อ่าน 12 นาที
เรตินา
เรตินัล (หรือที่รู้จักกันในชื่อ เรตินัลดีไฮด์ ) เป็น โครโมฟอร์โพ ลีอีน เรตินัลซึ่งจับกับโปรตีนที่เรียกว่า ออปซิน เป็นพื้นฐานทางเคมีของ การแปลงสัญญาณแสงในการมอง เห็น...
เรตินา
 | |
 | |
| ชื่อ | |
|---|---|
| ชื่อ IUPAC เรตินา | |
| ชื่อตามระบบ IUPAC (2 E ,4 E ,6 E ,8 E )-3,7-ไดเมทิล-9-(2,6,6-ไตรเมทิลไซโคลเฮกซ์-1-en-1-อิล)โนนา-2,4,6,8-เตตระอีนัล | |
ชื่ออื่นๆ
| |
| ตัวระบุ | |
| |
โมเดล 3 มิติ ( JSmol ) |
|
| ชอีบี |
|
| เคมสไปเดอร์ |
|
| บัตรข้อมูล ECHA | 100.003.760 |
PubChem CID |
|
| มหาวิทยาลัย |
|
แดชบอร์ด CompTox ( EPA ) |
|
| |
| |
| คุณสมบัติ | |
| C 20 H 28 O | |
| มวลโมลาร์ | 284.443 กรัม·โมล−1 |
| รูปร่าง | ผลึกสีส้มจากปิโตรเลียมอีเทอร์[ 1 ] |
| จุดหลอมเหลว | 61 ถึง 64 °C (142 ถึง 147 °F; 334 ถึง 337 K) [ 1 ] |
| แทบไม่ละลาย | |
| ความสามารถในการละลายในไขมัน | ละลายได้ |
| สารประกอบที่เกี่ยวข้อง | |
สารประกอบที่เกี่ยวข้อง | เรตินอล ; กรดเรติโนอิก ; เบต้าแคโรทีน ; ดี ไฮโดรเรตินัล ; 3-ไฮดรอกซีเรตินัล; 4-ไฮดรอกซีเรตินัล |
เว้นแต่จะระบุไว้เป็นอย่างอื่น ข้อมูลที่ให้ไว้เป็นข้อมูลสำหรับวัสดุในสภาวะมาตรฐาน (ที่อุณหภูมิ 25 °C [77 °F] ความดัน 100 kPa) ข้อมูลอ้างอิงในกล่องข้อมูล | |
เรตินัล (หรือที่รู้จักกันในชื่อเรตินัลดีไฮด์ ) เป็นโครโมฟอร์โพลีอีน เรตินัลซึ่งจับกับโปรตีนที่เรียกว่าออปซินเป็นพื้นฐานทางเคมีของการแปลงสัญญาณแสงในการมอง เห็น ซึ่งเป็นขั้นตอนการตรวจจับแสงในกระบวนการรับรู้ทางสายตา (การมองเห็น)
จุลินทรีย์บางชนิดใช้เรตินัลในการเปลี่ยนแสงให้เป็นพลังงานเมตาบอลิซึม การศึกษาหนึ่งชี้ให้เห็นว่าเมื่อประมาณสามพันล้านปีก่อน สิ่งมีชีวิตส่วนใหญ่บนโลกใช้เรตินัลแทนคลอโรฟิลล์ในการเปลี่ยนแสงแดดให้เป็นพลังงาน เนื่องจากเรตินัลดูดซับแสงสีเขียวเป็นส่วนใหญ่และส่งผ่านแสงสีม่วง จึงทำให้เกิดสมมติฐานโลกสีม่วงขึ้น[ 2 ]
เรตินัลนั้นถือเป็นวิตามินเอ ชนิดหนึ่ง เมื่อสัตว์กินเข้าไป มีวิตามินเอหลายรูปแบบ ซึ่งทั้งหมดจะถูกแปลงเป็นเรตินัล ซึ่งไม่สามารถสร้างขึ้นได้หากปราศจากวิตามินเอ จำนวนโมเลกุลที่แตกต่างกันที่สามารถแปลงเป็นเรตินัลนั้นแตกต่างกันไปในแต่ละสายพันธุ์ เดิมทีเรตินัลเรียกว่าเรตินีน[ 3 ] และได้รับการเปลี่ยนชื่อ[ 4 ]หลังจากที่ค้นพบว่าเป็นวิตามินเออัลดีไฮด์[ 5 ] [ 6 ]
สัตว์ มีกระดูกสันหลังได้รับเรตินัลโดยตรงจากเนื้อสัตว์ หรือพวกมันสร้างเรตินัลจากแคโรทีนอยด์ ไม่ว่าจะเป็นจากอัลฟาแคโรทีนหรือเบตาแคโรทีน ซึ่งทั้งสองอย่างเป็นแคโรทีนพวกมันยังสร้างเรตินัลจากเบตาคริปโทแซนทิน ซึ่งเป็น แซนโทฟิลล์ชนิด หนึ่ง ได้อีกด้วย แคโรทีนอยด์เหล่านี้ต้องได้รับจากพืชหรือ สิ่งมีชีวิต ที่สังเคราะห์แสงได้ สัตว์ไม่สามารถเปลี่ยนแคโรทีนอยด์ชนิดอื่นให้เป็นเรตินัลได้ สัตว์กินเนื้อบางชนิดไม่สามารถเปลี่ยนแคโรทีนอยด์ใดๆ ได้เลย วิตามินเอรูปแบบหลักอื่นๆ ได้แก่เรตินอลและกรดเรติโนอิก ซึ่งเป็นรูปแบบที่ออกฤทธิ์ได้บางส่วน อาจสร้างขึ้นจากเรตินัลได้เช่นกัน
สัตว์ไม่มีกระดูกสันหลังเช่นแมลงและปลาหมึกใช้เรตินัลในรูปแบบไฮดรอกซิเลตในระบบการมองเห็น ซึ่งได้มาจากการแปลงสภาพจากแซนโทฟิลล์ชนิด อื่น
การเผาผลาญวิตามินเอ
สิ่งมีชีวิตสร้างเรตินัลโดยการแตกตัวออกซิเดชันที่ไม่สามารถย้อนกลับได้ของแคโรทีนอยด์[ 7 ]
ตัวอย่างเช่น:
เร่งปฏิกิริยาโดยเบต้า-แคโรทีน 15,15'-โมโนออกซิเจเนส[ 8 ]หรือเบต้า-แคโรทีน 15,15'-ไดออกซิเจเนส[ 9 ]
เช่นเดียวกับที่แคโรทีนอยด์เป็นสารตั้งต้นของเรตินัล เรตินัลก็เป็นสารตั้งต้นของวิตามินเอในรูปแบบอื่นๆ เรตินัลสามารถเปลี่ยนรูปไปเป็นเรตินอลซึ่งเป็นรูปแบบการขนส่งและการเก็บรักษาวิตามินเอได้
เร่งปฏิกิริยาโดยเรตินอลดีไฮโดรจีเนส (RDHs) [ 10 ]และแอลกอฮอล์ดีไฮโดรจีเนส (ADHs) [ 11 ]
เรตินอลมีชื่อเรียกอีกอย่างว่า วิตามินเอแอลกอฮอล์หรือเรียกสั้นๆ ว่า วิตามินเอ นอกจากนี้ เรตินัลยังสามารถถูกออกซิไดซ์เป็นกรดเรติโนอิกได้ อีกด้วย
เร่งปฏิกิริยาโดยเรตินัลดีไฮโดรจีเนส[ 12 ]หรือที่รู้จักกันในชื่อเรตินัลดีไฮด์ดีไฮโดรจีเนส (RALDHs) [ 11 ]รวมถึงเรตินัลออกซิเดส[ 13 ]
กรดเรติโนอิก หรือบางครั้งเรียกว่า กรดวิตามินเอเป็นโมเลกุลส่งสัญญาณและฮอร์โมนที่สำคัญในสัตว์มีกระดูกสันหลัง
วิสัยทัศน์
เรตินัลเป็นโครโมฟอร์แบบคอนจูเกตในดวงตาของสัตว์มีกระดูกสันหลังเรตินัลเริ่มต้นในโครงสร้าง 11- ซิส -เรตินัล ซึ่งเมื่อจับโฟตอนที่มีความยาวคลื่นที่ถูกต้องได้ ก็จะยืดออกเป็นโครงสร้างออล- ทรานส์ -เรตินัล การเปลี่ยนแปลงโครงสร้างนี้จะไปกดกับโปรตีนออปซินในเรตินาทำให้เกิดการส่งสัญญาณทางเคมีเป็นลูกโซ่ ส่งผลให้ สมอง รับรู้แสงหรือภาพได้ สเปกตรัมการดูดกลืนแสงของโครโมฟอร์ขึ้นอยู่กับปฏิกิริยาของมันกับโปรตีนออปซินที่มันจับอยู่ ดังนั้นสารประกอบเรตินัล-ออปซินที่แตกต่างกันจะดูดกลืนโฟตอนที่มีความยาวคลื่นต่างกัน (เช่น สีของแสงที่แตกต่างกัน)
ออปซินส์

เรตินัลจับกับออปซินซึ่งเป็นตัวรับที่เชื่อมต่อกับโปรตีน G (GPCRs) [ 14 ] [ 15 ]ออปซิน เช่นเดียวกับ GPCRs อื่นๆ มีอัลฟาเฮลิกซ์แบบทรานส์เมมเบรน 7 อัน ที่เชื่อมต่อกันด้วยลูป 6 อัน พบได้ในเซลล์รับแสงในเรตินา ของดวงตา ออปซินใน เซลล์แท่งของสัตว์มี กระดูกสันหลัง คือโรดอปซิน เซลล์แท่งสร้างเป็นดิสก์ซึ่งมีโมเลกุลโรดอปซินอยู่ในเยื่อหุ้มเซลล์และอยู่ภายในเซลล์ทั้งหมด ส่วนหัว N-terminusของโมเลกุลยื่นเข้าไปภายในดิสก์ และ ส่วนหาง C-terminusยื่นเข้าไปในไซโตพลาสซึมของเซลล์ ออปซินในเซลล์กรวยคือOPN1SW , OPN1MWและOPN1LWเซลล์กรวยสร้างเป็นดิสก์ที่ไม่สมบูรณ์ซึ่งเป็นส่วนหนึ่งของเยื่อหุ้มพลาสมาดังนั้นส่วนหัว N-terminus จึงยื่นออกไปนอกเซลล์ ในออปซิน เรตินัลจะจับกับไลซีนแบบโควาเลนต์[ 16 ]ในเกลียวทรานส์เมมเบรนที่เจ็ด[ 17 ] [ 18 ] [ 19 ]ผ่านเบสชิฟฟ์ [ 20 ] [ 21 ] การสร้างพันธะเบสชิฟฟ์เกี่ยวข้องกับการกำจัดอะตอมออกซิเจนออกจากเรตินัลและอะตอมไฮโดรเจนสองอะตอมจากหมู่เอมีโนอิสระของไลซีน ทำให้เกิด H 2 O เรตินิลิดีนคือหมู่สองวาเลนต์ที่เกิดจากการกำจัดอะตอมออกซิเจนออกจากเรตินัล ดังนั้นออปซินจึงถูกเรียกว่าโปรตีนเรตินิลิดีน
ออปซินเป็นตัวรับที่เชื่อมโยงกับโปรตีน G (GPCR) ต้นแบบ [ 22 ]โรดอปซินของวัว ซึ่งเป็นออปซินของเซลล์แท่ง เป็น GPCR ตัวแรกที่มีลำดับกรดอะมิโน[ 23 ]และโครงสร้างสามมิติ (ผ่านการตกผลึกด้วยรังสีเอกซ์ ) ที่ได้รับการกำหนด[ 18 ] โรดอปซิน ของวัวประกอบด้วยกรดอะมิโน 348 ตัว เรตินัลจับเป็นโครโมฟอร์ที่ไลซีน296 [ 18 ] [ 23 ] ไลซีนนี้ได้รับการอนุรักษ์ไว้ในออปซินเกือบทั้งหมด มีเพียงออ ปซินบางส่วนเท่านั้นที่สูญเสียมันไปในระหว่างวิวัฒนาการ[ 24 ] ออ ปซินที่ไม่มีไลซีนที่จับกับเรตินัลจะไม่ไวต่อแสง[ 25 ] [ 26 ] [ 27 ]ออปซินดังกล่าวอาจมีหน้าที่อื่น[ 26 ] [ 24 ]
แม้ว่าสัตว์เลี้ยงลูกด้วยนมจะใช้เรตินัลเป็นโครโมฟอร์ออปซินเพียงอย่างเดียว แต่สัตว์กลุ่มอื่นๆ ยังใช้โครโมฟอร์อีกสี่ชนิดที่เกี่ยวข้องอย่างใกล้ชิดกับเรตินัล ได้แก่ 3,4-ไดดีไฮโดรเรตินัล (วิตามินเอ2 ), (3 R )-3-ไฮดรอกซีเรตินัล, (3 S )-3-ไฮดรอกซีเรตินัล (ทั้งสองเป็นวิตามินเอ3 ) และ (4 R )-4-ไฮดรอกซีเรตินัล (วิตามินเอ4 ) ปลาและสัตว์สะเทินน้ำสะเทินบกหลายชนิดใช้ 3,4-ไดดีไฮโดรเรตินัล หรือที่เรียกว่าดีไฮโดรเรตินัลยกเว้นแมลงวันในวงศ์ย่อยCyclorrhapha (ที่เรียกว่าแมลงวันชั้นสูง) แมลง ทั้งหมด ที่ได้รับการตรวจสอบใช้ ( R )- เอนันติโอเมอร์ของ 3-ไฮดรอกซีเรติ นัล ซึ่งคาดว่าจะพบได้หาก 3-ไฮด รอกซีเรตินัลถูกผลิตโดยตรงจาก แคโร ทีนอยด์แซนโทฟิลล์ ไซคลอร์ราแฟนส์ รวมถึงดรอโซฟิลาใช้ (3 S )-3-ไฮดรอกซีเรตินัล[ 28 ] [ 29 ] พบว่าปลาหมึกหิ่งห้อย ใช้ (4 R )-4-ไฮดรอกซีเรตินัล
วงจรภาพ
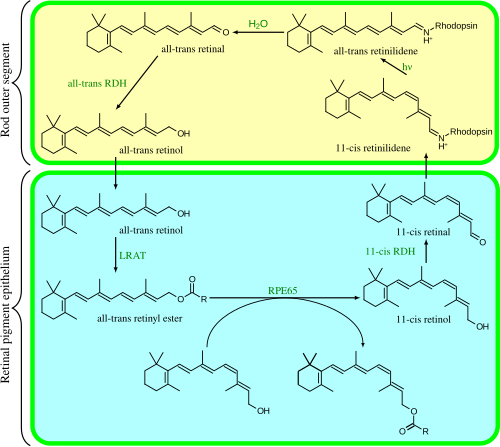
วงจรการมองเห็นเป็นเส้นทางเอนไซม์ แบบวงกลม ซึ่งเป็นส่วนหน้าของการแปลงสัญญาณแสง มันสร้าง 11- cis -retinal ขึ้นมาใหม่ ตัวอย่างเช่น วงจรการมองเห็นของเซลล์รูปแท่งในสัตว์เลี้ยงลูกด้วยนมมีดังนี้:
- all- trans- retinyl ester + H2O → 11- cis - retinol + กรดไขมัน ; RPE65 isomerohydrolases; [ 30 ]
- 11- ซิส -เรตินอล + NAD + → 11- ซิส -เรตินัล + NADH + H + ; 11- ซิส -เรตินอลดีไฮโดรจีเนส;
- 11- ซิส-เรตินัล + อะโพโรดอปซิน → โรดอปซิน + H2O ;เกิด พันธะ ชิฟฟ์เบสกับไลซีน -CH=N + H-;
- โรดอปซิน + hν → เมทาโรดอปซิน II (เช่น 11- ซิส โฟโตไอโซเมอไรเซชันเป็นออล- ทรานส์ ):
- (โรดอปซิน + hν → โฟโตโรดอปซิน → บาโธโรดอปซิน → ลูมิโรดอปซิน → เมทาโรดอปซิน I → เมทาโรดอปซิน II)
- เมทาโรดอปซิน II + H₂O →อะพอโรดอปซิน + อัลล์- ทรานส์ -เรตินัล;
- อัล-ทรานส์ -เรตินั ล + NADPH + H + → อัล- ทรานส์ -เรตินอล + NADP + ; อัล-ทรานส์ - เรตินอลดีไฮโดรจีเนส ;
- อัล-ทรานส์ -เรตินอล + กรดไขมัน → อัล-ทรานส์ -เรติ นิลเอสเทอร์ + H2O ;เลซิตินเรตินอลอะซิลทรานสเฟอเรส (LRATs) [ 31 ]
ขั้นตอนที่ 3, 4, 5 และ 6 เกิดขึ้นในส่วนนอกของเซลล์รูปแท่งส่วนขั้นตอนที่ 1, 2 และ 7 เกิดขึ้นใน เซลล์ เยื่อบุผิวเม็ดสีเรตินา (RPE)
ไอโซเมอโรไฮโดรเลส RPE65 มีความคล้ายคลึงกับเบตา-แคโรทีนโมโนออกซิเจเนส[ 7 ]เอนไซม์ ninaB ที่คล้ายคลึงกันในDrosophilaมีทั้งกิจกรรมแคโรทีนอยด์ออกซิเจเนสที่สร้างเรตินัลและกิจกรรมไอโซเมอเรส all- transถึง 11- cis [ 32 ]
โรดอปซินของจุลินทรีย์
ออล-ทราน ส์- เรตินัลยังเป็นส่วนประกอบสำคัญของ ออปซิน ของจุลินทรีย์เช่นแบคทีริโอโรดอปซินแช น เนลโรดอปซินและฮาโลโรดอปซินซึ่งมีความสำคัญในการสังเคราะห์แสงแบบไม่ใช้ออกซิเจน ของ แบคทีเรียและอาร์เคีย ในโมเลกุลเหล่านี้ แสงทำให้ออล-ท รานส์-เรติ นัลกลายเป็น 13- ซิส - เรตินัล จากนั้นจะวนกลับไปเป็นออล- ทรานส์-เรตินัลในสภาวะมืด โปรตีนเหล่านี้ไม่มีความสัมพันธ์ทางวิวัฒนาการกับออปซินของสัตว์และไม่ใช่ GPCR ข้อเท็จจริงที่ว่าทั้งสองใช้เรตินัลเป็นผลมาจาก วิวัฒนาการแบบ ลู่เข้า[ 33 ]
ประวัติศาสตร์
นักชีวเคมีชาวอเมริกันGeorge Waldและคนอื่นๆ ได้ร่างวงจรการมองเห็นไว้ตั้งแต่ปี 1958 จากผลงานของเขา Wald ได้รับรางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ร่วมกับHaldan Keffer HartlineและRagnar Granitใน ปี 1967 [ 34 ]
ดูเพิ่มเติม
อ่านเพิ่มเติม
- Fernald, Russell D. (2006). " การฉายแสงทางพันธุกรรมต่อวิวัฒนาการของดวงตา". Science . 313 (5795): 1914–1918 . Bibcode : 2006Sci...313.1914F . doi : 10.1126/science.1127889 . PMID 17008522. S2CID 84439732 .
- Amora, Tabitha L.; Ramos, Lavoisier S.; Galan, Jhenny F.; Birge, Robert R. (2008). "การปรับสเปกตรัมของเม็ดสีโคนสีแดงเข้ม" . ชีวเคมี . 47 (16): 4614– 20. doi : 10.1021/bi702069d . PMC 2492582 . PMID 18370404 .
- Barlow, HB; Levick, WR; Yoon, M. (1971). "การตอบสนองต่อควอนตัมแสงเดี่ยวในเซลล์แกงลีออนเรตินาของแมว" Vision Research . 11 (Supplement 3): 87– 101. doi : 10.1016/0042-6989(71)90033-2 . PMID 5293890 .
- Baylor, DA; Lamb, TD; Yau, KW (1979). "การตอบสนองของเซลล์รูปแท่งในจอประสาทตาต่อโฟตอนเดี่ยว" . Journal of Physiology . 288 : 613– 634. doi : 10.1113/jphysiol.1979.sp012716 . PMC 1281447 . PMID 112243 .
- Fan, Jie; Woodruff, Michael L; Cilluffo, Marianne C; Crouch, Rosalie K; Fain, Gordon L (2005). " การกระตุ้นการส่งสัญญาณของออปซินในเซลล์รูปแท่งของหนูที่เลี้ยงในที่มืดที่มียีน Rpe65 ถูกตัดออก"วารสารสรีรวิทยา568 (1): 83– 95. doi : 10.1113/jphysiol.2005.091942 . PMC 1474752 . PMID 15994181 .
- Hecht, Selig; Shlaer , Simon; Pirenne, Maurice Henri (1942). "พลังงาน ควอนตัม และการมองเห็น"วารสารสรีรวิทยาทั่วไป25 (6): 819– 840. doi : 10.1085/jgp.25.6.819 . PMC 2142545 . PMID 19873316 .
- Kawaguchi, Riki; Yu, Jiamei; Honda, Jane; Hu, Jane; Whitelegge, Julian; Ping, Peipei; Wiita, Patrick; Bok, Dean; Sun, Hui (2007). "ตัวรับเมมเบรนสำหรับโปรตีนที่จับกับเรตินอลเป็นตัวกลางในการดูดซึมวิตามินเอเข้าสู่เซลล์" . Science . 315 (5813): 820– 825. Bibcode : 2007Sci...315..820K . doi : 10.1126/science.1136244 . PMID 17255476 . S2CID 25258551 .
- Kloer, Daniel P.; Ruch, Sandra; Al-Babili, Salim; Beyer, Peter; Schulz, Georg E. (2005). "โครงสร้างของเอนไซม์ Carotenoid Oxygenase ที่สร้างเรตินา" Science . 308 (5719): 267– 269. Bibcode : 2005Sci...308..267K . doi : 10.1126/science.1108965 . PMID 15821095 . S2CID 6318853 .
- Luo, Dong-Gen; Xue, Tian; Yau, King-Wai (2008). "How vision begins: An odyssey" . Proceedings of the National Academy of Sciences . 105 (29): 9855– 9862. Bibcode : 2008PNAS..105.9855L . doi : 10.1073/pnas.0708405105 . PMC 2481352 . PMID 18632568 .การทบทวนประวัติศาสตร์ที่ดี
- Prado-Cabrero, Alfonso; Scherzinger, Daniel; Avalos, Javier; Al-Babili, Salim (2007). "การสังเคราะห์เรตินัลในเชื้อรา: ลักษณะเฉพาะของแคโรทีนอยด์ออกซิเจเนส CarX จาก Fusarium fujikuroi" . Eukaryotic Cell . 6 (4): 650– 657. doi : 10.1128/EC.00392-06 . PMC 1865656 . PMID 17293483 .
- Racker, Efraim; Stoeckenius, Walther (1974). "การสร้างเวสิเคิลเม มเบรนสีม่วงขึ้นใหม่เพื่อเร่งปฏิกิริยาการดูดซับโปรตอนที่ขับเคลื่อนด้วยแสงและการสร้างอะดีโนซีนไตรฟอสเฟต"วารสารเคมีชีวภาพ 249 (2): 662– 663. doi : 10.1016/S0021-9258(19)43080-9 . PMID 4272126 .
- Sadekar, Sumedha; Raymond, Jason; Blankenship, Robert E. (2006). "การอนุรักษ์โปรตีนเยื่อหุ้มเซลล์ที่เกี่ยวข้องห่างไกล: ศูนย์ปฏิกิริยาการสังเคราะห์แสงมีแกนโครงสร้างร่วมกัน". ชีววิทยาโมเลกุลและวิวัฒนาการ 23 ( 11): 2001– 2007. doi : 10.1093/molbev/msl079 . PMID 16887904 .
- Salom, David; Lodowski, David T.; Stenkamp, Ronald E.; Le Trong, Isolde; Golczak, Marcin; Jastrzebska, Beata; Harris, Tim; Ballesteros, Juan A.; Palczewski, Krzysztof (2006). "โครงสร้างผลึกของสารตัวกลางที่ถูกกระตุ้นด้วยแสงและถูกกำจัดโปรตอนของโรดอปซิน" Proceedings of the National Academy of Sciences . 103 (44): 16123– 16128. Bibcode : 2006PNAS..10316123S . doi : 10.1073/pnas.0608022103 . PMC 1637547 . PMID 17060607 .
- Schäfer, Günter; Engelhard, Martin; Müller, Volker (1999). "ชีวพลังงานของอาร์เคีย" . Microbiology and Molecular Biology Reviews . 63 (3): 570– 620. doi : 10.1128/MMBR.63.3.570-620.1999 . PMC 103747 . PMID 10477309 .
- Schmidt, Holger; Kurtzer, Robert; Eisenreich, Wolfgang; Schwab, Wilfried (2006). "เอนไซม์แคโรทีเนส AtCCD1 จาก Arabidopsis thaliana เป็นไดออกซิเจเนส"วารสารเคมีชีวภาพ 281 ( 15): 9845– 9851. doi : 10.1074/jbc.M511668200 . PMID 16459333 .
- Send, Robert; Sundholm, Dage (2007). "บันไดสู่จุดตัดรูปกรวย: การศึกษาเชิงคำนวณของการไอโซเมอไรเซชันของเรตินัล" วารสารเคมีฟิสิกส์ A . 111 (36): 8766– 8773. Bibcode : 2007JPCA..111.8766S . doi : 10.1021/jp073908l . PMID 17713894 .
- ซู, จิ๋หยิง; หลัว, ตงเก็น; เทราคิตะ, อากิฮิสะ; ชิจิดะ, โยชิโนริ; เหลียว, ซีเหวิน; คาซมี, มานิจา อ.; ซัคมาร์, โทมัส พี.; เหยา กิ่งไหว (2549) "ส่วนประกอบการถ่ายโอนแสงของ Parietal-Eye และผลกระทบเชิงวิวัฒนาการที่อาจเกิดขึ้น" ศาสตร์ . 311 (5767): 1617– 1621. Bibcode : 2006Sci...311.1617S . ดอย : 10.1126/science.1123802 . PMID16543463 . S2CID 28604455 .
- Venter, J. Craig ; Remington, K; Heidelberg, JF; Halpern, AL; Rusch, D; Eisen, JA; Wu, D; Paulsen, I; และคณะ (2004). "การจัดลำดับจีโนมแบบ Shotgun ของสิ่งแวดล้อมในทะเล Sargasso" Science . 304 ( 5667): 66–74 . Bibcode : 2004Sci...304...66V . CiteSeerX 10.1.1.124.1840 . doi : 10.1126/science.1093857 . PMID 15001713. S2CID 1454587 .มหาสมุทรเต็มไปด้วยโรดอปซินชนิดที่ 1
- Wang, Tao; Jiao, Yuchen; Montell, Craig (2007). "การวิเคราะห์เส้นทางที่จำเป็นสำหรับการสร้างวิตามินเอและสำหรับการแปลงสัญญาณแสงของแมลงหวี่"วารสาร ชีววิทยา ของเซลล์177 (2): 305– 316. doi : 10.1083/jcb.200610081 . PMC 2064138 . PMID 17452532 .
- Waschuk, Stephen A.; Bezerra, Arandi G.; Shi, Lichi; Brown, Leonid S. (2005). "Leptosphaeria rhodopsin : ปั๊มโปรตอนคล้ายแบคทีริโอโรดอปซินจากยูคาริโอต" Proceedings of the National Academy of Sciences . 102 (19): 6879– 6883. Bibcode : 2005PNAS..102.6879W . doi : 10.1073/pnas.0409659102 . PMC 1100770 . PMID 15860584 .
- Yokoyama, Shozo; Radlwimmer, F. Bernhard (2001). "พันธุศาสตร์ระดับโมเลกุลและวิวัฒนาการของการมองเห็นสีแดงและสีเขียวในสัตว์มีกระดูกสันหลัง"พันธุศาสตร์ 158 ( 4): 1697– 1710. doi : 10.1093/genetics/158.4.1697 . PMC 1461741 . PMID 11545071 .
- Briggs, Winslow R.; Spudich, John L., บรรณาธิการ (2005). คู่มือเกี่ยวกับตัวรับแสง . ไวลีย์. ISBN 978-3-527-31019-7.
- วอลด์, จอร์จ (1967). "ปาฐกถาโนเบล: พื้นฐานระดับโมเลกุลของการกระตุ้นการมองเห็น" (PDF) . สืบค้นเมื่อ23 กุมภาพันธ์ 2552 .
ลิงก์ภายนอก
- ก้าวแรกสู่การมองเห็น - พิพิธภัณฑ์สุขภาพแห่งชาติ
- การมองเห็นและการเปลี่ยนแปลงระดับโมเลกุลที่เกิดจากแสง
- กายวิภาคของจอประสาทตาและความสามารถในการมองเห็น
- เรตินา , อิมพีเรียลคอลเลจ v-chemlib
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เรตินา
เรตินัล (หรือที่รู้จักกันในชื่อ เรตินัลดีไฮด์ ) เป็น โครโมฟอร์โพ ลีอีน เรตินัลซึ่งจับกับโปรตีนที่เรียกว่า ออปซิน เป็นพื้นฐานทางเคมีของ การแปลงสัญญาณแสงในการมอง เห็น...
การเผาผลาญวิตามินเอ
สิ่งมีชีวิตสร้างเรตินัลโดยการแตกตัวออกซิเดชันที่ไม่สามารถย้อนกลับได้ของแคโรทีนอยด์ [ 7 ]
วิสัยทัศน์
เรตินัลเป็น โครโมฟอร์แบบคอนจูเกต ใน ดวงตาของสัตว์มีกระดูกสันหลัง เรตินัลเริ่มต้นในโครงสร้าง 11- ซิส -เรตินัล ซึ่งเมื่อจับ โฟตอน ที่มีความยาวคลื่นที่ถูกต้องได้ ก็จะยืดออกเป็นโครงสร้างออล- ทรานส์ -เรตินัล การเปลี่ยนแปลงโครงสร้างนี้จะไปกดกับโปรตีนออปซินใน เรตินา...
ออปซินส์
เรตินัลจับกับ ออปซิน ซึ่งเป็น ตัวรับที่เชื่อมต่อกับโปรตีน G (GPCRs) [ 14 ] [ 15 ] ออปซิน เช่นเดียวกับ GPCRs อื่นๆ มี อัลฟาเฮลิกซ์แบบทรานส์เมมเบรน 7 อัน ที่เชื่อมต่อกันด้วยลูป 6 อัน พบได้ใน เซลล์รับแสง ใน เรตินา ของดวงตา ออปซินใน เซลล์แท่ง ของสัตว์มี...