อ่าน 24 นาที
ตัวรับปัจจัยการเจริญเติบโตของเซลล์ตับ
ตัวรับปัจจัยการเจริญเติบโตของเซลล์ตับ ( ตัวรับ HGF ) [ 5 ] [ 6 ] เป็น โปรตีน ที่ในมนุษย์ถูกเข้ารหัสโดย ยีน MET โปรตีนนี้มีกิจกรรม ไทโรซีนไคเนส [ 7 ]...
ตัวรับปัจจัยการเจริญเติบโตของเซลล์ตับ
| เมท | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | MET , โปรโตออนโคยีน MET, ตัวรับไทโรซีนไคเนส, AUTS9, HGFR, RCCP2, c-Met, DFNB97, OSFD, c-met | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | โอมิม : 164860 ; เอ็มจีไอ : 96969 ; โฮโมโลยีน : 206 ; GeneCards : MET ; OMA : MET - ออร์โธโลจี | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




ตัวรับปัจจัยการเจริญเติบโตของเซลล์ตับ ( ตัวรับ HGF ) [ 5 ] [ 6 ]เป็นโปรตีนที่ในมนุษย์ถูกเข้ารหัสโดยยีนMET โปรตีนนี้มีกิจกรรมไทโรซีนไคเนส[ 7 ]โปรตีนพรีเคอร์เซอร์สายเดี่ยวหลักจะถูกตัดหลังการแปลเพื่อสร้างซับยูนิตอัลฟาและเบตา ซึ่งเชื่อมต่อกันด้วยพันธะไดซัลไฟด์เพื่อสร้างตัวรับที่สมบูรณ์
ตัวรับ HGF เป็นตัวรับไทโรซีนไคเนสแบบผ่านครั้งเดียวที่จำเป็นต่อการพัฒนาตัวอ่อน การสร้างอวัยวะ และการสมานแผล ปัจจัยการเจริญเติบโตของเซลล์ตับ/ปัจจัยการกระจายตัว (HGF/SF) และไอโซฟอร์มการตัดต่อ (NK1, NK2) เป็นลิแกนด์ที่รู้จักเพียงชนิดเดียวของตัวรับ HGF โดยปกติแล้ว MET จะแสดงออกในเซลล์ที่มี ต้นกำเนิดจาก เยื่อบุผิวในขณะที่การแสดงออกของ HGF/SF นั้นจำกัดอยู่เฉพาะในเซลล์ที่มี ต้นกำเนิด จากเนื้อเยื่อเกี่ยวพันเมื่อ HGF/SF จับกับตัวรับ MET ที่จำเพาะเจาะจง มันจะกระตุ้นให้เกิดการรวมตัวกันเป็นไดเมอร์ผ่านกลไกที่ยังไม่เป็นที่เข้าใจอย่างสมบูรณ์ ซึ่งนำไปสู่การกระตุ้นการทำงาน
บางครั้ง MET ถูกเข้าใจผิดว่าเป็นตัวย่อของการเปลี่ยนผ่านจากเนื้อเยื่อเกี่ยวพันเป็นเนื้อเยื่อบุผิว ซึ่งไม่ถูกต้อง ตัวอักษรสามตัวของ MET มาจากN -methyl-N'-nitro- N -nitrosoguanidine (MNNG) [ 8 ]
การทำงานของ MET ที่ผิดปกติในมะเร็งมีความสัมพันธ์กับการพยากรณ์โรคที่ไม่ดี โดย MET ที่ทำงานผิดปกติจะกระตุ้นการเจริญเติบโตของเนื้องอก การสร้างหลอดเลือดใหม่ ( angiogenesis ) ที่ให้สารอาหารแก่เนื้องอก และการแพร่กระจายของมะเร็งไปยังอวัยวะอื่น ( metastasis ) MET มีการทำงานผิดปกติในมะเร็งหลายชนิดในมนุษย์ รวมถึงมะเร็งไต ตับ กระเพาะอาหาร เต้านม และสมอง โดยปกติแล้ว มีเพียงเซลล์ต้นกำเนิดและเซลล์บรรพบุรุษ เท่านั้น ที่แสดงออก MET ซึ่งช่วยให้เซลล์เหล่านี้เจริญเติบโตแบบรุกรานเพื่อสร้างเนื้อเยื่อใหม่ในตัวอ่อนหรือฟื้นฟูเนื้อเยื่อที่เสียหายในผู้ใหญ่ อย่างไรก็ตาม เชื่อกันว่า เซลล์ต้นกำเนิดมะเร็งจะแย่งชิงความสามารถของเซลล์ต้นกำเนิดปกติในการแสดงออก MET และกลายเป็นสาเหตุของการคงอยู่และการแพร่กระจายของมะเร็งไปยังส่วนอื่น ๆ ของร่างกาย ทั้งการแสดงออกมากเกินไปของ Met/HGFR รวมถึง การกระตุ้น แบบออโตครีนโดยการแสดงออกร่วมกันของลิแกนด์ปัจจัยการเจริญเติบโตของเซลล์ตับ มีส่วนเกี่ยวข้องกับการเกิดมะเร็ง[ 9 ] [ 10 ]
การกลายพันธุ์ต่างๆ ในยีน MET เกี่ยวข้องกับ มะเร็งไต ชนิดpapillary [ 11 ]
ยีน
โปรโตออนโคยีน MET ( GeneID: 4233 ) มีความยาวทั้งหมด 125,982 bp และตั้งอยู่ในตำแหน่ง 7q31 ของโครโมโซม 7 [ 12 ] METถูกถอดรหัสเป็น mRNA ที่สมบูรณ์ขนาด 6,641 bp จากนั้นจึงแปลเป็นโปรตีน MET ที่มีกรดอะมิโน 1,390 ตัว
โปรตีน
MET เป็นตัวรับไทโรซีนไคเนส (RTK) ที่ผลิตเป็นสารตั้งต้นแบบสายเดี่ยว สารตั้งต้นจะถูกตัดด้วยเอนไซม์โปรตีเอส ที่ตำแหน่ง ฟูรินเพื่อให้ได้หน่วยย่อยอัลฟาภายนอกเซลล์ที่มีการเติมหมู่ไกลโคซิลจำนวนมากและหน่วยย่อยเบตาที่อยู่ภายในเยื่อหุ้มเซลล์ ซึ่งเชื่อมต่อกันด้วยพันธะไดซัลไฟด์[ 13 ]
ภายนอกเซลล์
- บริเวณที่มีความคล้ายคลึงกับเซมาฟอริน (โดเมนเซมา) ซึ่งรวมถึงสายอัลฟาแบบเต็มและส่วนปลายด้าน N ของสายเบตา
- ลำดับที่เกี่ยวข้องกับ MET ที่อุดมไปด้วยซิสเทอีน (โดเมน MRS)
- ส่วนที่ซ้ำกันซึ่งอุดมไปด้วยไกลซีนและโพรลีน (GP repeats)
- โครงสร้างคล้ายอิมมูโนโกลบูลินสี่โครงสร้าง (โดเมน Ig) ซึ่งเป็นบริเวณปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีนทั่วไป[ 13 ]
ภายในเซลล์
ส่วนประกอบที่อยู่ติดกับเยื่อหุ้มเซลล์ซึ่งประกอบด้วย:
- สารตกค้างซีรีน (Ser 985) ซึ่งยับยั้งกิจกรรมไคเนสของตัวรับเมื่อเกิดการฟอสโฟรีเลชัน[ 14 ]
- หมู่ไทโรซีน (Tyr 1003) ซึ่งมีหน้าที่รับผิดชอบในการเกิดโพลียูบิควิทิเนชันของ MET เอนโดไซโทซิสและการย่อยสลายเมื่อเกิดปฏิสัมพันธ์กับยูบิควิทินไลเกส CBL [ 15 ]
- โดเมนไทโรซีนไคเนส ซึ่งเป็นตัวกลางในการทำงานทางชีวภาพของ MET หลังจากการกระตุ้น MET จะเกิดการถ่ายโอนหมู่ฟอสเฟตไปยังไทโรซีน 1234 และไทโรซีน 1235
- บริเวณปลาย C ประกอบด้วยไทโรซีนที่สำคัญสองตัว (Tyr 1349 และ Tyr 1356) ซึ่งแทรกอยู่ในบริเวณเชื่อมต่อซับสเตรตหลายชนิด ซึ่งสามารถดึงดูดโปรตีนอะแดปเตอร์ปลายน้ำที่มี โดเมน Src homology-2 (SH2) ได้ [ 16 ]มีรายงานว่าไทโรซีนทั้งสองตัวในบริเวณเชื่อมต่อมีความจำเป็นและเพียงพอสำหรับการส่งสัญญาณทั้งในหลอดทดลอง[ 16 ] [ 17 ]
เส้นทางการส่งสัญญาณ MET
การกระตุ้น MET โดยลิแกนด์HGFทำให้เกิดกิจกรรมเร่งปฏิกิริยาของ MET kinase ซึ่งกระตุ้นการถ่ายโอนฟอสเฟตของไทโรซีน Tyr 1234 และ Tyr 1235 ไทโรซีนทั้งสองนี้จะเข้าไปมีส่วนร่วมกับตัวส่งสัญญาณต่างๆ[ 18 ] จึงเริ่มต้นกิจกรรมทางชีวภาพที่หลากหลายซึ่งขับเคลื่อนโดย MET ซึ่งโดยรวมเรียกว่าโปรแกรมการเจริญเติบโตแบบรุกราน ตัวส่งสัญญาณจะโต้ตอบกับตำแหน่งเชื่อมต่อหลายซับสเตรตภายในเซลล์ของ MET โดยตรง เช่นGRB2 , SHC [ 19 ] SRCและซับยูนิตควบคุม p85 ของฟอสฟาติดิลอินอซิทอล-3 ไคเนส ( PI3K ) [ 19 ]หรือโดยอ้อมผ่านโปรตีนโครงสร้าง Gab1 [ 20 ]
Tyr 1349 และ Tyr 1356 ของไซต์เชื่อมต่อซับสเตรตหลายชนิดมีส่วนเกี่ยวข้องในการโต้ตอบกับ GAB1, SRC และ SHC ในขณะที่ Tyr 1356 เพียงอย่างเดียวมีส่วนเกี่ยวข้องในการดึงดูด GRB2, ฟอสโฟลิเปส C γ (PLC-γ), p85 และ SHP2 [ 21 ]
GAB1 เป็นตัวประสานงานหลักของการตอบสนองของเซลล์ต่อ MET และจับกับบริเวณภายในเซลล์ของ MET ด้วยความแรง สูง แต่มีความสัมพันธ์ต่ำ[ 22 ]เมื่อเกิดปฏิกิริยากับ MET GAB1 จะถูกฟอสโฟรีเลตที่หมู่ไทโรซีนหลายหมู่ ซึ่งจะดึงดูดตัวส่งสัญญาณหลายตัว รวมถึงPI3K , SHP2 และ PLC-γ การฟอสโฟรีเลต GAB1 โดย MET ส่งผลให้เกิดสัญญาณต่อเนื่องที่ควบคุมเส้นทางการส่งสัญญาณปลายทางส่วนใหญ่[ 23 ]
การกระตุ้นการส่งสัญญาณ
การทำงานของ MET จะกระตุ้นเส้นทาง การส่งสัญญาณ หลายเส้นทาง:
- เส้นทางRAS เป็นตัวกลางในการส่งสัญญาณ การกระจายและการแพร่กระจายที่เกิดจาก HGF ซึ่งนำไปสู่การสร้างรูปร่างแบบแตกแขนง [ 24 ] ที่น่าสังเกตคือ HGF แตกต่างจากไมโทเจน ส่วนใหญ่ โดยกระตุ้นการทำงานของ RAS อย่างต่อเนื่อง และส่งผลให้กิจกรรมMAPK ยาวนานขึ้น [ 25 ]
- เส้นทางPI3Kถูกกระตุ้นได้สองวิธี: PI3K อาจอยู่ปลายทางของ RAS หรืออาจถูกดึงดูดโดยตรงผ่านไซต์เชื่อมต่ออเนกประสงค์[ 26 ]การกระตุ้นเส้นทาง PI3K ในปัจจุบันเกี่ยวข้องกับการเคลื่อนที่ของเซลล์ผ่านการปรับโครงสร้างการยึดเกาะกับเมทริกซ์นอกเซลล์ รวมถึงการดึงดูดทรานสดิวเซอร์ที่เกี่ยวข้องกับการจัดระเบียบโครงสร้างเซลล์ เช่นRAC1และPAK ในบริเวณเฉพาะที่ การกระตุ้น PI3K ยังกระตุ้น สัญญาณการ อยู่รอดเนื่องจากการกระตุ้นเส้นทางAKT [ 27 ]
- เส้นทาง STAT ร่วมกับการกระตุ้น MAPK อย่างต่อเนื่องมีความจำเป็นต่อ การเกิดรูปร่างแบบแตกแขนง ที่เกิด จาก HGF MET กระตุ้นปัจจัยการถอดรหัสSTAT3โดยตรงผ่านโดเมนSH2 [ 28 ]
- เส้นทางเบต้า-แคทเทนินซึ่งเป็นองค์ประกอบสำคัญของเส้นทางส่งสัญญาณ Wntจะเคลื่อนย้ายเข้าไปในนิวเคลียสหลังจากการกระตุ้น MET และมีส่วนร่วมในการควบคุมการถอดรหัสของยีนจำนวนมาก[ 29 ]
- เส้นทางNotchผ่านการกระตุ้นการถอดรหัสของลิแกนด์ Delta (ดูDLL3 ) [ 30 ] [ 31 ]
บทบาทในการพัฒนา
MET ทำหน้าที่เป็นตัวกลางของโปรแกรมที่ซับซ้อนที่เรียกว่าการเจริญเติบโตแบบรุกราน[ 27 ]การกระตุ้น MET จะกระตุ้นการแบ่งเซลล์และการสร้างรูปร่าง[ 32 ] [ 33 ]
ในระหว่างการพัฒนาของตัวอ่อน การเปลี่ยนแปลงของ แผ่นเนื้อเยื่อบุผิวแบนสองชั้นไปเป็นร่างกายสามมิติขึ้นอยู่กับการเปลี่ยนผ่านของเซลล์บางส่วนจากฟีโนไทป์ของเยื่อบุผิว ไปเป็นเซลล์รูปทรงกระสวยที่มีพฤติกรรมเคลื่อนที่ ซึ่งเป็นฟีโนไทป์ของเนื้อเยื่อเกี่ยวพันกระบวนการนี้เรียกว่าการเปลี่ยนผ่านจากเยื่อบุผิวเป็นเนื้อเยื่อเกี่ยวพัน ( EMT) [ 34 ]ต่อมาในการพัฒนาของตัวอ่อน MET มีความสำคัญต่อการเกิดแกสตรู เลชัน การสร้างหลอดเลือด การอพยพของไม โอบลาสต์ การปรับโครงสร้างกระดูกและ การงอก ของเส้นประสาทเป็นต้น[ 35 ] MET มีความสำคัญต่อการเกิดตัวอ่อนเนื่องจากหนูMET −/− ตาย ในครรภ์เนื่องจากความบกพร่องอย่างรุนแรงในการพัฒนาของรก[ 36 ]ร่วมกับEctodysplasin Aพบว่ามีส่วนเกี่ยวข้องกับการแยกความแตกต่างของแผ่นเนื้อเยื่อทางกายวิภาค ซึ่งเป็นสารตั้งต้นของเกล็ด ขน และรูขุมขนในสัตว์มีกระดูกสันหลัง[ 37 ]นอกจากนี้ MET ยังจำเป็นสำหรับกระบวนการที่สำคัญ เช่น การสร้าง ตับใหม่และการสมานแผลในวัยผู้ใหญ่[ 27 ]
แกน HGF/MET ยังมีส่วนเกี่ยวข้องกับการพัฒนาของกล้ามเนื้อหัวใจด้วย mRNA ของตัวรับ HGF และ MET จะถูกแสดงออกร่วมกันในเซลล์กล้ามเนื้อหัวใจตั้งแต่ E7.5 ซึ่งเป็นช่วงเวลาไม่นานหลังจากที่หัวใจได้รับการกำหนดแล้ว ไปจนถึง E9.5 ทรานสคริปต์สำหรับลิแกนด์และตัวรับ HGF จะถูกตรวจพบครั้งแรกก่อนที่จะเกิดการเต้นและการวนของหัวใจ และคงอยู่ตลอดระยะการวน ซึ่งเป็นช่วงที่รูปร่างของหัวใจเริ่มพัฒนา[ 38 ]ในการศึกษาในนก พบว่า HGF อยู่ในชั้นกล้ามเนื้อหัวใจของช่องเอทริโอเวนทริคูลาร์ ในระยะพัฒนาการที่เกิด การเปลี่ยนแปลงจากเยื่อบุผิวเป็นเนื้อเยื่อเกี่ยวพัน (EMT) ของ หมอนรองหัวใจ[ 39 ]อย่างไรก็ตาม MET ไม่จำเป็นต่อการพัฒนาของหัวใจ เนื่องจากหนู α-MHCMet-KO แสดงการพัฒนาของหัวใจปกติ[ 40 ]
การแสดงออก
การกระจายตัวของเนื้อเยื่อ
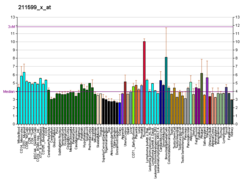
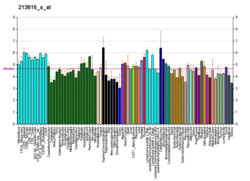
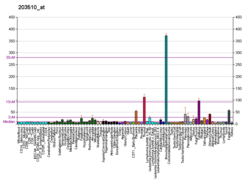
โดยปกติ MET จะแสดงออกโดยเซลล์เยื่อบุผิว[ 27 ]อย่างไรก็ตาม MET ยังพบได้ในเซลล์เยื่อบุหลอดเลือดเซลล์ประสาทเซลล์ตับ เซลล์เม็ดเลือดเซลล์เมลาโนไซต์และเซลล์กล้ามเนื้อหัวใจในทารกแรกเกิด[ 33 ] [ 41 ]การแสดงออกของ HGF ถูกจำกัดไว้เฉพาะเซลล์ที่มีต้นกำเนิดจากเนื้อเยื่อเกี่ยวพัน[ 34 ]
การควบคุมการถอดรหัส
การถอดรหัส METถูกกระตุ้นโดย HGF และปัจจัยการเจริญเติบโต หลาย ชนิด[ 42 ]โปรโมเตอร์MET มีตำแหน่งการจับที่คาดการณ์ได้สี่ตำแหน่งสำหรับEtsซึ่งเป็นตระกูลของปัจจัยการถอดรหัสที่ควบคุมยีนการเจริญเติบโตที่รุกรานหลายชนิด[ 42 ] ETS1กระตุ้นการถอดรหัสMET ในหลอดทดลอง [ 43 ] การ ถอดรหัส METถูกกระตุ้นโดยปัจจัยเหนี่ยวนำภาวะขาดออกซิเจน 1 (HIF1) ซึ่งถูกกระตุ้นโดยความเข้มข้นต่ำของออกซิเจนภายในเซลล์[ 44 ] HIF1 สามารถจับกับ องค์ประกอบการตอบสนองต่อภาวะขาดออกซิเจน (HREs) หนึ่งในหลายๆ ตัวในโปรโมเตอร์MET [ 34 ]ภาวะขาดออกซิเจนยังกระตุ้นปัจจัยการถอดรหัสAP-1ซึ่งเกี่ยวข้องกับการถอดรหัสMET [ 34 ]
ความสำคัญทางคลินิก
บทบาทในโรคมะเร็ง
วิถีการทำงานของ MET มีบทบาทสำคัญในการพัฒนาของโรคมะเร็งผ่านทาง:
- การกระตุ้นวิถีการเกิดมะเร็งที่สำคัญ ( RAS , PI3K , STAT3 , เบต้า-แคทเทนิน )
- การสร้างหลอดเลือดใหม่ (การงอกของหลอดเลือดใหม่จากหลอดเลือดที่มีอยู่เดิมเพื่อส่งสารอาหารไปเลี้ยงเนื้องอก)
- กระจาย (การแยกตัวของเซลล์เนื่องจาก การผลิต เมทัลโลโปรตีเอส ) ซึ่งมักนำไปสู่การแพร่กระจาย[ 45 ]
การควบคุมการลดระดับแบบประสานงานของทั้ง MET และตัวกระตุ้นปลายทาง extracellular signal-regulated kinase 2 (ERK2) โดยmiR-199a*อาจมีประสิทธิภาพในการยับยั้งไม่เพียงแต่การแพร่กระจายของเซลล์เท่านั้น แต่ยังรวมถึงความสามารถในการเคลื่อนที่และการรุกรานของเซลล์มะเร็ง ด้วย [ 46 ]
การขยายตัวของ MET ได้กลายเป็นตัวบ่งชี้ทางชีวภาพที่มีศักยภาพของชนิดย่อยของเนื้องอกเซลล์ใส[ 47 ]
การขยายตัวของตัวรับพื้นผิวเซลล์ MET มักทำให้เกิดความต้านทานต่อ การ บำบัดด้วยยาต้าน EGFRในมะเร็งลำไส้ใหญ่ [ 48 ]
บทบาทในภาวะออทิสติก
ฐานข้อมูล SFARIgene ระบุ MET ที่มี คะแนน ออทิสติก 2.0 ซึ่งบ่งชี้ว่าเป็นตัวเลือกที่แข็งแกร่งในการมีบทบาทในกรณีของออทิสติก ฐานข้อมูลยังระบุอย่างน้อยหนึ่งการศึกษาที่พบว่า MET มีบทบาทในกรณีของโรคจิตเภทยีนนี้ถูกเชื่อมโยงกับออทิสติกเป็นครั้งแรกในการศึกษาที่ระบุโพลีมอร์ฟิซึมในโปรโมเตอร์ของยีน MET [ 49 ]โพลีมอร์ฟิซึมลดการถอดรหัสลง 50% นอกจากนี้ ตัวแปรที่เป็นโพลีมอร์ฟิซึมความเสี่ยงต่อออทิสติกได้รับการยืนยันซ้ำ และแสดงให้เห็นว่ามีความเข้มข้นในเด็กที่เป็นออทิสติกและมีความผิดปกติของระบบทางเดินอาหาร[ 50 ]พบการกลายพันธุ์ที่หายากซึ่งปรากฏในสมาชิกครอบครัวสองคน คนหนึ่งเป็นออทิสติกและอีกคนหนึ่งมีความผิดปกติทางสังคมและการสื่อสาร[ 51 ]บทบาทของตัวรับในการพัฒนาสมองนั้นแตกต่างจากบทบาทในกระบวนการพัฒนาอื่นๆ การกระตุ้นตัวรับ MET ควบคุมการสร้างไซแนปส์[ 52 ] [ 53 ] [ 54 ] [ 55 ] [ 56 ]และสามารถส่งผลกระทบต่อการพัฒนาและการทำงานของวงจรที่เกี่ยวข้องกับพฤติกรรมทางสังคมและอารมณ์[ 57 ]
บทบาทในการทำงานของหัวใจ
ในหนูโตเต็มวัย MET จำเป็นต่อการปกป้องเซลล์กล้ามเนื้อหัวใจโดยการป้องกันความเครียดออกซิเดชันที่ เกี่ยวข้องกับอายุ การตายของเซลล์พังผืด และความผิดปกติของหัวใจ[ 40 ]นอกจากนี้ สารยับยั้ง MET เช่นcrizotinibหรือ PF-04254644 ได้รับการทดสอบโดยการรักษาระยะสั้นในแบบจำลองเซลล์และแบบจำลองก่อนคลินิก และแสดงให้เห็นว่าสามารถกระตุ้นการตายของเซลล์กล้ามเนื้อหัวใจผ่านการผลิต ROS การกระตุ้นแคสเปสการเปลี่ยนแปลงการเผาผลาญ และการปิดกั้นช่องไอออน[ 58 ] [ 59 ]
ในหัวใจที่ได้รับบาดเจ็บ แกน HGF/MET มีบทบาทสำคัญในการปกป้องหัวใจโดยการส่งเสริมผลในการอยู่รอด (ต่อต้านอะพอพโทซิสและต่อต้านออโตฟาจี ) ในเซลล์กล้ามเนื้อหัวใจ การสร้างหลอดเลือดใหม่ การยับยั้งการเกิดพังผืด สัญญาณต่อต้านการอักเสบและการปรับภูมิคุ้มกัน และการสร้างใหม่ผ่านการกระตุ้นเซลล์ต้นกำเนิดหัวใจ[ 60 ] [ 61 ]
ปฏิสัมพันธ์กับยีนยับยั้งเนื้องอก
พีทีน
PTEN (phosphatase and tensin homolog) เป็นยีนยับยั้งเนื้องอกที่เข้ารหัสโปรตีน PTEN ซึ่งมีกิจกรรมที่ขึ้นอยู่กับลิพิดและโปรตีนฟอสฟาเทส รวมถึงกิจกรรมที่ไม่ขึ้นอยู่กับฟอสฟาเทสด้วย [ 62 ] โปรตีน ฟอสฟาเทส PTENสามารถรบกวนการส่งสัญญาณ MET โดยการดีฟอสโฟรีเลต PIP 3ที่สร้างโดย PI3Kหรือไอโซฟอร์ม p52 ของ SHCการดีฟอสโฟรีเลต SHC ยับยั้งการดึงดูด อะแดปเตอร์ GRB2ไปยัง MET ที่ถูกกระตุ้น [ 30 ]
วีเอชแอล
มีหลักฐานความสัมพันธ์ระหว่างการปิดใช้งานยีนยับยั้งเนื้องอกVHL และการเพิ่มขึ้นของสัญญาณ MET ในมะเร็งเซลล์ไต (RCC) และการเปลี่ยนแปลงร้ายแรงของหัวใจ[ 63 ] [ 64 ]
การรักษาโรคมะเร็งโดยมุ่งเป้าไปที่ HGF/MET
เนื่องจากการรุกรานและการแพร่กระจายของเนื้องอกเป็นสาเหตุหลักของการเสียชีวิตในผู้ป่วยมะเร็ง การรบกวนการส่งสัญญาณของ MET จึงดูเหมือนจะเป็นแนวทางการรักษาที่มีแนวโน้มที่ดี สามารถดูรายชื่อยาที่อยู่ระหว่างการทดลองทางคลินิกในมนุษย์ที่มุ่งเป้าไปที่ HGF และ MET สำหรับการรักษามะเร็งได้ที่นี่
สารยับยั้ง MET ไคเนส
สารยับยั้งไคเนสเป็นโมเลกุลที่มีน้ำหนักโมเลกุลต่ำซึ่งป้องกัน การจับตัวของ ATPกับ MET จึงยับยั้งการถ่ายโอนฟอสเฟตของตัวรับและการดึงดูดตัวกระตุ้นปลายทาง ข้อจำกัดของสารยับยั้งไคเนส ได้แก่ ข้อเท็จจริงที่ว่ามันยับยั้งเฉพาะการกระตุ้น MET ที่ขึ้นอยู่กับไคเนสเท่านั้น และไม่มีสารยับยั้งไคเนสชนิดใดที่จำเพาะเจาะจงต่อ MET อย่างสมบูรณ์
- K252a ( Fermentek Biotechnology) เป็น อะนาล็อกของ สเตาโรสปอรีนที่แยกได้จากเชื้อรา ในดิน Nocardiopsis sp. และเป็นสารยับยั้งที่มีศักยภาพของตัวรับไทโรซีนไคเนส (RTK) ทั้งหมด ที่ความเข้มข้นระดับนาโนโมลาร์ K252a ยับยั้งการทำงานของ MET ทั้งแบบปกติและแบบกลายพันธุ์ (M1268T) [ 65 ]
- SU11274 ( SUGEN ) ยับยั้งกิจกรรมของ MET kinase และการส่งสัญญาณที่ตามมาโดยเฉพาะ SU11274 ยังเป็นตัวยับยั้งที่มีประสิทธิภาพของ MET กลายพันธุ์ M1268T และ H1112Y แต่ไม่ใช่ MET กลายพันธุ์ L1213V และ Y1248H [ 66 ]มีการแสดงให้เห็นว่า SU11274 ยับยั้งการเคลื่อนที่และการบุกรุกของเซลล์เยื่อบุผิวและเซลล์มะเร็งที่เกิดจาก HGF [ 67 ]
- PHA-665752 ( Pfizer ) ยับยั้งกิจกรรมของ MET kinase โดยเฉพาะ และได้รับการพิสูจน์แล้วว่าสามารถยับยั้งการฟอสโฟรีเลชันของ MET ทั้งแบบขึ้นอยู่กับ HGF และแบบปกติได้[ 68 ]นอกจากนี้ เนื้องอกบางชนิดที่มี การขยายตัวของ METยังมีความไวต่อการรักษาด้วย PHA-665752 สูง[ 69 ]
- ทิแวนตินิบ (ArQule) เป็นสารยับยั้ง MET แบบเลือกจำเพาะที่มีศักยภาพสูง ซึ่งเข้าสู่การทดลองทางคลินิกระยะที่ 2 ในปี 2551 (แต่ล้มเหลวในระยะที่ 3 ในปี 2560)
- ฟอเรตินิบ (XL880, Exelixis) ออกฤทธิ์ต่อตัวรับไทโรซีนไคเนส (RTK) หลายชนิดที่มีคุณสมบัติส่งเสริมการเจริญเติบโตและการสร้างหลอดเลือดใหม่ เป้าหมายหลักของฟอเรตินิบ ได้แก่ MET, VEGFR2และKDRฟอเรตินิบได้เสร็จสิ้นการทดลองทางคลินิกระยะที่ 2 โดยมีข้อบ่งชี้สำหรับการรักษามะเร็งไตชนิดพาพิลลารี มะเร็งกระเพาะ อาหาร และมะเร็ง ศีรษะ และลำคอ
- SGX523 (SGX Pharmaceuticals) ยับยั้ง MET โดยเฉพาะที่ความเข้มข้นระดับนาโนโมลาร์ต่ำ
- MP470 (SuperGen) เป็นสารยับยั้งตัวใหม่ของc-KIT , MET, PDGFR , Flt3 และAXLการทดลองทางคลินิกระยะที่ 1 ของ MP470 ได้รับการประกาศในปี 2550
- Vebreltinibได้รับการอนุมัติในประเทศจีนสำหรับการรักษาโรคมะเร็งปอดชนิดไม่ใช่เซลล์ขนาดเล็ก[ 70 ]
สารยับยั้ง HGF
เนื่องจาก HGF เป็นลิแกนด์เพียงชนิดเดียวที่รู้จักของ MET การปิดกั้นการสร้างสารเชิงซ้อน HGF:MET จึงปิดกั้นการทำงานทางชีวภาพ ของ MET ด้วยเหตุนี้ จึงมีการใช้ HGF ที่ถูกตัดทอน แอนติบอดีต้าน HGF และ HGF ในรูปแบบที่ไม่สามารถถูกตัดได้ ข้อจำกัดที่สำคัญของสารยับยั้ง HGF คือ พวกมันสามารถปิดกั้นเฉพาะการกระตุ้น MET ที่ขึ้นอยู่กับ HGF เท่านั้น
- NK4 แข่งขันกับ HGF เนื่องจากมันจับกับ MET โดยไม่กระตุ้นการทำงานของตัวรับ จึงทำหน้าที่เป็นตัวต้าน อย่างสมบูรณ์ NK4 เป็นโมเลกุลที่มีแฮร์พินที่ปลาย N และโดเมน kringle สี่โดเมน ของ HGF ยิ่งไปกว่านั้น NK4 ยังมีโครงสร้างคล้ายกับแอนจิโอสแตตินซึ่งเป็นเหตุผลว่าทำไมมันจึงมีฤทธิ์ต้านการสร้างหลอดเลือดใหม่[ 71 ]
- แอนติบอดีต่อต้าน HGF ที่ทำให้เป็นกลางได้รับการทดสอบเบื้องต้นในรูปแบบผสมผสาน และพบว่าจำเป็นต้อง ใช้ แอนติบอดี อย่างน้อยสามตัวที่ออกฤทธิ์บน อีพิโทป HGF ที่แตกต่างกัน เพื่อป้องกันการกระตุ้นไทโรซีนไคเนส MET [ 72 ]เมื่อไม่นานมานี้ ได้มีการแสดงให้เห็นว่าแอนติบอดีโมโนโคลนอล ของมนุษย์อย่างสมบูรณ์ สามารถจับและทำให้ HGF ของมนุษย์เป็นกลางได้ ซึ่งนำไปสู่การถดถอยของเนื้องอกในแบบจำลองหนู[ 73 ]ปัจจุบันมีแอนติบอดีต่อต้าน HGF สองชนิด ได้แก่ AV299 ที่เป็นมนุษย์ (AVEO) และ AMG102 ที่เป็นมนุษย์อย่างสมบูรณ์ ( Amgen )
- HGF ที่ไม่สามารถถูกตัดได้ (Uncleavable HGF) เป็นรูปแบบที่ถูกดัดแปลงของโปร-HGF โดยมีการเปลี่ยนแปลงกรดอะมิโนเพียงตัวเดียว ซึ่งป้องกันการเจริญเติบโตของโมเลกุล Uncleavable HGF สามารถยับยั้งการตอบสนองทางชีวภาพที่เกิดจาก MET โดยการจับกับ MET ด้วยความสัมพันธ์สูงและแทนที่ HGF ที่เจริญเต็มที่แล้ว ยิ่งไปกว่านั้น Uncleavable HGF ยังแข่งขันกับโปร-HGF ชนิดปกติในร่างกายเพื่อแย่งจับกับโดเมนเร่งปฏิกิริยาของเอนไซม์โปร ตีเอส ที่ทำหน้าที่ตัดสารตั้งต้นของ HGF การแสดงออกของ Uncleavable HGF ทั้งในระดับเฉพาะที่และทั่วร่างกายจะยับยั้งการเจริญเติบโตของเนื้องอก และที่สำคัญกว่านั้นคือป้องกันการแพร่กระจายของมะเร็ง
ล่อ MET
Decoy MET หมายถึงตัวรับ MET ที่ถูกตัดทอนซึ่งละลายได้ Decoy สามารถยับยั้งการกระตุ้น MET ที่เกิดจากกลไกที่ขึ้นอยู่กับ HGF และกลไกที่ไม่ขึ้นอยู่กับ HGF ได้ เนื่องจาก Decoy ป้องกันทั้งการจับกับลิแกนด์และการเกิดโฮโมไดเมอไรเซชันของตัวรับ MET CGEN241 ( Compugen ) เป็น Decoy MET ที่มีประสิทธิภาพสูงในการยับยั้งการเติบโตของเนื้องอกและป้องกันการแพร่กระจายในแบบจำลองสัตว์[ 74 ]
ภูมิคุ้มกันบำบัดที่มุ่งเป้าไปที่ MET
ยาที่ใช้ในการบำบัดด้วยภูมิคุ้มกันสามารถออกฤทธิ์ได้ทั้งแบบพาสซีฟโดยการเพิ่มการตอบสนองทางภูมิคุ้มกันต่อเซลล์มะเร็งที่แสดงออก MET หรือแบบแอคทีฟโดยการกระตุ้นเซลล์ภูมิคุ้มกันและเปลี่ยนแปลงการแยกตัว/การเจริญเติบโตของเซลล์มะเร็ง[ 75 ]
ภูมิคุ้มกันบำบัดแบบพาสซีฟ
การให้โมโนโคลนอลแอนติบอดี (mAbs) เป็นรูปแบบหนึ่งของภูมิคุ้มกันบำบัดแบบพาสซีฟ MAbs ช่วยให้เซลล์มะเร็งถูกทำลายโดยการทำลายเซลล์โดยอาศัยคอมพลีเมนต์ (CDC) และการทำลายเซลล์โดยอาศัยเซลล์ ( ADCC ) ใน CDC mAbs จะจับกับแอนติเจน จำเพาะ ทำให้เกิดการกระตุ้นของคอมพลีเมนต์แคสเคดซึ่งจะนำไปสู่การสร้างรูพรุนในเซลล์มะเร็ง ใน ADCC โดเมน Fab ของ mAb จะจับกับแอนติเจนของเนื้องอกและโดเมน Fc จะจับกับตัวรับ Fc ที่มีอยู่บนเซลล์เอฟเฟกเตอร์ ( เซลล์ฟาโกไซต์และเซลล์ NK ) จึงสร้างสะพานเชื่อมระหว่างเซลล์เอฟเฟกเตอร์และเซลล์เป้าหมาย สิ่งนี้กระตุ้นการทำงานของเซลล์เอฟเฟกเตอร์ นำไปสู่การกลืนกินเซลล์มะเร็งโดยนิวโทรฟิลและแมโครฟาจนอกจากนี้เซลล์ NKยังปล่อย โมเลกุล ที่เป็นพิษต่อเซลล์ซึ่งจะทำลายเซลล์มะเร็ง[ 75 ]
- DN30 เป็นแอนติบอดีโมโนโคลนอลต่อต้าน MET ที่รู้จักส่วนนอกเซลล์ของ MET DN30 กระตุ้นทั้งการหลุดออก ของ เอ็กโทโดเมนของ MET และการตัดส่วนภายในเซลล์ ซึ่งจะถูกย่อยสลายโดย กลไก โปรตีเอโซมตามลำดับ ผลที่ตามมาคือ MET จะถูกทำให้ไม่ทำงานในด้านหนึ่ง และในอีกด้านหนึ่ง ส่วนที่หลุดออกของ MET นอกเซลล์จะขัดขวางการกระตุ้นตัวรับ MET อื่นๆ โดยทำหน้าที่เป็นตัวล่อ DN30 ยับยั้งการเจริญเติบโตของเนื้องอกและป้องกันการแพร่กระจายในแบบจำลองสัตว์[ 76 ]
- OA-5D5 เป็นแอนติบอดีโมโนโคลนอลต่อต้าน MET แบบแขนเดียว ซึ่งได้รับการพิสูจน์แล้วว่าสามารถยับยั้งการเจริญเติบโตของเนื้องอกตับอ่อน แบบออร์โธโทปิก [ 77 ]และเนื้องอกสมองกลิโอบลาสโตมา[ 78 ] และช่วยเพิ่มอัตราการรอดชีวิตใน แบบจำลองเนื้องอกซีโนกราฟต์ OA-5D5 ผลิตเป็นโปรตีนรีคอมบิแนนท์ในEscherichia coliประกอบด้วยโดเมนตัวแปร ของหนู สำหรับ โซ่ หนักและโซ่เบาพร้อมด้วยโดเมนคงที่ IgG1 ของมนุษย์ แอนติบอดีนี้จะปิดกั้นการจับกันของ HGF กับ MET ในลักษณะการแข่งขัน
การรักษาด้วยภูมิคุ้มกันแบบแอคทีฟ
การบำบัดภูมิคุ้มกันแบบแอคทีฟต่อเนื้องอกที่แสดงออก MET สามารถทำได้โดยการให้ไซโตไคน์เช่นอินเตอร์เฟอรอน (IFN) และอินเตอร์ลิวคิน ( IL-2 ) ซึ่งกระตุ้นการทำงานของเซลล์ภูมิคุ้มกันจำนวนมากแบบไม่จำเพาะเจาะจง IFN ได้รับการทดสอบเป็นวิธีการรักษาสำหรับมะเร็งหลายชนิดและแสดงให้เห็นถึงประโยชน์ในการรักษา IL-2 ได้รับการอนุมัติจากสำนักงานคณะกรรมการอาหารและยาของสหรัฐอเมริกา (FDA) สำหรับการรักษามะเร็งเซลล์ไตและมะเร็งผิวหนังชนิดเมลาโนมาที่แพร่กระจาย ซึ่งมักมีการทำงานของ MET ที่ผิดปกติ[ 75 ]
ปฏิสัมพันธ์
มีการค้นพบว่า Met มีปฏิกิริยากับ:
ดูเพิ่มเติม
อ่านเพิ่มเติม
- Peruzzi B, Bottaro DP (2006). "การกำหนดเป้าหมายเส้นทางการส่งสัญญาณ c-Met ในมะเร็ง" . Clin. Cancer Res . 12 (12): 3657– 60. doi : 10.1158/1078-0432.CCR-06-0818 . PMID 16778093 .
- Birchmeier C, Birchmeier W, Gherardi E, Vande Woude GF (ธันวาคม 2003). "Met, metastasis, motility and more". Nat. Rev. Mol. Cell Biol . 4 (12): 915– 25. doi : 10.1038/nrm1261 . PMID 14685170 . S2CID 19330786 .
- Zhang YW, Vande Woude GF (กุมภาพันธ์ 2546). "การส่งสัญญาณ HGF/SF-met ในการควบคุมการสร้างกิ่งก้านสาขาและการรุกราน". J. Cell. Biochem . 88 (2): 408– 17. doi : 10.1002/jcb.10358 . PMID 12520544. S2CID 13212355 .
- Paumelle R, Tulasne D, Kherrouche Z, Plaza S, Leroy C, Reveneau S และคณะ (เมษายน 2545) "ปัจจัยการเจริญเติบโตของเซลล์ตับ/ปัจจัยกระจายตัวกระตุ้นปัจจัยการถอดรหัส ETS1 โดยผ่านเส้นทางการส่งสัญญาณ RAS-RAF-MEK-ERK" Oncogene . 21 (15): 2309– 19. doi : 10.1038/sj.onc.1205297 . PMID 11948414 . S2CID 22371025 .
- Comoglio PM (1993). "โครงสร้าง การสังเคราะห์ทางชีวภาพ และคุณสมบัติทางชีวเคมีของตัวรับ HGF ในเซลล์ปกติและเซลล์มะเร็ง" EXS . 65 : 131– 65. PMID 8380735 .
- Maulik G, Shrikhande A, Kijima T, Ma PC, Morrison PT, Salgia R (2002). "บทบาทของตัวรับปัจจัยการเจริญเติบโตของเซลล์ตับ c-Met ในการเกิดมะเร็งและศักยภาพในการยับยั้งการรักษา" Cytokine Growth Factor Rev . 13 (1): 41– 59. doi : 10.1016/S1359-6101(01)00029-6 . PMID 11750879 .
- Ma PC, Maulik G, Christensen J, Salgia R (2003). "c-Met: โครงสร้าง หน้าที่ และศักยภาพในการยับยั้งการรักษา" Cancer Metastasis Rev . 22 (4): 309– 25. doi : 10.1023/A:1023768811842 . PMID 12884908 . S2CID 23542507 .
- Knudsen BS, Edlund M (2004). "มะเร็งต่อมลูกหมากและตัวรับปัจจัยการเจริญเติบโตของเซลล์ตับ" . Adv. Cancer Res . Advances in Cancer Research. 91 : 31–67 . doi : 10.1016/S0065-230X(04)91002-0 . ISBN 978-0-12-006691-9. PMID 15327888 .
- Dharmawardana PG, Giubellino A, Bottaro DP (2004). "มะเร็งไตชนิด papillary ที่ถ่ายทอดทางพันธุกรรมประเภท I" . Curr. Mol. Med . 4 (8): 855– 68. doi : 10.2174/1566524043359674 . PMID 15579033 . S2CID 10928725 .
- Kemp LE, Mulloy B, Gherardi E (2006). "การส่งสัญญาณโดย HGF/SF และ Met: บทบาทของตัวรับร่วมเฮปารานซัลเฟต" Biochem. Soc. Trans . 34 (Pt 3): 414–7 . doi : 10.1042/BST0340414 . PMID 16709175 .
ลิงก์ภายนอก
- Proto-Oncogene+Proteins+c-met ที่ หัวข้อทางการ แพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
- ข้อมูลใน UniProtKB/Swiss-Prot หมายเลข P08581: MET_HUMANจากเซิร์ฟเวอร์โปรตีโอมิกส์ ExPASy (Expert Protein Analysis System) ของสถาบันชีวสารสนเทศแห่งสวิตเซอร์แลนด์ (SIB)
- ตาราง แสดงข้อมูลอ้างอิงเกี่ยว กับบทบาทสำคัญของ MET ในโรคมะเร็ง
- หน้าแสดงตำแหน่งจีโนม METของมนุษย์และ รายละเอียดเกี่ยวกับยีน METในUCSC Genome Browser
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ตัวรับปัจจัยการเจริญเติบโตของเซลล์ตับ
ตัวรับปัจจัยการเจริญเติบโตของเซลล์ตับ ( ตัวรับ HGF ) [ 5 ] [ 6 ] เป็น โปรตีน ที่ในมนุษย์ถูกเข้ารหัสโดย ยีน MET โปรตีนนี้มีกิจกรรม ไทโรซีนไคเนส [ 7 ]...
ยีน
โปรโตออนโคยีน MET ( GeneID: 4233 ) มีความยาวทั้งหมด 125,982 bp และตั้งอยู่ในตำแหน่ง 7q31 ของโครโมโซม 7 [ 12 ] MET ถูกถอดรหัสเป็น mRNA ที่สมบูรณ์ขนาด 6,641 bp จากนั้นจึงแปลเป็นโปรตีน MET ที่มีกรดอะมิโน 1,390 ตัว
โปรตีน
MET เป็น ตัวรับไทโรซีนไคเนส (RTK) ที่ผลิตเป็นสารตั้งต้นแบบสายเดี่ยว สารตั้งต้นจะถูกตัดด้วยเอนไซม์โปรตีเอส ที่ตำแหน่ง ฟูริน เพื่อให้ได้หน่วยย่อยอัลฟาภายนอกเซลล์ที่มีการเติมหมู่ไกลโคซิลจำนวนมากและหน่วยย่อยเบตาที่อยู่ภายในเยื่อหุ้มเซลล์ ซึ่งเชื่อมต่อกันด้วย...
ภายนอกเซลล์
บริเวณที่มีความคล้ายคลึงกับ เซมาฟอริน (โดเมนเซมา) ซึ่งรวมถึงสายอัลฟาแบบเต็มและส่วนปลายด้าน N ของสายเบตา ลำดับที่เกี่ยวข้องกับ MET ที่อุดมไปด้วยซิสเทอีน (โดเมน MRS) ส่วนที่ซ้ำกันซึ่งอุดมไปด้วยไกลซีนและโพรลีน (GP repeats)...