ปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีน

ปฏิสัมพันธ์ระหว่างโปรตีน ( PPIs ) คือการสัมผัสทางกายภาพที่มีความจำเพาะสูง ซึ่งเกิดขึ้นระหว่างโมเลกุล โปรตีนสองโมเลกุลขึ้นไปอันเป็นผลจากกระบวนการทางชีวเคมีที่ควบคุมโดยปฏิสัมพันธ์ต่างๆ เช่นแรงไฟฟ้าสถิตพันธะไฮโดรเจนและผลกระทบจากแรงดึงดูดระหว่างโมเลกุลที่ไม่ชอบน้ำ ปฏิสัมพันธ์เหล่านี้ส่วนใหญ่เป็นการสัมผัสทางกายภาพที่มีการเชื่อมโยงระดับโมเลกุลระหว่างสายโซ่ ซึ่งเกิดขึ้นในเซลล์หรือในสิ่งมีชีวิตในบริบททางชีวโมเลกุลที่เฉพาะเจาะจง
โปรตีนมักไม่ทำงานโดยลำพัง เนื่องจากหน้าที่ของพวกมันมักถูกควบคุม กระบวนการระดับโมเลกุลหลายอย่างภายในเซลล์ดำเนินการโดยเครื่องจักรระดับโมเลกุลที่สร้างขึ้นจากส่วนประกอบโปรตีนจำนวนมากซึ่งจัดระเบียบโดยปฏิกิริยาระหว่างโปรตีน (PPIs) ปฏิสัมพันธ์ทางสรีรวิทยาเหล่านี้ประกอบขึ้นเป็นสิ่งที่เรียกว่าปฏิสัมพันธ์เชิงปฏิสัมพันธ์ (interactomics)ของสิ่งมีชีวิต ในขณะที่ PPIs ที่ผิดปกติเป็นพื้นฐานของโรคที่เกี่ยวข้องกับการสะสมตัวหลายชนิด เช่น โรคครอยซ์เฟลด์-จาคอบและโรคอัลไซเมอร์
PPIs ได้รับการศึกษาด้วยวิธีการมากมายและจากมุมมองที่แตกต่างกัน เช่นชีวเคมีเคมีควอนตัมพลศาสตร์โมเลกุลการส่งสัญญาณและอื่นๆ[ 1 ] [ 2 ] [ 3 ]ข้อมูลทั้งหมดนี้ทำให้สามารถสร้างเครือข่ายปฏิสัมพันธ์ของโปรตีนขนาดใหญ่ได้[ 4 ]ซึ่งคล้ายกับ เครือข่าย เมตาบอลิซึมหรือเครือข่ายทางพันธุกรรม/เอพิเจเนติกส์ซึ่งช่วยเสริมความรู้ในปัจจุบันเกี่ยวกับกระบวนการทางชีวเคมีและสาเหตุทางโมเลกุลของโรค ตลอดจนการค้นพบเป้าหมายโปรตีนที่อาจมีประโยชน์ในการรักษา
ตัวอย่าง
โปรตีนถ่ายโอนอิเล็กตรอน
ในปฏิกิริยาเมตาบอลิซึมหลายอย่าง โปรตีนที่ทำหน้าที่เป็นตัวนำอิเล็กตรอนจะจับกับเอนไซม์ที่ทำหน้าที่เป็นรีดักเทสหลังจากได้รับอิเล็กตรอนแล้ว มันจะแยกตัวออกและจับกับเอนไซม์ตัวถัดไปที่ทำหน้าที่เป็นออกซิเดส (เช่น ตัวรับอิเล็กตรอน) ปฏิสัมพันธ์ระหว่างโปรตีนเหล่านี้ขึ้นอยู่กับการจับกันอย่างจำเพาะเจาะจงระหว่างโปรตีนเพื่อให้แน่ใจว่าการถ่ายโอนอิเล็กตรอนมีประสิทธิภาพ ตัวอย่างเช่น ส่วนประกอบของระบบห่วงโซ่ฟอสโฟรีเลชันแบบออกซิเดทีฟของไมโทคอนเดรีย ได้แก่ ไซโตโครมซีรีดักเทส / ไซโตโครมซี / ไซโตโครมซีออกซิเดส; ระบบ P450 ของไมโครโซมและไมโท คอนเดรีย [ 5 ]
ในกรณีของระบบ P450 ในไมโทคอนเดรีย พบว่าสารตกค้างเฉพาะที่เกี่ยวข้องกับการจับกันของโปรตีนถ่ายโอนอิเล็กตรอนแอดรีโน ดอกซินกับรีดักเทส ได้แก่ สารตกค้าง Arg พื้นฐาน 2 ตัวบนพื้นผิวของรีดักเทส และสารตกค้าง Asp ที่เป็นกรด 2 ตัวบนแอดรีโนดอกซิน [ 6 ] งานวิจัยล่าสุดเกี่ยวกับวิวัฒนาการของรีดักเทสแสดงให้เห็นว่าสารตกค้างเหล่านี้ที่เกี่ยวข้องกับปฏิกิริยาระหว่างโปรตีนกับโปรตีนได้รับการอนุรักษ์ไว้ตลอดวิวัฒนาการของเอนไซม์นี้[ 7 ]
การส่งสัญญาณ
การทำงานของเซลล์ถูกควบคุมโดยสัญญาณภายนอกเซลล์ การส่งสัญญาณภายในและ/หรือตามแนวภายในของเซลล์ขึ้นอยู่กับปฏิกิริยาระหว่างโปรตีน (PPIs) ระหว่างโมเลกุลส่งสัญญาณต่างๆ การรวมตัวของเส้นทางส่งสัญญาณผ่าน PPIs เรียกว่าการส่งสัญญาณ (signal transduction ) และมีบทบาทสำคัญในกระบวนการทางชีววิทยาหลายอย่าง รวมถึงโรคต่างๆ มากมาย เช่นโรคพาร์กินสันและมะเร็ง
การขนส่งผ่านเยื่อหุ้มเซลล์
โปรตีนหนึ่งอาจทำหน้าที่ขนส่งโปรตีนอีกชนิดหนึ่ง (ตัวอย่างเช่น จากไซโตพลาซึมไปยังนิวเคลียสหรือในทางกลับกัน ในกรณีของ โปรตีนนำเข้าของ รูพรุนนิวเคลียส )
การเผาผลาญของเซลล์
ในกระบวนการสังเคราะห์ทางชีวภาพหลายอย่างเอนไซม์จะทำปฏิกิริยากันเพื่อสร้างสารประกอบขนาดเล็กหรือโมเลกุลขนาดใหญ่อื่นๆ
การหดตัวของกล้ามเนื้อ
สรีรวิทยาของการหดตัวของกล้ามเนื้อเกี่ยวข้องกับปฏิสัมพันธ์หลายอย่าง เส้นใย ไมโอซินทำหน้าที่เป็นมอเตอร์ระดับโมเลกุลและการจับกับแอคตินทำให้เส้นใยสามารถเลื่อนได้[ 8 ]นอกจากนี้ สมาชิกใน กลุ่ม โปรตีนที่เกี่ยวข้องกับหยดไขมันในกล้ามเนื้อโครงร่าง ยังเชื่อมโยงกับโปรตีนอื่นๆ เช่น ตัวกระตุ้นของไลเปสไตรกลีเซอไรด์ในไขมันและตัวกระตุ้นร่วม ของมัน คือ ยีนเปรียบเทียบการระบุ-58 เพื่อควบคุมการสลายไขมันในกล้ามเนื้อโครงร่าง
ประเภท
เพื่ออธิบายประเภทของปฏิกิริยาระหว่างโปรตีน (PPIs) โปรตีนสามารถมีปฏิสัมพันธ์กันในลักษณะ "ชั่วคราว" (เพื่อสร้างผลเฉพาะบางอย่างในช่วงเวลาสั้น ๆ เช่น การส่งสัญญาณ) หรือมีปฏิสัมพันธ์กับโปรตีนอื่น ๆ ในลักษณะ "เสถียร" เพื่อสร้างคอมเพล็กซ์ที่กลายเป็นเครื่องจักรระดับโมเลกุลภายในระบบสิ่งมีชีวิต การประกอบคอมเพล็กซ์ของโปรตีนสามารถส่งผลให้เกิดการสร้างคอมเพล็กซ์แบบโฮโมโอลิโกเมอริกหรือเฮเทโรโอลิโกเมอริกได้นอกเหนือจากคอมเพล็กซ์แบบดั้งเดิม เช่น เอนไซม์-สารยับยั้ง และแอนติบอดี-แอนติเจนแล้ว ยังสามารถเกิดปฏิสัมพันธ์ระหว่างโดเมน-โดเมน และโดเมน-เปปไทด์ได้อีกด้วย ความแตกต่างที่สำคัญอีกประการหนึ่งในการระบุปฏิกิริยาระหว่างโปรตีนคือวิธีการที่ใช้ในการกำหนด เนื่องจากมีเทคนิคที่วัดปฏิสัมพันธ์ทางกายภาพโดยตรงระหว่างคู่โปรตีน เรียกว่าวิธีการ "ไบนารี" ในขณะที่มีเทคนิคอื่น ๆ ที่วัดปฏิสัมพันธ์ทางกายภาพระหว่างกลุ่มของโปรตีน โดยไม่ต้องกำหนดคู่โปรตีน เรียกว่าวิธีการ "โคคอมเพล็กซ์"
โอลิโกเมอร์ชนิดเดียวกัน เทียบกับ โอลิโกเมอร์ต่างชนิดกัน
โฮโมโอลิโกเมอร์เป็นสารประกอบโมเลกุลขนาดใหญ่ที่ประกอบด้วย หน่วยย่อยโปรตีนเพียงประเภทเดียวการประกอบหน่วยย่อยโปรตีนนั้นถูกชี้นำโดยการสร้างปฏิสัมพันธ์ที่ไม่ใช่พันธะโควาเลนต์ในโครงสร้างควอเทอร์นารีของโปรตีน การทำลายโฮโมโอลิโกเมอร์เพื่อให้กลับไปเป็นโมโนเมอร์ แต่ละตัวเริ่มต้น มักต้องอาศัยการเสียสภาพของสารประกอบ[ 9 ]เอนไซม์โปรตีนพาหะ โปรตีนโครงสร้าง และปัจจัยควบคุมการถอดรหัส หลายชนิดทำหน้าที่ในรูปของโฮโมโอลิโกเมอร์ หน่วยย่อยโปรตีนที่แตกต่างกันจะโต้ตอบกันในเฮเทอโรโอลิโกเมอร์ ซึ่งจำเป็นต่อการควบคุมการทำงานของเซลล์หลายอย่าง ความสำคัญของการสื่อสารระหว่างโปรตีนต่างชนิดกันนั้นเห็นได้ชัดเจนยิ่งขึ้นในระหว่างเหตุการณ์การส่งสัญญาณของเซลล์ และปฏิสัมพันธ์ดังกล่าวเป็นไปได้ก็ต่อเมื่อมีโดเมนโครงสร้างภายในโปรตีน (ดังที่อธิบายไว้ด้านล่าง)
ปฏิสัมพันธ์ที่เสถียรเทียบกับปฏิสัมพันธ์ชั่วคราว
ปฏิสัมพันธ์ที่เสถียรเกี่ยวข้องกับโปรตีนที่โต้ตอบกันเป็นเวลานาน โดยเป็นส่วนหนึ่งของคอมเพล็กซ์ถาวรในฐานะหน่วยย่อย เพื่อทำหน้าที่ตามบทบาท โดยทั่วไปแล้วจะเป็นกรณีของโฮโมโอลิโกเมอร์ (เช่น ไซโตโครมซี ) และโปรตีนเฮเทอโรโอลิโกเมอร์บางชนิด เช่น หน่วยย่อยของATPaseในทางกลับกัน โปรตีนอาจโต้ตอบกับโปรตีนอื่น ๆ ในระยะเวลาสั้น ๆ และในลักษณะที่ย้อนกลับได้ เฉพาะในบริบทของเซลล์บางอย่างเท่านั้น เช่นชนิดของเซลล์ ระยะของวงจรเซลล์ปัจจัยภายนอก การมีอยู่ของโปรตีนที่จับกันอื่น ๆ เป็นต้น เช่นเดียวกับโปรตีนส่วนใหญ่ที่เกี่ยวข้องกับกระบวนการทางชีวเคมีปฏิสัมพันธ์เหล่านี้เรียกว่าปฏิสัมพันธ์ชั่วคราว ตัวอย่างเช่น ตัวรับที่เชื่อมต่อกับโปรตีน G บางชนิดจะจับกับโปรตีน G ชั่วคราว เมื่อถูกกระตุ้นโดยลิแกนด์ภายนอกเซลล์[ 10 ]บางชนิดเช่น ตัวรับมัสคารินิก M3 จะเชื่อมต่อกับโปรตีน G ก่อนการจับกันระหว่างตัวรับและลิแกนด์[ 11 ]ปฏิสัมพันธ์ระหว่างบริเวณโปรตีนที่มีโครงสร้างไม่เป็นระเบียบโดยเนื้อแท้กับโดเมนโปรตีนทรงกลม (เช่นMoRF ) เป็นปฏิสัมพันธ์ชั่วคราว[ 12 ]
พันธะโควาเลนต์เทียบกับพันธะที่ไม่ใช่โควาเลนต์
ปฏิสัมพันธ์แบบโคเวเลนต์เป็นปฏิสัมพันธ์ที่มีการเชื่อมโยงที่แข็งแกร่งที่สุดและเกิดขึ้นจากพันธะไดซัลไฟด์หรือการแบ่งปันอิเล็กตรอนแม้ว่าจะหายาก แต่ปฏิสัมพันธ์เหล่านี้เป็นตัวกำหนดในการปรับเปลี่ยนหลังการแปล บางอย่าง เช่นยูบิควิติเนชันและซูโมอิเลชันพันธะที่ไม่ใช่โคเวเลนต์มักจะเกิดขึ้นระหว่างปฏิสัมพันธ์ชั่วคราวโดยการรวมกันของพันธะที่อ่อนกว่า เช่นพันธะไฮโดรเจนปฏิสัมพันธ์ไอออนิกแรงแวนเดอร์วาลส์หรือพันธะไฮโดรโฟบิก[ 13 ]
บทบาทของน้ำ
โมเลกุลของน้ำมีบทบาทสำคัญในการโต้ตอบระหว่างโปรตีน[ 14 ] [ 15 ]โครงสร้างผลึกของคอมเพล็กซ์ที่ได้มาด้วยความละเอียดสูงจากโปรตีนที่แตกต่างกันแต่เป็นโฮโมล็อกกัน แสดงให้เห็นว่าโมเลกุลของน้ำที่ส่วนต่อประสานบางส่วนได้รับการอนุรักษ์ไว้ระหว่างคอมเพล็กซ์โฮโมล็อกกัน โมเลกุลของน้ำที่ส่วนต่อประสานส่วนใหญ่สร้างพันธะไฮโดรเจนกับคู่ของแต่ละคอมเพล็กซ์ กรดอะมิโนที่ส่วนต่อประสานบางส่วนหรือกลุ่มอะตอมของโปรตีนคู่หนึ่งมีส่วนร่วมในการโต้ตอบทั้งโดยตรงและผ่านน้ำกับโปรตีนคู่อื่น การโต้ตอบทางอ้อมสองครั้งซึ่งผ่านโมเลกุลของน้ำสองโมเลกุลมีจำนวนมากกว่าในคอมเพล็กซ์โฮโมล็อกกันที่มีความสัมพันธ์ต่ำ[ 16 ]การทดลองการกลายพันธุ์ที่ดำเนินการอย่างระมัดระวัง เช่น การเปลี่ยนกรดอะมิโนไทโรซีนเป็นฟีนิลอะลานีน แสดงให้เห็นว่าการโต้ตอบผ่านน้ำสามารถมีส่วนช่วยในพลังงานของการโต้ตอบได้[ 17 ]ดังนั้น โมเลกุลของน้ำอาจช่วยอำนวยความสะดวกในการโต้ตอบและการจดจำข้ามระหว่างโปรตีน
โครงสร้าง
โครงสร้างโมเลกุลของโปรตีนเชิงซ้อนจำนวนมากได้รับการไขปริศนาโดยเทคนิคการตกผลึกด้วยรังสีเอกซ์[ 18 ] [ 19 ]โครงสร้างแรกที่ได้รับการแก้ไขด้วยวิธีนี้คือไมโอโกลบิน ของ วาฬสเปิร์ม โดยเซอร์จอห์น โควดรีย์ เคนดรูว์ [ 20 ] ในเทคนิคนี้ มุมและความเข้มของลำแสงรังสีเอกซ์ที่เลี้ยวเบนโดยอะตอมของผลึกจะถูกตรวจจับในฟิล์ม ทำให้เกิดภาพสามมิติของความหนาแน่นของอิเล็กตรอนภายในผลึก[ 21 ]
ต่อมาการเรโซแนนซ์แม่เหล็กนิวเคลียร์ก็เริ่มถูกนำมาใช้โดยมีเป้าหมายเพื่อไขโครงสร้างโมเลกุลของโปรตีนเชิงซ้อน ตัวอย่างแรกๆ คือโครงสร้างของโดเมนที่จับกับแคลโมดูลิน [ 22 ] [ 23 ] เทคนิคนี้อาศัยการศึกษาคุณสมบัติทางแม่เหล็กของนิวเคลียสอะตอม จึงสามารถกำหนดคุณสมบัติทางกายภาพและเคมีของอะตอมหรือโมเลกุลที่เกี่ยวข้องได้ การเรโซแนนซ์แม่เหล็กนิวเคลียร์มีข้อดีในการระบุลักษณะ PPIs ที่อ่อนแอ[ 24 ]
โดเมนปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีน
โปรตีนบางชนิดมีโครงสร้าง เฉพาะ หรือลำดับโมทีฟที่ช่วยในการจับกับโปรตีนอื่นๆ ตัวอย่างของโครงสร้างเฉพาะดังกล่าวมีดังนี้:
- โดเมน Src homology 2 (SH2)
- โดเมน SH2 ประกอบด้วยโครงสร้างแผ่นเบต้าบิดเกลียวสามสายประกบด้วยอัลฟาเฮลิกซ์สองอัน การมีอยู่ของช่องจับยึดที่ลึกซึ่งมีความสัมพันธ์สูงกับฟอสโฟไทโรซีนแต่ไม่ใช่ฟอสโฟเซอรีนหรือฟอสโฟทรีโอนีนเป็นสิ่งจำเป็นสำหรับการจดจำโปรตีนที่ถูกฟอสโฟริเลตด้วยไทโรซีน โดยเฉพาะอย่าง ยิ่งตัวรับปัจจัย การเจริญเติบโต ที่ถูกฟอสโฟริเลตด้วยตนเอง โปรตีนที่จับกับตัวรับปัจจัยการเจริญเติบโตและฟอสโฟลิเปส C γ เป็นตัวอย่างของโปรตีนที่มีโดเมน SH2 [ 25 ]
- โดเมน Src homology 3 (SH3)
- ในเชิงโครงสร้าง โดเมน SH3 ประกอบด้วยเบต้าบาร์เรลที่เกิดจากแผ่นเบต้าสองแผ่นที่ตั้งฉากกันและสายเบต้าสามสายที่ขนานกัน โดเมนเหล่านี้จดจำ ลำดับที่อุดมไป ด้วยโพรลีนเช่น โครงสร้างเกลียวโพลีโพรลีนประเภท II (โมทีฟ PXXP) ในโปรตีนส่งสัญญาณของเซลล์ เช่น โปรตีนไทโรซีนไคเนสและโปรตีนที่จับกับตัวรับปัจจัยการเจริญเติบโต 2 ( Grb2 ) [ 25 ]
- โดเมนจับฟอสโฟไทโรซีน (PTB)
- โดเมน PTB โต้ตอบกับลำดับที่มีกลุ่มฟอสโฟไทโรซีน โดเมนเหล่านี้สามารถพบได้ใน สาร ตั้งต้นของตัวรับอินซูลิน[ 25 ]
- โดเมน LIM
- โดเมน LIM ถูกระบุครั้งแรกในปัจจัยการถอดรหัสโฮมีโอโดเมน สามตัว (lin11, is11 และ mec3) นอกจากโปรตีนโฮมีโอโดเมน เหล่านี้ และโปรตีนอื่นๆ ที่เกี่ยวข้องกับการพัฒนาแล้ว โดเมน LIM ยังถูกระบุในโปรตีนที่ไม่ใช่โฮมีโอโดเมนที่มีบทบาทสำคัญในการจำแนกเซลล์การเชื่อมโยงกับโครงร่างเซลล์และการชราภาพโดเมนเหล่านี้มีโมทีฟนิ้วZn 2+ ที่อุดมไปด้วยซิสเทอีนแบบคู่ และครอบคลุมลำดับคอนเซนซัส CX2CX16-23HX2CX2CX2CX16-21CX2C/H/D โดเมน LIM จับกับโดเมน PDZ ปัจจัยการถอดรหัส bHLH และโดเมน LIM อื่นๆ[ 25 ]
- โดเมน Sterile alpha motif (SAM)
- โดเมน SAM ประกอบด้วยเกลียวห้าเกลียวที่ก่อตัวเป็นแพ็คเกจขนาดกะทัดรัดที่มีแกนไฮโดรโฟบิก ที่อนุรักษ์ไว้ โดเมนเหล่านี้ซึ่งพบได้ในตัวรับ Ephและโมเลกุลปฏิสัมพันธ์ของสโตรมัล ( STIM ) เป็นต้น จะจับกับโปรตีนที่ไม่มีโดเมน SAM และดูเหมือนว่าจะมีความสามารถในการจับกับRNAด้วย[ 25 ]
- โดเมน PDZ
- โดเมน PDZ ถูกระบุครั้งแรกในกัวนิเลตไคเนส 3 ชนิด ได้แก่ PSD-95, DlgA และ ZO-1 โดเมนเหล่านี้จดจำโมทีฟไตรเปปไทด์ปลายคาร์บอกซี (S/TXV) โดเมน PDZ อื่นๆ หรือโดเมน LIMและจับกับพวกมันผ่านลำดับเปปไทด์สั้นๆ ที่มี สารตกค้างไฮโดรโฟบิก ที่ปลาย Cโปรตีนบางชนิดที่ระบุว่ามีโดเมน PDZ เป็นโปรตีนโครงสร้างหรือดูเหมือนจะมีส่วนเกี่ยวข้องกับการประกอบตัวรับไอออนและการสร้างคอมเพล็กซ์ตัวรับ-เอนไซม์[ 25 ]
- โดเมน FERM
- โดเมน FERM ประกอบด้วยสารตกค้างพื้นฐานที่สามารถจับกับPtdIns(4,5)P ได้TalinและFocal Adhesion Kinase (FAK) เป็นโปรตีนสองชนิดที่มีโดเมน FERM [ 25 ]
- โดเมนความเหมือนของแคลโพนิน (CH)
- โดเมนโฮโมโลจีเพล็กสตริน
- โดเมน Pleckstrin homology จะจับกับฟอสโฟอิโนซิไทด์และโดเมนกรดในโปรตีนส่งสัญญาณ
- โดเมน WW
- โดเมน WW จะจับกับลำดับที่มีโพรลีนเป็นองค์ประกอบหลัก
- ลวดลาย WSxWS
- พบในตัวรับไซโตไคน์
คุณสมบัติของอินเทอร์เฟซ
การศึกษาโครงสร้างโมเลกุลสามารถให้รายละเอียดอย่างละเอียดเกี่ยวกับอินเทอร์เฟซที่ช่วยให้เกิดปฏิสัมพันธ์ระหว่างโปรตีน เมื่อทำการวิเคราะห์อินเทอร์เฟซ PPI สิ่งสำคัญคือต้องคำนึงถึงประเภทของคอมเพล็กซ์ด้วย[ 9 ]
พารามิเตอร์ที่ประเมิน ได้แก่ ขนาด (วัดเป็นมิติสัมบูรณ์Ųหรือพื้นที่ผิวที่เข้าถึงตัวทำละลายได้ (SASA) ) รูปร่าง ความเข้ากันได้ระหว่างพื้นผิว แนวโน้ม ของอินเทอร์เฟซตกค้าง ความชอบน้ำ การแบ่งส่วนและโครงสร้างทุติยภูมิ และการเปลี่ยนแปลงโครงสร้างเมื่อเกิดการสร้างเชิงซ้อน[ 9 ]
อินเทอร์เฟซ PPI ส่วนใหญ่สะท้อนถึงองค์ประกอบของพื้นผิวโปรตีนมากกว่าแกนกลางของโปรตีน แม้ว่าจะอุดมไปด้วยสารตกค้างที่ไม่ชอบน้ำ โดยเฉพาะอย่างยิ่งสารตกค้างอะโรมาติก[ 26 ]อินเทอร์เฟซ PPI มีพลวัตและมักจะเป็นระนาบ แม้ว่าอาจเป็นทรงกลมและยื่นออกมาได้เช่นกัน[ 27 ]จากโครงสร้างสามแบบ ได้แก่อินซูลินไดเม อร์ คอมเพล็กซ์ไตรป ซิน - สารยับยั้งไตร ปซินจากตับอ่อน และออก ซีฮีโมโกลบิน ไซรัส โชเทียและโจเอล จานิน พบว่าพื้นที่ผิวระหว่าง 1,130 ถึง 1,720 Ų ถูกกำจัดออกจากการสัมผัสกับน้ำ ซึ่งบ่งชี้ว่าความไม่ชอบน้ำเป็นปัจจัยสำคัญในการทำให้ PPI มีเสถียรภาพ[ 28 ]การศึกษาในภายหลังได้ปรับปรุงพื้นที่ผิวที่ถูกฝังของปฏิสัมพันธ์ส่วนใหญ่ให้เป็น 1,600±350 Ų อย่างไรก็ตามยังพบอินเทอร์เฟซปฏิสัมพันธ์ที่ใหญ่กว่ามาก และเกี่ยวข้องกับการเปลี่ยนแปลงที่สำคัญในโครงสร้างของคู่ปฏิสัมพันธ์หนึ่ง[ 18 ]อินเทอร์เฟซ PPIs แสดงให้เห็นถึงความสมบูรณ์แบบทั้งในด้านรูปร่างและไฟฟ้าสถิต[ 9 ] [ 11 ]
ระเบียบข้อบังคับ
- ความเข้มข้นของโปรตีน ซึ่งได้รับผลกระทบจากระดับการแสดงออกและอัตราการย่อยสลาย
- ความสัมพันธ์ของโปรตีนกับโปรตีนหรือลิแกนด์ที่จับกับโปรตีนอื่นๆ;
- ความเข้มข้นของลิแกนด์ ( สารตั้งต้นไอออนฯลฯ)
- มีโปรตีนกรดนิวคลีอิกและไอออน อื่นๆ อยู่ ด้วย
- สนามไฟฟ้าที่อยู่รอบโปรตีน
- การเกิดการดัดแปลงพันธะโควาเลนต์;
วิธีการทดลอง
มีวิธีการมากมายในการตรวจจับ[ 1 ] [ 29 ]แต่ละวิธีมีจุดแข็งและจุดอ่อนของตัวเอง โดยเฉพาะอย่างยิ่งในเรื่องความไวและความจำเพาะของวิธีการ วิธีการที่มีปริมาณงานสูงที่เป็นที่นิยมและใช้กันอย่างแพร่หลายมากที่สุด ได้แก่การคัดกรองยีสต์แบบทูไฮบริดและการทำให้บริสุทธิ์ ด้วยความสัมพันธ์ ร่วมกับการ วิเคราะห์มวลสาร

การคัดกรองยีสต์ทูไฮบริด
ระบบนี้ได้รับการอธิบายครั้งแรกในปี 1989 โดย Fields และ Song โดยใช้Saccharomyces cerevisiaeเป็นแบบจำลองทางชีวภาพ[ 30 ] [ 31 ]ยีสต์ทูไฮบริดช่วยให้สามารถระบุ PPIs แบบคู่ (วิธีไบนารี) ในร่างกายได้ โดยที่โปรตีนทั้งสองจะถูกทดสอบสำหรับการโต้ตอบโดยตรงทางชีวฟิสิกส์ Y2H ขึ้นอยู่กับการสร้างใหม่ของการทำงานของปัจจัยการถอดรหัสของยีสต์ Gal4 และการกระตุ้นตัวรายงานที่เลือกได้ เช่น His3 ในการทดสอบการโต้ตอบของโปรตีนสองชนิด จะมีการสร้างโครงสร้างการแสดงออกของโปรตีนสองแบบ: โปรตีนหนึ่ง (X) ถูกเชื่อมเข้ากับโดเมนการจับ DNA ของ Gal4 (DB) และโปรตีนที่สอง (Y) ถูกเชื่อมเข้ากับโดเมนการกระตุ้นของ Gal4 (AD) ในการทดสอบ เซลล์ยีสต์จะถูกแปลงสภาพด้วยโครงสร้างเหล่านี้ การถอดรหัสของยีนรายงานจะไม่เกิดขึ้นเว้นแต่เหยื่อ (DB-X) และตัวถูกล่า (AD-Y) จะโต้ตอบกันและสร้างปัจจัยการถอดรหัส Gal4 ที่ทำงานได้ ดังนั้น ปฏิสัมพันธ์ระหว่างโปรตีนสามารถอนุมานได้จากการปรากฏตัวของผลิตภัณฑ์ที่เกิดจากการแสดงออกของยีนรายงาน[ 13 ] [ 32 ]ในกรณีที่ยีนรายงานแสดงออกเอนไซม์ที่ช่วยให้ยีสต์สังเคราะห์กรดอะมิโนหรือนิวคลีโอไทด์ที่จำเป็น การเจริญเติบโตของยีสต์ภายใต้สภาวะสื่อคัดเลือกบ่งชี้ว่าโปรตีนทั้งสองที่ทดสอบมีปฏิสัมพันธ์กัน เมื่อเร็วๆ นี้ มีการเผยแพร่ซอฟต์แวร์เพื่อตรวจจับและจัดลำดับความสำคัญของปฏิสัมพันธ์ของโปรตีน[ 33 ] [ 34 ]
แม้ว่าระบบยีสต์ทูไฮบริดจะมีประโยชน์ แต่ก็มีข้อจำกัดอยู่บ้าง โดยใช้ยีสต์เป็นระบบโฮสต์หลัก ซึ่งอาจเป็นปัญหาเมื่อศึกษาโปรตีนที่มีการดัดแปลงหลังการแปลรหัสเฉพาะของสัตว์เลี้ยงลูกด้วยนม จำนวน PPIs ที่ระบุได้มักจะต่ำเนื่องจากอัตราการเกิดผลลบเท็จสูง[ 35 ]และยัง ประเมิน โปรตีนเยื่อหุ้มเซลล์ ต่ำกว่า ความเป็นจริงอีกด้วย[ 36 ] [ 37 ]
ในระหว่างการศึกษา นักชีววิทยาโมเลกุลAnton Yuryevใช้การคัดกรองยีสต์ทูไฮบริดเพื่อระบุปฏิสัมพันธ์ของโปรตีนใหม่ที่เชื่อมโยงการถอดรหัสกับการดัดแปลงหลังการถอดรหัส ซึ่งแสดงให้เห็นถึงประโยชน์ของวิธีการเหล่านี้ในการเปิดเผย PPIs ที่เฉพาะเจาะจงไอโซฟอร์ม[ 38 ] ในการศึกษาเบื้องต้นที่ใช้ Y2H การควบคุมที่เหมาะสมสำหรับผลบวกเท็จ (เช่น เมื่อ DB-X กระตุ้นยีนรายงานโดยไม่มี AD-Y) มักไม่ได้ดำเนินการ ทำให้มีอัตราผลบวกเท็จสูงกว่าปกติ ต้องมีการนำกรอบการทำงานเชิงประจักษ์มาใช้เพื่อควบคุมผลบวกเท็จเหล่านี้[ 39 ]ข้อจำกัดในการครอบคลุมโปรตีนเยื่อหุ้มเซลล์ที่ต่ำกว่าได้รับการแก้ไขโดยการเกิดขึ้นของยีสต์ทูไฮบริดรูปแบบต่างๆ เช่น เมมเบรนยีสต์ทูไฮบริด (MYTH) [ 37 ]และระบบสปลิตยูบิควิติน[ 32 ]ซึ่งไม่จำกัดเฉพาะปฏิสัมพันธ์ที่เกิดขึ้นในนิวเคลียส และระบบแบคทีเรียทูไฮบริดที่ดำเนินการในแบคทีเรีย[ 40 ]

การทำให้บริสุทธิ์โดยอาศัยความสัมพันธ์ร่วมกับการวิเคราะห์มวลสารด้วยสเปกโทรเมตรี
การทำให้บริสุทธิ์ด้วยแอฟฟินิตีร่วมกับแมสสเปกโทรเมตรีส่วนใหญ่จะตรวจจับปฏิสัมพันธ์ที่เสถียรและบ่งชี้ PPIs ที่มีฟังก์ชันในร่างกายได้ดีกว่า[ 41 ] [ 32 ]วิธีนี้เริ่มต้นด้วยการทำให้บริสุทธิ์ของโปรตีนที่มีแท็ก ซึ่งมักจะแสดงออกในเซลล์ที่ ความเข้มข้น ในร่างกายและโปรตีนที่ทำปฏิกิริยาด้วย (การทำให้บริสุทธิ์ด้วยแอฟฟินิตี) หนึ่งในวิธีการที่มีข้อได้เปรียบและใช้กันอย่างแพร่หลายที่สุดในการทำให้โปรตีนบริสุทธิ์โดยมีพื้นหลังปนเปื้อนต่ำมากคือการทำให้บริสุทธิ์ด้วยแอฟฟินิตีแบบคู่ขนานซึ่งพัฒนาโดย Bertrand Seraphin และ Matthias Mann และเพื่อนร่วมงานที่เกี่ยวข้อง จากนั้น PPIs สามารถวิเคราะห์ได้ด้วยแมสสเปกโทรเมตรีโดยใช้วิธีการต่างๆ ได้แก่ การรวมทางเคมี การรวมทางชีวภาพหรือเมตาบอลิซึม (SILAC) และวิธีการที่ไม่ใช้ฉลาก[ 9 ]นอกจากนี้ทฤษฎีเครือข่ายยังถูกนำมาใช้เพื่อศึกษาชุดปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีนที่ระบุทั้งหมดในเซลล์[ 4 ]
อาร์เรย์โปรตีนที่ตั้งโปรแกรมได้ด้วยกรดนิวคลีอิก (NAPPA)
ระบบนี้ได้รับการพัฒนาขึ้นครั้งแรกโดย LaBaer และคณะในปี 2547 โดยใช้ระบบการถอดรหัสและการแปลโปรตีนในหลอดทดลอง พวกเขาใช้แม่แบบ DNA ที่เข้ารหัสยีนที่สนใจซึ่งเชื่อมต่อกับโปรตีน GST และตรึงไว้บนพื้นผิวแข็ง แอนติบอดีต่อต้าน GST และพลาสมิด DNA ที่ติดไบโอตินถูกจับยึดไว้บนสไลด์ที่เคลือบด้วยอะมิโนโพรพิลไตรเอทอกซีไซเลน (APTES) BSA สามารถเพิ่มประสิทธิภาพการจับยึดของ DNA ได้ พลาสมิด DNA ที่ติดไบโอตินถูกจับยึดโดยอะวิดิน โปรตีนใหม่ถูกสังเคราะห์โดยใช้ระบบการแสดงออกที่ปราศจากเซลล์ เช่น ไลเซตของเรติคิวโลไซต์กระต่าย (RRL) จากนั้นโปรตีนใหม่จะถูกจับผ่านแอนติบอดีต่อต้าน GST ที่ยึดติดอยู่บนสไลด์ เพื่อทดสอบปฏิกิริยาระหว่างโปรตีนกับโปรตีน cDNA ของโปรตีนเป้าหมายและ cDNA ของโปรตีนที่ต้องการตรวจสอบจะถูกตรึงไว้บนสไลด์ที่เคลือบเดียวกัน โดยใช้ระบบการถอดรหัสและการแปลโปรตีนในหลอดทดลอง โปรตีนเป้าหมายและโปรตีนที่ต้องการตรวจสอบจะถูกสังเคราะห์จากสารสกัดเดียวกัน โปรตีนเป้าหมายถูกจับกับอาร์เรย์โดยแอนติบอดีที่เคลือบอยู่บนสไลด์ และใช้โปรตีนสอบถามเพื่อตรวจสอบอาร์เรย์ โปรตีนสอบถามถูกติดแท็กด้วยอีพิโทปเฮมากลูตินิน (HA) ดังนั้น ปฏิสัมพันธ์ระหว่างโปรตีนทั้งสองจึงถูกมองเห็นได้ด้วยแอนติบอดีต่อต้าน HA [ 42 ] [ 43 ]
การเสริมยีนภายใน
เมื่อพอลิเปปไทด์หลายชุดที่เข้ารหัสโดยยีนก่อตัวเป็นคอมเพล็กซ์ โครงสร้างโปรตีนนี้เรียกว่ามัลติเมอร์ เมื่อมัลติเมอร์ก่อตัวจากพอลิเปปไทด์ที่ผลิตโดย อัลลี ลกลายพันธุ์ สองแบบที่แตกต่างกัน ของยีนเฉพาะ มัลติเมอร์แบบผสมอาจแสดงกิจกรรมการทำงานที่มากกว่ามัลติเมอร์ที่ไม่ผสมซึ่งก่อตัวขึ้นจากอัลลีลกลายพันธุ์แต่ละแบบเพียงอย่างเดียว ในกรณีเช่นนี้ ปรากฏการณ์นี้เรียกว่าการเสริมกันภายในยีน (หรือเรียกว่าการเสริมกันระหว่างอัลลีล) การเสริมกันภายในยีนได้รับการพิสูจน์แล้วในยีนต่างๆ มากมายในสิ่งมีชีวิตหลากหลายชนิด รวมถึงเชื้อราNeurospora crassa , Saccharomyces cerevisiaeและSchizosaccharomyces pombe ; แบคทีเรียSalmonella typhimurium ; ไวรัสแบคทีริโอเฟจ T4 [ 44 ] ซึ่ง เป็นไวรัส RNA [ 45 ]และมนุษย์[ 46 ] ในการศึกษาดังกล่าว มักมีการแยกและแมป การกลายพันธุ์ จำนวนมาก ที่บกพร่องในยีนเดียวกันในลำดับเชิงเส้นตามความถี่ของการรวมตัวใหม่เพื่อสร้างแผนที่ทางพันธุกรรมของยีน มีการทดสอบตัวกลายพันธุ์แยกกันในรูปแบบคู่เพื่อวัดการเสริมกัน การวิเคราะห์ผลลัพธ์จากการศึกษาดังกล่าวทำให้ได้ข้อสรุปว่าการเสริมกันภายในยีนโดยทั่วไปเกิดขึ้นจากปฏิสัมพันธ์ของโมโนเมอร์โพลีเปปไทด์ที่บกพร่องต่างกันเพื่อสร้างมัลติเมอร์[ 47 ] ยีนที่เข้ารหัสโพลีเปปไทด์ที่สร้างมัลติเมอร์ดูเหมือนจะพบได้ทั่วไป การตีความข้อมูลอย่างหนึ่งคือ โมโนเมอร์โพลีเปปไทด์มักจะเรียงตัวกันในมัลติเมอร์ในลักษณะที่โพลีเปปไทด์กลายพันธุ์ที่บกพร่องในบริเวณใกล้เคียงในแผนที่ทางพันธุกรรมมีแนวโน้มที่จะสร้างมัลติเมอร์แบบผสมที่ทำงานได้ไม่ดี ในขณะที่โพลีเปปไทด์กลายพันธุ์ที่บกพร่องในบริเวณที่ห่างไกลมีแนวโน้มที่จะสร้างมัลติเมอร์แบบผสมที่ทำงานได้อย่างมีประสิทธิภาพมากขึ้น การโต้ตอบโดยตรงของโปรตีนเกิดใหม่สองตัวที่เกิดขึ้นจากไรโบโซม ที่อยู่ใกล้เคียง ดูเหมือนจะเป็นกลไกทั่วไปสำหรับการก่อตัวของโฮโมโอลิโกเมอร์ (มัลติเมอร์) [ 48 ] มีการระบุโอลิโกเมอร์โปรตีนหลายร้อยชนิดที่ประกอบกันในเซลล์มนุษย์โดยการโต้ตอบดังกล่าว[ 48 ] รูปแบบการโต้ตอบที่พบมากที่สุดคือระหว่างบริเวณปลาย N ของโปรตีนที่โต้ตอบกัน การก่อตัวของไดเมอร์ดูเหมือนจะสามารถเกิดขึ้นได้โดยไม่ขึ้นอยู่กับเครื่องจักรประกอบเฉพาะ แรงระหว่างโมเลกุลที่น่าจะรับผิดชอบต่อการจดจำตนเองและการก่อตัวของมัลติเมอร์ได้รับการกล่าวถึงโดย Jehle [ 49 ]
วิธีการอื่นๆ ที่เป็นไปได้
เทคนิคที่หลากหลายในการระบุ PPIs ได้เกิดขึ้นพร้อมกับความก้าวหน้าทางเทคโนโลยี ซึ่งรวมถึงการตกตะกอนร่วมภูมิคุ้มกันไมโครอาร์เรย์โปรตีน การปั่น เหวี่ยง ความเร็วสูงเชิงวิเคราะห์ การกระเจิงแสง สเปกโทรสโกปี ฟลูออเรสเซนซ์ การ ทำแผนที่ปฏิสัมพันธ์ของสัตว์เลี้ยงลูกด้วยนมโดยใช้การเรืองแสง (LUMIER) ระบบการถ่ายโอนพลังงานเรโซแนนซ์ กับดักปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีนของสัตว์เลี้ยงลูกด้วยนม พื้น ผิวชีวภาพที่สลับได้ด้วยไฟฟ้าการทดสอบการเสริมโปรตีน-ชิ้นส่วนตลอดจนการวัดแบบเรียลไทม์โดยไม่ต้องใช้ฉลากโดยใช้เรโซแนนซ์พลาสมอนบนพื้นผิวและแคลอริเมตรี[ 36 ] [ 37 ]
วิธีการคำนวณ
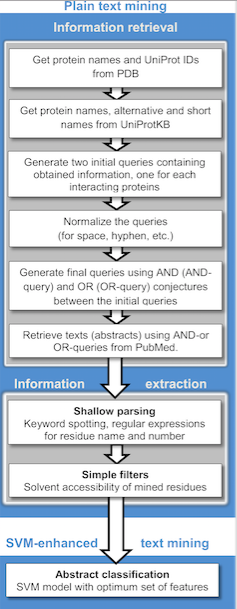
การตรวจจับและระบุลักษณะของปฏิกิริยาระหว่างโปรตีน (PPIs) ด้วยวิธีการทดลองนั้นต้องใช้แรงงานและเวลามาก อย่างไรก็ตาม PPIs จำนวนมากสามารถทำนายได้ด้วยวิธีการคำนวณ โดยปกติจะใช้ข้อมูลจากการทดลองเป็นจุดเริ่มต้น นอกจากนี้ ยังมีการพัฒนาวิธีการที่ช่วยให้สามารถทำนาย PPI ได้ตั้งแต่เริ่มต้น โดยไม่ต้องมีหลักฐานใดๆ เกี่ยวกับปฏิกิริยาเหล่านี้มาก่อน
วิธีการบริบททางจีโนม
วิธีการ Rosetta Stone หรือ Domain Fusion นั้นอิงตามสมมติฐานที่ว่าโปรตีนที่โต้ตอบกันบางครั้งจะถูกรวมเข้าเป็นโปรตีนเดียวในจีโนมอื่น [ 50 ]ดังนั้น เราจึงสามารถทำนายได้ว่าโปรตีนสองชนิดอาจโต้ตอบกันหรือไม่ โดยการพิจารณาว่าโปรตีนแต่ละชนิดมีความคล้ายคลึงกันของลำดับที่ไม่ทับซ้อนกันกับบริเวณของลำดับโปรตีนเดียวในจีโนมอื่นหรือไม่
วิธีการ Conserved Neighborhoodอิงตามสมมติฐานที่ว่า หากยีนที่เข้ารหัสโปรตีนสองชนิดอยู่ติดกันบนโครโมโซมในจีโนมจำนวนมาก แสดงว่ายีนเหล่านั้นน่าจะมีความสัมพันธ์กันในเชิงหน้าที่ (และอาจมีการโต้ตอบกันทางกายภาพด้วย) [ 51 ]
วิธีการ Phylogenetic Profileอิงตามสมมติฐานที่ว่า หากโปรตีนสองตัวขึ้นไปปรากฏหรือหายไปพร้อมกันในจีโนมหลายๆ จีโนม แสดงว่าโปรตีนเหล่านั้นน่าจะมีความสัมพันธ์กันในเชิงหน้าที่ [ 51 ]ดังนั้น โปรตีนที่อาจมีปฏิสัมพันธ์กันสามารถระบุได้โดยการตรวจสอบการปรากฏหรือไม่ปรากฏของยีนในจีโนมหลายๆ จีโนม และเลือกยีนที่ปรากฏหรือหายไปพร้อมกันเสมอ
วิธีการขุดค้นข้อมูลจากข้อความ
ข้อมูลที่เปิดเผยต่อสาธารณะจากเอกสารทางการแพทย์สามารถเข้าถึงได้ง่ายผ่านทางอินเทอร์เน็ต และกำลังกลายเป็นแหล่งข้อมูลที่มีประสิทธิภาพสำหรับการรวบรวมปฏิสัมพันธ์ระหว่างโปรตีน (PPIs) ที่รู้จัก การทำนาย PPI และการเชื่อมต่อโปรตีน การขุดค้นข้อความมีต้นทุนและใช้เวลาน้อยกว่าเมื่อเทียบกับเทคนิคที่มีปริมาณงานสูงอื่นๆ ปัจจุบัน วิธีการขุดค้นข้อความโดยทั่วไปจะตรวจจับความสัมพันธ์แบบไบนารีระหว่างโปรตีนที่โต้ตอบกันจากประโยคแต่ละประโยคโดยใช้การสกัดข้อมูลตามกฎ/รูปแบบและวิธีการเรียนรู้ของเครื่อง[ 52 ]มีแอปพลิเคชันการขุดค้นข้อความที่หลากหลายสำหรับการสกัดและ/หรือการทำนาย PPI ที่เปิดให้ใช้งานสาธารณะ รวมถึงคลังข้อมูลที่มักจะจัดเก็บ PPI ที่ได้รับการตรวจสอบด้วยตนเองและ/หรือทำนายโดยคอมพิวเตอร์ การขุดค้นข้อความสามารถดำเนินการได้สองขั้นตอน: การดึงข้อมูลซึ่งจะดึงข้อความที่มีชื่อของโปรตีนที่โต้ตอบกันอย่างใดอย่างหนึ่งหรือทั้งสองอย่าง และการสกัดข้อมูลซึ่งจะสกัดข้อมูลเป้าหมาย (โปรตีนที่โต้ตอบกัน สารตกค้างที่เกี่ยวข้อง ประเภทการโต้ตอบ ฯลฯ)
นอกจากนี้ยังมีการศึกษาวิจัยที่ใช้การสร้างโปรไฟล์ทางวิวัฒนาการโดยอาศัยทฤษฎีที่ว่าโปรตีนที่เกี่ยวข้องในวิถีร่วมกันจะวิวัฒนาการร่วมกันในลักษณะที่สัมพันธ์กันในสายพันธุ์ต่างๆ วิธีการขุดค้นข้อมูลข้อความที่ซับซ้อนกว่าบางวิธีใช้ เทคนิค การประมวลผลภาษาธรรมชาติ (NLP) ขั้นสูงและสร้างเครือข่ายความรู้ (ตัวอย่างเช่น การพิจารณาชื่อยีนเป็นโหนดและคำกริยาเป็นขอบ) การพัฒนาอื่นๆ เกี่ยวข้องกับวิธีการเคอร์เนลเพื่อทำนายปฏิสัมพันธ์ของโปรตีน[ 53 ]
วิธีการเรียนรู้ของเครื่องจักร

มีการเสนอและทบทวนวิธีการคำนวณหลายวิธีสำหรับการทำนายปฏิสัมพันธ์ระหว่างโปรตีน[ 54 ] [ 55 ] [ 56 ]แนวทางการทำนายสามารถจัดกลุ่มเป็นหมวดหมู่ตามหลักฐานการทำนายได้ เช่น ลำดับโปรตีนจีโนมิกส์เชิงเปรียบเทียบโดเมนโปรตีน โครงสร้างตติยภูมิของโปรตีน และโทโพโลยีเครือข่ายปฏิสัมพันธ์[ 54 ]การสร้างเซตบวก (คู่โปรตีนที่ทราบว่ามีปฏิสัมพันธ์กัน) และเซตลบ (คู่โปรตีนที่ไม่มีปฏิสัมพันธ์กัน) เป็นสิ่งจำเป็นสำหรับการพัฒนาแบบจำลองการทำนายด้วยคอมพิวเตอร์[ 55 ]แบบจำลองการทำนายโดยใช้เทคนิคการเรียนรู้ของเครื่องสามารถจำแนกได้เป็นสองกลุ่มหลัก ได้แก่ แบบมีผู้กำกับดูแลและแบบไม่มีผู้กำกับดูแล โดยพิจารณาจากการติดป้ายกำกับตัวแปรอินพุตตามผลลัพธ์ที่คาดหวัง[ 56 ]
ในปี 2548 โปรตีนเมมเบรนแบบบูรณาการของSaccharomyces cerevisiaeได้รับการวิเคราะห์โดยใช้ระบบยูบิควิตินแบบอาศัยการผสมพันธุ์ (mbSUS) ระบบนี้ตรวจจับปฏิสัมพันธ์ของโปรตีนเมมเบรนกับโปรตีนส่งสัญญาณภายนอกเซลล์[ 57 ]จากโปรตีนเมมเบรนแบบบูรณาการ 705 ชนิด มีการติดตามปฏิสัมพันธ์ที่แตกต่างกัน 1,985 รายการ ซึ่งเกี่ยวข้องกับโปรตีน 536 ชนิด ในการจัดเรียงและจำแนกปฏิสัมพันธ์นั้น ได้ใช้เครื่องสนับสนุนเวกเตอร์ (support vector machine) เพื่อกำหนดปฏิสัมพันธ์ที่มีความน่าเชื่อถือสูง ปานกลาง และต่ำ ระบบยีสต์ทูไฮบริดเมมเบรนแบบแยกยูบิควิติน (split-ubiquitin membrane yeast two-hybrid system) ใช้ตัวรายงานการถอดรหัสเพื่อระบุทรานส์ฟอร์แมนต์ของยีสต์ที่เข้ารหัสคู่ของโปรตีนที่มีปฏิสัมพันธ์กัน[ 58 ] ในปี 2549 พบว่า ป่าสุ่ม (random forest)ซึ่งเป็นตัวอย่างของเทคนิคแบบมีผู้กำกับดูแล เป็นวิธีการเรียนรู้ของเครื่องที่มีประสิทธิภาพมากที่สุดสำหรับการทำนายปฏิสัมพันธ์ของโปรตีน[ 59 ]วิธีการดังกล่าวถูกนำมาใช้เพื่อค้นหาปฏิสัมพันธ์ของโปรตีนบนอินเทอร์แอคโทมของมนุษย์ โดยเฉพาะอินเทอร์แอคโทมของโปรตีนเยื่อหุ้มเซลล์[ 60 ]และอินเทอร์แอคโทมของโปรตีนที่เกี่ยวข้องกับโรคจิตเภท[ 61 ]
ณ ปี 2020 โมเดลที่ใช้คลาสคลัสเตอร์ตกค้าง (RCCs) ซึ่งสร้างขึ้นจาก ฐานข้อมูล 3DIDและ Negatome ส่งผลให้สามารถจำแนกกรณีปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีนได้อย่างถูกต้อง 96-99% [ 62 ] RCCs เป็นพื้นที่เวกเตอร์เชิงคำนวณที่จำลองพื้นที่พับของโปรตีนและรวมชุดตกค้างที่สัมผัสกันพร้อมกันทั้งหมด ซึ่งสามารถใช้ในการวิเคราะห์ความสัมพันธ์ระหว่างโครงสร้างและหน้าที่ของโปรตีนและวิวัฒนาการ[ 63 ]
ฐานข้อมูล
การระบุ PPIs ในวงกว้างทำให้เกิดปฏิสัมพันธ์นับแสนรายการ ซึ่งถูกรวบรวมไว้ในฐานข้อมูลทางชีววิทยา เฉพาะทาง ที่ได้รับการอัปเดตอย่างต่อเนื่องเพื่อให้ได้อินแท็ก โทมที่สมบูรณ์ ฐานข้อมูลแรกเหล่านี้คือฐานข้อมูลโปรตีนที่มีปฏิสัมพันธ์ (DIP ) [ 64 ]
ฐานข้อมูลหลักรวบรวมข้อมูลเกี่ยวกับการปฏิสัมพันธ์ระหว่างโปรตีนที่ได้รับการตีพิมพ์และพิสูจน์แล้วว่ามีอยู่จริงผ่านวิธีการทดลองขนาดเล็กหรือขนาดใหญ่ ตัวอย่างเช่นDIP ( Biomolecular Interaction Network Database ), BIND (Biomolecular Interaction Network Database), BioGRID (Biological General Repository for Interaction Datasets), HPRD (Human Protein Reference Database), IntAct (IntAct Molecular Interaction Database), MINT (Molecular Interactions Database), MIPS-MPact (MIPS Protein Interaction Resource on Yeast) และ MIPS-MPPI (MIPS Mammalian Protein–Protein Interaction Database)
โดยปกติแล้ว เมตาฐานข้อมูลเกิดจากการบูรณาการข้อมูลจากฐานข้อมูลหลัก แต่ก็อาจรวบรวมข้อมูลดั้งเดิมบางส่วนไว้ด้วยเช่นกัน
ฐานข้อมูลการทำนายประกอบด้วย PPIs จำนวนมากที่ทำนายโดยใช้เทคนิคหลายอย่าง (บทความหลัก) ตัวอย่างเช่น ฐานข้อมูลการทำนายปฏิสัมพันธ์ระหว่างโปรตีนของมนุษย์ (PIPs) [ 65 ]ฐานข้อมูลปฏิสัมพันธ์ระหว่างโปรตีน (I2D) ปฏิสัมพันธ์ระหว่างโปรตีนที่ทราบและทำนายได้ (STRING-db)และ Unified Human Interactive (UniHI)
วิธีการคำนวณที่กล่าวมาข้างต้นทั้งหมดขึ้นอยู่กับฐานข้อมูลต้นทางซึ่งสามารถคาดการณ์ข้อมูลเพื่อทำนายปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีนใหม่ได้ความครอบคลุมแตกต่างกันอย่างมากระหว่างฐานข้อมูล โดยทั่วไป ฐานข้อมูลหลักจะมีจำนวนปฏิสัมพันธ์ของโปรตีนทั้งหมดที่บันทึกไว้น้อยที่สุด เนื่องจากไม่ได้รวมข้อมูลจากฐานข้อมูลอื่นๆ หลายฐานข้อมูล ในขณะที่ฐานข้อมูลการทำนายจะมีจำนวนมากที่สุด เพราะรวมหลักฐานรูปแบบอื่นๆ นอกเหนือจากการทดลอง ตัวอย่างเช่น ฐานข้อมูลหลัก IntAct มีปฏิสัมพันธ์ 572,063 รายการ[ 66 ]ฐานข้อมูลเมตา APID มีปฏิสัมพันธ์ 678,000 รายการ[ 67 ]และฐานข้อมูลการทำนาย STRING มีปฏิสัมพันธ์ 25,914,693 รายการ[ 68 ]อย่างไรก็ตาม ปฏิสัมพันธ์บางส่วนในฐานข้อมูล STRING นั้นได้รับการทำนายโดยวิธีการคำนวณเท่านั้น เช่น Genomic Context และยังไม่ได้รับการตรวจสอบโดยการทดลอง
เครือข่ายปฏิสัมพันธ์

ข้อมูลที่พบในฐานข้อมูล PPIs สนับสนุนการสร้างเครือข่ายปฏิสัมพันธ์ แม้ว่าเครือข่าย PPI ของโปรตีนคำถามที่กำหนดจะสามารถแสดงในตำราเรียนได้ แต่แผนภาพของ PPIs ของเซลล์ทั้งหมดนั้นค่อนข้างซับซ้อนและสร้างได้ยาก[ 69 ]
ตัวอย่างหนึ่งของแผนที่ปฏิสัมพันธ์ระดับโมเลกุลที่สร้างขึ้นด้วยตนเองคือแผนที่การควบคุมวงจรเซลล์ของ Kurt Kohn ในปี 1999 [ 70 ]โดยอ้างอิงจากแผนที่ของ Kohn นั้น Schwikowski และคณะได้ตีพิมพ์บทความเกี่ยวกับ PPIs ในยีสต์ในปี 2000 ซึ่งเชื่อมโยงโปรตีนที่โต้ตอบกัน 1,548 ตัวที่กำหนดโดยการคัดกรองแบบทูไฮบริด พวกเขาใช้วิธีการวาดกราฟแบบหลายชั้นเพื่อหาตำแหน่งเริ่มต้นของโหนด จากนั้นจึงปรับปรุงเค้าโครงโดยใช้อัลกอริทึมแบบอิงแรง[ 71 ]
เครื่องมือทางชีวสารสนเทศได้รับการพัฒนาขึ้นเพื่อลดความซับซ้อนของงานที่ยากลำบากในการแสดงภาพเครือข่ายปฏิสัมพันธ์ระดับโมเลกุลและเสริมด้วยข้อมูลประเภทอื่น ตัวอย่างเช่นCytoscapeเป็นซอฟต์แวร์โอเพนซอร์สที่ใช้กันอย่างแพร่หลายและมีปลั๊กอินมากมายให้ใช้งานในปัจจุบัน[ 72 ]ซอฟต์แวร์ Pajek มีข้อดีสำหรับการแสดงภาพและการวิเคราะห์เครือข่ายขนาดใหญ่มาก[ 51 ]
การระบุโมดูลการทำงานในเครือข่าย PPI เป็นความท้าทายที่สำคัญในชีวสารสนเทศ โมดูลการทำงานหมายถึงชุดของโปรตีนที่เชื่อมต่อกันอย่างมากในเครือข่าย PPI ซึ่งเป็นปัญหาที่คล้ายคลึงกับการตรวจจับชุมชนในเครือข่ายสังคมมีวิธีการบางอย่าง เช่น โมดูล Jactive [ 73 ]และ MoBaS [ 74 ]โมดูล Jactive ผสานรวมเครือข่าย PPI และ ข้อมูล การแสดงออกของยีนในขณะที่ MoBaS ผสานรวมเครือข่าย PPI และการศึกษาการเชื่อมโยงทั่วทั้งจีโนม
ความสัมพันธ์ระหว่างโปรตีนมักเป็นผลมาจากปฏิสัมพันธ์หลายประเภทหรืออนุมานจากแนวทางที่แตกต่างกัน รวมถึงการอยู่ร่วมกันในตำแหน่งเดียวกัน ปฏิสัมพันธ์โดยตรง ปฏิสัมพันธ์ทางพันธุกรรมแบบยับยั้ง ปฏิสัมพันธ์ทางพันธุกรรมแบบเสริม การเชื่อมโยงทางกายภาพ และการเชื่อมโยงอื่นๆ[ 75 ]
เครือข่ายปฏิสัมพันธ์ที่ลงนามแล้ว

ปฏิสัมพันธ์ระหว่างโปรตีนมักส่งผลให้โปรตีนที่ทำปฏิกิริยาตัวใดตัวหนึ่งถูก 'กระตุ้น' หรือ 'ยับยั้ง' ผลกระทบดังกล่าวสามารถระบุได้ในเครือข่าย PPI โดยใช้ "สัญญาณ" (เช่น "การกระตุ้น" หรือ "การยับยั้ง") แม้ว่าคุณลักษณะดังกล่าวจะถูกเพิ่มเข้าไปในเครือข่ายมานานแล้ว[ 77 ] Vinayagam et al. (2014) ก็ได้บัญญัติศัพท์ว่าเครือข่ายที่มีสัญญาณ (Signed network ) สำหรับเครือข่ายเหล่านี้ เครือข่ายที่มีสัญญาณมักแสดงโดยการติดป้ายกำกับปฏิสัมพันธ์ว่าเป็นบวกหรือลบ ปฏิสัมพันธ์ที่เป็นบวกคือปฏิสัมพันธ์ที่ส่งผลให้โปรตีนตัวใดตัวหนึ่งถูกกระตุ้น ในทางกลับกัน ปฏิสัมพันธ์ที่เป็นลบแสดงว่าโปรตีนตัวใดตัวหนึ่งถูกยับยั้ง[ 78 ]
เครือข่ายปฏิสัมพันธ์ระหว่างโปรตีนมักถูกสร้างขึ้นจากการทดลองในห้องปฏิบัติการ เช่น การคัดกรองยีสต์ทูไฮบริดหรือ 'การทำให้บริสุทธิ์ด้วยความสัมพันธ์และเทคนิคสเปกโทรเมตรีมวลในภายหลัง' [ 79 ]อย่างไรก็ตาม วิธีการเหล่านี้ไม่ได้ให้ข้อมูลที่จำเป็นในการกำหนดประเภทของปฏิสัมพันธ์ที่มีอยู่เพื่อให้สามารถระบุเครื่องหมายให้กับแผนภาพเครือข่ายได้
การคัดกรองการรบกวน RNA
การคัดกรองด้วย RNA interference (RNAi) (การยับยั้งโปรตีนแต่ละตัวระหว่างการถอดรหัสและการแปลรหัส) เป็นวิธีการหนึ่งที่สามารถนำมาใช้ในการค้นหาสัญญาณของการโต้ตอบระหว่างโปรตีน โดยจะทำการยับยั้งโปรตีนแต่ละตัวและวิเคราะห์ฟีโนไทป์ที่เกิดขึ้น ความสัมพันธ์ของฟีโนไทป์ที่สอดคล้องกัน (เช่น การยับยั้งโปรตีนตัวใดตัวหนึ่งในสองตัวส่งผลให้เกิดฟีโนไทป์เดียวกัน) บ่งชี้ถึงความสัมพันธ์เชิงบวกหรือการกระตุ้น ส่วนฟีโนไทป์ที่ไม่สอดคล้องกัน (เช่น การยับยั้งโปรตีนตัวใดตัวหนึ่งในสองตัวส่งผลให้เกิดฟีโนไทป์ที่แตกต่างกันสองแบบ) บ่งชี้ถึงความสัมพันธ์เชิงลบหรือการยับยั้ง หากโปรตีน A ขึ้นอยู่กับโปรตีน B ในการกระตุ้น การยับยั้งโปรตีน A หรือ B จะส่งผลให้เซลล์สูญเสียบริการที่ได้รับจากโปรตีน A และฟีโนไทป์จะเหมือนกันไม่ว่าจะเป็นการยับยั้ง A หรือ B ก็ตาม อย่างไรก็ตาม หากโปรตีน A ถูกยับยั้งโดยโปรตีน B ฟีโนไทป์จะแตกต่างกันไปขึ้นอยู่กับว่าโปรตีนใดถูกยับยั้ง (หากยับยั้งโปรตีน B โปรตีน B จะไม่สามารถยับยั้งโปรตีน A ได้อีกต่อไป ทำให้ A ยังคงทำงานอยู่ แต่หากยับยั้ง A โปรตีน B จะไม่มีอะไรให้กระตุ้นเนื่องจาก A ไม่ทำงาน และฟีโนไทป์จะเปลี่ยนไป) จำเป็นต้องทำการคัดกรอง RNAi หลายครั้ง เพื่อให้สามารถระบุสัญญาณของการโต้ตอบระหว่างโปรตีนกับโปรตีนได้อย่างน่าเชื่อถือ Vinayagam และคณะ ผู้คิดค้นเทคนิคนี้ระบุว่า จำเป็นต้องทำการคัดกรอง RNAi อย่างน้อยเก้าครั้ง โดยความมั่นใจจะเพิ่มขึ้นเมื่อทำการคัดกรองมากขึ้น[ 78 ]
ในฐานะเป้าหมายในการรักษา
การปรับเปลี่ยน PPI เป็นเรื่องท้าทายและได้รับความสนใจเพิ่มขึ้นจากชุมชนวิทยาศาสตร์[ 80 ]คุณสมบัติหลายประการของ PPI เช่น ตำแหน่งอัลโลสเตอริกและฮอตสปอต ได้ถูกนำมาใช้ในกลยุทธ์การออกแบบยา[ 81 ] [ 82 ]อย่างไรก็ตาม มี PPI เพียงไม่กี่ตัวเท่านั้นที่ถูกกำหนดเป้าหมายโดยตรงโดยสาร ยับยั้ง PPI โมเลกุลขนาดเล็กที่ได้รับการอนุมัติจาก FDAซึ่งเน้นย้ำถึงโอกาสอันมหาศาลที่ยังไม่ได้ใช้ประโยชน์สำหรับการค้นพบยา
ในปี 2014 Amit Jaiswal และคณะได้พัฒนาเปปไทด์ 30 ชนิดเพื่อยับยั้งการดึงดูดเทโลเมอเรสไปยังเทโลเมียร์โดยใช้การศึกษาปฏิสัมพันธ์ระหว่างโปรตีน[ 83 ] [ 84 ] Arkinและคณะได้พัฒนาสารยับยั้งที่ใช้ชิ้นส่วนแอนติบอดีเพื่อควบคุมปฏิสัมพันธ์ระหว่างโปรตีนที่เฉพาะเจาะจง[ 85 ]
เนื่องจากการ "ปรับเปลี่ยน" ของ PPIs ไม่เพียงแต่รวมถึงการยับยั้ง แต่ยังรวมถึงการทำให้โปรตีนเชิงซ้อนควอเทอร์นารี มีเสถียรภาพ ซึ่งเป็นแบบแผนที่นำเสนอโดยการค้นพบการก่อตัวของโปรตีนเชิงซ้อนที่เหนี่ยวนำโดยออกซิน โดย Ning Zheng [ 86 ] [ 87 ]โมเลกุลที่มีกลไกการทำงานนี้ (ที่เรียกว่ากาวโมเลกุล ) จึงได้รับการศึกษาอย่างกว้างขวางเช่นกัน[ 88 ]
ตัวอย่าง
สารยับยั้งปฏิกิริยาระหว่างโปรตีนที่ได้รับการพัฒนาเป็นยาและวางจำหน่ายในตลาด ได้แก่:
- ไทโรฟิบันซึ่งเป็นสารยับยั้งไกลโคโปรตีน IIb/IIIa ใช้เป็นยาสำหรับโรคหัวใจและหลอดเลือด[ 89 ]
- Maravirocซึ่งเป็นสารยับยั้งการโต้ตอบระหว่าง CCR5-gp120 ใช้เป็นยาต้านไวรัสเอชไอวี[ 90 ]
- เวเนโตแคล็กซ์[ 91 ]
- ไลฟิเทกราสต์[ 92 ]
- โซโตราซิบบ์[ 93 ]
- อะดากราซิบบ์[ 93 ]
สารยับยั้งที่อยู่ระหว่างการทดลองในการพัฒนายา ได้แก่:
- AMG-176, AZD5991, S64315 สารยับยั้ง โปรตีน เม็ดเลือดขาวชนิดไมอีลอยด์ 1 (Mcl-1) และปฏิกิริยาของมัน[ 94 ]
ดูเพิ่มเติม
- ปฏิสัมพันธ์ระหว่างไกลแคนและโปรตีน
- 3did
- อัลโลสเตอรี
- เครือข่ายชีวภาพ
- เครื่องจักรชีวภาพ
- ดีมา (ฐานข้อมูล)
- การเร่งปฏิกิริยาโดยเอนไซม์
- ฮิตพรีดิค
- ปฏิสัมพันธ์ของมนุษย์
- ไอโซเบส
- คอมเพล็กซ์โปรตีนหลายชนิด
- พลวัตของโดเมนโปรตีน
- ความยืดหยุ่นของโปรตีน
- โครงสร้างโปรตีน
- การทำนายปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีน
- การคัดกรองปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีน
- ชีววิทยาระบบ
อ่านเพิ่มเติม
- Stark C, Breitkreutz BJ, Reguly T, Boucher L, Breitkreutz A, Tyers M (มกราคม 2549). "BioGRID: คลังข้อมูลทั่วไปสำหรับชุดข้อมูลปฏิสัมพันธ์" . Nucleic Acids Research . 34 (ฉบับฐานข้อมูล): D535– D539. doi : 10.1093/nar/gkj109 . PMC 1347471 . PMID 16381927 .
- Peri S, Navarro JD, Kristiansen TZ, Amanchy R, Surendranath V, Muthusamy B และคณะ (มกราคม 2547). "ฐานข้อมูลอ้างอิงโปรตีนของมนุษย์เป็นแหล่งข้อมูลสำหรับการค้นพบในด้านโปรตีโอมิกส์" . Nucleic Acids Research . 32 (ฉบับฐานข้อมูล): D497– D501. doi : 10.1093/nar/gkh070 . PMC 308804 . PMID 14681466 .
- Hermjakob H, Montecchi-Palazzi L, Lewington C, Mudali S, Kerrien S, Orchard S, และคณะ (มกราคม 2547). "IntAct: ฐานข้อมูลปฏิสัมพันธ์โมเลกุลแบบโอเพ่นซอร์ส " การวิจัยกรดนิวคลีอิก32 (ปัญหาฐานข้อมูล): D452– D455 ดอย : 10.1093/nar/gkh052 . PMC 308786 . PMID 14681455 .
- Chatr-aryamontri A, Ceol A, Palazzi LM, Nardelli G, Schneider MV, Castagnoli L และคณะ (มกราคม 2550). "MINT: ฐานข้อมูลปฏิสัมพันธ์ระดับโมเลกุล" . Nucleic Acids Research . 35 (ฉบับฐานข้อมูล): D572– D574. doi : 10.1093/nar/gkl950 . PMC 1751541 . PMID 17135203 .
- Güldener U, Münsterkötter M, Oesterheld M, Pagel P, Ruepp A, Mewes HW และคณะ (มกราคม 2549). "MPact: แหล่งข้อมูลปฏิสัมพันธ์โปรตีน MIPS บนยีสต์" . Nucleic Acids Research . 34 (ฉบับฐานข้อมูล): D436– D441. doi : 10.1093/nar/gkj003 . PMC 1347366 . PMID 16381906 .
- Pagel P, Kovac S, Oesterheld M, Brauner B, Dunger-Kaltenbach I, Frishman G และคณะ (มีนาคม 2548). "ฐานข้อมูลปฏิสัมพันธ์ระหว่างโปรตีนของสัตว์เลี้ยงลูกด้วยนม MIPS" . Bioinformatics . 21 (6): 832– 834. doi : 10.1093/bioinformatics/bti115 . PMID 15531608 .
- Casado-Vela J, Matthiesen R, Sellés S, Naranjo JR (พฤษภาคม 2013). "ปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีน: ความซ้ำซ้อนของตัวย่อของยีนและข้อจำกัดในปัจจุบันที่ขัดขวางการบูรณาการข้อมูลอัตโนมัติ" . Proteomes . 1 (1): 3– 24. doi : 10.3390/proteomes1010003 . PMC 5314489 . PMID 28250396 .
- Robin V, Bodein A, Scott-Boyer MP, Leclercq M, Périn O, Droit A (2022). "ภาพรวมของวิธีการสำหรับการกำหนดลักษณะและแสดงภาพเครือข่ายปฏิสัมพันธ์ระหว่างโปรตีนในบริบทการบูรณาการหลายโอไมซ์" Frontiers in Molecular Biosciences 9 962799. doi : 10.3389 /fmolb.2022.962799 . PMC 494275 . PMID 36158572 .
ลิงก์ภายนอก
- ฐานข้อมูลปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีน
- คลังข้อมูลของตัวปรับแต่งปฏิสัมพันธ์ระหว่างโปรตีน (PPI)