อ่าน 14 นาที
ภูมิคุ้มกันตนเอง
ในทาง ภูมิคุ้มกันวิทยา ภาวะ ภูมิคุ้มกันทำลายตนเอง คือระบบ การตอบสนองทางภูมิคุ้มกัน ของสิ่งมีชีวิต ต่อ เซลล์ เนื้อเยื่อ และส่วนประกอบอื่นๆ ของร่างกายที่แข็งแรงของตนเอง [ 1 ] [ 2 ]...
ภูมิคุ้มกันตนเอง
| ภูมิคุ้มกันตนเอง | |
|---|---|
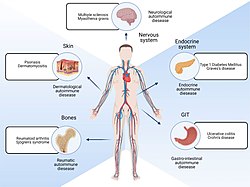 | |
| ส่วนต่างๆ ของร่างกายที่ได้รับผลกระทบจากโรคภูมิต้านทานตนเอง | |
| ความเชี่ยวชาญ | ภูมิคุ้มกันวิทยา |
ในทางภูมิคุ้มกันวิทยาภาวะภูมิคุ้มกันทำลายตนเองคือระบบการตอบสนองทางภูมิคุ้มกัน ของสิ่งมีชีวิต ต่อเซลล์เนื้อเยื่อและส่วนประกอบอื่นๆ ของร่างกายที่แข็งแรงของตนเอง[ 1 ] [ 2 ]โรคใดๆ ที่เกิดจากการตอบสนองทางภูมิคุ้มกันประเภทนี้เรียกว่า " โรคภูมิคุ้มกันทำลายตนเอง " ตัวอย่างที่สำคัญ ได้แก่โรคเซลิแอคโรคเบาหวานชนิดที่ 1โรคเฮนอค-เชินไลน์ เพอร์พูรา โรคซิสเต็มิก ลูปัส อี ริธีมาโตซั ส กลุ่มอาการ โจเกรน โรคอีโอซิโนฟิลิก แกรนูโลมาโตซิส ร่วมกับหลอดเลือดอักเสบ โรค ต่อมไทรอยด์อักเสบฮาชิโมโตะโรคเกรฟส์ โรคเกล็ดเลือดต่ำชนิด ไม่ ทราบ สาเหตุ โรคแอดดิสัน โรคข้ออักเสบรูมาต อยด์ โรคกระดูกสันหลังอักเสบ โรคกล้ามเนื้ออักเสบโรคผิวหนังอักเสบและโรคปลอกประสาทเสื่อมแข็งโรคภูมิคุ้มกันทำลายตนเองมักได้รับการรักษาด้วยสเตียรอยด์[ 3 ]
ภาวะภูมิคุ้มกันตนเองหมายถึงการมีแอนติบอดีหรือเซลล์ Tที่ทำปฏิกิริยากับโปรตีนของตนเองและมีอยู่ในบุคคลทุกคน แม้กระทั่งในภาวะสุขภาพปกติก็ตาม หากปฏิกิริยาต่อตนเองสามารถนำไปสู่ความเสียหายของเนื้อเยื่อได้ ก็จะทำให้เกิดโรคภูมิคุ้มกันตนเองขึ้น[ 4 ]
ประวัติศาสตร์
ในช่วงปลายศตวรรษที่ 19 มีความเชื่อว่าระบบภูมิคุ้มกันไม่สามารถตอบโต้เนื้อเยื่อของร่างกายเองได้พอล เออร์ลิชในช่วงต้นศตวรรษที่ 20 ได้เสนอแนวคิดเรื่อง " ความสยองขวัญจากปฏิกิริยาต่อต้านตนเอง" ( horror autotoxicus ) ต่อมาเออร์ลิชได้ปรับทฤษฎีของเขาเพื่อยอมรับความเป็นไปได้ของการโจมตีเนื้อเยื่อโดยระบบภูมิคุ้มกันต่อต้านตนเอง แต่เชื่อว่ากลไกการป้องกันโดยธรรมชาติบางอย่างจะป้องกันไม่ให้ปฏิกิริยาต่อต้านตนเองนั้นกลายเป็นพยาธิสภาพ
ในปี พ.ศ. 2447 ทฤษฎีนี้ถูกท้าทายด้วยการค้นพบสารในซีรั่มของผู้ป่วยที่เป็นโรคฮีโมโกลบินูเรียจากความเย็นแบบเฉียบพลันซึ่งทำปฏิกิริยากับเซลล์เม็ดเลือดแดง ในช่วงหลายทศวรรษต่อมา สภาวะต่างๆ จำนวนมากสามารถเชื่อมโยงกับการตอบสนองของระบบภูมิคุ้มกันอัตโนมัติได้ อย่างไรก็ตาม สถานะที่ได้รับการยอมรับของสมมติฐานของเออร์ลิชกลับเป็นอุปสรรคต่อความเข้าใจในผลการค้นพบเหล่านี้ ภูมิคุ้มกันวิทยาจึงกลายเป็นสาขาชีวเคมีมากกว่าสาขาทางคลินิก[ 5 ]ในช่วงทศวรรษ พ.ศ. 2493 ความเข้าใจสมัยใหม่เกี่ยวกับออโตแอนติบอดีและโรคภูมิคุ้มกันอัตโนมัติเริ่มแพร่หลาย[ 6 ]
เมื่อไม่นานมานี้ เป็นที่ยอมรับกันว่าการตอบสนองของภูมิคุ้มกันตนเองเป็นส่วนสำคัญของระบบภูมิคุ้มกัน ของสัตว์มีกระดูกสันหลัง (บางครั้งเรียกว่า "ภูมิคุ้มกันตนเองตามธรรมชาติ") [ 7 ]ภูมิคุ้มกันตนเองไม่ควรสับสนกับภูมิคุ้มกันต่างชนิด
ภาวะภูมิคุ้มกันบกพร่องระดับต่ำ
ภาวะภูมิคุ้มกันต่อต้านตนเองอาจมีบทบาทในการทำให้เกิดการตอบสนองทางภูมิคุ้มกันอย่างรวดเร็วในระยะเริ่มต้นของการติดเชื้อ เมื่อปริมาณของแอนติเจน แปลกปลอม มีจำกัด (เช่น เมื่อมีเชื้อโรคอยู่น้อย) ในการศึกษาของ Stefanova et al. (2002) พวกเขาฉีดแอนติบอดี ต่อต้าน MHC class II เข้าไปในหนูที่แสดงโมเลกุล MHC class II ชนิดเดียว (H-2 b ) เพื่อป้องกันการปฏิสัมพันธ์ระหว่างเซลล์ CD4+ T กับ MHC ชั่วคราวเซลล์ CD4+ T ที่ยังไม่ได้รับการกระตุ้น (เซลล์ที่ไม่เคยพบกับแอนติเจนที่ไม่ใช่ของตนเองมาก่อน) ที่เก็บได้จากหนูเหล่านี้ 36 ชั่วโมงหลังจากการให้แอนติบอดีต่อต้าน MHC แสดงให้เห็นถึงการตอบสนองต่อแอนติเจน pigeon cytochrome c peptide ที่ลดลง ซึ่งกำหนดโดยการฟอสโฟรีเลชัน ของ ZAP70การเพิ่มจำนวน และ การผลิต อินเตอร์ลิวคิน 2ดังนั้น Stefanova et al. (2002) แสดงให้เห็นว่าการรับรู้ MHC ของตนเอง (ซึ่งหากรุนแรงเกินไปอาจส่งผลให้เกิดโรคภูมิคุ้มกันตนเอง) ช่วยรักษาการตอบสนองของเซลล์ T CD4+ เมื่อไม่มีแอนติเจนจากภายนอก[ 8 ]
ความทนทานทางภูมิคุ้มกัน
งานบุกเบิกโดยNoel RoseและErnst Witebskyในนิวยอร์ก และRoittและDoniachที่University College Londonได้ให้หลักฐานที่ชัดเจนว่า อย่างน้อยในแง่ของเซลล์ B ที่สร้างแอนติบอดี (ลิมโฟไซต์ B) โรคต่างๆ เช่น โรคข้ออักเสบรูมาตอยด์และภาวะไทรอยด์เป็นพิษนั้นเกี่ยวข้องกับการสูญเสียความทนทานทางภูมิคุ้มกันซึ่งเป็นความสามารถของแต่ละบุคคลที่จะเพิกเฉยต่อ "ตนเอง" ในขณะที่ตอบสนองต่อ "สิ่งที่ไม่ใช่ตนเอง" การแตกหักนี้ทำให้ระบบภูมิคุ้มกันสร้างการตอบสนองทางภูมิคุ้มกันที่มีประสิทธิภาพและเฉพาะเจาะจงต่อแอนติเจนของตนเอง ต้นกำเนิดที่แท้จริงของความทนทานทางภูมิคุ้มกันยังคงคลุมเครือ แต่มีทฤษฎีหลายทฤษฎีที่ได้รับการเสนอตั้งแต่กลางศตวรรษที่ 20 เพื่ออธิบายที่มาของมัน[ 9 ]
มีสมมติฐานสามข้อที่ได้รับความสนใจอย่างกว้างขวางในหมู่นักภูมิคุ้มกันวิทยา:
- ทฤษฎีการกำจัดโคลน (Clonal deletion theory ) ที่เสนอโดยเบอร์เน็ต (Burnet ) ระบุว่าเซลล์น้ำเหลืองที่ตอบสนองต่อตนเองจะถูกทำลายไปในระหว่างการพัฒนาระบบภูมิคุ้มกันในแต่ละบุคคล จากผลงานของพวกเขา แฟรงค์ เอ็ม. เบอร์เน็ต และปีเตอร์ บี. เมดาวาร์ ได้รับรางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ประจำปี 1960 "สำหรับการค้นพบความทนทานทางภูมิคุ้มกันที่ได้มา" (acquired immunological tolerance)
- ทฤษฎีแอนเนอร์จีแบบโคลนัลที่เสนอโดยNossalซึ่งเซลล์ T หรือ B ที่ตอบสนองต่อตนเองจะถูกทำให้ไม่ทำงานในบุคคลปกติและไม่สามารถขยายการตอบสนองทางภูมิคุ้มกันได้[ 10 ]
- ทฤษฎีเครือข่ายไอดีโอไทป์ที่เสนอโดยJerneซึ่งเครือข่ายของแอนติบอดีที่สามารถทำให้แอนติบอดีที่ทำปฏิกิริยากับตัวเองเป็นกลางได้นั้นมีอยู่ตามธรรมชาติในร่างกาย[ 11 ]
นอกจากนี้ ยังมีทฤษฎีอีกสองทฤษฎีที่อยู่ระหว่างการตรวจสอบอย่างเข้มข้น:
- ทฤษฎีความไม่รู้ของโคลนซึ่งระบุว่าเซลล์ T ที่มีปฏิกิริยาต่อตนเองซึ่งไม่มีอยู่ในต่อมไทมัสจะเจริญเติบโตและอพยพไปยังบริเวณรอบนอก ซึ่งเซลล์เหล่านี้จะไม่พบแอนติเจนที่เหมาะสมเนื่องจากเป็นเนื้อเยื่อที่ไม่สามารถเข้าถึงได้ ส่งผลให้เซลล์ B ที่มีปฏิกิริยาต่อตนเองซึ่งรอดพ้นจากการกำจัดไม่สามารถหาแอนติเจนหรือเซลล์ T ช่วยเหลือที่เฉพาะเจาะจงได้[ 12 ]
- ทฤษฎีประชากรเซลล์ยับยั้งหรือเซลล์ทีควบคุม (Regulatory T cell theory ) ซึ่งเซลล์ทีลิมโฟไซต์ควบคุม (โดยทั่วไปคือเซลล์ CD4 + FoxP3 +และอื่นๆ) ทำหน้าที่ป้องกัน ลดระดับ หรือจำกัดการตอบสนองทางภูมิคุ้มกันที่ทำลายตัวเองในระบบภูมิคุ้มกัน
ภาวะทนทานต่อสิ่งแปลกปลอมยังสามารถแบ่งออกเป็น "ภาวะทนทานส่วนกลาง" และ "ภาวะทนทานส่วนปลาย" โดยพิจารณาจากว่ากลไกการตรวจสอบที่กล่าวมาข้างต้นทำงานในอวัยวะน้ำเหลืองส่วนกลาง (ต่อมไทมัสและไขกระดูก) หรืออวัยวะน้ำเหลืองส่วนปลาย (ต่อมน้ำเหลือง ม้าม ฯลฯ ซึ่งเซลล์บีที่ตอบสนองต่อสิ่งแปลกปลอมอาจถูกทำลาย) ต้องเน้นย้ำว่าทฤษฎีเหล่านี้ไม่ได้ขัดแย้งกัน และมีหลักฐานเพิ่มมากขึ้นที่บ่งชี้ว่ากลไกทั้งหมดเหล่านี้อาจมีส่วนร่วมอย่างแข็งขันต่อภาวะทนทานต่อสิ่งแปลกปลอมในสัตว์มีกระดูกสันหลัง
ลักษณะที่น่าสงสัยอย่างหนึ่งของการสูญเสียความทนทานที่พบในการเกิดโรคภูมิต้านตนเองในมนุษย์โดยธรรมชาติ คือ การสูญเสียความทนทานนั้นจำกัดอยู่เกือบทั้งหมดเฉพาะการตอบสนองของแอนติบอดีที่สร้างโดยลิมโฟไซต์บี การสูญเสียความทนทานโดยเซลล์ทีนั้นยากที่จะพิสูจน์ได้ และหากมีหลักฐานของการตอบสนองของเซลล์ทีที่ผิดปกติ มักจะไม่ใช่ต่อแอนติเจนที่แอนติบอดีจดจำได้ ดังนั้น ในโรคข้ออักเสบรูมาตอยด์ มีแอนติบอดีต่อ IgG Fc แต่ดูเหมือนจะไม่มีการตอบสนองของเซลล์ทีที่สอดคล้องกัน ในโรคแพ้ภูมิตัวเองชนิดลูปัส มีแอนติบอดีต่อ DNA ซึ่งไม่สามารถกระตุ้นการตอบสนองของเซลล์ทีได้ และหลักฐานที่จำกัดเกี่ยวกับการตอบสนองของเซลล์ทีนั้นชี้ไปที่แอนติเจนนิวคลีโอโปรตีน ในโรคเซลิแอค มีแอนติบอดีต่อทิชชูทรานส์กลูตามิเนส แต่การตอบสนองของเซลล์ทีนั้นกลับเป็นต่อโปรตีนแปลกปลอมอย่างกลูติน ความแตกต่างนี้ทำให้เกิดแนวคิดที่ว่าโรคภูมิคุ้มกันตนเองของมนุษย์ส่วนใหญ่ (โดยอาจมีข้อยกเว้น เช่น โรคเบาหวานประเภทที่ 1) เกิดจากการสูญเสียความทนทานของเซลล์ B ซึ่งใช้การตอบสนองของเซลล์ T ปกติต่อแอนติเจนแปลกปลอมในรูปแบบที่ผิดปกติต่างๆ[ 13 ]
ภาวะภูมิคุ้มกันบกพร่องและโรคภูมิต้านตนเอง
มีกลุ่มอาการภูมิคุ้มกันบกพร่องจำนวนมากที่แสดงลักษณะทางคลินิกและทางห้องปฏิบัติการของภาวะภูมิคุ้มกันทำลายตนเอง ความสามารถที่ลดลงของระบบภูมิคุ้มกันในการกำจัดการติดเชื้อในผู้ป่วยเหล่านี้อาจเป็นสาเหตุของการเกิดภาวะภูมิคุ้มกันทำลายตนเองผ่านการกระตุ้นระบบภูมิคุ้มกันอย่างต่อเนื่อง[ 14 ]
ตัวอย่างหนึ่งคือภาวะภูมิคุ้มกันบกพร่องแบบแปรผันทั่วไปซึ่งพบโรคภูมิต้านตนเองหลายชนิด เช่น โรคลำไส้อักเสบ โรคเกล็ดเลือดต่ำจากภูมิต้านตนเอง และโรคไทรอยด์จากภูมิต้านตนเอง[ 15 ]
โรคฮีโมฟาโก ไซติก ลิมโฟฮิสติโอซิสชนิดถ่ายทอดทางพันธุกรรม ( Familial hemophagocytic lymphohistiocytosis ) ซึ่งเป็นภาวะภูมิคุ้มกันบกพร่อง ปฐมภูมิแบบถ่ายทอดทางพันธุกรรมแบบยีนด้อย เป็นอีกตัวอย่างหนึ่ง ผู้ป่วยมักมีอาการเม็ดเลือดต่ำทุกชนิด ผื่นขึ้น ต่อมน้ำเหลืองบวม และตับและม้ามโตเชื่อว่าสาเหตุมาจากภาวะติดเชื้อไวรัสหลายชนิดที่ไม่ได้รับการรักษา เนื่องจากขาดโปรตีนเพอร์ฟอริน
นอกจากภาวะติดเชื้อเรื้อรังและ/หรือเกิดซ้ำแล้ว โรคภูมิต้านตนเองหลายชนิด เช่น โรคข้ออักเสบ โรคโลหิตจางจากภูมิคุ้มกันทำลายเม็ดเลือดแดง โรคหนังแข็ง และโรคเบาหวานชนิดที่ 1ก็พบได้ใน ผู้ป่วยโรค อะแกมมาโกลบูลินีเมียที่ถ่ายทอดทางโครโมโซม X (XLA) เช่นกัน การติดเชื้อแบคทีเรียและเชื้อราซ้ำๆ และการอักเสบเรื้อรังของลำไส้และปอดก็พบได้ในโรคแกรนูโลมาเรื้อรัง (CGD) ด้วย CGD เกิดจากการผลิตเอนไซม์นิโคตินาไมด์อะดีนีนไดนิวคลีโอไทด์ฟอสเฟต (NADPH) ออกซิเดสลดลงโดยนิวโทรฟิล การกลายพันธุ์แบบไฮโปมอร์ฟิกของยีน RAG พบได้ในผู้ป่วยโรคแกรนูโลมาบริเวณกึ่งกลางลำตัว ซึ่งเป็นโรคภูมิต้านตนเองที่พบได้บ่อยในผู้ป่วยโรคแกรนูโลมาโตซิสร่วมกับหลอดเลือดอักเสบและมะเร็งต่อมน้ำเหลืองชนิด NK/T เซลล์ ผู้ป่วยโรค วิสคอตต์-อัลดริช (WAS) ก็มีอาการผื่นผิวหนังอักเสบ อาการทางภูมิต้านตนเอง การติดเชื้อแบคทีเรียซ้ำๆ และมะเร็งต่อมน้ำเหลืองด้วย ในโรค autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophyภาวะภูมิคุ้มกันบกพร่องและการติดเชื้อเกิดขึ้นพร้อมกัน ได้แก่ การแสดงออกของภูมิคุ้มกันบกพร่องเฉพาะอวัยวะ (เช่น ภาวะ hypoparathyroidism และภาวะ adrenocortical failure) และ candidiasis เรื้อรังที่เยื่อบุผิวและเยื่อเมือก สุดท้ายภาวะขาด IgAบางครั้งก็เกี่ยวข้องกับการพัฒนาของปรากฏการณ์ภูมิคุ้มกันบกพร่องและภูมิแพ้[ 16 ]
สาเหตุ
ปัจจัยทางพันธุกรรม
บุคคลบางคนมีความเสี่ยงทางพันธุกรรมต่อการเกิดโรคภูมิคุ้มกันบกพร่อง ความเสี่ยงนี้เกี่ยวข้องกับยีนหลายตัวรวมถึงปัจจัยเสี่ยงอื่นๆ บุคคลที่มีความเสี่ยงทางพันธุกรรมไม่ได้เป็นโรคภูมิคุ้มกันบกพร่องเสมอไป ยีนหลักสามชุดถูกสงสัยว่าเกี่ยวข้องกับโรคภูมิคุ้มกันบกพร่องหลายชนิด ยีนเหล่านี้เกี่ยวข้องกับ: [ 17 ]
- อิมมูโนโกลบูลิน
- ตัวรับทีเซลล์
- คอมเพล็กซ์ฮิสโตคอมแพติบิลิตีหลัก (MHC)
สองส่วนแรกซึ่งเกี่ยวข้องกับการจดจำแอนติเจนนั้นมีความแปรปรวนและไวต่อการรวมตัวกันใหม่โดยธรรมชาติ ความแปรปรวนเหล่านี้ทำให้ระบบภูมิคุ้มกันสามารถตอบสนองต่อผู้บุกรุกได้หลากหลายชนิดมาก แต่ก็อาจทำให้เกิดลิมโฟไซต์ที่สามารถตอบสนองต่อตนเองได้ เช่นกัน
- HLA DR2 มีความสัมพันธ์เชิงบวกอย่างมากกับโรคแพ้ภูมิตัวเองชนิดลูปัสโรคนอนหลับผิดปกติ[ 18 ]และโรคปลอกประสาทเสื่อมแข็งและมีความสัมพันธ์เชิงลบกับโรคเบาหวานชนิดที่ 1
- HLA DR3 มีความสัมพันธ์อย่างมากกับโรค Sjögren syndrome , myasthenia gravis , lupus erythematosusและโรคเบาหวานชนิดที่ 1
- HLA DR4 มีความสัมพันธ์กับการเกิดโรคข้ออักเสบรูมาตอยด์โรคเบาหวานชนิดที่ 1 และโรคเพมฟิกัส วัลการิส
ความสัมพันธ์กับโมเลกุล MHC คลาส I นั้นมีน้อยกว่า ความสัมพันธ์ที่โดดเด่นและสม่ำเสมอที่สุดคือความสัมพันธ์ระหว่าง HLA B27 กับโรคข้ออักเสบกระดูกสันหลัง เช่นโรคกระดูกสันหลังอักเสบเรื้อรังและ โรค ข้อ อักเสบปฏิกิริยา ความสัมพันธ์อาจมีอยู่ระหว่างโพลีมอร์ฟิซึมภายในโปรโมเตอร์ MHC คลาส II กับโรคภูมิต้านตนเอง
บทบาทของยีนที่อยู่นอกเหนือกลุ่ม MHC ยังคงเป็นหัวข้อของการวิจัย ทั้งในแบบจำลองโรคในสัตว์ (เช่น การศึกษาทางพันธุกรรมอย่างละเอียดของ Linda Wicker เกี่ยวกับโรคเบาหวานในหนู NOD) และในผู้ป่วย (เช่น การวิเคราะห์ความเชื่อมโยงของ Brian Kotzin เกี่ยวกับความเสี่ยงต่อโรคแพ้ภูมิตัวเองชนิดลูปัส )
จากการศึกษาล่าสุด พบว่ายีนPTPN22เป็นปัจจัยสำคัญที่เชื่อมโยงกับโรคภูมิต้านตนเองหลายชนิด เช่น โรคเบาหวานชนิดที่ 1 โรคข้ออักเสบรูมาตอยด์ โรคลูปัสทั่วร่างกาย โรคต่อมไทรอยด์อักเสบฮาชิโมโตะ โรคเกรฟส์ โรคแอดดิสัน โรคกล้ามเนื้ออ่อนแรง โรคด่างขาว โรคหนังแข็งทั่วร่างกาย โรคข้ออักเสบไม่ทราบสาเหตุในเด็ก และโรคข้ออักเสบสะเก็ดเงิน[ 19 ] PTPN22 มีส่วนเกี่ยวข้องในการควบคุมการทำงานของเซลล์ภูมิคุ้มกัน ดังนั้นการเปลี่ยนแปลงในยีนนี้จึงอาจนำไปสู่การทำงานที่ผิดปกติของระบบภูมิคุ้มกัน ทำให้บุคคลมีความเสี่ยงต่อโรคภูมิต้านตนเองมากขึ้น[ 20 ] [ 21 ]
ปัจจัยเชิงอัตถิภาวะ (หรือเรียกอีกอย่างว่า ปัจจัยสิ่งแวดล้อมภายใน)
| อัตราส่วนการเกิด โรคภูมิต้านตนเอง ในเพศหญิงต่อเพศชาย | |
|---|---|
| โรคต่อมไทรอยด์อักเสบฮาชิโมโตะ | 10:1 [ 22 ] |
| โรคเกรฟส์ | 7:1 [ 22 ] |
| โรคปลอกประสาทเสื่อมแข็ง (MS) | 2:1 [ 22 ] |
| โรคกล้ามเนื้ออ่อนแรง | 2:1 [ 22 ] |
| โรคแพ้ภูมิตัวเองชนิดลูปัส | 9:1 [ 22 ] |
| โรคข้ออักเสบรูมาตอยด์ | 5:2 [ 22 ] |
| โรคท่อน้ำดีอักเสบเรื้อรังชนิดปฐมภูมิ | 1:2 |
เพศ
โรคภูมิต้านตนเองส่วนใหญ่เกี่ยวข้องกับเพศโดยรวมแล้ว ผู้หญิงมีแนวโน้มที่จะเป็นโรคภูมิต้านตนเองมากกว่าผู้ชาย การเป็นเพศหญิงเป็นปัจจัยเสี่ยงที่สำคัญที่สุดในการเกิดโรคภูมิต้านตนเองมากกว่าปัจจัยเสี่ยงทางพันธุกรรมหรือสิ่งแวดล้อมอื่นๆ ที่ค้นพบ[ 23 ] [ 24 ] โรค ภูมิต้านตนเองที่พบมากในผู้หญิง ได้แก่โรคลูปัสโรคท่อน้ำดีอักเสบปฐมภูมิโรค เกร ฟส์ โรคต่อมไทรอยด์อักเสบฮาชิโมโตะและ โรค ปลอกประสาทเสื่อมแข็งเป็นต้น โรคภูมิต้านตนเองบางโรคที่ผู้ชายมีโอกาสเป็นเท่าๆ กับผู้หญิงหรือมากกว่า ได้แก่โรคกระดูกสันหลังอักเสบ โรค เบาหวานชนิด ที่ 1 โรคหลอดเลือดอักเสบชนิดแกรนูโลมาโตซิสโรคท่อน้ำดีอักเสบปฐมภูมิและโรค สะเก็ดเงิน
เหตุผลเกี่ยวกับบทบาทของเพศในโรคภูมิต้านตนเองนั้นแตกต่างกันไป โดยทั่วไปแล้วผู้หญิงมักมีการตอบสนองต่อการอักเสบมากกว่าผู้ชายเมื่อระบบภูมิคุ้มกันของพวกเธอถูกกระตุ้น ซึ่งเพิ่มความเสี่ยงต่อโรคภูมิต้านตนเอง การมีส่วนร่วมของฮอร์โมนเพศบ่งชี้ได้จากข้อเท็จจริงที่ว่าโรคภูมิต้านตนเองหลายชนิดมีแนวโน้มที่จะผันผวนตามการเปลี่ยนแปลงของฮอร์โมน เช่น ในระหว่างตั้งครรภ์ ในรอบเดือน หรือเมื่อใช้ยาคุมกำเนิด ประวัติการตั้งครรภ์ยังดูเหมือนจะทำให้มีความเสี่ยงต่อโรคภูมิต้านตนเองเพิ่มขึ้นอย่างต่อเนื่อง มีการเสนอแนะว่าการแลกเปลี่ยนเซลล์โดยตรงเพียงเล็กน้อยระหว่างแม่และลูกในระหว่างตั้งครรภ์อาจกระตุ้นให้เกิดโรคภูมิต้านตนเองได้[ 25 ]ซึ่งจะทำให้ความสมดุลทางเพศเอนเอียงไปทางเพศหญิง
ทฤษฎีอีกทฤษฎีหนึ่งชี้ให้เห็นว่าแนวโน้มที่เพศหญิงจะเป็นโรคภูมิต้านตนเองสูงนั้นเกิดจากการปิดใช้งานโครโมโซม X ที่ไม่สมดุล[ 26 ]ทฤษฎีความเบี่ยงเบนของการปิดใช้งาน X ซึ่งเสนอโดย Jeff Stewart จากมหาวิทยาลัยพรินซ์ตัน ได้รับการยืนยันจากการทดลองในโรคหนังแข็งและโรคไทรอยด์อักเสบจากภูมิต้านตนเอง เมื่อ ไม่ นานมานี้ [ 27 ]กลไกความไวต่อพันธุกรรมที่ซับซ้อนอื่นๆ ที่เชื่อมโยงกับโครโมโซม X ได้รับการเสนอและอยู่ระหว่างการตรวจสอบ
ปัจจัยด้านสิ่งแวดล้อม
โรคติดเชื้อและปรสิต
มีความสัมพันธ์ผกผันที่น่าสนใจระหว่างโรคติดเชื้อและโรคภูมิต้านตนเอง ในพื้นที่ที่มีโรคติดเชื้อหลายชนิดระบาด โรคภูมิต้านตนเองมักพบได้น้อยมาก ในทางกลับกัน ดูเหมือนว่าความจริงจะเป็นเช่นนั้นในระดับหนึ่งสมมติฐานด้านสุขอนามัยอธิบายความสัมพันธ์เหล่านี้ว่าเป็นผลมาจากกลยุทธ์การควบคุมภูมิคุ้มกันของเชื้อโรค แม้ว่าการสังเกตดังกล่าวจะถูกเรียกว่าเป็นเท็จและไม่มีประสิทธิภาพในหลายแง่มุม แต่จากการศึกษาบางชิ้นพบว่าการติดเชื้อปรสิตมีความเกี่ยวข้องกับการลดลงของกิจกรรมของโรคภูมิต้านตนเอง[ 28 ] [ 29 ] [ 30 ]
กลไกที่สันนิษฐานไว้คือ ปรสิตจะลดทอนการตอบสนองทางภูมิคุ้มกันของโฮสต์เพื่อปกป้องตัวเอง ซึ่งอาจเป็นประโยชน์โดยบังเอิญต่อโฮสต์ที่มีโรคภูมิคุ้มกันบกพร่องอยู่แล้ว รายละเอียดของการปรับเปลี่ยนภูมิคุ้มกันของปรสิตยังไม่เป็นที่ทราบแน่ชัด แต่คาดว่าอาจรวมถึงการหลั่งสารต้านการอักเสบหรือการรบกวนการส่งสัญญาณภูมิคุ้มกันของโฮสต์
ข้อสังเกตที่ดูขัดแย้งกันอย่างหนึ่งคือ ความสัมพันธ์ที่แน่นแฟ้นระหว่างจุลินทรีย์บางชนิดกับโรคภูมิต้านตนเอง ตัวอย่างเช่น เชื้อKlebsiella pneumoniaeและไวรัส Coxsackievirus Bมีความสัมพันธ์อย่างมากกับโรคกระดูกสันหลังอักเสบและโรคเบาหวานชนิดที่ 1ตามลำดับ ซึ่งอธิบายได้จากแนวโน้มของจุลินทรีย์ที่ก่อให้เกิดการติดเชื้อในการสร้างซูเปอร์แอนติเจนที่สามารถกระตุ้นการทำงานของเซลล์ B-lymphocytes แบบหลายโคลนและการสร้างแอนติบอดีจำนวนมากที่มีความจำเพาะแตกต่างกัน ซึ่งบางส่วนอาจมีปฏิกิริยาต่อตนเอง (ดูด้านล่าง)
สารเคมีและยา
สารเคมีและยาบางชนิดอาจเกี่ยวข้องกับการเกิดโรคภูมิต้านตนเอง หรือภาวะที่เลียนแบบโรคภูมิต้านตนเอง ที่เห็นได้ชัดที่สุดคือ โรคลูปัสอีริธีมาโตซัสที่เกิดจากยาโดยปกติแล้ว การหยุดใช้ยาที่เป็นสาเหตุจะช่วยรักษาอาการในผู้ป่วยได้
ปัจจุบันเป็นที่ทราบกันดีว่าการสูบบุหรี่เป็นปัจจัยเสี่ยงสำคัญทั้งต่อการเกิดและความรุนแรงของโรคข้ออักเสบรูมาตอยด์ซึ่งอาจเกี่ยวข้องกับ การเกิด ซิโทรลลิเนชัน ที่ผิดปกติ ของโปรตีน เนื่องจากผลของการสูบบุหรี่มีความสัมพันธ์กับการมีแอนติบอดีต่อเปปไทด์ ซิโทรลลิ เนต
กลไกการเกิดโรคภูมิต้านตนเอง
เชื่อกันว่ามีกลไกหลายอย่างที่เกี่ยวข้องกับการเกิดโรคภูมิต้านตนเอง โดยมีปัจจัยทางพันธุกรรมและสิ่งแวดล้อมเข้ามาเกี่ยวข้อง บทความนี้ไม่สามารถอธิบายกลไกแต่ละอย่างได้อย่างละเอียด แต่จะสรุปกลไกที่สำคัญบางส่วนไว้ดังนี้:
- การข้ามขั้นตอนของเซลล์ T – ระบบภูมิคุ้มกันปกติจำเป็นต้องมีการกระตุ้นเซลล์ Bโดยเซลล์ Tก่อนที่เซลล์ B จะสามารถเปลี่ยนแปลงไปเป็นเซลล์พลาสมา B และผลิตแอนติบอดีในปริมาณมากได้ ในบางกรณีที่หายาก การข้ามขั้นตอนนี้อาจเกิดขึ้นได้ เช่น การติดเชื้อจากจุลินทรีย์ที่สร้างซูเปอร์แอนติเจนซึ่งสามารถกระตุ้นการทำงานของเซลล์ B หรือแม้แต่เซลล์ T ในหลายโคลนได้ โดยการจับกับหน่วยย่อยเบตาของตัวรับเซลล์ T โดยตรงในลักษณะที่ไม่จำเพาะเจาะจง
- ความไม่สอดคล้องกันระหว่างเซลล์ T และเซลล์ B – โดยปกติแล้ว การตอบสนองทางภูมิคุ้มกันจะเกี่ยวข้องกับการตอบสนองของเซลล์ B และเซลล์ T ต่อแอนติเจนเดียวกัน แม้ว่าเราจะรู้ว่าเซลล์ B และเซลล์ T จดจำสิ่งที่แตกต่างกันมาก เช่น โครงสร้างบนพื้นผิวของโมเลกุลสำหรับเซลล์ B และชิ้นส่วนเปปไทด์ที่ผ่านการประมวลผลแล้วของโปรตีนสำหรับเซลล์ T อย่างไรก็ตาม เท่าที่เราทราบ ยังไม่มีสิ่งใดที่กำหนดให้ต้องเป็นเช่นนั้น สิ่งที่จำเป็นคือ เซลล์ B ที่จดจำแอนติเจน X จะดูดซึมและประมวลผลโปรตีน Y (โดยปกติ = X) และนำเสนอต่อเซลล์ T Roosnek และ Lanzavecchia แสดงให้เห็นว่าเซลล์ B ที่จดจำ IgGFc สามารถได้รับความช่วยเหลือจากเซลล์ T ใดๆ ที่ตอบสนองต่อแอนติเจนที่ดูดซึมร่วมกับ IgG โดยเซลล์ B ซึ่งเป็นส่วนหนึ่งของสารเชิงซ้อนทางภูมิคุ้มกัน ในโรคเซลิแอค ดูเหมือนว่าเซลล์ B ที่จดจำทิชชูทรานส์กลูตามีนจะได้รับความช่วยเหลือจากเซลล์ T ที่จดจำกลูเตน
- การป้อนกลับที่ผิดปกติผ่านตัวรับเซลล์ B – ลักษณะเด่นของโรคภูมิต้านตนเองในมนุษย์คือ โรคนี้มักจำกัดอยู่เฉพาะกลุ่มแอนติเจนขนาดเล็ก ซึ่งหลายชนิดมีบทบาทในการส่งสัญญาณในระบบภูมิคุ้มกัน (DNA, C1q, IgGFc, Ro, ตัวรับ Con. A, ตัวรับ Peanut agglutinin (PNAR)) ข้อเท็จจริงนี้ทำให้เกิดแนวคิดที่ว่า ภาวะภูมิต้านตนเองที่เกิดขึ้นเองตามธรรมชาติ อาจเกิดขึ้นเมื่อการจับกันของแอนติบอดีกับแอนติเจนบางชนิด นำไปสู่การส่งสัญญาณที่ผิดปกติกลับไปยังเซลล์ B ต้นกำเนิดผ่านลิแกนด์ที่อยู่บนเยื่อหุ้มเซลล์ ลิแกนด์เหล่านี้ได้แก่ ตัวรับเซลล์ B (สำหรับแอนติเจน), ตัวรับ IgG Fc, CD21 ซึ่งจับกับคอมพลีเมนต์ C3d, ตัวรับ Toll-like receptors 9 และ 7 (ซึ่งสามารถจับกับ DNA และนิวคลีโอโปรตีน) และ PNAR นอกจากนี้ ยังสามารถจินตนาการถึงการกระตุ้น B เซลล์ที่ผิดปกติทางอ้อมเพิ่มเติมได้ด้วยออโตแอนติบอดีต่อตัวรับอะเซทิลโคลีน (บนเซลล์ไมออยด์ของต่อมไทมัส) และฮอร์โมนและโปรตีนที่จับกับฮอร์โมน ร่วมกับแนวคิดเรื่องความไม่สอดคล้องกันระหว่าง T-cell และ B-cell แนวคิดนี้ก่อให้เกิดพื้นฐานของสมมติฐานเกี่ยวกับเซลล์ B ที่มีปฏิกิริยาต่อตนเองที่ดำรงอยู่ต่อไป[ 31 ]เซลล์ B ที่มีปฏิกิริยาต่อตนเองในภาวะภูมิคุ้มกันต่อตนเองที่เกิดขึ้นเองตามธรรมชาติถือว่าอยู่รอดได้เนื่องจากการบิดเบือนทั้งเส้นทางช่วยเหลือของ T cell และสัญญาณป้อนกลับผ่านตัวรับ B cell จึงเอาชนะสัญญาณเชิงลบที่รับผิดชอบต่อความทนทานต่อตนเองของ B cell โดยไม่จำเป็นต้องสูญเสียความทนทานต่อตนเองของ T cell
- การเลียนแบบโมเลกุล –แอนติเจน จากภายนอก อาจมีโครงสร้างคล้ายคลึงกับแอนติเจนของโฮสต์บางชนิด ดังนั้น แอนติบอดีใดๆ ที่สร้างขึ้นต่อต้านแอนติเจนนี้ (ซึ่งเลียนแบบแอนติเจนของตนเอง) ก็สามารถจับกับแอนติเจนของโฮสต์และกระตุ้นการตอบสนองทางภูมิคุ้มกันได้ในทางทฤษฎี แนวคิดเรื่องการเลียนแบบโมเลกุลเกิดขึ้นในบริบทของไข้รูมาติกซึ่งเกิดจากการติดเชื้อแบคทีเรียกลุ่ม A เบต้า-ฮีโมไลติกสเต รปโต ค็อกคัส แม้ว่าไข้รูมาติกจะถูกกล่าวถึงว่าเป็นผลมาจากการเลียนแบบโมเลกุลมานานกว่าครึ่งศตวรรษแล้ว แต่ก็ยังไม่มีการระบุแอนติเจนอย่างเป็นทางการ (หากมีก็มีการเสนอแอนติเจนมากเกินไป) ยิ่งไปกว่านั้น การกระจายตัวของโรคในเนื้อเยื่อที่ซับซ้อน (หัวใจ ข้อต่อ ผิวหนัง ปมประสาทฐาน) ขัดแย้งกับความเป็นไปได้ที่โรคจะมีแอนติเจนเฉพาะที่หัวใจ จึงยังคงเป็นไปได้ว่าโรคนี้เกิดจากปฏิกิริยาที่ผิดปกติระหว่างสารเชิงซ้อนทางภูมิคุ้มกัน ส่วนประกอบของคอมพลีเมนต์ และเยื่อบุผนังหลอดเลือด เป็นต้น
- ปฏิกิริยาข้ามไอดีโอไทป์ – ไอ ดีโอไทป์เป็นอีพิโทปแอนติเจนที่พบในส่วนที่จับกับแอนติเจน (Fab) ของโมเลกุลอิมมูโนโกลบูลิน พล็อตซ์และโอลด์สโตนได้นำเสนอหลักฐานว่าภาวะภูมิคุ้มกันทำลายตนเองสามารถเกิดขึ้นได้จากปฏิกิริยาข้ามระหว่างไอดีโอไทป์บนแอนติบอดีต้านไวรัสและตัวรับเซลล์โฮสต์สำหรับไวรัสที่เกี่ยวข้อง ในกรณีนี้ ตัวรับเซลล์โฮสต์ถูกมองว่าเป็นภาพภายในของไวรัส และแอนติบอดีต่อไอดีโอไทป์สามารถทำปฏิกิริยากับเซลล์โฮสต์ได้
- การทำงานผิดปกติของไซโตไคน์ – เมื่อไม่นานมานี้ ไซโตไคน์ถูกแบ่งออกเป็นสองกลุ่มตามประชากรของเซลล์ที่พวกมันส่งเสริมการทำงาน ได้แก่ เซลล์ทีช่วยชนิดที่ 1 หรือชนิดที่ 2 ไซโตไคน์กลุ่มที่สอง ซึ่งรวมถึง IL-4, IL-10 และTGF-β (เป็นต้น) ดูเหมือนจะมีบทบาทในการป้องกันการตอบสนองทางภูมิคุ้มกันที่ก่อให้เกิดการอักเสบมากเกินไป
- การตายของเซลล์เดนไดรติก – เซลล์ระบบภูมิคุ้มกันที่เรียกว่าเซลล์เดนไดรติกจะนำเสนอแอนติเจนให้กับลิมโฟไซต์ ที่ทำงานอยู่ เซลล์เดนไดรติกที่บกพร่องในการตายของเซลล์อาจนำไปสู่การกระตุ้นลิมโฟไซต์ทั่วร่างกายที่ไม่เหมาะสมและส่งผลให้ความทนทานต่อตนเองลดลง[ 32 ]
- การแพร่กระจายของเอพิโทปหรือการเปลี่ยนแปลงของเอพิโทป – เมื่อปฏิกิริยาภูมิคุ้มกันเปลี่ยนจากการกำหนดเป้าหมายเอพิโทป หลัก ไปเป็นการกำหนดเป้าหมายเอพิโทปอื่นๆ ด้วย[ 33 ]ในทางตรงกันข้ามกับการเลียนแบบโมเลกุล เอพิโทปอื่นๆ ไม่จำเป็นต้องมีโครงสร้างคล้ายกับเอพิโทปหลัก
- การดัดแปลงเอพิโทปหรือการเปิดเผยเอพิโทปแบบซ่อนเร้น – กลไกของโรคภูมิต้านตนเองนี้มีความพิเศษตรงที่ไม่ได้เกิดจากความบกพร่องในระบบเม็ดเลือด แต่โรคเกิดจากการเปิดเผยพันธะ N-ไกลแคน (พอลิแซ็กคาไรด์) แบบซ่อนเร้นซึ่งพบได้ทั่วไปในยูคาริโอตและโปรคาริโอตระดับต่ำบนไกลโคโปรตีนของเซลล์และอวัยวะที่ไม่ใช่เม็ดเลือดของสัตว์เลี้ยงลูกด้วยนม[ 34 ] การเปิดเผยไกลแคนดั้งเดิมทางวิวัฒนาการนี้จะกระตุ้นตัวรับ เซลล์ภูมิคุ้มกันโดยกำเนิด ของสัตว์ เลี้ยงลูกด้วยนมอย่างน้อยหนึ่งตัวเพื่อเหนี่ยวนำให้เกิดภาวะอักเสบเรื้อรังที่ไม่ติดเชื้อ ในกรณีที่มีความเสียหายของเซลล์เรื้อรังและอักเสบ ระบบภูมิคุ้มกันแบบปรับตัวจะถูกดึงเข้ามา และความทนทานต่อตนเองจะสูญเสียไปพร้อมกับการผลิตแอนติบอดีต่อต้านตนเองที่เพิ่มขึ้น ในรูปแบบของโรคนี้ การขาดลิมโฟไซต์สามารถเร่งความเสียหายของอวัยวะ และการให้ IgG ทางหลอดเลือดดำสามารถรักษาได้ แม้ว่ากลไกการเกิดโรคภูมิต้านตนเองนี้อาจเป็นสาเหตุของโรคเสื่อมต่างๆ แต่ในปัจจุบันยังไม่มีวิธีการวินิจฉัยกลไกการเกิดโรคนี้ ดังนั้นบทบาทของมันในโรคภูมิต้านตนเองในมนุษย์จึงยังไม่เป็นที่ทราบแน่ชัด
บทบาทของเซลล์ควบคุมภูมิคุ้มกันชนิดพิเศษ เช่นเซลล์ T ควบคุม (regulatory T cells) , เซลล์ NKTและเซลล์ T γδในการเกิดโรคภูมิต้านตนเองกำลังอยู่ระหว่างการศึกษา
การจำแนกประเภท
โรคภูมิต้านตนเองสามารถแบ่งออกได้เป็นโรคภูมิต้านตนเองแบบทั่วร่างกาย และโรคภูมิต้านตนเองเฉพาะอวัยวะหรือเฉพาะที่ ขึ้นอยู่กับลักษณะทางคลินิกและพยาธิวิทยาหลักของแต่ละโรค
- โรคภูมิต้านตนเองแบบทั่วร่างกายได้แก่โรคเซลิแอคโรคลูปัสอีริธีมาโตซัส กลุ่มอาการโจเกรนโรคหนังแข็งโรคข้ออักเสบรูมาตอยด์ โรคหลอดเลือด อักเสบจากไครโอโกลบูลิ นโรคผิวหนังอักเสบจากกล้ามเนื้อ และอื่นๆ อีกมากมาย โรคเหล่านี้มักเกี่ยวข้องกับแอนติบอดีต่อต้านแอนติเจนที่ไม่จำเพาะต่อเนื้อเยื่อ ดังนั้น แม้ว่าโรคกล้ามเนื้ออักเสบ หลายส่วน จะมีลักษณะเฉพาะต่อเนื้อเยื่อมากหรือน้อย แต่ก็อาจถูกรวมอยู่ในกลุ่มนี้ได้ เนื่องจากแอนติเจนต่อต้านตนเองมักเป็นเอนไซม์ t-RNA synthetase ที่พบได้ทั่วไป
- กลุ่มอาการ เฉพาะที่ซึ่งส่งผลกระทบต่ออวัยวะหรือเนื้อเยื่อเฉพาะส่วน:
- ระบบต่อมไร้ท่อ: โรคเบาหวานชนิดที่ 1 , โรคต่อมไทรอยด์อักเสบเรื้อรัง (ฮาชิโมโตะ) , โรคแอดดิสัน
- ระบบทางเดินอาหาร: โรคโลหิตจางชนิดร้ายแรง
- โรคผิวหนัง: โรคเพมฟิกัส วัลกา ริส , โรคด่างขาว
- ระบบโลหิตวิทยา: โรคโลหิตจางจากเม็ดเลือดแดงแตกจากภูมิคุ้มกันตนเอง , โรคเกล็ดเลือดต่ำชนิดไม่ทราบสาเหตุ
- โรคทางระบบประสาท: โรคปลอกประสาทเสื่อมแข็ง , โรคกล้ามเนื้ออ่อนแรง , โรคสมองอักเสบจากภูมิคุ้มกันตนเอง , โรคเสียการทรงตัวจากกลูเตน
การใช้แผนการจำแนกประเภทแบบดั้งเดิม "เฉพาะอวัยวะ" และ "ไม่เฉพาะอวัยวะ" ทำให้โรคหลายชนิดถูกรวมไว้ด้วยกันภายใต้ร่มเงาของโรคภูมิต้านตนเอง อย่างไรก็ตาม ความผิดปกติของการอักเสบเรื้อรังในมนุษย์หลายชนิดขาดความสัมพันธ์ที่ชัดเจนของพยาธิวิทยาภูมิคุ้มกันที่ขับเคลื่อนโดยเซลล์ B และ T ในช่วงทศวรรษที่ผ่านมา มีการยืนยันอย่างแน่ชัดแล้วว่า "การอักเสบของเนื้อเยื่อต่อต้านตนเอง " ไม่จำเป็นต้องอาศัยการตอบสนองของเซลล์ T และ B ที่ผิดปกติ[ 35 ]
สิ่งนี้นำไปสู่ข้อเสนอเมื่อเร็ว ๆ นี้ว่าควรพิจารณาสเปกตรัมของภูมิคุ้มกันตนเองตาม "ความต่อเนื่องของโรคทางภูมิคุ้มกัน" โดยมีโรคภูมิคุ้มกันตนเองแบบคลาสสิกอยู่ที่ปลายด้านหนึ่ง และโรคที่เกิดจากระบบภูมิคุ้มกันโดยกำเนิดอยู่ที่ปลายอีกด้านหนึ่ง ภายในแผนผังนี้ สามารถรวมสเปกตรัมทั้งหมดของภูมิคุ้มกันตนเองได้ โรคภูมิคุ้มกันตนเองทั่วไปในมนุษย์หลายโรคสามารถมองเห็นได้ว่ามีพยาธิสภาพทางภูมิคุ้มกันที่เกิดจากระบบภูมิคุ้มกันโดยกำเนิดอย่างมีนัยสำคัญโดยใช้แผนผังใหม่นี้ แผนผังการจำแนกประเภทใหม่นี้มีนัยสำคัญต่อการทำความเข้าใจกลไกของโรคและการพัฒนาการรักษา[ 35 ]
การวินิจฉัย
การวินิจฉัยโรคภูมิต้านตนเองส่วนใหญ่ขึ้นอยู่กับประวัติและการตรวจร่างกายของผู้ป่วยอย่างแม่นยำ และการสงสัยในระดับสูงเมื่อพบความผิดปกติบางอย่างในการตรวจทางห้องปฏิบัติการทั่วไป (เช่นโปรตีนซี-รีแอคทีฟ สูงขึ้น )
ในโรคที่ส่งผลต่อระบบต่างๆ ในร่างกายหลายชนิด สามารถใช้ การตรวจทางซีรั่มวิทยาเพื่อตรวจหา แอนติบอดีจำเพาะ ได้ ส่วนโรคที่เกิดขึ้นเฉพาะที่นั้น การวินิจฉัยที่ดีที่สุดคือการตรวจด้วยวิธี อิมมูโนฟลูออเรสเซนซ์จากชิ้นเนื้อตัวอย่าง
แอนติบอดีต่อต้านตนเองถูกนำมาใช้ในการวินิจฉัยโรคภูมิต้านตนเองหลายชนิด โดยจะวัดระดับแอนติบอดีต่อต้านตนเองเพื่อประเมินความคืบหน้าของโรค
การรักษา
การรักษาโรคภูมิต้านตนเองแบบดั้งเดิมมักเป็นการกดภูมิคุ้มกันต้านการอักเสบหรือบรรเทาอาการ [ 12 ] การจัดการการอักเสบเป็นสิ่งสำคัญในโรคภูมิต้านตนเอง[ 36 ]การรักษาที่ไม่เกี่ยวกับภูมิคุ้มกัน เช่น การทดแทนฮอร์โมนในโรคต่อมไทรอยด์อักเสบฮาชิโมโตะหรือโรคเบาหวานชนิดที่ 1 จะรักษาผลลัพธ์ของการตอบสนองที่ทำลายตนเอง ดังนั้นจึงเป็นการรักษาแบบบรรเทาอาการ การควบคุมอาหารช่วยลดความรุนแรงของโรคเซลิแอค การรักษาด้วยสเตียรอยด์หรือ NSAID ช่วยลดอาการอักเสบของโรคหลายชนิดIVIGใช้สำหรับCIDPและGBS การรักษา ด้วยภูมิคุ้มกันบำบัดเฉพาะเช่น สารต้าน TNFα (เช่นetanercept ) สารลดจำนวนเซลล์ B rituximabสารต้านตัวรับ IL-6 tocilizumabและสารปิดกั้นการกระตุ้นร่วมabataceptได้รับการพิสูจน์แล้วว่ามีประโยชน์ในการรักษาโรคข้อ อักเสบรูมาตอยด์ การรักษาด้วยภูมิคุ้มกันบำบัดบางวิธีอาจเกี่ยวข้องกับความเสี่ยงที่เพิ่มขึ้นของผลข้างเคียง เช่น ความเสี่ยงต่อการติดเชื้อ
การบำบัดด้วยพยาธิ เป็นวิธีการทดลองที่เกี่ยวข้องกับการฉีด พยาธิในลำไส้(helminths) เฉพาะชนิดเข้าไปในผู้ป่วย ปัจจุบันมีวิธีการรักษาที่เกี่ยวข้องกันอย่างใกล้ชิดอยู่ 2 วิธี ได้แก่ การฉีด Necator americanus ซึ่งรู้จักกันทั่วไปในชื่อพยาธิปากขอหรือ Trichuris Suis Ova ซึ่งรู้จักกันทั่วไปในชื่อไข่พยาธิแส้หมู[ 37 ] [ 38 ] [ 39 ] [ 40 ] [ 41 ]
การฉีดวัคซีนทีเซลล์กำลังอยู่ระหว่างการศึกษาเพื่อเป็นวิธีการรักษาที่เป็นไปได้ในอนาคตสำหรับโรคภูมิคุ้มกันบกพร่อง[ 42 ]
โภชนาการและภูมิคุ้มกันตนเอง
วิตามินดี/แสงแดด
- เนื่องจากเซลล์และเนื้อเยื่อของมนุษย์ส่วนใหญ่มีตัวรับวิตามินดี รวมถึงเซลล์ T และเซลล์ B ระดับวิตามินดีที่เพียงพอจึงสามารถช่วยในการควบคุมระบบภูมิคุ้มกันได้[ 43 ] วิตามินดีมีบทบาทในการทำงานของระบบภูมิคุ้มกันโดยการออกฤทธิ์ต่อ เซลล์ T และ เซลล์นักฆ่าตามธรรมชาติ[ 44 ] งานวิจัยแสดงให้เห็นถึงความสัมพันธ์ระหว่างวิตามินดีในซีรั่มต่ำกับโรคภูมิต้านตนเอง รวมถึงโรค ปลอกประสาทเสื่อมแข็งโรค เบาหวานชนิดที่ 1และ โรคแพ้ภูมิต้านตนเองชนิดลูปัส (โดยทั่วไปเรียกว่าลูปัส) [ 44 ] [ 45 ] [ 46 ] อย่างไรก็ตาม เนื่องจาก ภาวะไวต่อแสง เกิดขึ้นในลูปัส ผู้ป่วยจึงควรหลีกเลี่ยงแสงแดด ซึ่งอาจเป็นสาเหตุของการขาดวิตามินดีที่พบในโรคนี้[ 44 ] [ 45 ] [ 46 ] โพลี มอร์ฟิซึม ใน ยีน ตัวรับวิตามินดี มักพบในผู้ที่เป็นโรคภูมิต้านตนเอง ซึ่งเป็นกลไกหนึ่งที่เป็นไปได้สำหรับบทบาทของวิตามินดีในโรคภูมิต้านตนเอง[ 44 ] [ 45 ] มีหลักฐานที่หลากหลายเกี่ยวกับผลของการเสริมวิตามินดีในโรคเบาหวานประเภทที่ 1 โรคลูปัส และโรคปลอกประสาทเสื่อมแข็ง[ 44 ] [ 45 ] [ 46 ]
กรดไขมันโอเมก้า 3
- การศึกษาแสดงให้เห็นว่าการบริโภคกรดไขมันโอเมก้า-3 ในปริมาณที่เพียงพอสามารถต่อต้านผลกระทบของกรดอะราคิโดนิก ซึ่งเป็นสาเหตุของอาการของโรคภูมิคุ้มกันบกพร่อง การทดลองในมนุษย์และสัตว์แสดงให้เห็นว่าโอเมก้า-3 เป็นวิธีการรักษา ที่มีประสิทธิภาพ สำหรับโรคข้ออักเสบรูมาตอยด์ โรคลำไส้อักเสบ โรคหอบหืด และโรคสะเก็ดเงินหลายกรณี[ 47 ]
- แม้ว่าภาวะซึมเศร้าขั้นรุนแรงจะไม่ใช่โรคภูมิต้านตนเองเสมอไป แต่อาการทางสรีรวิทยาบางอย่างมีลักษณะเป็นการอักเสบและภูมิต้านตนเอง โอเมก้า 3 อาจยับยั้งการผลิตอินเตอร์เฟรอนแกมมาและไซโตไคน์อื่นๆ ซึ่งเป็นสาเหตุของอาการทางสรีรวิทยาของภาวะซึมเศร้า นี่อาจเป็นเพราะความไม่สมดุลของกรดไขมันโอเมก้า 3 และโอเมก้า 6 ซึ่งมีผลตรงกันข้าม มีบทบาทสำคัญในสาเหตุของภาวะซึมเศร้าขั้นรุนแรง[ 47 ]
โปรไบโอติกส์/จุลินทรีย์ประจำถิ่น
- แบคทีเรียและจุลินทรีย์หลายชนิดที่มีอยู่ในผลิตภัณฑ์นมหมัก โดยเฉพาะLactobacillus caseiได้รับการพิสูจน์แล้วว่าสามารถกระตุ้นการตอบสนองทางภูมิคุ้มกันต่อเนื้องอกในหนูและควบคุมการทำงานของภูมิคุ้มกัน ชะลอหรือป้องกันการเกิดโรคเบาหวานชนิดไม่อ้วนได้ โดยเฉพาะอย่างยิ่งสายพันธุ์ Shirota ของL. casei (LcS) สายพันธุ์ LcS พบได้ส่วนใหญ่ในโยเกิร์ตและผลิตภัณฑ์ที่คล้ายคลึงกันในยุโรปและญี่ปุ่น และพบได้น้อยในที่อื่นๆ[ 48 ]
- มีการตั้งทฤษฎีว่าอนุมูลอิสระมีส่วนทำให้เกิดโรคเบาหวานประเภทที่ 1 ในทารกและเด็กเล็ก ดังนั้นความเสี่ยงจึงอาจลดลงได้ด้วยการบริโภคสารต้านอนุมูลอิสระในปริมาณสูงระหว่างตั้งครรภ์ อย่างไรก็ตาม การศึกษาที่ดำเนินการในโรงพยาบาลแห่งหนึ่งในฟินแลนด์ตั้งแต่ปี 1997 ถึง 2002 สรุปว่าไม่มีความสัมพันธ์ทางสถิติอย่างมีนัยสำคัญระหว่างการบริโภคสารต้านอนุมูลอิสระและความเสี่ยงต่อโรคเบาหวาน[ 49 ]การศึกษานี้เกี่ยวข้องกับการติดตามการบริโภคอาหารผ่านแบบสอบถาม และประมาณการการบริโภคสารต้านอนุมูลอิสระบนพื้นฐานนี้ แทนที่จะใช้การวัดที่แม่นยำหรือการใช้ผลิตภัณฑ์เสริมอาหาร
ดูเพิ่มเติม
ลิงก์ภายนอก
- สมาคมโรคภูมิคุ้มกันบกพร่องแห่งอเมริกา: องค์กรไม่แสวงหาผลกำไรที่ให้การสนับสนุน
- เครือข่ายความทนทานต่อภูมิคุ้มกัน: แหล่งข้อมูลที่เน้นการวิจัย
- รางวัลโนเบล – รางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ประจำปี 1960 มอบให้แก่ แฟรงค์ เอ็ม. เบอร์เน็ต และ ปีเตอร์ บี. เมดาวาร์ "สำหรับการค้นพบภาวะทนต่อภูมิคุ้มกันที่เกิดขึ้นภายหลัง"
- ฐานข้อมูลและพอร์ทัลการวิเคราะห์ภูมิคุ้มกันวิทยา – แหล่งข้อมูลฐานข้อมูลที่ได้รับการสนับสนุนจาก NIAID ซึ่งรวบรวมข้อมูลอ้างอิงและข้อมูลการทดลองครอบคลุมสาขาภูมิคุ้มกันวิทยาทั้งหมด
- ทำความเข้าใจเกี่ยวกับโรคภูมิต้านตนเอง - สถาบันโรคข้ออักเสบ โรคกระดูกและกล้ามเนื้อ และโรคผิวหนังแห่งชาติของสหรัฐอเมริกา
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ภูมิคุ้มกันตนเอง
ในทาง ภูมิคุ้มกันวิทยา ภาวะ ภูมิคุ้มกันทำลายตนเอง คือระบบ การตอบสนองทางภูมิคุ้มกัน ของสิ่งมีชีวิต ต่อ เซลล์ เนื้อเยื่อ และส่วนประกอบอื่นๆ ของร่างกายที่แข็งแรงของตนเอง [ 1 ] [ 2 ]...
ประวัติศาสตร์
ในช่วงปลายศตวรรษที่ 19 มีความเชื่อว่าระบบภูมิคุ้มกันไม่สามารถตอบโต้เนื้อเยื่อของร่างกายเองได้ พอล เออร์ลิช ในช่วงต้นศตวรรษที่ 20 ได้เสนอแนวคิดเรื่อง " ความสยองขวัญจากปฏิกิริยาต่อต้านตนเอง" ( horror autotoxicus )...
ภาวะภูมิคุ้มกันบกพร่องระดับต่ำ
ภาวะภูมิคุ้มกันต่อต้านตนเองอาจมีบทบาทในการทำให้เกิดการตอบสนองทางภูมิคุ้มกันอย่างรวดเร็วในระยะเริ่มต้นของการติดเชื้อ เมื่อปริมาณของ แอนติเจน แปลกปลอม มีจำกัด (เช่น เมื่อมี เชื้อโรค อยู่น้อย) ในการศึกษาของ Stefanova et al.
ความทนทานทางภูมิคุ้มกัน
งานบุกเบิกโดย Noel Rose และ Ernst Witebsky ในนิวยอร์ก และ Roitt และ Doniach ที่ University College London ได้ให้หลักฐานที่ชัดเจนว่า อย่างน้อยในแง่ของเซลล์ B ที่สร้างแอนติบอดี (ลิมโฟไซต์ B) โรคต่างๆ เช่น...