อ่าน 17 นาที
ยูเอสเอฟ1
ปัจจัยกระตุ้นต้นน้ำ 1 เป็น โปรตีน ที่ในมนุษย์ถูกเข้ารหัสโดย ยีน USF1 [ 5 ] [ 6 ]
ยูเอสเอฟ1
| ยูเอสเอฟ1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | USF1 , FCHL, FCHL1, HYPLIP1, MLTF, MLTFI, UEF, bHLHb11, ปัจจัยการถอดรหัสต้นน้ำ 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | โอมิม : 191523 ; เอ็มจีไอ : 99542 ; โฮโมโลยีน : 31426 ; การ์ดยีน : USF1 ; OMA : USF1 - ออโธโลจี | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




ปัจจัยกระตุ้นต้นน้ำ 1เป็นโปรตีนที่ในมนุษย์ถูกเข้ารหัสโดยยีนUSF1 [ 5 ] [ 6 ]
ยีน

ยีนปัจจัยกระตุ้นต้นน้ำ (USF) เข้ารหัสปัจจัยการถอดรหัส USF ซึ่งอยู่ใน ตระกูล โปรโตออนโคยีนMYCและมี โมทิฟ เบสิกเฮลิกซ์-ลูป-เฮลิกซ์ลิวซีนซิปเปอร์ (bHLH-LZ)ในโครงสร้างโปรตีน[ 7 ]เดิมที USF ถูกระบุว่าควบคุมโปรโมเตอร์ปลายหลักของอะดีโนไวรัสและการวิจัยล่าสุดได้เปิดเผยบทบาทของมันในการปกป้องเนื้อเยื่อเพิ่มเติม[ 7 ]โมทิฟ bHLH-LZ ช่วยให้โปรตีน USF มีความสามารถ ใน การกระตุ้นการถอดรหัส ผ่านการโต้ตอบกับ องค์ประกอบเริ่มต้น (Inr) และ โมทิฟ E-boxบน DNA ที่จับอยู่[ 7 ] [ 8 ]ในบริบทของกิจกรรม USF ที่เกิดจากอินซูลินและกลูโคส โมทิฟ E-box เหล่านั้นสามารถทำหน้าที่เป็นองค์ประกอบที่ตอบสนองต่อกลูโคส (GRE) และเป็นส่วนหนึ่งขององค์ประกอบการตอบสนองต่อคาร์โบไฮเดรต (ChoRE) เพื่อโต้ตอบกับปัจจัยการถอดรหัส[ 8 ]
ไอโซฟอร์ม
USF ประกอบด้วย ไอโซฟอร์มหลัก 2 ชนิดได้แก่ USF1 และUSF2ยีน USF1 ตั้งอยู่บนบริเวณโครโมโซม 1q22-q23 ทั้งในมนุษย์และหนู ส่วนยีน USF2 ตั้งอยู่บนโครโมโซม 19q13 ในมนุษย์และโครโมโซม 19q7 ในหนู ตามลำดับ[ 9 ]ทั้งทรานสคริปต์ USF1 และ USF2 ประกอบด้วยเอ็กซอน 10 เอ็กซอนและสามารถเกิดการตัดเอ็กซอน 4 ออกได้ในระหว่าง การสไปลซิ งแบบทางเลือก[ 7 ] [ 9 ]จากมุมมองของการควบคุมตนเอง ผลิตภัณฑ์จากการตัดเอ็กซอน 4 ออกเหล่านี้ทำหน้าที่เป็นตัวควบคุมเชิงลบที่เด่น และพบว่าสามารถยับยั้งการแสดงออกของยีนที่ขึ้นอยู่กับ USF ได้[ 7 ] [ 9 ]
โปรตีน

แม้ว่า USF1 และ USF2 จะมีลำดับกรดอะมิโนร่วมกันถึง 70% ในบริเวณ bHLH-LZ แต่โปรตีนแบบเต็มความยาวกลับมีความคล้ายคลึงกันเพียง 40% เท่านั้น นอกจากนี้ USF1 และ USF2 ยังแสดงปริมาณโปรตีนที่แตกต่างกันไปตามชนิดของเซลล์[ 7 ]พบว่าการแสดงออกของ USF1 และ USF2 เพิ่มขึ้นในระหว่างการสร้างความแตกต่างของเซลล์เม็ดเลือดแดง[ 10 ]แม้ว่าไอโซฟอร์มทั้งสองจะแสดงออกอย่างแพร่หลาย แต่ USF1 และ USF2 ก็ทำหน้าที่ควบคุมกระบวนการทางชีวภาพและหน้าที่ต่างๆ ในเซลล์ที่แตกต่างกัน ในขณะที่ USF1 ทำหน้าที่ควบคุมการเผาผลาญการตอบสนองของระบบภูมิคุ้มกันและการปกป้องเนื้อเยื่อ USF2 จะควบคุมการพัฒนาของตัวอ่อนการทำงานของสมองการเผาผลาญธาตุเหล็กและความสามารถในการ สืบพันธุ์เป็นหลัก [ 7 ]โครงสร้าง bHLH-LZ ที่มีการอนุรักษ์สูงบนปลาย C ของ USF ทำให้เกิดความจำเพาะในการจับสูงและส่งเสริมการก่อตัวของโฮโมไดเมอร์ USF1 หรือเฮเทโรไดเมอร์ USF1-USF2 สำหรับการจับกับ DNA [ 7 ] [ 9 ] [ 11 ]ในทางกลับกัน บริเวณเฉพาะของ USF (USR) บนบริเวณปลาย N ช่วยอำนวยความสะดวกในการเคลื่อนย้ายไปยังนิวเคลียสและการกระตุ้นของ USF1
การทำงาน
ยีน USF1 เข้ารหัสสมาชิกของ ตระกูล เบสิกเฮลิกซ์-ลูป-เฮลิกซ์ลิวซีนซิปเปอร์และสามารถทำหน้าที่เป็นปัจจัยการถอดรหัส ของเซลล์ได้ โปรตีนที่เข้ารหัสสามารถกระตุ้นการถอดรหัสผ่าน องค์ประกอบ เริ่มต้นที่ อุดมด้วยไพริมิดีน (Inr) และ โมทีฟ E-boxยีนนี้เชื่อมโยงกับภาวะไขมันในเลือดสูงแบบผสมในครอบครัว (FCHL) มีการระบุ ตัวแปรการถอดรหัสสองแบบที่เข้ารหัสไอโซฟอร์ม ที่แตกต่างกัน สำหรับยีนนี้[ 6 ]
การศึกษาในหนูชี้ให้เห็นว่าระดับ USF1 ที่ลดลงจะเพิ่มการเผาผลาญในไขมันสีน้ำตาล[ 12 ]
ระเบียบข้อบังคับ
การปรับเปลี่ยนความสัมพันธ์ในการจับกับ DNA
ลวดลาย E-box ที่สมมาตรเป็นเป้าหมายหลักของปัจจัยการถอดรหัส bHLH-LZ และ USF1 มีความสัมพันธ์ในการจับกับลำดับแกนกลาง CACGTG ในลวดลายสูง[ 9 ] กิจกรรมการจับกับ DNA ของ USF1 สามารถปรับเปลี่ยนได้โดย การเมทิลเล ชัน และ การอะเซ ทิเลชันของ DNA ที่จำเพาะต่อชนิดของเซลล์บนลวดลาย E-box หรือโดยการดัดแปลงหลังการถอดรหัสของโปรตีน USF1 ตัวอย่างเช่น การเมทิลเลชัน CpGบนลวดลาย E-box ตรงกลางจะยับยั้งการสร้างคอมเพล็กซ์ของ USF1 กับปัจจัยการถอดรหัสร่วม และด้วยเหตุนี้จึงลดการแสดงออกของยีนที่เกี่ยวข้องในเซลล์มะเร็งต่อมน้ำ เหลืองของหนู [ 9 ]ในทางตรงกันข้าม การฟอสโฟรีเลชันของ USF1 โดยโปรตีนไคเนสที่กระตุ้นด้วยไมโทเจน p38โปรตีนไคเนส Aหรือโปรตีนไคเนส Cจะเพิ่มการจับกับลวดลาย E-box และกระตุ้นการถอดรหัสยีน[ 9 ]
การฟอสฟอริเลชัน
โปรตีนไคเนสที่กระตุ้นด้วยไมโทเจน (MAPK) จะฟอสโฟรีเลตหมู่เซรินและทรีโอนีนของโปรตีนที่เป็นสารตั้งต้น และแปลงสัญญาณภายนอกเซลล์ที่เกิดจากปัจจัยการเจริญเติบโต ไมโทเจน หรือไซโตไคน์ ให้เป็นลำดับการฟอสโฟรีเลตภายในเซลล์ ซึ่งควบคุมการเพิ่มจำนวนเซลล์ การแบ่งเซลล์ การตอบสนองต่อความเครียด และอะพอพโทซิส (การตายของเซลล์ตามโปรแกรม ) [ 7 ]
การฟอสโฟรีเลชันโดย MAPK ทำให้เกิดการเปลี่ยนแปลงโครงสร้างของโปรตีน USF และเปิดเผยโดเมนการจับกับ DNA เพื่อการโต้ตอบ การเปิดเผยโครงสร้างที่เพิ่มขึ้นนี้ช่วยเพิ่มการจับกับ DNA และด้วยเหตุนี้จึงเพิ่มกิจกรรมการถอดรหัสของ USF [ 13 ]
- ERK1 (หรือที่รู้จักกันในชื่อMAPK3 ) และ ERK2 (หรือที่รู้จักกันในชื่อMAPK1 ) ฟอสโฟรีเลต USF1 เพื่อตอบสนองต่อการส่งสัญญาณ TFG-β ในเซลล์กล้ามเนื้อเรียบของหลอดเลือด[ 13 ]การส่งสัญญาณ SMAD2 และ SMAD3 หลังจากการกระตุ้นตัวรับ TFG-β ยังสามารถทำงานร่วมกับเส้นทาง EGFR/ERK เพื่อกระตุ้น USF1 ซึ่งจะควบคุมการแสดงออกของยีนของสารยับยั้งการทำงานของพลาสมิโนเจน-1 (PAI-1) ซึ่งเป็นตัวบ่งชี้ทางชีวภาพที่สำคัญและเป็นตัวทำนายการเสียชีวิตที่เกี่ยวข้องกับโรคหัวใจและหลอดเลือด[ 13 ]และเป็นเครื่องหมายของการพยากรณ์โรคที่ไม่ดีในมะเร็งเต้านม[ 7 ]
- เคซีนไคเนส 2หรือ CK-II (CK2) เป็นเอนไซม์เตตระเมอริกที่ประกอบด้วยซับยูนิตเร่งปฏิกิริยา 2 ซับยูนิตและซับยูนิตควบคุม 2 ซับยูนิต ในเซลล์ตับอ่อน CK2 จะฟอสโฟรีเลต USF1, PDX1และMST1เพื่อยับยั้งการแสดงออกของอินซูลิน[ 14 ]
| โปรตีนที่เป็นตัวกลางในการปรับเปลี่ยน USF1 | |
|---|---|
| การฟอสฟอริเลชัน | p38, pKA และ pKC, [ 9 ] ERK1/2, [ 13 ] DNA-PK [ 11 ] |
| อะเซทิเลชัน | PCAF [ 11 ] |
| เมทิลเลชัน' | SET7/92 [ 9 ] |
| โปรตีนที่ทำปฏิกิริยากับ USF1 | |
| ปัจจัยร่วมในการถอดรหัส | USF2, [ 7 ] SP1, PEA3, MTF1, [ 9 ] SREBP1-c, [ 11 ] MED17, [ 15 ] BAF60 [ 15 ] |
การถอดรหัสยีน
- ทรานส์ฟอร์มมิงโกรทแฟคเตอร์ β 1 ( TGF beta 1 ) ถูกเข้ารหัสโดยยีน TFGB1 ซึ่งมี E-box อยู่ภายในบริเวณโปรโมเตอร์ และมีส่วนเกี่ยวข้องกับการสะสมของเมทริกซ์นอกเซลล์มากเกินไปภายใต้สภาวะที่มีกลูโคสสูง[ 8 ]พบว่าการแสดงออกมากเกินไปของ USF1 หรือ USF2 จะเพิ่มกิจกรรมของโปรโมเตอร์ TFGB1 ในเซลล์ไตของตัวอ่อนมนุษย์ อย่างไรก็ตาม การแสดงออกมากเกินไปของ USF1 เท่านั้นที่นำไปสู่การหลั่ง TGF-β1 ที่เพิ่มขึ้น
- Thrombospondin 1 (TSP1) มีส่วนเกี่ยวข้องกับการพัฒนาของโรคไตจากเบาหวาน USF1/2 จับกับโมทีฟ E-box (CAGATG) บนโปรโมเตอร์ THBS1 ของมนุษย์และควบคุมการแสดงออกของ TSP1 ที่เกิดจากกลูโคสสูงในเซลล์เมแซงเจียล [ 8 ] พบว่าการแสดงออกเกินของ USF2 ช่วยเพิ่มกิจกรรมของโปรโมเตอร์ THBS1 และการแสดงออกของ TSP1 การเพิ่มขึ้นของการแสดงออกของ TSP1 ที่เกิดขึ้นจะส่งเสริมการสร้าง TGF-β ที่ออกฤทธิ์ต่อไป[ 8 ]
- ปัจจัยถอดรหัส AP-1 (AP-1) หมายถึงกลุ่มของปัจจัยถอดรหัสแบบไดเมอร์ที่ประกอบด้วยc-Jun , c-Fosหรือตัวกระตุ้นการถอดรหัส

ปัจจัย (ATF) ที่จับกับไซต์การจับ AP-1บน DNA [ 16 ]ไดเมอร์ cJun-cJun / cJun-cFos จะจับกับองค์ประกอบที่ตอบสนองต่อฟอร์บอล12-O-เตตราเดคาโนอิลฟอร์บอล-13-อะซิเตต (TPA) (บริเวณ TRE, TGACTCA) เป็นหลัก ในขณะที่ไดเมอร์ cJun-ATF และโฮโมไดเมอร์ ATF จะจับกับองค์ประกอบที่ตอบสนองต่อ cAMP ( CRE , TGACGTCA) เป็นหลัก [ 16 ]คอมเพล็กซ์ AP-1 จะถูกกระตุ้นเมื่อตอบสนองต่อกลูโคสสูง ความเครียดออกซิเดชันไลโปโปรตีนความหนาแน่นต่ำ (LDL) และ LDL ที่ถูกออกซิไดซ์ มีรายงานว่าระดับกลูโคสสูงจะเพิ่มการทำงานของการจับ USF และ AP-1 รวมถึงระดับโปรตีนของ cFos ด้วย[ 8 ]
ปฏิสัมพันธ์ระหว่าง USF1 และปัจจัยการถอดรหัสอื่นๆ รวมถึงSP1 , PEA3 (หรือที่รู้จักกันในชื่อETV4 ) และMTF1ยังนำไปสู่การควบคุมการถอดรหัสแบบร่วมมือกัน ตัวอย่างเช่น ลวดลาย leucine zipperของ USF1 จะดึงดูด PEA3 เพื่อสร้างคอมเพล็กซ์สามส่วนและควบคุมการถอดรหัสของBAXซึ่งเป็นตัวควบคุมอะพอพโทซิส ร่วมกัน [ 9 ]เป้าหมายที่ถูกควบคุมโดย USF1 อีกประการหนึ่งคือtopoisomerase III (hTOP3α) ซึ่งเร่งปฏิกิริยาการเปลี่ยนแปลงทางโทโพโลยีของ DNA ปรับเปลี่ยนโครงสร้างซูเปอร์คอยล์ของ DNA และเพิ่มการเข้าถึงโครมาตินสำหรับการแสดงออกของยีน[ 9 ]ปฏิสัมพันธ์ที่คล้ายกันมีอยู่ระหว่าง USF1 และJMJD1CหรือH3K9 demethylaseซึ่งปฏิสัมพันธ์ระดับโมเลกุลจะเปลี่ยนการเข้าถึงโครมาตินและเพิ่มการถอดรหัสของยีนสร้างไขมันหลายชุด รวมถึงยีนสำหรับfatty acid synthase (FASN), acetyl-CoA carboxylase (ACC), ATP citrate synthase (ACLY) และsterol regulatory element-binding protein 1 (SREBP1) [ 15 ]
ขอบเขตโครโมโซมโดย USF

โดยทั่วไปแล้ว โครโมโซมจะถูกจัดประเภทเป็นยูโครมาตินหรือเฮเทอโรโครมาตินที่มีการดัดแปลงฮิสโตน ระดับการอัดแน่น และรูปแบบการแสดงออกของยีนที่แตกต่างกัน เฮเทอโรโครมาตินเป็นโดเมนโครมาตินที่อัดแน่นและถูกยับยั้งการถอดรหัส ซึ่งมีลักษณะเฉพาะด้วยการดัดแปลงฮิสโตนหลังการแปลที่แตกต่างกัน[ 17 ]เฮเทอโรโครมาตินมีความจำเป็นต่อความเสถียรของจีโนมและการควบคุมการแสดงออกของยีน อย่างไรก็ตาม มันสามารถแพร่กระจายไปยังบริเวณ DNA ที่อยู่ใกล้เคียงและทำให้การแสดงออกของยีนไม่ทำงาน[ 17 ] [ 18 ]ดังนั้น องค์ประกอบขอบเขตของโครโมโซมจึงจำเป็นในการปิดกั้นการแพร่กระจายแบบสุ่มของเฮเทอโรโครมาตินและรักษาการแสดงออกของยีนให้คงที่[ 19 ]พบว่า USF1 และ USF2 ดึงดูดคอมเพล็กซ์ที่ดัดแปลงฮิสโตนต่างๆ รวมถึงคอมเพล็กซ์เมทิลทรานสเฟอเรสฮิสโตน H3 Set1และเมทิลทรานสเฟอเรสอาร์จินีน 3 H4 PRMT1โดยที่ PRMT1 เป็นที่ทราบกันว่าสร้างโดเมนโครมาตินที่ทำงานได้[ 19 ] การจับกันของ USF1/USF2 ทำให้เกิดการดัดแปลงฮิสโตนที่กระตุ้นการทำงานในระดับสูงบนนิวคลีโอโซมที่อยู่ติดกัน และด้วยเหตุนี้จึงป้องกันการแพร่กระจายของการดัดแปลงโครมาตินที่ทำให้เงียบจากเฮเทอโรโครมาติน เช่น การเมทิลเลชัน H3K9 และ K27 [ 19 ]

การดัดแปลงโครมาตินอื่นๆ ที่เกี่ยวข้องกับ USF1/USF2 ได้แก่ การดึงดูด E3 ubiquitin ligase, RNF20เพื่อเติมโมโนยูบิควิตินให้กับ ฮิส โตนH2B [ 19 ]พบว่าการสูญเสีย RNF20 ทำให้เกิดการขยายการดัดแปลงการปิดเสียงจากโดเมนเฮเทอโรโครมาติน 16 kbไปยังตำแหน่งβ-globin [ 19 ]นอกจากนี้ USF1 และ USF2 ยังสามารถจับกับไซต์ที่ไวต่อ DNase I 5' HS4 และดึงดูด H3 acetyltransferase, PCAFซึ่งจะปิดกั้นการแพร่กระจายของเฮเทอโรโครมาตินไปยังตำแหน่ง β-globin [ 18 ]
FASN กระตุ้นการสร้างไขมัน

USF เป็นที่ทราบกันดีว่าจับกับโปรโมเตอร์ของ ไพรูเวทไคเนสชนิด L บน DNA ในระดับกลูโคสและอินซูลินสูง อินซูลินที่มากเกินไปจะกระตุ้นไคเนสและฟอสฟาเทสที่ปรับเปลี่ยน USF, โปรตีนที่จับกับองค์ประกอบควบคุมสเตอรอล 1C (SREBP1C), โปรตีนที่จับกับองค์ประกอบตอบสนองต่อคาร์โบไฮเดรต (ChREBP) และตัวรับ Liver X (LXRs) หลังการแปล [ 11 ]เมื่อได้รับการกระตุ้นจากอินซูลิน USF1 และ USF2 จะจับกับ E-box ที่ -332 และ -65 ในบริเวณโปรโมเตอร์ของ FASN ซึ่งเข้ารหัสเอนไซม์สังเคราะห์กรดไขมัน (FAS) สำหรับการสร้างไขมัน[ 11 ]
การดัดแปลงหลังการแปลของ USF1 ที่แตกต่างกันจะกำหนดกิจกรรมและเส้นทางการส่งสัญญาณ และอาจส่งผลต่อกระบวนการสร้างไขมัน การเพิ่มขึ้นที่ผิดปกติของ การสังเคราะห์กรดไขมัน ใหม่ ที่เกิดจาก USF พบว่าทำให้เกิดการสะสมกรดไขมันภายในเซลล์และทำให้การแสดงออกของยีนและกระบวนการของเซลล์ เช่น การอยู่รอดของเซลล์มะเร็ง ผิดปกติ[ 20 ]
เส้นทางการสร้างไขมัน
- เมื่อระดับอินซูลินเพิ่มสูงขึ้นDNA-protein kinase (DNA-PK) ที่เกี่ยวข้องกับการซ่อมแซมความเสียหายของ DNA จะถูกดีฟอสโฟรีเลตและเปิดใช้งาน[ 11 ] DNA-PK ในรูปแบบที่ใช้งานได้จะฟอสโฟรีเลต USF1 ที่ S262 โดยอ้อมผ่านAMP-activated protein kinase (AMPK) การฟอสโฟรีเลตที่ S262 จะเพิ่มปฏิสัมพันธ์ของ USF1 กับ SREBP1C ใกล้กับองค์ประกอบควบคุมสเตอรอล (SRE) และอำนวยความสะดวกในการเปิดใช้งานร่วมกันของ SREBP1C และการถอดรหัสของยีนสร้างไขมันปลายน้ำ[ 11 ]
- การฟอสโฟรีเลชันของ USF1 S262 ยังดึงดูดPCAFมาอะเซทิเลต USF1 ที่ไซต์ K237 ทั้งการฟอสโฟรีเลชันของ S262 และการอะเซทิเลชันของ K237 ช่วยเพิ่มกิจกรรมของ USF1 และการกระตุ้นการถอดรหัสของ ยีน สังเคราะห์กรดไขมัน (FASN) ในภายหลัง [ 11 ]เอนไซม์สังเคราะห์กรดไขมัน (FAS) ร่วมกับอะเซทิล-โคเอ คาร์บอกซิเลส (ACC) ผลิตมาโลนิล-โคเอเปลี่ยนเป็นกรดไขมันสายยาว และส่งเสริมการสังเคราะห์กรดไขมันใหม่ เพื่อจัดหาพลังงานและการเติมไขมัน ให้กับโปรตีน[ 21 ] [ 20 ]
- USF1 ที่ได้รับการดัดแปลงด้วยการฟอสโฟรีเลชันที่ S262 และการอะเซทิเลชันที่ K237 ยังดึงดูดปัจจัยที่เกี่ยวข้องกับ BGR1 (หรือที่รู้จักกันในชื่อSMARCA4 ) 60c (BAF60c) [ 15 ]จากนั้น BAF60c จะถูกฟอสโฟรีเลตโดยโปรตีนไคเนส C ที่ผิดปกติ (aPKC) ที่ S257 ทำให้สามารถสร้างคอมเพล็กซ์ LipoBAF ที่โปรโมเตอร์ของยีนสร้างไขมันเพื่อควบคุมโครงสร้างโครมาตินและการถอดรหัสยีน[ 15 ]
- ในทางตรงกันข้ามHDAC9จะทำการดีอะเซทิเลต USF1 ในระหว่างการอดอาหารของเซลล์ ป้องกันการดึงดูดปัจจัยที่โต้ตอบกับ USF1 และยับยั้งการกระตุ้นการถอดรหัสของยีนสร้างไขมัน[ 11 ]
ในระยะเริ่มต้นของการพัฒนาตัวอ่อน
การถอดรหัส USF1 มีการเปลี่ยนแปลงอย่างกระฉับกระเฉงในระหว่างการแบ่งเซลล์แบบไมโอซิส โดยที่ mRNA ของ USF1 จะเพิ่มขึ้นอย่างมีนัยสำคัญเมื่อเซลล์ 2-8 ถูกสร้างขึ้น จากนั้นจะลดลงจนตรวจไม่พบในระยะบลาสโตซิสต์ ซึ่งบ่งชี้ถึงบทบาทของมันในการกระตุ้นจีโนมของตัวอ่อน[ 22 ]การกำจัด USF1 ด้วย siRNA แสดงให้เห็นว่าส่งผลกระทบต่ออัตราการเกิดบลาสโตซิสต์และทำให้การถอดรหัสของโปรตีนที่เกี่ยวข้องกับทวิสต์ 2 (เพิ่มขึ้น) และปัจจัยการเจริญเติบโตและการแยกความแตกต่าง-9และฟอลลิสแตติน (ลดลง) ผิดปกติ โดยส่งผลกระทบต่อบริเวณ E-box ขององค์ประกอบการจับโปรโมเตอร์ในระหว่างการเจริญเติบโตของโอโอไซต์[ 22 ]
ความสำคัญทางคลินิก
โรคไตจากเบาหวาน

โรคไตจากเบาหวาน (DKD) (หรือโรคไตจากเบาหวาน ) เป็น โรค ไมโครอัลบูมินูเรีย ที่ดำเนินไปอย่างต่อเนื่อง โดยมีการสูญเสียอัลบูมินในปัสสาวะเล็กน้อย (30–300 มิลลิกรัมต่อวัน) DKD ถูกมองว่าเป็นภาวะแทรกซ้อนของเบาหวานที่เกี่ยวข้องกับความผิดปกติของหลอดเลือด ขนาดเล็ก ในไต[ 23 ]ในการตรวจชิ้นเนื้อไต DKD มีลักษณะเฉพาะคือ การหนาตัวของฐานโกลเมอรูลัสและท่อไตการขยายตัวของเมแซงเจียมโกล เมอรูโลส เคลอโรซิสการเปลี่ยนแปลง ของโพโดไซต์ และการสูญเสียเนฟรอน[ 24 ] DKD เกิดขึ้นในผู้ป่วยเบาหวานร้อยละ 30-50 และนำไปสู่ภาวะไตวายในผู้ป่วยเบาหวานประเภทที่ 1 มากถึงร้อยละ 20 [ 24 ]อย่างไรก็ตาม ผู้ป่วย DKD จำนวนมากไม่มีอาการอัลบูมิ นูเรี ย[ 24 ]กลไกการเกิดโรค DKD เกิดจากการขนส่งกลูโคสที่ผิดปกติในระดับกลูโคสที่สูงขึ้นและการไหลเข้าของกลูโคสภายในเซลล์มากเกินไปในเซลล์เยื่อบุหลอดเลือด[ 23 ]ระดับกลูโคสที่สูงขึ้นจะคงอยู่ควบคู่ไปกับลักษณะทางเมตาบอลิซึมหลายอย่าง เช่นกรดไขมัน ส่วนเกิน และความเครียดออกซิเดชันรวมถึงแรงเฉือนที่เกิดจาก ความดัน โลหิตสูงและการไหลเวียน เลือดมากเกินไป ซึ่งอาจนำไปสู่การลดจำนวน ของหลอดเลือดขนาดเล็ก ภาวะขาดออกซิเจน และการปรับตัวที่ไม่เหมาะสมในการสร้าง หลอดเลือดใหม่ ใน ไต [ 23 ]
USF1 ซึ่งเป็นปัจจัยการถอดรหัสที่ไวต่ออินซูลินและทำงานเมื่อระดับกลูโคสสูง จะส่งเสริมการกระตุ้นการทำงานของยีนที่เกี่ยวข้องกับการเผาผลาญไขมัน รวมถึงเฮปาติกไลเปส (LIPC), เฮปาโตไซต์นิวเคลียร์แฟคเตอร์ 4 อัลฟา (HNF4A), อะโพลิโปโปรตีน AI (APOA1), อะโพลิโปโปรตีน L1 ( APOL1 ) และโปรตีนที่เกี่ยวข้องกับแฮปโตโกลบิน (HPR) [ 25 ] โดยเฉพาะอย่างยิ่ง APOL1 เป็นที่ทราบกันดีว่าสร้างคอมเพล็กซ์กับ APOA-I และ HDL เพื่ออำนวยความสะดวก ใน การเกิดออโตฟาจี ของเซลล์ เพื่อตอบสนองต่อการบาดเจ็บและเพื่อป้องกันโรคไต อย่างไรก็ตาม ตัวแปรความเสี่ยงของ APOL1 ที่เฉพาะเจาะจงกับโพโดไซต์จะยับยั้งการเกิดออโตฟาจีของเซลล์และสามารถกระตุ้นให้เกิดโรคไตได้[ 25 ]
มะเร็ง
การสังเคราะห์ไขมันใหม่ที่เกิดจาก FASN เพิ่มขึ้น
เซลล์มะเร็งแสดงฟีโนไทป์หลายประการ รวมถึงการเพิ่มขึ้นอย่างเห็นได้ชัดของไกลโคไลซิส แบบใช้ออกซิเจน การผลิตกรดแลคติก (ที่รู้จักกันในชื่อปรากฏการณ์วาร์เบิร์ก)การสังเคราะห์โปรตีนและดีเอ็นเอที่สูงขึ้น และการสังเคราะห์กรดไขมันแบบ de novoหรือแบบเอนโดเจนัส ที่เพิ่มขึ้น โดยเอนไซม์สังเคราะห์กรดไขมัน (FAS) [ 20 ] FAS สังเคราะห์พาลมิเตตจากมาโลนิล-CoA เป็นหลักซึ่งจะถูกเอสเทอริฟายด์ต่อไปเป็นไตรกลีเซอไรด์เพื่อเก็บพลังงาน โดยปกติแล้ว ยีนสังเคราะห์กรดไขมัน (FASN) จะทำงานในช่วงการเกิดตัวอ่อนและในปอดของทารกในครรภ์เพื่อผลิตสารหล่อลื่น อย่างไรก็ตาม ยีนนี้จะมีการแสดงออกในระดับต่ำทางสรีรวิทยาในเซลล์ผู้ใหญ่ที่ไม่เป็นมะเร็ง ในทางตรงกันข้าม การแสดงออกของ FASN ที่ผิดปกติมากเกินไปจะตรวจพบในมะเร็งหลายชนิด ได้แก่มะเร็งเต้านมมะเร็งลำไส้ใหญ่ มะเร็ง ต่อมลูกหมาก มะเร็งตับอ่อนและมะเร็งรังไข่[ 26 ] การสังเคราะห์ไขมัน แบบ de novoที่เกิดจาก FASN คิดเป็นมากกว่า 93% ของไตรกลีเซอไรด์ในเซลล์เนื้องอก[ 20 ]โดยเฉพาะอย่างยิ่ง เซลล์มะเร็งชอบใช้ไกลโคไลซิสมากกว่าออกซิเดชันในการใช้พลังงาน และเปลี่ยนเส้นทางผลิตภัณฑ์ไกลโคไลซิสไปสู่การสังเคราะห์กรดไขมันใหม่ เพื่อจัดหาไขมันสำหรับการสร้างเยื่อหุ้มเซลล์และ การเติมไขมันให้กับโปรตีนเพื่อการแพร่กระจายของเซลล์อย่างรวดเร็ว[ 20 ] ตัวอย่างเช่นพบ ว่า เส้นทาง PI3K-AKT เพิ่มขึ้นในเซลล์มะเร็งต่อมลูกหมาก LNCaPเพื่อกระตุ้นการแสดงออกของ FASN มากเกินไป ในขณะเดียวกัน การแสดงออกของเอนไซม์สังเคราะห์กรดไขมันมากเกินไปยังได้รับการสนับสนุนหลังการแปลโดย การลดยูบิควิตินเนชันที่เกิดจาก USP2aซึ่งทำให้ FAS มีเสถียรภาพสำหรับการส่งสัญญาณแบบต่อเนื่อง[ 20 ]นอกเหนือจาก การสร้างไขมัน ใหม่แล้ว FAS ยังส่งเสริมการกำหนดตำแหน่งของ VEGFR-2 ไปยังแพไขมันของเยื่อหุ้มเซลล์บุผนังหลอดเลือด และด้วยเหตุนี้จึงช่วยเพิ่มการสร้างหลอดเลือดใหม่ในการพัฒนาของเนื้องอก[ 26 ]ในขณะเดียวกัน การกระตุ้นร่วมกันระหว่างสัญญาณ FAS และ ERBB2 ( HER2 ) ยังช่วยส่งเสริมการเกิดเนื้องอก ซึ่งการขยายตัวของ ERBB2 เกี่ยวข้องกับอัตราการรอดชีวิตและการแพร่กระจายของเซลล์มะเร็งที่สูงขึ้น และการพยากรณ์โรคที่ไม่ดีในมะเร็งเต้านมและมะเร็งกระเพาะอาหาร โดยเฉพาะอย่างยิ่ง การเพิ่มขึ้นของ ERBB2 มีส่วนทำให้เกิดมะเร็งเต้านม 18-25% [ 27 ] ในเซลล์มะเร็งต่อมลูกหมากและเซลล์มะเร็งเม็ดเลือดขาวชนิดโปรไมอีโลไซต์ การกระตุ้น USF1 ยังทำให้ระดับ PAI-1สูงขึ้นด้วยการแสดงออกและยับยั้งการเกิดอะพอพโทซิสโดยธรรมชาติหรือที่เกิดจากแคมป์โทเทซิน[ 13 ]
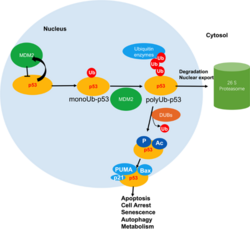
ปฏิสัมพันธ์ระหว่าง USF1 และ p53 ลดลง และความไม่เสถียรของ p53 เพิ่มขึ้น
การพยากรณ์โรคที่ไม่ดีของมะเร็งกระเพาะอาหารเกี่ยวข้องกับการแสดงออกของ USF1 และ p53 ที่ต่ำ[ 28 ]ในบรรดาผู้ป่วยมะเร็งกระเพาะอาหาร 88% ของผู้ป่วยได้รับการวินิจฉัยว่าติด เชื้อ H. pyloriและครึ่งหนึ่งของผู้ป่วยแสดงการแสดงออกของ USF1 ที่ต่ำกว่าในเนื้อเยื่อเนื้องอก ในเชิงกลไกH. pyloriกระตุ้นให้เกิดการไฮเปอร์เมทิลเลชันของ DNA ในบริเวณโปรโมเตอร์ของ USF1 และ USF2 และยับยั้งการแสดงออก การแสดงออกที่ลดลงจะลดการโต้ตอบระหว่าง USF1 และ p53 เมื่อเกิดความเสียหายของ DNA ทำให้ p53 เชื่อมโยงกับ E3-ubiquitin ligase HDM2 (หรือที่รู้จักกันในชื่อMDM2 ) บ่อยขึ้น และเพิ่มความไม่เสถียรของ p53 ในเซลล์มะเร็ง[ 28 ]
ภาวะไขมันในเลือดสูงแบบผสมทางพันธุกรรม
ภาวะไขมันในเลือดสูงแบบผสมในครอบครัว(FCHL) ถูกใช้ครั้งแรกเพื่ออธิบายความผิดปกติของไขมันในสมาชิกครอบครัวซีแอตเติล 47 คนที่มีภาวะคอเลสเตอรอลสูงและไตรกลีเซอไรด์สูง[ 29 ]โปรไฟล์ไขมันหลักของ FCHL มีลักษณะเด่นคือระดับคอเลสเตอรอล/ไตรกลีเซอไรด์ในซีรั่มสูงอะโพลิโปโปรตีน บี (APOB) และ LDL สูง หลักฐานทางพันธุกรรมชี้ให้เห็นตำแหน่งที่เกี่ยวข้องกับ FCHL บนโครโมโซมมนุษย์ 1q21-q23 ซึ่งเชื่อมโยงกับกลุ่มอาการเมตาบอลิก[ 30 ]การทำแผนที่อย่างละเอียดของบริเวณที่เชื่อมโยงเหล่านั้นระบุ USF1 เป็นยีนที่ถูกโคลนตามตำแหน่งตัวแรกสำหรับ FCHL และเป็นเป้าหมายสำหรับการรักษา FCHL นอกจากนี้ปัจจัยนิวเคลียร์ของเซลล์ตับ 4 อัลฟา (HNF4A) ยังเกี่ยวข้องกับระดับไขมันสูงและกลุ่มอาการเมตาบอลิกด้วย ผลกระทบร่วมกันของ USF1 และ HNF4A ได้รับการแสดงให้เห็นว่าควบคุมการแสดงออกของอะโพลิโปโปรตีน A-II (APOA2) และอะโพลิโปโปรตีน C-III (APOC3) [ 30 ]การกลายพันธุ์ใน USF1, HNF4A และอะโพลิโปโปรตีนยังเพิ่มความเสี่ยงของผู้ป่วยต่อ FCHL อีกด้วย[ 30 ]ยีนเพิ่มเติมที่อยู่ภายใต้การควบคุมของ USF1 และเกี่ยวข้องกับการเผาผลาญกลูโคส/ไขมัน ได้แก่ อะโพลิโปโปรตีน A5 ( APOA5 ), อะโพลิโปโปรตีน E (APOE), ไลเปสที่ไวต่อฮอร์โมน (LIPE ), ไลเป สในตับ (LIPC), กลูโคไคเนส (GCK), โปรตีนที่เกี่ยวข้องกับหน่วยย่อยเร่งปฏิกิริยาของกลูโคส-6-ฟอสฟาเตสเฉพาะเกาะ (IGRP), อินซูลิน, ตัวรับกลูคากอน (GCGR) และตัวขนส่งแคสเซ็ตที่จับกับ ATP A1 ( ABCA1 ) [ 30 ]
ปฏิสัมพันธ์
USF1 ( ยีนของมนุษย์) ได้รับการแสดงให้เห็นว่ามีปฏิสัมพันธ์กับUSF2 [ 31 ] [ 32 ] FOSL1 [ 33 ]และGTF2I [ 34 ] [ 35 ]
อ่านเพิ่มเติม
- Corre S, Galibert MD (มกราคม 2549). "[USF เป็นองค์ประกอบควบคุมที่สำคัญของการแสดงออกของยีน]" . Médecine/Sciences . 22 (1): 62– 67. doi : 10.1051/medsci/200622162 . PMID 16386222 .
- ลี เจซี, ลูซิส เอเจ, ปาจูกันตา พี (เมษายน 2549) "ภาวะไขมันในเลือดสูงรวมในครอบครัว: ปัจจัยการถอดรหัสต้นน้ำ 1 และมากกว่านั้น" ความคิดเห็นปัจจุบันในด้าน Lipidology . 17 (2): 101– 109. ดอย : 10.1097/01.mol.0000217890.54875.13 . PMID16531745 . S2CID 20122462 .
- Roy AL, Meisterernst M, Pognonec P, Roeder RG (พฤศจิกายน 1991). "ปฏิสัมพันธ์แบบร่วมมือของปัจจัยเริ่มต้นการถอดรหัสที่จับกับตัวเริ่มต้นและตัวกระตุ้นเฮลิกซ์-ลูป-เฮลิกซ์ USF" Nature . 354 (6350): 245– 248. Bibcode : 1991Natur.354..245R . doi : 10.1038/354245a0 . PMID 1961251 . S2CID 4260885 .
- Gregor PD, Sawadogo M, Roeder RG (ตุลาคม 1990). "ปัจจัยการถอดรหัสล่าช้าหลักของอะดีโนไวรัส USF เป็นสมาชิกของกลุ่มโปรตีนควบคุมแบบเฮลิกซ์-ลูป-เฮลิกซ์และจับกับ DNA ในรูปไดเมอร์" Genes & Development . 4 (10): 1730– 1740. doi : 10.1101/gad.4.10.1730 . PMID 2249772 .
- Henrion AA, Martinez A, Mattei MG, Kahn A, Raymondjean M (มกราคม 1995). "โครงสร้าง ลำดับ และตำแหน่งโครโมโซมของยีนสำหรับปัจจัยการถอดรหัส USF2 ในหนู" Genomics . 25 (1): 36– 43. doi : 10.1016/0888-7543(95)80107-W . PMID 7774954 .
- Ferré-D'Amaré AR, Pognonec P, Roeder RG, Burley SK (มกราคม 1994). "โครงสร้างและหน้าที่ของโดเมน b/HLH/Z ของ USF"วารสารEMBO 13 ( 1): 180– 189. doi : 10.1002/j.1460-2075.1994.tb06247.x . PMC 394791 . PMID 8306960 .
- Viollet B, Lefrançois-Martinez AM, Henrion A, Kahn A, Raymondjean M, Martinez A (มกราคม 1996). "ลักษณะทางภูมิเคมีและคุณสมบัติการโต้ตอบของไอโซฟอร์มปัจจัยกระตุ้นต้นน้ำ"วารสารเคมีชีวภาพ 271 ( 3): 1405– 1415. doi : 10.1074/jbc.271.3.1405 . PMID 8576131 .
- Ghosh AK, Datta PK, Jacob ST (กุมภาพันธ์ 1997). "บทบาทคู่ของโปรตีนเฮลิกซ์-ลูป-เฮลิกซ์-ซิปเปอร์ USF ในการถอดรหัสยีนไรโบโซมอาร์เอ็นเอในร่างกาย" Oncogene . 14 (5): 589– 594. doi : 10.1038/sj.onc.1200866 . PMID 9053857 . S2CID 23764497 .
- Pognonec P, Boulukos KE, Aperlo C, Fujimoto M, Ariga H, Nomoto A, Kato H (พฤษภาคม 1997). "ปฏิสัมพันธ์ข้ามตระกูลระหว่างโปรตีน bHLHZip USF และ bZip Fra1 ส่งผลให้กิจกรรมของ AP1 ลดลง" . Oncogene . 14 (17): 2091– 2098. doi : 10.1038/sj.onc.1201046 . PMID 9160889 .
- Roy AL, Du H, Gregor PD, Novina CD, Martinez E, Roeder RG (ธันวาคม 1997). "การโคลนโปรตีนที่จับกับ inr และ E-box, TFII-I ซึ่งมีปฏิสัมพันธ์ทางกายภาพและเชิงฟังก์ชันกับ USF1"วารสารEMBO . 16 (23): 7091– 7104. doi : 10.1093/emboj/16.23.7091 . PMC 1170311 . PMID 9384587 .
- Pajukanta P, Nuotio I, Terwilliger JD, Porkka KV, Ylitalo K, Pihlajamäki J และคณะ (เมษายน 2541). "การเชื่อมโยงภาวะไขมันในเลือดสูงรวมในครอบครัวกับโครโมโซม 1q21-q23" พันธุศาสตร์ธรรมชาติ . 18 (4): 369– 373. ดอย : 10.1038/ng0498-369 . PMID9537421 . S2CID 5905269 .
- Hartley JL, Temple GF, Brasch MA (พฤศจิกายน 2000). "การโคลน DNA โดยใช้การรวมตัวใหม่เฉพาะตำแหน่งในหลอดทดลอง" . Genome Research . 10 (11): 1788– 1795. doi : 10.1101/gr.143000 . PMC 310948 . PMID 11076863 .
- Wiemann S, Weil B, Wellenreuther R, Gassenhuber J, Glassl S, Ansorge W และคณะ (มีนาคม 2544). "สู่แคตตาล็อกของยีนและโปรตีนของมนุษย์: การจัดลำดับและการวิเคราะห์ cDNA ของมนุษย์ที่เข้ารหัสโปรตีนอย่างสมบูรณ์ 500 รายการใหม่" . Genome Research . 11 (3): 422– 435. doi : 10.1101/gr.GR1547R . PMC 311072 . PMID 11230166 .
- Bengtsson SH, Madeyski-Bengtson K, Nilsson J, Bjursell G (กรกฎาคม 2545). "การควบคุมการถอดรหัสของยีนคาร์บอกซิลเอสเทอร์ไลเปสของมนุษย์ในโมโนไซต์ THP-1: E-box ที่จำเป็นสำหรับการกระตุ้นจะจับกับปัจจัยกระตุ้นต้นน้ำ 1 และ 2" วารสารชีวเคมี 365 ( ตอนที่ 2 ): 481–488 . doi : 10.1042/BJ20020223 . PMC 1222691. PMID 11945176 .
- Villavicencio EH, Yoon JW, Frank DJ, Füchtbauer EM, Walterhouse DO, Iannaccone PM (เมษายน 2545). "การควบคุม E-box ร่วมกันของ GLI1 ในมนุษย์โดย TWIST และ USF" Genesis . 32 (4): 247– 258. doi : 10.1002/gene.10078 . PMID 11948912 . S2CID 12132097 .
- Coulson JM, Edgson JL, Marshall-Jones ZV, Mulgrew R, Quinn JP, Woll PJ (กุมภาพันธ์ 2546). "ปัจจัยกระตุ้นต้นน้ำกระตุ้น โปรโมเตอร์ ของวาโซเพรสซิ นผ่านโมทีฟหลายแบบ รวมถึง E-box ที่ไม่เป็นไปตามแบบแผน"วารสารชีวเคมี 369 (ตอนที่ 3): 549–561 . doi : 10.1042/BJ20021176 . PMC 1223122. PMID 12403649 .
- Salero E, Giménez C, Zafra F (มีนาคม 2546). "การระบุโมทีฟ E-box ที่ไม่เป็นไปตามแบบแผนว่าเป็นองค์ประกอบควบคุมในบริเวณโปรโมเตอร์ส่วนต้นของยีนอะโพลิโปโปรตีน E"วารสารชีวเคมี 370 ( ตอน ที่ 3): 979–986 . doi : 10.1042/BJ20021142 . PMC 1223214. PMID 12444925 .
- Pickwell GV, Shih H, Quattrochi LC (เมษายน 2546). "การโต้ตอบของโปรตีนปัจจัยกระตุ้นต้นน้ำกับ E-box ที่ตั้งอยู่ภายในยีน CYP1A2 5'-flanking ของมนุษย์มีส่วนช่วยในการกระตุ้นการถอดรหัสยีนพื้นฐาน" Biochemical Pharmacology . 65 (7): 1087– 1096. doi : 10.1016/S0006-2952(03)00037-6 . PMID 12663044 .
ลิงก์ภายนอก
- แฟคทอรบุ๊คยูเอสเอฟ1
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ยูเอสเอฟ1
ปัจจัยกระตุ้นต้นน้ำ 1 เป็น โปรตีน ที่ในมนุษย์ถูกเข้ารหัสโดย ยีน USF1 [ 5 ] [ 6 ]
ยีน
ยีนปัจจัยกระตุ้นต้นน้ำ (USF) เข้ารหัส ปัจจัยการถอดรหัส USF ซึ่งอยู่ใน ตระกูล โปรโตออนโคยีน MYC และมี โมทิฟ เบสิกเฮลิกซ์-ลูป-เฮลิกซ์ ลิวซีนซิปเปอร์ (bHLH-LZ) ในโครงสร้างโปรตีน [ 7 ] เดิมที USF ถูกระบุว่าควบคุมโปรโมเตอร์ปลายหลักของ อะดีโนไวรัส...
ไอโซฟอร์ม
USF ประกอบด้วย ไอโซฟอร์ม หลัก 2 ชนิดได้แก่ USF1 และ USF2 ยีน USF1 ตั้งอยู่บนบริเวณโครโมโซม 1q22-q23 ทั้งในมนุษย์และหนู ส่วนยีน USF2 ตั้งอยู่บนโครโมโซม 19q13 ในมนุษย์และโครโมโซม 19q7 ในหนู ตามลำดับ [ 9 ] ทั้งทรานสคริปต์ USF1 และ USF2 ประกอบด้วยเอ็กซอน 10...
โปรตีน
แม้ว่า USF1 และ USF2 จะมีลำดับกรดอะมิโนร่วมกันถึง 70% ในบริเวณ bHLH-LZ แต่โปรตีนแบบเต็มความยาวกลับมีความคล้ายคลึงกันเพียง 40% เท่านั้น นอกจากนี้ USF1 และ USF2 ยังแสดงปริมาณโปรตีนที่แตกต่างกันไปตามชนิดของเซลล์ [ 7 ] พบว่าการแสดงออกของ USF1 และ USF2...