อ่าน 17 นาที
แอกซอน
แอกซอน (จากภาษากรีกโบราณ : ἄξων , โรมัน : áxōn , แปลตรงตัวว่า ' แกน' ; เรียกอีกอย่างว่าเส้นใยประสาทหรือไฟเบอร์ ) คือ...
แอกซอน
| แอกซอน | |
|---|---|
 แอกซอนของเซลล์ประสาทหลายขั้ว | |
| ตัวระบุ | |
| เมช | D001369 |
| เอฟเอ็มเอ | 67308 |
| ศัพท์ทางกายวิภาคศาสตร์ | |
แอกซอน (จากภาษากรีกโบราณ : ἄξων , โรมัน : áxōn , แปลตรงตัวว่า ' แกน' ; เรียกอีกอย่างว่าเส้นใยประสาทหรือไฟเบอร์ ) คือ ส่วนที่ยื่นออกมาเป็นเส้นยาวและเรียวของเซลล์ประสาทหรือนิวรอนที่พบในสัตว์ ส่วนใหญ่ ซึ่งโดยทั่วไปจะนำกระแสไฟฟ้าที่เรียกว่าศักย์ไฟฟ้ากระทำ ( action potential)ออกจากตัวเซลล์ประสาทหน้าที่ของแอกซอนคือการส่งข้อมูลไปยังนิวรอน กล้ามเนื้อ และต่อมต่างๆ ใน นิวรอน รับความรู้สึก บางชนิด ( นิวรอน แบบซูโดอูนิโพ ลาร์ ) เช่น นิวรอนที่รับสัมผัสและความอบอุ่น แอกซอนจะเรียกว่าเส้นใยประสาทนำเข้า (afferent nerve fiber ) และกระแสไฟฟ้าจะเดินทางไปตามเส้นใยเหล่านี้จากส่วนปลาย ไปยังตัวเซลล์ และจากตัวเซลล์ไปยังไขสันหลังตามกิ่งก้านอีกกิ่งหนึ่งของแอกซอนเดียวกัน การทำงานผิดปกติของแอกซอนอาจเป็นสาเหตุของ ความผิดปกติทางระบบประสาทที่ถ่ายทอดทางพันธุกรรมและที่เกิดขึ้นภายหลังหลายอย่าง ซึ่งส่งผลกระทบ ต่อทั้ง นิวรอน ส่วนปลายและส่วนกลางเส้นใยประสาทแบ่งออกเป็นสามประเภท ได้แก่เส้นใยประสาทกลุ่ม A เส้นใยประสาท กลุ่ม Bและ เส้นใยประสาท กลุ่ม Cกลุ่ม A และ B มีปลอกไมอีลินหุ้มส่วนกลุ่ม C ไม่มีปลอกไมอีลินหุ้ม กลุ่มเหล่านี้รวมถึงทั้งเส้นใยประสาทรับความรู้สึกและเส้นใยประสาทสั่งการ อีกการจำแนกประเภทหนึ่งแบ่งเฉพาะเส้นใยประสาทรับความรู้สึกออกเป็นสี่ประเภท ได้แก่ ประเภทที่ 1 ประเภทที่ 2 ประเภทที่ 3 และประเภทที่ 4
แอกซอนเป็นส่วนยื่นของไซโตพลาสซึมจากตัวเซลล์ของเซลล์ประสาทอยู่ 2 ชนิด อีกชนิดหนึ่งคือเดนไดรต์แอกซอนแตกต่างจากเดนไดรต์ด้วยลักษณะหลายประการ ได้แก่ รูปร่าง (เดนไดรต์มักจะเรียวลง ในขณะที่แอกซอนมักจะมีรัศมีคงที่) ความยาว (เดนไดรต์จำกัดอยู่ในบริเวณเล็กๆ รอบตัวเซลล์ ในขณะที่แอกซอนอาจยาวกว่ามาก) และหน้าที่ (เดนไดรต์รับสัญญาณ ในขณะที่แอกซอนส่งสัญญาณ) เซลล์ประสาทบางชนิดไม่มีแอกซอนและส่งสัญญาณจากเดนไดรต์ ในบางชนิด แอกซอนสามารถงอกออกมาจากเดนไดรต์ที่เรียกว่าเดนไดรต์ที่นำแอกซอน[ 1 ]ไม่มีเซลล์ประสาทใดมีแอกซอนมากกว่าหนึ่งเส้น อย่างไรก็ตาม ในสัตว์ไม่มีกระดูกสันหลังเช่นแมลงหรือปลิงแอกซอนบางครั้งประกอบด้วยหลายส่วนที่ทำงานแยกจากกันมากหรือน้อย[ 2 ]
แอกซอนถูกหุ้มด้วยเยื่อหุ้มเซลล์ที่เรียกว่าแอกโซเลมมา ส่วนไซโตพ ลาซึมภายในแอกซอน เรียกว่า แอกโซ พลา ซึม แอกซอนส่วนใหญ่แตกแขนงออกไป ในบางกรณีอาจแตกแขนงอย่างมากมาย ปลายแขนงของแอกซอนเรียกว่าเทโลเดนเดรีย ปลายที่บวมของเทโลเดนเดรียเรียกว่าปลายแอกซอนหรือ ปลายเท้า ซึ่งจะเชื่อมต่อกับเดนไดรต์หรือตัวเซลล์ของเซลล์ประสาทอื่นเพื่อสร้าง การเชื่อมต่อแบบ ไซแนปส์โดยปกติแอกซอนจะสัมผัสกับเซลล์ประสาทอื่นที่จุดเชื่อมต่อที่เรียกว่าไซแนปส์แต่ก็สามารถสัมผัสกับเซลล์กล้ามเนื้อหรือเซลล์ต่อมได้เช่นกัน ในบางกรณี แอกซอนของเซลล์ประสาทหนึ่งอาจสร้างไซแนปส์กับเดนไดรต์ของเซลล์ประสาทเดียวกัน ทำให้เกิดออโตไซแนปส์ ที่ไซแนปส์เยื่อหุ้มของแอกซอนจะอยู่ติดกับเยื่อหุ้มของเซลล์เป้าหมายอย่างใกล้ชิด และโครงสร้างโมเลกุลพิเศษจะทำหน้าที่ส่งสัญญาณไฟฟ้าหรือสัญญาณไฟฟ้าเคมีข้ามช่องว่าง จุดเชื่อมต่อไซแนปส์บางจุดปรากฏขึ้นตามความยาวของแอกซอนขณะที่มันทอดยาวออกไป สิ่งเหล่านี้เรียกว่าปุ่มประสาทแบบผ่าน ("ปุ่มประสาทแบบผ่าน") และอาจมีเป็นร้อยหรือหลายพันปุ่มตามแนวแกนแอกซอนเดียว[ 3 ]ซินแนปส์อื่นๆ ปรากฏเป็นเทอร์มินัลที่ปลายกิ่งของแอกซอน
แอกซอนเดี่ยว เมื่อรวมกิ่งก้านทั้งหมดเข้าด้วยกัน สามารถกำหนดเป้าหมายไปยังส่วนต่างๆ ของสมองได้หลายส่วน และสร้างปลายประสาทไซแนปส์ได้หลายพันแห่ง กลุ่มของแอกซอนก่อให้เกิดเส้นทางประสาทในระบบประสาทส่วนกลาง [ 4 ]และกลุ่มเส้นใยประสาทในระบบประสาทส่วนปลายในสัตว์เลี้ยงลูกด้วยนมที่มี รก เส้นทาง เนื้อขาว ที่ ใหญ่ที่สุดในสมองคือคอร์ปัสคัลโลซัมซึ่งประกอบด้วยแอกซอนประมาณ 200 ล้านเส้นในสมองของมนุษย์[ 4 ]
กายวิภาคศาสตร์
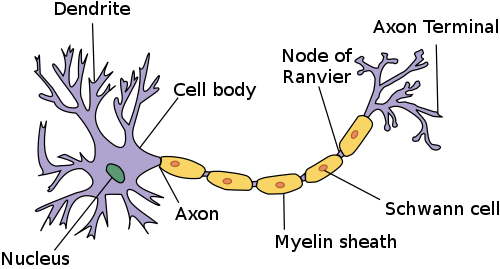

แอกซอนเป็นเส้นทางการส่งสัญญาณหลักของระบบประสาทและเมื่อรวมกันเป็นมัดจะเกิดเป็นเส้นประสาทในระบบประสาทส่วนปลาย หรือเป็นเส้นใยประสาทในระบบประสาทส่วนกลาง (CNS) แอกซอนบางเส้นอาจยาวได้ถึงหนึ่งเมตรหรือมากกว่านั้น ในขณะที่บางเส้นอาจยาวเพียงหนึ่งมิลลิเมตร แอกซอนที่ยาวที่สุดในร่างกายมนุษย์คือแอกซอนของเส้นประสาทไซแอติกซึ่งทอดยาวจากฐานของไขสันหลังไปยังนิ้วหัวแม่เท้าแต่ละข้าง เส้นผ่านศูนย์กลางของแอกซอนก็แตกต่างกันไป แอกซอนส่วนใหญ่มีขนาดเล็กมาก (โดยทั่วไปประมาณหนึ่งไมโครเมตร (μm)) แอกซอนที่ใหญ่ที่สุดในสัตว์เลี้ยงลูกด้วยนมอาจมีเส้นผ่านศูนย์กลางได้ถึง 20 μm แอกซอนยักษ์ของปลาหมึกซึ่งมีความเชี่ยวชาญในการส่งสัญญาณอย่างรวดเร็ว มีเส้นผ่านศูนย์กลางเกือบ 1 มิลลิเมตร ขนาดเท่ากับไส้ดินสอขนาดเล็ก จำนวนของเทโลเดนเดรีย (โครงสร้างที่แตกแขนงที่ปลายแอกซอน) ก็อาจแตกต่างกันไปในแต่ละเส้นใยประสาทด้วย โดยทั่วไปแล้ว แอกซอนในระบบประสาทส่วนกลางจะมีเทโลเดนเดรียหลายอัน และมีจุดเชื่อมต่อไซแนปส์จำนวนมาก ในทางตรงกันข้าม แอกซอน ของเซลล์เม็ดเล็กในสมองส่วนซีรีเบลลัมมีลักษณะเฉพาะคือมีกิ่งก้านรูปตัว T เพียงกิ่งเดียว ซึ่ง มี เส้นใยขนานสองเส้นยื่นออกมา การแตกแขนงที่ซับซ้อน นี้ช่วยให้สามารถส่งข้อความไปยัง เซลล์ประสาทเป้าหมายจำนวนมากพร้อมกันในบริเวณเดียวของสมองได้
ในระบบประสาทมีแอกซอนอยู่ 2 ประเภท ได้แก่ แอกซอนที่มีไมอีลินหุ้มและแอกซอนที่ไม่มีไม อีลินหุ้ม [ 5 ]ไมอีลินเป็นชั้นของสารฉนวนไขมัน ซึ่งเกิดจากเซลล์เกลีย 2 ประเภท ได้แก่เซลล์ชวานน์และโอลิโกเดนโดรไซต์ในระบบประสาทส่วนปลายเซลล์ชวานน์จะสร้างปลอกไมอีลินของแอกซอนที่มีไมอีลินหุ้ม ส่วนโอลิโกเดนโดรไซต์จะสร้างไมอีลินที่เป็นฉนวนในระบบประสาทส่วนกลาง ตามเส้นใยประสาทที่มีไมอีลินหุ้ม จะมีช่องว่างในปลอกไมอีลินที่เรียกว่าปมของแรนเวียร์เกิดขึ้นในระยะห่างเท่าๆ กัน การสร้างไมอีลินช่วยให้เกิดการส่งสัญญาณไฟฟ้าอย่างรวดเร็วเป็นพิเศษที่เรียกว่า การนำไฟฟ้า แบบกระโดด (saltatory conduction )
แอกซอนที่มีไมอีลินหุ้มจากเซลล์ประสาทในคอร์เทกซ์ก่อตัวเป็นเนื้อเยื่อประสาทส่วนใหญ่ที่เรียกว่าเนื้อขาวในสมอง ไมอีลินทำให้เนื้อเยื่อ มีลักษณะสีขาว ซึ่งแตกต่างจากเนื้อเทาของคอร์เทกซ์สมองที่ประกอบด้วยตัวเซลล์ประสาท การจัดเรียงที่คล้ายกันนี้พบได้ในซีรีเบลลัมกลุ่มของแอกซอนที่มีไมอีลินหุ้มประกอบเป็นเส้นประสาทในระบบประสาทส่วนกลาง และบริเวณที่พวกมันข้ามเส้นกลางของสมองเพื่อเชื่อมต่อบริเวณตรงข้ามกันเรียกว่าคอมมิสซูร์ คอมมิสซูร์ที่ใหญ่ที่สุดคือคอร์ปัสคัลโลซัมที่เชื่อมต่อซีกสมอง ทั้งสองข้าง และมีแอกซอนประมาณ 20 ล้านเส้น[ 4 ]
โครงสร้างของเซลล์ประสาทนั้นประกอบด้วยสองส่วนที่ทำหน้าที่แยกจากกัน หรือสองส่วนย่อย คือ ส่วนลำตัวเซลล์พร้อมกับเดนไดรต์เป็นส่วนหนึ่ง และส่วนแอกซอนเป็นอีกส่วนหนึ่ง
บริเวณแอกซอน
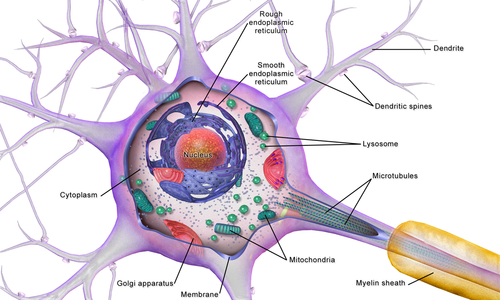
บริเวณหรือช่องแอกซอน ประกอบด้วยแอกซอนฮิลล็อก ส่วนเริ่มต้น ส่วนที่เหลือของแอกซอน และเทโลเดนเดรียของแอกซอน รวมถึงปลายแอกซอน นอกจากนี้ยังรวมถึงปลอกไมอีลินด้วยไม่มีนิสเซิลบอดี้ ที่สร้างโปรตีนประสาทในบริเวณแอกซอน [ 3 ]โปรตีนที่จำเป็นสำหรับการเจริญเติบโตของแอกซอนและการกำจัดของเสีย จำเป็นต้องมีโครงสร้างสำหรับการขนส่งการขนส่งแอกซอน นี้ เกิดขึ้นในแอกโซพลาสม์โดยการจัดเรียงของไมโครทิวบูลและเส้นใยระดับกลางประเภท IVที่เรียกว่านิวโรฟิลาเมนต์
เนินแอกซอน
แอกซอนฮิลล็อกคือบริเวณที่เกิดจากตัวเซลล์ของเซลล์ประสาทขณะที่มันขยายตัวกลายเป็นแอกซอน มันอยู่ก่อนหน้าส่วนเริ่มต้นสัญญาณศักย์ไฟฟ้าที่ ได้รับและ รวมกันในเซลล์ประสาทจะถูกส่งไปยังแอกซอนฮิลล็อกเพื่อสร้างศักย์ไฟฟ้าจากส่วนเริ่มต้น
ส่วนเริ่มต้นของแอกซอน
ส่วนเริ่มต้นของแอกซอน (AIS) เป็นไมโครโดเมนของแอกซอนที่มีโครงสร้างและหน้าที่แยกจากกัน[ 6 ] [ 7 ]หน้าที่อย่างหนึ่งของส่วนเริ่มต้นคือการแยกส่วนหลักของแอกซอนออกจากส่วนที่เหลือของเซลล์ประสาท อีกหน้าที่หนึ่งคือการช่วยเริ่มต้นศักยภาพการกระทำ[ 8 ]หน้าที่ทั้งสองนี้สนับสนุนขั้วของเซลล์ ประสาท ซึ่งเดนไดรต์ (และในบางกรณีโซมา ) ของเซลล์ประสาทจะรับสัญญาณอินพุตที่บริเวณฐาน และที่บริเวณปลาย แอกซอนของเซลล์ประสาทจะให้สัญญาณเอาต์พุต[ 9 ]
ส่วนเริ่มต้นของแอกซอนไม่มีปลอกไมอีลินและประกอบด้วยโปรตีนเชิงซ้อนพิเศษ มีความยาวประมาณ 20 ถึง 60 ไมโครเมตร และทำหน้าที่เป็นจุดเริ่มต้นของศักยภาพการกระทำ[ 10 ] [ 11 ]ทั้งตำแหน่งบนแอกซอนและความยาวของ AIS สามารถเปลี่ยนแปลงได้ แสดงให้เห็นถึงความยืดหยุ่นในระดับหนึ่งที่สามารถปรับแต่งผลลัพธ์ของเซลล์ประสาทได้อย่างละเอียด[ 10 ] [ 12 ] AIS ที่ยาวขึ้นสัมพันธ์กับความตื่นตัวที่มากขึ้น[ 12 ]ความยืดหยุ่นยังพบได้ในความสามารถของ AIS ในการเปลี่ยนแปลงการกระจายตัวและรักษาการทำงานของวงจรประสาทให้อยู่ในระดับคงที่[ 13 ]
AIS มีความเชี่ยวชาญสูงในการนำกระแสประสาท อย่างรวดเร็ว ซึ่งทำได้โดยความเข้มข้นสูงของช่องโซเดียมที่ควบคุมด้วยแรงดันไฟฟ้าในส่วนเริ่มต้นซึ่งเป็นจุดเริ่มต้นของศักยภาพการกระทำ[ 13 ] ช่องไอออนเหล่านี้มาพร้อมกับ โมเลกุลการยึดเกาะเซลล์และโปรตีนโครงสร้างจำนวนมากที่ยึดพวกมันไว้กับโครงร่างเซลล์[ 10 ]ปฏิสัมพันธ์กับแอนไคริน-จีมีความสำคัญเนื่องจากเป็นตัวจัดระเบียบหลักใน AIS [ 10 ]
ในกรณีอื่นๆ ดังที่พบในการศึกษาในหนู แอกซอนจะกำเนิดจากเดนไดรต์ แอกซอนดังกล่าวเรียกว่ามี "ต้นกำเนิดจากเดนไดรต์" แอกซอนบางส่วนที่มีต้นกำเนิดจากเดนไดรต์จะมีส่วนเริ่มต้น "ส่วนต้น" ที่เริ่มต้นตรงจุดกำเนิดของแอกซอน ในขณะที่บางส่วนมีส่วนเริ่มต้น "ส่วนปลาย" ที่แยกออกจากจุดกำเนิดของแอกซอนอย่างชัดเจน[ 14 ]ในหลายๆ สปีชีส์ เซลล์ประสาทบางส่วนมีแอกซอนที่แผ่ออกมาจากเดนไดรต์ ไม่ใช่จากตัวเซลล์ และสิ่งเหล่านี้เรียกว่าเดนไดรต์ที่นำพาแอกซอน[ 1 ]ในหลายกรณี แอกซอนจะกำเนิดที่เนินแอกซอนบนตัวเซลล์ แอกซอนดังกล่าวเรียกว่ามี "ต้นกำเนิดจากตัวเซลล์" แอกซอนบางส่วนที่มีต้นกำเนิดจากตัวเซลล์จะมีส่วนเริ่มต้น "ส่วนต้น" ที่อยู่ติดกับเนินแอกซอน ในขณะที่บางส่วนมีส่วนเริ่มต้น "ส่วนปลาย" ที่แยกออกจากตัวเซลล์โดยเนินแอกซอนที่ยื่นออกมา[ 14 ]
การลำเลียงแอกซอน
แอ็กโซพลาสม์เทียบเท่ากับไซโตพลาสม์ในเซลล์ ไมโครทิวบูลก่อตัวขึ้นในแอ็กโซพลาสม์ที่แอกซอนฮิลล็อก พวกมันเรียงตัวตามความยาวของแอกซอนในส่วนที่ซ้อนทับกัน และทั้งหมดชี้ไปในทิศทางเดียวกัน – ไปยังปลายแอกซอน[ 15 ]สิ่งนี้สังเกตได้จากปลายบวกของไมโครทิวบูล การจัดเรียงที่ซ้อนทับกันนี้เป็นเส้นทางสำหรับการขนส่งวัสดุต่างๆ จากตัวเซลล์[ 15 ]การศึกษาเกี่ยวกับแอ็กโซพลาสม์แสดงให้เห็นการเคลื่อนที่ของเวสิเคิลจำนวนมากทุกขนาดตามเส้นใยโครงร่างเซลล์ – ไมโครทิวบูลและนิวโรฟิลาเมนต์ในทั้งสองทิศทางระหว่างแอกซอนและปลายแอกซอนกับตัวเซลล์
การขนส่งแบบแอนเทอโรเกรดขาออกจากตัวเซลล์ไปตามแอกซอน จะนำไมโทคอนเดรียและโปรตีนเยื่อ หุ้มเซลล์ ที่จำเป็นต่อการเจริญเติบโตไปยังปลาย แอกซอน การขนส่งแบบรีโทรเกรด ขาเข้า จะนำของเสียจากเซลล์จากปลายแอกซอนไปยังตัวเซลล์[ 16 ]เส้นทางขาออกและขาเข้าใช้โปรตีนมอเตอร์ชุดที่แตกต่างกัน[ 15 ]การขนส่งขาออกดำเนินการโดยไคเนซินและการขนส่งขาเข้าแบบย้อนกลับดำเนินการโดยไดเนอิน ไดเนอินมุ่งไปยังปลายลบ[ 16 ]มีโปรตีนมอเตอร์ไคเนซินและไดเนอินหลายรูปแบบ และแต่ละรูปแบบเชื่อว่าบรรทุกสินค้าที่แตกต่างกัน[ 15 ]การศึกษาเกี่ยวกับการขนส่งในแอกซอนนำไปสู่การตั้งชื่อไคเนซิน[ 15 ]
การสร้างไมอีลิน


ในระบบประสาท แอกซอนอาจมีไมอีลินหุ้มหรือไม่มีไมอีลินหุ้มก็ได้ นี่คือการสร้างชั้นฉนวนที่เรียกว่าปลอกไมอีลิน เยื่อไมอีลินมีความพิเศษตรงที่มีอัตราส่วนของไขมันต่อโปรตีนค่อนข้างสูง[ 17 ]
ในระบบประสาทส่วนปลาย แอกซอนจะถูกหุ้มด้วยไมอีลินโดยเซลล์เกลียที่เรียกว่าเซลล์ชวานน์ในระบบประสาทส่วนกลาง ปลอกไมอีลินจะถูกสร้างขึ้นโดยเซลล์เกลียอีกประเภทหนึ่ง คือ โอลิโกเดนโดรไซต์เซลล์ชวานน์จะหุ้มไมอีลินให้กับแอกซอนเพียงเส้นเดียว โอลิโกเดนโดรไซต์สามารถหุ้มไมอีลินให้กับแอกซอนได้มากถึง 50 เส้น[ 18 ]
องค์ประกอบของไมอีลินแตกต่างกันในระบบประสาทส่วนกลางและระบบประสาทส่วนปลาย ในระบบประสาทส่วนกลาง โปรตีนไมอีลินหลักคือโปรตีนโปรตีโอลิปิดในขณะที่ในระบบประสาทส่วนปลายโปรตีนไมอีลินหลักคือโปรตีนไมอีลินเบสิก
โหนดของ Ranvier
โหนดของ Ranvier (หรือที่รู้จักกันในชื่อช่องว่างปลอกไมอีลิน ) คือส่วนสั้นๆ ที่ไม่มีไมอีลินหุ้มของแอกซอนที่มีไมอีลินหุ้ม ซึ่งพบแทรกเป็นระยะๆ ระหว่างส่วนของปลอกไมอีลิน ดังนั้น ณ จุดของโหนดของ Ranvier เส้นผ่านศูนย์กลางของแอกซอนจะลดลง[ 19 ]โหนดเหล่านี้เป็นบริเวณที่สามารถสร้างศักย์ไฟฟ้าได้ ในการนำไฟฟ้าแบบกระโดดกระแสไฟฟ้าที่เกิดขึ้นที่แต่ละโหนดของ Ranvier จะถูกส่งผ่านไปยังโหนดถัดไปโดยมีการลดทอนเพียงเล็กน้อย ซึ่งกระแสไฟฟ้ายังคงแรงพอที่จะสร้างศักย์ไฟฟ้าอีกครั้ง ดังนั้นในแอกซอนที่มีไมอีลินหุ้ม ศักย์ไฟฟ้าจึง "กระโดด" จากโหนดหนึ่งไปยังอีกโหนดหนึ่งอย่างมีประสิทธิภาพ โดยข้ามส่วนที่มีไมอีลินหุ้มระหว่างกัน ส่งผลให้ความเร็วในการแพร่กระจายเร็วกว่าแอกซอนที่ไม่มีไมอีลินหุ้มที่เร็วที่สุดเสียอีก
ปลายแอกซอน
แอกซอนสามารถแตกแขนงออกเป็นหลายสาขาเรียกว่าเทโลเดนเดรีย (ภาษากรีกแปลว่า 'ปลายต้นไม้') ที่ปลายของเทโลเดนเดรีย แต่ละอัน จะมีปลายแอกซอน (เรียกอีกอย่างว่าปลายปุ่มหรือปุ่มไซแนปส์ หรือปลายเท้า ) [ 20 ]ปลายแอกซอนมีถุงไซแนปส์ที่เก็บสารสื่อประสาท ไว้ เพื่อปล่อยออกมาที่ไซแนปส์ทำให้สามารถเชื่อมต่อไซแนปส์กับเซลล์ประสาทอื่นได้หลายจุด บางครั้งแอกซอนของเซลล์ประสาทอาจสร้างไซแนปส์กับเดนไดรต์ของเซลล์ประสาทเดียวกัน ซึ่งเรียกว่าออแทปส์ จุดเชื่อมต่อไซแนปส์บางจุดปรากฏตามความยาวของแอกซอนขณะที่มันขยายออกไป เรียกว่าปุ่มเอนพาสซองต์ ("ปุ่มที่ผ่านไป") และอาจมีเป็นร้อยหรือหลายพันจุดตามแอกซอนเดียว[ 3 ]
เส้นเลือดโป่งพองของแอกซอน
ในสมองที่พัฒนาตามปกติ บริเวณแกนของแอกซอนบางส่วนจะมีปุ่มประสาทก่อนซินแนปส์ หรือที่รู้จักกันในชื่อปุ่มแอกซอนและพบปุ่มเหล่านี้ในบริเวณของฮิปโปแคมปัสที่ทำหน้าที่ในการปล่อยสารสื่อประสาท[ 21 ]อย่างไรก็ตาม ปุ่มแอกซอนยังพบได้ในโรคทางระบบประสาทเสื่อม ซึ่งจะขัดขวางการนำกระแสประสาท ปุ่มแอกซอนยังเป็นลักษณะเด่นของการบาดเจ็บที่สมอง[ 21 ] [ 22 ] ความเสียหายของแอกซอนมักเกิดขึ้นกับโครงสร้างไซโตสเกเลตันของแอกซอน ทำให้การขนส่งหยุด ชะงักส่งผลให้โปรตีนสะสม เช่นโปรตีนพรีเคอร์เซอร์อะไมลอยด์เบต้าสามารถก่อตัวเป็นก้อนบวม ทำให้เกิดปุ่มจำนวนมากตามแนวแอกซอน[ 21 ] [ 22 ]
ศักยภาพการกระทำ


แอกซอนส่วนใหญ่ส่งสัญญาณในรูปของศักย์ไฟฟ้าแอคชั่น ซึ่งเป็นแรงกระตุ้นทางไฟฟ้าเคมีที่ไม่ต่อเนื่องกันและเดินทางอย่างรวดเร็วไปตามแอกซอน โดยเริ่มต้นจากตัวเซลล์และสิ้นสุดที่จุดที่แอกซอนสร้างการเชื่อมต่อไซแนปส์กับเซลล์เป้าหมาย คุณลักษณะที่สำคัญของศักย์ไฟฟ้าแอคชั่นคือ "ทั้งหมดหรือไม่มีเลย" – ศักย์ไฟฟ้าแอคชั่นทุกตัวที่แอกซอนสร้างขึ้นจะมีขนาดและรูปร่างเหมือนกันโดยพื้นฐาน คุณลักษณะ แบบทั้งหมดหรือไม่มีเลย นี้ ทำให้ศักย์ไฟฟ้าแอคชั่นสามารถส่งผ่านจากปลายด้านหนึ่งของแอกซอนยาวไปยังอีกด้านหนึ่งได้โดยไม่ลดขนาดลง อย่างไรก็ตาม มีเซลล์ประสาทบางชนิดที่มีแอกซอนสั้นซึ่งส่งสัญญาณทางไฟฟ้าเคมีแบบไล่ระดับที่มีแอมพลิจูดแปรผันได้
เมื่อศักย์ไฟฟ้ากระตุ้น (action potential) ไปถึงปลายประสาทก่อนซินแนปส์ (presynaptic terminal) มันจะกระตุ้นกระบวนการส่งสัญญาณประสาท ขั้นตอนแรกคือการเปิดช่องไอออนแคลเซียมในเยื่อหุ้มแอกซอนอย่างรวดเร็ว ทำให้ไอออนแคลเซียมไหลเข้าสู่ภายในเซลล์ผ่านเยื่อหุ้มเซลล์ การเพิ่มขึ้นของความเข้มข้นของแคลเซียมภายในเซลล์ส่งผลให้ถุงซินแนปส์ (ภาชนะขนาดเล็กที่ล้อมรอบด้วยเยื่อไขมัน) ที่บรรจุสารสื่อประสาทรวมตัวกับเยื่อหุ้มแอกซอนและปล่อยสารสื่อประสาทออกสู่ช่องว่างภายนอกเซลล์ สารสื่อประสาทจะถูกปล่อยออกจากปลายประสาทก่อนซินแนปส์ผ่าน กระบวนการ เอ็กโซไซโทซิส (exocytosis ) จากนั้นสารสื่อประสาทจะแพร่กระจายไปยังตัวรับที่อยู่บนเยื่อหุ้มเซลล์เป้าหมาย สารสื่อประสาทจะจับกับตัวรับเหล่านี้และกระตุ้นการทำงาน ขึ้นอยู่กับชนิดของตัวรับที่ถูกกระตุ้น ผลกระทบต่อเซลล์เป้าหมายอาจเป็นการกระตุ้นเซลล์เป้าหมาย ยับยั้งการทำงาน หรือเปลี่ยนแปลงกระบวนการเผาผลาญในบางลักษณะ ลำดับเหตุการณ์ทั้งหมดนี้มักเกิดขึ้นในเวลาน้อยกว่าหนึ่งในพันของวินาที หลังจากนั้น ภายในปลายประสาทก่อนซินแนปส์ ชุดเวสิเคิลใหม่จะถูกเคลื่อนย้ายไปยังตำแหน่งถัดจากเยื่อหุ้มเซลล์ พร้อมที่จะปล่อยออกมาเมื่อศักยภาพการกระทำครั้งต่อไปมาถึง ศักยภาพการกระทำเป็นขั้นตอนทางไฟฟ้าขั้นสุดท้ายในการรวมข้อความซินแนปส์ในระดับเซลล์ประสาท[ 5 ]
การบันทึกการแพร่กระจายศักยภาพการกระทำภายนอกเซลล์ในแอกซอนได้รับการสาธิตในสัตว์ที่เคลื่อนไหวได้อย่างอิสระ ในขณะที่ศักยภาพการกระทำของเซลล์ร่างกายภายนอกเซลล์ถูกนำมาใช้เพื่อศึกษาการทำงานของเซลล์ในสัตว์ที่เคลื่อนไหวได้อย่างอิสระ เช่นเซลล์ตำแหน่ง การทำงานของแอกซอนในทั้งเนื้อขาวและเนื้อเทาก็สามารถบันทึกได้เช่นกัน การบันทึกการแพร่กระจายศักยภาพการกระทำของแอกซอนภายนอกเซลล์นั้นแตกต่างจากศักยภาพการกระทำของเซลล์ร่างกายในสามประการ: 1. สัญญาณมีระยะเวลาพีค-รูทที่สั้นกว่า (~150 μs) เมื่อเทียบกับเซลล์พีระมิด (~500 μs) หรืออินเตอร์นิวรอน (~250 μs) 2. การเปลี่ยนแปลงแรงดันไฟฟ้าเป็นแบบสามเฟส 3. การทำงานที่บันทึกบนเทโทรดจะเห็นได้บนสายบันทึกเพียงเส้นเดียวจากสี่เส้น ในการบันทึกจากหนูที่เคลื่อนไหวได้อย่างอิสระ สัญญาณแอกซอนได้รับการแยกในเส้นใยเนื้อขาวรวมถึงอัลเวียสและคอร์ปัสคัลโลซัม ตลอดจนเนื้อเทาของฮิปโปแคมปัส[ 23 ]
ในความเป็นจริง การสร้างศักยภาพการกระทำในร่างกายมีลักษณะเป็นลำดับ และสไปค์ที่เป็นลำดับเหล่านี้ประกอบขึ้นเป็นรหัสดิจิทัลในเซลล์ประสาท แม้ว่าการศึกษาก่อนหน้านี้จะบ่งชี้ถึงต้นกำเนิดของแอกซอนของสไปค์เดี่ยวที่ถูกกระตุ้นโดยพัลส์ระยะสั้น แต่สัญญาณทางสรีรวิทยาในร่างกายจะกระตุ้นการเริ่มต้นของสไปค์ที่เป็นลำดับที่ตัวเซลล์ของเซลล์ประสาท[ 24 ] [ 25 ]
นอกจากการส่งต่อศักยภาพการกระทำไปยังปลายแอกซอนแล้ว แอกซอนยังสามารถขยายศักยภาพการกระทำ ซึ่งทำให้มั่นใจได้ว่าการส่งต่อศักยภาพการกระทำตามลำดับไปยังปลายแอกซอนจะปลอดภัย ในแง่ของกลไกโมเลกุลช่องโซเดียมที่ควบคุมด้วยแรงดันไฟฟ้าในแอกซอนมีเกณฑ์ ที่ต่ำกว่า และระยะเวลาการไม่ตอบสนอง ที่สั้นกว่า เมื่อตอบสนองต่อพัลส์ระยะสั้น[ 26 ]
การพัฒนาและการเติบโต
การพัฒนา
การพัฒนาของแอกซอนไปยังเป้าหมายเป็นหนึ่งในหกขั้นตอนหลักในการพัฒนาโดยรวมของระบบประสาท[ 27 ]การศึกษาที่ทำกับเซลล์ประสาทฮิปโปแคมปัสที่เพาะเลี้ยงแสดงให้เห็นว่าเซลล์ประสาทในตอนแรกสร้างนิวไรต์ หลายเส้น ที่เทียบเท่ากัน แต่มีเพียงนิวไรต์เส้นเดียวเท่านั้นที่จะกลายเป็นแอกซอน[ 28 ]ยังไม่ชัดเจนว่าการกำหนดแอกซอนเกิดขึ้นก่อนการยืดตัวของแอกซอนหรือในทางกลับกัน[ 29 ]แม้ว่าหลักฐานล่าสุดจะชี้ไปทางหลังก็ตาม หากแอกซอนที่ยังไม่พัฒนาเต็มที่ถูกตัด ขั้วอาจเปลี่ยนแปลงและนิวไรต์อื่นๆ อาจกลายเป็นแอกซอนได้ การเปลี่ยนแปลงขั้วนี้จะเกิดขึ้นก็ต่อเมื่อแอกซอนถูกตัดสั้นกว่านิวไรต์อื่นๆ อย่างน้อย 10 ไมโครเมตร หลังจากทำการตัดแล้ว นิวไรต์ที่ยาวที่สุดจะกลายเป็นแอกซอนในอนาคต และนิวไรต์อื่นๆ ทั้งหมด รวมถึงแอกซอนเดิม จะกลายเป็นเดนไดรต์[ 30 ]การออกแรงภายนอกกระทำต่อนิวไรต์ ทำให้มันยืดออกและกลายเป็นแอกซอน[ 31 ]อย่างไรก็ตาม การพัฒนาของแอกซอนเกิดขึ้นจากการทำงานร่วมกันที่ซับซ้อนระหว่างการส่งสัญญาณภายนอกเซลล์ การส่งสัญญาณภายในเซลล์ และพลวัต ของไซโตสเกเลตัน
การส่งสัญญาณภายนอกเซลล์
สัญญาณนอกเซลล์ที่แพร่กระจายผ่านเมทริกซ์นอกเซลล์ที่ล้อมรอบเซลล์ประสาทมีบทบาทสำคัญในการพัฒนาแอกซอน[ 32 ]โมเลกุลส่งสัญญาณเหล่านี้รวมถึงโปรตีนปัจจัยนิวโรโทรฟิกและเมทริกซ์นอกเซลล์และโมเลกุลยึดเกาะ เนทริน (หรือที่รู้จักกันในชื่อ UNC-6) ซึ่งเป็นโปรตีนที่หลั่งออกมา ทำหน้าที่ในการสร้างแอกซอน เมื่อ ตัวรับเนทริน UNC-5เกิดการกลายพันธุ์ นิวไรต์หลายเส้นจะยื่นออกมาจากเซลล์ประสาทอย่างไม่เป็นระเบียบ และในที่สุดแอกซอนเส้นเดียวจะยื่นออกมาทางด้านหน้า[ 33 ] [ 34 ] [ 35 ] [ 36 ]ปัจจัยนิวโรโทรฟิก ได้แก่ปัจจัยการเจริญเติบโตของเส้นประสาท (NGF) ปัจจัยนิวโรโทรฟิกที่ได้จากสมอง (BDNF) และนิวโรโทรฟิน-3 (NTF3) ก็มีส่วนเกี่ยวข้องในการพัฒนาแอกซอนและจับกับตัวรับ Trkด้วย[ 37 ]
เอนไซม์แปลงแกงกลิโอไซด์ไซอาลิเด สของเยื่อหุ้มพลาสมา (PMGS) ซึ่งเกี่ยวข้องกับการกระตุ้นTrkAที่ปลายของนิวไรต์ จำเป็นสำหรับการยืดตัวของแอกซอน PMGS กระจายตัวอย่างไม่สมมาตรไปยังปลายของนิวไรต์ซึ่งจะกลายเป็นแอกซอนในอนาคต[ 38 ]
การส่งสัญญาณภายในเซลล์
ในระหว่างการพัฒนาแอกซอน กิจกรรมของPI3Kจะเพิ่มขึ้นที่ปลายของแอกซอนที่กำหนด การขัดขวางกิจกรรมของ PI3K จะยับยั้งการพัฒนาแอกซอน การกระตุ้น PI3K ส่งผลให้เกิดการผลิตฟอสฟาติดิลอิโนซิทอล (3,4,5)-ไตรฟอสเฟต (PtdIns) ซึ่งสามารถทำให้นิวไรต์ยืดออกอย่างมากและเปลี่ยนเป็นแอกซอนได้ ดังนั้น การแสดงออกมากเกินไปของฟอสฟาเทสที่กำจัดฟอสเฟตออกจาก PtdIns จึงนำไปสู่ความล้มเหลวของการสร้างขั้ว[ 32 ]
พลวัตของโครงร่างเซลล์
เส้นประสาทที่มีปริมาณเส้นใย แอคตินต่ำที่สุดจะกลายเป็นแอกซอน ความเข้มข้นของ PGMS และ ปริมาณ f-actinมีความสัมพันธ์แบบผกผัน เมื่อ PGMS มีความเข้มข้นมากขึ้นที่ปลายเส้นประสาท ปริมาณ f-actin จะลดลงอย่างมาก[ 38 ]นอกจากนี้ การสัมผัสกับยาที่ทำให้แอคตินสลายตัวและสารพิษ B (ซึ่งทำให้การส่งสัญญาณ Rho ไม่ทำงาน ) ทำให้เกิดการสร้างแอกซอนหลายเส้น ดังนั้น การขัดจังหวะเครือข่ายแอคตินในกรวยเจริญเติบโตจะส่งเสริมให้เส้นประสาทนั้นกลายเป็นแอกซอน[ 39 ]
การเจริญเติบโต
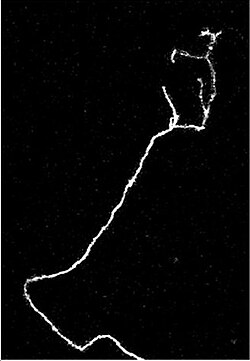
แอกซอนที่กำลังเติบโตจะเคลื่อนที่ผ่านสภาพแวดล้อมโดยผ่านกรวยการเจริญเติบโตซึ่งอยู่ที่ปลายของแอกซอน กรวยการเจริญเติบโตมีส่วนขยายคล้ายแผ่นกว้างที่เรียกว่าลาเมลลิโพเดียมซึ่งมีส่วนยื่นที่เรียกว่าฟิโลโพเดียฟิโลโพเดียเป็นกลไกที่ทำให้กระบวนการทั้งหมดเกาะติดกับพื้นผิวและสำรวจสภาพแวดล้อมโดยรอบ แอคตินมีบทบาทสำคัญในการเคลื่อนที่ของระบบนี้ สภาพแวดล้อมที่มีโมเลกุลการยึดเกาะเซลล์ (CAMs) ในระดับสูงจะสร้างสภาพแวดล้อมที่เหมาะสมสำหรับการเจริญเติบโตของแอกซอน ดูเหมือนว่าจะให้พื้นผิวที่ "เหนียว" สำหรับแอกซอนในการเจริญเติบโต ตัวอย่างของ CAMs ที่เฉพาะเจาะจงกับระบบประสาท ได้แก่N-CAM , TAG-1 ซึ่งเป็น ไกลโคโปรตีน ของแอกซอน[ 40 ] และMAGซึ่งทั้งหมดเป็นส่วนหนึ่งของ ตระกูล อิมมูโนโกลบูลิน โมเลกุลอีกชุดหนึ่งที่เรียกว่าเมทริกซ์นอกเซลล์ - โมเลกุลการยึดเกาะยังให้พื้นผิวที่เหนียวสำหรับแอกซอนในการเจริญเติบโตอีกด้วย ตัวอย่างของโมเลกุลเหล่านี้ ได้แก่ลามินินไฟโบรเนกตินเทนาซินและเพอร์เลแคนบางชนิดยึดติดอยู่กับพื้นผิวของเซลล์ จึงทำหน้าที่เป็นสารดึงดูดหรือสารขับไล่ในระยะสั้น ในขณะที่บางชนิดเป็นลิแกนด์ที่แพร่กระจายได้ จึงสามารถออกฤทธิ์ในระยะไกลได้
เซลล์ที่เรียกว่าเซลล์นำทางช่วยในการนำทางการเจริญเติบโตของแอกซอนประสาท เซลล์เหล่านี้ที่ช่วยนำทางแอกซอนมักจะเป็นเซลล์ประสาทอื่น ๆ ซึ่งบางครั้งอาจยังไม่เจริญเต็มที่ เมื่อแอกซอนเจริญเติบโตเสร็จสมบูรณ์ที่จุดเชื่อมต่อกับเป้าหมาย เส้นผ่านศูนย์กลางของแอกซอนสามารถเพิ่มขึ้นได้ถึงห้าเท่า ขึ้นอยู่กับความเร็วในการนำกระแสที่ต้องการ[ 41 ]
จากการวิจัยยังพบว่า หากแอกซอนของเซลล์ประสาทได้รับความเสียหาย ตราบใดที่โซมา (ตัวเซลล์ของเซลล์ประสาท) ไม่ได้รับความเสียหาย แอกซอนจะงอกใหม่และสร้างการเชื่อมต่อไซแนปส์กับเซลล์ประสาทใหม่โดยอาศัยเซลล์นำทางซึ่งเรียกอีกอย่างว่าการงอกใหม่ของ เซลล์ประสาท [ 42 ]
Nogo-Aเป็นส่วนประกอบยับยั้งการเจริญเติบโตของเส้นประสาทชนิดหนึ่งที่มีอยู่ในเยื่อหุ้มไมอีลินของระบบประสาทส่วนกลาง (พบในแอกซอน) มีบทบาทสำคัญในการจำกัดการงอกใหม่ของแอกซอนในระบบประสาทส่วนกลางของสัตว์เลี้ยงลูกด้วยนมที่โตเต็มวัย จากการศึกษาล่าสุดพบว่า หาก Nogo-A ถูกปิดกั้นและทำให้เป็นกลาง จะสามารถกระตุ้นการงอกใหม่ของแอกซอนในระยะไกล ซึ่งนำไปสู่การเพิ่มการฟื้นตัวของหน้าที่การทำงานในไขสันหลังของหนูและหนูทดลอง อย่างไรก็ตาม ยังไม่ได้ทำการทดลองในมนุษย์[ 43 ]การศึกษาล่าสุดยังพบว่ามาโครฟาจที่ถูกกระตุ้นผ่านเส้นทางการอักเสบเฉพาะที่ถูกกระตุ้นโดย ตัวรับ Dectin-1สามารถส่งเสริมการฟื้นตัวของแอกซอนได้ แต่ก็ทำให้เกิดความเป็นพิษต่อเซลล์ประสาท ด้วย [ 44 ]
การควบคุมความยาว
แอกซอนมีความยาวแตกต่างกันอย่างมาก ตั้งแต่ไม่กี่ไมโครเมตรไปจนถึงหลายเมตรในสัตว์บางชนิด สิ่งนี้เน้นย้ำว่าต้องมีกลไกการควบคุมความยาวของเซลล์ที่ช่วยให้เซลล์ประสาทสามารถรับรู้ความยาวของแอกซอนและควบคุมการเจริญเติบโตได้ตามนั้น มีการค้นพบว่าโปรตีนมอเตอร์มีบทบาทสำคัญในการควบคุมความยาวของแอกซอน[ 45 ]จากการสังเกตนี้ นักวิจัยได้พัฒนารูปแบบที่ชัดเจนสำหรับการเจริญเติบโตของแอกซอน โดยอธิบายว่าโปรตีนมอเตอร์สามารถส่งผลต่อความยาวของแอกซอนในระดับโมเลกุลได้อย่างไร[ 46 ] [ 47 ] [ 48 ] [ 49 ]การศึกษาเหล่านี้ชี้ให้เห็นว่าโปรตีนมอเตอร์นำโมเลกุลส่งสัญญาณจากโซมาไปยังกรวยเจริญเติบโตและในทางกลับกัน ซึ่งความเข้มข้นจะแกว่งไปมาตามเวลาด้วยความถี่ที่ขึ้นอยู่กับความยาว
การจำแนกประเภท
แอกซอนของเซลล์ประสาทในระบบประสาทส่วนปลาย ของมนุษย์ สามารถจำแนกได้ตามลักษณะทางกายภาพและคุณสมบัติการนำสัญญาณ แอกซอนเป็นที่ทราบกันว่ามีความหนาต่างกัน (ตั้งแต่ 0.1 ถึง 20 μm) [ 3 ]และความแตกต่างเหล่านี้เชื่อว่าเกี่ยวข้องกับความเร็วที่ศักยภาพการกระทำสามารถเดินทางไปตามแอกซอนได้ ซึ่งก็คือความเร็วในการนำสัญญาณ Erlanger และ Gasser ได้พิสูจน์สมมติฐานนี้ และระบุเส้นใยประสาทหลายประเภท โดยสร้างความสัมพันธ์ระหว่างเส้นผ่านศูนย์กลางของแอกซอนและความเร็วในการนำสัญญาณประสาท พวกเขาตีพิมพ์ผลการค้นพบของพวกเขาในปี 1941 ซึ่งเป็นการจำแนกแอกซอนครั้งแรก
แอกซอนถูกจำแนกออกเป็นสองระบบ ระบบแรกที่เสนอโดย Erlanger และ Gasser แบ่งเส้นใยออกเป็นสามกลุ่มหลักโดยใช้ตัวอักษร A, B และ C กลุ่มเหล่านี้ ได้แก่กลุ่ม A , กลุ่ม Bและกลุ่ม Cซึ่งรวมถึงทั้งเส้นใยรับความรู้สึก ( afferents ) และเส้นใยสั่งการ ( efferents ) กลุ่ม A แรกนั้นแบ่งย่อยออกเป็นเส้นใยอัลฟา เบตา แกมมา และเดลตา – Aα, Aβ, Aγ และ Aδ ตามลำดับ เซลล์ประสาทสั่งการของเส้นใยสั่งการต่างๆ เหล่านั้น คือเซลล์ประสาทสั่งการระดับล่าง – เซลล์ประสาทสั่งการอัลฟาเซลล์ประสาทสั่งการเบตาและเซลล์ประสาทสั่งการแกมมาซึ่งมีเส้นใยประสาท Aα, Aβ และ Aγ ตามลำดับ
ต่อมานักวิจัยกลุ่มอื่นได้ค้นพบว่าเส้นใย Aa สองกลุ่มเป็นเส้นใยรับความรู้สึก จากนั้นจึงนำเส้นใยเหล่านี้ไปจัดอยู่ในระบบ (การจำแนกประเภทของลอยด์) ซึ่งรวมเฉพาะเส้นใยรับความรู้สึกเท่านั้น (แม้ว่าบางส่วนจะเป็นเส้นประสาทผสมและเป็นเส้นใยสั่งการด้วยก็ตาม) ระบบนี้เรียกกลุ่มเส้นใยรับความรู้สึกว่า ประเภท และใช้เลขโรมัน ได้แก่ ประเภท Ia, ประเภท Ib, ประเภท II, ประเภท III และประเภท IV
มอเตอร์
เซลล์ประสาทสั่งการส่วนล่างมีเส้นใยอยู่สองชนิด:
| พิมพ์ | การจำแนกประเภทErlanger-Gasser | เส้นผ่านศูนย์กลาง(ไมโครเมตร) | ไมอีลิน | ความเร็วในการนำกระแสประสาท(เมตร/วินาที) | เส้นใยกล้ามเนื้อที่เกี่ยวข้อง |
|---|---|---|---|---|---|
| เซลล์ประสาทสั่งการอัลฟา (α) | เอα | 13–20 | ใช่ | 80–120 | เส้นใยกล้ามเนื้อนอกฟิวซัล |
| เซลล์ประสาทสั่งการเบต้า (β) | เอบีเอ | ||||
| เซลล์ประสาทสั่งการแกมมา (γ) | เอγ | 5-8 | ใช่ | 4–24 [ 50 ] [ 51 ] | เส้นใยกล้ามเนื้ออินทราฟิวซัล |
ประสาทสัมผัส
ตัวรับความรู้สึกต่าง ๆจะถูกควบคุมโดยเส้นใยประสาทชนิดต่าง ๆ ตัวรับความรู้สึกเกี่ยวกับการ ทรงตัว (Proprioceptors)ถูกควบคุมโดยเส้นใยประสาทรับความรู้สึกชนิด Ia, Ib และ II ตัวรับความรู้สึกเกี่ยวกับแรงกล (Mechanoreceptors) ถูกควบคุมโดยเส้นใยประสาทรับความรู้สึกชนิด II และ III และตัวรับความรู้สึกเกี่ยวกับความ เจ็บปวด (Nociceptors ) และ ตัวรับความรู้สึกเกี่ยวกับ อุณหภูมิ (Thermoreceptors)ถูกควบคุมโดยเส้นใยประสาทรับความรู้สึกชนิด III และ IV
| พิมพ์ | การจำแนกประเภทErlanger-Gasser | เส้นผ่านศูนย์กลาง(ไมโครเมตร) | ไมอีลิน | ความเร็ว ในการนำไฟฟ้า(เมตร/วินาที) | ตัวรับความรู้สึกที่เกี่ยวข้อง | ตัวรับรู้ตำแหน่ง | ตัวรับเชิงกล | ตัวรับความเจ็บปวดและตัวรับความร้อน |
|---|---|---|---|---|---|---|---|---|
| เอีย | เอα | 13–20 | ใช่ | 80–120 | ตัวรับหลักของกล้ามเนื้อสปินเดิล (ปลายแอนนูโลสไปรัล) | ✔ | ||
| อิบ | เอα | 13–20 | ใช่ | 80–120 | อวัยวะเอ็นกอลจิ | |||
| 2. | เอบีเอ | 6–12 | ใช่ | 33–75 | ตัวรับทุติยภูมิของเส้นใยกล้ามเนื้อ (ปลายรูปดอกไม้) ตัวรับกลไกทางผิวหนังทั้งหมด | ✔ | ||
| 3. | เอδ | 1–5 | บาง | 3–30 | ปลายประสาทอิสระ ที่รับรู้ การสัมผัสและแรงกดตัวรับความเจ็บปวดของวิถีประสาทสไปโนทาลามิกด้านข้าง ตัวรับอุณหภูมิเย็น | ✔ | ||
| IV | ซี | 0.2–1.5 | เลขที่ | 0.5–2.0 | ตัวรับความเจ็บปวดของเส้นทางสไปโนทาลามิกด้านหน้าตัวรับความอบอุ่น |
ระบบประสาทอัตโนมัติ
ระบบประสาทอัตโนมัติมีเส้นใยประสาทส่วนปลายอยู่สองชนิด:
| พิมพ์ | การจำแนกประเภทErlanger-Gasser | เส้นผ่านศูนย์กลาง[ 52 ] (μm) | ไมอีลิน[ 53 ] | ความเร็ว ในการนำไฟฟ้า(เมตร/วินาที) |
|---|---|---|---|---|
| เส้นใยพรีแกงกลิโอนิก | บี | 0.5–3 | ใช่ | 3–15 |
| เส้นใยหลังปมประสาท | ซี | 0.2–1.4 | เลขที่ | 0.5–2.0 |
ความสำคัญทางคลินิก
การบาดเจ็บของเส้นประสาทในระบบประสาทส่วนปลายสามารถอธิบายได้ตามลำดับความรุนแรงว่า เรียกว่านิวราแพรกเซีย แอ็กโซโนทเมซิสหรือนิวโรทเมซิสการ กระทบกระเทือนถือเป็นรูปแบบที่ไม่รุนแรงของการบาดเจ็บของแอกซอนแบบกระจาย[ 54 ]การบาดเจ็บของแอกซอนยังสามารถทำให้เกิด โครมาโตไลซิ สส่วนกลาง ได้อีกด้วย การทำงานผิดปกติของแอกซอนในระบบประสาทเป็นสาเหตุสำคัญประการหนึ่งของ ความผิดปกติทางระบบประสาทที่ถ่ายทอดทางพันธุกรรมและที่เกิดขึ้นภายหลังซึ่งส่งผลกระทบต่อทั้งเซลล์ประสาทส่วนปลายและส่วนกลาง[ 5 ]
เมื่อแอกซอนถูกบดขยี้ กระบวนการเสื่อมสภาพของแอกซอนจะเกิดขึ้นอย่างรวดเร็วที่ส่วนของแอกซอนที่อยู่ไกลจากตัวเซลล์มากที่สุด การเสื่อมสภาพนี้เกิดขึ้นอย่างรวดเร็วหลังจากได้รับบาดเจ็บ โดยส่วนของแอกซอนจะถูกปิดกั้นที่เยื่อหุ้มเซลล์และถูกทำลายโดยแมโครฟาจ ซึ่งเรียกว่าการเสื่อมสภาพแบบวอลเลเรียน [ 55 ] การตายของแอกซอนส่วนท้ายยังสามารถเกิดขึ้นได้ในโรคทางระบบประสาทเสื่อม หลายชนิด โดยเฉพาะอย่างยิ่งเมื่อการขนส่งแอกซอนบกพร่อง ซึ่งเรียกว่าการเสื่อมสภาพแบบคล้ายวอลเลเรียน[ 56 ]การศึกษาชี้ให้เห็นว่าการเสื่อมสภาพเกิดขึ้นเนื่องจากโปรตีนแอกซอนNMNAT2ถูกขัดขวางไม่ให้ไปถึงแอกซอนทั้งหมด[ 57 ]
การเสื่อมของปลอกไมอีลินของแอกซอนทำให้เกิดอาการทางระบบประสาทมากมายที่พบในโรคปลอกประสาทเสื่อมแข็ง
ภาวะไมอี ลินผิดปกติคือการสร้างปลอกไมอีลินที่ผิดปกติ ซึ่งเกี่ยวข้องกับโรคเม็ดเลือดขาวผิด ปกติหลายชนิด รวมถึงโรคจิตเภทด้วย[ 58 ] [ 59 ] [ 60 ]
การบาดเจ็บ รุนแรงที่สมองอาจส่งผลให้เกิดรอยโรคกระจายไปทั่วเส้นประสาท ทำให้แอกซอนเสียหายในภาวะที่เรียกว่าการบาดเจ็บของแอกซอนแบบกระจายซึ่งอาจนำไปสู่ภาวะหมดสติถาวรได้[ 61 ] จากการศึกษาใน หนูพบ ว่า ความเสียหายของแอกซอนจากการบาดเจ็บที่สมองเล็กน้อยเพียงครั้งเดียว อาจทำให้หนูมีความเสี่ยงต่อ การบาดเจ็บเพิ่มเติมได้ แม้จะได้รับบาดเจ็บที่สมองเล็กน้อยซ้ำๆ ก็ตาม[ 62 ]
ท่อช่วยนำทางเส้นประสาทเป็นวิธีการประดิษฐ์ขึ้นเพื่อนำทางการเจริญเติบโตของแอกซอนเพื่อช่วยให้เกิดการฟื้นฟูระบบประสาทและเป็นหนึ่งในวิธีการรักษามากมายที่ใช้สำหรับอาการบาดเจ็บของเส้นประสาท ประเภท ต่างๆ
ศัพท์เฉพาะ
พจนานุกรมทั่วไปบางเล่มให้คำจำกัดความของ "เส้นใยประสาท" ว่าเป็นกระบวนการของเซลล์ประสาท ใดๆ รวมถึงแอกซอนและเดนไดรต์ [ 63 ] [ 64 ] อย่างไรก็ตามแหล่งข้อมูลทางการแพทย์โดยทั่วไปใช้คำว่า "เส้นใยประสาท" เพื่อหมายถึงแอกซอนเท่านั้น[ 65 ] [ 66 ]
ประวัติศาสตร์
โดยทั่วไปแล้ว Otto Friedrich Karl Deitersนักกายวิภาคชาวเยอรมันได้รับการยกย่องว่าเป็นผู้ค้นพบแอกซอนโดยการแยกแยะแอกซอนออกจากเดนไดรต์[ 5 ] Rüdolf Albert von KöllikerชาวสวิสและRobert Remak ชาวเยอรมัน เป็นคนแรกที่ระบุและกำหนดลักษณะของส่วนเริ่มต้นของแอกซอน Kölliker ตั้งชื่อแอกซอนในปี 1896 [ 67 ] Louis-Antoine Ranvierเป็นคนแรกที่อธิบายช่องว่างหรือปมที่พบในแอกซอน และด้วยผลงานนี้ คุณลักษณะของแอกซอนเหล่านี้จึงถูกเรียกโดยทั่วไปว่า ป มของ Ranvier Santiago Ramón y Cajalนักกายวิภาคชาวสเปน เสนอว่าแอกซอนเป็นส่วนประกอบขาออกของเซลล์ประสาท โดยอธิบายถึงการทำงานของเซลล์ประสาท[ 5 ]โจเซฟ เออร์แลงเกอร์และเฮอร์เบิร์ต กัสเซอร์ได้พัฒนาระบบการจำแนกประเภทเส้นใยประสาทส่วนปลายก่อนหน้านี้[ 68 ]โดยอิงจากความเร็วในการนำกระแสประสาทของแอกซอน การสร้าง ไมอีลินขนาดของเส้นใย ฯลฯอลัน ฮอดจ์กินและแอนดรูว์ ฮักซ์ลีย์ยังได้ใช้แอกซอนยักษ์ของปลาหมึก (1939) และในปี 1952 พวกเขาได้คำอธิบายเชิงปริมาณที่สมบูรณ์เกี่ยวกับพื้นฐานไอออนของศักยภาพการกระทำ ซึ่งนำไปสู่การกำหนดรูปแบบของแบบจำลองฮอดจ์กิน-ฮักซ์ลีย์ฮอดจ์กินและฮักซ์ลีย์ได้รับรางวัลโนเบล ร่วมกัน สำหรับงานนี้ในปี 1963 สูตรที่ให้รายละเอียดเกี่ยวกับการนำกระแสประสาทของแอกซอนได้รับการขยายไปยังสัตว์มีกระดูกสันหลังในสมการแฟรงเคนเฮาเซอร์-ฮักซ์ลีย์ ความเข้าใจเกี่ยวกับพื้นฐานทางชีวเคมีสำหรับการแพร่กระจายของศักยภาพการกระทำได้ก้าวหน้าไปอีกขั้น และรวมถึงรายละเอียดมากมายเกี่ยวกับช่องไอออน แต่ละ ช่อง
สัตว์อื่นๆ
แอกซอนในสัตว์ไม่มีกระดูกสันหลังได้รับการศึกษาอย่างกว้างขวางปลาหมึกครีบยาวชายฝั่งซึ่งมักใช้เป็นสิ่งมีชีวิตต้นแบบมีแอกซอนที่ยาวที่สุดเท่าที่รู้จัก[ 69 ]ปลาหมึกยักษ์มีแอกซอนที่ใหญ่ที่สุดเท่าที่รู้จัก ขนาดของมันมีเส้นผ่านศูนย์กลางตั้งแต่ 0.5 (โดยทั่วไป) ถึง 1 มม. และใช้ในการควบคุม ระบบ ขับเคลื่อนแบบเจ็ทความเร็วในการนำกระแสที่บันทึกไว้ที่เร็วที่สุดคือ 210 ม./วินาที พบในแอกซอนที่มีปลอกหุ้มของกุ้งเพเนอิด บางชนิดที่อาศัยอยู่ในทะเลเปิด [ 70 ]และช่วงปกติจะอยู่ระหว่าง 90 ถึง 200 เมตร/วินาที[ 71 ] ( เทียบกับ 100–120 ม./วินาที สำหรับแอกซอนของสัตว์มีกระดูกสันหลังที่มีไมอีลินหุ้มที่เร็วที่สุด)
รูปภาพเพิ่มเติม
 การบันทึกข้อมูลในฮิปโปแคมปัสจากเซลล์และแอกซอนประเภทต่างๆ
การบันทึกข้อมูลในฮิปโปแคมปัสจากเซลล์และแอกซอนประเภทต่างๆ

ดูเพิ่มเติม
- สรีรวิทยาไฟฟ้า
- เนินปมประสาท
- โรคเส้นประสาทแอกซอนขนาดใหญ่
- การติดตามเส้นประสาท
- ไพโอเนียร์ แอ็กซอน
- การบันทึกแบบหน่วยเดียว
ลิงก์ภายนอก
- ภาพทางจุลพยาธิวิทยา: 3_09ที่ศูนย์วิทยาศาสตร์สุขภาพ มหาวิทยาลัยโอคลาโฮมา – "สไลด์ที่ 3 ไขสันหลัง "
{kind=link}
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ แอกซอน
แอกซอน (จากภาษากรีกโบราณ : ἄξων , โรมัน : áxōn , แปลตรงตัวว่า ' แกน' ; เรียกอีกอย่างว่าเส้นใยประสาทหรือไฟเบอร์ ) คือ...
กายวิภาคศาสตร์
แอกซอนเป็นเส้นทางการส่งสัญญาณหลักของ ระบบประสาท และเมื่อรวมกันเป็นมัดจะเกิดเป็น เส้นประสาท ในระบบประสาทส่วนปลาย หรือ เป็นเส้นใยประสาท ใน ระบบประสาทส่วนกลาง (CNS) แอกซอนบางเส้นอาจยาวได้ถึงหนึ่งเมตรหรือมากกว่านั้น ในขณะที่บางเส้นอาจยาวเพียงหนึ่งมิลลิเมตร...
บริเวณแอกซอน
บริเวณหรือช่องแอกซอน ประกอบด้วยแอกซอนฮิลล็อก ส่วนเริ่มต้น ส่วนที่เหลือของแอกซอน และเทโลเดนเดรียของแอกซอน รวมถึงปลายแอกซอน นอกจากนี้ยังรวมถึงปลอกไมอีลินด้วยไม่มี นิสเซิลบอดี้ ที่สร้างโปรตีนประสาทในบริเวณแอกซอน [ 3 ]...
การลำเลียงแอกซอน
แอ็ กโซพลาสม์ เทียบเท่ากับ ไซโตพลาสม์ ในเซลล์ ไมโครทิวบูลก่อตัวขึ้นในแอ็กโซพลาสม์ที่แอกซอนฮิลล็อก พวกมันเรียงตัวตามความยาวของแอกซอนในส่วนที่ซ้อนทับกัน และทั้งหมดชี้ไปในทิศทางเดียวกัน – ไปยังปลายแอกซอน [ 15 ] สิ่งนี้สังเกตได้จากปลายบวกของไมโครทิวบูล...