อ่าน 14 นาที
การเปลี่ยนแปลงจำนวนสำเนา
ความแปรผันของจำนวนสำเนา ( CNV ) เป็นปรากฏการณ์ที่ส่วนต่างๆ ของจีโนมมีการทำซ้ำ และจำนวนการทำซ้ำในจีโนมจะแตกต่างกันไปในแต่ละบุคคล [ 1 ] ความแปรผันของจำนวนสำเนาเป็น...
การเปลี่ยนแปลงจำนวนสำเนา

ความแปรผันของจำนวนสำเนา ( CNV ) เป็นปรากฏการณ์ที่ส่วนต่างๆ ของจีโนมมีการทำซ้ำ และจำนวนการทำซ้ำในจีโนมจะแตกต่างกันไปในแต่ละบุคคล[ 1 ] ความแปรผันของจำนวนสำเนาเป็น ความแปรผันเชิงโครงสร้างชนิดหนึ่งโดยเฉพาะอย่างยิ่ง เป็น เหตุการณ์ การทำซ้ำหรือการลบที่ส่งผลกระทบต่อเบสคู่จำนวนมาก[ 2 ]ประมาณสองในสามของจีโนมมนุษย์ทั้งหมดอาจประกอบด้วยส่วนที่ทำซ้ำ[ 3 ]และ 4.8–9.5% ของจีโนมมนุษย์สามารถจัดเป็นความแปรผันของจำนวนสำเนาได้[ 4 ]ในสัตว์เลี้ยงลูกด้วยนมความแปรผันของจำนวนสำเนามีบทบาทสำคัญในการสร้างความแปรผันที่จำเป็นในประชากร รวมถึงลักษณะอาการ ของโรค ด้วย[ 1 ]
ความแปรผันของจำนวนสำเนาสามารถแบ่งออกได้เป็นสองกลุ่มหลักๆ คือ สำเนาซ้ำสั้นและสำเนาซ้ำยาว อย่างไรก็ตาม ไม่มีขอบเขตที่ชัดเจนระหว่างสองกลุ่มนี้ และการจัดประเภทขึ้นอยู่กับลักษณะของตำแหน่งที่สนใจ สำเนาซ้ำสั้นส่วนใหญ่ประกอบด้วย สำเนาซ้ำ ไดนิวคลี โอไท ด์ (นิวคลีโอไทด์ที่ซ้ำกันสองตัว เช่น ACACAC...) และสำเนาซ้ำไตรนิวคลีโอไทด์สำเนาซ้ำยาวประกอบด้วยสำเนาซ้ำของยีนทั้งหมด การจัดประเภทนี้โดยพิจารณาจากขนาดของสำเนาซ้ำเป็นการจัดประเภทที่ชัดเจนที่สุด เนื่องจากขนาดเป็นปัจจัยสำคัญในการตรวจสอบกลไกประเภทต่างๆ ที่น่าจะก่อให้เกิดสำเนาซ้ำ[ 5 ]ดังนั้นจึงอาจส่งผลต่อฟีโนไทป์ของสำเนาซ้ำเหล่านี้
ประเภทและการจัดเรียงโครโมโซมใหม่
หนึ่งในตัวอย่างที่รู้จักกันดีที่สุดของการเปลี่ยนแปลงจำนวนสำเนาสั้นๆ คือการทำซ้ำของไตรนิวคลีโอไทด์ของคู่เบส CAG ในยีนฮันติงตินซึ่งเป็นสาเหตุของโรคฮันติงตัน ซึ่งเป็นความผิดปกติทางระบบประสาท [ 6 ]สำหรับกรณีนี้ เมื่อไตรนิวคลีโอไทด์ CAG ทำซ้ำมากกว่า 36 ครั้งในการขยายตัวของการทำซ้ำไตรนิวคลีโอไทด์โรคฮันติงตันมีแนวโน้มที่จะเกิดขึ้นในบุคคลนั้นและมีแนวโน้มที่จะถ่ายทอดไปยังลูกหลานของเขาหรือเธอ[ 6 ]จำนวนการทำซ้ำของไตรนิวคลีโอไทด์ CAG มีความสัมพันธ์ผกผันกับอายุที่เริ่มมีอาการของโรคฮันติงตัน[ 7 ]การทำซ้ำสั้นๆ ประเภทนี้มักคิดว่าเกิดจากข้อผิดพลาดในกิจกรรมของพอลิเมอเรส ในระหว่าง การจำลองแบบรวมถึงการลื่นไถลของพอลิเมอเรส การสลับแม่แบบ และการสลับส้อม ซึ่งจะกล่าวถึงรายละเอียดในภายหลัง ขนาดการทำซ้ำสั้นๆ ของความแปรผันของจำนวนสำเนาเหล่านี้เอื้อต่อข้อผิดพลาดในพอลิเมอเรส เนื่องจากบริเวณที่ทำซ้ำเหล่านี้มีแนวโน้มที่จะถูกพอลิเมอเรสจดจำผิดพลาด และบริเวณที่จำลองแบบอาจถูกจำลองแบบอีกครั้ง ทำให้เกิดสำเนาเพิ่มเติมของการทำซ้ำ[ 8 ]นอกจากนี้ หากการทำซ้ำของไตรนิวคลีโอไทด์เหล่านี้อยู่ในเฟรมการอ่าน เดียวกัน ใน ส่วน การเข้ารหัสของยีน อาจนำไปสู่สายยาวของกรดอะมิโน ชนิดเดียวกัน ซึ่งอาจสร้างโปรตีนรวมกลุ่มในเซลล์[ 7 ]และหากการทำซ้ำสั้นๆ เหล่านี้ตกอยู่ในส่วนที่ไม่เข้ารหัสของยีน อาจส่งผลต่อการแสดงออกและการควบคุมยีนในทางกลับกัน การทำซ้ำจำนวนแปรผันของยีนทั้งหมดนั้นพบได้น้อยกว่าในจีโนม ตัวอย่างหนึ่งของการทำซ้ำยีนทั้งหมดคือ ยีน อัลฟา-อะไมเลส 1 ( AMY1 ) ซึ่งเข้ารหัสอัลฟา-อะไมเลส ซึ่งมีความแปรผันของจำนวนสำเนาอย่างมีนัยสำคัญระหว่างประชากรต่างๆ ที่มีอาหารแตกต่างกัน[ 9 ]แม้ว่ากลไกเฉพาะที่ทำให้ ยีน AMY1เพิ่มหรือลดจำนวนสำเนาของมันยังคงเป็นหัวข้อถกเถียงกันอยู่ แต่สมมติฐานบางอย่างชี้ให้เห็นว่าการเชื่อมต่อปลายที่ไม่เหมือนกันหรือการเชื่อมต่อปลายที่เกิดจากไมโครโฮโมโลจีน่าจะเป็นสาเหตุของการทำซ้ำยีนทั้งหมดเหล่านี้[ 9 ]การทำซ้ำของยีนทั้งหมดมีผลโดยตรงต่อการแสดงออกของยีนนั้นๆ และข้อเท็จจริงที่ว่าความแปรผันของจำนวนสำเนาของAMY1ยีนที่เกี่ยวข้องกับอาหารเป็นตัวอย่างที่น่าทึ่งของการปรับตัวทางวิวัฒนาการของมนุษย์เมื่อไม่นานมานี้[ 9 ]แม้ว่านี่จะเป็นกลุ่มทั่วไปที่การเปลี่ยนแปลงจำนวนสำเนาถูกจัดกลุ่มไว้ แต่จำนวนเบสคู่ที่แน่นอนที่การเปลี่ยนแปลงจำนวนสำเนาส่งผลกระทบนั้นขึ้นอยู่กับตำแหน่งที่สนใจเฉพาะ ปัจจุบัน จากข้อมูลของการเปลี่ยนแปลงจำนวนสำเนาที่รายงานทั้งหมด ขนาดเฉลี่ยของตัวแปรจำนวนสำเนาอยู่ที่ประมาณ 118kb และค่ามัธยฐานอยู่ที่ประมาณ 18kb [ 10 ]
ในแง่ของโครงสร้างสถาปัตยกรรมของการแปรผันจำนวนสำเนา งานวิจัยได้แนะนำและกำหนดบริเวณฮอตสปอตในจีโนมซึ่งการแปรผันจำนวนสำเนามีความเข้มข้นมากกว่าถึงสี่เท่า[ 2 ]บริเวณฮอตสปอตเหล่านี้ถูกกำหนดให้เป็นบริเวณที่มีการทำซ้ำยาวที่คล้ายคลึงกัน 90–100% ซึ่งรู้จักกันในชื่อการทำซ้ำแบบแบ่งส่วนไม่ว่าจะ เป็นแบบ เรียงต่อกันหรือแบบกระจาย และที่สำคัญที่สุดคือ บริเวณฮอตสปอตเหล่านี้มีอัตราการจัดเรียงโครโมโซมใหม่ ที่เพิ่ม ขึ้น[ 2 ]เชื่อกันว่าการจัดเรียงโครโมโซมใหม่ขนาดใหญ่เหล่านี้ก่อให้เกิดความแปรผัน ปกติ และโรคทางพันธุกรรมรวมถึงการแปรผันจำนวนสำเนา[ 1 ]ยิ่งไปกว่านั้น ฮอตสปอตการแปรผันจำนวนสำเนาเหล่านี้มีความสอดคล้องกันในประชากรจำนวนมากจากทวีปต่างๆ ซึ่งหมายความว่าฮอตสปอตเหล่านี้อาจถูกได้รับมาโดยอิสระโดยประชากรทั้งหมดและส่งต่อผ่านรุ่นสู่รุ่น หรืออาจถูกได้รับมาในวิวัฒนาการของมนุษย์ยุค แรก ก่อนที่ประชากรจะแยกตัว ซึ่งอย่างหลังดูเหมือนจะเป็นไปได้มากกว่า[ 1 ]สุดท้ายนี้ ดูเหมือนว่าจะไม่มีอคติเชิงพื้นที่ของตำแหน่งที่การเปลี่ยนแปลงจำนวนสำเนากระจายตัวหนาแน่นที่สุดในจีโนม[ 1 ]แม้ว่าเดิมทีจะตรวจพบโดยการผสมแบบฟลูออเรสเซนต์ในแหล่งกำเนิดและการวิเคราะห์ไมโครแซทเทลไลต์ว่าการทำซ้ำจำนวนสำเนาอยู่ในบริเวณที่มีการทำซ้ำสูง เช่นเทโลเมียร์ เซนโทรเมียร์และเฮเทอโรโครมาติน [ 11 ] แต่การศึกษาทั่วทั้งจีโนมเมื่อเร็วๆ นี้ได้สรุปเป็นอย่างอื่น[ 2 ]กล่าวคือ บริเวณ ซับเทโลเมียร์และบริเวณเพอริเซนโทรเมียร์เป็นบริเวณที่พบฮอตสปอตการจัดเรียงโครโมโซมใหม่มากที่สุด และไม่มีการเพิ่มขึ้นอย่างมีนัยสำคัญของการเปลี่ยนแปลงจำนวนสำเนาในบริเวณนั้น[ 2 ]ยิ่งไปกว่านั้น บริเวณฮอตสปอตการจัดเรียงโครโมโซมใหม่เหล่านี้ไม่ได้มีจำนวนยีนลดลง ซึ่งหมายความว่ามีอคติเชิงพื้นที่น้อยที่สุดของตำแหน่งจีโนมของการเปลี่ยนแปลงจำนวนสำเนา[ 2 ]
การตรวจจับและการระบุตัวตน
ในตอนแรก การเปลี่ยนแปลงจำนวนสำเนาถูกคิดว่าครอบครองส่วนเล็ก ๆ ของจีโนมที่น้อยมากและไม่สำคัญเมื่อสังเกต ทาง เซลล์พันธุศาสตร์[ 12 ]โดยทั่วไปแล้ว การเปลี่ยนแปลงจำนวนสำเนาจะเกี่ยวข้องกับลำดับซ้ำแบบเรียงต่อกันขนาดเล็กหรือความผิดปกติทางพันธุกรรมเฉพาะ[ 13 ]ดังนั้น ในตอนแรก การเปลี่ยนแปลงจำนวนสำเนาจึงถูกตรวจสอบเฉพาะในแง่ของตำแหน่งเฉพาะเท่านั้น อย่างไรก็ตาม การพัฒนาทางเทคโนโลยีทำให้มีวิธีการระบุและศึกษาการเปลี่ยนแปลงจำนวนสำเนาที่แม่นยำมากขึ้นเรื่อย ๆ การเปลี่ยนแปลงจำนวนสำเนาได้รับการศึกษาครั้งแรกโดยเทคนิคทางเซลล์พันธุศาสตร์ ซึ่งเป็นเทคนิคที่ช่วยให้สามารถสังเกตโครงสร้างทางกายภาพของโครโมโซมได้[ 12 ]หนึ่งในเทคนิคเหล่านี้คือการไฮบริดไดเซชันในแหล่งกำเนิดด้วยฟลูออเรสเซนต์ (FISH) ซึ่งเกี่ยวข้องกับการใส่โพรบฟลูออเรสเซนต์ที่ต้องการ ความสมบูรณ์ในระดับสูงในจีโนมสำหรับการจับ[ 10 ]การไฮบริดไดเซชันทางจีโนมเชิงเปรียบเทียบก็ถูกใช้กันทั่วไปในการตรวจจับการเปลี่ยนแปลงจำนวนสำเนาโดย การมองเห็น ฟลูออโรฟอร์แล้วเปรียบเทียบความยาวของโครโมโซม[ 10 ]
ความก้าวหน้าล่าสุดใน เทคโนโลยี จีโนมิกส์ทำให้เกิดวิธีการสำคัญมากมายที่มีความละเอียดทางจีโนมสูงมาก และส่งผลให้มีการรายงานความแปรผันของจำนวนสำเนาในจีโนมเพิ่มมากขึ้น[ 10 ]ในขั้นต้น ความก้าวหน้าเหล่านี้เกี่ยวข้องกับการใช้ อาร์เรย์ โครโมโซมเทียมแบคทีเรีย (BAC) ที่มีช่วงห่างประมาณ 1 เมกะเบสตลอดทั้งยีน[ 14 ] BAC ยังสามารถตรวจจับความแปรผันของจำนวนสำเนาในฮอตสปอตการจัดเรียงใหม่ ทำให้สามารถตรวจจับความแปรผันของจำนวนสำเนาใหม่ได้ 119 รายการ[ 2 ]การจัดลำดับจีโนมแบบความเร็วสูงได้ปฏิวัติวงการจีโนมิกส์ของมนุษย์ และ มีการศึกษา ในคอมพิวเตอร์เพื่อตรวจจับความแปรผันของจำนวนสำเนาในจีโนม[ 2 ]ลำดับอ้างอิงได้รับการเปรียบเทียบกับลำดับอื่นๆ ที่น่าสนใจโดยใช้ฟอสมีดโดยควบคุมโคลนฟอสมีดให้มีขนาด 40 กิโลเบสอย่างเคร่งครัด[ 15 ]การจัดลำดับปลายอ่านจะให้ข้อมูลที่เพียงพอในการจัดเรียงลำดับอ้างอิงกับลำดับที่สนใจ และการจัดเรียงที่ไม่ถูกต้องจะสังเกตเห็นได้ง่าย จึงสรุปได้ว่าเป็นการเปลี่ยนแปลงจำนวนสำเนาภายในบริเวณนั้นของโคลน[ 15 ]เทคนิคการตรวจจับประเภทนี้ให้ความละเอียดของจีโนมสูงและระบุตำแหน่งซ้ำในจีโนมได้อย่างแม่นยำ และยังสามารถตรวจจับการเปลี่ยนแปลงโครงสร้างประเภทอื่น ๆ เช่น การผกผันได้อีกด้วย[ 10 ]
นอกจากนี้ อีกวิธีหนึ่งในการตรวจจับความแปรผันของจำนวนสำเนาคือการใช้โพลีมอร์ฟิซึมของนิวคลีโอไทด์เดี่ยว (SNP) [ 10 ]เนื่องจากมีข้อมูล SNP ของมนุษย์จำนวนมาก ทิศทางการตรวจจับความแปรผันของจำนวนสำเนาจึงเปลี่ยนไปใช้ SNP เหล่านี้[ 16 ]โดยอาศัยข้อเท็จจริงที่ว่าการรวมตัวใหม่ ของมนุษย์ ค่อนข้างหายาก และเหตุการณ์การรวมตัวใหม่จำนวนมากเกิดขึ้นในบริเวณเฉพาะของจีโนมที่เรียกว่าฮอตสปอตการรวมตัวใหม่ความไม่สมดุลของการเชื่อมโยงสามารถใช้เพื่อระบุความแปรผันของจำนวนสำเนาได้[ 16 ]มีความพยายามในการเชื่อมโยงความแปรผันของจำนวนสำเนากับ SNP ของแฮปโลไทป์ เฉพาะ โดยการวิเคราะห์ความไม่สมดุลของการเชื่อมโยง โดยใช้การเชื่อมโยงเหล่านี้ เราสามารถระบุความแปรผันของจำนวนสำเนาในจีโนมโดยใช้ SNP เป็นเครื่องหมาย เทคนิค การจัดลำดับรุ่นต่อไปรวมถึงการจัดลำดับแบบอ่านสั้นและอ่านยาว กำลังถูกนำมาใช้มากขึ้นในปัจจุบันและเริ่มเข้ามาแทนที่เทคนิคแบบอาร์เรย์ในการตรวจจับความแปรผันของจำนวนสำเนา[ 17 ] [ 18 ]
กลไกโมเลกุล
There are two main types of molecular mechanism for the formation of copy number variations: homologous based and non-homologous based.[5] Although many suggestions have been put forward, most of these theories are speculations and conjecture. There is no conclusive evidence that correlates a specific copy number variation to a specific mechanism.

หนึ่งในทฤษฎีที่ได้รับการยอมรับมากที่สุดที่นำไปสู่ความแปรผันของจำนวนสำเนา รวมถึงการลบและการผกผัน คือการรวมตัวกันใหม่ของโครโมโซมคู่เหมือนที่ไม่ใช่แอลลีล [ 19 ] ในระหว่างการรวมตัวกันใหม่ ของไมโอ ซิส โครโมโซมคู่เหมือนจะจับคู่กันและสร้างรอยแตกสองสายที่ปลายทั้งสองข้าง ทำให้เกิดฮอลลิเดย์จังก์ชัน อย่างไรก็ตาม ในกลไกที่ผิดปกติ ในระหว่างการสร้างฮอลลิเดย์จังก์ชัน รอยแตกสองสายจะไม่อยู่ในแนวเดียวกัน และการไขว้กันจะเกิดขึ้นในตำแหน่งที่ไม่ใช่แอลลีลบนโครโมโซมเดียวกัน เมื่อฮอลลิเดย์จังก์ชันได้รับการแก้ไข เหตุการณ์การไขว้กันที่ไม่เท่ากันจะทำให้เกิดการถ่ายโอนสารพันธุกรรมระหว่างโครโมโซมคู่เหมือนทั้งสอง และเป็นผลให้ส่วนหนึ่งของ DNA บนโครโมโซมคู่เหมือนทั้งสองถูกทำซ้ำ[ 19 ]เนื่องจากบริเวณที่ทำซ้ำไม่ได้แยกตัวอย่างอิสระ อีกต่อ ไป บริเวณที่ทำซ้ำของโครโมโซมจึงถูกถ่ายทอดทางพันธุกรรม กลไกการรวมตัวกันของโฮโมโลกัสอีกประเภทหนึ่งที่สามารถนำไปสู่การเปลี่ยนแปลงจำนวนสำเนาได้เรียกว่าการจำลองแบบที่เกิดจากการแตกหัก[ 20 ]เมื่อเกิดการแตกหักของสายคู่ในจีโนมโดยไม่คาดคิด เซลล์จะกระตุ้นเส้นทางที่ทำหน้าที่ซ่อมแซมการแตกหัก[ 20 ]ข้อผิดพลาดในการซ่อมแซมการแตกหัก เช่นเดียวกับการรวมตัวกันของโฮโมโลกัสที่ไม่ใช่แอลลีล สามารถนำไปสู่การเพิ่มจำนวนสำเนาของบริเวณเฉพาะในจีโนมได้ ในระหว่างการซ่อมแซมการแตกหักของสายคู่ ปลายที่แตกหักสามารถบุกรุกโครโมโซมโฮโมโลกัสแทนที่จะเชื่อมต่อกับสายเดิม[ 20 ]เช่นเดียวกับกลไกการรวมตัวกันของโฮโมโลกัสที่ไม่ใช่แอลลีล สำเนาพิเศษของบริเวณเฉพาะจะถูกถ่ายโอนไปยังโครโมโซมอื่น ทำให้เกิดเหตุการณ์การทำซ้ำ นอกจากนี้ ยังพบว่า โปรตีน โคฮีซินช่วยในระบบการซ่อมแซมการแตกหักของสายคู่โดยการหนีบปลายทั้งสองข้างให้อยู่ใกล้กัน ซึ่งป้องกันการบุกรุกระหว่างโครโมโซมของปลายทั้งสองข้าง[ 21 ]หากด้วยเหตุผลใดก็ตาม เช่น การกระตุ้นของไรโบโซมอลอาร์เอ็นเอ กิจกรรมของโคฮีซินได้รับผลกระทบ อาจทำให้เกิดข้อผิดพลาดในการซ่อมแซมสายคู่ที่เพิ่มขึ้นในบริเวณนั้น[ 21 ]
กลไกอีกประเภทหนึ่งที่คาดว่าจะนำไปสู่การเปลี่ยนแปลงจำนวนสำเนาคือกลไกที่ไม่เหมือนกัน เพื่อแยกแยะระหว่างกลไกนี้กับกลไกที่เหมือนกัน เราต้องเข้าใจแนวคิดของความเหมือนกัน การจับคู่โครโมโซมที่เหมือนกันเกี่ยวข้องกับการใช้สาย DNA ที่มีความคล้ายคลึงกันสูง (~97%) และสายเหล่านี้ต้องมีความยาวมากกว่าความยาวที่กำหนดเพื่อหลีกเลี่ยงการจับคู่ที่สั้นแต่มีความคล้ายคลึงกันสูง[ 5 ]ในทางกลับกัน การจับคู่ที่ไม่เหมือนกันอาศัยความคล้ายคลึงกันของเบสคู่เพียงไม่กี่คู่ระหว่างสองสาย ดังนั้นจึงเป็นไปได้ที่วัสดุทางพันธุกรรมจะถูกแลกเปลี่ยนหรือทำซ้ำในกระบวนการซ่อมแซมสายคู่ที่ไม่เหมือนกัน[ 5 ]
กลไกประเภทหนึ่งที่ไม่เหมือนกันคือกลไกการเชื่อมต่อปลายที่ไม่เหมือนกันหรือกลไกการเชื่อมต่อปลายไมโครโฮโมโลจี[ 22 ]กลไกเหล่านี้ยังเกี่ยวข้องกับการซ่อมแซมการแตกของสายคู่ แต่ไม่จำเป็นต้องมีความเหมือนกันหรือมีความเหมือนกันในระดับจำกัด[ 5 ]เมื่อสายเหล่านี้ได้รับการซ่อมแซม มักจะมีการลบหรือการแทรกขนาดเล็กเพิ่มเข้าไปในสายที่ได้รับการซ่อมแซม เป็นไปได้ว่าเรโทรทรานสโพซอนจะถูกแทรกเข้าไปในจีโนมผ่านระบบการซ่อมแซมนี้[ 22 ]หากเรโทรทรานสโพซอนถูกแทรกเข้าไปในตำแหน่งที่ไม่ใช่แอลลีลบนโครโมโซม การรวมตัวใหม่แบบไมโอซิสสามารถผลักดันให้การแทรกนั้นรวมตัวใหม่เข้ากับสายเดียวกันกับสำเนาที่มีอยู่แล้วของบริเวณเดียวกัน กลไกอีกอย่างหนึ่งคือวงจรการแตก-หลอมรวม-เชื่อมต่อ ซึ่งเกี่ยวข้องกับโครมาทิดคู่ที่สูญเสียบริเวณเทโลเมอร์เนื่องจากการแตกของสายคู่[ 23 ]มีการเสนอว่าโครมาทิดคู่แฝดเหล่านี้จะรวมกันเพื่อสร้างโครโมโซมไดเซนทริก หนึ่งตัว จากนั้นจะแยกตัวออกเป็นนิวเคลียสที่แตกต่างกันสองนิวเคลียส[ 23 ]เนื่องจากการดึงโครโมโซมไดเซนทริกออกจากกันทำให้เกิดการแตกของสายคู่ บริเวณปลายสามารถรวมเข้ากับการแตกของสายคู่อื่นๆ และทำซ้ำวงจรได้[ 23 ]การรวมตัวของโครมาทิดคู่แฝดสองตัวสามารถทำให้เกิดการทำซ้ำแบบกลับด้าน และเมื่อเหตุการณ์เหล่านี้เกิดขึ้นซ้ำๆ ตลอดวงจร บริเวณที่กลับด้านจะถูกทำซ้ำ ทำให้จำนวนสำเนาเพิ่มขึ้น[ 23 ]กลไกสุดท้ายที่สามารถนำไปสู่ความแปรผันของจำนวนสำเนาคือการลื่นไถลของพอลิเมอเรส ซึ่งเรียกอีกอย่างว่าการสลับแม่แบบ[ 24 ]ในระหว่างการจำลองดีเอ็นเอตามปกติ พอลิเมอเรสบนสายที่ล้าหลังจะต้องคลายและยึดบริเวณการจำลองอย่างต่อเนื่อง[ 24 ]เมื่อมีการทำซ้ำขนาดเล็กในลำดับ DNA อยู่แล้ว โพลีเมอเรสอาจ 'สับสน' เมื่อมันจับยึดอีกครั้งเพื่อดำเนินการจำลองแบบต่อไป และแทนที่จะจับยึดกับคู่เบสที่ถูกต้อง มันอาจเลื่อนไปสองสามคู่เบสและจำลองส่วนหนึ่งของบริเวณที่ซ้ำกันอีกครั้ง[ 24 ]โปรดทราบว่าถึงแม้จะได้รับการสังเกตจากการทดลองและเป็นกลไกที่ได้รับการยอมรับอย่างกว้างขวาง แต่ปฏิสัมพันธ์ระดับโมเลกุลที่นำไปสู่ข้อผิดพลาดนี้ยังคงไม่เป็นที่ทราบแน่ชัด นอกจากนี้ เนื่องจากกลไกประเภทนี้ต้องการให้พอลิเมอเรสกระโดดไปรอบๆ สาย DNA และเป็นไปได้ยากที่พอลิเมอเรสจะสามารถจับยึดที่ตำแหน่งอื่นที่อยู่ห่างกันหลายกิโลเบสได้ ดังนั้นจึงเหมาะสมกว่าสำหรับลำดับซ้ำสั้นๆ เช่น ลำดับซ้ำของไดนิวคลีโอไทด์หรือไตรนิวคลีโอไทด์[ 25 ]
ยีนอัลฟา-อะไมเลส

อะไมเลสเป็นเอนไซม์ในน้ำลายที่ทำหน้าที่ย่อยสลายแป้งให้เป็นโมโนแซ็กคาไรด์และอะไมเลสชนิดหนึ่งถูกเข้ารหัสโดยยีนอัลฟา-อะไมเลส ( AMY1 ) [ 9 ]ตำแหน่งAMY1เช่นเดียวกับเอนไซม์อะไมเลส เป็นหนึ่งในยีนที่ได้รับการศึกษาและจัดลำดับอย่างกว้างขวางที่สุดในจีโนมมนุษย์ โฮโมล็อกของมันยังพบในไพรเมตอื่นๆ ด้วย ดังนั้นจึงเป็นไปได้ว่า ยีน AMY1 ของไพรเมต เป็นบรรพบุรุษของ ยีน AMY1 ของมนุษย์ และได้รับการปรับตัวในช่วงต้นของการวิวัฒนาการของไพรเมต[ 9 ] AMY1เป็นหนึ่งในยีนที่ได้รับการศึกษาอย่างดีที่สุด ซึ่งมีจำนวนสำเนาที่หลากหลายในประชากรมนุษย์ที่แตกต่างกัน[ 9 ]ยีนAMY1ยังเป็นหนึ่งในยีนไม่กี่ตัวที่ได้รับการศึกษาซึ่งแสดงหลักฐานที่น่าเชื่อถือซึ่งเชื่อมโยงการทำงานของโปรตีนกับจำนวนสำเนาของมัน[ 9 ]เป็นที่ทราบกันดีว่าจำนวนสำเนาสามารถเปลี่ยนแปลง ระดับ การถอดรหัสและการแปลของยีนเฉพาะได้ อย่างไรก็ตาม งานวิจัยแสดงให้เห็นว่าความสัมพันธ์ระหว่างระดับโปรตีนและจำนวนสำเนานั้นมีความแปรปรวน[ 26 ]ใน ยีน AMY1ของชาวอเมริกันเชื้อสายยุโรป พบว่าความเข้มข้นของอะไมเลสในน้ำลายมีความสัมพันธ์อย่างใกล้ชิดกับจำนวนสำเนาของยีนAMY1 [ 9 ]ด้วยเหตุนี้ จึงมีการตั้งสมมติฐานว่าจำนวนสำเนาของ ยีน AMY1มีความสัมพันธ์อย่างใกล้ชิดกับหน้าที่ของโปรตีน ซึ่งก็คือการย่อยแป้ง[ 9 ]
พบว่าจำนวนสำเนาของยีน AMY1 มีความสัมพันธ์กับระดับแป้งที่แตกต่างกันในอาหารของประชากรกลุ่มต่างๆ[ 9 ]ประชากร8กลุ่มจากทวีปต่างๆ ถูกจัดประเภทเป็นอาหารที่มีแป้งสูงและอาหารที่มีแป้งต่ำ และจำนวนสำเนาของ ยีน AMY1ถูกแสดงให้เห็นโดยใช้ FISH และqPCRที่ มีความละเอียดสูง [ 9 ]พบว่าประชากรที่รับประทานอาหารที่มีแป้งสูง ซึ่งประกอบด้วยชาวญี่ปุ่น ชาวฮัดซา และชาวอเมริกันเชื้อสายยุโรป มีจำนวนสำเนาของยีนAMY1 เฉลี่ยสูงกว่าอย่างมีนัยสำคัญ (สูงกว่าสองเท่า) เมื่อเทียบกับประชากรที่รับประทานอาหารที่มีแป้งต่ำ ซึ่งรวมถึงชาวบิอากา ชาวมบูติ ชาวดาต็อก และชาวยาคุต[ 9 ]มีการตั้งสมมติฐานว่าระดับของแป้งในอาหารปกติ ซึ่งเป็นสารตั้งต้นของ AMY1 สามารถส่งผลโดยตรงต่อจำนวนสำเนาของยีนAMY1 ได้ [ 9 ]เนื่องจากสรุปได้ว่าจำนวนสำเนาของAMY1มีความสัมพันธ์โดยตรงกับอะไมเลสในน้ำลาย[ 9 ]ยิ่งมีแป้งในอาหารประจำวันของประชากรมากเท่าไร การมีสำเนาของยีนAMY1 หลายสำเนาก็ยิ่งเอื้อต่อวิวัฒนาการมากขึ้นเท่านั้น ยีน AMY1เป็นยีนแรกที่ให้หลักฐานที่ชัดเจนสำหรับการวิวัฒนาการในระดับพันธุศาสตร์โมเลกุล[ 26 ]ยิ่งไปกว่านั้น การใช้การผสมแบบจีโนมเปรียบเทียบ ได้มีการเปรียบเทียบความแปรผันของจำนวนสำเนาของจีโนมทั้งหมดของประชากรญี่ปุ่นกับประชากรยาคุต[ 9 ]พบว่าความแปรผันของจำนวนสำเนาของ ยีน AMY1แตกต่างอย่างมีนัยสำคัญจากความแปรผันของจำนวนสำเนาในยีนหรือบริเวณอื่นๆ ของจีโนม ซึ่งบ่งชี้ว่ายีนAMY1อยู่ภายใต้แรงกดดันการคัดเลือกที่รุนแรงซึ่งมีอิทธิพลน้อยหรือไม่มีผลต่อความแปรผันของจำนวนสำเนาอื่นๆ[ 9 ]สุดท้ายนี้ ได้มีการเปรียบเทียบความแปรปรวนของความยาวของไมโครแซทเทลไลต์ 783 ตัว ระหว่างประชากรทั้งสองกลุ่มกับความแปรปรวนของจำนวนสำเนาของ ยีน AMY1พบว่า ช่วงจำนวนสำเนาของยีน AMY1มีขนาดใหญ่กว่าไมโครแซทเทลไลต์ที่ตรวจสอบมากกว่า 97% [ 9 ]ซึ่งหมายความว่าการคัดเลือกโดยธรรมชาติมีบทบาทสำคัญในการกำหนดจำนวนเฉลี่ยของ ยีน AMY1ในประชากรทั้งสองกลุ่มนี้[ 9 ]อย่างไรก็ตาม เนื่องจากมีการศึกษาเพียงหกกลุ่มประชากร จึงเป็นสิ่งสำคัญที่จะต้องพิจารณาความเป็นไปได้ว่าอาจมีปัจจัยอื่นๆ ในอาหารหรือวัฒนธรรมของพวกเขาที่ส่งผลต่อ จำนวนสำเนาของยีน AMY1นอกเหนือจากแป้ง
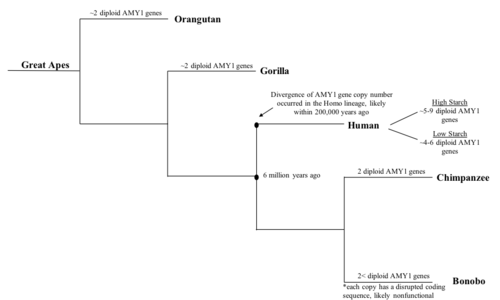
แม้ว่าจะไม่ชัดเจนว่า จำนวนสำเนาของยีน AMY1เริ่มเพิ่มขึ้นเมื่อใด แต่ก็เป็นที่ทราบและยืนยันแล้วว่า ยีน AMY1มีอยู่ในไพรเมตยุคแรกพบว่าชิมแปนซี ซึ่งเป็นญาติทางวิวัฒนาการที่ใกล้ชิดที่สุดของมนุษย์ มี สำเนาของ ยีน AMY1 แบบดิ พลอยด์สองชุด ที่มีความยาวเท่ากับยีน AMY1 ของมนุษย์[ 9 ]ซึ่งน้อยกว่าของมนุษย์อย่างมีนัยสำคัญ ในทางกลับกันพบว่าโบโนโบ ซึ่งเป็นญาติใกล้ชิดของมนุษย์ยุคใหม่เช่นกัน มีสำเนาของ ยีนAMY1 แบบดิพลอยด์มากกว่าสองชุด [ 9 ]อย่างไรก็ตาม ยีน AMY1 ของโบโนโบ ได้รับการจัดลำดับและวิเคราะห์ และพบว่าลำดับการเข้ารหัสของ ยีน AMY1ถูกรบกวน ซึ่งอาจนำไปสู่การผลิตอะไมเลสในน้ำลายที่ทำงานผิดปกติ[ 9 ]จากผลลัพธ์สามารถอนุมานได้ว่าการเพิ่มขึ้นของ จำนวนสำเนาของยีน AMY1 ในโบโนโบ ไม่น่าจะสัมพันธ์กับปริมาณแป้งในอาหารของพวกมัน มีการตั้งสมมติฐานเพิ่มเติมว่าการเพิ่มจำนวนสำเนาเริ่มต้นขึ้นเมื่อไม่นานมานี้ในช่วงวิวัฒนาการของโฮมินิน ยุคแรก เนื่องจาก ลิงใหญ่ ไม่มีสำเนาของยีน AMY1มากกว่าสองสำเนาที่สร้างโปรตีนที่ใช้งานได้[ 9 ]นอกจากนี้ ยังมีการคาดการณ์ว่าการเพิ่ม จำนวนสำเนาของ AMY1เริ่มต้นขึ้นเมื่อประมาณ 20,000 ปีที่แล้ว เมื่อมนุษย์เปลี่ยนจาก วิถีชีวิต แบบล่าสัตว์และเก็บเกี่ยวมาเป็น สังคม เกษตรกรรมซึ่งเป็นช่วงเวลาที่มนุษย์พึ่งพาพืชหัวที่มีแป้งสูง เป็นอย่างมาก [ 9 ]สมมติฐานนี้ แม้ว่าจะสมเหตุสมผล แต่ก็ขาดหลักฐานเชิงทดลองเนื่องจากความยากลำบากในการรวบรวมข้อมูลเกี่ยวกับการเปลี่ยนแปลงของอาหารมนุษย์ โดยเฉพาะอย่างยิ่งพืชหัวที่มีแป้งสูง เนื่องจากไม่สามารถสังเกตหรือทดสอบได้โดยตรง ความก้าวหน้าล่าสุดในการจัดลำดับดีเอ็นเอทำให้นักวิจัยสามารถจัดลำดับดีเอ็นเอที่เก่ากว่า เช่น ดีเอ็นเอของมนุษย์นีแอนเดอร์ ทาล ได้ในระดับความแม่นยำระดับหนึ่ง บางทีการจัดลำดับดีเอ็นเอของมนุษย์นีแอนเดอร์ทาลอาจให้เครื่องหมายบอกเวลาที่ จำนวนสำเนาของยีน AMY1เพิ่มขึ้นและให้ข้อมูลเชิงลึกเกี่ยวกับอาหารของมนุษย์และวิวัฒนาการของยีน
ปัจจุบันยังไม่ทราบกลไกที่ทำให้เกิดการจำลองยีนอะไมเลสในขั้นต้น และอาจหมายความว่าการแทรกของ ลำดับ เรโทรไวรัสเกิดจากการเชื่อมต่อปลายที่ไม่เหมือนกัน ซึ่งทำให้เกิดการจำลองยีนAMY1 [ 27 ]อย่างไรก็ตาม ปัจจุบันยังไม่มีหลักฐานสนับสนุนทฤษฎีนี้ ดังนั้นสมมติฐานนี้จึงยังคงเป็นเพียงการคาดเดา การกำเนิดของ ยีน AMY1 หลายสำเนาเมื่อไม่นานมานี้ บ่งชี้ว่าขึ้นอยู่กับสภาพแวดล้อม จำนวนสำเนาของยีน AMY1สามารถเพิ่มขึ้นและลดลงได้อย่างรวดเร็วเมื่อเทียบกับยีนที่ไม่โต้ตอบกับสภาพแวดล้อมโดยตรง[ 26 ]ยีนAMY1เป็นตัวอย่างที่ดีเยี่ยมของวิธีที่ปริมาณยีนส่งผลต่อการอยู่รอดของสิ่งมีชีวิตในสภาพแวดล้อมที่กำหนด สำเนาหลายชุดของ ยีน AMY1ทำให้ผู้ที่พึ่งพาอาหารที่มีแป้งสูงมีข้อได้เปรียบทางวิวัฒนาการ ดังนั้นจำนวนสำเนาของยีนที่สูงจึงคงอยู่ในประชากร[ 26 ]
เซลล์สมอง
ในบรรดาเซลล์ประสาทในสมองของมนุษย์การเปลี่ยนแปลงจำนวนสำเนาที่ได้มาจากโซมาติกนั้นเกิดขึ้นบ่อยครั้ง[ 28 ]การเปลี่ยนแปลงจำนวนสำเนาแสดงให้เห็นถึงความแปรปรวนที่กว้าง (9 ถึง 100% ของเซลล์ประสาทในสมองในการศึกษาต่างๆ) การเปลี่ยนแปลงส่วนใหญ่มีขนาดระหว่าง 2 ถึง 10 Mb โดยการลบมีจำนวนมากกว่าการเพิ่มจำนวนอย่างมาก[ 28 ]
การเพิ่มจำนวนจีโนมและการเพิ่มจำนวนสามเท่าของยีนดูเหมือนจะเป็นสาเหตุที่หายากของโรคพาร์กินสันแม้ว่าจะพบได้บ่อยกว่าการกลายพันธุ์แบบจุดก็ตาม[ 29 ]
การเปลี่ยนแปลงจำนวนสำเนาใน ยีน RCL1เกี่ยวข้องกับ ฟีโนไทป์ ทางระบบประสาทและจิตเวช หลายประเภท ในเด็ก[ 30 ]
ตระกูลยีนและการคัดเลือกโดยธรรมชาติ

Recently, there had been discussion connecting copy number variations to gene families. Gene families are defined as a set of related genes that serve similar functions but have minor temporal or spatial differences and these genes likely derived from one ancestral gene.[26] The main reason copy number variations are connected to gene families is that there is a possibility that genes in a family may have derived from one ancestral gene which got duplicated into different copies.[26] Mutations accumulate through time in the genes and with natural selection acting on the genes, some mutations lead to environmental advantages allowing those genes to be inherited and eventually clear gene families are separated out. An example of a gene family that may have been created due to copy number variations is the globin gene family. The globin gene family is an elaborate network of genes consisting of alpha and beta globin genes including genes that are expressed in both embryos and adults as well as pseudogenes.[31] These globin genes in the globin family are all well conserved and only differ by a small portion of the gene, indicating that they were derived from a common ancestral gene, perhaps due to duplication of the initial globin gene.[31]
Research has shown that copy number variations are significantly more common in genes that encode proteins that directly interact with the environment than proteins that are involved in basic cellular activities.[32] It was suggested that the gene dosage effect accompanying copy number variation may lead to detrimental effects if essential cellular functions are disrupted, therefore proteins involved in cellular pathways are subjected to strong purifying selection.[32] In addition, proteins function together and interact with proteins of other pathways, therefore it is important to view the effects of natural selection on bio-molecular pathways rather than on individual proteins. With that being said, it was found that proteins in the periphery of the pathway are enriched in copy number variations whereas proteins in the center of the pathways are depleted in copy number variations.[33] It was explained that proteins in the periphery of the pathway interact with fewer proteins and so a change in protein dosage affected by a change in copy number may have a smaller effect on the overall outcome of the cellular pathway.[33]
See also
- CaSNP, a database
- Comparative genomics
- Copy number analysis
- Human genome
- Inparanoid
- Molecular evolution
- Pseudogenes
- Segmental duplication
- Tandem exon duplication
- Virtual karyotype
อ่านเพิ่มเติม
- Pollack JR, Perou CM, Alizadeh AA, Eisen MB, Pergamenschikov A, Williams CF, Jeffrey SS, Botstein D, Brown PO (กันยายน 1999). "การวิเคราะห์การเปลี่ยนแปลงจำนวนสำเนา DNA ทั่วทั้งจีโนมโดยใช้ไมโครอาร์เรย์ cDNA" Nature Genetics . 23 (1): 41– 6. doi : 10.1038/12640 . PMID 10471496 . S2CID 997032 .
- "ความแปรผันทางพันธุกรรมมหาศาลในคนที่มีสุขภาพดี"นิวไซเอนทิสต์ 7 สิงหาคม 2547
- Carter NP (กันยายน 2547). "ปกติที่สุดเท่าที่จะเป็นไปได้?" Nature Genetics . 36 (9): 931– 2. doi : 10.1038/ng0904-931 . PMID 15340426 .
- ตรวจสอบ E (ตุลาคม 2548) "จีโนมมนุษย์: ผู้คนที่ประกอบขึ้นจากชิ้นส่วนต่างๆ" Nature 437 ( 7062 ): 1084– 6. Bibcode : 2005Natur.437.1084C . doi : 10.1038/4371084a . PMID 16237414 . S2CID 8211641 .
- "การเพิ่มจำนวนยีนอาจเป็นตัวกำหนดว่าคุณเป็นใคร"นิวไซเอนทิสต์ 22 พฤศจิกายน 2549
- "ดีเอ็นเอมีความแตกต่างกันอย่างมากในแต่ละบุคคล แผนที่ทางพันธุกรรมเผยให้เห็น"เนชั่นแนล จีโอแกรฟิก 22 พฤศจิกายน 2549 เก็บถาวรจากต้นฉบับเมื่อ 25 พฤศจิกายน 2549
- "การเลือกเลนส์ที่เหมาะสม" (PDF) . Nature Genetics. 1 กรกฎาคม 2550.
- Lam HY, Mu XJ, Stütz AM, Tanzer A, Cayting PD, Snyder M และคณะ (มกราคม 2010). "การวิเคราะห์โครงสร้างตัวแปรด้วยความละเอียดระดับนิวคลีโอไทด์โดยใช้ BreakSeq และไลบรารีจุดแตกหัก" Nature Biotechnology 28 ( 1). Nature Biotechnology: 47– 55. doi : 10.1038/nbt.1600 . PMC 2951730 . PMID 20037582 .
- "งานวิจัยใหม่เผยสาเหตุทางพันธุกรรมของออทิสติก" Singularity Hub. 15 มิถุนายน 2010. เก็บถาวรจากต้นฉบับเมื่อ 18 มิถุนายน 2010. เรียกดูเมื่อ15 มิถุนายน 2010 .
ลิงก์ภายนอก
- โครงการความแปรผันของจำนวนสำเนาสถาบันแซงเกอร์
- ข้ออ้าง: ฝาแฝดเหมือนมีดีเอ็นเอเหมือนกันทุกประการ
- แพลตฟอร์มการระบุข้อมูลแบบบูรณาการสำหรับความแปรผันของจำนวนสำเนาในมนุษย์
- บรรณานุกรมเกี่ยวกับความแปรผันของจำนวนสำเนา
- ฐานข้อมูลความแปรผันทางพันธุกรรม (Database of Genomic Variants ) คือฐานข้อมูลของความแปรผันเชิงโครงสร้างในจีโนมมนุษย์
- การตรวจจับความแปรผันของจำนวนสำเนาผ่านการระบุจีโนไทป์ SNP ความหนาแน่นสูง
- เทคโนโลยีพันธุกรรมอ็อกซ์ฟอร์ด
- หมายเลขสำเนา BioDiscovery Nexus
- การทำแผนที่ความละเอียดสูงของความแปรผันของจำนวนสำเนาในบุคคลที่มีสุขภาพดี 2,026 คน
- IGSR: แหล่งรวบรวมตัวอย่างจีโนมระหว่างประเทศ
- cn.FARMS: โมเดลตัวแปรแฝงสำหรับตรวจจับความแปรผันของจำนวนสำเนาในข้อมูลไมโครอาร์เรย์ด้วยอัตราการค้นพบที่ผิดพลาดต่ำ แพ็กเกจ R —ซอฟต์แวร์
- cn.MOPS: การผสมผสานของ Poisson สำหรับการค้นหาความแปรผันของจำนวนสำเนาในข้อมูลการจัดลำดับรุ่นใหม่ — ซอฟต์แวร์
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การเปลี่ยนแปลงจำนวนสำเนา
ความแปรผันของจำนวนสำเนา ( CNV ) เป็นปรากฏการณ์ที่ส่วนต่างๆ ของจีโนมมีการทำซ้ำ และจำนวนการทำซ้ำในจีโนมจะแตกต่างกันไปในแต่ละบุคคล [ 1 ] ความแปรผันของจำนวนสำเนาเป็น...
ประเภทและการจัดเรียงโครโมโซมใหม่
หนึ่งในตัวอย่างที่รู้จักกันดีที่สุดของการเปลี่ยนแปลงจำนวนสำเนาสั้นๆ คือการทำซ้ำของไตรนิวคลีโอไทด์ของคู่เบส CAG ใน ยีนฮันติงติน ซึ่งเป็นสาเหตุของ โรคฮันติงตัน ซึ่งเป็นความผิดปกติทางระบบประสาท [ 6 ] สำหรับกรณีนี้ เมื่อไตรนิวคลีโอไทด์ CAG ทำซ้ำมากกว่า 36 ครั้งใน...
การตรวจจับและการระบุตัวตน
ในตอนแรก การเปลี่ยนแปลงจำนวนสำเนาถูกคิดว่าครอบครองส่วนเล็ก ๆ ของจีโนมที่น้อยมากและไม่สำคัญเมื่อสังเกต ทาง เซลล์พันธุศาสตร์ [ 12 ] โดยทั่วไปแล้ว การเปลี่ยนแปลงจำนวนสำเนาจะเกี่ยวข้องกับลำดับซ้ำแบบเรียงต่อกันขนาดเล็กหรือความผิดปกติทางพันธุกรรมเฉพาะ [ 13 ]...
กลไกโมเลกุล
There are two main types of molecular mechanism for the formation of copy number variations: homologous based and non-homologous based. [ 5 ] Although many suggestions have been put forward, most of these theories are speculations and conjecture.