อ่าน 19 นาที
การจำลองดีเอ็นเอ
การจำลองดีเอ็นเอ เป็น กระบวนการ ที่ เซลล์ สร้างสำเนา ดีเอ็นเอ ของตัวเองอย่างแม่นยำ[ 1 ] [ 2 ] [ 3 ] [ 4 ] กระบวนการ นี้เกิดขึ้นใน สิ่งมีชีวิต ทุกชนิด และมีความสำคัญต่อ...
การจำลองดีเอ็นเอ

การจำลองดีเอ็นเอเป็นกระบวนการที่เซลล์ สร้างสำเนา ดีเอ็นเอของตัวเองอย่างแม่นยำ[ 1 ] [ 2 ] [ 3 ] [ 4 ] กระบวนการนี้เกิดขึ้นในสิ่งมีชีวิต ทุกชนิด และมีความสำคัญต่อการถ่ายทอดทางชีววิทยาการแบ่งเซลล์และการซ่อมแซม เนื้อเยื่อที่เสียหาย การจำลองดีเอ็นเอทำให้มั่นใจได้ว่า เซลล์ลูกที่แบ่งตัวใหม่แต่ละเซลล์จะได้รับสำเนาของโมเลกุลดีเอ็นเอแต่ละโมเลกุล[ 5 ]
ดีเอ็นเอส่วนใหญ่มักอยู่ในรูปสายคู่ ซึ่งประกอบด้วยสายเสริม สองสาย ที่ยึดติดกันด้วยการจับคู่เบสของนิวคลีโอไทด์ที่ประกอบเป็นแต่ละสาย สายเชิงเส้นสองสายของโมเลกุลดีเอ็นเอสายคู่มักจะบิดเข้าด้วยกันในรูป ทรง เกลียวคู่[ 6 ]ในระหว่างการจำลองแบบ สายทั้งสองจะแยกออกจากกัน และแต่ละสายของโมเลกุลดีเอ็นเอเดิมจะทำหน้าที่เป็นแม่แบบสำหรับการสร้างสายคู่เสริม ซึ่งเป็นกระบวนการที่เรียกว่าการจำลองแบบกึ่งอนุรักษ์ส่งผลให้โมเลกุลดีเอ็นเอที่จำลองแบบแต่ละโมเลกุลประกอบด้วยสายดีเอ็นเอเดิมหนึ่งสายและสายที่สังเคราะห์ขึ้นใหม่หนึ่งสาย[ 7 ] กลไก การตรวจสอบ ความถูกต้องและการตรวจสอบข้อผิดพลาด ของเซลล์ ทำให้ การจำลองแบบดีเอ็นเอมีความแม่นยำเกือบสมบูรณ์แบบ[ 8 ] [ 9 ]
การจำลองดีเอ็นเอมักจะเริ่มต้นที่ตำแหน่งเฉพาะที่เรียกว่าจุดเริ่มต้นของการจำลอง[ 10 ]ซึ่งกระจายอยู่ทั่วจีโนม [ 11 ] การคลายเกลียวของดีเอ็นเอที่จุดเริ่มต้นจะเกิดขึ้นโดยเอนไซม์ที่เรียกว่าเฮลิเคสและส่งผลให้เกิดส้อมการจำลองที่เติบโตไปในสองทิศทางจากจุดเริ่มต้นโปรตีน จำนวนมาก เกี่ยวข้องกับส้อมการจำลองเพื่อช่วยในการเริ่มต้นและต่อเนื่องของการสังเคราะห์ดีเอ็นเอที่โดดเด่นที่สุด คือ ดีเอ็นเอพอลิเมอเรสซึ่งสังเคราะห์สายใหม่โดยการรวมนิวคลีโอไทด์ที่เข้าคู่กับนิวคลีโอไทด์ของสายแม่แบบ การจำลองดีเอ็นเอเกิดขึ้นในช่วงระยะ S (การสังเคราะห์) ของระยะอินเตอร์เฟส[ 12 ]
การจำลองดีเอ็นเอสามารถทำได้ในหลอดทดลอง (โดยเทียม นอกเซลล์) [ 13 ]เอนไซม์ดีเอ็นเอพอลิเมอเรสที่แยกได้จากเซลล์และไพรเมอร์ดีเอ็นเอเทียมสามารถใช้เพื่อเริ่มต้นการสังเคราะห์ดีเอ็นเอที่ลำดับที่ทราบในโมเลกุลดีเอ็นเอแม่แบบปฏิกิริยาลูกโซ่พอลิเมอเรส (PCR) ปฏิกิริยาลูกโซ่ไลเกส (LCR) และการขยายสัญญาณผ่านการถอดรหัส (TMA) ล้วนเป็นตัวอย่างทั่วไปของเทคนิคนี้ ในเดือนมีนาคม พ.ศ. 2564 นักวิจัยรายงานหลักฐานที่ชี้ให้เห็นว่ารูปแบบเบื้องต้นของอาร์เอ็นเอถ่ายโอนซึ่งเป็นองค์ประกอบที่จำเป็นของการแปล (การสังเคราะห์ทางชีวภาพของ โปรตีนใหม่ตามรหัสพันธุกรรม ) อาจเป็นโมเลกุลจำลองตัวเองในการกำเนิดสิ่งมีชีวิตยุค แรกเริ่ม [ 14 ] [ 15 ]
โครงสร้างดีเอ็นเอ

ดีเอ็นเอเป็นโครงสร้างแบบสองสาย โดยทั้งสองสายจะขดเข้าด้วยกันเพื่อสร้างเกลียวคู่ ที่เป็นลักษณะเฉพาะ แต่ละสายเดี่ยวของดีเอ็นเอเป็นโซ่ของนิวคลีโอไทด์สี่ชนิดนิ วคลีโอไท ด์ในดีเอ็นเอประกอบด้วยน้ำตาลดีออกซีไรโบส ฟอสเฟตและนิวคลีโอเบส นิวคลีโอไทด์ สี่ชนิดนี้สอดคล้องกับ นิว คลีโอเบส สี่ชนิด ได้แก่อะดีนีนไซโตซีนกัวนีนและไทมีนซึ่งโดยทั่วไปจะย่อเป็น A, C, G และ T ตามลำดับ อะดีนีนและกัวนีนเป็น นิวคลีโอเบส พิวรีน[ 16 ]ในขณะที่ไซโตซีนและไทมีนเป็นไพริมิดีน นิวคลีโอไท ด์เหล่านี้สร้างพันธะฟอสโฟไดเอสเทอร์ ทำให้เกิดโครงสร้างฟอสเฟต-ดีออกซีไรโบสของเกลียวคู่ดีเอ็นเอโดยมีนิวคลีโอเบสชี้เข้าด้านใน (เช่น ไปทางสายตรงข้าม) นิวคลีโอเบสที่เสริมกันจะจับคู่กันระหว่างสายผ่านพันธะไฮโดรเจนเพื่อสร้างคู่เบส อะดีนีนจับคู่กับไทมีน (พันธะไฮโดรเจนสองพันธะ) และกัวนีนจับคู่กับไซโตซีน ( พันธะไฮโดรเจน สามพันธะ ) [ 17 ]
สาย DNA มีทิศทางและปลายที่แตกต่างกันของสายเดี่ยวเรียกว่า "ปลาย 3′ (สามไพรม์)" และ "ปลาย 5′ (ห้าไพรม์)" ตามธรรมเนียม หากลำดับเบสของสาย DNA เดี่ยวถูกกำหนด ปลายด้านซ้ายของลำดับจะเป็นปลาย 5′ ในขณะที่ปลายด้านขวาของลำดับจะเป็นปลาย 3′ สายของเกลียวคู่จะขนานกันในทิศทางตรงข้าม โดยสายหนึ่งเป็น 5′ ถึง 3′ และสายตรงข้ามเป็น 3′ ถึง 5′ คำศัพท์เหล่านี้อ้างอิงถึงธรรมเนียมทางเคมีในการกำหนดหมายเลขอะตอมคาร์บอนที่ประกอบเป็นโมเลกุลดีออกซีไรโบส และระบุอะตอมคาร์บอนเฉพาะที่ฟอสเฟตตัวถัดไปในโซ่จะยึดติด ทิศทางมีผลต่อการสังเคราะห์ DNA เนื่องจาก DNA polymerase สามารถสังเคราะห์ DNA ได้ในทิศทางเดียวเท่านั้นโดยการเพิ่มนิวคลีโอไทด์ที่ปลาย 3′ ของสาย DNA [ 18 ]
การจับคู่เบสที่เสริมกันใน DNA (ผ่านพันธะไฮโดรเจน ) หมายความว่าข้อมูลที่บรรจุอยู่ภายในแต่ละสายนั้นซ้ำซ้อน พันธะฟอสโฟไดเอสเทอร์ (ภายในสาย) แข็งแรงกว่าพันธะไฮโดรเจน (ระหว่างสาย) หน้าที่ที่แท้จริงของพันธะฟอสโฟไดเอสเทอร์คือการเชื่อมต่ออะตอมคาร์บอน 5' ของนิวคลีโอไทด์หนึ่งกับอะตอมคาร์บอน 3' ของนิวคลีโอไทด์อีกตัวหนึ่ง ในขณะที่พันธะไฮโดรเจนทำให้เกลียวคู่ของ DNA มีเสถียรภาพในแนวขวางแกนเกลียว แต่ไม่ใช่ในทิศทางของแกนตามยาว[ 19 ]ทำให้สามารถแยกสายออกจากกันได้ ดังนั้นนิวคลีโอไทด์บนสายเดี่ยวจึงสามารถใช้สร้างนิวคลีโอไทด์บนสายคู่ที่สังเคราะห์ขึ้นใหม่ได้[ 20 ]
ดีเอ็นเอพอลิเมอเรส

ดีเอ็นเอพอลิเมอเรสเป็นกลุ่มของเอนไซม์ที่ทำหน้าที่ในการจำลองดีเอ็นเอทุกรูปแบบ[ 22 ]โดยทั่วไปแล้ว ดีเอ็นเอพอลิเมอเรสไม่สามารถเริ่มต้นการสังเคราะห์สายใหม่ได้ แต่สามารถขยายสายดีเอ็นเอหรืออาร์เอ็นเอที่มีอยู่ซึ่งจับคู่กับสายแม่แบบได้เท่านั้น ในการเริ่มต้นการสังเคราะห์ จะต้องสร้างชิ้นส่วนอาร์เอ็นเอสั้นๆ ที่เรียกว่าไพรเมอร์และจับคู่กับสายดีเอ็นเอแม่แบบ
เอนไซม์ดีเอ็นเอพอลิเมอเรสจะเพิ่มสายดีเอ็นเอใหม่โดยการต่อปลาย 3′ ของสายนิวคลีโอไทด์ที่มีอยู่แล้ว โดยเพิ่มนิวคลีโอไทด์ ใหม่ ที่เข้าคู่กับสายแม่แบบทีละตัว ผ่านการสร้างพันธะฟอสโฟไดเอสเทอร์พลังงานสำหรับกระบวนการพอลิเมอไรเซชันของดีเอ็นเอมาจากการไฮโดรไลซิสของพันธะฟอสเฟต (ฟอสโฟแอนไฮไดรด์) พลังงานสูงระหว่างฟอสเฟตสามตัวที่ติดอยู่กับเบส แต่ละตัวที่ยังไม่ได้ รวมเข้าด้วยกัน เบสอิสระที่มีโมเลกุลน้ำตาลติดอยู่ (ดีออกซีไรโบสในกรณีของดีเอ็นเอ) เรียกว่านิวคลี โอไซด์ และนิวคลีโอไซด์ที่มีหมู่ฟอสเฟตติดอยู่หนึ่งหมู่หรือมากกว่านั้นเรียก ว่า นิ วคลีโอไท ด์โดยเฉพาะอย่างยิ่ง นิวคลีโอไซด์ที่มีหมู่ฟอสเฟตติดอยู่สามหมู่เรียกว่านิวคลีโอไซด์ไตรฟอสเฟตเมื่อมีการเพิ่มนิวคลีโอไทด์อิสระเข้าไปในสาย DNA ที่กำลังเติบโต การเกิดพันธะฟอสโฟไดเอสเทอร์ระหว่างฟอสเฟตด้านใกล้ของนิวคลีโอไทด์อิสระกับดีออกซีไรโบสของนิวคลีโอไทด์อีกตัวหนึ่งในสายที่กำลังเติบโตนั้น จะเกิดขึ้นพร้อมกับการไฮโดรไลซิสของพันธะฟอสเฟตพลังงานสูง ส่งผลให้ฟอสเฟตสองหมู่ด้านไกลของนิวคลีโอไทด์อิสระถูกปลดปล่อยออกมาเป็นไพโรฟอสเฟตการไฮโดรไลซิสด้วยเอนไซม์ของไพโรฟอสเฟต ที่เกิดขึ้นให้ กลายเป็นฟอสเฟตอนินทรีย์จะใช้พันธะฟอสเฟตพลังงานสูงอีกพันธะหนึ่ง ทำให้ปฏิกิริยานี้ไม่สามารถย้อนกลับได้อย่างมีประสิทธิภาพ[หมายเหตุ 1 ]
โดยทั่วไป ดีเอ็นเอพอลิเมอเรสมีความแม่นยำสูง โดยมีอัตราความผิดพลาดโดยธรรมชาติที่น้อยกว่าหนึ่งความผิดพลาดต่อทุกๆ 10⁷ นิวคลีโอไทด์ที่เพิ่มเข้ามา[ 23 ]ดีเอ็นเอพอลิเมอเรสบางชนิดยังสามารถลบนิวคลีโอไทด์ออกจากปลายสายที่กำลังพัฒนาเพื่อแก้ไขเบสที่ไม่ตรงกัน ซึ่งเรียกว่าการตรวจสอบความถูกต้อง (proofreading) สุดท้าย กลไกการซ่อมแซมความไม่ตรงกันหลังการจำลองแบบจะตรวจสอบดีเอ็นเอเพื่อหาข้อผิดพลาด โดยสามารถแยกแยะความไม่ตรงกันในสายดีเอ็นเอที่สังเคราะห์ขึ้นใหม่จากลำดับสายเดิมได้ ขั้นตอนการแยกแยะทั้งสามนี้ร่วมกันทำให้ความแม่นยำในการจำลองแบบน้อยกว่าหนึ่งความผิดพลาดต่อทุกๆ 10⁹ นิวคลีโอไทด์ที่เพิ่มเข้ามา[ 23 ]
อัตราการจำลองดีเอ็นเอในเซลล์ที่มีชีวิตถูกวัดครั้งแรกโดยอัตราการยืดตัวของดีเอ็นเอของฟาจ T4 ในE. coli ที่ติดเชื้อฟา จ[ 24 ]ในช่วงระยะเวลาของการเพิ่มขึ้นของดีเอ็นเอแบบทวีคูณที่ 37 °C อัตราดังกล่าวคือ 749 นิวคลีโอไทด์ต่อวินาที อัตราการกลายพันธุ์ต่อคู่เบสต่อการจำลองในระหว่างการสังเคราะห์ดีเอ็นเอของฟาจ T4 คือ 1.7 ต่อ10 8 [ 25 ]
กระบวนการจำลองแบบ


การจำลองดีเอ็นเอ เช่นเดียวกับกระบวนการพอลิเมอไรเซชันทางชีวภาพทั้งหมด ดำเนินไปในสามขั้นตอนที่เร่งปฏิกิริยาและประสานงานกันโดยเอนไซม์ ได้แก่ การเริ่มต้น การยืดออก และการสิ้นสุด[ 26 ]
การเริ่มต้น


เพื่อให้เซลล์แบ่งตัวได้ เซลล์จะต้องจำลอง DNA ก่อน[ 27 ]การจำลอง DNA เป็นกระบวนการแบบทั้งหมดหรือไม่มีเลย เมื่อการจำลองเริ่มต้นขึ้น มันจะดำเนินต่อไปจนเสร็จสมบูรณ์ เมื่อการจำลองเสร็จสมบูรณ์แล้ว จะไม่เกิดขึ้นอีกในวงจรเซลล์เดียวกัน สิ่งนี้เป็นไปได้ด้วยการแบ่งตัวของการเริ่มต้นของคอมเพล็กซ์ก่อนการจำลอง
คอมเพล็กซ์ก่อนการจำลองแบบ
ในช่วงปลายไมโทซิสและต้นเฟส G1คอมเพล็กซ์ขนาดใหญ่ของโปรตีนเริ่มต้นจะประกอบกันเป็นคอมเพล็กซ์ก่อนการจำลองแบบที่จุดเฉพาะใน DNA ซึ่งเรียกว่า " ต้นกำเนิด " [ 11 ] [ 10 ]ในE. coliโปรตีนเริ่มต้นหลักคือDNA Aในยีสต์นี่คือคอมเพล็กซ์การจดจำต้นกำเนิด[ 28 ] ลำดับที่ใช้โดยโปรตีนเริ่มต้นมักจะเป็น "AT-rich" (อุดมไปด้วยเบสอะดีนีนและไทมีน) เนื่องจากคู่เบส AT มีพันธะไฮโดรเจนสองพันธะ (แทนที่จะเป็นสามพันธะที่เกิดขึ้นในคู่ CG) และจึงแยกสายได้ง่ายกว่า[ 29 ] ในยูคาริโอต คอมเพล็กซ์การจดจำต้นกำเนิด (ORC) จะเร่งปฏิกิริยาการประกอบโปรตีนเริ่มต้นเข้ากับคอมเพล็กซ์ก่อนการจำลองแบบ นอกจากนี้ รายงานล่าสุดยังชี้ให้เห็นว่า ORC ของยีสต์ที่ กำลัง แตกหน่อจะเกิดการรวมตัวเป็นไดเมอร์ในลักษณะที่ขึ้นอยู่กับวงจรเซลล์เพื่อควบคุมการอนุญาต[ 30 ] [ 31 ] ในทางกลับกัน กระบวนการสร้างไดเมอร์ของ ORC จะถูกควบคุมโดยวงจรการสร้างไดเมอร์ของ Noc3p ที่ขึ้นอยู่กับวัฏจักรเซลล์ในร่างกาย และบทบาทของ Noc3p นี้สามารถแยกออกจากบทบาทของมันในการสร้างไรโบโซมได้ วงจรการสร้างไดเมอร์ของ Noc3p ที่จำเป็นจะควบคุมการสร้างดับเบิลเฮกซาเมอร์ของ ORC ในการอนุญาตการจำลองแบบและ Noc3p จะจับกับโครมาตินอย่างต่อเนื่องตลอดวัฏจักรเซลล์[ 32 ] จากนั้น Cdc6และCdt1จะเชื่อมโยงกับคอมเพล็กซ์การจดจำต้นกำเนิดที่จับอยู่ที่ต้นกำเนิดเพื่อสร้างคอมเพล็กซ์ขนาดใหญ่ขึ้นซึ่งจำเป็นต่อการโหลดคอมเพล็กซ์ Mcmลงบน DNA ในยูคาริโอต คอมเพล็กซ์ Mcm คือเฮลิเคสที่จะแยกเกลียว DNA ที่จุดแยกและต้นกำเนิดของการจำลองแบบ คอมเพล็กซ์ Mcm จะถูกดึงดูดในช่วงปลายระยะ G1 และถูกโหลดโดยคอมเพล็กซ์ ORC-Cdc6-Cdt1 ลงบน DNA ผ่านการปรับโครงสร้างโปรตีนที่ขึ้นอยู่กับ ATP การโหลดคอมเพล็กซ์ MCM ลงบน DNA ต้นกำเนิดถือเป็นการเสร็จสิ้นการก่อตัวของคอมเพล็กซ์ก่อนการจำลองแบบ[ 33 ]
หากสภาพแวดล้อมเหมาะสมในช่วงปลายระยะ G1 คอมเพล็กซ์ ไซคลิน G1 และ G1/S - Cdkจะถูกกระตุ้น ซึ่งจะกระตุ้นการแสดงออกของยีนที่เข้ารหัสส่วนประกอบของเครื่องจักรสังเคราะห์ DNA การกระตุ้น G1/S-Cdk ยังส่งเสริมการแสดงออกและการกระตุ้นคอมเพล็กซ์ S-Cdk ซึ่งอาจมีบทบาทในการกระตุ้นจุดเริ่มต้นการจำลองแบบขึ้นอยู่กับชนิดและประเภทของเซลล์ การควบคุม Cdk เหล่านี้แตกต่างกันไปตามประเภทของเซลล์และระยะการพัฒนา การควบคุมนี้เข้าใจได้ดีที่สุดในยีสต์ที่กำลังแตกหน่อซึ่งไซคลิน S Clb5และClb6มีหน้าที่หลักในการจำลองแบบ DNA [ 34 ]คอมเพล็กซ์ Clb5,6-Cdk1 กระตุ้นการทำงานของจุดเริ่มต้นการจำลองแบบโดยตรง ดังนั้นจึงจำเป็นต้องมีตลอดระยะ S เพื่อกระตุ้นจุดเริ่มต้นแต่ละจุดโดยตรง[ 33 ]
ในทำนองเดียวกันCdc7ก็มีความจำเป็นในช่วงS phaseเพื่อกระตุ้นจุดเริ่มต้นของการจำลองแบบ Cdc7 ไม่ได้ทำงานตลอดวงจรเซลล์ และการกระตุ้นของมันจะถูกกำหนดเวลาอย่างเคร่งครัดเพื่อหลีกเลี่ยงการเริ่มต้นการจำลองแบบ DNA ก่อนกำหนด ในช่วงปลาย G1 กิจกรรมของ Cdc7 เพิ่มขึ้นอย่างฉับพลันอันเป็นผลมาจากการเชื่อมโยงกับหน่วยย่อยควบคุมDBF4ซึ่งจับกับ Cdc7 โดยตรงและส่งเสริมกิจกรรมโปรตีนไคเนสของมัน พบว่า Cdc7 เป็นตัวควบคุมอัตราการทำงานของจุดเริ่มต้น โดยรวมแล้ว G1/S-Cdks และ/หรือ S-Cdks และ Cdc7 ทำงานร่วมกันเพื่อกระตุ้นจุดเริ่มต้นของการจำลองแบบโดยตรง นำไปสู่การเริ่มต้นการสังเคราะห์ DNA [ 33 ]
คอมเพล็กซ์ก่อนการเริ่มต้น
ในระยะ S ตอนต้น การกระตุ้น S-Cdk และ Cdc7 นำไปสู่การประกอบคอมเพล็กซ์ก่อนการเริ่มต้น ซึ่งเป็นคอมเพล็กซ์โปรตีนขนาดใหญ่ที่เกิดขึ้นที่ต้นกำเนิด การก่อตัวของคอมเพล็กซ์ก่อนการเริ่มต้นทำให้ Cdc6 และ Cdt1 หลุดออกจากคอมเพล็กซ์การจำลองแบบที่ต้นกำเนิด ทำให้คอมเพล็กซ์ก่อนการจำลองแบบไม่ทำงานและสลายตัว การโหลดคอมเพล็กซ์ก่อนการเริ่มต้นลงบนต้นกำเนิดจะกระตุ้นเฮลิเคส Mcm ทำให้เกิดการคลายเกลียวของ DNA คอมเพล็กซ์ก่อนการเริ่มต้นยังโหลดα-primaseและพอลิเมอเรส DNA อื่นๆ ลงบน DNA ด้วย[ 33 ]
หลังจากที่ α-primase สังเคราะห์ไพรเมอร์ตัวแรกแล้ว จุดเชื่อมต่อไพรเมอร์-แม่แบบจะทำปฏิกิริยากับตัวโหลดแคลมป์ ซึ่งจะโหลดแคลมป์แบบเลื่อนลงบน DNA เพื่อเริ่มต้นการสังเคราะห์ DNA ส่วนประกอบของคอมเพล็กซ์ก่อนการเริ่มต้นจะยังคงเชื่อมโยงกับส้อมการจำลองแบบในขณะที่เคลื่อนออกจากจุดเริ่มต้น[ 33 ]
การยืดตัว
เอนไซม์ดีเอ็นเอพอลิเมอเรสมีกิจกรรมในทิศทาง 5′–3′ ระบบการจำลองดีเอ็นเอที่รู้จักทั้งหมดต้องการ หมู่ ไฮดรอกซิล 3′ ที่ว่าง อยู่ก่อนจึงจะเริ่มการสังเคราะห์ได้ (หมายเหตุ: แม่แบบดีเอ็นเอจะถูกอ่านในทิศทาง 3′ ไป 5′ ในขณะที่สายใหม่จะถูกสังเคราะห์ในทิศทาง 5′ ไป 3′ ซึ่งมักทำให้เกิดความสับสน) มีกลไกการสังเคราะห์ดีเอ็นเอที่แตกต่างกันสี่แบบที่ได้รับการยอมรับ:
- สิ่งมีชีวิตเซลล์เดียวทุกชนิด รวมถึงไวรัส ดีเอ็นเอ แบค ทีริโอ เฟจและพลาสมิด จำนวนมาก ใช้ เอนไซม์ ไพรเมสในการสังเคราะห์ไพรเมอร์อาร์เอ็นเอขนาดสั้นที่มีหมู่ 3′ OH อิสระ ซึ่งต่อมาจะถูกต่อให้ยาวขึ้นโดยเอนไซม์ดีเอ็นเอพอลิเมอเรส
- เรโทรเอเลเมนต์ (รวมถึงเรโทรไวรัส ) ใช้ทรานสเฟอร์อาร์เอ็นเอที่เป็นตัวเริ่มต้นการจำลองดีเอ็นเอโดยการให้หมู่ 3′ OH อิสระ ซึ่งจะถูกนำไปใช้ในการยืดสายโดยรีเวอร์สทรานสคริปเทส
- ในอะดีโนไวรัสและแบคทีริโอเฟจตระกูล φ29 หมู่ 3′ OH มาจากโซ่ข้างของกรดอะมิโนในโปรตีนที่ยึดติดกับจีโนม (โปรตีนปลายทาง) ซึ่งนิวคลีโอไทด์จะถูกเติมโดยดีเอ็นเอพอลิเมอเรสเพื่อสร้างสายใหม่
- ในไวรัสดีเอ็นเอสายเดี่ยว ซึ่งเป็นกลุ่มที่รวมถึงเซอร์โคไวรัสเจมินีไวรัสพาร์โวไวรัสและอื่นๆ รวมถึงฟาจและพลาสมิด จำนวนมาก ที่ใช้กลไกการจำลองแบบวงกลม (RCR) เอนโดนิวคลีเอส RCR จะสร้างรอยบากในสายจีโนม (ไวรัสสายเดี่ยว) หรือสายดีเอ็นเอสายใดสายหนึ่ง (พลาสมิด) ปลาย 5′ ของสายที่ถูกตัดจะถูกถ่ายโอนไปยัง หมู่ ไทโรซีนบนนิวคลีเอส และหมู่ 3′ OH ที่เป็นอิสระจะถูกใช้โดยดีเอ็นเอพอลิเมอเรสเพื่อสังเคราะห์สายใหม่
สิ่งมีชีวิตเซลล์เดียวใช้กลไกแรกนี้ ซึ่งเป็นกลไกที่รู้จักกันดีที่สุด ในกลไกนี้ เมื่อสายดีเอ็นเอทั้งสองแยกออกจากกันแล้วเอนไซม์ไพรเมสจะเพิ่มไพรเมอร์ RNA เข้าไปในสายแม่แบบ สายนำจะได้รับไพรเมอร์ RNA หนึ่งตัว ในขณะที่สายตามจะได้รับหลายตัว สายนำจะถูกต่อขยายอย่างต่อเนื่องจากไพรเมอร์โดยเอนไซม์ดีเอ็นเอพอลิเมอเรสที่มีประสิทธิภาพ สูง ในขณะที่สายตามจะถูกต่อขยายอย่างไม่ต่อเนื่องจากไพรเมอร์แต่ละตัว ทำให้เกิดชิ้นส่วนโอคาซากิ เอนไซม์อา ร์ เอ็นเอสจะกำจัดชิ้นส่วนไพรเมอร์ RNA ออกไป และเอนไซม์ดีเอ็นเอพอลิเมอเรสที่มีประสิทธิภาพต่ำ ซึ่งแตกต่างจากพอลิเมอเรสที่ใช้ในการจำลองแบบ จะเข้ามาเติมช่องว่าง เมื่อกระบวนการนี้เสร็จสมบูรณ์ จะพบรอยบิ่นหนึ่งรอยบนสายนำและรอยบิ่นหลายรอยบนสายตาม เอนไซม์ไลเกสจะทำหน้าที่เติมรอยบิ่นเหล่านี้ ทำให้โมเลกุลดีเอ็นเอที่จำลองแบบใหม่เสร็จสมบูรณ์
ไพรเมสที่ใช้ในกระบวนการนี้แตกต่างกันอย่างมากระหว่างแบคทีเรียและอาร์เคีย / ยูคาริโอตแบคทีเรียใช้ไพรเมสที่อยู่ใน กลุ่มโปรตีนซูเปอร์แฟมิลี DnaGซึ่งมีโดเมนเร่งปฏิกิริยาแบบพับ TOPRIM [ 35 ]โครงสร้างพับ TOPRIM ประกอบด้วยแกน α/β ที่มีสายอนุรักษ์สี่สายใน โทโพโลยี แบบ Rossmannโครงสร้างนี้ยังพบในโดเมนเร่งปฏิกิริยาของโทโพไอโซเมอเรส Ia โทโพไอโซเมอเรส II นิวคลีเอสในกลุ่ม OLD และโปรตีนซ่อมแซม DNA ที่เกี่ยวข้องกับโปรตีน RecR
ในทางตรงกันข้าม ไพรเมสที่ใช้โดยอาร์เคียและยูคาริโอตนั้นมีเวอร์ชันที่ดัดแปลงมาจากโมทีฟการจดจำ RNA (RRM) ไพรเมสนี้มีโครงสร้างคล้ายกับพอลิเมอเรส RNA ที่ขึ้นอยู่กับไวรัสหลายชนิด รีเวิร์สทรานสคริปเทส ไซเคลสที่สร้างนิวคลีโอไทด์แบบวงจร และพอลิเมอเรส DNA ของตระกูล A/B/Y ที่เกี่ยวข้องกับการจำลองแบบและการซ่อมแซม DNA ในการจำลองแบบของยูคาริโอต ไพรเมสจะสร้างคอมเพล็กซ์กับ Pol α [ 36 ]
เอนไซม์ DNA polymerase หลายชนิดมีบทบาทแตกต่างกันในกระบวนการจำลอง DNA ในแบคทีเรีย E. coliเอนไซม์DNA Pol IIIเป็นเอนไซม์ polymerase หลักที่รับผิดชอบในการจำลอง DNA มันจะรวมตัวกันเป็นคอมเพล็กซ์การจำลองที่จุดแยกการจำลอง ซึ่งมีประสิทธิภาพในการทำงานสูงมาก และคงสภาพเดิมตลอดวงจรการจำลอง ในทางตรงกันข้ามDNA Pol Iเป็นเอนไซม์ที่รับผิดชอบในการแทนที่ไพรเมอร์ RNA ด้วย DNA DNA Pol I มี กิจกรรม exonuclease จาก 5′ ไป 3′ นอกเหนือจากกิจกรรม polymerase และใช้กิจกรรม exonuclease ในการย่อยสลายไพรเมอร์ RNA ที่อยู่ข้างหน้าขณะที่มันขยายสาย DNA ที่อยู่ข้างหลัง ในกระบวนการที่เรียกว่าการแปลนิค (nick translation ) Pol I มีประสิทธิภาพในการทำงานน้อยกว่า Pol III มาก เนื่องจากหน้าที่หลักของมันในการจำลอง DNA คือการสร้างบริเวณ DNA สั้นๆ จำนวนมาก แทนที่จะสร้างบริเวณยาวๆ เพียงไม่กี่บริเวณ
ในยูคาริโอตเอนไซม์ที่มีกระบวนการต่ำอย่าง Pol α ช่วยเริ่มต้นการจำลองแบบเนื่องจากมันสร้างคอมเพล็กซ์กับไพรเมส[ 37 ] ในยูคาริโอต เชื่อกันว่าการสังเคราะห์สายนำจะดำเนินการโดย Pol ε อย่างไรก็ตาม มุมมองนี้เพิ่งถูกท้าทายเมื่อเร็ว ๆ นี้ โดยชี้ให้เห็นถึงบทบาทของ Pol δ [ 38 ] การกำจัดไพรเมอร์เสร็จสมบูรณ์โดย Pol δ [ 39 ]ในขณะที่การซ่อมแซม DNA ระหว่างการจำลองแบบเสร็จสมบูรณ์โดย Pol ε
เมื่อการสังเคราะห์ DNA ดำเนินต่อไป สาย DNA ดั้งเดิมจะคลายตัวออกที่แต่ละด้านของฟองอากาศ ทำให้เกิดส้อมการจำลองแบบที่มีสองง่าม ในแบคทีเรียซึ่งมีต้นกำเนิดการจำลองแบบเพียงจุดเดียวบนโครโมโซมวงกลม กระบวนการนี้จะสร้าง " โครงสร้างธีตา " (คล้ายกับอักษรกรีกธีตา: θ) ในทางตรงกันข้าม ยูคาริโอตมีโครโมโซมเชิงเส้นที่ยาวกว่าและเริ่มต้นการจำลองแบบที่ต้นกำเนิดหลายจุดภายในโครโมโซมเหล่านี้[ 40 ]
การจำลองแบบแยก


โครงสร้างการจำลองแบบ (replication fork) เป็นโครงสร้างที่เกิดขึ้นภายใน DNA เกลียวยาวในระหว่างการจำลองแบบ DNA เกิดจากเอนไซม์ที่เรียกว่าเฮลิเคส ซึ่งทำลายพันธะไฮโดรเจนที่ยึดสาย DNA เข้าด้วยกันเป็นเกลียว โครงสร้างที่ได้จะมี "กิ่ง" สองกิ่ง แต่ละกิ่งประกอบด้วยสาย DNA เพียงสายเดียว สายทั้งสองนี้ทำหน้าที่เป็นแม่แบบสำหรับสายนำและสายตาม ซึ่งจะถูกสร้างขึ้นเมื่อ DNA โพลีเมอเรสจับคู่นิวคลีโอไทด์ที่เสริมกับแม่แบบ แม่แบบเหล่านี้อาจเรียกได้ว่าแม่แบบสายนำและแม่แบบสายตามอย่างถูกต้อง[ 41 ]
ดีเอ็นเอจะถูกอ่านโดยเอนไซม์ดีเอ็นเอพอลิเมอเรสในทิศทาง 3′ ไป 5′ ซึ่งหมายความว่าสายใหม่จะถูกสังเคราะห์ในทิศทาง 5' ไป 3'เนื่องจากแม่แบบสายนำและสายตามวางตัวในทิศทางตรงกันข้ามที่จุดแยกการจำลองแบบ ปัญหาสำคัญคือจะสังเคราะห์ดีเอ็นเอสายตามใหม่ได้อย่างไร เนื่องจากทิศทางการสังเคราะห์ของสายตามนั้นตรงกันข้ามกับทิศทางการเติบโตของจุดแยกการจำลองแบบ
สายนำ
สายนำคือสาย DNA ใหม่ซึ่งถูกสังเคราะห์ในทิศทางเดียวกับส้อมการจำลองแบบที่กำลังเติบโต การจำลองแบบ DNA ประเภทนี้เกิดขึ้นอย่างต่อเนื่อง[ 42 ]
สายล้าหลัง
สายที่ล้าหลังคือสาย DNA ใหม่ที่มีทิศทางการสังเคราะห์ตรงข้ามกับทิศทางของส้อมการจำลองแบบที่กำลังเติบโต แม้ว่าปฏิกิริยาทางเคมีจะเหมือนกันทุกประการก็ตาม เนื่องจากทิศทางดังกล่าว การจำลองแบบของสายที่ล้าหลังจึงซับซ้อนกว่าเมื่อเทียบกับสายนำหน้า ผลที่ตามมาคือ DNA polymerase บนสายนี้จึงดูเหมือน "ล้าหลัง" สายอื่น[ 43 ]
สายล้าหลังจะถูกสังเคราะห์เป็นชิ้นส่วนสั้นๆ บนแม่แบบ สายล้าหลัง ไพร เมสจะ " อ่าน" DNA แม่แบบและเริ่มต้นการสังเคราะห์ ไพรเมอร์ RNA เสริมสั้นๆ ดีเอ็นเอพอ ลิเมอเรสจะขยายชิ้นส่วนไพรเมอร์ ทำให้เกิดชิ้นส่วนโอคาซากิจากนั้นไพรเมอร์ RNA จะถูกกำจัดออกโดยดีเอ็นเอพอลิเมอเรสอีกตัวหนึ่งและแทนที่ด้วย DNA และชิ้นส่วนของ DNA จะถูกเชื่อมต่อกันโดยดีเอ็นเอไลเกส[ 43 ]
พลวัตที่จุดแยกการจำลองแบบ
ในทุกกรณี เฮลิเคสประกอบด้วยโพลีเปปไทด์หกตัวที่พันรอบสาย DNA เพียงสายเดียวที่กำลังจำลองแบบ โพลีเมอเรสสองตัวจะจับกับเฮกซาเมอร์ของเฮลิเคส ในยูคาริโอต เฮลิเคสจะพันรอบสายนำ และในโปรคาริโอต เฮลิเคสจะพันรอบสายตาม[ 44 ]
เมื่อเฮลิเคสคลายเกลียว DNA ที่จุดแยกการจำลอง DNA ที่อยู่ข้างหน้าจะถูกบังคับให้หมุน กระบวนการนี้ส่งผลให้เกิดการบิดตัวสะสมใน DNA ที่อยู่ข้างหน้า[ 45 ]การบิดตัวสะสมนี้สร้างภาระแรงบิดที่จะหยุดจุดแยกการจำลองในที่สุด โทโปไอโซเมอเรสเป็นเอนไซม์ที่ทำลายสาย DNA ชั่วคราว บรรเทาความตึงเครียดที่เกิดจากการคลายเกลียวของสาย DNA สองสาย โทโปไอโซเมอเรส (รวมถึงDNA ไจเรส ) ทำเช่นนี้โดยการเพิ่มซูเปอร์คอยล์ เชิง ลบให้กับ DNA [ 46 ]
ดีเอ็นเอสายเดี่ยวเปล่ามีแนวโน้มที่จะพับกลับเข้าหากันเอง ทำให้เกิดโครงสร้างทุติยภูมิโครงสร้างเหล่านี้สามารถขัดขวางการเคลื่อนที่ของดีเอ็นเอพอลิเมอเรสได้ เพื่อป้องกันสิ่งนี้โปรตีนที่จับกับสายเดี่ยวจะจับกับดีเอ็นเอจนกว่าจะมีการสังเคราะห์สายที่สองขึ้นมา ซึ่งจะป้องกันการเกิดโครงสร้างทุติยภูมิ[ 47 ]
ดีเอ็นเอแบบสองสายจะขดตัวรอบฮิสโตนซึ่งมีบทบาทสำคัญในการควบคุมการแสดงออกของยีน ดังนั้นดีเอ็นเอที่จำลองแบบจะต้องขดตัวรอบฮิสโตนในตำแหน่งเดียวกับดีเอ็นเอเดิม[ 48 ]เพื่อให้แน่ใจในเรื่องนี้ ฮิสโตนชาเปอโรน จะแยก โครมาตินออกก่อนที่จะจำลองแบบและแทนที่ฮิสโตนในตำแหน่งที่ถูกต้อง ขั้นตอนบางอย่างในการประกอบใหม่นี้ยังเป็นการคาดเดาอยู่บ้าง[ 49 ]
โปรตีนแคลมป์ทำหน้าที่เป็นแคลมป์เลื่อนบน DNA ทำให้ DNA โพลีเมอเรสสามารถจับกับแม่แบบและช่วยในกระบวนการทำงาน ด้านในของแคลมป์ช่วยให้ DNA สามารถลอดผ่านได้ เมื่อโพลีเมอเรสไปถึงปลายแม่แบบหรือตรวจพบ DNA สองสาย แคลมป์เลื่อนจะเกิดการเปลี่ยนแปลงโครงสร้างที่ปล่อย DNA โพลีเมอเรส โปรตีนโหลดแคลมป์ใช้ในการโหลดแคลมป์ในขั้นต้น โดยจดจำจุดเชื่อมต่อระหว่างแม่แบบและไพรเมอร์ RNA [ 9 ] :274-5
โปรตีนที่เกี่ยวข้องกับการจำลองดีเอ็นเอ
ที่จุดแยกการจำลองแบบ เอนไซม์การจำลองแบบจำนวนมากจะประกอบกันบน DNA กลายเป็นเครื่องจักรโมเลกุลที่ซับซ้อนที่เรียกว่ารีพลิโซมต่อไปนี้คือรายชื่อเอนไซม์การจำลองแบบ DNA หลักที่เข้าร่วมในรีพลิโซม: [ 50 ]
| เอนไซม์ | หน้าที่ในการจำลองดีเอ็นเอ |
|---|---|
| ดีเอ็นเอเฮลิเคส | เฮลิเคส หรือที่รู้จักกันในชื่อเอนไซม์ที่ทำให้โครงสร้างเกลียวดีเอ็นเอไม่เสถียร ทำหน้าที่แยกสายดีเอ็นเอสองสายออกจากกันที่ จุดแยกสายดีเอ็นเอ (Replication Fork)ด้านหลังเอนไซม์โทโปไอโซเมอเรส |
| ดีเอ็นเอพอลิเมอเรส | เอนไซม์ที่ทำหน้าที่เร่งปฏิกิริยาการเติมสารตั้งต้นนิวคลีโอไทด์ลงในดีเอ็นเอในทิศทาง 5′ ไป 3′ ระหว่างการจำลองดีเอ็นเอ นอกจากนี้ยังทำหน้าที่ตรวจสอบความถูกต้องและแก้ไขข้อผิดพลาดด้วย มีดีเอ็นเอพอลิเมอเรสหลายประเภท ซึ่งแต่ละประเภททำหน้าที่แตกต่างกันในเซลล์ประเภทต่างๆ |
| ตัวยึดดีเอ็นเอ | โปรตีนที่ป้องกันไม่ให้เอนไซม์ DNA polymerase ที่กำลังยืดสาย DNA แยกตัวออกจากสาย DNA ต้นแบบ |
| โปรตีนที่จับกับดีเอ็นเอสายเดี่ยว | จับกับ ssDNA และป้องกันไม่ให้เกลียวคู่ของ DNA กลับมาเชื่อมต่อกันอีกครั้งหลังจากที่ DNA helicase คลายเกลียวออก จึงช่วยรักษาการแยกตัวของสาย DNA และอำนวยความสะดวกในการสังเคราะห์สาย DNA ใหม่ |
| โทโปไอโซเมอเรส | ช่วยคลายเกลียว DNA ที่ขดตัวอย่างแน่นหนา |
| ดีเอ็นเอไจเรส | ช่วยลดภาระจากการคลายเกลียวของดีเอ็นเอโดยเฮลิเคส ซึ่งเป็นโทโปไอโซเมอเรสชนิดหนึ่งโดยเฉพาะ |
| ดีเอ็นเอไลเกส | ทำการรีแอนนีลสายดีเอ็นเอแบบกึ่งอนุรักษ์ และเชื่อมต่อชิ้นส่วนโอคาซากิของสายดีเอ็นเอที่ล้าหลังเข้า ด้วยกัน |
| ไพรเมส | ทำหน้าที่เป็นจุดเริ่มต้นของ RNA (หรือ DNA) เพื่อให้เอนไซม์ DNA polymerase เริ่มสังเคราะห์สาย DNA ใหม่ |
| เทโลเมอเรส | เพิ่มความยาวของ DNA เทโลเมียร์โดยการเพิ่มลำดับนิวคลีโอไทด์ซ้ำๆ ที่ปลายโครโมโซมยูคาริโอตซึ่งช่วยให้เซลล์สืบพันธุ์และเซลล์ต้นกำเนิดหลีกเลี่ยงข้อจำกัดของ Hayflickในการแบ่งเซลล์[ 51 ] |
การทดลองโมเลกุลเดี่ยวในหลอดทดลอง (โดยใช้แหนบแสงและแหนบแม่เหล็ก ) พบปฏิสัมพันธ์แบบเสริมฤทธิ์กันระหว่างเอนไซม์รีพลิโซม (เฮลิเคสพอลิเมอเรสและโปรตีนที่จับกับดีเอ็นเอสายเดี่ยว ) และกับส้อมการจำลองดีเอ็นเอ ซึ่งช่วยเพิ่มการคลายเกลียวดีเอ็นเอและการจำลองดีเอ็นเอ [ 13 ]ผลลัพธ์เหล่านี้นำไปสู่การพัฒนารูปแบบจลนศาสตร์ที่อธิบายถึงปฏิสัมพันธ์แบบเสริมฤทธิ์กันและความเสถียรของพวกมัน [ 13 ]
เครื่องจักรจำลอง

รีพลิโซม หรือระบบการจำลองดีเอ็นเอประกอบด้วยปัจจัยที่เกี่ยวข้องกับการจำลองดีเอ็นเอและเชื่อมโยงกับแม่แบบ ssDNA รีพลิโซมประกอบด้วยดีเอ็นเอพอลิเมอเรส ดีเอ็นเอเฮลิเคส ดีเอ็นเอแคลมป์ และดีเอ็นเอโทโปไอโซเมอเรส รวมถึงโปรตีนการจำลอง เช่น โปรตีนจับดีเอ็นเอสายเดี่ยว (SSB) ในระหว่างการจำลองดีเอ็นเอ ส่วนประกอบเหล่านี้จะประสานงานกันในกิจกรรมของพวกมัน ในแบคทีเรียส่วนใหญ่ ปัจจัยทั้งหมดที่เกี่ยวข้องกับการจำลองดีเอ็นเอจะอยู่ที่จุดแยกการจำลองในระหว่างการจำลองดีเอ็นเอ ในเซลล์ยูคาริโอตและแบคทีเรียบางชนิด รีพลิโซมจะไม่เกิดขึ้น[ 52 ]
ในปี พ.ศ. 2523 บรูซ อัลเบิร์ตส์ ได้เสนอ "แบบจำลองทรอมโบน" ของการจำลองดีเอ็นเอเพื่อแสดงภาพการประสานงานระหว่างสายนำและสายตามของการสังเคราะห์ดีเอ็นเอ[ 53 ]ในแบบจำลองนี้ กลไกการจำลองจะรวมกลุ่มกันในขณะที่การสังเคราะห์สายนำไหลผ่าน และสายตามขยายเข้าและออกเหมือนสไลด์บนทรอมโบน ปีเตอร์ ไมสเตอร์และคณะได้สังเกตตำแหน่งการจำลองโดยตรงในยีสต์ที่กำลังแตกหน่อโดยการตรวจสอบ พอลิเมอเรสดีเอ็นเอ α ที่ติดแท็ก โปรตีนเรืองแสงสีเขียว (GFP) พวกเขาตรวจพบการจำลองดีเอ็นเอของคู่ของตำแหน่งที่ติดแท็กซึ่งเว้นระยะห่างกันอย่างสมมาตรจากจุดเริ่มต้นของการจำลอง และพบว่าระยะห่างระหว่างคู่ลดลงอย่างเห็นได้ชัดเมื่อเวลา ผ่านไป [ 54 ]การค้นพบนี้ชี้ให้เห็นว่ากลไกการจำลองดีเอ็นเอสอดคล้องกับแบบจำลองทรอมโบนที่มีกลุ่มของรีพลิโซมที่บรรจุอยู่บนจุดเริ่มต้นของการจำลอง ดำเนินการในทั้งสองทิศทาง การค้นพบของไมสเตอร์เป็นหลักฐานโดยตรงแรกของแบบจำลองทรอมโบน การวิจัยในภายหลังแสดงให้เห็นว่า DNA helicase ก่อตัวเป็นไดเมอร์ในเซลล์ยูคาริโอตจำนวนมาก และ replisome ของแบคทีเรียจะอยู่ในตำแหน่งภายในนิวเคลียสเดียวในระหว่างการสังเคราะห์ DNA [ 55 ]
โรงงานจำลองแบบแยกโครมาทิดคู่แฝด การแยกโครมาทิดเป็นสิ่งจำเป็นสำหรับการกระจายโครมาทิดไปยังเซลล์ลูกหลังจากการจำลองดีเอ็นเอ เนื่องจากโครมาทิดคู่แฝดหลังจากการจำลองดีเอ็นเอจะยึดกันด้วย วงแหวน โค ฮีซิ น โอกาสเดียวที่จะเกิดการแยกโครมาทิดคือการจำลองดีเอ็นเอ การแก้ไขกลไกการจำลองแบบให้เป็นโรงงานจำลองแบบสามารถปรับปรุงอัตราความสำเร็จของการจำลองดีเอ็นเอได้ หากส้อมจำลองแบบเคลื่อนที่ได้อย่างอิสระในโครโมโซม การเชื่อมต่อของนิวเคลียสจะรุนแรงขึ้นและขัดขวางการแยกตัวแบบไมโทซิส[ 54 ] [ 56 ]
การเลิกจ้าง
เซลล์ยูคาริโอตเริ่มต้นการจำลองดีเอ็นเอที่หลายจุดบนโครโมโซม ดังนั้นส้อมการจำลองจึงมาบรรจบกันและสิ้นสุดที่หลายจุดบนโครโมโซม เนื่องจากเซลล์ยูคาริโอตมีโครโมโซมเป็นเส้นตรง การจำลองดีเอ็นเอจึงไม่สามารถไปถึงปลายสุดของโครโมโซมได้ ด้วยเหตุนี้ ดีเอ็นเอจึงสูญหายไปในแต่ละรอบการจำลองจากปลายโครโมโซม เทโลเมียร์เป็นบริเวณของดีเอ็นเอที่ซ้ำกันใกล้กับปลายโครโมโซมและช่วยป้องกันการสูญเสียยีนเนื่องจากการหดตัวนี้ การหดตัวของเทโลเมียร์เป็นกระบวนการปกติในเซลล์ร่างกายซึ่งทำให้เทโลเมียร์ของโครโมโซมดีเอ็นเอของเซลล์ลูกสั้นลง ผลก็คือ เซลล์สามารถแบ่งตัวได้เพียงจำนวนครั้งที่จำกัดก่อนที่การสูญเสียดีเอ็นเอจะขัดขวางการแบ่งตัวต่อไป (สิ่งนี้เรียกว่าขีดจำกัดของเฮย์ฟลิค ) ภายใน สาย เซลล์สืบพันธุ์ซึ่งส่งต่อดีเอ็นเอไปยังรุ่นต่อไปเอนไซม์เทโลเมอเรสจะขยายลำดับที่ซ้ำกันของบริเวณเทโลเมียร์เพื่อป้องกันการเสื่อมสภาพ เทโลเมอเรสอาจทำงานผิดปกติในเซลล์ร่างกาย ซึ่งบางครั้งอาจนำไปสู่ การเกิด มะเร็งการเพิ่มขึ้นของกิจกรรมเทโลเมอเรสเป็นหนึ่งในลักษณะเด่นของมะเร็ง[ 57 ]
การยุติต้องทำให้ความคืบหน้าของการจำลองดีเอ็นเอหยุดลงหรือถูกบล็อก การยุติที่ตำแหน่งเฉพาะ เมื่อเกิดขึ้น จะเกี่ยวข้องกับปฏิสัมพันธ์ระหว่างองค์ประกอบสองส่วน: (1) ลำดับไซต์การยุติในดีเอ็นเอ และ (2) โปรตีนที่จับกับลำดับนี้เพื่อหยุดการจำลองดีเอ็นเอ ในแบคทีเรียหลายชนิด โปรตีนนี้เรียกว่าโปรตีนที่จับกับไซต์การยุติการจำลองดีเอ็นเอ หรือโปรตีนTer [ 58 ]
เนื่องจากแบคทีเรียมีโครโมโซมเป็นวงกลม การสิ้นสุดของการจำลองแบบจะเกิดขึ้นเมื่อส้อมการจำลองแบบทั้งสองมาบรรจบกันที่ปลายตรงข้ามของโครโมโซมแม่E. coliควบคุมกระบวนการนี้โดยใช้ลำดับการสิ้นสุด ซึ่งเมื่อจับกับโปรตีน Tusจะทำให้ส้อมการจำลองแบบผ่านไปได้เพียงทิศทางเดียวเท่านั้น ส่งผลให้ส้อมการจำลองแบบถูกจำกัดให้มาบรรจบกันภายในบริเวณการสิ้นสุดของโครโมโซมเสมอ[ 59 ]
ระเบียบข้อบังคับ
ยูคาริโอต
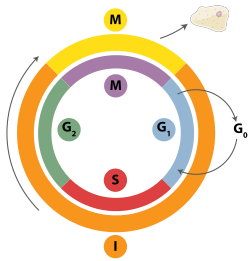
ภายในยูคาริโอต การจำลองดีเอ็นเอจะถูกควบคุมภายในบริบทของวงจรเซลล์เมื่อเซลล์เจริญเติบโตและแบ่งตัว มันจะดำเนินไปตามขั้นตอนต่างๆ ในวงจรเซลล์ การจำลองดีเอ็นเอเกิดขึ้นในช่วงระยะ S (ระยะสังเคราะห์) ความคืบหน้าของเซลล์ยูคาริโอตผ่านวงจรจะถูกควบคุมโดยจุดตรวจสอบวงจรเซลล์ความคืบหน้าผ่านจุดตรวจสอบจะถูกควบคุมผ่านปฏิสัมพันธ์ที่ซับซ้อนระหว่างโปรตีนต่างๆ รวมถึงไซคลินและไคเนสที่ขึ้นอยู่กับไซคลิน [ 60 ] แตกต่างจากแบคทีเรีย ดีเอ็นเอของยูคาริโอตจะจำลองตัวเองภายในนิวเคลียส[ 61 ]
จุดตรวจสอบ G1/S (จุดตรวจสอบการจำกัด) ควบคุมว่าเซลล์ยูคาริโอติกจะเข้าสู่กระบวนการจำลองดีเอ็นเอและการแบ่งตัวในภายหลังหรือไม่ เซลล์ที่ไม่ผ่านจุดตรวจสอบนี้จะยังคงอยู่ในระยะ G0 และไม่จำลองดีเอ็นเอ[ 62 ]
เมื่อ DNA ผ่านการทดสอบ "G1/S" แล้ว จะสามารถคัดลอกได้เพียงครั้งเดียวในแต่ละรอบเซลล์ เมื่อคอมเพล็กซ์ Mcm เคลื่อนตัวออกจากจุดเริ่มต้น คอมเพล็กซ์ก่อนการจำลองแบบจะถูกแยกส่วน เนื่องจากคอมเพล็กซ์ Mcm ใหม่ไม่สามารถโหลดที่จุดเริ่มต้นได้จนกว่าหน่วยย่อยก่อนการจำลองแบบจะถูกกระตุ้นใหม่ จุดเริ่มต้นของการจำลองแบบหนึ่งจุดจึงไม่สามารถใช้ซ้ำได้สองครั้งในรอบเซลล์เดียวกัน[ 33 ]
การเปิดใช้งาน S-Cdks ในระยะ S ตอนต้นจะส่งเสริมการทำลายหรือการยับยั้งส่วนประกอบของคอมเพล็กซ์ก่อนการจำลองแบบแต่ละส่วน ป้องกันการประกอบใหม่ในทันที S และ M-Cdks ยังคงปิดกั้นการประกอบคอมเพล็กซ์ก่อนการจำลองแบบแม้หลังจากระยะ S เสร็จสมบูรณ์แล้ว ทำให้มั่นใจได้ว่าการประกอบจะไม่เกิดขึ้นอีกจนกว่ากิจกรรม Cdk ทั้งหมดจะลดลงในระยะไมโทซิสตอนปลาย[ 33 ]
ในยีสต์ที่กำลังแตกหน่อ การยับยั้งการประกอบเกิดจากการฟอสโฟรีเลชันของส่วนประกอบของคอมเพล็กซ์ก่อนการจำลองแบบโดยอาศัย Cdk ในช่วงเริ่มต้นของระยะ S การฟอสโฟรีเลชันของ Cdc6 โดยCdk1ทำให้ Cdc6 จับกับโปรตีนไลเกสยูบิควิตินSCF ซึ่งทำให้เกิดการทำลาย Cdc6 ด้วยเอนไซม์โปรตีเอส การฟอสโฟรีเลชันของโปรตีน Mcm โดยอาศัย Cdk ส่งเสริมการส่งออกโปรตีน Mcm ออกจากนิวเคลียสพร้อมกับ Cdt1 ในระหว่างระยะ S ป้องกันการโหลดคอมเพล็กซ์ Mcm ใหม่ที่ต้นกำเนิดในระหว่างรอบเซลล์เดียว การฟอสโฟรีเลชันของคอมเพล็กซ์การจำลองแบบที่ต้นกำเนิดโดย Cdk ยังยับยั้งการประกอบคอมเพล็กซ์ก่อนการจำลองแบบด้วย การมีกลไกใดกลไกหนึ่งในสามกลไกนี้ก็เพียงพอที่จะยับยั้งการประกอบคอมเพล็กซ์ก่อนการจำลองแบบ อย่างไรก็ตาม การกลายพันธุ์ของโปรตีนทั้งสามชนิดในเซลล์เดียวกันจะกระตุ้นการเริ่มต้นใหม่ที่ต้นกำเนิดการจำลองแบบหลายแห่งภายในรอบเซลล์เดียว[ 33 ] [ 63 ]
ในเซลล์สัตว์ โปรตีนgemininเป็นตัวยับยั้งที่สำคัญของการประกอบคอมเพล็กซ์ก่อนการจำลองแบบ Geminin จับกับ Cdt1 ป้องกันไม่ให้ Cdt1 จับกับคอมเพล็กซ์การจดจำต้นกำเนิด ในระยะ G1 ระดับของ geminin จะถูกควบคุมให้ต่ำโดย APC ซึ่งจะทำการ ubiquitinate geminin เพื่อกำหนดเป้าหมายสำหรับการย่อยสลาย เมื่อ geminin ถูกทำลาย Cdt1 จะถูกปล่อยออกมา ทำให้สามารถทำหน้าที่ในการประกอบคอมเพล็กซ์ก่อนการจำลองแบบได้ ในตอนท้ายของระยะ G1 APC จะถูกปิดใช้งาน ทำให้ geminin สะสมและจับกับ Cdt1 ได้[ 33 ]
การจำลองแบบของจีโนมคลอโรพลาสต์และไมโตคอนเดรียเกิดขึ้นโดยอิสระจากวงจรเซลล์ ผ่านกระบวนการจำลองแบบD -loop [ 64 ]
จุดเน้นการจำลองแบบ
ในเซลล์ของสัตว์มีกระดูกสันหลัง ตำแหน่งการจำลองแบบจะรวมตัวกันที่ตำแหน่งที่เรียกว่าจุดรวมการจำลองแบบ [ 54 ] สามารถตรวจจับตำแหน่งการจำลองแบบได้โดยการย้อมภูมิคุ้มกันของสายลูกสาวและเอนไซม์การจำลองแบบ และการตรวจสอบปัจจัยการจำลองแบบที่ติดแท็ก GFP ด้วยวิธีการเหล่านี้ พบว่าจุดรวมการจำลองแบบที่มีขนาดและตำแหน่งที่แตกต่างกันปรากฏขึ้นในระยะ S ของการแบ่งเซลล์ และจำนวนของจุดรวมการจำลองแบบต่อหนึ่งนิวเคลียสนั้นน้อยกว่าจำนวนของส้อมการจำลองแบบจีโนมมาก
P. Heun และคณะติดตามจุดโฟกัสการจำลองแบบที่ติดแท็ก GFP ในเซลล์ยีสต์ที่กำลังแตกหน่อ และพบว่าต้นกำเนิดการจำลองแบบเคลื่อนที่อย่างต่อเนื่องในระยะ G1 และ S และพลวัตลดลงอย่างมีนัยสำคัญในระยะ S [ 54 ]ตามธรรมเนียมแล้ว ตำแหน่งการจำลองแบบจะถูกตรึงไว้บนโครงสร้างเชิงพื้นที่ของโครโมโซมโดยเมทริกซ์นิวเคลียร์หรือลามินผลลัพธ์ของ Heun ปฏิเสธแนวคิดดั้งเดิม ยีสต์ที่กำลังแตกหน่อไม่มีลามิน และสนับสนุนว่าต้นกำเนิดการจำลองแบบประกอบตัวเองและก่อตัวเป็นจุดโฟกัสการจำลองแบบ[ 65 ]
การก่อตัวของจุดโฟกัสการจำลองแบบถูกควบคุมโดยการยิงของต้นกำเนิดการจำลองแบบ ซึ่งควบคุมทั้งในเชิงพื้นที่และเวลา DA Jackson และคณะ (1998) เปิดเผยว่าต้นกำเนิดที่อยู่ใกล้เคียงจะยิงพร้อมกันในเซลล์ของสัตว์เลี้ยงลูกด้วยนม[ 54 ]การเรียงตัวกันในเชิงพื้นที่ของไซต์การจำลองแบบทำให้เกิดการรวมกลุ่มของส้อมการจำลองแบบ การรวมกลุ่มนี้ช่วยกู้คืนส้อมการจำลองแบบที่หยุดชะงักและส่งเสริมความคืบหน้าตามปกติของส้อมการจำลองแบบ ความคืบหน้าของส้อมการจำลองแบบถูกยับยั้งโดยหลายปัจจัย เช่น การชนกับโปรตีนหรือกับสารประกอบที่จับกับ DNA อย่างแน่นหนา การขาดแคลน dNTPs รอยแตกบน DNA แม่แบบ และอื่นๆ หากส้อมการจำลองแบบติดอยู่และลำดับที่เหลือจากส้อมที่ติดอยู่ไม่ได้ถูกคัดลอก สายลูกสาวจะได้รับไซต์ที่ไม่ได้รับการจำลองแบบที่มีรอยแตก ไซต์ที่ไม่ได้รับการจำลองแบบบนสายแม่สายหนึ่งจะยึดสายอื่นไว้ด้วยกัน แต่ไม่ใช่สายลูกสาว ดังนั้น โครมาทิดคู่ที่เกิดขึ้นจึงไม่สามารถแยกออกจากกันและไม่สามารถแบ่งตัวเป็นเซลล์ลูก 2 เซลล์ได้ เมื่อต้นกำเนิดที่อยู่ใกล้เคียงเริ่มทำงานและส้อมจากต้นกำเนิดหนึ่งหยุดชะงัก ส้อมจากต้นกำเนิดอื่นจะเข้าถึงในทิศทางตรงกันข้ามกับส้อมที่หยุดชะงักและทำสำเนาไซต์ที่ยังไม่ได้ทำสำเนา กลไกการกู้คืนอื่น ๆ คือการใช้ต้นกำเนิดการจำลองแบบที่อยู่เฉยๆซึ่งต้นกำเนิดส่วนเกินจะไม่ทำงานในการจำลองแบบ DNA ปกติ[ 66 ]
แบคทีเรีย

แบคทีเรียส่วนใหญ่ไม่ได้ผ่านวงจรเซลล์ที่กำหนดไว้อย่างชัดเจน แต่จะคัดลอก DNA อย่างต่อเนื่อง ในระหว่างการเจริญเติบโตอย่างรวดเร็ว อาจส่งผลให้เกิดการจำลองแบบหลายรอบพร้อมกัน[ 67 ] ในE. coliซึ่งเป็นแบคทีเรียที่ได้รับการศึกษาอย่างละเอียดที่สุด การจำลองแบบ DNA ถูกควบคุมผ่านกลไกหลายอย่าง ได้แก่ การเติมหมู่เมทิลครึ่งหนึ่งและการกักเก็บลำดับต้นกำเนิด อัตราส่วนของอะดีโนซีนไตรฟอสเฟต (ATP)ต่ออะดีโนซีนไดฟอสเฟต (ADP)และระดับของโปรตีน DnaA ทั้งหมดนี้ควบคุมการจับกันของโปรตีนเริ่มต้นกับลำดับต้นกำเนิด[ 68 ]
เนื่องจากE. coli ทำการเมทิลเลชันลำดับ DNA GATC การสังเคราะห์ DNA จึงส่งผลให้เกิดลำดับเฮมิเมทิลเลชัน DNA เฮมิเมทิลเลชันนี้ได้รับการจดจำโดยโปรตีนSeqAซึ่งจะจับและกักเก็บลำดับต้นกำเนิด นอกจากนี้ DnaA (ซึ่งจำเป็นสำหรับการเริ่มต้นการจำลองแบบ) จะจับกับ DNA เฮมิเมทิลเลชันได้น้อยลง ส่งผลให้ต้นกำเนิดที่จำลองแบบใหม่ไม่สามารถเริ่มต้นการจำลองแบบ DNA รอบต่อไปได้ทันที[ 69 ]
ATP สะสมขึ้นเมื่อเซลล์อยู่ในสภาพแวดล้อมที่อุดมสมบูรณ์ ซึ่งจะกระตุ้นการจำลอง DNA เมื่อเซลล์มีขนาดถึงขนาดที่กำหนด ATP จะแข่งขันกับ ADP ในการจับกับ DnaA และคอมเพล็กซ์ DnaA-ATP จะสามารถเริ่มต้นการจำลองได้ นอกจากนี้ยังต้องการโปรตีน DnaA จำนวนหนึ่งสำหรับการจำลอง DNA — ทุกครั้งที่มีการคัดลอกต้นกำเนิด จำนวนตำแหน่งการจับของ DnaA จะเพิ่มขึ้นเป็นสองเท่า ทำให้ต้องสังเคราะห์ DnaA เพิ่มขึ้นเพื่อให้สามารถเริ่มต้นการจำลองอีกครั้งได้[ 70 ]
ในแบคทีเรียที่เติบโตอย่างรวดเร็ว เช่นE. coliการจำลองโครโมโซมใช้เวลานานกว่าการแบ่งเซลล์ แบคทีเรียแก้ปัญหานี้โดยเริ่มการจำลองรอบใหม่ก่อนที่รอบก่อนหน้าจะสิ้นสุดลง[ 71 ]การจำลองรอบใหม่นี้จะสร้างโครโมโซมของเซลล์ที่เกิดในสองรุ่นหลังจากเซลล์ที่แบ่งตัว กลไกนี้สร้างวงจรการจำลองที่ทับซ้อนกัน
ปัญหาเกี่ยวกับการจำลองดีเอ็นเอ


มีเหตุการณ์หลายอย่างที่ก่อให้เกิดความเครียดในการจำลองแบบ รวมถึง: [ 72 ]
- การใส่ไรโบนิวคลีโอไทด์ผิดตำแหน่ง
- โครงสร้าง DNAที่ผิดปกติ
- ความขัดแย้งระหว่างการจำลองแบบและการถอดรหัส
- ความไม่เพียงพอของปัจจัยการจำลองแบบที่จำเป็น
- จุดเปราะบางทั่วไป
- การแสดงออกมากเกินไปหรือการทำงานอย่างต่อเนื่องของยีนก่อ มะเร็ง
- โครมาตินไม่สามารถเข้าถึงได้
ปฏิกิริยาลูกโซ่พอลิเมอเรส
นักวิจัยมักจำลอง DNA ในหลอดทดลองโดยใช้ปฏิกิริยาลูกโซ่พอลิเมอเรส (PCR) PCR ใช้ ไพรเมอร์คู่หนึ่งเพื่อครอบคลุมบริเวณเป้าหมายใน DNA แม่แบบ จากนั้นจึงพอลิเมอไรซ์สายคู่ในแต่ละทิศทางจากไพรเมอร์เหล่านี้โดยใช้DNA พอลิเมอเรสที่ทนความร้อนการทำซ้ำกระบวนการนี้ผ่านหลายรอบจะขยายบริเวณ DNA เป้าหมาย ในตอนเริ่มต้นของแต่ละรอบ ส่วนผสมของแม่แบบและไพรเมอร์จะถูกให้ความร้อนเพื่อแยกโมเลกุลที่สังเคราะห์ขึ้นใหม่และแม่แบบ จากนั้นเมื่อส่วนผสมเย็นลง ทั้งสองอย่างจะกลายเป็นแม่แบบสำหรับการจับคู่ของไพรเมอร์ใหม่ และพอลิเมอเรสจะขยายจากสิ่งเหล่านี้ ส่งผลให้จำนวนสำเนาของบริเวณเป้าหมายเพิ่มขึ้นเป็นสองเท่าในแต่ละรอบเพิ่มขึ้นแบบทวีคูณ[ 73 ]
ดูเพิ่มเติม
- ออโตโพเอซิส
- เซลล์ (ชีววิทยา)
- การแบ่งเซลล์
- การแยกโครโมโซม
- อุปกรณ์จัดเก็บข้อมูล
- ยีน
- การแสดงออกของยีน
- เอพิเจเนติกส์
- จีโนม
- ดีเอ็นเอฮาจิโมจิ
- ชีวิต
- การจำลองแบบ (การคำนวณ)
- การจำลองตัวเอง
หมายเหตุ
- ^พลังงาน ของกระบวนการนี้อาจช่วยอธิบายทิศทางการสังเคราะห์ได้เช่นกัน—หาก DNA ถูกสังเคราะห์ในทิศทาง 3′ ไป 5′ พลังงานสำหรับกระบวนการจะมาจากปลาย 5′ ของสายที่กำลังเติบโต แทนที่จะ มาจากนิวคลีโอไทด์อิสระ ปัญหาคือ หากไตรฟอสเฟตที่มีพลังงานสูงอยู่บนสายที่กำลังเติบโตและไม่ได้อยู่บนนิวคลีโอไทด์อิสระ การตรวจสอบความถูกต้องโดยการกำจัดนิวคลีโอไทด์ปลายที่ไม่ตรงกันจะเป็นปัญหา: เมื่อมีการเพิ่มนิวคลีโอไทด์ ไตรฟอสเฟตจะหายไปและฟอสเฟตเดี่ยวจะยังคงอยู่บนโครงสร้างหลักระหว่างนิวคลีโอไทด์ใหม่กับส่วนที่เหลือของสาย หากนิวคลีโอไทด์ที่เพิ่มเข้ามาไม่ตรงกัน การกำจัดจะส่งผลให้สาย DNA สิ้นสุดด้วยโมโนฟอสเฟตที่ปลายของ "สายที่กำลังเติบโต" แทนที่จะเป็นไตรฟอสเฟตที่มีพลังงานสูง สายนั้นจึงจะติดอยู่และไม่สามารถเติบโตได้อีกต่อไป ในความเป็นจริง ไตรฟอสเฟตพลังงานสูงที่ถูกไฮโดรไลซ์ในแต่ละขั้นตอนนั้นมาจากนิวคลีโอไทด์อิสระ ไม่ใช่จากสายพอลิเมอร์ ดังนั้นปัญหาดังกล่าวจึงไม่มีอยู่จริง
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การจำลองดีเอ็นเอ
การจำลองดีเอ็นเอ เป็น กระบวนการ ที่ เซลล์ สร้างสำเนา ดีเอ็นเอ ของตัวเองอย่างแม่นยำ[ 1 ] [ 2 ] [ 3 ] [ 4 ] กระบวนการ นี้เกิดขึ้นใน สิ่งมีชีวิต ทุกชนิด และมีความสำคัญต่อ...
โครงสร้างดีเอ็นเอ
ดีเอ็นเอเป็นโครงสร้างแบบสองสาย โดยทั้งสองสายจะขดเข้าด้วยกันเพื่อสร้าง เกลียวคู่ ที่เป็นลักษณะเฉพาะ แต่ละสายเดี่ยวของดีเอ็นเอเป็นโซ่ของนิวคลีโอไทด์สี่ชนิด นิ วคลีโอไท ด์ในดีเอ็นเอประกอบด้วยน้ำตาล ดีออกซีไร โบส ฟอสเฟต และ นิวคลีโอเบส นิวคลีโอไทด์...
ดีเอ็นเอพอลิเมอเรส
ดีเอ็นเอพอลิเมอเรส เป็นกลุ่มของ เอนไซม์ ที่ทำหน้าที่ในการจำลองดีเอ็นเอทุกรูปแบบ [ 22 ] โดยทั่วไปแล้ว ดีเอ็นเอพอลิเมอเรสไม่สามารถเริ่มต้นการสังเคราะห์สายใหม่ได้ แต่สามารถขยายสายดีเอ็นเอหรืออาร์เอ็นเอที่มีอยู่ซึ่งจับคู่กับสายแม่แบบได้เท่านั้น...
กระบวนการจำลองแบบ
การจำลองดีเอ็นเอ เช่นเดียวกับกระบวนการพอลิเมอไรเซชันทางชีวภาพทั้งหมด ดำเนินไปในสามขั้นตอนที่เร่งปฏิกิริยาและประสานงานกันโดยเอนไซม์ ได้แก่ การเริ่มต้น การยืดออก และการสิ้นสุด [ 26 ]