อ่าน 11 นาที
เซลล์ทีที่เป็นพิษต่อเซลล์
เซลล์ T นักฆ่า(เรียกอีกอย่างว่า T C , เซลล์ T ลิมโฟไซต์ที่เป็นพิษต่อเซลล์ , CTL , เซลล์ T นักฆ่า , เซลล์ T ที่ทำลายเซลล์ , เซลล์ T CD8 + หรือ cd8 ) คือ เซลล์ T ลิมโฟไซต์ (...
เซลล์ทีที่เป็นพิษต่อเซลล์

เซลล์ Tนักฆ่า(เรียกอีกอย่างว่าT C , เซลล์ T ลิมโฟไซต์ที่เป็นพิษต่อเซลล์ , CTL , เซลล์ T นักฆ่า , เซลล์ T ที่ทำลายเซลล์ , เซลล์ T CD8 +หรือcd8 ) คือเซลล์ T ลิมโฟไซต์ ( เม็ดเลือดขาวชนิดหนึ่ง) ที่ฆ่า เซลล์ มะเร็งเซลล์ที่ติดเชื้อจากเชื้อโรค ภายในเซลล์ เช่นไวรัสหรือแบคทีเรียหรือเซลล์ที่เสียหายในรูปแบบอื่นๆ[ 1 ]
เซลล์ทีชนิดทำลายเซลล์ส่วนใหญ่มีตัวรับเซลล์ที (TCR) ที่สามารถจดจำแอนติเจน จำเพาะได้ แอนติเจนคือโมเลกุลที่สามารถกระตุ้นการตอบสนองทางภูมิคุ้มกันและมักผลิตโดยเซลล์มะเร็งไวรัส แบคทีเรีย หรือสัญญาณภายในเซลล์ แอนติเจนภายในเซลล์จะจับกับ โมเลกุล MHC คลาส Iและถูกนำไปยังพื้นผิวของเซลล์โดยโมเลกุล MHC คลาส I ซึ่งเซลล์ทีสามารถจดจำได้ หาก TCR จำเพาะต่อแอนติเจนนั้น มันจะจับกับสารประกอบของโมเลกุล MHC คลาส I และแอนติเจน และเซลล์ทีจะทำลายเซลล์นั้น
เพื่อให้ TCR สามารถจับกับโมเลกุล MHC คลาส I ได้นั้น TCR จะต้องมีไกลโคโปรตีนที่เรียกว่าCD8 อยู่ด้วย ซึ่งจะจับกับส่วนคงที่ของโมเลกุล MHC คลาส I ดังนั้น เซลล์ T เหล่านี้จึงเรียกว่าเซลล์ T CD8 +
ความสัมพันธ์ระหว่าง CD8 และโมเลกุล MHC ทำให้เซลล์ T Cและเซลล์เป้าหมายยึดติดกันอย่างใกล้ชิดในระหว่างการกระตุ้นด้วยแอนติเจนจำเพาะ เซลล์ T ที่มี CD8 +จะถูกจำแนกเป็นเซลล์ T Cเมื่อถูกกระตุ้น และโดยทั่วไปจะถูกจัดประเภทว่ามีบทบาทในการทำลายเซลล์เป้าหมายตามที่กำหนดไว้ล่วงหน้าภายในระบบภูมิคุ้มกัน อย่างไรก็ตาม เซลล์ T ที่มี CD8 +ยังมีความสามารถในการสร้างไซโตไคน์ บางชนิด เช่นTNF-αและIFN-γซึ่งมีฤทธิ์ต้านมะเร็งและต้านจุลชีพ
การพัฒนา
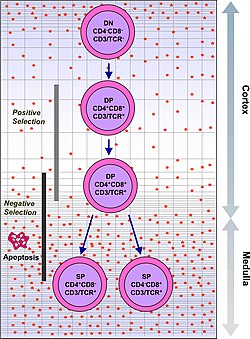
ระบบภูมิคุ้มกันต้องจดจำแอนติเจนที่มีศักยภาพมากกว่าหนึ่งพันล้านรายการ ร่างกายมนุษย์มียีนน้อยกว่า 30,000 ยีน ดังนั้นจึงเป็นไปไม่ได้ที่จะมียีนหนึ่งตัวสำหรับแอนติเจนทุกตัว ในทางกลับกัน DNA ในเซลล์เม็ดเลือดขาวหลายล้านเซลล์ในไขกระดูกจะถูกสับเปลี่ยนเพื่อสร้างเซลล์ที่มีตัวรับเฉพาะ ซึ่งแต่ละตัวรับสามารถจับกับแอนติเจนที่แตกต่างกันได้ ตัวรับบางตัวจับกับเนื้อเยื่อในร่างกายมนุษย์เอง ดังนั้นเพื่อป้องกันไม่ให้ร่างกายโจมตีตัวเอง เซลล์เม็ดเลือดขาวที่ทำปฏิกิริยากับตัวเองเหล่านั้นจะถูกทำลายในระหว่างการพัฒนาต่อไปในต่อมไทมัสซึ่งไอโอดีนมีความจำเป็นต่อการพัฒนาและการทำงานของมัน[ 2 ]
TCR ประกอบด้วยสองส่วน โดยปกติจะเป็นโซ่แอลฟาและโซ่เบตา (TCR บางชนิดมีโซ่แกมมาและโซ่เดลตา พวกมันมีหน้าที่ต่อต้านความเครียดและเป็นส่วนหนึ่งของสิ่งกีดขวางเยื่อบุผิว[ 3 ] ) เซลล์ต้นกำเนิดเม็ดเลือดในไขกระดูกจะอพยพเข้าไปในต่อมไทมัสซึ่งพวกมันจะผ่านกระบวนการV(D)J recombination ของ DNA TCRโซ่เบตาเพื่อสร้างโปรตีน TCR ในรูปแบบพัฒนาการที่เรียกว่า pre-TCR หากการจัดเรียงใหม่นั้นประสบความสำเร็จ เซลล์ก็จะจัดเรียง DNA TCR โซ่แอลฟาใหม่เพื่อสร้างคอมเพล็กซ์ TCR แอลฟา-เบตาที่ใช้งานได้ ผลิตภัณฑ์การจัดเรียงทางพันธุกรรมที่มีความแปรปรวนสูงในยีน TCR นี้ช่วยสร้างเซลล์ T ที่แตกต่างกันหลายล้านเซลล์ที่มี TCR ที่แตกต่างกัน ช่วยให้ระบบภูมิคุ้มกันของร่างกายตอบสนองต่อโปรตีนของผู้บุกรุก ได้แทบทุกชนิด เซลล์ Tส่วนใหญ่แสดงออกถึง TCR อัลฟา-เบตา (เซลล์ T αβ) แต่เซลล์ T บางส่วนในเนื้อเยื่อเยื่อบุผิว (เช่น ลำไส้) แสดงออกถึง TCR แกมมา-เดลตา ( เซลล์ T แกมมาเดลตา ) ซึ่งรู้จักแอนติเจนที่ไม่ใช่โปรตีน เซลล์เหล่านี้มีลักษณะเฉพาะคือความสามารถในการจดจำแอนติเจนที่ไม่ได้ถูกนำเสนอ นอกจากนี้ พวกมันยังสามารถจดจำโปรตีนช็อกพิษของจุลินทรีย์และโปรตีนความเครียดของเซลล์ตัวเองได้[ 4 ]เซลล์ T γδ มีความยืดหยุ่นในการทำงานอย่างกว้างขวางหลังจากจดจำเซลล์ที่ติดเชื้อหรือเปลี่ยนแปลงไป เนื่องจากพวกมันสามารถผลิตไซโตไคน์ (IFN-γ, TNF-α, IL-17) และเคโมไคน์ (IP-10, ลิมโฟแทกติน) กระตุ้นการสลายเซลล์เป้าหมาย (เพอร์ฟอริน, แกรนไซม์...) และโต้ตอบกับเซลล์อื่นๆ เช่น เซลล์เยื่อบุผิว โมโนไซต์ เซลล์เดนดริติก นิวโทรฟิล และเซลล์ B ในการติดเชื้อบางชนิด เช่นไซโตเมกาไวรัสของมนุษย์จะมีการขยายตัวแบบโคลนของเซลล์ γδ T ในส่วนปลายที่มี TCR เฉพาะ ซึ่งบ่งชี้ถึงลักษณะการปรับตัวของการตอบสนองทางภูมิคุ้มกันที่เกิดจากเซลล์เหล่านี้[ 5 ]
เซลล์ T ที่มี TCR ที่เสถียรในเชิงฟังก์ชันจะแสดงออกทั้งตัวรับร่วมCD4และCD8 ดังนั้นจึงเรียกว่าเซลล์ T "ดับเบิลโพซิทีฟ" (DP) (CD4 + CD8 + ) เซลล์ T ดับเบิลโพซิทีฟจะสัมผัสกับแอนติเจนของตัวเองที่หลากหลายในต่อมไทมัสและผ่านเกณฑ์การคัดเลือกสองเกณฑ์:
- การคัดเลือกเชิงบวก (Positive selection ) คือการคัดเลือกเซลล์ T ที่มีคุณสมบัติเป็นบวกทั้งสองอย่าง (double-positive T cells) ซึ่งสามารถจับกับแอนติเจนจากภายนอกได้ในขณะที่มี MHC ของตัวเองอยู่ด้วย เซลล์เหล่านี้จะพัฒนาไปเป็น CD4 +หรือ CD8 +ขึ้นอยู่กับว่า MHC ใดเกี่ยวข้องกับแอนติเจนที่นำเสนอ (MHC1 สำหรับ CD8, MHC2 สำหรับ CD4) ในกรณีนี้ เซลล์จะได้รับแอนติเจนในบริบทของ MHC1 การคัดเลือกเชิงบวกหมายถึงการเลือก TCR ที่สามารถจดจำโมเลกุล MHC ของตัวเองได้
- การคัดเลือกเชิงลบคือกระบวนการที่เซลล์ T ที่มีคุณสมบัติเป็นบวกสองเท่าซึ่งจับกับแอนติเจนของตนเองที่นำเสนอโดยMHC อย่างแน่นหนาเกินไปจะ เกิด ภาวะอะพอพโทซิสเนื่องจากมิเช่นนั้นอาจกลายเป็นเซลล์ที่ตอบสนองต่อตนเอง นำไปสู่โรคภูมิต้านตนเองได้
เฉพาะเซลล์ T ที่จับกับคอมเพล็กซ์ MHC-self-antigen อย่างอ่อนๆ เท่านั้นที่จะถูกคัดเลือกในเชิงบวก เซลล์เหล่านั้นที่รอดพ้นจากการคัดเลือกทั้งในเชิงบวกและเชิงลบจะพัฒนาไปเป็นเซลล์ T ที่มีตัวรับเดี่ยว (CD4 +หรือ CD8 + ) ขึ้นอยู่กับว่า TCR ของพวกมันจดจำแอนติเจนที่นำเสนอโดย MHC คลาส I (CD8) หรือ แอนติเจนที่นำเสนอโดย MHC คลาส II (CD4) เซลล์ T ที่มี CD8 + เหล่านี้ จะเจริญเติบโตเต็มที่และกลายเป็นเซลล์ T ที่มีฤทธิ์ทำลายเซลล์เป้าหมายหลังจากได้รับการกระตุ้นด้วยแอนติเจนที่จำกัดโดยคลาส I
การเปิดใช้งาน
เซลล์ T ที่โตเต็มวัยจะผ่านขั้นตอนต่างๆ ขึ้นอยู่กับจำนวนครั้งที่สัมผัสกับแอนติเจน ในขั้นแรก เซลล์ T ที่ยังไม่ได้รับการกระตุ้น (เซลล์ T บริสุทธิ์) เป็นระยะเริ่มต้นของเซลล์ T ที่อยู่ในต่อมไทมัส ซึ่งยังไม่เคยพบกับแอนติเจนที่มีความสัมพันธ์กับ TCR มาก่อน เซลล์ T ที่เคยสัมผัสกับแอนติเจนอย่างน้อยหนึ่งครั้ง แต่ต่อมากลับสู่สภาวะสงบหรือไม่ทำงาน เรียกว่าเซลล์ T หน่วยความจำ เซลล์ระยะนี้พร้อมที่จะตอบสนองต่อแอนติเจนจำเพาะที่กระตุ้นอีกครั้ง สุดท้าย เมื่อมีการกระตุ้นการตอบสนองทางภูมิคุ้มกัน เซลล์ T ที่ยังไม่ได้รับการกระตุ้นและเซลล์ T หน่วยความจำเหล่านี้จะถูกกระตุ้น ทำให้เกิดเซลล์ T ตัวทำหน้าที่ที่มีความสามารถในการฆ่าเชื้อโรคหรือเซลล์มะเร็ง[ 6 ] [ 7 ]
ระดับการกระตุ้นของเซลล์เหล่านี้สูงมาก และกระบวนการนี้สามารถเกิดขึ้นได้สองทาง คือ แบบไม่ขึ้นกับต่อมไทมัส (โดยAPC ที่ติดเชื้อ ) หรือแบบขึ้นกับต่อมไทมัส (โดยเซลล์ T CD4+ ) ในเส้นทางที่ไม่ขึ้นกับต่อมไทมัส เนื่องจาก APC ติดเชื้อ จึงมีการกระตุ้นสูงและแสดงตัวรับร่วมจำนวนมากสำหรับการกระตุ้นร่วม หาก APC ไม่ติดเชื้อ เซลล์ CD4 จะต้องเข้ามาเกี่ยวข้อง ไม่ว่าจะเพื่อกระตุ้น APC โดยการกระตุ้นร่วม (พบได้บ่อยกว่า) หรือเพื่อกระตุ้นเซลล์ Tc โดยตรงโดยการหลั่งIL- 2
หากเกิดการกระตุ้น ลิมโฟไซต์จะจัดเรียงเม็ดเล็กๆ ไปทางบริเวณไซแนปส์และปล่อยออกมา ทำให้เกิด "การโจมตีที่ทำให้ถึงแก่ชีวิต" ณ จุดนี้ มันจะแยกตัวออกจากเซลล์เป้าหมาย และสามารถเคลื่อนไปยังเซลล์อื่นและเซลล์อื่นต่อไปได้ เซลล์เป้าหมายจะตายภายในเวลาประมาณ 6 ชั่วโมง โดยปกติแล้วด้วยกระบวนการอะพอพโทซิส[ 8 ]
MHC คลาส I พบได้ใน เซลล์ เจ้าบ้าน ทุก เซลล์ ยกเว้นเซลล์ที่ไม่มีนิวเคลียสเช่นเม็ดเลือดแดงเมื่อเซลล์เหล่านี้ติดเชื้อจากเชื้อก่อโรคภายใน เซลล์ เซลล์จะย่อยสลายโปรตีนแปลกปลอมผ่านกระบวนการประมวลผลแอนติเจนส่งผลให้เกิดชิ้นส่วนเปปไทด์ ซึ่งบางส่วนจะถูกนำเสนอโดย MHC คลาส I ไปยังตัวรับแอนติเจนของเซลล์ T (TCR) บน เซลล์ T ชนิด CD8 +
การกระตุ้นเซลล์ทีชนิดทำลายเซลล์เป้าหมายนั้นขึ้นอยู่กับปฏิกิริยาพร้อมกันหลายอย่างระหว่างโมเลกุลที่แสดงออกบนพื้นผิวของเซลล์ทีและโมเลกุลบนพื้นผิวของเซลล์นำเสนอแอนติเจน (APC) ตัวอย่างเช่น พิจารณาแบบจำลองสัญญาณสองแบบสำหรับการกระตุ้นเซลล์ ที ซี
| สัญญาณ | เซลล์ที | เอพีซี | คำอธิบาย |
|---|---|---|---|
| สัญญาณแรก | ทีซีอาร์ | โมเลกุลMHC คลาส Iที่จับกับเปปไทด์ | มีการปฏิสัมพันธ์ครั้งที่สองระหว่าง ตัวรับร่วม CD8และโมเลกุล MHC คลาส I เพื่อทำให้สัญญาณนี้มีความเสถียร |
| สัญญาณที่สอง | โมเลกุล CD28บนเซลล์ T | CD80หรือCD86 ( เรียกอีกอย่างว่า B7-1 และ B7-2) | CD80 และ CD86 เป็นที่รู้จักกันในชื่อโคสติมูเลเตอร์สำหรับการกระตุ้นเซลล์ T สัญญาณที่สองนี้สามารถเสริม (หรือทดแทน) ได้โดยการกระตุ้นเซลล์ T Cด้วยไซโตไคน์ที่ปล่อยออกมาจากเซลล์ T ตัวช่วย |
การกระตุ้นเซลล์ T CD8 + ที่ยังไม่ได้รับการกระตุ้นอย่างง่าย ๆ จำเป็นต้องมีการโต้ตอบกับเซลล์นำเสนอแอนติเจนแบบมืออาชีพ โดยเฉพาะอย่างยิ่งเซลล์เดนดริติก ที่เจริญเต็มที่ เพื่อสร้างเซลล์ T หน่วยความจำ ที่คงอยู่ยาวนาน และเพื่อให้สามารถกระตุ้นเซลล์ T ที่เป็นพิษต่อเซลล์ซ้ำ ๆ ได้ เซลล์เดนดริติกจะต้องโต้ตอบกับทั้งเซลล์ T ตัวช่วย CD4 + ที่ถูกกระตุ้น และเซลล์ T CD8 + [ 9 ] [ 7 ]ในระหว่างกระบวนการนี้ เซลล์ T ตัวช่วย CD4 +จะ "อนุญาต" ให้เซลล์เดนดริติกส่งสัญญาณกระตุ้นที่มีประสิทธิภาพไปยังเซลล์ T CD8 + ที่ยังไม่ได้รับการกระตุ้น [ 10 ]การอนุญาตเซลล์นำเสนอแอนติเจนโดยเซลล์ T ตัวช่วย CD4 + นี้ เกิดขึ้นโดยการส่งสัญญาณระหว่าง CD154/CD40L บนเซลล์ T ตัวช่วยและตัวรับ CD40 บนเซลล์นำเสนอแอนติเจนในระหว่างการสร้างไซแนปส์ทางภูมิคุ้มกัน[ 11 ]
ในขณะที่ในกรณีส่วนใหญ่ การกระตุ้นจะขึ้นอยู่กับการรับรู้แอนติเจนของ TCR แต่ก็มีการอธิบายเส้นทางการกระตุ้นทางเลือกอื่นๆ ไว้แล้ว ตัวอย่างเช่น พบว่าเซลล์ T ที่เป็นพิษต่อเซลล์จะถูกกระตุ้นเมื่อถูกกำหนดเป้าหมายโดยเซลล์ T CD8 อื่นๆ ซึ่งนำไปสู่การสร้างความทนทานต่อเซลล์ T CD8 เหล่านั้น[ 12 ]
เมื่อถูกกระตุ้นแล้ว เซลล์ T Cจะเกิดการเพิ่มจำนวนแบบโคลนโดยอาศัยไซโตไคน์อินเตอร์ลิวคิน 2 (IL-2) ซึ่งเป็นปัจจัยการเจริญเติบโตและการเปลี่ยนแปลงรูปร่างของเซลล์ T การเพิ่มจำนวนนี้จะทำให้จำนวนเซลล์ที่จำเพาะต่อแอนติเจนเป้าหมายเพิ่มขึ้น จากนั้นเซลล์เหล่านี้จะสามารถเดินทางไปทั่วร่างกายเพื่อค้นหาเซลล์ร่างกายที่ มีแอนติเจนเป็นบวก ได้
หน้าที่ของตัวกระตุ้น
เมื่อสัมผัสกับเซลล์ร่างกายที่ติดเชื้อ/ทำงานผิดปกติ เซลล์ T Cจะปล่อยสารพิษไซโตท็อกซิน ได้แก่ เพอร์ ฟอ รินแกรนไซม์และแกรนูลิซินโดยผ่านการทำงานของเพอร์ฟอริน แกรนไซม์จะเข้าสู่ไซโตพลาสซึมของเซลล์เป้าหมาย และการทำงาน ของ เซรินโปรตีเอส จะกระตุ้นแค สเปสแคสเคด ซึ่งเป็นชุดของซิสเทอีนโปรตีเอสที่จะนำไปสู่อะพอพโทซิส (การตายของเซลล์ตามโปรแกรม) ในที่สุด สิ่งนี้เรียกว่า "การโจมตีที่ทำให้ถึงแก่ชีวิต" และทำให้สามารถสังเกตการตายของเซลล์เป้าหมายเป็นระลอกคลื่นได้[ 13 ]เนื่องจากมีลิพิดเรียงตัวสูงและฟอสฟาติดิลเซอรีนที่มีประจุลบอยู่ในเยื่อหุ้มพลาสมา เซลล์ T Cจึงทนต่อผลกระทบของสารพิษไซโตท็อกซินเพอร์ฟอรินและแกรนไซม์ได้[ 14 ]
วิธีที่สองในการชักนำให้เกิดอะพอพโทซิสคือผ่านการปฏิสัมพันธ์ระหว่างพื้นผิวเซลล์ของเซลล์ T Cและเซลล์ที่ติดเชื้อ เมื่อเซลล์ T Cถูกกระตุ้น มันจะเริ่มแสดงโปรตีนบนพื้นผิว เซลล์ที่เรียกว่า FAS ligand (FasL)(Apo1L)(CD95L) ซึ่งสามารถจับกับ โมเลกุล Fas (Apo1)(CD95) ที่แสดงออกบนเซลล์เป้าหมายได้ อย่างไรก็ตาม การปฏิสัมพันธ์ระหว่าง Fas กับ Fas ligand นี้เชื่อว่ามีความสำคัญต่อการกำจัดลิมโฟไซต์ T ที่ไม่ต้องการ ในระหว่างการพัฒนาหรือต่อกิจกรรมการทำลายล้างของเซลล์ T H บางชนิด มากกว่ากิจกรรมการทำลายล้างของเซลล์ T C ที่ทำหน้าที่เป็น เซลล์เอฟเฟกเตอร์ การจับกันของ Fas กับ FasL ช่วยให้เกิดการดึงดูดคอมเพล็กซ์ส่งสัญญาณที่เหนี่ยวนำให้เกิดการตาย (DISC) [ 15 ] โดเมนการตายที่เกี่ยวข้องกับ Fas (FADD) เคลื่อนย้ายไปพร้อมกับ DISC ทำให้สามารถดึงดูดโปรแคสเปส 8 และ 10 ได้[ 15 ]จากนั้นแคสเปสเหล่านี้จะกระตุ้นแคสเปสตัวกระตุ้น 3, 6 และ 7 ซึ่งนำไปสู่การตัดสารตั้งต้นของการตาย เช่นลามิน A , ลามิน B1, ลามิน B2, PARP ( โพลี ADP ไรโบสพอลิเมอเรส ) และDNA-PKcs (โปรตีนไคเนสที่กระตุ้นด้วย DNA) ผลลัพธ์สุดท้ายคืออะพอพโทซิสของเซลล์ที่แสดง Fas เซลล์ T CD8 ยังสามารถแสดงการตายของเซลล์ที่เกิดจากการกระตุ้นหรือ AICD ซึ่งถูกควบคุมโดยคอมเพล็กซ์ตัวรับ CD3 เมื่อเร็วๆ นี้ โปรตีน TLT-1 ที่ปล่อยออกมาจากเกล็ดเลือดได้รับการแสดงให้เห็นว่าสามารถกระตุ้นการตายของเซลล์แบบ AICD ในเซลล์ T CD8 ได้[ 16 ]
มีการเสนอแนะว่า ปัจจัยการถอดรหัสEomesoderminมีบทบาทสำคัญในการทำงานของเซลล์ CD8 + T โดยทำหน้าที่เป็นยีนควบคุมในการตอบสนองภูมิคุ้มกันแบบปรับตัว[ 17 ]การศึกษาที่ตรวจสอบผลของการสูญเสียการทำงานของ Eomesodermin พบว่าการลดลงของการแสดงออกของปัจจัยการถอดรหัสนี้ส่งผลให้ปริมาณของเพอร์ฟอรินที่ผลิตโดยเซลล์ CD8 + T ลดลง [ 17 ]
บทบาทในการเกิดโรค
ต่างจากแอนติบอดีซึ่งมีประสิทธิภาพต่อทั้ง การติดเชื้อ ไวรัสและแบคทีเรียเซลล์ T ที่เป็นพิษต่อเซลล์ส่วนใหญ่มีประสิทธิภาพต่อไวรัส[ 18 ]
ในระหว่าง การติดเชื้อ ไวรัสตับอักเสบ B (HBV) เซลล์ T ที่เป็นพิษต่อเซลล์จะฆ่าเซลล์ที่ติดเชื้อและผลิตไซโตไคน์ต้านไวรัสที่สามารถกำจัด HBV ออกจากเซลล์ตับที่ยังมีชีวิตอยู่ได้ นอกจากนี้ เซลล์เหล่านี้ยังมีบทบาทสำคัญในการก่อโรค โดยมีส่วนทำให้เกิดความเสียหายต่อตับเกือบทั้งหมดที่เกี่ยวข้องกับการติดเชื้อ HBV [ 19 ] พบว่า เกล็ดเลือดช่วยอำนวยความสะดวกในการสะสมของเซลล์ T ที่เป็นพิษต่อเซลล์ที่จำเพาะต่อไวรัสในตับที่ติดเชื้อ[ 20 ]ในการศึกษาบางส่วนกับหนู การฉีด เซลล์ CXCR5+ CD8+T แสดงให้เห็นว่า HBsAgลดลงอย่างมีนัยสำคัญนอกจากนี้ การเพิ่มขึ้นของ ระดับ CXCL13ยังช่วยอำนวยความสะดวกในการดึงดูดเซลล์ CXCR5+CD8+T ภายในตับ และเซลล์ประเภทนี้ผลิตอินเตอร์เฟรอน (IFN)-γ และIL-21 ที่จำเพาะต่อ HBV ในระดับสูง ซึ่งสามารถช่วยปรับปรุงการควบคุมการติดเชื้อ HBV เรื้อรังได้[ 21 ]
เซลล์ T ที่เป็นพิษต่อเซลล์มีส่วนเกี่ยวข้องกับความก้าวหน้าของโรคข้ออักเสบการมีส่วนร่วมหลักของโรคข้ออักเสบรูมาตอยด์คือการมีส่วนร่วม ของข้อต่อ เยื่อ หุ้มข้อมีลักษณะเฉพาะคือ การเพิ่ม จำนวนเซลล์ การเพิ่มขึ้นของหลอดเลือด และการแทรกซึมของเซลล์อักเสบ โดยส่วนใหญ่เป็นลิมโฟไซต์ T CD4+ ซึ่งเป็นผู้จัดระเบียบหลักของการตอบสนองทางภูมิคุ้มกันแบบอาศัยเซลล์ ในการศึกษาต่างๆ โรคข้ออักเสบรูมาตอยด์มีความเชื่อมโยงอย่างมากกับแอนติเจน MHC คลาส II เซลล์เดียวในร่างกายที่แสดงแอนติเจน MHC คลาส II คือเซลล์นำเสนอแอนติเจนแบบคงที่ สิ่งนี้ชี้ให้เห็นอย่างชัดเจนว่าโรคข้ออักเสบรูมาตอยด์เกิดจากแอนติเจนที่ก่อให้เกิดโรคข้ออักเสบที่ยังไม่ได้รับการระบุ แอนติเจนนั้นอาจเป็นแอนติเจนภายนอกใดๆ เช่น โปรตีนไวรัส หรือโปรตีนภายในร่างกาย[ 22 ]เมื่อเร็วๆ นี้ มีการระบุแอนติเจนภายในร่างกายที่เป็นไปได้จำนวนหนึ่ง เช่น ไกลโคโปรตีนกระดูกอ่อนของมนุษย์ 39 โปรตีนที่จับกับโซ่หนัก และโปรตีนที่ถูกซิโทรลิน เซลล์ T CD4+ ที่ถูกกระตุ้นจะกระตุ้นโมโนไซต์ แมโครฟาจ และไฟโบรบลาสต์ในเยื่อหุ้มข้อให้สร้างไซโตไคน์อินเตอร์ลิวคิน-1 อินเตอร์ลิวคิน-6และทูมอร์เนโครซิสแฟคเตอร์อัลฟา (TNFa) และหลั่งเมทัลโลโปรตีเนส ซึ่งสามชนิดแรกเป็นกุญแจสำคัญในการกระตุ้นการอักเสบในโรคข้ออักเสบรูมาตอยด์ เซลล์ลิมโฟไซต์ที่ถูกกระตุ้นเหล่านี้ยังกระตุ้นเซลล์ B ให้ผลิตอิมมูโนโกลบูลิน รวมถึงรูมาตอยด์แฟคเตอร์ด้วย[ 23 ]บทบาทที่ก่อให้เกิดโรคยังไม่เป็นที่ทราบแน่ชัด แต่อาจเกิดจาก การ กระตุ้นคอมพลีเมนต์ผ่านการสร้างสารเชิงซ้อนภูมิคุ้มกัน นอกจากนี้ การศึกษาในสัตว์หลายชิ้นชี้ให้เห็นว่าเซลล์ T ที่เป็นพิษต่อเซลล์อาจมีผลต่อการอักเสบเป็นหลักในโรคนี้ นอกจากนี้ยังมีการศึกษาว่าการผลิตไซโตไคน์โดยเซลล์ CD8+ อาจเร่งการดำเนินไปของโรคข้ออักเสบ[ 24 ]
พบว่าเซลล์CD8 + T มีบทบาทในการติดเชื้อ HIV HIV ได้พัฒนากลยุทธ์มากมายเพื่อหลีกเลี่ยงระบบภูมิคุ้มกันของเซลล์เจ้าบ้าน ตัวอย่างเช่น HIV ได้ใช้อัตราการกลายพันธุ์ที่สูงมากเพื่อให้สามารถหลบเลี่ยงการจดจำโดยเซลล์ CD8 + T ได้[ 25 ]นอกจากนี้ยังสามารถลดการแสดงออกของโปรตีน MHC Class I บนพื้นผิวของเซลล์ที่ติดเชื้อ เพื่อหลีกเลี่ยงการทำลายโดยเซลล์ CD8 + T ได้อีกด้วย [ 25 ]หากเซลล์ CD8 + T ไม่สามารถค้นหา จดจำ และจับกับเซลล์ที่ติดเชื้อได้ ไวรัสจะไม่ถูกทำลายและจะเจริญเติบโตต่อไป
นอกจากนี้ เซลล์ CD8 + T อาจมีส่วนเกี่ยวข้องกับ โรค เบาหวานประเภทที่ 1 [ 26 ]การศึกษาในแบบจำลองหนูที่เป็นเบาหวานแสดงให้เห็นว่าเซลล์ CD4+ มีส่วนรับผิดชอบต่อการแทรกซึมของเม็ดเลือดขาวโมโนนิวเคลียร์จำนวนมากเข้าไปในเกาะตับอ่อนอย่างไรก็ตาม เซลล์ CD8+ ได้รับการพิสูจน์แล้วว่ามีบทบาทในการทำลายเซลล์เบต้าของเกาะตับอ่อนในที่สุด อย่างไรก็ตาม ในการศึกษากับหนู NOD ที่มีการกลายพันธุ์แบบไม่มีการทำงานที่ ตำแหน่ง เบต้า-2 ไมโครโกลบูลิน (B2M) และขาดโมเลกุลของคอมเพล็กซ์ความเข้ากันได้ทางเนื้อเยื่อหลักคลาส I และเซลล์ CD8+ T พบว่าพวกมันไม่เป็นโรคเบาหวาน[ 27 ]
เซลล์ CD8 + T อาจจำเป็นต่อการแก้ไขภาวะเส้นประสาทส่วนปลายอักเสบที่เกิดจากเคมีบำบัด (CIPN) [ 28 ] [ 29 ]หนูที่ไม่มีเซลล์ CD8 + T จะแสดงอาการ CIPN ที่ยาวนานกว่าเมื่อเทียบกับหนูปกติ และการฉีดเซลล์ CD8 + T ที่ได้รับการฝึกฝนจะช่วยแก้ไขหรือป้องกัน CIPN ได้
เซลล์ T ที่เป็นพิษต่อเซลล์มีส่วนเกี่ยวข้องกับการพัฒนาของโรคและความผิดปกติต่างๆ เช่นการปฏิเสธการปลูกถ่าย (เซลล์ T ที่เป็นพิษต่อเซลล์จะโจมตีอวัยวะใหม่หลังจากตรวจพบว่าเป็นสิ่งแปลกปลอมเนื่องจากความแตกต่างของ HLA ระหว่างผู้บริจาคและผู้รับ) [ 30 ]การผลิตไซโตไคน์มากเกินไปใน การติดเชื้อ SARS-CoV-2 อย่างรุนแรง (เนื่องจากการตอบสนองของลิมโฟไซต์ที่มากเกินไป ทำให้เกิด ไซโตไคน์ที่ก่อให้เกิดการอักเสบจำนวนมากซึ่งเป็นอันตรายต่อผู้ป่วย) [ 31 ] [ 32 ]โรคอักเสบและเสื่อมของระบบประสาทส่วนกลาง เช่น โรคปลอกประสาทเสื่อมแข็ง (เซลล์ T จะไวต่อโปรตีนบางชนิด เช่นไมอีลินโจมตีเซลล์ที่แข็งแรงและดึงดูดเซลล์ภูมิคุ้มกันมากขึ้น ทำให้โรครุนแรงขึ้น) [ 33 ]
ดูเพิ่มเติม
ลิงก์ภายนอก
 สื่อที่เกี่ยวข้องกับเซลล์ทีชนิดทำลายเซลล์เป้าหมาย (Cytotoxic T cells)ในวิกิมีเดียคอมมอนส์
สื่อที่เกี่ยวข้องกับเซลล์ทีชนิดทำลายเซลล์เป้าหมาย (Cytotoxic T cells)ในวิกิมีเดียคอมมอนส์- กลุ่มทีเซลล์ – มหาวิทยาลัยคาร์ดิฟฟ์
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เซลล์ทีที่เป็นพิษต่อเซลล์
เซลล์ T นักฆ่า(เรียกอีกอย่างว่า T C , เซลล์ T ลิมโฟไซต์ที่เป็นพิษต่อเซลล์ , CTL , เซลล์ T นักฆ่า , เซลล์ T ที่ทำลายเซลล์ , เซลล์ T CD8 + หรือ cd8 ) คือ เซลล์ T ลิมโฟไซต์ (...
การพัฒนา
ระบบภูมิคุ้มกันต้องจดจำแอนติเจนที่มีศักยภาพมากกว่าหนึ่งพันล้านรายการ ร่างกายมนุษย์มียีนน้อยกว่า 30,000 ยีน ดังนั้นจึงเป็นไปไม่ได้ที่จะมียีนหนึ่งตัวสำหรับแอนติเจนทุกตัว ในทางกลับกัน DNA...
การเปิดใช้งาน
เซลล์ T ที่โตเต็มวัยจะผ่านขั้นตอนต่างๆ ขึ้นอยู่กับจำนวนครั้งที่สัมผัสกับแอนติเจน ในขั้นแรก เซลล์ T ที่ยังไม่ได้รับการกระตุ้น (เซลล์ T บริสุทธิ์) เป็นระยะเริ่มต้นของเซลล์ T ที่อยู่ในต่อมไทมัส ซึ่งยังไม่เคยพบกับแอนติเจนที่มีความสัมพันธ์กับ TCR มาก่อน เซลล์ T...
หน้าที่ของตัวกระตุ้น
เมื่อสัมผัสกับเซลล์ร่างกายที่ติดเชื้อ/ทำงานผิดปกติ เซลล์ T C จะปล่อยสารพิษไซโตท็อกซิน ได้แก่ เพอร์ ฟอ ริน แกรนไซม์ และ แกรนูลิซิน โดยผ่านการทำงานของเพอร์ฟอริน แกรนไซม์จะเข้าสู่ไซโตพลาสซึมของเซลล์เป้าหมาย และการทำงาน ของ เซรินโปรตีเอส จะกระตุ้นแค สเปส แคสเคด...