แบบจำลองเซลล์ประสาททางชีววิทยา
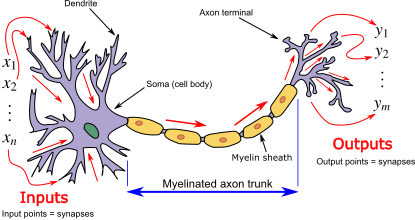

แบบจำลองเซลล์ประสาททางชีววิทยาหรือที่รู้จักกันในชื่อแบบจำลองเซลล์ประสาทแบบสไปค์ [ 1 ] เป็น คำอธิบายทางคณิตศาสตร์ของการนำสัญญาณไฟฟ้าในเซลล์ประสาทเซลล์ประสาท (หรือเซลล์ประสาท) เป็นเซลล์ ที่สามารถ ถูกกระตุ้นด้วยไฟฟ้า ภายใน ระบบประสาทสามารถส่งสัญญาณไฟฟ้าที่เรียกว่าศักย์การกระทำข้ามเครือข่ายประสาทแบบจำลองทางคณิตศาสตร์เหล่านี้อธิบายบทบาทของลักษณะทางชีวฟิสิกส์และเรขาคณิตของเซลล์ประสาทในการนำกิจกรรมทางไฟฟ้า
หัวใจสำคัญของแบบจำลองเหล่านี้คือการอธิบายว่าศักย์เยื่อหุ้มเซลล์ (นั่นคือ ความแตกต่างของศักย์ไฟฟ้าระหว่างภายในและภายนอกเซลล์ ชีวภาพ ) ข้ามเยื่อหุ้มเซลล์เปลี่ยนแปลงไปอย่างไรเมื่อเวลาผ่านไป ในการทดลอง การกระตุ้นเซลล์ประสาทด้วยกระแสไฟฟ้าจะสร้างศักย์การกระทำ (หรือสไปค์) ที่แพร่กระจายไปตาม แอกซอนของเซลล์ประสาทแอกซอนนี้สามารถแตกแขนงออกและเชื่อมต่อกับเซลล์ประสาทปลายทางจำนวนมากที่บริเวณที่เรียกว่าไซแนปส์ที่ไซแนปส์เหล่านี้ สไปค์สามารถทำให้เกิดการปล่อยสารสื่อประสาทซึ่งในทางกลับกันสามารถเปลี่ยนแปลงศักย์ไฟฟ้าของเซลล์ประสาทปลายทาง การเปลี่ยนแปลงนี้อาจนำไปสู่สไปค์ที่มากขึ้นในเซลล์ประสาทปลายทางเหล่านั้น จึงเป็นการส่งสัญญาณต่อไป เซลล์ประสาทมากถึง 95% ในนีโอคอร์เทกซ์ซึ่งเป็นชั้นนอกสุดของสมองสัตว์เลี้ยงลูกด้วยนม ประกอบด้วยเซลล์ประสาทพีระมิด ที่กระตุ้น [ 2 ] [ 3 ] และเซลล์ประสาทพีระมิดแต่ละ เซลล์จะได้รับอินพุตหลายหมื่นรายการจากเซลล์ประสาทอื่น ๆ[ 4 ]ดังนั้น เซลล์ประสาทที่ส่งสัญญาณจึงเป็นหน่วยประมวลผลข้อมูลหลักของระบบประสาท
ตัวอย่างหนึ่งของแบบจำลองเซลล์ประสาทแบบสไปค์อาจเป็นแบบจำลองทางคณิตศาสตร์ที่มีรายละเอียดสูงซึ่งรวมถึงสัณฐานวิทยา เชิงพื้นที่ อีกตัวอย่างหนึ่งอาจเป็นแบบจำลองเซลล์ประสาทแบบอิงการนำไฟฟ้าที่มองเซลล์ประสาทเป็นจุดและอธิบายพลวัตของแรงดันไฟฟ้าของเยื่อหุ้มเซลล์เป็นฟังก์ชันของกระแสไฟฟ้าข้ามเยื่อหุ้มเซลล์ แบบจำลอง "รวมและยิง" ที่เรียบง่ายกว่าทางคณิตศาสตร์ช่วยลดความซับซ้อนของการอธิบายช่องไอออนและพลวัตของศักย์เยื่อหุ้มเซลล์ได้อย่างมาก (ซึ่งศึกษาครั้งแรกโดย Lapique ในปี 1907) [ 5 ] [ 6 ]
พื้นฐานทางชีววิทยา การจำแนกประเภท และเป้าหมายของแบบจำลองเซลล์ประสาท
เซลล์ที่ไม่สร้างสไปค์ เซลล์ที่สร้างสไปค์ และการวัดค่าของเซลล์เหล่านั้น
ไม่ใช่เซลล์ทั้งหมดในระบบประสาทจะสร้างสัญญาณไฟฟ้าแบบที่กำหนดขอบเขตของแบบจำลองเซลล์ประสาทที่สร้างสัญญาณไฟฟ้าได้ ตัวอย่างเช่นเซลล์ขนในหูชั้นใน เซลล์รับสัญญาณในจอประสาทตาและเซลล์ไบโพลาร์ในจอประสาทตา ไม่สร้างสัญญาณไฟฟ้า นอกจากนี้ เซลล์จำนวนมากในระบบประสาทไม่ได้ถูกจัดประเภทเป็นเซลล์ประสาท แต่ถูกจัดประเภทเป็น เซลล์เกลียแทน
กิจกรรมของเซลล์ประสาทสามารถวัดได้ด้วยเทคนิคการทดลองต่างๆ เช่น เทคนิคการวัดแบบ "เซลล์ทั้งหมด" ซึ่งบันทึกกิจกรรมการส่งสัญญาณของเซลล์ประสาทเดี่ยวและสร้างศักยภาพการกระทำที่มีแอมพลิจูดเต็มที่
ด้วยเทคนิคการวัดนอกเซลล์ จะมีการวางอิเล็กโทรดหนึ่งตัวหรือมากกว่านั้นในช่องว่างนอกเซลล์สัญญาณกระตุ้น ซึ่งมักมาจากแหล่งกำเนิดสัญญาณกระตุ้นหลายแหล่ง ขึ้นอยู่กับขนาดของอิเล็กโทรดและความใกล้ชิดกับแหล่งกำเนิดสัญญาณ สามารถระบุได้ด้วยเทคนิคการประมวลผลสัญญาณ การวัดนอกเซลล์มีข้อดีหลายประการ:
- สามารถหาได้ง่ายกว่าโดยวิธีการทดลอง
- มันแข็งแรงทนทานและใช้งานได้นานกว่า
- มันสามารถสะท้อนถึงผลกระทบที่เด่นชัด โดยเฉพาะอย่างยิ่งเมื่อทำการทดสอบในบริเวณทางกายวิภาคที่มีเซลล์ที่คล้ายคลึงกันจำนวนมาก
ภาพรวมของแบบจำลองเซลล์ประสาท
แบบจำลองนิวรอนสามารถแบ่งออกได้เป็นสองประเภทตามหน่วยทางกายภาพของส่วนต่อประสานของแบบจำลอง แต่ละประเภทสามารถแบ่งย่อยออกไปได้อีกตามระดับนามธรรม/รายละเอียด:
- แบบจำลองแรงดันไฟฟ้าขาเข้า-ขาออกของเยื่อหุ้มเซลล์ – แบบจำลองเหล่านี้สร้างการทำนายแรงดันไฟฟ้าขาออกของเยื่อหุ้มเซลล์เป็นฟังก์ชันของการกระตุ้นทางไฟฟ้าที่ให้มาในรูปของกระแสหรือแรงดันไฟฟ้าขาเข้า แบบจำลองต่างๆ ในหมวดหมู่นี้แตกต่างกันในความสัมพันธ์เชิงฟังก์ชันที่แน่นอนระหว่างกระแสขาเข้าและแรงดันไฟฟ้าขาออก และในระดับรายละเอียด แบบจำลองบางแบบในหมวดหมู่นี้ทำนายเฉพาะช่วงเวลาที่เกิดสไปค์ขาออก (หรือที่เรียกว่า "ศักย์การกระทำ") เท่านั้น ในขณะที่แบบจำลองอื่นๆ มีรายละเอียดมากกว่าและคำนึงถึงกระบวนการระดับเซลล์ย่อย แบบจำลองในหมวดหมู่นี้อาจเป็นได้ทั้งแบบกำหนดหรือแบบความน่าจะเป็น
- แบบจำลองเซลล์ประสาทที่รับสิ่งเร้า จากธรรมชาติหรือสารเภสัชกรรม – แบบจำลองในหมวดหมู่นี้เชื่อมต่อสิ่งเร้าขาเข้า ซึ่งอาจเป็นสารเภสัชกรรมหรือสิ่งเร้าจากธรรมชาติ เข้ากับความน่าจะเป็นของการเกิดสไปค์ ขั้นตอนการป้อนข้อมูลของแบบจำลองเหล่านี้ไม่ใช่ไฟฟ้า แต่มีหน่วยเป็นความเข้มข้นของสารเภสัชกรรม (สารเคมี) หรือหน่วยทางกายภาพที่บ่งบอกลักษณะของสิ่งเร้าภายนอก เช่น แสง เสียง หรือแรงกดทางกายภาพรูปแบบอื่นๆ นอกจากนี้ ขั้นตอนการส่งออกจะแสดงถึงความน่าจะเป็นของการเกิดสไปค์ ไม่ใช่แรงดันไฟฟ้า
แม้ว่าในวิทยาศาสตร์และวิศวกรรมศาสตร์ การมีแบบจำลองเชิงพรรณนาหลายแบบสำหรับระดับนามธรรม/รายละเอียดที่แตกต่างกันนั้นไม่ใช่เรื่องผิดปกติ แต่จำนวนแบบจำลองเซลล์ประสาททางชีววิทยาที่แตกต่างกันและบางครั้งขัดแย้งกันนั้นสูงมากเป็นพิเศษ สถานการณ์นี้เป็นผลส่วนหนึ่งมาจากการตั้งค่าการทดลองที่หลากหลาย และความยากลำบากในการแยกคุณสมบัติที่แท้จริงของเซลล์ประสาทแต่ละเซลล์ออกจากผลกระทบจากการวัดและการปฏิสัมพันธ์ของเซลล์จำนวนมาก ( ผลกระทบ จากเครือข่าย )
จุดมุ่งหมายของแบบจำลองเซลล์ประสาท
ในที่สุด แบบจำลองเซลล์ประสาททางชีววิทยามุ่งที่จะอธิบายกลไกที่อยู่เบื้องหลังการทำงานของระบบประสาท อย่างไรก็ตาม สามารถแยกแยะแนวทางต่างๆ ได้หลายแบบ ตั้งแต่แบบจำลองที่สมจริงมากขึ้น (เช่น แบบจำลองเชิงกลไก) ไปจนถึงแบบจำลองที่ใช้งานได้จริงมากขึ้น (เช่น แบบจำลองเชิงปรากฏการณ์) [ 7 ]การสร้างแบบจำลองช่วยในการวิเคราะห์ข้อมูลการทดลองและตอบคำถามต่างๆ แบบจำลองยังมีความสำคัญในบริบทของการฟื้นฟูการทำงานของสมองที่สูญเสียไปผ่านอุปกรณ์ประสาทเทียม
แบบจำลองแรงดันไฟฟ้าของเมมเบรนขาเข้า-ขาออกทางไฟฟ้า
แบบจำลองในหมวดหมู่นี้อธิบายความสัมพันธ์ระหว่างกระแสเมมเบรนประสาทที่ขั้นตอนอินพุตและแรงดันไฟฟ้าเมมเบรนที่ขั้นตอนเอาต์พุต หมวดหมู่นี้รวมถึงแบบจำลองอินทิเกรตแอนด์ไฟร์ (ทั่วไป) และแบบจำลองทางชีวฟิสิกส์ที่ได้รับแรงบันดาลใจจากงานของ Hodgkin–Huxley ในช่วงต้นทศวรรษ 1950 โดยใช้การตั้งค่าการทดลองที่เจาะเมมเบรนเซลล์และอนุญาตให้บังคับแรงดันไฟฟ้า/กระแสเมมเบรนที่เฉพาะเจาะจง[ 8 ] [ 9 ] [ 10 ] [ 11 ]
อินเทอร์เฟซประสาทไฟฟ้าสมัยใหม่ส่วนใหญ่ใช้การกระตุ้นไฟฟ้าภายนอกเซลล์เพื่อหลีกเลี่ยงการเจาะเยื่อหุ้มเซลล์ ซึ่งอาจนำไปสู่การตายของเซลล์และความเสียหายของเนื้อเยื่อ ดังนั้นจึงไม่ชัดเจนว่าแบบจำลองเซลล์ประสาทไฟฟ้าจะใช้ได้กับการกระตุ้นภายนอกเซลล์ในระดับใด (ดูเช่น[ 12 ] )
ฮอดจ์กิน-ฮักซ์ลีย์
| ทรัพย์สินของแบบจำลอง H&H | อ้างอิง |
|---|---|
| รูปร่างของหนามแต่ละอัน | [ 8 ] [ 9 ] [ 10 ] [ 11 ] |
| เอกลักษณ์ของไอออนที่เกี่ยวข้อง | [ 8 ] [ 9 ] [ 10 ] [ 11 ] |
| ความเร็วของการเกิดสไปค์ข้ามแอกซอน | [ 8 ] |
แบบจำลอง Hodgkin–Huxley (แบบจำลอง H&H) [ 8 ] [ 9 ] [ 10 ] [ 11 ] เป็นแบบจำลองความสัมพันธ์ระหว่างการไหลของกระแสไอออนิกข้ามเยื่อหุ้มเซลล์ประสาทและแรงดันไฟฟ้าของเยื่อหุ้มเซลล์[ 8 ] [ 9 ] [ 10 ] [ 11 ]ประกอบด้วยชุดสมการเชิงอนุพันธ์ไม่เชิงเส้นที่อธิบายพฤติกรรมของช่องไอออนที่แทรกซึมผ่านเยื่อหุ้มเซลล์ของแอกซอนยักษ์ของปลาหมึก Hodgkin และ Huxley ได้รับรางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ในปี 1963 จากผลงานนี้
สิ่งสำคัญคือต้องสังเกตความสัมพันธ์ระหว่างแรงดันไฟฟ้าและกระแสไฟฟ้า โดยมีกระแสไฟฟ้าหลายค่าที่ขึ้นอยู่กับแรงดันไฟฟ้าซึ่งทำหน้าที่ชาร์จเยื่อหุ้มเซลล์จนมีความจุC

สมการข้างต้นคืออนุพันธ์เทียบกับเวลาของกฎความจุ Q = CVโดยที่การเปลี่ยนแปลงของประจุรวมต้องอธิบายเป็นผลรวมของกระแสไฟฟ้าแต่ละกระแส โดยแต่ละกระแสมีค่าดังนี้

โดยที่g ( t , V )คือค่าการนำไฟฟ้าหรือค่าผกผันของความต้านทาน ซึ่งสามารถขยายได้ในรูปของค่าการนำไฟฟ้าสูงสุดḡ และเศษส่วนการกระตุ้นและการยับยั้งmและhตามลำดับ ซึ่งเป็นตัวกำหนดว่าไอออนจำนวนเท่าใดสามารถไหลผ่านช่องเมมเบรนที่มีอยู่ได้ การขยายนี้กำหนดโดย

และเศษส่วนของเราเป็นไปตามจลนศาสตร์อันดับหนึ่ง

โดยมีพลวัตที่คล้ายกันสำหรับhซึ่งเราสามารถใช้τและm หรือαและβเพื่อกำหนดสัดส่วนเกตของเราได้
แบบจำลองของ Hodgkin–Huxley สามารถขยายให้ครอบคลุมกระแสไอออนเพิ่มเติมได้ โดยทั่วไปแล้ว กระแสเหล่านี้จะรวมถึงกระแสขาเข้าของ Ca 2+และ Na +รวมถึงกระแสขาออกของ K + หลายประเภท ซึ่งรวมถึงกระแส "รั่ว" ด้วย
ผลลัพธ์อาจมีพารามิเตอร์น้อยที่สุดถึง 20 ตัว ซึ่งจำเป็นต้องประมาณหรือวัดเพื่อให้ได้แบบจำลองที่แม่นยำ ในแบบจำลองของระบบเซลล์ประสาท ที่ซับซ้อน การคำนวณเชิงตัวเลขของสมการนั้นมีค่าใช้จ่ายสูงดังนั้นจึงจำเป็นต้องมีการลดรูปแบบจำลองของ Hodgkin–Huxley อย่างระมัดระวัง
แบบจำลองสามารถลดเหลือสองมิติได้ด้วยความสัมพันธ์แบบไดนามิกที่สามารถสร้างขึ้นระหว่างตัวแปรเกตติ้ง[ 13 ]นอกจากนี้ยังสามารถขยายเพื่อพิจารณาถึงวิวัฒนาการของความเข้มข้น (ซึ่งถือว่าคงที่ในแบบจำลองดั้งเดิม) [ 14 ] [ 15 ]
การผสานและจุดไฟที่สมบูรณ์แบบ
หนึ่งในแบบจำลองแรกสุดของเซลล์ประสาทคือแบบจำลองอินทิเกรตแอนด์ไฟที่สมบูรณ์แบบ (เรียกอีกอย่างว่าอินทิเกรตแอนด์ไฟที่ไม่รั่ว) ซึ่งได้รับการศึกษาครั้งแรกในปี พ.ศ. 2450 โดยLouis Lapicque [ 16 ]เซลล์ประสาทแสดงด้วยแรงดันไฟฟ้าของเยื่อหุ้มเซลล์Vซึ่งเปลี่ยนแปลงไปตามเวลาในระหว่างการกระตุ้นด้วยกระแสอินพุตI(t)ตาม

ซึ่งก็คือ อนุพันธ์เทียบกับเวลาของกฎความจุ Q = CVเมื่อกระแสไฟฟ้าไหลเข้า แรงดันไฟฟ้าของเยื่อหุ้มเซลล์จะเพิ่มขึ้นตามเวลาจนกระทั่งถึงค่าเกณฑ์คงที่Vth จุดนั้น จะเกิดสไปค์ แบบฟังก์ชันเดลต้าและแรงดันไฟฟ้าจะถูกรีเซ็ตกลับไปที่ศักย์พักหลังจากนั้นแบบจำลองจะทำงานต่อไปความถี่ในการยิงของแบบจำลองจึงเพิ่มขึ้นเป็นเส้นตรงอย่างไม่มีขีดจำกัดเมื่อกระแสไฟฟ้าขาเข้าเพิ่มขึ้น
แบบจำลองนี้สามารถทำให้แม่นยำยิ่งขึ้นได้โดยการเพิ่มช่วงเวลาพักt ที่จำกัดความถี่การยิงของเซลล์ประสาทโดยป้องกันไม่ให้เซลล์ประสาทยิงในช่วงเวลานั้น สำหรับอินพุตคงที่I(t)=Iแรงดันไฟฟ้าเกณฑ์จะถึงหลังจากเวลาการรวมt =CV /Iหลังจากเริ่มต้นจากศูนย์ หลังจากรีเซ็ต ช่วงเวลาพักจะทำให้เกิดช่วงเวลาหยุดทำงาน ดังนั้นเวลาทั้งหมดจนกว่าจะเกิดการยิงครั้งต่อไปคือt + t ความถี่การยิงคือส่วนกลับของช่วงเวลาระหว่างการยิงทั้งหมด (รวมถึงช่วงเวลาหยุดทำงาน) ดังนั้นความถี่การยิงเป็นฟังก์ชันของกระแสอินพุตคงที่จึงเป็นดังนี้

ข้อเสียของแบบจำลองนี้คือ มันไม่ได้อธิบายถึงการปรับตัวหรือการรั่วไหล หากแบบจำลองได้รับพัลส์กระแสไฟฟ้าสั้นๆ ที่ต่ำกว่าเกณฑ์ในบางช่วงเวลา มันจะคงแรงดันไฟฟ้าที่เพิ่มขึ้นนั้นไว้ตลอดไป จนกว่าจะมีอินพุตอื่นมากระตุ้นให้มันทำงานในภายหลัง ลักษณะนี้ไม่สอดคล้องกับพฤติกรรมของเซลล์ประสาทที่สังเกตได้ การปรับปรุงเพิ่มเติมต่อไปนี้ทำให้แบบจำลองการรวมและการทำงาน (integrate-and-fire model) มีความน่าเชื่อถือมากขึ้นจากมุมมองทางชีววิทยา
การผสานรวมและการยิงที่รั่วไหล
แบบจำลองอินทิเกรตและไฟแบบรั่ว ซึ่งสามารถสืบย้อนไปถึงLouis Lapicque ได้ [ 16 ] มีเทอม "รั่ว" ในสมการศักย์เยื่อหุ้มเซลล์ที่สะท้อนถึงการแพร่กระจายของไอออนผ่านเยื่อหุ้มเซลล์ ซึ่งแตกต่างจากแบบจำลองอินทิเกรตและไฟแบบไม่รั่ว สมการแบบจำลองมีลักษณะดังนี้[ 1 ]


โดยที่Vmคือแรงดันไฟฟ้าคร่อมเยื่อหุ้มเซลล์ และ Rm คือ ต้านทานของเยื่อหุ้มเซลล์ (แบบจำลองอินทิเกรตแอนด์ไฟที่ไม่รั่วซึมจะถูกเรียกคืนเมื่อRm เข้าใกล้ค่าอนันต์ เกณฑ์Vthหลังจากนั้นศักย์ไฟฟ้าของเยื่อหุ้มเซลล์จะถูกรีเซ็ต
สำหรับการป้อนข้อมูลคงที่ ค่าการป้อนข้อมูลขั้นต่ำที่จะถึงเกณฑ์คือI = V / R โดยสมมติว่ามีการรีเซ็ตเป็นศูนย์ ความถี่ในการยิงจึงเป็นดังนี้
![{\displaystyle f(I)={\begin{cases}0,&I\leq I_{\mathrm {th} }\\\left[t_{\mathrm {ref} }-R_{\mathrm {m} }C_{\mathrm {m} }\log \left(1-{\tfrac {V_{\mathrm {th} }}{IR_{\mathrm {m} }}}\right)\right]^{-1},&I>I_{\mathrm {th} }\end{cases}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/3d2b9ee97278ff7f139dc76afecd9d3336a35a5b)
ซึ่งลู่เข้าสู่แบบจำลองที่ไม่มีการรั่วไหลก่อนหน้านี้ด้วยช่วงเวลาการไม่ตอบสนองสำหรับกระแสอินพุตขนาดใหญ่[ 17 ]แบบจำลองนี้ยังสามารถใช้กับเซลล์ประสาทที่ยับยั้งได้อีกด้วย[ 18 ] [ 19 ]
ข้อเสียที่สำคัญที่สุดของแบบจำลองนี้คือไม่มีการปรับตัวของเซลล์ประสาท ดังนั้นจึงไม่สามารถอธิบายชุดสไปค์ที่วัดได้จากการทดลองเพื่อตอบสนองต่อกระแสอินพุตคงที่ได้[ 20 ]ข้อเสียนี้ถูกกำจัดออกไปในแบบจำลองอินทิเกรตแอนด์ไฟร์แบบทั่วไปซึ่งมีตัวแปรการปรับตัวหนึ่งตัวหรือมากกว่า และสามารถทำนายเวลาสไปค์ของเซลล์ประสาทคอร์เทกซ์ภายใต้การฉีดกระแสได้อย่างแม่นยำสูง[ 21 ] [ 22 ] [ 23 ]
การบูรณาการและการยิงแบบปรับตัวได้
| แบบจำลองการบูรณาการและการยิงแบบปรับตัวได้ | อ้างอิง |
|---|---|
| แรงดันต่ำกว่าเกณฑ์สำหรับกระแสอินพุตที่ขึ้นอยู่กับเวลา | [ 22 ] [ 23 ] |
| เวลาการจุดระเบิดสำหรับกระแสอินพุตที่ขึ้นอยู่กับเวลา | [ 22 ] [ 23 ] |
| รูปแบบการยิงตอบสนองต่อกระแสไฟฟ้าแบบขั้นบันได | [ 24 ] [ 25 ] [ 26 ] |
การปรับตัวของเซลล์ประสาทหมายถึงข้อเท็จจริงที่ว่า แม้จะมีกระแสไฟฟ้าคงที่ฉีดเข้าไปในตัวเซลล์ประสาท ช่วงเวลาระหว่างการส่งสัญญาณออกก็จะเพิ่มขึ้น แบบจำลองเซลล์ประสาทแบบปรับตัวและส่งสัญญาณ (adaptive integrate-and-fire neuron model) ผสมผสานการรวมแบบรั่วไหลของแรงดันไฟฟ้าV เข้าด้วยกันโดยมีตัวแปรการปรับตัวหนึ่งตัวหรือมากกว่าw (ดูบทที่ 6.1 ในตำรา Neuronal Dynamics [ 27 ] )
![{\displaystyle \tau _{\mathrm {m} }{\frac {dV_{\mathrm {m} }(t)}{dt}}=RI(t)-[V_{\mathrm {m} }(t)-E_{\mathrm {m} }]-R\sum _{k}w_{k}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/9f8345759b126e0ae20788d2be0b5f74b89dd598)
![{\displaystyle \tau _{k}{\frac {dw_{k}(t)}{dt}}=-a_{k}[V_{\mathrm {m} }(t)-E_{\mathrm {m} }]-w_{k}+b_{k}\tau _{k}\sum _{f}\delta (tt^{f})}](https://wikimedia.org/api/rest_v1/media/math/render/svg/8561a663fc284164af5004d49dda606de65f3153)
ที่ไหน คือค่าคงที่เวลาของเยื่อหุ้มเซลล์, w k คือกระแสการปรับตัว โดยมีดัชนีkw คือค่าคงที่เวลาของกระแสการปรับตัว, E คือศักย์พัก และt fคือเวลาการยิงของเซลล์ประสาท และเดลต้าของกรีกหมายถึงฟังก์ชันเดลต้าของ Dirac เมื่อใดก็ตามที่แรงดันไฟฟ้าถึงเกณฑ์การยิง แรงดันไฟฟ้าจะถูกรีเซ็ตเป็นค่าV ที่ต่ำกว่าเกณฑ์การยิง ค่ารีเซ็ตเป็นหนึ่งในพารามิเตอร์ที่สำคัญของแบบจำลอง แบบจำลองการปรับตัวที่ง่ายที่สุดมีตัวแปรการปรับตัวเพียงตัวเดียวคือwและผลรวมเหนือ k จะถูกลบออก[ 28 ]



เซลล์ประสาทแบบ Integrate-and-fire ที่มีตัวแปรการปรับตัวหนึ่งตัวหรือมากกว่านั้น สามารถอธิบายรูปแบบการยิงของเซลล์ประสาทได้หลากหลายรูปแบบเพื่อตอบสนองต่อการกระตุ้นอย่างต่อเนื่อง รวมถึงการปรับตัว การระเบิด และการระเบิดเริ่มต้น[ 24 ] [ 25 ] [ 26 ]ยิ่งไปกว่านั้น เซลล์ประสาทแบบ integrate-and-fire ที่ปรับตัวได้ซึ่งมีตัวแปรการปรับตัวหลายตัว สามารถทำนายเวลาการเกิดสไปค์ของเซลล์ประสาทในเยื่อหุ้มสมองภายใต้การฉีดกระแสไฟฟ้าแบบขึ้นอยู่กับเวลาเข้าไปในโซมาได้[ 22 ] [ 23 ]
การรวมและยิงแบบรั่วไหลลำดับเศษส่วน
ความก้าวหน้าล่าสุดในแคลคูลัสเศษส่วน เชิงคำนวณและทฤษฎี นำไปสู่รูปแบบใหม่ของโมเดลที่เรียกว่า Fractional-order leaky integrate-and-fire [ 29 ] [ 30 ] ข้อดีของโมเดลนี้คือสามารถจับผลกระทบของการปรับตัวด้วยตัวแปรเดียว โมเดลมีรูปแบบดังต่อไปนี้[ 30 ]

เมื่อแรงดันไฟฟ้าถึงเกณฑ์ ระบบจะรีเซ็ต การรวมเศษส่วนถูกนำมาใช้เพื่ออธิบายการปรับตัวของเซลล์ประสาทในข้อมูลการทดลอง[ 29 ]
'Exponential integrate-and-fire' และ 'adaptive exponential integrate-and-fire'
| การรวมและยิงแบบเอ็กซ์โปเนนเชียลที่ปรับตัวได้ | อ้างอิง |
|---|---|
| ความสัมพันธ์ระหว่างกระแสและแรงดันที่ต่ำกว่าเกณฑ์ | [ 31 ] |
| รูปแบบการยิงตอบสนองต่อกระแสไฟฟ้าแบบขั้นบันได | [ 26 ] |
| การดื้อยาและการปรับตัว | [ 32 ] |
ใน แบบ จำลองการรวมและการยิงแบบ เอกซ์โปเนนเชียล [ 33 ]การสร้างสไปค์เป็นแบบเอกซ์โปเนนเชียลตามสมการ:
![{\displaystyle {\frac {dV}{dt}}-{\frac {R}{\tau _{m}}}I(t)={\frac {1}{\tau _{m}}}\left[E_{m}-V+\Delta _{T}\exp \left({\frac {V-V_{T}}{\Delta _{T}}}\right)\right].}](https://wikimedia.org/api/rest_v1/media/math/render/svg/2681d0764425e62cdbe8563763546145ac871888)
ที่ไหนคือศักย์เยื่อหุ้มเซลล์คือค่าศักย์ไฟฟ้าของเยื่อหุ้มเซลล์ที่แท้จริง คือค่าคงที่เวลาของเยื่อหุ้มเซลล์คือศักยภาพขณะพัก และคือความคมชัดของการเริ่มต้นศักยภาพการกระทำ โดยปกติจะอยู่ที่ประมาณ 1 mV สำหรับเซลล์ประสาทพีระมิดในคอร์เทกซ์[ 31 ]เมื่อศักยภาพของเยื่อหุ้มเซลล์ข้ามผ่านมันลู่เข้าสู่อนันต์ในเวลาจำกัด[ 34 ]ในการจำลองเชิงตัวเลข การรวมจะหยุดลงหากศักย์เยื่อหุ้มเซลล์ถึงเกณฑ์ที่กำหนด (ซึ่งมากกว่ามาก)) ซึ่งศักย์เยื่อหุ้มเซลล์จะถูกรีเซ็ตเป็นค่าV ค่าแรงดันไฟฟ้ารีเซ็ตV เป็นหนึ่งในพารามิเตอร์ที่สำคัญของแบบจำลอง ที่สำคัญคือ ด้านขวาของสมการข้างต้นมีลักษณะไม่เป็นเชิงเส้นที่สามารถดึงออกมาได้โดยตรงจากข้อมูลการทดลอง[ 31 ]ในแง่นี้ ความไม่เป็นเชิงเส้นแบบเอกซ์โปเนนเชียลได้รับการสนับสนุนอย่างมากจากหลักฐานการทดลอง




ในเซลล์ประสาทแบบปรับตัวแบบเอกซ์โพเนนเชียลแบบรวมและยิง[ 32 ]ความไม่เป็นเชิงเส้นแบบเอกซ์โพเนนเชียลข้างต้นของสมการแรงดันไฟฟ้าจะถูกรวมเข้ากับตัวแปรการปรับตัว w
![{\displaystyle \tau _{m}{\frac {dV}{dt}}=RI(t)+\left[E_{m}-V+\Delta _{T}\exp \left({\frac {V-V_{T}}{\Delta _{T}}}\right)\right]-Rw}](https://wikimedia.org/api/rest_v1/media/math/render/svg/dc57795f1765df722678bfca445163588b4898aa)
![{\displaystyle \tau {\frac {dw(t)}{dt}}=-a[V_{\mathrm {m} }(t)-E_{\mathrm {m} }]-w+b\tau \delta (tt^{f})}](https://wikimedia.org/api/rest_v1/media/math/render/svg/3ad1061c0bd8fa3f0894982dfd5913c5114471f9)

ที่ไหนwแสดงถึงกระแสการปรับตัวตามมาตราส่วนเวลาพารามิเตอร์โมเดลที่สำคัญ ได้แก่ ค่ารีเซ็ตแรงดัน V และค่าเกณฑ์ภายในค่าคงที่เวลาและรวมถึงพารามิเตอร์การเชื่อมต่อด้วยและขแบบจำลองการรวมและการยิงแบบปรับตัวแบบเอกซ์โปเนนเชียลสืบทอดความไม่เป็นเชิงเส้นของแรงดันไฟฟ้าที่ได้มาจากการทดลอง[ 31 ]ของแบบจำลองการรวมและการยิงแบบเอกซ์โปเนนเชียล แต่เหนือกว่าแบบจำลองนี้ มันยังสามารถอธิบายรูปแบบการยิงของเซลล์ประสาทที่หลากหลายในการตอบสนองต่อการกระตุ้นอย่างต่อเนื่อง รวมถึงการปรับตัว การระเบิด และการระเบิดเริ่มต้น[ 26 ]อย่างไรก็ตาม เนื่องจากการปรับตัวอยู่ในรูปของกระแสไฟฟ้า อาจเกิดภาวะไฮเปอร์โพลาไรเซชันที่ผิดปกติขึ้นได้ ปัญหานี้ได้รับการแก้ไขโดยการแสดงออกมาในรูปของค่าการนำไฟฟ้า[ 35 ]

แบบจำลองเซลล์ประสาทเกณฑ์ปรับได้
ในแบบจำลองนี้ ฟังก์ชันที่ขึ้นอยู่กับเวลาจะถูกเพิ่มเข้าไปในค่าเกณฑ์คงที่หลังจากเกิดสัญญาณกระตุ้นแต่ละครั้ง จะทำให้เกิดการปรับตัวของค่าเกณฑ์ ค่าศักยภาพเกณฑ์นั้นค่อยๆ กลับคืนสู่ค่าคงที่ในระยะยาว โดยขึ้นอยู่กับค่าคงที่เวลาในการปรับตัวของเกณฑ์[ 36 ]นี่เป็นหนึ่งในเทคนิคที่ง่ายกว่าในการปรับความถี่สไปค์[ 37 ]นิพจน์สำหรับเกณฑ์การปรับตัวมีดังนี้:




![{\displaystyle v_{th}(t)=v_{th0}+{\frac {\sum \theta (t-t_{f})}{f}}=v_{th0}+{\frac {\sum \theta _{0}\exp \left[-{\frac {(t-t_{f})}{\tau _{\theta }}}\right]}{f}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/3092291632a18a195d5cb17f97c121cac909b46b)
ที่ไหนถูกกำหนดโดย:
![{\displaystyle \theta (t)=\theta _{0}\exp \left[-{\frac {t}{\tau _{\theta }}}\right]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/5dcd1b31f7ffad6905bffe2135e8160cbab229a5)
เมื่อศักย์ไฟฟ้าของเยื่อหุ้มเซลล์เมื่อถึงเกณฑ์ที่กำหนด ระบบจะรีเซ็ตเป็นค่าเริ่มต้น:



เวอร์ชันที่ง่ายกว่านี้ที่มีค่าคงที่เวลาเดียวในการลดเกณฑ์ด้วยเซลล์ประสาท LIF ได้รับการนำไปใช้ใน[ 38 ]เพื่อให้ได้เครือข่ายประสาทแบบสไปค์วนซ้ำคล้าย LSTM เพื่อให้ได้ความแม่นยำใกล้เคียงกับ ANN ในงานเชิงพื้นที่และเวลาเพียงไม่กี่อย่าง
ค่าเกณฑ์ปรับได้แบบเลขชี้กำลังสองเท่า (DEXAT)
แบบจำลองเซลล์ประสาท DEXAT เป็นแบบจำลองเซลล์ประสาทแบบปรับตัวได้ชนิดหนึ่ง ซึ่งแรงดันเกณฑ์จะลดลงแบบเลขชี้กำลังสองเท่าที่มีค่าคงที่เวลาสองค่าการลดลงแบบเลขชี้กำลัง สองเท่านี้ ถูกควบคุมโดยการลดลงอย่างรวดเร็วในช่วงเริ่มต้น แล้วจึงลดลงช้าลงในช่วงเวลาที่ยาวนานขึ้น[ 39 ] [ 40 ]เซลล์ประสาทนี้ที่ใช้ใน SNN ผ่านการไล่ระดับตัวแทนจะสร้างอัตราการเรียนรู้แบบปรับตัวได้ ซึ่งให้ความแม่นยำสูงขึ้น การบรรจบกันที่เร็วขึ้น และหน่วยความจำระยะสั้นระยะยาว ที่ยืดหยุ่นกว่า เมื่อเทียบกับแบบจำลองที่มีอยู่แล้วในวรรณกรรม พลวัตของศักย์เยื่อหุ้มเซลล์อธิบายได้ด้วยสมการ และกฎการปรับตัวของเกณฑ์คือ:

พลวัตของและได้รับจาก


,

,

ที่ไหนและ.
![{\displaystyle p_{j1}=\exp \left[-{\frac {\delta t}{\tau _{b1}}}\right]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/5417632ba41e7bf5dd3f3698b17d3f4ca39c7d6e)
![{\displaystyle p_{j2}=\exp \left[-{\frac {\delta t}{\tau _{b2}}}\right]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/6a1f4efbe00768358ce5e4ba3143407cbf3594a5)
นอกจากนี้ โมเดลเซลล์ประสาทเกณฑ์ปรับตัวหลายช่วงเวลาแสดงไดนามิกที่ซับซ้อนมากขึ้นใน[ 41 ]
แบบจำลองเชิงสุ่มของแรงดันเยื่อหุ้มเซลล์และจังหวะการเกิดสไปค์
แบบจำลองในหมวดหมู่นี้เป็นแบบจำลองการรวมและการยิงแบบทั่วไปที่รวมเอาความสุ่มในระดับหนึ่งไว้ด้วย เซลล์ประสาทในคอร์เทกซ์ในการทดลองพบว่าตอบสนองต่ออินพุตที่ขึ้นอยู่กับเวลาได้อย่างน่าเชื่อถือ แม้ว่าจะมีความแปรผันเล็กน้อยระหว่างการทดลองหนึ่งกับการทดลองถัดไปหากมีการกระตุ้นซ้ำแบบเดียวกัน[ 42 ] [ 43 ]ความสุ่มในเซลล์ประสาทมีแหล่งที่มาสำคัญสองแหล่ง ประการแรก แม้ในการทดลองที่มีการควบคุมอย่างดีซึ่งกระแสอินพุตถูกฉีดเข้าไปในโซมาโดยตรง ช่องไอออนจะเปิดและปิดแบบสุ่ม[ 44 ]และสัญญาณรบกวนของช่องนี้ทำให้เกิดความแปรปรวนเล็กน้อยในค่าที่แน่นอนของศักย์เยื่อหุ้มเซลล์และเวลาที่แน่นอนของสไปค์เอาต์พุต ประการที่สอง สำหรับเซลล์ประสาทที่ฝังอยู่ในเครือข่ายคอร์เทกซ์ เป็นเรื่องยากที่จะควบคุมอินพุตที่แน่นอนเนื่องจากอินพุตส่วนใหญ่มาจากเซลล์ประสาทที่มองไม่เห็นที่อื่นในสมอง[ 27 ]
ความสุ่มได้ถูกนำมาใช้ในแบบจำลองเซลล์ประสาทแบบสไปค์ในสองรูปแบบที่แตกต่างกันโดยพื้นฐาน: (i) กระแสอินพุตที่มีเสียงรบกวน ถูกเพิ่มเข้าไปในสมการเชิงอนุพันธ์ของแบบจำลองเซลล์ประสาท[ 45 ]หรือ (ii) กระบวนการสร้างสไปค์มีเสียงรบกวน[ 46 ]ในทั้งสองกรณี ทฤษฎีทางคณิตศาสตร์สามารถพัฒนาสำหรับเวลาต่อเนื่องได้ ซึ่งหากต้องการใช้ในการจำลองด้วยคอมพิวเตอร์ ก็สามารถแปลงเป็นแบบจำลองเวลาไม่ต่อเนื่องได้
ความสัมพันธ์ของสัญญาณรบกวนในแบบจำลองเซลล์ประสาทกับความแปรปรวนของชุดสไปค์และรหัสประสาทได้รับการกล่าวถึงในการเข้ารหัสประสาทและในบทที่ 7 ของตำราเรียนพลวัตของเซลล์ประสาท[ 27 ]
แบบจำลองอินพุตที่มีสัญญาณรบกวน (สัญญาณรบกวนแบบแพร่กระจาย)
เซลล์ประสาทที่ฝังอยู่ในเครือข่ายจะรับสัญญาณสไปค์จากเซลล์ประสาทอื่นๆ เนื่องจากเวลาการมาถึงของสไปค์นั้นไม่สามารถควบคุมได้โดยนักทดลอง จึงสามารถพิจารณาได้ว่าเป็นแบบสุ่ม ดังนั้นแบบจำลองอินทิเกรตแอนด์ไฟร์ (ซึ่งอาจไม่เป็นเชิงเส้น) ที่มีฟังก์ชันไม่เชิงเส้น f(v) จะได้รับอินพุตสองอย่าง: อินพุต ควบคุมโดยนักทดลองและกระแสอินพุตที่มีสัญญาณรบกวนซึ่งอธิบายถึงข้อมูลป้อนเข้าจากพื้นหลังที่ไม่สามารถควบคุมได้



แบบจำลองของ Stein [ 45 ]เป็นกรณีพิเศษของเซลล์ประสาทแบบอินทิเกรตแอนด์ไฟที่รั่วและกระแสสัญญาณรบกวนสีขาวแบบคงที่โดยมีค่าเฉลี่ยเป็นศูนย์และความแปรปรวนเป็นหนึ่ง ในสภาวะต่ำกว่าเกณฑ์ สมมติฐานเหล่านี้จะนำไปสู่สมการของกระบวนการออร์นสไตน์-อูห์เลนเบ็ ค

![{\displaystyle \tau _{m}{\frac {dV}{dt}}=[E_{m}-V]+RI(t)+R\xi (t)}](https://wikimedia.org/api/rest_v1/media/math/render/svg/2471c95b51d6f1a179aaeacc81d4680cbc66d53f)
อย่างไรก็ตาม เมื่อเปรียบเทียบกับกระบวนการ Ornstein–Uhlenbeck มาตรฐาน แรงดันไฟฟ้าของเยื่อหุ้มเซลล์จะถูกรีเซ็ตเมื่อใดก็ตามที่ถึงเกณฑ์การยิงVth [ 45 ]การคำนวณการกระจายช่วงเวลาของแบบจำลอง Ornstein–Uhlenbeck สำหรับอินพุตคงที่ที่มีเกณฑ์นำไปสู่ปัญหาเวลาผ่านครั้งแรก[ 45 ] [ 47 ]แบบจำลองเซลล์ประสาทของ Stein และรูปแบบต่างๆ ของแบบจำลองนี้ถูกนำมาใช้เพื่อปรับการกระจายช่วงเวลาระหว่างสไปค์ของชุดสไปค์จากเซลล์ประสาทจริงภายใต้กระแสอินพุตคงที่[ 47 ]
ในเอกสารทางคณิตศาสตร์ สมการข้างต้นของกระบวนการออร์นสไตน์-อูห์เลนเบ็ค เขียนอยู่ในรูปแบบดังนี้
![{\displaystyle dV=[E_{m}-V+RI(t)]{\frac {dt}{\tau _{m}}}+\sigma \,dW}](https://wikimedia.org/api/rest_v1/media/math/render/svg/c77157af02d1ea4ea0e312efa41ed7083ff17525)
ที่ไหนคือแอมพลิจูดของสัญญาณรบกวนขาเข้า และdWคือส่วนเพิ่มของกระบวนการ Wienerสำหรับการใช้งานแบบเวลาไม่ต่อเนื่องด้วยขั้นตอนเวลา dt การอัปเดตแรงดันไฟฟ้าคือ[ 27 ]

![{\displaystyle \Delta V=[E_{m}-V+RI(t)]{\frac {\Delta t}{\tau _{m}}}+\sigma {\sqrt {\tau _{m}}}y}](https://wikimedia.org/api/rest_v1/media/math/render/svg/ac593d7a28da8c87a36c135c66923293472f26df)
โดยที่ y ถูกสุ่มมาจากการแจกแจงแบบเกาส์เซียนที่มีค่าเฉลี่ยเป็นศูนย์ และ ความแปรปรวน หนึ่ง แรงดันไฟฟ้าจะถูกรีเซ็ตเมื่อถึงเกณฑ์การกระตุ้นVth
แบบจำลองอินพุตที่มีสัญญาณรบกวนยังสามารถใช้ในแบบจำลองการรวมและการยิงแบบทั่วไปได้อีกด้วย ตัวอย่างเช่น แบบจำลองการรวมและการยิงแบบเอกซ์ponential ที่มีอินพุตที่มีสัญญาณรบกวนมีดังนี้

สำหรับการป้อนข้อมูลแบบกำหนดค่าคงที่สามารถคำนวณอัตราการยิงเฉลี่ยเป็นฟังก์ชันของ[ 48 ]สิ่งนี้สำคัญเพราะความสัมพันธ์ระหว่างความถี่และกระแส (เส้นโค้ง fI) มักถูกใช้โดยนักทดลองเพื่อกำหนดลักษณะของเซลล์ประสาท


การรวมและการยิงที่รั่วไหลพร้อมอินพุตที่มีเสียงรบกวนถูกนำมาใช้กันอย่างแพร่หลายในการวิเคราะห์เครือข่ายของเซลล์ประสาทแบบสไปค์[ 49 ]อินพุตที่มีเสียงรบกวนยังเรียกว่า 'เสียงรบกวนแบบแพร่กระจาย' เนื่องจากทำให้เกิดการแพร่กระจายของศักยภาพเยื่อหุ้มเซลล์ที่ต่ำกว่าเกณฑ์รอบวิถีที่ปราศจากเสียงรบกวน (Johannesma, [ 50 ]ทฤษฎีของเซลล์ประสาทแบบสไปค์ที่มีอินพุตที่มีเสียงรบกวนได้รับการทบทวนในบทที่ 8.2 ของตำราNeuronal Dynamics [ 27 ]
แบบจำลองเอาต์พุตที่มีสัญญาณรบกวน (สัญญาณรบกวนที่หลุดออกไป)
ในแบบจำลองอินทิเกรตแอนด์ไฟแบบกำหนดค่าได้ จะเกิดสไปค์ขึ้นหากศักย์เยื่อหุ้มเซลล์V (t)ถึงเกณฑ์ที่กำหนดในแบบจำลองเอาต์พุตที่มีเสียงรบกวน เกณฑ์ที่เข้มงวดจะถูกแทนที่ด้วยเกณฑ์ที่มีเสียงรบกวนดังต่อไปนี้ ในแต่ละช่วงเวลา t จะมีการสร้างสไปค์แบบสุ่มด้วยความเข้มแบบสุ่มทันทีหรือ'อัตราการหลุดออก' [ 27 ]


ซึ่งขึ้นอยู่กับความแตกต่างชั่วขณะระหว่างแรงดันไฟฟ้าของเยื่อหุ้มเซลล์V (t)และเกณฑ์[ 46 ] ตัว เลือกทั่วไปสำหรับ'อัตราการหลบหนี' (ซึ่งสอดคล้องกับข้อมูลทางชีววิทยา[ 22 ] ) คือ

![{\displaystyle f(V-V_{th})={\frac {1}{\tau _{0}}}\exp[\beta (V-V_{th})]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/e0e6a01b14e279fb530c4c37647a0f4dcf60b1b1)

ที่ไหนเป็นค่าคงที่เวลาที่อธิบายว่าสัญญาณไฟฟ้าเกิดขึ้นเร็วแค่ไหนเมื่อศักย์เยื่อหุ้มเซลล์ถึงระดับเกณฑ์ และเป็นพารามิเตอร์ความคมชัด สำหรับเกณฑ์จะคมชัดขึ้น และการยิงสไปค์จะเกิดขึ้นอย่างแน่นอนในขณะที่ศักย์เยื่อหุ้มเซลล์กระทบกับเกณฑ์จากด้านล่าง ค่าความคมชัดที่พบในการทดลอง[ 22 ]คือซึ่งหมายความว่าการส่งสัญญาณประสาทจะเริ่มมีนัยสำคัญมากขึ้นทันทีที่ศักย์เยื่อหุ้มเซลล์ต่ำกว่าเกณฑ์การส่งสัญญาณอย่างเป็นทางการเพียงไม่กี่มิลลิโวลต์




กระบวนการอัตราการหลบหนีผ่านเกณฑ์อ่อนได้รับการทบทวนในบทที่ 9 ของตำราNeuronal Dynamics [ 27 ]
สำหรับแบบจำลองในเวลาแบบไม่ต่อเนื่อง การเกิดสไปค์จะเกิดขึ้นด้วยความน่าจะเป็น
![{\displaystyle P_{F}(t_{n})=F[V(t_{n})-V_{th}]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/2ef050261c3ab3b90026f75a47810dc554c3fbec)
ซึ่งขึ้นอยู่กับความแตกต่างชั่วขณะระหว่างแรงดันไฟฟ้าของเยื่อหุ้มเซลล์Vในเวลานั้นและเกณฑ์[ 55 ]ฟังก์ชัน F มักถูกนำมาใช้เป็นฟังก์ชันซิกมอยด์มาตรฐาน ด้วยพารามิเตอร์ความชัน[ 46 ]คล้ายกับไดนามิกการอัปเดตในเครือข่ายประสาทเทียม แต่รูปแบบฟังก์ชันของ F ยังสามารถหาได้จากความเข้มแบบสุ่มในเวลาต่อเนื่องตามที่ได้กล่าวไว้ข้างต้นที่ไหนคือระยะทางเกณฑ์[ 46 ]

![{\displaystyle F(x)=0.5[1+\tanh(\gamma x)]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/506d2b8f3cb095fc95e2993bea559a297013a5ed)

![{\displaystyle F(y_{n})\approx 1-\exp[y_{n}\Delta t]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/cafeec2276d1633d2d283c3ba07623c0b3f206d5)

แบบจำลอง Integrate-and-fire ที่มีสัญญาณรบกวนเอาต์พุตสามารถใช้ในการทำนายฮิสโตแกรมเวลาตอบสนองต่อสิ่งเร้า (PSTH) ของเซลล์ประสาทจริงภายใต้การป้อนข้อมูลที่ขึ้นอยู่กับเวลาแบบใดก็ได้[ 22 ]สำหรับเซลล์ประสาท Integrate-and-fire ที่ไม่ปรับตัว การกระจายช่วงเวลาภายใต้การกระตุ้นคงที่สามารถคำนวณได้จากทฤษฎีการต่ออายุ แบบคง ที่[ 27 ]
แบบจำลองการตอบสนองของสไปค์ (SRM)
| แบบจำลองการตอบสนองแบบสไปค์ | อ้างอิง |
|---|---|
| แรงดันต่ำกว่าเกณฑ์สำหรับกระแสอินพุตที่ขึ้นอยู่กับเวลา | [ 23 ] [ 22 ] |
| เวลาการจุดระเบิดสำหรับกระแสอินพุตที่ขึ้นอยู่กับเวลา | [ 23 ] [ 22 ] |
| รูปแบบการยิงตอบสนองต่อกระแสไฟฟ้าแบบขั้นบันได | [ 56 ] [ 57 ] |
| การกระจายช่วงเวลาระหว่างสไปค์ | [ 56 ] [ 46 ] |
| ศักย์หลังการเกิดสไปค์ | [ 23 ] |
| ภาวะดื้อต่อการกระตุ้นและเกณฑ์การกระตุ้นแบบไดนามิก | [ 23 ] [ 22 ] |
บทความหลัก : แบบจำลองการตอบสนองแบบสไปค์
แบบจำลองการตอบสนองสไปค์ (SRM) เป็นแบบจำลองเชิงเส้นทั่วไปสำหรับแรงดันไฟฟ้าของเยื่อหุ้มเซลล์ที่ต่ำกว่าเกณฑ์ รวมกับกระบวนการเสียงรบกวนเอาต์พุตที่ไม่เป็นเชิงเส้นสำหรับการสร้างสไปค์[ 46 ] [ 58 ] [ 56 ]แรงดันไฟฟ้าของเยื่อหุ้มเซลล์V (t)ณ เวลาtคือ

โดยที่t fคือเวลาการยิงของสไปค์หมายเลข f ของเซลล์ประสาท, V คือแรงดันไฟฟ้าขณะพักในกรณีที่ไม่มีอินพุต, I(ts)คือกระแสไฟฟ้าขาเข้า ณ เวลา ts และเป็นตัวกรองเชิงเส้น (หรือเรียกว่าเคอร์เนล) ที่อธิบายถึงการมีส่วนร่วมของพัลส์กระแสอินพุต ณ เวลา ts ต่อแรงดันไฟฟ้า ณ เวลา t การมีส่วนร่วมต่อแรงดันไฟฟ้าที่เกิดจากสไปค์ ณ เวลา tอธิบายโดยแกนกลางที่ทนความร้อนโดยเฉพาะอย่างยิ่งอธิบายถึงการรีเซ็ตหลังจากสไปค์และช่วงเวลาของศักย์หลังสไปค์ ดังนั้นจึงแสดงถึงผลที่ตามมาของภาวะดื้อต่อการกระตุ้นและการปรับตัว[ 46 ] [ 23 ]แรงดันไฟฟ้า V(t) สามารถตีความได้ว่าเป็นผลลัพธ์ของการรวมสมการเชิงอนุพันธ์ของแบบจำลองอินทิเกรตและจุดไฟแบบรั่วที่เชื่อมโยงกับตัวแปรการปรับตัวที่กระตุ้นด้วยสไปค์จำนวนเท่าใดก็ได้[ 24 ]



การเกิดสไปค์ (Spike firing) เป็นแบบสุ่ม และเกิดขึ้นด้วยความเข้มข้นแบบสุ่มที่ขึ้นอยู่กับเวลา (อัตราทันที)
![{\displaystyle f(V-\vartheta (t))={\frac {1}{\tau _{0}}}\exp[\beta (V-\vartheta (t))]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/2b0258498f0e91cf25539b60de5af5a5b24dcfcb)
ด้วยพารามิเตอร์และและเกณฑ์แบบไดนามิกมอบให้โดย


ที่นี่คือเกณฑ์การกระตุ้นการทำงานของเซลล์ประสาทที่ไม่ทำงาน และอธิบายถึงการเพิ่มขึ้นของเกณฑ์หลังจากเกิดการพุ่งขึ้น ณ เวลา[ 22 ] [ 23 ]ในกรณีที่มีเกณฑ์คงที่ หนึ่งตั้งค่า. สำหรับกระบวนการเกณฑ์เป็นแบบกำหนดได้[ 27 ]



ลำดับเวลาของตัวกรองพารามิเตอร์ที่บ่งบอกลักษณะของแบบจำลองการตอบสนองสไปค์สามารถสกัดได้โดยตรงจากข้อมูลการทดลอง[ 23 ]ด้วยพารามิเตอร์ที่เหมาะสมที่สุด SRM จะอธิบายลำดับเวลาของแรงดันไฟฟ้าเมมเบรนต่ำกว่าเกณฑ์สำหรับอินพุตที่ขึ้นอยู่กับเวลาด้วยความแม่นยำ 2 mV และสามารถทำนายจังหวะเวลาของสไปค์เอาต์พุตส่วนใหญ่ด้วยความแม่นยำ 4 ms [ 22 ] [ 23 ] SRM มีความเกี่ยวข้องอย่างใกล้ชิดกับแบบจำลองแคสเคดเชิงเส้น-ไม่เชิงเส้น-ปัวซอง (เรียกอีกอย่างว่าแบบจำลองเชิงเส้นทั่วไป) [ 54 ] การประมาณค่าพารามิเตอร์ของ แบบจำลองนิวรอนเชิงความน่าจะเป็น เช่น SRM โดยใช้วิธีการที่พัฒนาขึ้นสำหรับแบบจำลองเชิงเส้นทั่วไป[ 59 ]ได้รับการกล่าวถึงในบทที่ 10 ของตำราNeuronal Dynamics [ 27 ]


ชื่อ " แบบจำลองการตอบสนองแบบสไปค์"มาจากข้อเท็จจริงที่ว่า ในเครือข่าย กระแสอินพุตสำหรับเซลล์ประสาท i ถูกสร้างขึ้นโดยสไปค์ของเซลล์ประสาทอื่น ๆ ดังนั้นในกรณีของเครือข่าย สมการแรงดันไฟฟ้าจึงกลายเป็น

ที่ไหนคือช่วงเวลาการยิงของเซลล์ประสาท j (เช่น ชุดสัญญาณสไปค์ของมัน)อธิบายถึงลำดับเวลาของการเกิดสไปค์และศักย์หลังสไปค์สำหรับเซลล์ประสาท i; และ และอธิบายแอมพลิจูดและช่วงเวลาของการเปลี่ยนแปลงศักย์ไฟฟ้าหลังไซแนปส์ (PSP) แบบกระตุ้นหรือยับยั้งที่เกิดจากสไปค์ของเซลล์ประสาทก่อนซิแนปส์ j. ลำดับเวลาผลลัพธ์ของ PSP มาจากการรวมกันของกระแสโพสต์ไซแนปส์เกิดจากการมาถึงของสัญญาณกระตุ้นก่อนซินแนปส์จากเซลล์ประสาท j พร้อมกับตัวกรองเยื่อหุ้มเซลล์[ 27 ]





เอสอาร์เอ็มโอ
SRM 0 [ 56 ] [ 60 ] [ 61 ] เป็นแบบจำลองเซลล์ประสาทแบบสุ่มที่เกี่ยวข้องกับทฤษฎีการต่ออายุแบบ ไม่เชิงเส้นที่ขึ้นอยู่กับเวลาและเป็นการทำให้แบบจำลองการตอบสนองของสไปค์ (SRM) ง่ายขึ้น ความแตกต่างหลักจากสมการแรงดันไฟฟ้าของ SRM ที่กล่าวมาข้างต้นคือในเทอมที่มีเคอร์เนลการหักเหไม่มีเครื่องหมายผลรวมเหนือยอดแหลมที่ผ่านมา: มีเพียงยอดแหลมล่าสุด เท่านั้น (ระบุเป็นเวลา)) มีความสำคัญ ความแตกต่างอีกประการหนึ่งคือค่าเกณฑ์คงที่ โมเดล SRM0 สามารถกำหนดได้ในเวลาแบบไม่ต่อเนื่องหรือแบบต่อเนื่อง ตัวอย่างเช่น ในเวลาแบบต่อเนื่อง สมการของเซลล์ประสาทเดี่ยวคือ



และสมการเครือข่ายของ SRM 0 คือ[ 56 ]

ที่ไหนคือเวลาการยิงครั้งสุดท้ายของเซลล์ประสาท i โปรดสังเกตว่าช่วงเวลาของศักยภาพหลังไซแนปส์นอกจากนี้ยังอนุญาตให้ขึ้นอยู่กับเวลาตั้งแต่สไปค์สุดท้ายของนิวรอน i เพื่ออธิบายการเปลี่ยนแปลงการนำไฟฟ้าของเยื่อหุ้มเซลล์ในช่วงระยะเวลาที่ไม่ตอบสนอง[ 60 ]อัตราการยิงทันที (ความเข้มแบบสุ่ม) คือ


![{\displaystyle f(V-\vartheta )={\frac {1}{\tau _{0}}}\exp[\beta (V-V_{th})]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/9bd95a6edcc2e90476af83affbc51d150042d33b)
ที่ไหนเป็นเกณฑ์การกระตุ้นคงที่ ดังนั้นการยิงสไปค์ของเซลล์ประสาท i จึงขึ้นอยู่กับอินพุตและเวลาตั้งแต่เซลล์ประสาท i ยิงสไปค์ครั้งล่าสุดเท่านั้น
ด้วย SRM 0การกระจายช่วงเวลาระหว่างสไปค์สำหรับอินพุตคงที่สามารถเชื่อมโยงทางคณิตศาสตร์กับรูปร่างของเคอร์เนลที่ไม่ตอบสนองได้[ 46 ] [ 56 ] ยิ่งไปกว่านั้น ความสัมพันธ์ระหว่างความถี่และกระแสคง ที่ สามารถคำนวณได้จากอัตราการหลุดออกเมื่อรวมกับแกนกลางที่หักเหแสง[ 46 ] [ 56 ]ด้วยการเลือกเคอร์เนลที่เหมาะสม SRM 0จะประมาณไดนามิกของแบบจำลอง Hodgkin-Huxley ได้อย่างแม่นยำสูง[ 60 ] ยิ่งไปกว่านั้น การตอบสนองของ PSTH ต่ออินพุตที่ขึ้นอยู่กับเวลาใดๆ ก็สามารถทำนายได้[ 56 ]

แบบจำลองของเล่นเพื่อการเรียนรู้เกี่ยวกับแรงดันไฟฟ้าของเยื่อหุ้มเซลล์
แบบจำลองในหมวดหมู่นี้เป็นแบบจำลองของเล่นที่เรียบง่ายมาก ซึ่งอธิบายแรงดันไฟฟ้าของเยื่อหุ้มเซลล์ในเชิงคุณภาพโดยขึ้นอยู่กับอินพุต ส่วนใหญ่ใช้เพื่อวัตถุประสงค์ทางการสอน แต่ไม่ถือว่าเป็นแบบจำลองเซลล์ประสาทที่ถูกต้องสำหรับการจำลองขนาดใหญ่หรือการปรับข้อมูลให้เข้ากับแบบจำลอง
ฟิตซ์ฮิวจ์–นากูโมะ
การลดความซับซ้อนครั้งใหญ่ของ Hodgkin–Huxley ได้รับการนำเสนอโดย FitzHugh และ Nagumo ในปี 1961 และ 1962 โดยพยายามอธิบาย "การกระตุ้นตัวเองแบบสร้างใหม่" ด้วยแรงดันเมมเบรนป้อนกลับเชิงบวกแบบไม่เชิงเส้นและการฟื้นตัวด้วยแรงดันเกตป้อนกลับเชิงลบแบบเชิงเส้น พวกเขาได้พัฒนารูปแบบที่อธิบายโดย[ 62 ]

โดยที่เรามีแรงดันไฟฟ้าและกระแสอินพุตที่คล้ายเยื่อหุ้มเซลล์อีกครั้ง พร้อมด้วยแรงดันไฟฟ้าเกตทั่วไปที่ช้าลงwและพารามิเตอร์ที่กำหนดจากการทดลองa = -0.7, b = 0.8, τ = 1/0.08แม้ว่าจะไม่สามารถอนุมานได้จากชีววิทยา แต่แบบจำลองนี้ช่วยให้เกิดพลวัตที่ง่ายขึ้นและพร้อมใช้งานได้ทันที โดยไม่ทำให้ง่ายขึ้นอย่างผิวเผิน[ 63 ]การสนับสนุนจากการทดลองนั้นอ่อนแอ แต่แบบจำลองนี้มีประโยชน์ในฐานะเครื่องมือการสอนเพื่อแนะนำพลวัตของการสร้างสไปค์ผ่าน การวิเคราะห์ ระนาบเฟสดูบทที่ 7 ในตำรา วิธีการ สร้างแบบจำลองเซลล์ประสาท[ 64 ]
มอร์ริส-เลการ์
ในปี 1981 มอร์ริสและเลการ์ได้รวมแบบจำลองของฮอดจ์กิน-ฮักซ์ลีย์และฟิตซ์ฮิวจ์-นากูโมเข้าด้วยกันเป็นแบบจำลองช่องแคลเซียมที่ควบคุมด้วยแรงดันไฟฟ้า โดยมีช่องโพแทสเซียม แบบปรับแก้ล่าช้า แทนด้วย

ที่ไหน[ 17 ]การสนับสนุนเชิงทดลองของแบบจำลองนั้นอ่อนแอ แต่แบบจำลองนี้มีประโยชน์ในฐานะเครื่องมือการสอนเพื่อแนะนำพลวัตของการสร้างสไปค์ผ่านการ วิเคราะห์ ระนาบเฟสดูบทที่ 7 [ 65 ]ในตำรา วิธีการสร้างแบบ จำลองประสาท[ 64 ]

แบบจำลองเซลล์ประสาทสองมิติที่คล้ายกับแบบจำลอง Morris-Lecar มาก สามารถสร้างขึ้นทีละขั้นตอนโดยเริ่มจากแบบจำลอง Hodgkin-Huxley ดูบทที่ 4.2 ในตำรา Neuronal Dynamics [ 27 ]
ฮินด์มาร์ช-โรส
โดยอาศัยแบบจำลอง FitzHugh–Nagumo, Hindmarsh และ Rose ได้เสนอแบบจำลองกิจกรรมของเซลล์ประสาทในปี 1984 [ 66 ]ซึ่งอธิบายโดยสมการเชิงอนุพันธ์อันดับหนึ่งที่เชื่อมโยงกันสามสมการ:

โดยที่r 2 = x 2 + y 2 + z 2และr ≈ 10 −2ดังนั้น ตัวแปร zจึงเปลี่ยนแปลงช้ามาก ความซับซ้อนทางคณิตศาสตร์เพิ่มเติมนี้ช่วยให้เกิดพฤติกรรมไดนามิกที่หลากหลายสำหรับศักย์เยื่อหุ้มเซลล์ ซึ่งอธิบายโดย ตัวแปร xของแบบจำลอง ซึ่งรวมถึงไดนามิกแบบอลวน สิ่งนี้ทำให้แบบจำลองเซลล์ประสาท Hindmarsh–Rose มีประโยชน์มาก เนื่องจากยังคงเรียบง่าย ช่วยให้สามารถอธิบายรูปแบบการยิงที่แตกต่างกันมากมายของศักย์การกระทำ โดยเฉพาะอย่างยิ่งการระเบิด ที่สังเกตได้ในการทดลอง อย่างไรก็ตาม มันยังคงเป็นแบบจำลองของเล่นและยังไม่ได้ปรับให้เข้ากับข้อมูลการทดลอง มันถูกใช้กันอย่างแพร่หลายเป็นแบบจำลองอ้างอิงสำหรับไดนามิกการระเบิด[ 66 ]
แบบจำลองเธต้าและการรวมและยิงแบบกำลังสอง
แบบจำลองเธต้าหรือ แบบจำลอง แคนอนิกประเภท I ของ Ermentrout–Kopell นั้นเทียบเท่าทางคณิตศาสตร์กับแบบจำลองการรวมและยิงแบบกำลังสอง ซึ่งเป็นการประมาณค่าของแบบจำลองการรวมและยิงแบบเอกซ์โพเนนเชียลและแบบจำลอง Hodgkin-Huxley เรียกว่าแบบจำลองแคนอนิกเพราะเป็นหนึ่งในแบบจำลองทั่วไปสำหรับอินพุตคงที่ใกล้จุดแยกสาขา ซึ่งหมายความว่าใกล้การเปลี่ยนจากการยิงแบบเงียบไปเป็นการยิงซ้ำ[ 67 ] [ 68 ]
สูตรมาตรฐานของแบบจำลองเธต้าคือ[ 27 ] [ 67 ] [ 68 ]
![{\displaystyle {\frac {d\theta (t)}{dt}}=(I-I_{0})[1+\cos(\theta )]+[1-\cos(\theta )]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/ecbba69c0c299fc9ba2a49a79be5d607200af4c3)
สมการสำหรับแบบจำลองการรวมและการยิงกำลังสองคือ (ดูบทที่ 5.3 ในตำรา Neuronal Dynamics [ 27 ] )
![{\displaystyle \tau _{\mathrm {m} }{\frac {dV_{\mathrm {m} }(t)}{dt}}=(I-I_{0})R+[V_{\mathrm {m} }(t)-E_{\mathrm {m} }][V_{\mathrm {m} }(t)-V_{\mathrm {T} }]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/e8392a8de9357950491d89517ec59294d6ef35e5)
ตัวอย่างเช่น ความเท่าเทียมกันของแบบจำลองเธต้าและการรวมและการยิงกำลังสองได้รับการทบทวนในบทที่ 4.1.2.2 ของแบบจำลองเซลล์ประสาทสไปค์[ 1 ]
สำหรับการป้อนข้อมูลหากมีการเปลี่ยนแปลงตามเวลาหรืออยู่ห่างจากจุดแยกสาขา ควรใช้โมเดลการรวมและการยิงแบบเอกซ์โปเนนเชียล (หากต้องการอยู่ในกลุ่มโมเดลเซลล์ประสาทแบบหนึ่งมิติ) เนื่องจากเซลล์ประสาทจริงแสดงคุณสมบัติไม่เป็นเชิงเส้นของโมเดลการรวมและการยิงแบบเอกซ์โปเนนเชียล[ 31 ]
แบบจำลองเซลล์ประสาทเข้ารหัสสิ่งเร้าจากข้อมูลป้อนเข้าทางประสาทสัมผัส
แบบจำลองในหมวดหมู่นี้ได้มาจากการทดลองที่เกี่ยวข้องกับการกระตุ้นตามธรรมชาติ เช่น แสง เสียง การสัมผัส หรือกลิ่น ในการทดลองเหล่านี้ รูปแบบการส่งสัญญาณประสาทที่เกิดขึ้นจากการกระตุ้นแต่ละครั้งจะแตกต่างกันไปในแต่ละครั้ง แต่การตอบสนองโดยเฉลี่ยจากหลายๆ ครั้งมักจะบรรจบกันเป็นรูปแบบที่ชัดเจน ดังนั้น แบบจำลองในหมวดหมู่นี้จึงสร้างความสัมพันธ์เชิงความน่าจะเป็นระหว่างการกระตุ้นที่ป้อนเข้าไปกับการเกิดการส่งสัญญาณประสาท ที่สำคัญคือ เซลล์ประสาทที่บันทึกไว้มักจะอยู่หลังจากเซลล์ประสาทรับความรู้สึกหลายขั้นตอนการประมวลผล ดังนั้นแบบจำลองเหล่านี้จึงสรุปผลกระทบของลำดับขั้นตอนการประมวลผลในรูปแบบที่กระชับ
แบบจำลองกระบวนการปัวซงที่ไม่เป็นเอกพันธ์ (ซีเบิร์ต)
Siebert [ 69 ] [ 70 ]ได้สร้างแบบจำลองรูปแบบการยิงสไปค์ของเซลล์ประสาทโดยใช้ แบบจำลอง กระบวนการปัวซง ที่ไม่ เป็นเนื้อเดียวกัน โดยอ้างอิงจากการทดลองที่เกี่ยวข้องกับระบบการได้ยิน[ 69 ] [ 70 ]ตามที่ Siebert กล่าว ความน่าจะเป็นของเหตุการณ์สไปค์ในช่วงเวลาเป็นสัดส่วนกับฟังก์ชันที่ไม่เป็นลบ, ที่ไหนคือสิ่งเร้าดิบ:
![{\displaystyle [t,t+\Delta _{t}]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/7a5f74c8e9f1b771d10e66322f05d9692676551b)
![{\displaystyle g[s(t)]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/79f02d6c931d32be9a417598bb7164a38b18e7c7)

![{\displaystyle P_{\text{spike}}(t\in [t',t'+\Delta _{t}])=\Delta _{t}\cdot g[s(t)]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/6c518881857d805263d6674a39ae79d3b830a06c)
ซีเบิร์ตพิจารณาหน้าที่หลายประการดังนี้, รวมทั้งสำหรับระดับความเข้มของสิ่งเร้าต่ำ
![{\displaystyle g[s(t)]\propto s^{2}(t)}](https://wikimedia.org/api/rest_v1/media/math/render/svg/279ad33854b65b37667308913d562014bdb7f30a)
ข้อดีหลักของแบบจำลองของซีเบิร์ตคือความเรียบง่าย ส่วนข้อเสียของแบบจำลองนี้คือไม่สามารถสะท้อนปรากฏการณ์ต่อไปนี้ได้อย่างถูกต้อง:
- การเพิ่มขึ้นชั่วคราวของกิจกรรมการส่งสัญญาณประสาทเพื่อตอบสนองต่อสิ่งเร้าแบบขั้นบันได
- ภาวะอิ่มตัวของอัตราการยิง
- ค่าของ ฮิสโตแกรมช่วงเวลาระหว่างการเกิดสไปค์ในช่วงเวลาสั้นๆ (ใกล้ศูนย์)
ข้อบกพร่องเหล่านี้ได้รับการแก้ไขโดยแบบจำลองกระบวนการจุดที่ขึ้นอยู่กับอายุและแบบจำลองมาร์คอฟสองสถานะ[ 71 ] [ 72 ] [ 73 ]
แบบจำลองกระบวนการจุดที่ไม่ตอบสนองต่อสิ่งเร้าและอายุ
Berry และ Meister [ 74 ]ศึกษาภาวะดื้อต่อการกระตุ้นของเซลล์ประสาทโดยใช้แบบจำลองสุ่มที่ทำนายการเกิดสไปค์เป็นผลคูณของสองเทอม คือ ฟังก์ชัน f(s(t)) ที่ขึ้นอยู่กับสิ่งเร้า s(t) ที่ขึ้นอยู่กับเวลา และฟังก์ชันการฟื้นตัว ขึ้นอยู่กับระยะเวลาตั้งแต่การพุ่งขึ้นครั้งล่าสุด


แบบจำลองนี้ยังเรียกว่ากระบวนการช่วงเวลา Markov ที่ไม่เหมือนกัน (IMI) [ 75 ] แบบ จำลองที่คล้ายกันนี้ถูกใช้มาหลายปีแล้วในประสาทวิทยาศาสตร์การได้ยิน[ 76 ] [ 77 ] [ 78 ]เนื่องจากแบบจำลองนี้เก็บความทรงจำของเวลาสไปค์สุดท้ายไว้ จึงไม่ใช่แบบปัวซงและจัดอยู่ในกลุ่มของแบบจำลองการต่ออายุที่ขึ้นอยู่กับเวลา[ 27 ]มันมีความเกี่ยวข้องอย่างใกล้ชิดกับแบบจำลอง SRM0ที่มีอัตราการหลบหนีแบบเอกซ์โพเนนเชียล[ 27 ]ที่สำคัญคือ สามารถปรับพารามิเตอร์ของแบบจำลองกระบวนการจุดที่ขึ้นอยู่กับอายุได้ เพื่ออธิบายไม่เพียงแค่การตอบสนอง PSTH เท่านั้น แต่ยังรวมถึงสถิติช่วงเวลาระหว่างสไปค์ด้วย[ 75 ] [ 76 ] [ 78 ]
แบบจำลอง Poisson แบบเรียงลำดับเชิงเส้น-ไม่เชิงเส้น และ GLM
แบบจำลองแคสเคดเชิงเส้น-ไม่เชิงเส้น-ปัวซงเป็นแคสเคดของกระบวนการกรองเชิงเส้นตามด้วยขั้นตอนการสร้างสไปค์ที่ไม่เชิงเส้น[ 79 ]ในกรณีที่สไปค์เอาต์พุตป้อนกลับผ่านกระบวนการกรองเชิงเส้น เราจะได้แบบจำลองที่รู้จักกันในสาขาวิทยาศาสตร์ประสาทว่าแบบจำลองเชิงเส้นทั่วไป (GLM) [ 54 ] [ 59 ] GLM เทียบเท่าทางคณิตศาสตร์กับแบบจำลองการตอบสนองสไปค์ (SRM) ที่มีสัญญาณรบกวนแบบหลบหนี แต่ในขณะที่ใน SRM ตัวแปรภายในถูกตีความว่าเป็นศักย์เยื่อหุ้มเซลล์และเกณฑ์การยิง ใน GLM ตัวแปรภายในเป็นปริมาณนามธรรมที่สรุปผลสุทธิของอินพุต (และสไปค์เอาต์พุตล่าสุด) ก่อนที่จะสร้างสไปค์ในขั้นตอนสุดท้าย[ 27 ] [ 54 ]
แบบจำลองมาร์คอฟสองสถานะ (นอสเซนสันและเมสเซอร์)
แบบจำลองเซลล์ประสาทแบบสไปค์โดย Nossenson & Messer [ 71 ] [ 72 ] [ 73 ]สร้างความน่าจะเป็นที่เซลล์ประสาทจะยิงสไปค์เป็นฟังก์ชันของสิ่งเร้าภายนอกหรือสิ่งเร้าทางเภสัชวิทยา[ 71 ] [ 72 ] [ 73 ]แบบจำลองนี้ประกอบด้วยแบบจำลองชั้นตัวรับและแบบจำลองเซลล์ประสาทแบบสไปค์ที่เรียงลำดับกัน ดังแสดงในรูปที่ 4 การเชื่อมต่อระหว่างสิ่งเร้าภายนอกกับความน่าจะเป็นของการยิงสไปค์เกิดขึ้นในสองขั้นตอน: ขั้นแรก แบบจำลองเซลล์ตัวรับจะแปลงสิ่งเร้าภายนอกดิบเป็นความเข้มข้นของสารสื่อประสาท จากนั้น แบบจำลองเซลล์ประสาทแบบสไปค์จะเชื่อมต่อความเข้มข้นของสารสื่อประสาทกับอัตราการยิง (ความน่าจะเป็นของการยิงสไปค์) ดังนั้น แบบจำลองเซลล์ประสาทแบบสไปค์จึงขึ้นอยู่กับความเข้มข้นของสารสื่อประสาทในขั้นตอนการป้อนข้อมูล[ 71 ] [ 72 ] [ 73 ]


คุณลักษณะสำคัญของแบบจำลองนี้คือการทำนายรูปแบบอัตราการยิงของเซลล์ประสาท ซึ่งสามารถจับภาพการตอบสนองที่เน้นขอบลักษณะเฉพาะของเซลล์ประสาทต่อพัลส์กระตุ้นได้โดยใช้พารามิเตอร์อิสระจำนวนน้อย ดังแสดงในรูปที่ 5 อัตราการยิงถูกระบุทั้งในรูปของความน่าจะเป็นแบบนอร์มาไลซ์สำหรับการยิงสไปค์ของเซลล์ประสาท และในรูปของปริมาณที่แปรผันตามกระแสของสารสื่อประสาทที่ปล่อยออกมาจากเซลล์ สูตรสำหรับอัตราการยิงมีรูปแบบดังต่อไปนี้:
![{\displaystyle R_{\text{fire}}(t)={\frac {P_{\text{spike}}(t;\Delta _{t})}{\Delta _{t}}}=[y(t)+R_{0}]\cdot P_{0}(t)}](https://wikimedia.org/api/rest_v1/media/math/render/svg/1038693094674ba67b2e424be6eae51db081721e)
ที่ไหน,
- P0 คือความน่าจะเป็นที่เซลล์ประสาทจะ "พร้อมใช้งาน" และพร้อมที่จะส่งสัญญาณ โดยกำหนดโดยสมการเชิงอนุพันธ์ต่อไปนี้:
![{\displaystyle {\dot {P}}_{0}=-[y(t)+R_{0}+R_{1}]\cdot P_{0}(t)+R_{1}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/830fe4d93362af37a7b4d97c1902b7db1826f9e1)
โดยทั่วไป P0 สามารถคำนวณแบบวนซ้ำได้โดยใช้วิธีออยเลอร์ แต่ในกรณีของพัลส์ของสิ่งเร้า จะได้นิพจน์แบบปิดที่เรียบง่าย[ 71 ] [ 80 ]
- y ( t ) คืออินพุตของแบบจำลอง และถูกตีความว่าเป็นความเข้มข้นของสารสื่อประสาทในเซลล์โดยรอบ (ในกรณีส่วนใหญ่คือกลูตาเมต) สำหรับสิ่งเร้าภายนอก สามารถประมาณค่าได้ผ่านแบบจำลองชั้นตัวรับ:

กับโดยเป็นค่าเฉลี่ยของกำลังกระตุ้นในช่วงเวลาสั้นๆ (ระบุเป็นวัตต์หรือพลังงานอื่นๆ ต่อหน่วยเวลา)

- R สอดคล้องกับอัตราการยิงกระแสประสาทโดยธรรมชาติของเซลล์ประสาท
- R คืออัตราการฟื้นตัวของเซลล์ประสาทจากสภาวะที่ไม่ตอบสนอง
การคาดการณ์อื่นๆ จากแบบจำลองนี้ ได้แก่:
1) ศักยภาพการตอบสนองที่ถูกกระตุ้นโดยเฉลี่ย (ERP) เนื่องมาจากประชากรของเซลล์ประสาทจำนวนมากในการวัดที่ไม่ได้กรองนั้นคล้ายกับอัตราการยิง[ 73 ]
2) ความแปรปรวนของแรงดันไฟฟ้าของกิจกรรมเนื่องจากกิจกรรมของเซลล์ประสาทหลายเซลล์มีลักษณะคล้ายกับอัตราการยิง (เรียกอีกอย่างว่าพลังงานกิจกรรมหลายหน่วยหรือ MUA) [ 72 ] [ 73 ]
3) การกระจายความน่าจะเป็นระหว่างช่วงเวลาสไปค์มีรูปแบบเป็นฟังก์ชันคล้ายการกระจายแกมมา[ 71 ] [ 80 ]
| ทรัพย์สินของแบบจำลองโดย Nossenson & Messer | อ้างอิง | คำอธิบายหลักฐานเชิงทดลอง |
|---|---|---|
| รูปทรงของอัตราการยิงตอบสนองต่อพัลส์กระตุ้นทางเสียง | [ 81 ] [ 82 ] [ 83 ] [ 84 ] [ 85 ] | อัตราการยิงมีรูปร่างเหมือนกับรูปที่ 5 |
| รูปทรงของอัตราการยิงตอบสนองต่อพัลส์กระตุ้นทางสายตา | [ 86 ] [ 87 ] [ 88 ] [ 89 ] | อัตราการยิงมีรูปร่างเหมือนกับรูปที่ 5 |
| รูปทรงของอัตราการยิงตอบสนองต่อพัลส์กระตุ้นทางกลิ่น | [ 90 ] | อัตราการยิงมีรูปร่างเหมือนกับรูปที่ 5 |
| รูปทรงของอัตราการยิงสัญญาณตอบสนองต่อสิ่งเร้าทางประสาทสัมผัส | [ 91 ] | อัตราการยิงมีรูปร่างเหมือนกับรูปที่ 5 |
| การเปลี่ยนแปลงอัตราการยิงของเซลล์ประสาทตอบสนองต่อการให้สารสื่อประสาท (ส่วนใหญ่คือกลูตาเมต) | [ 92 ] [ 93 ] | การเปลี่ยนแปลงอัตราการยิงของเซลล์ประสาทตอบสนองต่อการให้สารสื่อประสาท (กลูตาเมต) |
| ความสัมพันธ์กำลังสองระหว่างแรงดันกระตุ้นทางเสียงและอัตราการยิงของเซลล์ประสาท | [ 94 ] | ความสัมพันธ์กำลังสองระหว่างแรงดันของสิ่งเร้าทางเสียงและอัตราการยิง (- ความสัมพันธ์เชิงเส้นในกำลังสองของแรงดัน (กำลัง)) |
| ความสัมพันธ์กำลังสองระหว่างสนามไฟฟ้าของสิ่งเร้าทางสายตา (โวลต์) และอัตราการยิงของเซลล์ประสาท | [ 87 ] | ความสัมพันธ์แบบกำลังสองระหว่างสนามไฟฟ้าของสิ่งเร้าทางสายตา (โวลต์) กับความสัมพันธ์เชิงเส้นระหว่างกำลังของ สิ่งเร้าทางสายตา และอัตราการยิงของเซลล์ประสาท |
| รูปทรงของสถิติช่วงเวลาระหว่างการเกิดสไปค์ (Inter-Spike-Interval Statistics หรือ ISI) | [ 95 ] | รูปร่างของ ISI คล้ายกับฟังก์ชันแกมมา |
| ERP มีลักษณะคล้ายกับอัตราการยิงในข้อมูลการวัดที่ไม่ผ่านการกรอง | [ 96 ] | รูปร่างของศักยภาพการตอบสนองที่เกิดขึ้นโดยเฉลี่ยเมื่อตอบสนองต่อสิ่งเร้า มีลักษณะคล้ายกับอัตราการยิง (รูปที่ 5) |
| พลังงาน MUA มีลักษณะคล้ายกับอัตราการยิง | [ 73 ] [ 97 ] | รูปทรงของความแปรปรวนเชิงประจักษ์ของการวัดนอกเซลล์ที่ตอบสนองต่อพัลส์กระตุ้นนั้นคล้ายคลึงกับอัตราการยิง (รูปที่ 5) |
แบบจำลองเซลล์ประสาทที่ได้รับสิ่งกระตุ้นทางเภสัชวิทยา
แบบจำลองในหมวดหมู่นี้สร้างการคาดการณ์สำหรับการทดลองที่เกี่ยวข้องกับการกระตุ้นทางเภสัชวิทยา
การส่งสัญญาณประสาท (โคชและเซเกฟ)
ตามแบบจำลองของ Koch และ Segev [ 17 ]การตอบสนองของเซลล์ประสาทต่อสารสื่อประสาทแต่ละชนิดสามารถจำลองได้เป็นส่วนขยายของแบบจำลอง Hodgkin–Huxley แบบคลาสสิกที่มีทั้งกระแสจลนศาสตร์มาตรฐานและไม่มาตรฐาน สารสื่อประสาทสี่ชนิดมีอิทธิพลต่อระบบประสาทส่วนกลางเป็นหลัก ตัวรับ AMPA/kainateเป็น ตัวกลาง กระตุ้น อย่างรวดเร็ว ในขณะที่ตัวรับ NMDAเป็นตัวกลางของกระแสที่ช้ากว่ามาก กระแส ยับยั้ง อย่างรวดเร็ว ไหลผ่านตัวรับGABA ในขณะที่ตัวรับGABA เป็นตัวกลางผ่านช่องโพแทสเซียมที่เปิดใช้งานโดยโปรตีน Gรองช่วงของการเป็นตัวกลางนี้ทำให้เกิดพลวัตของกระแสดังต่อไปนี้:
![{\displaystyle I_{\mathrm {AMPA} }(t,V)={\bar {g}__{\mathrm {AMPA} }\cdot [O]\cdot (V(t)-E_{\mathrm {AMPA} })}](https://wikimedia.org/api/rest_v1/media/math/render/svg/a00bcdac49e857cbf0e2440b47d7760a17d7bdc5)
![{\displaystyle I_{\mathrm {NMDA} }(t,V)={\bar {g}}_{\mathrm {NMDA} }\cdot B(V)\cdot [O]\cdot (V(t)-E_{\mathrm {NMDA} })}](https://wikimedia.org/api/rest_v1/media/math/render/svg/4d0bedfd5fbcaada1bfb385795d52950e6429e10)
![{\displaystyle I_{\mathrm {GABA_{A}} }(t,V)={\bar {g}__{\mathrm {GABA_{A}} }\cdot ([O_{1}]+[O_{2}])\cdot (V(t)-E_{\mathrm {Cl} })}](https://wikimedia.org/api/rest_v1/media/math/render/svg/a0a4898469d8c0f8a2a1b4fd1620c3f69795f85b)
![{\displaystyle I_{\mathrm {GABA_{B}} }(t,V)={\bar {g}__{\mathrm {GABA_{B}} }\cdot {\tfrac {[G]^{n}}{[G]^{n}+K_{\mathrm {d} }}}\cdot (V(t)-E_{\mathrm {K} })}](https://wikimedia.org/api/rest_v1/media/math/render/svg/09fdce1c61e28bac586e48a928523772a0f4187f)
โดยที่ḡ คือค่าการนำไฟฟ้าสูงสุด[ 8 ] [ 17 ] (ประมาณ 1 S ) และEคือศักยภาพสมดุลของไอออนหรือตัวส่งสัญญาณที่กำหนด (AMDA, NMDA, ClหรือK ) ในขณะที่[ O ]อธิบายถึงสัดส่วนของตัวรับที่เปิดอยู่ สำหรับ NMDA จะมีผลกระทบอย่างมีนัยสำคัญของการปิดกั้นแมกนีเซียมซึ่งขึ้นอยู่กับความเข้มข้นของแมกนีเซียมภายในเซลล์แบบซิกมอยด์โดยB ( V ) สำหรับ GABA B [ G ] คือความเข้มข้นโปรตีนGและ K dอธิบายถึงแยกตัวของGในการจับกับประตูโพแทสเซียม
พลวัตของแบบจำลองที่ซับซ้อนกว่านี้ได้รับการศึกษาอย่างละเอียดถี่ถ้วนในเชิงทดลอง และให้ผลลัพธ์ที่สำคัญในแง่ของการเสริมสร้างและลดทอนความแข็งแรงของไซแนปส์ อย่างรวดเร็ว ซึ่งก็คือการเรียนรู้ระยะสั้นที่รวดเร็ว
แบบ จำลองสุ่มของ Nossenson และ Messerแปลงความเข้มข้นของสารสื่อประสาทที่ขั้นตอนอินพุตเป็นความน่าจะเป็นของการปล่อยสารสื่อประสาทที่ขั้นตอนเอาต์พุต[ 71 ] [ 72 ] [ 73 ]สำหรับคำอธิบายโดยละเอียดเพิ่มเติมของแบบจำลองนี้ โปรดดูส่วนแบบจำลองมาร์คอฟสองสถานะด้านบน
แบบจำลองเซลล์ประสาท HTM
แบบจำลองเซลล์ประสาท HTM พัฒนาขึ้นโดยเจฟฟ์ ฮอว์กินส์และนักวิจัยจากนูเมนตา โดยอิงจากทฤษฎีที่เรียกว่าหน่วยความจำเชิงลำดับชั้น (Hierarchical Temporal Memory ) ซึ่งอธิบายไว้ในหนังสือOn Intelligence เป็นครั้งแรก แบบจำลองนี้อิงจากประสาทวิทยาศาสตร์สรีรวิทยา และปฏิสัมพันธ์ของเซลล์ประสาทพีระมิดในเปลือกสมองส่วนหน้าของมนุษย์
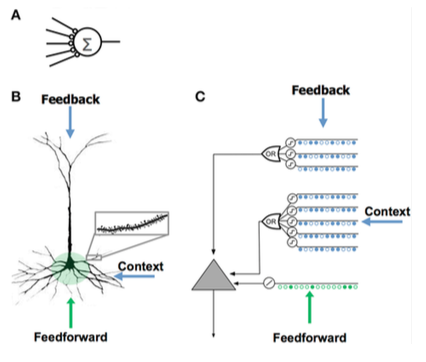
โครงข่ายประสาทเทียม (ANN) | เซลล์ประสาทพีระมิดในเปลือกสมองส่วนหน้า (เซลล์ประสาททางชีววิทยา) | เซลล์ประสาทโมเดล HTM |
| - มีไซแนปส์น้อย - ไม่มีเดนไดรต์ - ผลรวมของอินพุต x น้ำหนัก - เรียนรู้โดยการปรับเปลี่ยนน้ำหนักของไซแนปส์ | - มีจุดเชื่อมต่อประสาท (synapses) นับพันจุดบนเดนไดรต์ - เดนไดรต์ที่ทำงานอยู่: เซลล์สามารถจดจำรูปแบบเฉพาะได้หลายร้อยแบบ - การกระตุ้นพร้อมกันของกลุ่มไซแนปส์บนส่วนเดนไดรต์ทำให้เกิดการกระตุ้น NMDA และการเกิดดีโพลาไรเซชันที่ตัวเซลล์ - แหล่งที่มาของข้อมูลป้อนเข้าสู่เซลล์:
- เรียนรู้โดยการสร้างเซลล์ประสาทใหม่ | - ได้รับแรงบันดาลใจจากเซลล์รูปพีระมิดในชั้นที่ 2/3 และ 5 ของเปลือกสมองส่วนหน้า - ซินแนปส์หลายพันตัว - เดนไดรต์ที่ทำงานอยู่: เซลล์สามารถจดจำรูปแบบเฉพาะได้หลายร้อยแบบ - แบบจำลองเดนไดรต์และสไปค์ NMDA โดยแต่ละอาร์เรย์ของตัวตรวจจับที่เกิดขึ้นพร้อมกันจะมีชุดของไซแนปส์ - เรียนรู้โดยการจำลองการเติบโตของไซแนปส์ใหม่ |
แอปพลิเคชัน
แบบจำลองเซลล์ประสาทแบบสไปค์ถูกนำไปใช้ในแอปพลิเคชันที่หลากหลายซึ่งต้องการการเข้ารหัสหรือการถอดรหัสจากชุดสไปค์ของเซลล์ประสาทในบริบทของประสาทเทียมและอินเทอร์เฟซระหว่างสมองกับคอมพิวเตอร์เช่นประสาทเทียมจอประสาทตา : [ 12 ] [ 98 ] [ 99 ] [ 100 ]หรือการควบคุมและการรับรู้แขนขาเทียม[ 101 ] [ 102 ] [ 103 ]แอปพลิเคชันเหล่านี้ไม่ได้เป็นส่วนหนึ่งของบทความนี้ สำหรับข้อมูลเพิ่มเติมเกี่ยวกับหัวข้อนี้ โปรดดูบทความหลัก
ความสัมพันธ์ระหว่างแบบจำลองเซลล์ประสาทเทียมและเซลล์ประสาทชีวภาพ
แบบจำลองพื้นฐานที่สุดของเซลล์ประสาทประกอบด้วยอินพุตที่มี เวกเตอร์ น้ำหนักไซแนปส์และฟังก์ชันการกระตุ้นหรือฟังก์ชันถ่ายโอนภายในเซลล์ประสาทซึ่งกำหนดเอาต์พุต นี่คือโครงสร้างพื้นฐานที่ใช้สำหรับเซลล์ประสาทเทียม ซึ่งในเครือข่ายประสาทมักมีลักษณะดังนี้

โดยที่y คือเอาต์พุตของ นิวรอนที่ i , x คือสัญญาณอินพุตของนิวรอนที่j , w คือน้ำหนักของไซแนปส์ (หรือความแข็งแรงของการเชื่อมต่อ) ระหว่างนิวรอนiและjและφคือฟังก์ชันการกระตุ้นแม้ว่าแบบจำลองนี้จะประสบความสำเร็จในการประยุกต์ใช้ในด้านการเรียนรู้ของเครื่อง แต่ก็เป็นแบบจำลองที่ไม่ดีสำหรับนิวรอนจริง (ทางชีววิทยา) เนื่องจากขาดการพึ่งพาเวลาในอินพุตและเอาต์พุต
เมื่ออินพุตถูกเปิดใช้งานในเวลา t และคงที่หลังจากนั้น เซลล์ประสาททางชีวภาพจะปล่อยชุดสไปค์ออกมา สิ่งสำคัญคือ ชุดสไปค์นี้ไม่สม่ำเสมอ แต่แสดงโครงสร้างเชิงเวลาที่มีลักษณะเฉพาะด้วยการปรับตัว การระเบิด หรือการระเบิดเริ่มต้นตามด้วยการยิงสไปค์อย่างสม่ำเสมอ โมเดลการรวมและยิงแบบทั่วไป เช่น โมเดลการรวมและยิงแบบเอ็กซ์โปเนนเชียลแบบปรับตัว โมเดลการตอบสนองสไปค์ หรือโมเดลการรวมและยิงแบบปรับตัว (เชิงเส้น) สามารถจับรูปแบบการยิงของเซลล์ประสาทเหล่านี้ได้[ 24 ] [ 25 ] [ 26 ]
ยิ่งไปกว่านั้น อินพุตของเซลล์ประสาทในสมองขึ้นอยู่กับเวลา อินพุตที่ขึ้นอยู่กับเวลาจะถูกแปลงโดยตัวกรองเชิงเส้นและไม่เชิงเส้นที่ซับซ้อนให้กลายเป็นชุดสไปค์ในเอาต์พุต อีกครั้ง โมเดลการตอบสนองสไปค์หรือโมเดลอินทิเกรตแอนด์ไฟร์แบบปรับตัวได้ช่วยให้สามารถทำนายชุดสไปค์ในเอาต์พุตสำหรับอินพุตที่ขึ้นอยู่กับเวลาใดๆ ก็ได้[ 22 ] [ 23 ]ในขณะที่เซลล์ประสาทเทียมหรืออินทิเกรตแอนด์ไฟร์แบบรั่วธรรมดาไม่สามารถทำได้
หากเราใช้แบบจำลอง Hodkgin-Huxley เป็นจุดเริ่มต้น แบบจำลอง integrate-and-fire ทั่วไปสามารถได้มาอย่างเป็นระบบในขั้นตอนการลดความซับซ้อนทีละขั้น ซึ่งได้แสดงให้เห็นอย่างชัดเจนสำหรับ แบบจำลอง integrate-and-fire แบบเอกซ์โป เนนเชียล [ 33 ]และแบบจำลองการตอบสนองสไปค์[ 60 ]
ในกรณีของการสร้างแบบจำลองเซลล์ประสาททางชีววิทยา จะใช้แบบจำลองทางกายภาพแทนนามธรรม เช่น "น้ำหนัก" และ "ฟังก์ชันการถ่ายโอน" เซลล์ประสาทเต็มไปด้วยและล้อมรอบด้วยไอออนที่มีน้ำเป็นองค์ประกอบ ซึ่งนำพาประจุไฟฟ้า เซลล์ประสาทถูกล้อมรอบด้วยเยื่อหุ้มเซลล์ที่เป็นฉนวน และสามารถรักษาระดับความเข้มข้นของไอออนที่มีประจุไว้ได้ทั้งสองด้าน ซึ่งกำหนดค่าความจุC การทำงานของเซลล์ประสาทเกี่ยวข้องกับการเคลื่อนที่ของไอออนเข้าไปในเซลล์ ซึ่งเกิดขึ้นเมื่อสารสื่อประสาททำให้ช่องไอออนบนเยื่อหุ้มเซลล์เปิดออก เราอธิบายสิ่งนี้ด้วยกระแสไฟฟ้า ทางกายภาพที่ขึ้นอยู่กับเวลา I ( t )พร้อมกับการเปลี่ยนแปลงของแรงดันไฟฟ้าหรือความแตกต่างของพลังงานศักย์ไฟฟ้าระหว่างเซลล์กับสิ่งแวดล้อม ซึ่งบางครั้งพบว่าส่งผลให้เกิดแรงดันไฟฟ้าพุ่งสูงขึ้นเรียกว่าศักย์การกระทำ ซึ่งเดินทางไปตามความยาวของเซลล์และกระตุ้นการปล่อยสารสื่อประสาทเพิ่มเติม ดังนั้น แรง ดันไฟฟ้าจึงเป็นปริมาณที่น่าสนใจและกำหนดโดยV ( t ) [ 19 ]
หากกระแสไฟฟ้าขาเข้าคงที่ เซลล์ประสาทส่วนใหญ่จะปล่อยสัญญาณไฟฟ้าออกมาอย่างสม่ำเสมอหลังจากช่วงเวลาปรับตัวหรือการระเบิดเริ่มต้น ความถี่ของการยิงสัญญาณอย่างสม่ำเสมอเพื่อตอบสนองต่อกระแสไฟฟ้าคงที่Iนั้น อธิบายได้ด้วยความสัมพันธ์ระหว่างความถี่และกระแสไฟฟ้า ซึ่งสอดคล้องกับฟังก์ชันถ่ายโอนของเครือข่ายประสาทเทียม ในทำนองเดียวกัน สำหรับแบบจำลองเซลล์ประสาทแบบสไปค์ทั้งหมด ฟังก์ชันการถ่ายโอนสามารถคำนวณได้ด้วยวิธีเชิงตัวเลข (หรือเชิงวิเคราะห์)

ทฤษฎีสายเคเบิลและแบบจำลองแบบแบ่งส่วน
แบบจำลองเชิงกำหนดข้างต้นทั้งหมดเป็นแบบจำลองเซลล์ประสาทแบบจุด เนื่องจากไม่ได้พิจารณาโครงสร้างเชิงพื้นที่ของเซลล์ประสาท อย่างไรก็ตาม เดนไดรต์มีส่วนช่วยในการแปลงอินพุตเป็นเอาต์พุต[ 104 ] [ 64 ] แบบจำลองเซลล์ประสาทแบบจุดเป็นคำอธิบายที่ถูกต้องในสามกรณี (i) หากกระแสอินพุตถูกฉีดเข้าไปในโซมาโดยตรง (ii) หากอินพุตไซแนปส์มาถึงที่หรือใกล้กับโซมาเป็นส่วนใหญ่ (ความใกล้ชิดถูกกำหนดโดยมาตราส่วนความยาว)แนะนำไว้ด้านล่าง (iii) หากไซแนปส์มาถึงที่ใดก็ได้บนเดนไดรต์ แต่เดนไดรต์เป็นเส้นตรงโดยสมบูรณ์ ในกรณีสุดท้าย สายเคเบิลทำหน้าที่เป็นตัวกรองเชิงเส้น คุณสมบัติของตัวกรองเชิงเส้นเหล่านี้สามารถรวมอยู่ในสูตรของแบบจำลองอินทิเกรตแอนด์ไฟร์ทั่วไป เช่นแบบจำลองการตอบสนองของสไปค์

คุณสมบัติของตัวกรองสามารถคำนวณได้จาก สม การของสายเคเบิล
ลองพิจารณาเยื่อหุ้มเซลล์ในรูปทรงของสายเคเบิลทรงกระบอก ตำแหน่งบนสายเคเบิลแทนด้วย x และแรงดันไฟฟ้าคร่อมเยื่อหุ้มเซลล์แทนด้วย V สายเคเบิลนี้มีลักษณะเฉพาะคือความต้านทานตามแนวยาว ต่อหน่วยความยาวและความต้านทานของเยื่อหุ้มเซลล์ถ้าทุกอย่างเป็นแบบเชิงเส้น แรงดันไฟฟ้าจะเปลี่ยนแปลงไปตามฟังก์ชันของเวลา


| 19 |

เรานำเสนอมาตราส่วนความยาว ทางด้านซ้ายและค่าคงที่เวลา ทางด้านขวาสมการของสายเคเบิลสามารถเขียนได้ในรูปแบบที่อาจเป็นที่รู้จักมากที่สุดดังนี้:


| 20 |

สมการสายเคเบิลข้างต้นใช้ได้กับสายเคเบิลทรงกระบอกเดี่ยวเท่านั้น
ทฤษฎีสายเคเบิลเชิงเส้นอธิบาย โครงสร้าง เดนไดรต์ของเซลล์ประสาทว่าเป็นโครงสร้างทรงกระบอกที่มีการแตกแขนง อย่างเป็นระเบียบ คล้ายกับกิ่งไม้ สำหรับทรงกระบอกเดี่ยวหรือต้นไม้ทั้งต้น ค่าการนำไฟฟ้าขาเข้าแบบคงที่ที่ฐาน (จุดที่ต้นไม้เชื่อมต่อกับตัวเซลล์หรือขอบเขตใดๆ) ถูกกำหนดดังนี้
- ,

โดยที่Lคือความยาวอิเล็กโทรนิกของทรงกระบอก ซึ่งขึ้นอยู่กับความยาว เส้นผ่านศูนย์กลาง และความต้านทาน อัลกอริทึมแบบเรียกซ้ำอย่างง่ายจะปรับขนาดเชิงเส้นตามจำนวนกิ่ง และสามารถใช้ในการคำนวณค่าการนำไฟฟ้าที่มีประสิทธิภาพของต้นไม้ได้ ซึ่งกำหนดโดย

โดยที่A = πldคือพื้นที่ผิวทั้งหมดของต้นไม้ที่มีความยาวทั้งหมดlและL คือความยาวอิเล็กโทรโทนิกทั้งหมด สำหรับเซลล์ประสาททั้งเซลล์ซึ่งค่าการนำไฟฟ้าของตัวเซลล์คือG และค่าการนำไฟฟ้าของเยื่อหุ้มเซลล์ต่อหน่วยพื้นที่คือG = G / Aเราจะพบค่าการนำไฟฟ้าทั้งหมด ของเซลล์ประสาท G สำหรับ ต้นเดนไดรต์ n ต้นโดยการบวกค่าการนำไฟฟ้าของต้นไม้และตัวเซลล์ทั้งหมดเข้าด้วยกัน ซึ่งกำหนดโดย

โดยที่เราสามารถหาค่าตัวประกอบการแก้ไขทั่วไปF ได้จากการทดลอง โดยสังเกตว่าG = G A F
แบบจำลองสายเคเบิลเชิงเส้นทำให้ง่ายขึ้นหลายประการเพื่อให้ได้ผลลัพธ์เชิงวิเคราะห์แบบปิด กล่าวคือ กิ่งก้านสาขาของเดนไดรต์จะต้องแตกแขนงเป็นคู่ที่ลดลงในรูปแบบคงที่ และเดนไดรต์เป็นเส้นตรง แบบจำลองแบบแบ่งส่วน[ 64 ]อนุญาตให้มีโทโพโลยีต้นไม้ที่ต้องการใดๆ ก็ได้ โดยมีกิ่งก้านและความยาวตามอำเภอใจ รวมถึงความไม่เป็นเชิงเส้นตามอำเภอใจด้วย โดยพื้นฐานแล้วมันคือการนำเดนไดรต์ที่ไม่เป็นเชิงเส้นมาใช้ในการคำนวณแบบไม่ต่อเนื่อง
แต่ละส่วนหรือช่องของเดนไดรต์จะถูกจำลองโดยทรงกระบอกตรงที่มีความยาวlและเส้นผ่านศูนย์กลางd ที่กำหนด ซึ่งเชื่อมต่อกับทรงกระบอกแตกแขนงจำนวนใดก็ได้ด้วยความต้านทานคงที่ เรากำหนดอัตราส่วนการนำไฟฟ้าของทรงกระบอกที่i เป็น B = G / G โดยที่และR คือความต้านทานระหว่างช่องปัจจุบันกับช่องถัดไป เราจะได้ชุดสมการสำหรับอัตราส่วนการนำไฟฟ้าเข้าและออกจากช่องโดยทำการแก้ไขค่าไดนามิกปกติB = B ดังนี้




โดยสมการสุดท้ายเกี่ยวข้องกับพ่อแม่และลูกสาวที่สาขาต่างๆ และเราสามารถวนซ้ำสมการเหล่านี้ไปเรื่อยๆ ผ่านโครงสร้างต้นไม้ จนกระทั่งถึงจุดที่เดนไดรต์เชื่อมต่อกับตัวเซลล์ (โซมา) ซึ่งอัตราส่วนการนำไฟฟ้าคือB จากนั้นค่าการนำไฟฟ้ารวมของเซลล์ประสาทสำหรับอินพุตคงที่จะได้รับจาก


ที่สำคัญคือ อินพุตแบบคงที่เป็นกรณีพิเศษมาก ในทางชีววิทยา อินพุตจะขึ้นอยู่กับเวลา นอกจากนี้ เดนไดรต์ก็ไม่ได้เป็นเส้นตรงเสมอไป
แบบจำลองแบบแบ่งส่วนช่วยให้สามารถรวมความไม่เป็นเชิงเส้นผ่านช่องไอออนที่วางไว้ที่ตำแหน่งใดๆ ตามเดนไดรต์ได้[ 104 ] [ 105 ]สำหรับอินพุตแบบคงที่ บางครั้งสามารถลดจำนวนส่วน (เพิ่มความเร็วในการคำนวณ) และยังคงรักษาคุณลักษณะทางไฟฟ้าที่โดดเด่นไว้ได้[ 106 ]
ข้อสันนิษฐานเกี่ยวกับบทบาทของเซลล์ประสาทในบริบทที่กว้างขึ้นของหลักการทำงานของสมอง
แผนการตรวจจับพลังงานโดยอาศัยสารสื่อประสาท
แผนการตรวจจับพลังงานตามสารสื่อประสาท[ 73 ] [ 80 ]ชี้ให้เห็นว่าเนื้อเยื่อประสาทดำเนินการตามขั้นตอนการตรวจจับแบบเรดาร์ทางเคมี

ดังแสดงในรูปที่ 6 แนวคิดหลักของสมมติฐานนี้คือการพิจารณาความเข้มข้นของสารสื่อประสาท การสร้างสารสื่อประสาท และอัตราการกำจัดสารสื่อประสาทเป็นปริมาณสำคัญในการดำเนินการตรวจจับ ในขณะที่อ้างถึงศักยภาพทางไฟฟ้าที่วัดได้เป็นผลข้างเคียงซึ่งสอดคล้องกับวัตถุประสงค์เชิงหน้าที่ของแต่ละขั้นตอนเฉพาะในบางเงื่อนไขเท่านั้น โครงร่างการตรวจจับคล้ายกับ "การตรวจจับพลังงาน" แบบเรดาร์ เนื่องจากประกอบด้วยการยกกำลังสองของสัญญาณ การรวมเชิงเวลา และกลไกการสลับเกณฑ์ เช่นเดียวกับเครื่องตรวจจับพลังงาน แต่ยังรวมถึงหน่วยที่เน้นขอบของสิ่งเร้าและความยาวหน่วยความจำที่แปรผันได้ (หน่วยความจำแปรผัน) ตามสมมติฐานนี้ ค่าเทียบเท่าทางสรีรวิทยาของสถิติการทดสอบพลังงานคือความเข้มข้นของสารสื่อประสาท และอัตราการยิงสอดคล้องกับกระแสสารสื่อประสาท ข้อดีของการตีความนี้คือ นำไปสู่คำอธิบายที่สอดคล้องกับหน่วย ซึ่งช่วยเชื่อมโยงระหว่างการวัดทางสรีรวิทยาไฟฟ้า การวัดทางชีวเคมี และผลลัพธ์ทางจิตวิทยา
หลักฐานที่ตรวจสอบใน[ 73 ] [ 80 ]ชี้ให้เห็นถึงความสัมพันธ์ระหว่างการทำงานกับการจำแนกประเภททางเนื้อเยื่อวิทยาดังต่อไปนี้:
- การแปลงสัญญาณกระตุ้นเป็นกำลังสองน่าจะเกิดขึ้นโดยเซลล์รับสัญญาณ
- เซลล์ประสาททำหน้าที่เน้นขอบของสิ่งเร้าและส่งสัญญาณผ่านสิ่งเร้า
- เซลล์เกลียทำหน้าที่สะสมสารสื่อประสาทตามช่วงเวลา การสะสมสารสื่อประสาทในระยะสั้นอาจเกิดขึ้นในเซลล์ประสาทบางชนิดด้วยเช่นกัน
- การสลับการทำงานเชิงตรรกะเกิดขึ้นในเซลล์เกลีย และเป็นผลมาจากการที่ความเข้มข้นของสารสื่อประสาทเกินระดับเกณฑ์ การข้ามเกณฑ์นี้ยังมาพร้อมกับการเปลี่ยนแปลงอัตราการรั่วไหลของสารสื่อประสาทด้วย
- การสลับการเคลื่อนไหวแบบเต็มรูปแบบทางกายภาพเกิดจากเซลล์กล้ามเนื้อและเป็นผลมาจากการที่ความเข้มข้นของสารสื่อประสาทในบริเวณรอบกล้ามเนื้อเกินเกณฑ์ที่กำหนด
โปรดทราบว่า แม้ว่าสัญญาณทางไฟฟ้าสรีรวิทยาในรูปที่ 6 มักจะคล้ายกับสัญญาณการทำงาน (กำลังสัญญาณ/ความเข้มข้นของสารสื่อประสาท/แรงกล้ามเนื้อ) แต่ก็มีบางขั้นตอนที่การสังเกตทางไฟฟ้าแตกต่างจากวัตถุประสงค์การทำงานของขั้นตอนนั้นๆ โดยเฉพาะอย่างยิ่ง Nossenson และคณะได้เสนอว่า การข้ามเกณฑ์ของเซลล์เกลียมีการทำงานที่แตกต่างอย่างสิ้นเชิงเมื่อเทียบกับสัญญาณทางไฟฟ้าสรีรวิทยาที่แผ่กระจายออกมา และสัญญาณหลังอาจเป็นเพียงผลข้างเคียงของการแตกของเซลล์เกลียเท่านั้น
ข้อคิดเห็นทั่วไปเกี่ยวกับมุมมองสมัยใหม่ของแบบจำลองทางวิทยาศาสตร์และวิศวกรรม
- แบบจำลองข้างต้นยังคงเป็นแบบจำลองในอุดมคติ ต้องมีการแก้ไขสำหรับพื้นที่ผิวเมมเบรนที่เพิ่มขึ้นเนื่องจากหนามเดนไดรต์จำนวนมาก อุณหภูมิที่สูงกว่าข้อมูลการทดลองที่อุณหภูมิห้องอย่างมีนัยสำคัญ และความไม่สม่ำเสมอในโครงสร้างภายในของเซลล์[ 17 ]ผลกระทบที่สังเกตได้บางอย่างไม่สอดคล้องกับแบบจำลองเหล่านี้ ตัวอย่างเช่น การเปลี่ยนแปลงอุณหภูมิ (โดยมีอุณหภูมิสุทธิเพิ่มขึ้นน้อยที่สุด) ของเมมเบรนระหว่างการแพร่กระจายศักย์ไฟฟ้าไม่สอดคล้องกับแบบจำลองที่อาศัยการจำลองเมมเบรนเป็นความต้านทานที่ต้องกระจายพลังงานเมื่อกระแสไหลผ่าน การหนาตัวชั่วคราวของเมมเบรนระหว่างการแพร่กระจายศักย์ไฟฟ้าก็ไม่ได้ถูกทำนายโดยแบบจำลองเหล่านี้เช่นกัน รวมถึงความจุที่เปลี่ยนแปลงและแรงดันไฟฟ้าที่พุ่งสูงขึ้นอันเป็นผลมาจากการหนาตัวนี้ก็ไม่ได้ถูกรวมเข้าไว้ในแบบจำลองเหล่านี้ การทำงานของยาสลบบางชนิด เช่น ก๊าซเฉื่อย ก็เป็นปัญหาสำหรับแบบจำลองเหล่านี้เช่นกัน แบบจำลองใหม่ เช่นแบบจำลองโซลิตอนพยายามอธิบายปรากฏการณ์เหล่านี้ แต่ยังไม่ได้รับการพัฒนาเท่าแบบจำลองเก่าและยังไม่ได้นำไปใช้อย่างกว้างขวาง
- มุมมองสมัยใหม่เกี่ยวกับบทบาทของแบบจำลองทางวิทยาศาสตร์ชี้ให้เห็นว่า "แบบจำลองทั้งหมดนั้นผิดพลาด แต่บางแบบก็มีประโยชน์" (Box and Draper, 1987; Gribbin, 2009; Paninski et al., 2009)
- ข้อสันนิษฐานล่าสุดชี้ให้เห็นว่าเซลล์ประสาทแต่ละเซลล์อาจทำหน้าที่เป็นชุดของหน่วยเกณฑ์อิสระ มีการเสนอแนะว่าเซลล์ประสาทอาจถูกกระตุ้นแบบไม่สมมาตรตามต้นกำเนิดของสัญญาณที่มาถึงเยื่อหุ้มเซลล์ผ่านทางกิ่งก้านเดนไดรต์ นอกจากนี้ยังมีการเสนอว่ารูปคลื่นสไปค์ขึ้นอยู่กับต้นกำเนิดของสิ่งเร้าด้วย[ 107 ]
ลิงก์ภายนอก
- พลวัตของเซลล์ประสาท: จากเซลล์ประสาทเดี่ยวไปจนถึงเครือข่ายและแบบจำลองของการรับรู้ (W. Gerstner, W. Kistler, R. Naud, L. Paninski, Cambridge University Press, 2014) [ 27 ] โดย เฉพาะบทที่ 6 - 10 เวอร์ชันออนไลน์ html
- แบบจำลองเซลล์ประสาทสไปค์[ 1 ] (W. Gerstner และ W. Kistler, สำนักพิมพ์มหาวิทยาลัยเคมบริดจ์, 2002)