อ่าน 26 นาที
ไรโบโซม
ไรโบโซม ( / ˈ r aɪ b ə z oʊ m , - s oʊ m / ) คืออนุภาคไรโบนิวคลีโอโปรตีนที่พบในเซลล์ ทุกชนิด
ไรโบโซม
| ไรโบโซม | |
|---|---|
 หน่วยย่อยขนาดใหญ่ (สีแดง) และหน่วยย่อยขนาดเล็ก (สีน้ำเงิน) ของไรโบโซม | |
| รายละเอียด | |
| ส่วนหนึ่งของ | เซลล์ (ชีววิทยา) |
| ตัวระบุ | |
| เมช | D012270 |
| เอฟเอ็มเอ | 66867 |
| คำศัพท์ทางกายวิภาคศาสตร์ของจุลกายวิภาคศาสตร์ | |
| ชีววิทยาของเซลล์ | |
|---|---|
| แผนภาพเซลล์สัตว์ | |
 ส่วนประกอบของเซลล์สัตว์ทั่วไป:
|
ไรโบโซม ( / ˈ r aɪ b ə z oʊ m , - s oʊ m / ) คืออนุภาคไรโบนิวคลีโอโปรตีนที่พบในเซลล์ ทุกชนิด ทั้งโปรคาริโอตและยูคาริโอตมีหน้าที่ในการสังเคราะห์โปรตีนไรโบโซมทำหน้าที่เป็นเครื่องจักรระดับโมเลกุลในการแปลรหัสของสายอาร์เอ็นเอส่งสาร (mRNA) และสร้างโปรตีนไรโบโซมเชื่อมต่อกรดอะมิโนเข้าด้วยกันตามลำดับที่กำหนดโดยโคดอนของโมเลกุล mRNA เพื่อสร้างสายโพลีเปปไทด์ ไรโบโซมประกอบด้วยหน่วยย่อยขนาดใหญ่และขนาดเล็ก แต่ละหน่วยประกอบด้วย โมเลกุล อาร์เอ็นเอไรโบโซมหนึ่งโมเลกุล หรือมากกว่า และโปรตีนไรโบโซม จำนวนมาก ไรโบโซมและโมเลกุลที่เกี่ยวข้องยังเรียกอีกอย่างว่าเครื่องมือการแปลรหัส
การสร้างไรโบโซมเป็นกระบวนการสร้างไรโบโซม กระบวนการนี้ใช้พลังงานสูงและเป็นกระบวนการแบบไดนามิก โดยต้องสังเคราะห์โปรตีนประมาณ 200 ชนิดในการประมวลผล RNA ของไรโบโซม และประกอบเข้ากับโปรตีนของไรโบโซมเพื่อสร้างหน่วยย่อยของไรโบโซม
ภาพรวม
ลำดับของRNAที่เข้ารหัสลำดับของกรดอะมิโนในโปรตีนจะถูกถอดรหัสเป็น สาย mRNA ( messenger RNA ) ไรโบโซมจะจับกับโมเลกุล mRNA และใช้ลำดับนิวคลีโอไทด์ ของ RNA เพื่อกำหนดลำดับของกรดอะมิโนที่จำเป็นในการสร้างโปรตีน กรดอะมิโนจะถูกเลือกและนำไปยังไรโบโซมโดย โมเลกุล tRNA ( transfer RNA) ซึ่งเข้าไปในไรโบโซมและจับกับสาย mRNA ผ่านทาง ลูปก้าน แอนติโคดอนสำหรับแต่ละรหัสสามตัว ( โคดอน ) ใน mRNA จะมี tRNA ที่ไม่ซ้ำกันซึ่งต้องมีแอนติโคดอนที่ตรงกันอย่างแม่นยำ และนำกรดอะมิโนที่ถูกต้องเพื่อรวมเข้ากับ สาย โพลีเปปไทด์ ที่กำลังเติบโต เมื่อโปรตีนถูกผลิตขึ้นแล้ว โปรตีนนั้นจะสามารถพับตัวเพื่อสร้างโครงสร้างสามมิติ ที่ใช้งานได้ [ 1 ] [ 2 ]
ในระหว่างการแปล การสังเคราะห์โปรตีนจากหน่วยสร้างจะเกิดขึ้นในสี่ขั้นตอน ได้แก่ การเริ่มต้น การยืดตัว การสิ้นสุด และการรีไซเคิลไรโบโซม[ 3 ]รหัสเริ่มต้นในโมเลกุล mRNA ทั้งหมดมีลำดับ AUG รหัสหยุดอาจเป็น UAA, UAG หรือ UGA เนื่องจากไม่มีโมเลกุล tRNA ที่รู้จักรหัสเหล่านี้ ไรโบโซมจึงรับรู้ว่าการแปลเสร็จสมบูรณ์แล้ว[ 4 ] เมื่อไรโบโซมอ่านโมเลกุล mRNA เสร็จแล้ว หน่วยย่อยทั้งสองจะแยกออกจากกันและมักจะถูกทำลาย แต่สามารถนำกลับมาใช้ใหม่ได้ ไรโบโซมเป็น เอนไซม์ชนิดหนึ่งเรียกว่าไรโบไซม์เนื่องจาก กิจกรรม การถ่ายโอนเปปไทด์ที่เร่งปฏิกิริยา ซึ่งเชื่อมโยงกรดอะมิโนเข้าด้วยกันนั้นดำเนินการโดย RNA ของไรโบโซม[ 5 ]
ไรโบโซมจากแบคทีเรียอาร์เคียและยูคาริโอต (ในระบบสามโดเมน ) มีความคล้ายคลึงกันอย่างน่าทึ่ง ซึ่งเป็นหลักฐานบ่งชี้ถึงต้นกำเนิดร่วมกัน แต่แตกต่างกันในขนาด ลำดับ โครงสร้าง และอัตราส่วนของโปรตีนต่ออาร์เอ็นเอ ความแตกต่างในโครงสร้างนี้ทำให้ยาปฏิชีวนะ บางชนิด สามารถฆ่าแบคทีเรียได้โดยการยับยั้งไรโบโซมของแบคทีเรีย ในขณะที่ไรโบโซมของมนุษย์ไม่ได้รับผลกระทบ ในทุกโดเมน โพลีโซม ที่ประกอบด้วยไร โบโซมสองตัวขึ้นไปอาจเคลื่อนที่ไปตามสายเอ็มอาร์เอ็นเอเส้นเดียวในเวลาเดียวกัน โดยแต่ละตัวจะอ่านลำดับเฉพาะและสร้างโมเลกุลโปรตีนที่สอดคล้องกัน
ไรโบโซมไมโตคอนเดรีย (ไมโตริโบโซม) ของเซลล์ยูคาริโอตนั้นแตกต่างจากไรโบโซมอื่นๆ ไรโบโซมเหล่านี้ทำหน้าที่คล้ายกับไรโบโซมในแบคทีเรีย ซึ่งสะท้อนถึงต้นกำเนิดทางวิวัฒนาการของไมโตคอนเดรียในฐานะแบคทีเรียเอนโดซิมไบโอติก[ 6 ] [ 7 ]
การค้นพบ
ไรโบโซมถูกสังเกตครั้งแรกในช่วงกลางทศวรรษ 1950 ในรูปของอนุภาคหรือเม็ดที่มีความหนาแน่นสูงโดยนักชีววิทยาเซลล์ชาวโรมาเนีย-อเมริกันGeorge Emil Paladeโดยใช้กล้องจุลทรรศน์อิเล็กตรอน[ 8 ] ในตอนแรกเรียกว่า เม็ด Paladeเนื่องจากโครงสร้างที่เป็นเม็ด[ 9 ]คำว่า "ไรโบโซม" ถูกเสนอในปี 1958 โดย Howard M. Dintzis: [ 10 ]
ในระหว่างการประชุมสัมมนา เกิดปัญหาทางด้านความหมายขึ้น ผู้เข้าร่วมบางคนเข้าใจว่า "ไมโครโซม" หมายถึงอนุภาคไรโบโปรตีนในส่วนของไมโครโซมที่ปนเปื้อนด้วยโปรตีนและไขมันชนิดอื่น ในขณะที่ผู้เข้าร่วมคนอื่นๆ เข้าใจว่าไมโครโซมประกอบด้วยโปรตีนและไขมันที่ปนเปื้อนด้วยอนุภาค วลี "อนุภาคไมโครโซม" ดูไม่เหมาะสม และ "อนุภาคไรโบโปรตีนในส่วนของไมโครโซม" ก็ฟังดูไม่เป็นธรรมชาติ ในระหว่างการประชุม มีการเสนอคำว่า "ไรโบโซม" ซึ่งเป็นชื่อที่ฟังดูดีและน่าพอใจ ความสับสนในปัจจุบันจะหมดไปหากนำคำว่า "ไรโบโซม" มาใช้เรียกอนุภาคไรโบโปรตีนที่มีขนาดตั้งแต่ 35 ถึง100 ไมโครเมตร
— อัลเบิร์ต คลอดด์ อนุภาคไมโครโซมและการสังเคราะห์โปรตีน[ 11 ]
อัลเบิร์ต คลอดด์ , คริสเตียน เดอ ดูฟและจอร์จ เอมิล พาเลดได้รับรางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ ร่วมกัน ในปี 1974 จากการค้นพบไรโบโซม[ 12 ]รางวัลโนเบลสาขาเคมีประจำปี 2009 มอบให้แก่เวนคัตรามัน รามาคริชนัน , โทมัส เอ. สไตทซ์และเอดา อี. โยนาธสำหรับการกำหนดโครงสร้างและกลไกโดยละเอียดของไรโบโซม[ 13 ]
โครงสร้าง

ไรโบโซมเป็นอนุภาคไรโบนิวคลีโอโปรตีนที่สร้างขึ้นจากสารประกอบของไรโบโซมอาร์เอ็นเอและโปรตีน จัดเรียงเป็นสองหน่วยย่อยไรโบโซม หน่วยหนึ่งใหญ่และอีกหน่วยหนึ่งเล็ก ไรโบโซมเป็นเครื่องจักรโมเลกุล ที่ซับซ้อนซึ่งมีอยู่ใน เซลล์ทั้งหมดทั้งโปรคาริโอตและยู คาริโอต หน่วยย่อยไรโบโซมของโปรคาริโอตและยูคาริโอตค่อนข้างคล้ายกัน[ 14 ]
ไรโบโซมส่วนใหญ่ประกอบด้วยRNA ไรโบโซม (rRNA) ที่ไม่เข้ารหัส เฉพาะทาง รวมถึงโปรตีนไรโบโซม ที่แตกต่างกันหลายสิบชนิด (จำนวนจะแตกต่างกันเล็กน้อยในแต่ละสายพันธุ์) โปรตีนไรโบโซมและ rRNA จะถูกจัดเรียงเป็นหน่วยย่อยไรโบโซมสองหน่วยที่แตกต่างกัน คือ หน่วยใหญ่หนึ่งหน่วยและหน่วยเล็กหนึ่งหน่วย หน่วยย่อยเหล่านี้จะเข้าคู่กันโดยล็อกรอบสาย mRNA และทำงานร่วมกันเพื่อแปล mRNA เป็นสายโพลีเปปไทด์ในระหว่าง การ สังเคราะห์โปรตีน[ 15 ]
โปรคาริโอต
แบคทีเรีย
ไรโบโซม ของแบคทีเรียมีเส้นผ่านศูนย์กลางประมาณ 20 นาโนเมตร (200 อังสตรอม ) และประกอบด้วย rRNA 65% และโปรตีนไรโบโซม 35% [ 16 ]ไรโบโซมของยูคาริโอตมีเส้นผ่านศูนย์กลางระหว่าง 25 ถึง 30 นาโนเมตร (250–300 อังสตรอม) โดยมีอัตราส่วน rRNA ต่อโปรตีนที่ใกล้เคียงกับ 1 [ 17 ] งานวิจัยเชิงผลึกศาสตร์[ 18 ]แสดงให้เห็นว่าไม่มีโปรตีนไรโบโซมอยู่ใกล้กับบริเวณปฏิกิริยาสำหรับการสังเคราะห์พอลิเปปไทด์ ซึ่งชี้ให้เห็นว่าส่วนประกอบโปรตีนของไรโบโซมไม่ได้มีส่วนร่วมโดยตรงในการเร่งปฏิกิริยาการสร้างพันธะเปปไทด์ แต่โปรตีนเหล่านี้ทำหน้าที่เป็นโครงสร้างค้ำยันที่อาจช่วยเพิ่มความสามารถของ rRNA ในการสังเคราะห์โปรตีน[ 19 ]

หน่วยวัดที่ใช้ในการอธิบายหน่วยย่อยของไรโบโซมและชิ้นส่วน rRNA คือหน่วยสเวดเบิร์ก (Svedberg unit)ซึ่งเป็นการวัดอัตราการตกตะกอนในการปั่นเหวี่ยงไม่ใช่ขนาด นี่คือเหตุผลที่ชื่อของชิ้นส่วนต่างๆ ไม่สอดคล้องกัน ตัวอย่างเช่น ไรโบโซม 70S ของแบคทีเรียประกอบด้วยหน่วยย่อย 50S และ 30S
โปรคาริโอตมีไรโบโซม 70 Sแต่ละอันประกอบด้วยซับยูนิตขนาดเล็ก ( 30S ) และซับยูนิตขนาดใหญ่ ( 50S ) ตัวอย่างเช่นE. coli มีซับยูนิต RNA 16S (ประกอบด้วยนิวคลีโอไทด์ 1540 ตัว) ที่จับกับโปรตีน 21 ตัว ซับยูนิตขนาดใหญ่ประกอบด้วย ซับยูนิต RNA 5S (นิวคลีโอไทด์ 120 ตัว) ซับยูนิต RNA 23S (นิวคลีโอไทด์ 2900 ตัว) และโปรตีน 31 ตัว[ 14 ]
ไรโบโซมของE. coli (แบคทีเรีย) [ 21 ] : 962 ไรโบโซม หน่วยย่อย อาร์อาร์เอ็นเอ โปรตีนอาร์ ยุค 70 50S 23S (2904 นิวคลี โอไทด์ ) 31 5S (120 นิวคลีโอไทด์) 30S 16S (1542 นิวคลีโอไทด์) 21
การติดฉลากความสัมพันธ์สำหรับตำแหน่งการจับ tRNA บน ไรโบโซม ของ E. coliทำให้สามารถระบุโปรตีนตำแหน่ง A และ P ที่น่าจะเกี่ยวข้องกับกิจกรรมเปปทิดิลทรานสเฟอเรสได้[ 5 ]โปรตีนที่ติดฉลากคือ L27, L14, L15, L16, L2 อย่างน้อย L27 ตั้งอยู่ที่ตำแหน่งผู้ให้ ดังที่แสดงโดย E. Collatz และ AP Czernilofsky [ 22 ] [ 23 ]การวิจัยเพิ่มเติมแสดงให้เห็นว่าโปรตีน S1 และ S21 ที่เกี่ยวข้องกับปลาย 3′ ของ RNA ไรโบโซม 16S มีส่วนเกี่ยวข้องในการเริ่มต้นการแปล[ 24 ]
อาร์เคีย
โดยทั่วไปแล้ว ไรโบโซม ของอาร์ เคีย จะถูกระบุว่ามีขนาดใกล้เคียงกับไรโบโซมของแบคทีเรีย โดยเป็นไรโบโซม 70S ที่ประกอบด้วยซับยูนิตขนาดใหญ่ 50S และซับยูนิตขนาดเล็ก 30S [ 25 ]สายโซ่ rRNA มักถูกเรียกว่า 16S, 23S และ 5S เช่นกัน แม้ว่าจะมีแหล่งข้อมูลน้อยมาก (หรืออาจไม่มีเลย) ที่วัดค่าสัมประสิทธิ์การตกตะกอนของพวกมันอย่างแท้จริง[ 26 ]อย่างไรก็ตามในระดับลำดับและโครงสร้าง พวกมันมีความใกล้เคียงกับของยูคาริโอตมากกว่าของแบคทีเรีย โปรตีนไรโบโซมพิเศษทุกตัวที่อาร์เคียมีเมื่อเทียบกับแบคทีเรียจะมีคู่ที่เทียบเท่าในยูคาริโอต ในขณะที่ไม่มีความสัมพันธ์ดังกล่าวระหว่างอาร์เคียและแบคทีเรีย[ 27 ] [ 28 ] [ 29 ]
ไรโบโซมของPyrococcus furiosus (อาร์เคีย) [ 30 ] [ 31 ] ไรโบโซม หน่วยย่อย อาร์อาร์เอ็นเอ โปรตีนอาร์ ยุค 70 50S 23S (3049 นิวคลี โอไทด์ ) 42 5S (120 นิวคลีโอไทด์) 30S 16S (1495 นิวคลีโอไทด์) 26
ยูคาริโอต
เซลล์ยูคาริโอติกมีไรโบโซม 80Sอยู่ในไซโตซอล โดยแต่ละไรโบโซมประกอบด้วย ซับยูนิต ขนาดเล็ก (40S)และขนาดใหญ่ (60S)ซับยูนิต 40S ประกอบด้วยRNA 18S (1900 นิวคลีโอไทด์) และโปรตีน 33 ชนิด[ 32 ] [ 33 ]ซับยูนิตขนาดใหญ่ประกอบด้วยRNA 5S (120 นิวคลีโอไทด์), RNA 28S (4700 นิวคลีโอไทด์), RNA 5.8S (160 นิวคลีโอไทด์) และโปรตีน 49 ชนิด[ 14 ] [ 32 ] [ 34 ]
ยูคาริโอตไซโตซิลิกไรโบโซม ( R. norvegicus ) [ 21 ] : 65 ไรโบโซม หน่วยย่อย อาร์อาร์เอ็นเอ โปรตีนอาร์ ยุค 80 60S 28S (4718 นิวคลีโอไทด์) 49 5.8 วินาที (160 นิวคลีโอไทด์) 5S (120 นิวคลีโอไทด์) 40S 18S (1874 nt) 33
ในปี พ.ศ. 2520 Czernilofsky ได้ตีพิมพ์งานวิจัยที่ใช้การติดฉลากความสัมพันธ์เพื่อระบุตำแหน่งการจับ tRNA บนไรโบโซมของตับหนู โปรตีนหลายชนิด รวมถึง L32/33, L36, L21, L23, L28/29 และ L13 มีส่วนเกี่ยวข้องว่าอยู่ที่หรือใกล้กับศูนย์กลางการถ่ายโอนเปปไทด์[ 35 ]
พลาสโตริโบโซมและไมโตริโบโซม
ในยูคาริโอต ไรโบโซมมีอยู่ในไมโทคอนเดรีย (บางครั้งเรียกว่าไมโทไรโบโซม ) และในพลาสติดเช่นคลอโรพลาสต์ (เรียกอีกอย่างว่า พลาสโทไรโบโซม) พวกมันประกอบด้วยหน่วยย่อยขนาดใหญ่และขนาดเล็กที่เชื่อมต่อกันด้วยโปรตีนเป็นอนุภาค 70S หนึ่งอนุภาค[ 14 ]ไรโบโซมเหล่านี้คล้ายกับของแบคทีเรีย และเชื่อกันว่าออร์แกเนลล์เหล่านี้มีต้นกำเนิดมาจากแบคทีเรียแบบพึ่งพาอาศัยกัน [ 14 ] ในบรรดาทั้งสอง ไรโบโซมของคลอโรพลาสต์มีความใกล้เคียงกับของแบคทีเรียมากกว่าไรโบโซมของไมโทคอนเดรีย ชิ้นส่วนของ RNA ไรโบโซมจำนวนมากในไมโทคอนเดรียสั้นลง และในกรณีของ5S rRNAจะถูกแทนที่ด้วยโครงสร้างอื่นในสัตว์และเชื้อรา[ 36 ]โดยเฉพาะอย่างยิ่งLeishmania tarentolaeมีชุด rRNA ไมโทคอนเดรียที่ลดขนาดลง[ 37 ]ในทางตรงกันข้าม ไมโตริโบโซมของพืชมีทั้ง rRNA ที่ขยายออกและโปรตีนเพิ่มเติมเมื่อเปรียบเทียบกับแบคทีเรีย โดยเฉพาะอย่างยิ่งโปรตีนที่มีการทำซ้ำของเพนตาไตรโคพีไทด์จำนวนมาก[ 38 ]
สาหร่ายคริปโตโมนาดและคลอราแรคนิโอไฟต์อาจมีนิวคลีโอเมอร์ฟที่มีลักษณะคล้ายนิวเคลียสยูคาริโอตที่เหลืออยู่[ 39 ]ไรโบโซมยูคาริโอต 80S อาจมีอยู่ในช่องที่มีนิวคลีโอเมอร์ฟ
การใช้ประโยชน์จากความแตกต่าง
ความแตกต่างระหว่างไรโบโซมของแบคทีเรียและยูคาริโอตถูกนำมาใช้ประโยชน์โดยนักเคมีเภสัชกรรมในการสร้างยาปฏิชีวนะที่สามารถทำลายการติดเชื้อแบคทีเรียได้โดยไม่ทำอันตรายต่อเซลล์ของผู้ติดเชื้อ เนื่องจากความแตกต่างในโครงสร้าง ไรโบโซม 70S ของแบคทีเรียจึงอ่อนแอต่อยาปฏิชีวนะเหล่านี้ ในขณะที่ไรโบโซม 80S ของยูคาริโอตไม่เป็นเช่นนั้น[ 40 ]แม้ว่าไมโทคอน เดรีย จะมีไรโบโซมที่คล้ายกับของแบคทีเรีย แต่ไมโทคอนเดรียก็ไม่ได้รับผลกระทบจากยาปฏิชีวนะเหล่านี้ เนื่องจากมีเยื่อหุ้มสองชั้นล้อมรอบซึ่งไม่ยอมให้ยาปฏิชีวนะเหล่านี้เข้าไปในออร์แกเนลล์ได้ง่าย[ 41 ]ตัวอย่างที่น่าสนใจคือยาปฏิชีวนะต้านมะเร็งคลอแรมเฟนิคอลซึ่งยับยั้งไรโบโซม 50S ของแบคทีเรียและไรโบโซม 50S ของไมโทคอนเดรียยูคาริโอต[ 42 ]อย่างไรก็ตาม ไรโบโซมในคลอโรพลาสต์นั้นแตกต่างกัน: ความต้านทานต่อยาปฏิชีวนะในโปรตีนไรโบโซมของคลอโรพลาสต์เป็นลักษณะที่ต้องนำมาใช้เป็นเครื่องหมายด้วยวิศวกรรมพันธุกรรม[ 43 ]
คุณสมบัติทั่วไป
ไรโบโซมชนิดต่างๆ มีโครงสร้างหลักที่คล้ายคลึงกันมาก แม้จะมีขนาดแตกต่างกันมากก็ตาม อาร์เอ็นเอส่วนใหญ่มีการจัดระเบียบอย่างสูงเป็นโครงสร้างตติยภูมิ หลายรูปแบบ เช่นพсевдокнотที่แสดงการเรียงซ้อนแบบแกนร่วม อาร์ เอ็นเอ ส่วนเกินในไรโบโซมขนาดใหญ่จะอยู่ในรูปของการแทรกที่ยาวต่อเนื่องกัน[ 44 ]ทำให้เกิดลูปออกจากโครงสร้างหลักโดยไม่รบกวนหรือเปลี่ยนแปลงโครงสร้าง[ 14 ]กิจกรรมเร่งปฏิกิริยาทั้งหมดของไรโบโซมดำเนินการโดยอาร์เอ็นเอโปรตีนจะอยู่บนพื้นผิวและดูเหมือนจะช่วยรักษาเสถียรภาพของโครงสร้าง[ 14 ]
โครงสร้างความละเอียดสูง

โครงสร้างโมเลกุลทั่วไปของไรโบโซมเป็นที่รู้จักมาตั้งแต่ช่วงต้นทศวรรษ 1970 ในช่วงต้นทศวรรษ 2000 โครงสร้าง ดังกล่าวได้รับการกำหนดที่ความละเอียดสูงถึงระดับไม่กี่อังสตรอม[ 46 ]
เอกสารฉบับแรกที่ให้โครงสร้างของไรโบโซมที่ความละเอียดระดับอะตอมได้รับการตีพิมพ์เกือบพร้อมกันในช่วงปลายปี 2000 หน่วยย่อย 50S (โปรคาริโอตขนาดใหญ่) ได้รับการกำหนดจากอาร์เคียHaloarcula marismortui [ 45 ]และแบคทีเรียDeinococcus radioduransและโครงสร้างของหน่วยย่อย 30S ได้รับการกำหนดจากแบคทีเรียThermus thermophilus [ 20 ] [ 47 ] การศึกษาโครงสร้างเหล่านี้ได้รับรางวัลโนเบลสาขาเคมีในปี 2009 ในเดือนพฤษภาคม 2001 พิกัดเหล่านี้ถูกนำมาใช้เพื่อสร้าง อนุภาค T. thermophilus 70S ทั้งหมดขึ้นใหม่ที่ ความละเอียด 5.5 Å [ 48 ]
ในเดือนพฤศจิกายน พ.ศ. 2548 มีการตีพิมพ์เอกสารสองฉบับที่มีโครงสร้างของไรโบโซม 70S ของ Escherichia coli โดยโครงสร้างของไรโบโซมที่ว่างเปล่าได้รับการกำหนดที่ความละเอียด 3.5 Å โดยใช้การตกผลึกด้วยรังสีเอกซ์ [ 49 ] จากนั้นอีกสองสัปดาห์ต่อมา โครงสร้างที่ได้จากกล้องจุลทรรศน์อิเล็กตรอนแบบไครโอถูกตีพิมพ์[ 50 ]ซึ่งแสดงให้เห็นไรโบโซมที่ความละเอียด 11–15 Åในขณะที่กำลังส่งสายโปรตีนที่สังเคราะห์ขึ้นใหม่เข้าไปในช่องทางนำโปรตีน
โครงสร้างอะตอมแรกของไรโบโซมที่ซับซ้อนกับ โมเลกุล tRNAและmRNAได้รับการแก้ไขโดยใช้การตกผลึกด้วยรังสีเอกซ์โดยสองกลุ่มอย่างอิสระ ที่ความละเอียด 2.8 Å [ 51 ]และ 3.7 Å [ 52 ] โครงสร้างเหล่า นี้ทำให้สามารถมองเห็นรายละเอียดของการโต้ตอบของ ไรโบโซม Thermus thermophilusกับmRNAและกับtRNAที่จับอยู่ที่ไซต์ไรโบโซมแบบคลาสสิก การโต้ตอบของไรโบโซมกับ mRNA ยาวที่มีลำดับ Shine-Dalgarno ได้รับการแสดงให้เห็นในเวลาต่อมาไม่นานที่ ความละเอียด4.5–5.5 Å [ 53 ]ในปี 2023 การศึกษาด้วยกล้องจุลทรรศน์อิเล็กตรอนแบบไครโอรายงานโครงสร้าง 1.55 Å ของ ไรโบโซม Escherichia coli 70S ในสถานะการแปล ซึ่งให้รายละเอียดใกล้เคียงอะตอมของการดัดแปลง rRNA การโต้ตอบระหว่าง tRNA-mRNA และการประสานงานของไอออน แผนที่ความละเอียดสูงช่วยให้สามารถระบุตำแหน่งโพลีมอร์ฟิซึมของไรโบโซมและมองเห็นสถานะไฮบริดไคเมอริกชั่วคราวที่เกี่ยวข้องกับการเคลื่อนย้าย tRNA ที่ความละเอียดประมาณ 2 Å ผลการค้นพบเหล่านี้ช่วยปรับปรุงความเข้าใจเชิงโครงสร้างของบริเวณการทำงานของไรโบโซมและให้ข้อมูลเชิงลึกที่มีค่าสำหรับการออกแบบยาปฏิชีวนะ[ 54 ]
ในปี 2011 โครงสร้างอะตอมที่สมบูรณ์แรกของไรโบโซม 80S ของยูคาริโอตจากยีสต์Saccharomyces cerevisiae ได้รับจากการศึกษาผลึกศาสตร์[ 32 ]แบบจำลองนี้เผยให้เห็นสถาปัตยกรรมขององค์ประกอบเฉพาะของยูคาริโอตและการโต้ตอบกับแกนกลางที่อนุรักษ์ไว้ทั่วโลก ในขณะเดียวกัน แบบจำลองที่สมบูรณ์ของโครงสร้างไรโบโซม 40S ของยูคาริโอตในTetrahymena thermophilaได้รับการตีพิมพ์และอธิบายโครงสร้างของหน่วยย่อย 40Sรวมถึงรายละเอียดมากมายเกี่ยวกับการโต้ตอบของหน่วยย่อย 40S กับeIF1ในระหว่าง การเริ่มต้น การแปล[ 33 ]ในทำนองเดียวกัน โครงสร้าง ของหน่วยย่อย 60S ของยูคาริโอต ก็ได้รับการกำหนดจากTetrahymena thermophilaในเชิงซ้อนกับeIF6ด้วย[ 34 ]นอกจากนี้ โครงสร้าง cryo-EM ความละเอียดสูงของไรโบโซม 80S ยูคาริโอตที่ชอบอุณหภูมิสูงซึ่งถูกบันทึกในสองสถานะการหมุนที่ความละเอียด ~2.9 Å และ ~3.0 Å เผยให้เห็นรายละเอียดระดับอะตอมของกลไกการเคลื่อนย้ายยูคาริโอตและพลวัตเชิงโครงสร้างของ eEF2 ระหว่างการไฮโดรไลซิส GTP [ 55 ]
การทำงาน
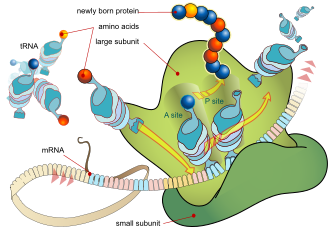
ไรโบโซมเป็นอนุภาคขนาดเล็กที่ประกอบด้วย RNA และโปรตีนที่เกี่ยวข้อง ทำหน้าที่สังเคราะห์โปรตีน โปรตีนมีความจำเป็นต่อการทำงานหลายอย่างของเซลล์ เช่น การซ่อมแซมความเสียหาย หรือการควบคุมกระบวนการทางเคมี ไรโบโซมอาจลอยอยู่ได้อย่างอิสระในไซโตพลาสซึม หรืออาจเกาะติดอยู่กับเอนโดพลาสมิกเรติคูลัมแบบหยาบหน้าที่หลักของไรโบโซมคือการแปลงรหัสพันธุกรรมให้เป็นลำดับกรดอะมิโน และสร้างพอลิเมอร์โปรตีนจากโมโนเมอร์ของกรดอะมิโน
ไรโบโซมทำหน้าที่เป็นตัวเร่งปฏิกิริยาในกระบวนการทางชีววิทยาที่สำคัญอย่างยิ่งสองกระบวนการ ได้แก่ การถ่ายโอนเปปไทด์และการไฮโดรไลซิสเปปไทด์
โดยสรุปแล้ว ไรโบโซมมีหน้าที่หลักสองประการ ได้แก่ การถอดรหัสข้อความ และการสร้างพันธะเปปไทด์ หน้าที่ทั้งสองนี้อยู่ในหน่วยย่อยของไรโบโซม แต่ละหน่วยย่อยประกอบด้วย rRNA หนึ่งตัวหรือมากกว่า และโปรตีน r จำนวนมาก หน่วยย่อยขนาดเล็ก (30S ในแบคทีเรียและอาร์เคีย, 40S ในยูคาริโอต) มีหน้าที่ในการถอดรหัส ในขณะที่หน่วยย่อยขนาดใหญ่ (50S ในแบคทีเรียและอาร์เคีย, 60S ในยูคาริโอต) ทำหน้าที่เร่งปฏิกิริยาการสร้างพันธะเปปไทด์ ซึ่งเรียกว่ากิจกรรมเปปทิดิลทรานสเฟอเรส หน่วยย่อยขนาดเล็กของแบคทีเรีย (และอาร์เคีย) ประกอบด้วย rRNA 16S และโปรตีน r 21 ตัว ( Escherichia coli ) ในขณะที่หน่วยย่อยขนาดเล็กของยูคาริโอตประกอบด้วย rRNA 18S และโปรตีน r 32 ตัว (Saccharomyces cerevisiae แม้ว่าจำนวนจะแตกต่างกันไปในแต่ละสายพันธุ์) หน่วยย่อยขนาดใหญ่ของแบคทีเรียประกอบด้วย rRNA 5S และ 23S และโปรตีน r 34 ตัว ( E. coli ) ในขณะที่หน่วยย่อยขนาดใหญ่ของยูคาริโอตประกอบด้วย rRNA 5S, 5.8S และ 25S/28S และโปรตีน r 46 ตัว ( S. cerevisiae ; อีกครั้ง จำนวนที่แน่นอนจะแตกต่างกันไปในแต่ละสายพันธุ์) [ 56 ]
การแปล
ไรโบโซมเป็นสถานที่ทำงานของการสังเคราะห์โปรตีนซึ่งเป็นกระบวนการแปลmRNAเป็นโปรตีน mRNA ประกอบด้วยชุดของโคดอนซึ่งไรโบโซมจะถอดรหัสเพื่อสร้างโปรตีน โดยใช้ mRNA เป็นแม่แบบ ไรโบโซมจะเคลื่อนที่ไปตามแต่ละโคดอน (3 นิวคลีโอไทด์ ) ของ mRNA และจับคู่กับกรดอะมิโนที่เหมาะสมซึ่งจัดหาโดยอะมิโนเอซิล-tRNAอะมิโนเอซิล-tRNA มีแอนติโคดอน ที่เสริมกัน ที่ปลายด้านหนึ่งและกรดอะมิโนที่เหมาะสมที่ปลายอีกด้านหนึ่ง เพื่อการจดจำ tRNA ที่เหมาะสมอย่างรวดเร็วและแม่นยำ ไรโบโซมจะใช้การเปลี่ยนแปลงโครงสร้างขนาดใหญ่ ( การตรวจสอบความถูกต้องเชิงโครงสร้าง ) [ 57 ]หน่วยย่อยไรโบโซมขนาดเล็ก ซึ่งโดยทั่วไปจะจับกับอะมิโนเอซิล-tRNA ที่มีกรดอะมิโนตัวแรกคือเม ไทโอนีน จะจับกับโคดอน AUG บน mRNA และดึงดูดหน่วยย่อยไรโบโซมขนาดใหญ่ ไรโบโซมมีตำแหน่งการจับ RNA สามตำแหน่ง ได้แก่ A, P และ E ตำแหน่ง Aจับกับอะมิโนเอซิล-tRNA หรือปัจจัยปลดปล่อยการยุติ[ 58 ] [ 59 ]ตำแหน่ง Pจับกับเปปทิดิล-tRNA (tRNA ที่จับกับสายโพลีเปปไทด์) และตำแหน่ง E (ทางออก) จับกับ tRNA อิสระ การสังเคราะห์โปรตีนเริ่มต้นที่รหัสเริ่มต้น AUG ใกล้ปลาย 5′ของ mRNA mRNA จับกับตำแหน่ง P ของไรโบโซมก่อน ไรโบโซมจดจำรหัสเริ่มต้นโดยใช้ลำดับ Shine-Dalgarnoของ mRNA ในโปรคาริโอตและกล่อง Kozakในยูคาริโอต
แม้ว่าการเร่งปฏิกิริยาของพันธะเปปไทด์ จะเกี่ยวข้องกับ ไฮดรอกซิล C2 ของอะดีโนซีน ในตำแหน่ง P ของ RNA ในกลไกการเคลื่อนย้ายโปรตอน แต่ขั้นตอนอื่นๆ ในการสังเคราะห์โปรตีน (เช่น การเคลื่อนย้าย) เกิดจากการเปลี่ยนแปลงในโครงสร้างของโปรตีน เนื่องจากแกนเร่งปฏิกิริยา ของพวกมัน ทำจาก RNA ไรโบโซมจึงถูกจัดประเภทเป็น " ไรโบไซม์ " [ 60 ]และเชื่อกันว่าพวกมันอาจเป็นส่วนที่เหลือของโลก RNA [ 61 ]

ในภาพที่ 5 ทั้งสองหน่วยย่อยของไรโบโซม ( ขนาดเล็กและขนาดใหญ่ ) จะประกอบกันที่โคดอนเริ่มต้น (ทางด้าน 5' ของmRNA ) ไรโบโซมจะใช้tRNAที่ตรงกับโคดอนปัจจุบัน (ไตรเพล็ต) บน mRNA เพื่อเติมกรดอะมิโนลงในสายโพลีเปปไทด์ กระบวนการนี้จะเกิดขึ้นกับไตรเพล็ตแต่ละอันบน mRNA ในขณะที่ไรโบโซมเคลื่อนที่ไปทางด้าน 3' ของ mRNA โดยปกติในเซลล์แบคทีเรีย ไรโบโซมหลายตัวจะทำงานพร้อมกันบน mRNA เดียวกัน ทำให้เกิดสิ่งที่เรียกว่าโพลีไรโบโซมหรือโพลีโซม
การพับตัวแบบโคทรานสเลชัน
เป็นที่ทราบกันดีว่าไรโบโซมมีส่วนร่วมอย่างแข็งขันในการพับโปรตีน[ 62 ] [ 63 ]โครงสร้างที่ได้ด้วยวิธีนี้มักจะเหมือนกับโครงสร้างที่ได้ระหว่างการพับโปรตีนทางเคมี อย่างไรก็ตาม เส้นทางที่นำไปสู่ผลิตภัณฑ์สุดท้ายอาจแตกต่างกัน[ 64 ] [ 65 ]ในบางกรณี ไรโบโซมมีความสำคัญอย่างยิ่งในการสร้างโปรตีนที่มีรูปแบบการทำงาน ตัวอย่างเช่น หนึ่งในกลไกที่เป็นไปได้ของการพับโปรตีนที่มีปม ลึกนั้น อาศัยไรโบโซมในการผลักโซ่ผ่านห่วงที่ติดอยู่[ 66 ]
การเติมกรดอะมิโนที่ไม่ขึ้นกับการแปลรหัส
การมีอยู่ของโปรตีนควบคุมคุณภาพของไรโบโซม Rqc2 เกี่ยวข้องกับการยืดตัวของโปรตีนที่ไม่ขึ้นกับ mRNA [ 67 ] [ 68 ] การยืดตัวนี้เป็นผลมาจากการเพิ่ม หางCATของไรโบโซม (ผ่าน tRNA ที่นำมาโดย Rqc2) : ไรโบโซมจะขยายปลายCของโปรตีนที่หยุดชะงักด้วยลำดับแบบสุ่มที่ไม่ขึ้นกับการแปลของอะลานีนและทรีโอนีน[ 69 ] [ 70 ]
ตำแหน่งของไรโบโซม
ไรโบโซมถูกจัดประเภทเป็น "อิสระ" หรือ "ยึดติดกับเยื่อหุ้มเซลล์" [ 71 ] ไรโบโซมอิสระและไรโบโซมที่ยึดติดกับเยื่อหุ้มเซลล์แตกต่างกันเพียงแค่การกระจายตัวในเชิงพื้นที่เท่านั้น โครงสร้างของพวกมันเหมือนกัน การที่ไรโบโซมอยู่ในสถานะอิสระหรือยึดติดกับเยื่อหุ้มเซลล์นั้นขึ้นอยู่กับการมีลำดับสัญญาณเป้าหมาย ERบนโปรตีนที่กำลังสังเคราะห์ ดังนั้นไรโบโซมแต่ละตัวอาจยึดติดกับเยื่อหุ้มเซลล์เมื่อมันกำลังสร้างโปรตีนชนิดหนึ่ง แต่เป็นอิสระในไซโตโซลเมื่อมันกำลังสร้างโปรตีนอีกชนิดหนึ่ง
บางครั้งไรโบโซมถูกเรียกว่าออร์แกเนลล์แต่การใช้คำว่าออร์แกเนลล์มักจำกัดอยู่เฉพาะการอธิบายส่วนประกอบย่อยของเซลล์ที่มีเยื่อฟอสโฟลิปิด ซึ่งไรโบโซมเป็นอนุภาคทั้งหมดจึงไม่มี ด้วยเหตุนี้ บางครั้งไรโบโซมจึงอาจถูกเรียกว่า "ออร์แกเนลล์ที่ไม่มีเยื่อหุ้ม" [ 72 ]
ไรโบโซมอิสระ
ไรโบโซมอิสระสามารถเคลื่อนที่ไปได้ทุกที่ในไซโตซอลแต่จะไม่สามารถเข้าไปในนิวเคลียสและออร์แกเนลล์อื่นๆ ของเซลล์ได้ โปรตีนที่เกิดขึ้นจากไรโบโซมอิสระจะถูกปล่อยออกมาในไซโตซอลและนำไปใช้ภายในเซลล์ เนื่องจากไซโตซอลมีกลูตาไธโอน ในความเข้มข้นสูง และเป็นสภาพแวดล้อมแบบรีดิวซ์โปรตีนที่มีพันธะไดซัลไฟด์ซึ่งเกิดจากหมู่ซิสเทอีนที่ถูกออกซิไดซ์ จึงไม่สามารถผลิตขึ้นภายในไซโตซอลได้
ไรโบโซมที่ยึดติดกับเยื่อหุ้มเซลล์
เมื่อไรโบโซมเริ่มสังเคราะห์โปรตีนที่จำเป็นในออร์แกเนลล์บางชนิด ไรโบโซมที่สร้างโปรตีนนี้สามารถ "ยึดติดกับเยื่อหุ้มเซลล์" ได้ ในเซลล์ยูคาริโอต สิ่งนี้เกิดขึ้นในบริเวณของเอนโดพลาสมิกเรติคูลัม (ER)ที่เรียกว่า "ER แบบหยาบ" สายโซ่โพลีเปปไทด์ที่ผลิตขึ้นใหม่จะถูกแทรกเข้าไปใน ER โดยตรงโดยไรโบโซมที่ทำการสังเคราะห์แบบเวกเตอร์จากนั้นจะถูกขนส่งไปยังปลายทางผ่านทางเส้นทางการหลั่งไรโบโซมที่ยึดติดมักจะผลิตโปรตีนที่ใช้ภายในเยื่อหุ้มพลาสมาหรือถูกขับออกจากเซลล์ผ่านทางเอ็กโซไซโทซิส[ 73 ]
ไบโอเจเนซิส
ในเซลล์แบคทีเรีย ไรโบโซมจะถูกสังเคราะห์ในไซโตพลาสซึมผ่านการถอดรหัสของโอเปรอน ยีนไรโบโซมหลายตัว ในยูคาริโอต กระบวนการนี้เกิดขึ้นทั้งในไซโตพลาสซึมของเซลล์และในนิวคลีโอลัสซึ่งเป็นบริเวณภายในนิวเคลียสของเซลล์กระบวนการประกอบเกี่ยวข้องกับการทำงานที่ประสานกันของโปรตีนมากกว่า 200 ชนิดในการสังเคราะห์และการประมวลผลของ rRNA ทั้งสี่ชนิด รวมถึงการประกอบ rRNA เหล่านั้นเข้ากับโปรตีนไรโบโซม[ 74 ]
ต้นทาง
ไรโบโซมอาจมีต้นกำเนิดมาจากโปรโตริโบโซม[ 75 ]ซึ่งอาจมีศูนย์ถ่ายโอนเปปไทด์ (PTC) ในโลกของ อาร์เอ็นเอ โดยปรากฏเป็นคอมเพล็กซ์ที่จำลองตัวเองได้ ซึ่งต่อมาได้พัฒนาความสามารถในการสังเคราะห์โปรตีนเมื่อกรดอะมิโนเริ่มปรากฏขึ้น[ 76 ]การศึกษาชี้ให้เห็นว่าไรโบโซมโบราณที่สร้างขึ้นจากrRNA เพียงอย่างเดียว อาจพัฒนาความสามารถในการสังเคราะห์พันธะเปปไทด์ได้[ 77 ] [ 78 ] [ 79 ] [ 80 ] [ 81 ]นอกจากนี้ หลักฐานยังชี้ให้เห็นอย่างชัดเจนว่าไรโบโซมโบราณเป็นคอมเพล็กซ์ที่จำลองตัวเองได้ โดยที่ rRNA ในไรโบโซมมีวัตถุประสงค์ด้านข้อมูล โครงสร้าง และเร่งปฏิกิริยา เนื่องจากสามารถเข้ารหัสtRNAและโปรตีนที่จำเป็นสำหรับการจำลองตัวเองของไรโบโซมได้[ 82 ]สิ่งมีชีวิตเซลล์สมมุติที่มี RNA ที่สามารถจำลองตัวเองได้แต่ไม่มี DNA เรียกว่าไรโบไซต์ (หรือไรโบเซลล์) [ 83 ] [ 84 ]
เมื่อกรดอะมิโนค่อยๆ ปรากฏขึ้นในโลกของ RNA ภายใต้สภาวะก่อนกำเนิดสิ่งมีชีวิต[ 85 ] [ 86 ]ปฏิสัมพันธ์ของกรดอะมิโนกับ RNA ที่เป็นตัวเร่งปฏิกิริยาจะเพิ่มทั้งขอบเขตและประสิทธิภาพของการทำงานของโมเลกุล RNA ที่เป็นตัวเร่งปฏิกิริยา[ 76 ]ดังนั้น แรงผลักดันสำหรับการวิวัฒนาการของไรโบโซมจากเครื่องจักรจำลองตัว เองโบราณ ไปสู่รูปแบบปัจจุบันที่เป็นเครื่องจักรการแปลอาจเป็นแรงกดดันในการคัดเลือกเพื่อรวมโปรตีนเข้ากับกลไกการจำลองตัวเองของไรโบโซม เพื่อเพิ่มความสามารถในการจำลองตัวเอง[ 82 ] [ 87 ] [ 88 ]
ไรโบโซมต่างชนิดกัน
ในปี พ.ศ. 2491 ฟรานซิส คริก ได้เสนอสมมติฐาน "หนึ่งยีน-หนึ่งไรโบโซม-หนึ่งโปรตีน" ซึ่งไรโบโซมแต่ละตัวจะบรรจุข้อมูลทางพันธุกรรมที่จำเป็นในการเข้ารหัสโปรตีนเพียงตัวเดียว แม้ว่าสมมติฐานนี้จะถูกปฏิเสธในเวลานั้น แต่จากการค้นพบโรคไรโบโซมผิดปกติชนิดแรก คือ โรคโลหิตจางไดมอนด์-แบล็กแฟน ในปี พ.ศ. 2542 ไรโบโซมได้เปลี่ยนจากเครื่องจักรโมเลกุลแบบพาสซีฟไปเป็นเครื่องจักรโมเลกุลขนาดใหญ่แบบไดนามิก[ 89 ] [ 90 ]ไรโบโซมมีองค์ประกอบที่แตกต่างกันระหว่างสายพันธุ์และแม้กระทั่งภายในเซลล์เดียวกัน ดังที่เห็นได้จากการมีอยู่ของไรโบโซมในไซโตพลาสซึมและไมโทคอนเดรียภายในเซลล์ยูคาริโอติกเดียวกัน นักวิจัยบางคนเสนอว่าความแตกต่างในองค์ประกอบของโปรตีนไรโบโซมในสัตว์เลี้ยงลูกด้วยนมมีความสำคัญต่อการควบคุมยีน กล่าวคือสมมติฐานไรโบโซมเฉพาะทาง[ 91 ] [ 92 ]อย่างไรก็ตาม สมมติฐานนี้เป็นที่ถกเถียงและเป็นหัวข้อของการวิจัยอย่างต่อเนื่อง[ 93 ] [ 94 ]
ความไม่สม่ำเสมอในองค์ประกอบของไรโบโซมได้รับการเสนอครั้งแรกว่ามีส่วนเกี่ยวข้องกับการควบคุมการแปลของการสังเคราะห์โปรตีนโดย Vince Mauro และGerald Edelman [ 95 ] พวกเขาเสนอสมมติฐานตัวกรองไรโบโซมเพื่ออธิบายหน้าที่การควบคุมของไรโบโซม หลักฐานชี้ให้เห็นว่าไรโบโซมเฉพาะที่เฉพาะเจาะจงกับประชากรเซลล์ที่แตกต่างกันอาจส่งผลต่อวิธีการแปลยีน[ 96 ]โปรตีนไรโบโซมบางชนิดแลกเปลี่ยนจากคอมเพล็กซ์ที่ประกอบขึ้นกับสำเนาในไซโตโซล[ 97 ]ซึ่งบ่งชี้ว่าโครงสร้างของ ไรโบโซม ในร่างกายสามารถเปลี่ยนแปลงได้โดยไม่ต้องสังเคราะห์ไรโบโซมใหม่ทั้งหมด
โปรตีนไรโบโซมบางชนิดมีความสำคัญอย่างยิ่งต่อการดำรงชีวิตของเซลล์ ในขณะที่บางชนิดไม่สำคัญ ในยีสต์ที่กำลังแตกหน่อโปรตีนไรโบโซม 14/78 ชนิดไม่จำเป็นต่อการเจริญเติบโต ในขณะที่ในมนุษย์นั้นขึ้นอยู่กับเซลล์ที่ศึกษา[ 98 ]รูปแบบอื่นๆ ของความแตกต่างกัน ได้แก่ การดัดแปลงหลังการแปลของโปรตีนไรโบโซม เช่น การอะเซทิเลชัน การเมทิลเลชัน และการฟอสโฟรีเลชัน[ 99 ] อาราบิโดปซิส [ 100 ] [ 101 ] [ 102 ] [ 103 ]ตำแหน่งการเข้าสู่ไรโบโซมภายในของไวรัส ( IRES ) อาจเป็นตัวกลางในการแปลโดยไรโบโซมที่มีองค์ประกอบแตกต่างกัน ตัวอย่างเช่น หน่วยไรโบโซม 40S ที่ไม่มีeS25ในยีสต์และเซลล์สัตว์เลี้ยงลูกด้วยนมไม่สามารถดึงดูดCrPV IGR IRESได้[ 104 ]
ความหลากหลายของการดัดแปลง RNA ไรโบโซมมีบทบาทสำคัญในการรักษาโครงสร้างและ/หรือการทำงาน และการดัดแปลง mRNA ส่วนใหญ่พบในบริเวณที่มีการอนุรักษ์สูง[ 105 ] [ 106 ]การดัดแปลง rRNA ที่พบบ่อยที่สุดคือpseudouridylationและ2'-O-methylationของไรโบส[ 107 ]
ดูเพิ่มเติม
ลิงก์ภายนอก
- คอมพิวเตอร์ในห้องปฏิบัติการจำลองการเคลื่อนที่ของไรโบโซม
- บทบาทของไรโบโซมโดย กเวน วี. ไชลด์ส คัดลอกมาไว้ที่นี่
- ไรโบโซมในโปรตีโอพีเดีย — สารานุกรมสามมิติแบบร่วมมือกันฟรีเกี่ยวกับโปรตีนและโมเลกุลอื่นๆ
- ตระกูลโปรตีนไรโบโซมใน ExPASy เก็บถาวรเมื่อ 30 เมษายน 2554 ที่Wayback Machine
- โมเลกุลประจำเดือนเก็บถาวรเมื่อ 27 ตุลาคม 2009 ที่Wayback Machine © RCSB Protein Data Bank :
- ไรโบโซมถูกเก็บถาวรเมื่อวันที่ 14 พฤศจิกายน 2010 ที่Wayback Machine
- ปัจจัยการยืดตัว (Elongation Factors) ถูกเก็บถาวรเมื่อวันที่ 16 มีนาคม 2011 ที่Wayback Machine
- พระราชวัง
- โครงสร้างสามมิติของไรโบโซมจากกล้องจุลทรรศน์อิเล็กตรอนในฐานข้อมูล EM Data Bank (EMDB)
 บทความนี้ได้นำเนื้อหาที่เป็นสาธารณสมบัติจากScience PrimerของNCBI มา ใช้ โดยเก็บถาวรจากต้นฉบับเมื่อวันที่ 8 ธันวาคม 2552
บทความนี้ได้นำเนื้อหาที่เป็นสาธารณสมบัติจากScience PrimerของNCBI มา ใช้ โดยเก็บถาวรจากต้นฉบับเมื่อวันที่ 8 ธันวาคม 2552
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ไรโบโซม
ไรโบโซม ( / ˈ r aɪ b ə z oʊ m , - s oʊ m / ) คืออนุภาคไรโบนิวคลีโอโปรตีนที่พบในเซลล์ ทุกชนิด
ภาพรวม
ลำดับของ RNA ที่เข้ารหัสลำดับของ กรดอะมิโน ในโปรตีนจะถูกถอดรหัสเป็น สาย mRNA ( messenger RNA ) ไรโบโซมจะจับกับโมเลกุล mRNA และใช้ลำดับนิว คลีโอไทด์ ของ RNA เพื่อกำหนดลำดับของกรดอะมิโนที่จำเป็นในการสร้างโปรตีน กรดอะมิโนจะถูกเลือกและนำไปยังไรโบโซมโดย โมเลกุล...
การค้นพบ
ไรโบโซมถูกสังเกตครั้งแรกในช่วงกลางทศวรรษ 1950 ในรูปของอนุภาคหรือเม็ดที่มีความหนาแน่นสูงโดยนักชีววิทยาเซลล์ ชาวโรมาเนีย-อเมริกัน George Emil Palade โดยใช้กล้องจุลทรรศน์ อิเล็กตรอน [ 8 ] ในตอนแรกเรียกว่า เม็ด Palade เนื่องจากโครงสร้างที่เป็นเม็ด [ 9 ] คำว่า...
โครงสร้าง
ไรโบโซมเป็น อนุภาคไรโบนิวคลีโอโปรตีน ที่สร้างขึ้นจาก สารประกอบ ของไรโบโซมอาร์เอ็นเอและโปรตีน จัดเรียงเป็นสองหน่วยย่อยไรโบโซม หน่วยหนึ่งใหญ่และอีกหน่วยหนึ่งเล็ก ไรโบโซมเป็น เครื่องจักรโมเลกุล ที่ซับซ้อนซึ่งมีอยู่ใน เซลล์ ทั้งหมดทั้ง โปรคาริโอต และ ยู คาริโอต...