวเคลียส โครโมโซมเหล่านี้มี ฮิสโตนจำนวนน้อยลงแทนที่ฮิสโตน นิวเคลียสของไดโนแฟลเจลเลตจะมีโปรตีนนิวเคลียร์ตระกูลใหม่ที่เด่น ซึ่งดูเหมือนจะมีต้นกำเนิดมาจากไวรัส จึงเรียกว่าโปรตีนนิวคลีโอโปรตีนไวรัสไดโนแฟลเจลเลต (DVNPs) ซึ่งมีคุณสมบัติเป็นเบสสูง จับกับ DNA ด้วยความสัมพันธ์ที่คล้ายกับฮิสโตน และพบได้ในรูปแบบที่ได้รับการดัดแปลงหลังการแปลหลายรูปแบบ[ 30 ]นิวเคลียสของไดโนแฟลเจลเลตยังคงควบแน่นตลอดระยะอินเตอร์เฟส แทนที่จะเป็นเฉพาะในช่วงไมโทซิส ซึ่งเป็นระยะปิดและเกี่ยวข้องกับ แกนไมโทซิสที่อยู่นอกนิวเคลียสโดยเฉพาะ[ 31 ]นิวเคลียสชนิดนี้เคยถูกพิจารณาว่าเป็นตัวกลางระหว่างบริเวณนิวคลีออยด์ของโปรคาริโอตและนิวเคลียสที่แท้จริงของยูคาริโอตจึงถูกเรียกว่า " เมโซคาริโอต " แต่ปัจจุบันถือว่าเป็นลักษณะที่พัฒนาแล้วมากกว่าลักษณะดั้งเดิม (เช่น บรรพบุรุษของไดโนแฟลเจลเลตมีนิวเคลียสแบบยูคาริโอตทั่วไป) นอกจากไดโนคาริโอตแล้ว DVNP ยังสามารถพบได้ในกลุ่มของไดโนแฟลเจลเลตพื้นฐาน (ที่รู้จักกันในชื่อ Marine Alveolatesหรือ "MALVs") ซึ่งแตกแขนงเป็นกลุ่มพี่น้องกับไดโนคาริโอต ( Syndiniales ) [ 32 ]
การจำแนกประเภท
ความทั่วไป
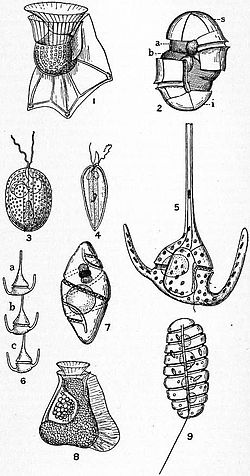
ไดโนแฟลเจลเลตเป็นโปรติสต์ และได้รับการจัดจำแนกโดยใช้ทั้งประมวลกฎการตั้งชื่อทางพฤกษศาสตร์สากล (ICBN ซึ่งปัจจุบันเปลี่ยนชื่อเป็น ICN) และประมวลกฎการตั้งชื่อทางสัตววิทยาสากล (ICZN) ประมาณครึ่งหนึ่งของสายพันธุ์ไดโนแฟลเจลเลตที่ยังมีชีวิตอยู่เป็นออโตโทรฟที่มีคลอโรพลาสต์ และอีกครึ่งหนึ่งเป็นเฮเทอโรโทรฟที่ไม่สังเคราะห์แสง
ไดโน แฟลเจลเลต เพอริดินินซึ่งตั้งชื่อตามพลาสติดเพอริดินินของพวกมัน ดูเหมือนจะเป็นบรรพบุรุษของสายพันธุ์ไดโนแฟลเจลเลต เกือบครึ่งหนึ่งของสายพันธุ์ที่รู้จักทั้งหมดมีคลอโรพลาสต์ ซึ่งเป็นพลาสติดเพอริดินินดั้งเดิมหรือพลาสติดใหม่ที่ได้รับมาจากสายพันธุ์อื่นของสาหร่ายเซลล์เดียวผ่านเอนโดซิมไบโอซิส สายพันธุ์ที่เหลือได้สูญเสียความสามารถในการสังเคราะห์แสงและปรับตัวให้เข้ากับวิถีชีวิต แบบเฮเทอโรโทรฟิก ปรสิต หรือ เคล็ปโตพลา สต์ [ 33 ] [ 34 ]
ไดโนแฟลเจลเลตส่วนใหญ่ (แต่ไม่ใช่ทั้งหมด) มีไดโนคาริออ น ซึ่งจะอธิบายรายละเอียดด้านล่าง (ดู: วงจรชีวิตด้านล่าง) ไดโนแฟลเจลเลตที่มีไดโนคาริออนจัดอยู่ในกลุ่มไดโนคาริโอตาในขณะที่ไดโนแฟลเจลเลตที่ไม่มีไดโนคาริออนจัดอยู่ในกลุ่มซินดินิอาเลส
แม้ว่าจะถูกจัดอยู่ในกลุ่มยูคาริโอตแต่นิวเคลียสของไดโนแฟลเจลเลตก็ไม่ได้มีลักษณะเฉพาะของยูคาริโอต เนื่องจากบางส่วนขาดฮิสโตนและนิวคลีโอโซมและยังคงรักษาโครโมโซมที่ควบแน่นอย่างต่อเนื่องในระหว่างการแบ่ง เซลล์ แบบไมโทซิส นิวเคลียสของไดโนแฟลเจลเลตถูกเรียกว่า 'เมโซคาริโอต' โดยดอดจ์ (1966) [ 35 ]เนื่องจากมีลักษณะที่เป็นตัวกลางระหว่างบริเวณดีเอ็นเอที่ขดตัวของแบคทีเรียโปรคาริโอตและนิวเคลียสยูคาริโอตที่กำหนดไว้อย่างชัดเจน อย่างไรก็ตาม กลุ่มนี้ก็มีออร์แกเนลล์ ที่เป็นลักษณะเฉพาะของยูคาริโอต เช่น กอลจิ บอดี้ ไมโตคอนเดรีย และคลอโรพลาสต์[ 36 ]
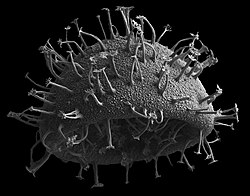
Jakob Schiller (1931–1937) ได้ให้คำอธิบายเกี่ยวกับสปีชีส์ทั้งหมด ทั้งในทะเลและน้ำจืด ที่รู้จักในเวลานั้น[ 37 ]ต่อมา Alain Sournia (1973, 1978, 1982, 1990, 1993) ได้จัดทำรายการอนุกรมวิธานใหม่ที่ตีพิมพ์หลังจาก Schiller (1931–1937) [ 38 ] [ 39 ] [ 40 ] [ 41 ] [ 42 ] Sournia (1986) ได้ให้คำอธิบายและภาพประกอบของสกุลไดโนแฟลเจลเลตในทะเล โดยไม่รวมข้อมูลในระดับสปีชีส์[ 43 ]ดัชนีล่าสุดเขียนโดย Gómez [ 11 ]
การระบุตัวตน
มีการตีพิมพ์โมโนกราฟอนุกรมวิธานภาษาอังกฤษที่ครอบคลุมจำนวนสายพันธุ์จำนวนมากสำหรับอ่าวเม็กซิโก[ 44 ]มหาสมุทรอินเดีย[ 45 ]หมู่เกาะอังกฤษ[ 46 ]ทะเลเมดิเตอร์เรเนียน[ 47 ]และทะเลเหนือ[ 48 ]
แหล่งข้อมูลหลักสำหรับการระบุไดโนแฟลเจลเลตน้ำจืดคือSüsswasser Flora [ 49 ]
สามารถใช้แคลโคฟลูออร์ไวท์ ในการย้อมแผ่นเทคาลในไดโนแฟลเจลเลตที่มีเกราะได้ [ 50 ]
นิเวศวิทยาและสรีรวิทยา
แหล่งที่อยู่อาศัย
ไดโนแฟลเจลเลตพบได้ในสภาพแวดล้อมทางน้ำทุกประเภท ได้แก่ น้ำทะเล น้ำกร่อย และน้ำจืด รวมถึงในหิมะหรือน้ำแข็ง นอกจากนี้ยังพบได้ทั่วไปในสภาพแวดล้อมใต้ทะเลและน้ำแข็งในทะเล
เอนโดซิมไบออนต์
Zooxanthellaeทั้งหมดเป็นไดโนแฟลเจลเลต และส่วนใหญ่เป็นสมาชิกในวงศ์ Symbiodiniaceae (เช่น สกุลSymbiodinium ) [ 51 ]ความสัมพันธ์ระหว่างSymbiodiniumกับปะการัง ที่สร้างแนวปะการัง เป็นที่รู้จักกันอย่างกว้างขวาง อย่างไรก็ตามZooxanthellae ที่เป็นเอนโดซิมไบออน อาศัยอยู่ในสัตว์ไม่มีกระดูกสันหลังและโปรติสต์อื่นๆ อีกมากมาย เช่นดอกไม้ทะเลแมงกะพรุนทากทะเล หอยยักษ์Tridacnaและเรดิโอลาเรียนและฟอรามินิเฟอแรนหลาย ชนิด [ 52 ] ได โน แฟล เจลเลตที่มีอยู่ในปัจจุบันหลายชนิดเป็นปรสิต (ในที่นี้หมายถึงสิ่งมีชีวิตที่กินเหยื่อจากภายใน เช่นเอนโดปรสิตหรือที่ยังคงเกาะติดกับเหยื่อเป็นเวลานาน เช่น เอ็กโตปรสิต) พวกมันสามารถเป็นปรสิตในโฮสต์ที่เป็นสัตว์หรือโปรติสต์ได้Protoodinium, Crepidoodinium, PiscinoodiniumและBlastodiniumยังคงมีพลาสติดอยู่ในตัวขณะที่กินแพลงก์ตอนสัตว์หรือปลาเป็นอาหาร ในไดโนแฟลเจลเลตที่เป็นปรสิตส่วนใหญ่ ระยะติดเชื้อจะมีลักษณะคล้ายเซลล์ไดโนแฟลเจลเลตที่เคลื่อนที่ได้ทั่วไป
กลยุทธ์ด้านโภชนาการ
ไดโนแฟลเจลเลตมีกลยุทธ์ทางโภชนาการ 3 แบบ ได้แก่การสังเคราะห์แสงการผสมและการพึ่งพา อาหารจากสิ่งมีชีวิตอื่น ได โน แฟลเจลเลตที่สังเคราะห์แสงได้อาจเป็นโฟโตออโตโทรฟหรือออโซโทรฟ ไดโนแฟลเจลเลตแบบผสมสามารถสังเคราะห์แสงได้ แต่ก็พึ่งพาอาหารจากสิ่งมีชีวิตอื่นด้วย ไดโนแฟลเจลเลตแบบผสมที่สามารถสังเคราะห์แสงหรือพึ่งพาอาหารจากสิ่งมีชีวิตอื่นได้เพียงพอต่อโภชนาการ จะถูกจัดเป็นแอมฟิโทรฟิก หากสิ่งมีชีวิตทั้งสองรูปแบบมีความจำเป็น สิ่งมีชีวิตนั้นจะเรียกว่า ผสมแบบเคร่งครัด (mixotrophic sensu stricto ) ไดโนแฟลเจลเลตที่ดำรงชีวิตอิสระบางชนิดไม่มีคลอโรพลาสต์ แต่มีเอนโดซิมไบออนที่สังเคราะห์แสงได้ ไดโนแฟลเจลเลตบางชนิดอาจใช้คลอโรพลาสต์จากสิ่งมีชีวิตอื่น (cleptochloroplasts) ที่ได้มาจากอาหาร ( kleptoplasty ) ไดโนแฟลเจลเลตบางชนิดอาจกินสิ่งมีชีวิตอื่นเป็นอาหารในฐานะผู้ล่าหรือปรสิต[ 53 ]
ส่วนประกอบของอาหารประกอบด้วยแบคทีเรีย สาหร่ายสีเขียวแกมน้ำเงิน ไดอะตอม ซิลิเอต และไดโนแฟลเจลเลตอื่นๆ[ 54 ] [ 55 ] [ 56 ] [ 57 ] [ 58 ] [ 59 ] [ 60 ]
กลไกการจับและการ กินในไดโนแฟลเจลเลตมีความหลากหลายมาก ไดโนแฟลเจลเลตหลายชนิด ทั้งแบบมีเปลือกหุ้ม (เช่นCeratium hirundinella [ 59 ] Peridinium globulus [ 57 ] ) และแบบไม่มีเปลือกหุ้ม (เช่นOxyrrhis marina [ 55 ] Gymnodinium sp. [ 61 ]และKofoidinium spp. [ 62 ] ) จะดึงเหยื่อไปยังบริเวณร่องของเซลล์ (ไม่ว่าจะผ่านกระแสน้ำที่สร้างขึ้นโดยแฟลเจลลาหรือผ่านส่วนขยายของซูโดโพเดีย) และกินเหยื่อผ่านร่องนั้น ในProtoperidinium spp. หลายชนิด เช่นP. conicumจะมีการยื่นแผ่นกรองอาหารขนาดใหญ่ ซึ่งเป็นซูโดโพเดียที่เรียกว่าพัลเลียม เพื่อจับเหยื่อซึ่งจะถูกย่อยภายนอกเซลล์ ในภายหลัง (= การกินแบบพัลเลียม) [ 63 ] [ 64 ] Oblea , ZygabikodiniumและDiplopsalisเป็นเพียงสกุลไดโนแฟลเจลเลตอื่น ๆ ที่ทราบว่าใช้กลไกการกินอาหารแบบนี้[ 64 ] [ 65 ] [ 66 ] Gymnodinium fungiformeซึ่งมักพบเป็นสิ่งปนเปื้อนในวัฒนธรรมสาหร่ายหรือซีลิเอต จะกินอาหารโดยการเกาะติดกับเหยื่อและดูดกลืนไซโตพลาสซึมของเหยื่อผ่านก้านที่ยืดหดได้[ 67 ]สกุลที่เกี่ยวข้องสองสกุลคือPolykrikosและNeatodiniumจะยิงออร์แกเนลล์ที่มีลักษณะคล้ายฉมวกเพื่อจับเหยื่อ[ 68 ]
ไดโนแฟลเจลเลตแบบมิกโซโทรฟิกบางชนิดสามารถผลิตสารพิษต่อระบบประสาทที่มีฤทธิ์ต้านการกินโคพีพอดขนาดใหญ่ และเพิ่มความสามารถของไดโนแฟลเจลเลตในการล่าโคพีพอดขนาดใหญ่ได้ สายพันธุ์ที่เป็นพิษของKarlodinium veneficum ผลิตคาร์โลทอกซินที่ฆ่าผู้ล่าที่กินเข้าไป จึงลดจำนวนประชากรผู้ล่าและทำให้เกิดการเพิ่มจำนวนของ K. veneficumทั้งสายพันธุ์ที่เป็นพิษและไม่เป็นพิษนอกจากนี้ การผลิตคาร์โลทอกซินยังช่วยเพิ่มความสามารถในการล่าของK. veneficumโดยทำให้เหยื่อขนาดใหญ่เป็นอัมพาต[ 69 ] K. armigerมีแนวโน้มที่จะล่าโคพีพอดมากกว่าโดยการปล่อยสารพิษต่อระบบประสาทที่มีฤทธิ์รุนแรงซึ่งทำให้เหยื่อเป็นอัมพาตเมื่อสัมผัส เมื่อK. armigerมีอยู่เป็นจำนวนมากพอ พวกมันสามารถกำจัดประชากรโคพีพอดที่เป็นเหยื่อได้ทั้งหมด[ 70 ]
กลไกการกินอาหารของไดโนแฟลเจลเลตในมหาสมุทรยังคงไม่เป็นที่รู้จัก แม้ว่าจะมีการสังเกตส่วนขยายของซูโดโพเดียในPodolampas bipesก็ตาม[ 71 ]
รงควัตถุในไดโนแฟลเจลเลต
ไดโนแฟลเจลเลตมีชุดรงควัตถุสังเคราะห์แสงที่โดดเด่น ซึ่งช่วยให้พวกมันสามารถอยู่รอดและเติบโตได้ในสภาพแวดล้อมทางน้ำที่หลากหลาย เช่นเดียวกับแพลงก์ตอนพืชอื่นๆ ไดโนแฟลเจลเลตหลายชนิดมีคลอโรฟิลล์เอและคลอโรฟิลล์ซี ซึ่งจำเป็นต่อการสังเคราะห์แสงและการจับพลังงานแสง[ 72 ]อย่างไรก็ตาม แตกต่างจากสาหร่ายสีเขียวและพืชชั้นสูง พวกมันไม่มีคลอโรฟิลล์บี แต่พวกมันใช้คลอโรฟิลล์ซี2 ซึ่งมีประสิทธิภาพมากกว่าในการดูดซับแสงสีฟ้าอมเขียว ทำให้พวกมันปรับตัวได้ดีในสภาพแสงน้อยหรือในน้ำลึก[ 73 ]รงควัตถุเหล่านี้ร่วมกับแคโรทีนอยด์มีส่วนทำให้ไดโนแฟลเจลเลตมีสีที่เป็นเอกลักษณ์ ซึ่งมีตั้งแต่สีน้ำตาลทองไปจนถึงสีแดง
เม็ดสีเฉพาะในไดโนแฟลเจลเลตคือเพอริดินินซึ่งเป็นแคโรทีนอยด์ชนิดพิเศษที่มีบทบาทสำคัญในการเก็บเกี่ยวแสงและการถ่ายโอนพลังงานไปยังคลอโรฟิลล์เอ[ 74 ]เพอริดินินมีประสิทธิภาพสูงในการดักจับแสงสีฟ้า ซึ่งทะลุทะลวงลงไปในน้ำได้ลึกกว่า ทำให้ไดโนแฟลเจลเลตหลายชนิดได้เปรียบในการแข่งขันในสภาพแวดล้อมที่มีการแบ่งชั้นหรือขุ่น[ 75 ]นอกจากนี้ ไดโนแฟลเจลเลตยังมีแคโรทีนอยด์อื่นๆ เช่น ไดอะดินอกแซนทินและไดนอกแซนทิน ซึ่งมีบทบาทสำคัญในการป้องกันแสงโดยการกระจายพลังงานแสงส่วนเกินและป้องกันความเครียดจากออกซิเดชันภายใต้ความเข้มแสงสูง[ 76 ] เม็ดสีเหล่านี้จำเป็นสำหรับการปรับตัวให้เข้ากับแสง ทำให้ไดโนแฟลเจลเลตสามารถอยู่รอดได้ภายใต้สภาวะแสงที่ผันผวน
ไดโนแฟลเจลเลตไม่ได้พึ่งพาเม็ดสีสังเคราะห์แสงเพียงอย่างเดียวในการให้พลังงาน หลายชนิดเป็นเฮเทอโรโทรฟิกหรือมิกโซโทรฟิก ซึ่งหมายความว่าพวกมันสามารถได้รับสารอาหารทั้งจากการสังเคราะห์แสงและการล่าเหยื่อ[ 77 ]ไดโนแฟลเจลเลตแบบพึ่งพาอาศัยกัน เช่น Symbiodinium มีบทบาททางนิเวศวิทยาที่สำคัญโดยการสร้างความสัมพันธ์แบบพึ่งพาซึ่งกันและกันกับปะการัง ซึ่งเม็ดสีของพวกมันเป็นตัวขับเคลื่อนการสังเคราะห์แสงและการผลิตพลังงานที่ช่วยรักษาระบบนิเวศแนวปะการัง[ 78 ]องค์ประกอบของเม็ดสีที่เป็นเอกลักษณ์ของไดโนแฟลเจลเลตยังก่อให้เกิดปรากฏการณ์ขนาดใหญ่ เช่น การระบาดของสาหร่ายที่เป็นอันตรายและปรากฏการณ์น้ำแดง ซึ่งความเข้มข้นสูงของเซลล์ที่มีเม็ดสีทำให้เกิดการเปลี่ยนสีอย่างมากของน้ำทะเลชายฝั่งและอาจก่อให้เกิดผลกระทบที่เป็นพิษได้[ 79 ]
บุปผา
การแนะนำ
โดยทั่วไปแล้ว การแพร่กระจายของไดโนแฟลเจลเลตนั้นคาดเดาได้ยาก มีระยะเวลาสั้น มีความหลากหลายของชนิดพันธุ์ต่ำ และมีการเปลี่ยนแปลงของชนิดพันธุ์น้อย[ 80 ]ความหลากหลายของชนิดพันธุ์ที่ต่ำอาจเกิดจากหลายปัจจัย วิธีหนึ่งที่การขาดความหลากหลายอาจเกิดขึ้นในการแพร่กระจายคือการลดลงของการล่าเหยื่อและการลดลงของการแข่งขัน อย่างแรกอาจเกิดขึ้นได้จากการที่ผู้ล่าปฏิเสธไดโนแฟลเจลเลต เช่น โดยการลดปริมาณอาหารที่มันสามารถกินได้ นอกจากนี้ยังช่วยป้องกันการเพิ่มขึ้นของแรงกดดันจากการล่าเหยื่อในอนาคตโดยทำให้ผู้ล่าที่ปฏิเสธมันขาดพลังงานในการสืบพันธุ์ จากนั้นชนิดพันธุ์หนึ่งๆ ก็สามารถยับยั้งการเติบโตของคู่แข่งได้ จึงทำให้เกิดการครอบงำ[ 81 ]
การแพร่กระจายของสาหร่ายที่เป็นอันตราย
บางครั้งไดโนแฟลเจลเลตจะเพิ่มจำนวนขึ้นอย่างหนาแน่นจนมีมากกว่าหนึ่งล้านเซลล์ต่อมิลลิลิตร ในสภาวะเช่นนี้ พวกมันสามารถผลิตสารพิษ (โดยทั่วไปเรียกว่าไดโนท็อกซิน ) ในปริมาณที่สามารถฆ่าปลาได้ และสะสมอยู่ในสัตว์ที่กรองอาหาร เช่นหอยซึ่งอาจส่งต่อไปยังคนที่กินหอยเหล่านั้นได้ ปรากฏการณ์นี้เรียกว่าน้ำแดงเนื่องจากสีแดงที่เกิดจากการเพิ่มจำนวนของไดโนแฟลเจลเลตในน้ำ ไดโนแฟลเจลเลตที่ไม่มีสีบางชนิดก็อาจก่อให้เกิดการเพิ่มจำนวนที่เป็นพิษได้เช่นกัน เช่นPfiesteriaการเพิ่มจำนวนของไดโนแฟลเจลเลตบางชนิดไม่เป็นอันตราย แสงสีฟ้าที่มองเห็นในน้ำทะเลในเวลากลางคืนมักมาจากการเพิ่มจำนวนของ ไดโนแฟลเจลเลต เรือง แสง ซึ่งจะปล่อยแสงวาบสั้นๆ เมื่อถูกรบกวน

ปรากฏการณ์น้ำทะเลสีแดงเกิดขึ้นเนื่องจากไดโนแฟลเจลเลตสามารถขยายพันธุ์ได้อย่างรวดเร็วและมีจำนวนมากอันเป็นผลมาจากสารอาหารที่อุดมสมบูรณ์ในน้ำ พวกมันมีสารพิษที่ส่งผลกระทบต่อสิ่งมีชีวิตในทะเลโดยรอบและผู้คนที่บริโภคพวกมัน[ 82 ]พาหะที่สำคัญอย่างหนึ่งคือหอยซึ่งสามารถนำมาซึ่งโรคทั้งที่ไม่ร้ายแรงและร้ายแรงได้ สารพิษชนิดหนึ่งคือแซกซิโทซินซึ่ง เป็น สารพิษต่อระบบประสาทที่ทำให้เป็นอัมพาต อย่างรุนแรง[ 83 ] [ 84 ] [ 85 ]
การป้อน ฟอสเฟตของมนุษย์ยิ่งกระตุ้นให้เกิดปรากฏการณ์น้ำแดง ดังนั้นจึงมีความสนใจอย่างมากในการเรียนรู้เพิ่มเติมเกี่ยวกับไดโนแฟลเจลเลต ทั้งจากมุมมองทางการแพทย์และเศรษฐกิจ ไดโนแฟลเจลเลตเป็นที่รู้จักกันดีว่ามีความสามารถพิเศษในการดักจับฟอสฟอรัสอินทรีย์ที่ละลายน้ำเพื่อใช้เป็นสารอาหาร P พบว่า HAS หลายชนิดมีความหลากหลายและมีกลไกการทำงานที่แตกต่างกันในการใช้ DOP ประเภทต่างๆ[ 83 ] [ 84 ] [ 85 ]นิเวศวิทยาของการแพร่กระจายของสาหร่ายที่เป็นอันตรายได้รับการศึกษาอย่างกว้างขวาง[ 86 ]
การเรืองแสงทางชีวภาพ

ในเวลากลางคืน น้ำอาจมีลักษณะเป็นประกายระยิบระยับเนื่องจากการเรืองแสงทางชีวภาพของไดโนแฟลเจลเลต[ 87 ] [ 88 ]ไดโนแฟลเจลเลตมากกว่า 18 สกุลสามารถเรืองแสงได้[ 89 ]และส่วนใหญ่จะปล่อยแสงสีฟ้าอมเขียว[ 90 ]สปีชีส์เหล่านี้มีสซินทิลลอน ซึ่งเป็นส่วนประกอบ ของไซโตพลาสซึมแต่ละส่วน (มีเส้นผ่านศูนย์กลางประมาณ 0.5 ไมโครเมตร) กระจายอยู่ส่วนใหญ่ในบริเวณคอร์เทกซ์ของเซลล์ ซึ่งเป็นส่วนที่ยื่นออกมาจากแวคิวโอลหลักของเซลล์ พวกมันมีไดโนแฟลเจลเลตลูซิเฟอเรส ซึ่งเป็น เอนไซม์หลักที่เกี่ยวข้องกับการเรืองแสงทางชีวภาพของไดโนแฟลเจลเลต และลูซิเฟอรินซึ่งเป็นวงแหวนเตตระไพโรลที่ได้จากคลอโรฟิลล์ ทำหน้าที่เป็นสารตั้งต้นในปฏิกิริยาการสร้างแสง การเรืองแสงเกิดขึ้นเป็นแสงวาบสีฟ้าสั้นๆ (0.1 วินาที) (สูงสุด 476 นาโนเมตร) เมื่อถูกกระตุ้น โดยปกติเกิดจากการรบกวนทางกล ดังนั้น เมื่อได้รับการกระตุ้นทางกล เช่น จากเรือ การว่ายน้ำ หรือคลื่น จึงสามารถมองเห็นแสงสีฟ้าระยิบระยับเปล่งออกมาจากผิวน้ำทะเลได้[ 91 ]
การเรืองแสงของไดโนแฟลเจลเลตถูกควบคุมโดยนาฬิกาชีวภาพและเกิดขึ้นเฉพาะในเวลากลางคืนเท่านั้น[ 92 ]สายพันธุ์ที่เรืองแสงและไม่เรืองแสงสามารถเกิดขึ้นได้ในสปีชีส์เดียวกัน จำนวนแสงระยิบระยับจะสูงกว่าในเวลากลางคืนมากกว่าในเวลากลางวัน และจะลดลงในช่วงท้ายของคืน ซึ่งเป็นช่วงเวลาที่มีการเรืองแสงสูงสุด[ 93 ]
ปฏิกิริยาลูซิเฟอริน-ลูซิเฟอเรสที่ทำให้เกิดการเรืองแสงทางชีวภาพนั้นมีความไวต่อค่า pH [ 91 ]เมื่อค่า pH ลดลง ลูซิเฟอเรสจะเปลี่ยนรูปร่าง ทำให้ลูซิเฟอริน โดยเฉพาะเตตระไพโรล สามารถจับได้[ 91 ]ซึ่งเกิดขึ้นจากความปั่นป่วนทางกลที่เหนี่ยวนำให้เกิดศักย์ไฟฟ้าในเยื่อหุ้มเซลล์[ 94 ]บางชนิดยังมีโปรตีนที่จับกับลูซิเฟอริน ซึ่งจะจับกับลูซิเฟอรินก่อนที่จะเกิดความเป็นกรดเพื่อป้องกันการเกิดออกซิเดชันเอง จากนั้นจึงปล่อยลูซิเฟอรินออกมาเมื่อโครงสร้างเปลี่ยนไป ทำให้ลูซิเฟอรินทำงานได้[ 95 ]
"สมมติฐานการป้องกันออกซิเจน" ชี้ให้เห็นว่าการเรืองแสงทางชีวภาพวิวัฒนาการขึ้นครั้งแรกเพื่อตอบสนองต่อเหตุการณ์ออกซิเดชันครั้งใหญ่เมื่อออกซิเจนสะสมอยู่ในชั้นบรรยากาศ สิ่งมีชีวิตต้องปรับตัวให้เข้ากับความเครียดจากออกซิเดชันจากสารออกซิเจนที่ว่องไว ส่งผลให้เกิดการปล่อยแสงเป็นผลพลอยได้[ 96 ]แม้ว่าการเรืองแสงทางชีวภาพจะมีต้นกำเนิดมาจากการปรับตัวเข้ากับสิ่งแวดล้อม แต่ปฏิสัมพันธ์ทางนิเวศวิทยาก็ได้เปลี่ยนบทบาทของมันไป
ไดโนแฟลเจลเลตสามารถใช้ไบโอเรืองแสงเป็นกลไกป้องกันตัวได้ พวกมันสามารถทำให้ผู้ล่าตกใจด้วยแสงวาบ ใช้เป็นสัญญาณเตือนภัยเพื่อเตือนถึงความเป็นพิษ หรือขับไล่ผู้ล่าที่อาจเกิดขึ้นด้วยผลกระทบทางอ้อม เช่น "สัญญาณเตือนภัย" [ 97 ]ในสมมติฐาน "สัญญาณเตือนภัย" ไบโอเรืองแสงดึงดูดความสนใจไปยังไดโนแฟลเจลเลตและผู้โจมตี ทำให้ผู้ล่ามีความเสี่ยงต่อการถูกล่าจากระดับโภชนาการที่สูงกว่ามากขึ้น[ 91 ]แม้ว่านี่จะเป็นสมมติฐานที่ได้รับการยอมรับมากที่สุด แต่ก็มีกลไกอื่นที่เป็นไปได้ซึ่งมุ่งเน้นไปที่พฤติกรรมของผู้กินพืช พบว่าโคพีพอดมีอัตราการกระโดดที่เพิ่มขึ้นเมื่อมีไบโอเรืองแสง ซึ่งทำให้ผู้ล่าระดับสูงกว่าสามารถตรวจจับพวกมันได้ง่ายขึ้น[ 98 ]สิ่งนี้ชี้ให้เห็นว่าพฤติกรรมของโคพีพอดเป็นสิ่งที่ทำให้เกิดผลกระทบ "สัญญาณเตือนภัย" มากกว่าไบโอเรืองแสงเพียงอย่างเดียว สมมติฐานทั้งหมดนี้บ่งชี้ว่าการเรืองแสงทางชีวภาพเป็นประโยชน์ต่อการอยู่รอด ซึ่งเป็นเหตุผลว่าทำไมจึงเป็นลักษณะที่พบได้ทั่วไปในไดโนแฟลเจลเลต
อ่าวระบบนิเวศไดโนแฟลเจลเลตเรืองแสงเป็นหนึ่งในระบบนิเวศที่หายากและเปราะบางที่สุด[ 99 ]โดยอ่าวที่มีชื่อเสียงที่สุด ได้แก่ อ่าวเรืองแสงในลาปาร์เกรา ลาฮาส เปอร์โตริโก; อ่าวมอสquito ในวีเกส เปอร์โตริโก ; และเขตอนุรักษ์ธรรมชาติลาสกาเบซัสเดซานฮวนฟาจาร์โด เปอร์โตริโกนอกจากนี้ ยังมีทะเลสาบเรืองแสงอยู่ใกล้กับอ่าวมอนเตโก ประเทศจาเมกา และท่าเรือเรืองแสงล้อมรอบเมืองคาสทีน รัฐเมน[ 100 ]ในสหรัฐอเมริกา ฟลอริดาตอนกลางเป็นที่ตั้งของทะเลสาบอินเดียนริเวอร์ซึ่งอุดมไปด้วยไดโนแฟลเจลเลตในฤดูร้อนและซีเทโนฟอร์เรืองแสงในฤดูหนาว[ 101 ]
การผลิตไขมันและสเตอรอล
ไดโนแฟลเจลเลตผลิตลิปิดและสเตอรอลที่มีลักษณะเฉพาะ[ 102 ]สเตอรอลชนิดหนึ่งเป็นลักษณะเฉพาะของไดโนแฟลเจลเลตและเรียกว่าไดโนสเตอรอล
ขนส่ง
เปลือกหุ้มของไดโนแฟลเจลเลตสามารถจมลงสู่พื้นทะเลอย่างรวดเร็วในหิมะทะเล[ 103 ]
วงจรชีวิต
การแนะนำ
ไดโนแฟลเจลเลตมีวงจรชีวิตแบบแฮพลอยด์โดยอาจมีข้อยกเว้นคือNoctilucaและญาติของมัน[ 3 ] วงจรชีวิตมักเกี่ยวข้องกับการสืบพันธุ์แบบไม่อาศัยเพศโดยวิธีการไมโทซิส ไม่ว่าจะผ่านdesmoschisisหรือeleutheroschisisวงจรชีวิตที่ซับซ้อนกว่านั้นเกิดขึ้น โดยเฉพาะอย่างยิ่งกับไดโนแฟลเจลเลตที่เป็นปรสิต การสืบพันธุ์แบบอาศัยเพศก็เกิดขึ้นเช่นกัน[ 104 ]แม้ว่าวิธีการสืบพันธุ์แบบนี้จะพบได้ในไดโนแฟลเจลเลตเพียงเล็กน้อยเท่านั้น[ 105 ]การสืบพันธุ์แบบนี้เกิดขึ้นโดยการรวมตัวของสองตัวเพื่อสร้างไซโกต ซึ่งอาจเคลื่อนที่ได้ตามปกติของไดโนแฟลเจลเลต และเรียกว่า planozygote ไซโกตนี้อาจสร้างระยะพักตัวหรือ hypnozygoteในภายหลังซึ่งเรียกว่าซีสต์ไดโนแฟลเจล เลต หรือไดโนซิสต์หลังจาก (หรือก่อน) การงอกของซีสต์ ลูกอ่อนจะผ่านกระบวนการไมโอซิส เพื่อสร้าง เซลล์แฮพลอยด์ใหม่ไดโนแฟลเจลเลตดูเหมือนจะสามารถดำเนิน การกระบวนการ ซ่อมแซม DNA หลายอย่าง ที่สามารถจัดการกับความเสียหายของ DNA ประเภทต่างๆ ได้[ 106 ]


ซีสต์ของไดโนแฟลเจลเลต
วงจรชีวิตของไดโนแฟลเจลเลตหลายชนิดประกอบด้วยระยะเบนทิกที่ไม่มีแฟลเจลเลตอย่างน้อยหนึ่งระยะในรูปของซีสต์ ซีสต์ของไดโนแฟลเจลเลตประเภทต่างๆ ส่วนใหญ่ถูกกำหนดโดยอาศัยความแตกต่างทางด้านสัณฐานวิทยา (จำนวนและชนิดของชั้นในผนังเซลล์) และหน้าที่การทำงาน (ความทนทานในระยะยาวหรือระยะสั้น) ในตอนแรกคิดว่าลักษณะเหล่านี้สามารถแยกแยะ ซีสต์ แบบเพลลิเคิล (ผนังบาง) ออกจาก ซีสต์ แบบพักตัว (ผนังสองชั้น) ของไดโนแฟลเจลเลตได้อย่างชัดเจน ซีสต์แบบแรกถือว่าเป็นซีสต์ระยะสั้น (ชั่วคราว) และซีสต์แบบหลังถือว่าเป็นซีสต์ระยะยาว (พักตัว) อย่างไรก็ตาม ในช่วงสองทศวรรษที่ผ่านมา ความรู้เพิ่มเติมได้เน้นให้เห็นถึงความซับซ้อนอย่างมากของประวัติชีวิตของไดโนแฟลเจลเลต[ 107 ]

มากกว่า 10% ของสายพันธุ์ไดโนแฟลเจลเลตในทะเลที่รู้จักกันประมาณ 2,000 ชนิด สร้างซีสต์เป็นส่วนหนึ่งของวงจรชีวิต (ดูแผนภาพทางด้านขวา) ระยะเบนทิกเหล่านี้มีบทบาทสำคัญในระบบนิเวศของสายพันธุ์ โดยเป็นส่วนหนึ่งของการเชื่อมโยงระหว่างแพลงก์ตอนและเบนทิก ซึ่งซีสต์จะคงอยู่ในชั้นตะกอนในช่วงที่สภาวะไม่เอื้ออำนวยต่อการเจริญเติบโตแบบพืช และจากนั้นก็จะแพร่พันธุ์กลับสู่มวลน้ำเมื่อสภาวะที่เอื้ออำนวยกลับคืนมา[ 107 ]
อันที่จริง ในระหว่างวิวัฒนาการของไดโนแฟลเจลเลต ความจำเป็นในการปรับตัวให้เข้ากับสภาพแวดล้อมที่ผันผวนและ/หรือฤดูกาลนั้น เชื่อกันว่าเป็นแรงผลักดันให้เกิดการพัฒนาของระยะวงจรชีวิตนี้ โปรติสต์ส่วนใหญ่จะสร้างซีสต์ที่อยู่ในสภาวะพักตัวเพื่อทนต่อการอดอาหารและความเสียหายจากรังสียูวี[ 108 ]อย่างไรก็ตาม มีความแตกต่างอย่างมากในคุณสมบัติทางฟีโนไทป์ สรีรวิทยา และความต้านทานหลักของซีสต์ของไดโนแฟลเจลเลตแต่ละชนิด ซึ่งแตกต่างจากในพืชชั้นสูง ความแปรปรวนส่วนใหญ่ เช่น ใน ช่วงเวลา พักตัวยังไม่ได้รับการพิสูจน์ว่าเกิดจากการปรับตัวตามละติจูดหรือขึ้นอยู่กับลักษณะวงจรชีวิตอื่นๆ[ 109 ] [ 110 ]ดังนั้น แม้จะมีความก้าวหน้าล่าสุดในการทำความเข้าใจประวัติชีวิตของไดโนแฟลเจลเลตหลายชนิด รวมถึงบทบาทของระยะซีสต์ แต่ก็ยังมีช่องว่างมากมายในความรู้เกี่ยวกับต้นกำเนิดและหน้าที่การทำงานของพวกมัน[ 107 ]
การรับรู้ถึงความสามารถของไดโนแฟลเจลเลตในการสร้างซีสต์มีมาตั้งแต่ต้นศตวรรษที่ 20 ใน การศึกษา ทางชีวธรณีวิทยาของซีสต์ไดโนแฟลเจลเลตฟอสซิลพอล ไรน์ชเป็นคนแรกที่ระบุซีสต์ว่าเป็นซากดึกดำบรรพ์ของไดโนแฟลเจลเลต[ 111 ]ต่อมา มีการรายงานการก่อตัวของซีสต์จากการรวมตัวของเซลล์สืบพันธุ์ ซึ่งนำไปสู่ข้อสรุปว่าการสร้างซีสต์เกี่ยวข้องกับการสืบพันธุ์แบบอาศัยเพศ[ 104 ]การสังเกตเหล่านี้ยังสนับสนุนแนวคิดที่ว่าการสร้างซีสต์ของไมโครอัลเกเป็นกระบวนการที่ไซโกตเตรียมตัวสำหรับช่วงเวลาพักตัว[ 112 ]เนื่องจากซีสต์พักตัวที่ศึกษาจนถึงเวลานั้นมาจากกระบวนการสืบพันธุ์แบบอาศัยเพศ การพักตัวจึงเกี่ยวข้องกับเพศสัมพันธ์ ซึ่งเป็นข้อสันนิษฐานที่คงอยู่มาหลายปี การระบุลักษณะนี้สอดคล้องกับทฤษฎีวิวัฒนาการเกี่ยวกับต้นกำเนิดของการรวมตัวของเซลล์ยูคาริโอตและการสืบพันธุ์แบบอาศัยเพศ ซึ่งตั้งสมมติฐานถึงข้อดีสำหรับสายพันธุ์ที่มีระยะพักตัวแบบดิพลอยด์ ในความสามารถในการทนต่อความเครียดจากสารอาหารและรังสี UV ที่ทำให้เกิดการกลายพันธุ์ผ่านการซ่อมแซมแบบรีคอมบิเนชัน และสำหรับสายพันธุ์ที่มีระยะเจริญเติบโตแบบแฮพลอยด์ เนื่องจากการแบ่งตัวแบบไม่ใช้เพศจะเพิ่มจำนวนเซลล์เป็นสองเท่า[ 108 ]อย่างไรก็ตาม สภาพแวดล้อมบางอย่างอาจจำกัดข้อดีของรีคอมบิเนชันและการสืบพันธุ์แบบอาศัยเพศ[ 113 ]เช่น ในเชื้อรา การผสมผสานที่ซับซ้อนของวัฏจักรแฮพลอยด์และดิพลอยด์ได้วิวัฒนาการขึ้น ซึ่งรวมถึงระยะพักตัวแบบไม่ใช้เพศและแบบอาศัยเพศ[ 114 ] [ 107 ]
อย่างไรก็ตาม ในวงจรชีวิตทั่วไปของไดโนแฟลเจลเลตที่สร้างซีสต์ตามที่ระบุไว้ในช่วงทศวรรษ 1960 และ 1970 ซีสต์พักตัวถูกสันนิษฐานว่าเป็นชะตากรรมของการสืบพันธุ์แบบอาศัยเพศ[ 104 ] [ 115 ]ซึ่งถือเป็นการตอบสนองต่อความเครียดหรือสภาวะที่ไม่เอื้ออำนวย การสืบพันธุ์แบบอาศัยเพศเกี่ยวข้องกับการรวมตัวของแกมีตแฮพลอยด์จากระยะพืชแพลงก์ตอนที่เคลื่อนที่ได้เพื่อสร้างแพลโนไซโกตดิพลอยด์ซึ่งในที่สุดจะก่อตัวเป็นซีสต์หรือไฮปโนไซโกตซึ่งการงอกของซีสต์เหล่านี้ขึ้นอยู่กับการควบคุมทั้งภายในและภายนอกภายในแล้ว ระยะเวลาการเจริญเติบโตทางสรีรวิทยาขั้นต่ำเฉพาะชนิด (ระยะพักตัว) เป็นสิ่งจำเป็นก่อนที่การงอกจะเกิดขึ้นได้ ดังนั้น ไฮปโนไซโกตจึงถูกเรียกว่าซีสต์ "พักตัว" หรือ "ทนทาน" โดยอ้างอิงถึงลักษณะทางสรีรวิทยานี้และความสามารถของซีสต์เหล่านี้หลังจากระยะพักตัวที่จะยังคงมีชีวิตอยู่ได้ในตะกอนเป็นเวลานาน การงอกจะเกิดขึ้นได้เฉพาะภายในช่วงเวลาของสภาพแวดล้อมที่เหมาะสมเท่านั้น[ 107 ]
อย่างไรก็ตาม การค้นพบว่าแพลโนไซโกตสามารถแบ่งตัวได้นั้น ทำให้เห็นได้ชัดว่าความซับซ้อนของวงจรชีวิตของไดโนแฟลเจลเลตนั้นมากกว่าที่คิดไว้แต่เดิม[ 116 ] [ 117 ]หลังจากการยืนยันพฤติกรรมนี้ในหลายสปีชีส์ ความสามารถของระยะสืบพันธุ์แบบอาศัยเพศของไดโนแฟลเจลเลตในการฟื้นฟูระยะเจริญเติบโตโดยไม่ต้องผ่านการสร้างซีสต์ ก็ได้รับการยอมรับอย่างกว้างขวาง[ 118 ] [ 119 ]นอกจากนี้ ในปี 2549 Kremp และ Parrow ได้แสดงให้เห็นว่าซีสต์พักตัวของไดโนแฟลเจลเลตน้ำเย็นในทะเลบอลติกScrippsiella hangoeiและGymnodinium sp. เกิดจากการสร้างซีสต์โดยตรงของเซลล์เจริญเติบโตแบบแฮพลอยด์ นั่นคือแบบไม่ใช้เพศ[ 120 ]ยิ่งไปกว่านั้น สำหรับซีสต์ไซโกตของPfiesteria piscicidaการพักตัวนั้นไม่จำเป็น[ 121 ] [ 107 ]
จีโนมิกส์
หนึ่งในคุณสมบัติที่โดดเด่นที่สุดของไดโนแฟลเจลเลตคือปริมาณ DNA ในเซลล์จำนวนมากที่พวกมันมีอยู่ สาหร่ายยูคาริโอตส่วนใหญ่มี DNA เฉลี่ยประมาณ 0.54 pg/เซลล์ ในขณะที่การประมาณปริมาณ DNA ของไดโนแฟลเจลเลตมีตั้งแต่ 3–250 pg/เซลล์[ 31 ]ซึ่งสอดคล้องกับประมาณ 3,000–215,000 Mb (เมื่อเปรียบเทียบ จีโนมของมนุษย์แบบแฮพลอยด์มีขนาด 3,180 Mb และ ข้าวสาลี Triticum แบบเฮกซาพลอยด์ มีขนาด 16,000 Mb) ภาวะโพลีพลอยด์หรือโพลีเทนีอาจเป็นสาเหตุของปริมาณ DNA ในเซลล์จำนวนมากนี้[ 122 ]แต่การศึกษาจลนศาสตร์การรวมตัวใหม่ของ DNA ก่อนหน้านี้และการวิเคราะห์จีโนมล่าสุดไม่สนับสนุนสมมติฐานนี้[ 123 ]แต่สมมติฐานนี้ถูกตั้งขึ้นโดยอาศัยการย้อนกลับตำแหน่งอย่างแพร่หลายที่พบในจีโนมของไดโนแฟลเจลเลต[ 124 ] [ 125 ]
นอกจากจีโนมที่มีขนาดใหญ่เกินสัดส่วนแล้ว นิวเคลียสของไดโนแฟลเจลเลตยังมีลักษณะเฉพาะในด้านรูปร่าง การควบคุม และองค์ประกอบ ดีเอ็นเอของพวกมันถูกบรรจุไว้อย่างแน่นหนาจนยังไม่แน่ชัดว่าพวกมันมีโครโมโซมกี่คู่[ 126 ]
ไดโนแฟลเจลเลตมีโครงสร้างจีโนมไมโทคอนเดรียที่ผิดปกติร่วมกับญาติของพวกมันคือ อะ พิคอม เพล็กซา[ 127 ]ทั้งสองกลุ่มมีจีโนมไมโทคอนเดรียที่ลดลงมาก (ประมาณ 6 กิโลเบส (kb) ในอะพิคอมเพล็กซา เทียบกับ ~16kb สำหรับไมโทคอนเดรียของมนุษย์) สายพันธุ์หนึ่งคืออะมีโบฟริอาเซราติได้สูญเสียจีโนมไมโทคอนเดรียไปอย่างสมบูรณ์ แต่ยังคงมีไมโทคอนเดรียที่ทำงานได้[ 128 ]ยีนในจีโนมของไดโนแฟลเจลเลตได้ผ่านการจัดระเบียบใหม่หลายครั้ง รวมถึงการขยายจีโนมและการรวมตัวใหม่จำนวนมาก ซึ่งส่งผลให้มีสำเนาหลายชุดของแต่ละยีนและชิ้นส่วนยีนที่เชื่อมโยงกันในรูปแบบต่างๆ มากมาย การสูญเสียรหัสหยุดมาตรฐาน การต่อเชื่อม mRNA สำหรับ mRNA ของ cox3 และการแก้ไข RNA อย่างกว้างขวางเพื่อเข้ารหัสยีนส่วนใหญ่ได้เกิดขึ้นแล้ว[ 129 ] [ 130 ]สาเหตุของการเปลี่ยนแปลงนี้ยังไม่เป็นที่ทราบแน่ชัด ในกลุ่มไดโนแฟลเจลเลตขนาดเล็กที่เรียกว่า 'ไดโนทอม' (Durinskia และ Kryptoperidinium) เอนโดซิมไบออนต์ (ไดอะตอม) ยังคงมีไมโทคอนเดรีย ทำให้พวกมันเป็นสิ่งมีชีวิตเพียงกลุ่มเดียวที่มีไมโทคอนเดรียสองชนิดที่แตกต่างกันในเชิงวิวัฒนาการ[ 131 ]
ในสายพันธุ์ส่วนใหญ่ จีโนมพลาสติดประกอบด้วยยีนเพียง 14 ยีน[ 132 ]
ดีเอ็นเอของพลาสติดในไดโนแฟลเจลเลตที่มีเพอริดินินนั้นบรรจุอยู่ในวงกลมเล็กๆ ที่เรียกว่ามินิเซอร์เคิล [ 133 ] แต่ละวงกลมประกอบด้วยยีนโพลีเปปไทด์หนึ่งหรือสองยีน ยีนสำหรับโพลีเปปไทด์เหล่านี้เป็นยีนเฉพาะของคลอโรพลาสต์ เนื่องจากโฮโมล็อกของยีนเหล่านี้จากยูคาริโอตสังเคราะห์แสงอื่นๆ จะถูกเข้ารหัสเฉพาะในจีโนมของคลอโรพลาสต์เท่านั้น ภายในแต่ละวงกลมจะมีบริเวณ 'แกนกลาง' ที่สามารถแยกแยะได้ ยีนจะอยู่ในทิศทางเดียวกันเสมอเมื่อเทียบกับบริเวณแกนกลางนี้
ในแง่ของบาร์โค้ดดีเอ็นเอลำดับ ITS สามารถใช้เพื่อระบุชนิดได้[ 134 ]โดยสามารถใช้ระยะทางทางพันธุกรรม p≥0.04 เพื่อกำหนดขอบเขตของชนิดได้[ 135 ]ซึ่งได้ถูกนำมาใช้สำเร็จในการแก้ไขความสับสนทางอนุกรมวิธานที่มีมายาวนาน เช่น ในกรณีของการแบ่งกลุ่ม Alexandrium tamarense ออกเป็นห้าชนิด[ 136 ]การศึกษาล่าสุด[ 137 ]เปิดเผยว่ายีนของไดโนแฟลเจลเลตจำนวนมากเข้ารหัสสำหรับหน้าที่ที่ไม่รู้จัก และยีนเหล่านี้อาจได้รับการอนุรักษ์และจำเพาะต่อสายพันธุ์
ประวัติวิวัฒนาการ
ไดโนแฟลเจลเลตส่วนใหญ่พบในรูปฟอสซิลไดโนซิสต์ซึ่งมีบันทึกทางธรณีวิทยายาวนาน โดยพบน้อยที่สุดในช่วงกลางยุคไทรแอสสิก [ 138 ]ในขณะที่ตัวบ่งชี้ทางเคมีบ่งชี้ว่ามีอยู่ตั้งแต่ต้นยุคแคมเบรียน[ 139 ]หลักฐานบางอย่างบ่งชี้ว่าไดโนสเตอรอยด์ใน หิน ยุคพาลีโอโซอิกและพรีแคมเบรียน จำนวนมาก อาจเป็นผลผลิตของไดโนแฟลเจลเลตบรรพบุรุษ (โปรโตไดโนแฟลเจลเลต) [ 140 ] [ 141 ] ไดโนแฟลเจลเลตแสดงให้เห็นถึงการแพร่กระจายของรูปร่างอย่างคลาสสิกในช่วงปลายยุคไทรแอสสิกจนถึงกลางยุคจูรา สสิก [ 142 ] [ 143 ] [ 144 ]รูปแบบที่ดูทันสมัยมากขึ้นแพร่หลายในช่วงปลายยุคจูราสสิกและยุคครีเทเชียส [ 142 ] แนวโน้มนี้ยังคงดำเนินต่อไปในยุคซีโนโซอิกแม้ว่าจะมีการสูญเสียความหลากหลายไปบ้าง[ 142 ] [ 138 ]
การวิเคราะห์วิวัฒนาการระดับโมเลกุลแสดงให้เห็นว่าไดโนแฟลเจลเลตถูกจัดกลุ่มร่วมกับซีลิเอตและอะพิคอมเพล็กซาน (=สปอโรซัว) ในกลุ่มที่มีความสัมพันธ์กันอย่างดี คือ อัลวี โอเลตญาติที่ใกล้ชิดที่สุดของไดโนแฟลเจลเลตไดโนคาริโอตดูเหมือนจะเป็นอะพิคอม เพล็ กซาน เพอร์กินซัส พาร์วิลู ซิเฟรา ซิน ดิเนียน และออกซีร์ริส[ 145 ]การวิเคราะห์วิวัฒนาการระดับโมเลกุลมีความคล้ายคลึงกับการวิเคราะห์วิวัฒนาการตามสัณฐานวิทยา[ 146 ] [ 147 ]
ระยะแรกสุดของการวิวัฒนาการของไดโนแฟลเจลเลตดูเหมือนจะถูกครอบงำโดยสายพันธุ์ปรสิต เช่น เพอร์กินซิดและซินดิเนียน (เช่นอะมีโบฟริอาและเฮมาโทดิเนียม ) [ 148 ] [ 149 ] [ 150 ] [ 151 ]
ไดโนแฟลเจลเลตทั้งหมดมีพลาสติดของสาหร่ายสีแดงหรือออร์แกเนลล์ที่เหลืออยู่ (ที่ไม่สังเคราะห์แสง) จากต้นกำเนิดของสาหร่ายสีแดง[ 152 ] อย่างไรก็ตาม ไดโนแฟลเจลเลตปรสิตHematodiniumไม่มีพลาสติดเลย[ 153 ]บางกลุ่มที่สูญเสียคุณสมบัติการสังเคราะห์แสงของพลาสติดสาหร่ายสีแดงดั้งเดิมไป ได้รับพลาสติดสังเคราะห์แสงใหม่ (คลอโรพลาสต์) ผ่านกระบวนการที่เรียกว่าเอนโดซิมไบโอซิสแบบอนุกรม ทั้งแบบทุติยภูมิและตติยภูมิ:
- Lepidodiniumมีพลาสติดที่ได้มาจากสาหร่ายสีเขียวอย่างผิดปกติ (พลาสติดที่ได้มาตามลำดับอื่นๆ ทั้งหมดสามารถสืบย้อนกลับไปถึงสาหร่ายสีแดงได้) [ 154 ] พลาสติดนี้มีความเกี่ยวข้องมากที่สุดกับ Pedinomonasที่ดำรงชีวิตอิสระ(ดังนั้นจึงน่าจะเป็นพลาสติดรอง) ไดโนแฟลเจลเลตสองชนิดที่ไม่เคยมีการอธิบายมาก่อน ("MGD" และ "TGD") มีพลาสติดที่เกี่ยวข้องอย่างใกล้ชิด [ 155 ]
- Karenia , KarlodiniumและTakayamaมีพลาสติดที่มีต้นกำเนิดจากแฮปโทไฟต์ซึ่งผลิตขึ้นในสามเหตุการณ์ที่แยกจากกัน[ 156 ]
- "ไดโนทอม" ( ดูรินสเกียและคริปโทเพอริดิเนียม ) มีพลาสติดที่ได้มาจากไดอะตอม[ 157 ] [ 158 ]
สัตว์บางชนิดยังทำการผ่าตัดลอกคราบ ด้วย :
- ไดโนฟิซิสมีพลาสติดจากคริปโตโมนาดเนื่องจากการขโมยพลาสติดจากเหยื่อซิลิตา [ 159 ]
- Kareniaceae (ซึ่งประกอบด้วยสกุลที่มี haptophyte สามสกุล) มีกรณี kleptoplasty แยกกันสองกรณี[ 160 ] [ 156 ]
วิวัฒนาการของไดโนแฟลเจลเลตได้รับการสรุปเป็นประเภทการจัดระเบียบหลัก 5 ประเภท ได้แก่ โปรโรเซนทรอยด์ ไดโนฟิซอยด์ กอนยาอูลาคอยด์ เพอริดินิออยด์ และจิมโนดินอยด์[ 161 ] การเปลี่ยนแปลงของสายพันธุ์ในทะเลไปสู่น้ำจืดเป็นเหตุการณ์ที่เกิดขึ้นบ่อยครั้งในระหว่างการเพิ่มความหลากหลายของไดโนแฟลเจลเลตและเพิ่งเกิดขึ้นเมื่อไม่นานมานี้[ 162 ]
ไดโนแฟลเจลเลตหลายชนิดยังมีความสัมพันธ์แบบพึ่งพาอาศัยกันกับไซยาโนแบคทีเรีย เรียกว่า ไซยาโนไบออนต์ ซึ่งมีจีโนมที่ลดลงและไม่พบอยู่นอกโฮสต์ของพวกมัน ไดโนแฟลเจลเลตกลุ่มไดโนฟิซอยด์มีสองสกุล ได้แก่ แอมฟิโซเลเนียและไตรโพโซเลเนีย ที่มีไซยาโนไบออนต์ภายในเซลล์ และสี่สกุล ได้แก่ ซิธาริสเตส ฮิสติโอเนียส พาราฮิสติโอเนียส และออร์นิโทเซอร์คัส ที่มีไซยาโนไบออนต์ภายนอกเซลล์[ 163 ] ไซยาโนไบ ออนต์ส่วนใหญ่ใช้สำหรับการตรึงไนโตรเจน ไม่ใช่สำหรับการสังเคราะห์แสง แต่บางชนิดก็ไม่มีความสามารถในการตรึงไนโตรเจน ไดโนแฟลเจลเลต ออร์นิโทเซอร์คัส แมกนิฟิคัสเป็นโฮสต์ของไซยาโนไบออนต์ซึ่งอาศัยอยู่ในห้องภายนอกเซลล์ แม้ว่าจะยังไม่ทราบแน่ชัดว่าไดโนแฟลเจลเลตได้รับประโยชน์จากสิ่งนี้อย่างไร แต่ก็มีการเสนอแนะว่ามันเพาะเลี้ยงไซยาโนแบคทีเรียในห้องพิเศษและย่อยสลายไซยาโนแบคทีเรียบางส่วนเป็นประจำ[ 164 ]
เมื่อเร็วๆ นี้ฟอสซิลมีชีวิตDapsilidinium pastielsiiถูกพบว่าอาศัยอยู่ในIndo-Pacific Warm Poolซึ่งทำหน้าที่เป็นแหล่งหลบภัยสำหรับไดโนแฟลเจลเลตที่ชอบอุณหภูมิสูง[ 165 ]และสิ่งมีชีวิตอื่นๆ เช่นCalciodinellum operosumและPosoniella tricarinelloidesก็ได้รับการอธิบายจากฟอสซิลก่อนที่จะถูกพบว่ามีชีวิตในภายหลัง[ 166 ] [ 167 ]
ตัวอย่าง


 ซีเรียมมาโครเซรอส (Dinophyceae)
ซีเรียมมาโครเซรอส (Dinophyceae) ซีเรียมเฟอร์คอยด์ (Dinophyceae)
ซีเรียมเฟอร์คอยด์ (Dinophyceae) ไดโนแฟลเจลเลตชนิดที่ไม่ทราบชนิดภายใต้กล้องจุลทรรศน์อิเล็กตรอนแบบสแกน (Dinophyceae)
ไดโนแฟลเจลเลตชนิดที่ไม่ทราบชนิดภายใต้กล้องจุลทรรศน์อิเล็กตรอนแบบสแกน (Dinophyceae) Pfiesteria shumwayae (Dinophyceae)
Pfiesteria shumwayae (Dinophyceae) Symbiodinium sp. (Dinophyceae):ซูแซนเทลลา จุลินทรีย์ร่วมอาศัยภายในปะการัง
Symbiodinium sp. (Dinophyceae):ซูแซนเทลลา จุลินทรีย์ร่วมอาศัยภายในปะการัง








ดูเพิ่มเติม
บรรณานุกรม
ลิงก์ภายนอก
- สมาคมระหว่างประเทศเพื่อการศึกษาเกี่ยวกับสาหร่ายที่เป็นอันตราย
- เอกสารทางวิชาการเกี่ยวกับไดโนแฟลเจลเลตฉบับคลาสสิกเก็บถาวรเมื่อวันที่ 16 กรกฎาคม 2012 ที่Wayback Machine
- แหล่งข้อมูลไดโนแฟลเจลเลตของญี่ปุ่นเก็บถาวรเมื่อวันที่ 12 พฤษภาคม 2013 ที่Wayback Machine
- Noctiluca scintillans —คู่มือแพลงก์ตอนสัตว์ทะเลแห่งออสเตรเลียตะวันออกเฉียงใต้สถาบันการเพาะเลี้ยงสัตว์น้ำและการประมงแทสเมเนียน
- ต้นไม้แห่งชีวิต ไดโนแฟลเจลเลตเก็บถาวรเมื่อ 13 ตุลาคม 2012 ที่Wayback Machine
- ศูนย์ความเป็นเลิศด้านการจำแนกประเภทไดโนไฟต์ (CEDiT)
- ไดโนแฟลเจลเลตถูกเก็บถาวรเมื่อวันที่ 13 ตุลาคม 2012 ที่Wayback Machine
- Judson O (5 มกราคม 2010). "เรื่องราวของแฟลเจลลาสองอัน" . เดอะนิวยอร์กไทมส์ .
- Tomas CK (1997), "บทนำ", การระบุแพลงก์ตอนพืชในทะเล , Elsevier, หน้า 585–589 , doi : 10.1016/b978-012693018-4/50006-9 , ISBN 978-0-12-693018-4
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ไดโนแฟลเจลเลต
ไดโนแฟลเจลเลต (จากภาษากรีกโบราณδῖνος ( dînos ) ' หมุนวน'และภาษาละตินflagellum ' แส้, แส้' ) หรือที่เรียกว่าไดโนไฟต์เป็น กลุ่ม โมโนฟิเลติก ของ...
นิรุกติศาสตร์
คำว่า "ไดโนแฟลเจลเลต" มาจากการรวมกันของคำว่า dinos ในภาษากรีก และ flagellum ในภาษา ละติน Dinos หมายถึง "การหมุนวน" และหมายถึงลักษณะเฉพาะของการว่ายน้ำของไดโนแฟลเจลเลตที่ถูกสังเกต Flagellum หมายถึง "แส้" และหมายถึง แฟลเจลลา ของ พวกมัน [ 12 ]
ประวัติศาสตร์
ในปี ค.ศ. 1753 ไดโนแฟลเจลเลตสมัยใหม่ชนิดแรก ( Noctiluca แม้ว่าจะยังไม่ได้รับการตั้งชื่ออย่างเป็นทางการจนกระทั่งปี ค.ศ.
สัณฐานวิทยา
ไดโนแฟลเจลเลตเป็นสิ่งมีชีวิตเซลล์เดียวและมีแฟลเจลลาสองแบบที่ไม่เหมือนกันซึ่งเกิดขึ้นจากด้านท้องของเซลล์ (การสร้างแฟลเจลลาแบบไดโนคอนต์) พวกมันมีแฟลเจลลัมตามขวางที่มีลักษณะคล้ายริบบิ้นที่มีหลายคลื่นซึ่งโบกไปทางซ้ายของเซลล์...