อ่าน 18 นาที
การส่งสัญญาณของเซลล์
การส่งสัญญาณของเซลล์ ( cell signalling ใน ภาษาอังกฤษแบบ บริติช ) คือ กระบวนการทางชีววิทยา ที่ เซลล์ มีปฏิสัมพันธ์กับตัวเอง กับเซลล์อื่น ๆ และกับสิ่งแวดล้อม...
การส่งสัญญาณของเซลล์
การส่งสัญญาณของเซลล์ ( cell signallingในภาษาอังกฤษแบบ บริติช ) คือกระบวนการทางชีววิทยาที่เซลล์มีปฏิสัมพันธ์กับตัวเอง กับเซลล์อื่น ๆ และกับสิ่งแวดล้อม การส่งสัญญาณของเซลล์เป็นคุณสมบัติพื้นฐานของสิ่งมีชีวิตทุกรูปแบบโดยทั่วไป กระบวนการส่งสัญญาณประกอบด้วยองค์ประกอบสามส่วน ได้แก่ ตัวส่งสารตัวแรก ( ligand ) ตัวรับและสัญญาณเอง[ 1 ]
ในทางชีววิทยา สัญญาณส่วนใหญ่มีลักษณะทางเคมี แต่ก็อาจเป็นสัญญาณทางกายภาพได้เช่น กันเช่นความดัน แรงดันไฟฟ้าอุณหภูมิหรือแสง สัญญาณทางเคมีคือโมเลกุลที่มีความสามารถในการจับและกระตุ้นตัวรับเฉพาะ โมเลกุลเหล่านี้เรียกอีกอย่างว่าลิแกนด์ มีความหลากหลายทางเคมี รวมถึงไอออน (เช่น Na + , K +และ Ca2 + ), ลิปิด (เช่น สเตียรอยด์, โปรสตาแกลนดิน), เปปไทด์ (เช่น อินซูลิน, ACTH), คาร์โบไฮเดรต, โปรตีนที่ถูกไกลโคซิเลต (โปรตีโอไกลแคน), กรดนิวคลีอิก เป็นต้น ลิแกนด์เปปไทด์และลิปิดมีความสำคัญเป็นพิเศษ เนื่องจากฮอร์โมนส่วนใหญ่อยู่ในกลุ่มสารเคมีเหล่านี้ เปปไทด์มักเป็นโมเลกุลที่มีขั้วและชอบน้ำ ดังนั้นจึงไม่สามารถแพร่ผ่านชั้นไขมันสองชั้นของเยื่อหุ้มเซลล์ได้อย่างอิสระ การทำงานของมันจึงต้องอาศัยตัวรับที่อยู่บนเยื่อหุ้มเซลล์ ในทางกลับกัน สารเคมีที่ละลายในไขมัน เช่น ฮอร์โมนสเตียรอยด์ สามารถแพร่ผ่านเยื่อหุ้มเซลล์ได้โดยไม่จำเป็นต้องใช้พลังงาน และมีปฏิสัมพันธ์กับตัวรับภายในเซลล์
การส่งสัญญาณของเซลล์สามารถจำแนกได้เป็นแบบออโตครีนอินทราครีนจุกซ์ตาครีนพาราค รีน หรือเอนโดครีนการส่งสัญญาณแบบออโตครีนเกิดขึ้นเมื่อสัญญาณเคมีออกฤทธิ์ต่อเซลล์เดียวกันกับที่ผลิตสารเคมีส่งสัญญาณนั้น[ 2 ]การส่งสัญญาณแบบอินทราครีนเกิดขึ้นเมื่อสัญญาณเคมีที่ผลิตโดยเซลล์หนึ่งออกฤทธิ์ต่อตัวรับที่อยู่ในไซโตพลาสซึมหรือนิวเคลียสของเซลล์เดียวกัน[ 3 ]การส่งสัญญาณแบบจุกซ์ตาครีนเกิดขึ้นระหว่างเซลล์ที่อยู่ติดกัน[ 4 ]การส่งสัญญาณแบบพาราครีนเกิดขึ้นระหว่างเซลล์ที่อยู่ใกล้เคียงกัน การส่งสัญญาณแบบเอนโดครีนเกิดขึ้นระหว่างเซลล์ที่อยู่ห่างไกลกัน โดยสัญญาณเคมี ( ฮอร์โมน ) มักถูกลำเลียงโดยกระแสเลือด[ 5 ]
ตัวรับสัญญาณเป็นโปรตีนที่ซับซ้อนหรือกลุ่มโปรตีนที่ยึดติดกันอย่างแน่นหนา ตั้งอยู่บนเยื่อหุ้มเซลล์หรือภายในเซลล์ เช่น ในไซโตพลาซึมออร์แกเนลล์และนิวเคลียสตัวรับสัญญาณมีความสามารถในการตรวจจับสัญญาณได้โดยการจับกับสารเคมีจำเพาะ หรือโดยการเปลี่ยนแปลงโครงสร้างเมื่อมีปฏิสัมพันธ์กับสารทางกายภาพ ความจำเพาะของปฏิกิริยาทางเคมีระหว่างลิแกนด์กับตัวรับสัญญาณทำให้เกิดความสามารถในการกระตุ้นการตอบสนองของเซลล์ที่จำเพาะ ตัวรับสัญญาณสามารถแบ่งออกเป็นสองประเภทใหญ่ๆ คือ ตัวรับสัญญาณบนเยื่อหุ้มเซลล์และตัวรับสัญญาณภายในเซลล์

ตัวรับบนเยื่อหุ้มเซลล์สามารถจำแนกเพิ่มเติมได้เป็นตัวรับที่เชื่อมโยงกับช่องไอออน ตัวรับที่เชื่อมโยงกับโปรตีนจี และตัวรับที่เชื่อมโยงกับเอนไซม์
- ตัวรับช่องไอออนเป็นโปรตีนขนาดใหญ่ที่อยู่ภายในเยื่อหุ้มเซลล์ มีหน้าที่เป็นประตูที่ถูกกระตุ้นด้วยลิแกนด์ เมื่อตัวรับเหล่านี้ถูกกระตุ้น พวกมันอาจยอมให้หรือปิดกั้นการผ่านของไอออนเฉพาะบางชนิดผ่านเยื่อหุ้มเซลล์ ตัวรับส่วนใหญ่ที่ถูกกระตุ้นด้วยสิ่งเร้าทางกายภาพ เช่น ความดันหรืออุณหภูมิ จัดอยู่ในประเภทนี้
- ตัวรับโปรตีนจีเป็นโปรตีนหลายหน่วยที่ฝังอยู่ในเยื่อหุ้มเซลล์ ตัวรับเหล่านี้มีโดเมนภายนอกเซลล์ โดเมนที่ทะลุผ่านเยื่อหุ้มเซลล์ และโดเมน ภายใน เซลล์ โดเมนภายนอกเซลล์มีหน้าที่ในการจับกับลิแกนด์จำเพาะ ส่วนโดเมนภายในเซลล์มีหน้าที่ในการเริ่มต้นปฏิกิริยาเคมีแบบต่อเนื่อง ซึ่งจะกระตุ้นการทำงานของเซลล์ตามที่ตัวรับควบคุมไว้ในที่สุด
- ตัวรับที่เชื่อมโยงกับเอนไซม์เป็นโปรตีนที่อยู่บนเยื่อหุ้มเซลล์ โดยมีโดเมนภายนอกเซลล์ที่ทำหน้าที่จับกับลิแกนด์จำเพาะ และโดเมนภายในเซลล์ที่มีฤทธิ์ทางเอนไซม์หรือเร่งปฏิกิริยา เมื่อถูกกระตุ้น ส่วนที่เป็นเอนไซม์จะทำหน้าที่ส่งเสริมปฏิกิริยาเคมีภายในเซลล์ที่จำเพาะ
ตัวรับภายในเซลล์มีกลไกการทำงาน ที่แตกต่างออก ไป โดยปกติแล้วจะจับกับลิแกนด์ที่ละลายในไขมันซึ่งแพร่ผ่านเยื่อหุ้มเซลล์ได้เอง เช่น ฮอร์โมนสเตียรอยด์ ลิแกนด์เหล่านี้จะจับกับตัวขนส่งเฉพาะในไซโตพลาสซึม ซึ่งจะนำพาสารประกอบฮอร์โมน-ตัวขนส่งเข้าไปในนิวเคลียส ที่ซึ่งยีนเฉพาะจะถูกกระตุ้นและส่งเสริมการสังเคราะห์โปรตีนเฉพาะ
ส่วนประกอบที่ทำหน้าที่ส่งสัญญาณเริ่มต้นด้วยการถ่ายทอดสัญญาณในกระบวนการนี้ สัญญาณจะทำปฏิกิริยากับตัวรับ และเริ่มต้นเหตุการณ์ระดับโมเลกุลต่างๆ ภายในเซลล์ ซึ่งนำไปสู่ผลลัพธ์สุดท้ายของกระบวนการส่งสัญญาณ โดยทั่วไป ผลลัพธ์สุดท้ายจะประกอบด้วยการกระตุ้นช่องไอออน ( ช่องไอออนที่ควบคุมด้วยลิแกนด์ ) หรือการเริ่มต้นของระบบตัวส่งสัญญาณรองที่ส่งสัญญาณไปทั่วเซลล์ ระบบตัวส่งสัญญาณรองสามารถขยายหรือปรับเปลี่ยนสัญญาณได้ โดยการกระตุ้นตัวรับเพียงไม่กี่ตัวจะส่งผลให้ตัวส่งสัญญาณรองหลายตัวถูกกระตุ้น ทำให้สัญญาณเริ่มต้น (ตัวส่งสัญญาณแรก) ถูกขยายให้แรงขึ้นผลกระทบที่ตามมาของเส้นทางการส่งสัญญาณเหล่านี้อาจรวมถึงกิจกรรมของเอนไซม์เพิ่มเติม เช่นการแตกตัวของโปรตีนการ ฟอส ฟอริเลชัน การเมทิลเลชันและการยูบิควิตินิเลชัน
โมเลกุลส่งสัญญาณสามารถสังเคราะห์ได้จากกระบวนการสังเคราะห์ทางชีวภาพต่างๆ และถูกปล่อยออกมาผ่านการขนส่งแบบพาสซีฟหรือ แอคทีฟ หรือแม้กระทั่งจากความ เสียหายของเซลล์
เซลล์แต่ละเซลล์ถูกตั้งโปรแกรมให้ตอบสนองต่อโมเลกุลสัญญาณภายนอก เซลล์ ที่เฉพาะเจาะจง ซึ่งเป็นพื้นฐานของการพัฒนาการซ่อมแซมเนื้อเยื่อภูมิคุ้มกันและภาวะสมดุลของร่างกายความผิดพลาดในปฏิสัมพันธ์ของสัญญาณอาจทำให้เกิดโรคต่างๆ เช่นมะเร็ง โรคภูมิต้านตนเองและเบาหวาน
ขอบเขตทางอนุกรมวิธาน
ในสิ่งมีชีวิตขนาดเล็กหลาย ชนิดเช่นแบคทีเรียการรับรู้จำนวนประชากรช่วยให้แต่ละบุคคลสามารถเริ่มกิจกรรมได้ก็ต่อเมื่อประชากรมีขนาดใหญ่เพียงพอ การส่งสัญญาณระหว่างเซลล์นี้ถูกสังเกตครั้งแรกในแบคทีเรียในทะเลAliivibrio fischeriซึ่งผลิตแสงเมื่อประชากรมีความหนาแน่นเพียงพอ[ 6 ]กลไกนี้เกี่ยวข้องกับการผลิตและการตรวจจับโมเลกุลส่งสัญญาณ และการควบคุมการถอดรหัสยีนเพื่อตอบสนอง การรับรู้จำนวนประชากรทำงานในแบคทีเรียทั้งแกรมบวกและแกรมลบ และทั้งภายในและระหว่างสายพันธุ์[ 7 ]
ในราเมือกเซลล์แต่ละเซลล์จะรวมตัวกันเพื่อสร้างโครงสร้างผลและในที่สุดก็กลายเป็นสปอร์ ภายใต้อิทธิพลของสัญญาณทางเคมีที่เรียกว่าอะครา ซิน เซลล์จะเคลื่อนที่โดยเคโมแท็กซิส กล่าว คือ พวกมันถูกดึงดูดโดยความลาดชันทางเคมี บางชนิดใช้ไซคลิก AMPเป็นสัญญาณ ในขณะที่บางชนิด เช่นPolysphondylium violaceumใช้ไดเปปไทด์ที่เรียกว่ากลอริน[ 8 ]
ในพืชและสัตว์ การส่งสัญญาณระหว่างเซลล์เกิดขึ้นได้ทั้งโดยการปล่อยสารเข้าสู่ช่องว่างนอกเซลล์ซึ่งแบ่งออกเป็นการส่งสัญญาณแบบพาราครีน (ในระยะทางสั้นๆ) และการส่งสัญญาณแบบเอนโดครีน (ในระยะทางไกล) หรือโดยการสัมผัสโดยตรง ซึ่งเรียกว่าการส่งสัญญาณแบบจุกซ์ตาครีน เช่นการส่งสัญญาณแบบน็อตช์[ 9 ]การส่งสัญญาณแบบออโตครีนเป็นกรณีพิเศษของการส่งสัญญาณแบบพาราครีน โดยที่เซลล์ที่หลั่งสารมีความสามารถในการตอบสนองต่อโมเลกุลส่งสัญญาณที่หลั่งออกมา[ 10 ] การส่งสัญญาณ แบบไซแนปส์เป็นกรณีพิเศษของการส่งสัญญาณแบบพาราครีน (สำหรับไซแนปส์เคมี ) หรือการส่งสัญญาณแบบจุกซ์ตาครีน (สำหรับไซแนปส์ไฟฟ้า ) ระหว่างเซลล์ประสาทและเซลล์เป้าหมาย
สัญญาณภายนอกเซลล์
การสังเคราะห์และการปล่อย

สัญญาณเซลล์จำนวนมากถูกส่งผ่านโมเลกุลที่ถูกปล่อยออกมาจากเซลล์หนึ่งและเคลื่อนที่ไปสัมผัสกับเซลล์อื่น โมเลกุลส่งสัญญาณสามารถอยู่ในกลุ่มสารเคมีหลายประเภท ได้แก่ลิปิด ฟอ สโฟลิปิด กรดอะมิโน โมโนอะมีน โปรตีน ไกลโคโปรตีน หรือก๊าซโมเลกุลส่งสัญญาณที่จับกับตัวรับบนพื้นผิวโดยทั่วไปจะมีขนาดใหญ่และชอบน้ำ (เช่นTRHวาโซเพรสซินอะเซทิลโคลีน ) ในขณะที่โมเลกุลที่เข้าสู่เซลล์โดยทั่วไปจะมีขนาดเล็กและไม่ชอบน้ำ (เช่นกลูโคคอร์ติคอยด์ ฮอร์โมนไทรอยด์โคลีแคลซิเฟอรอลกรดเรติโนอิก ) แต่ก็มีข้อยกเว้นที่สำคัญมากมาย และโมเลกุลเดียวกันสามารถออกฤทธิ์ได้ทั้งผ่านตัวรับบนพื้นผิวหรือในลักษณะภายในเซลล์เพื่อให้เกิดผลที่แตกต่างกัน[ 10 ]ในเซลล์สัตว์เซลล์ต่อมไร้ท่อ เฉพาะ จะปล่อยฮอร์โมนเหล่านี้และส่งผ่านระบบไหลเวียนโลหิตไปยังส่วนอื่นๆ ของร่างกาย จากนั้นฮอร์โมนเหล่านี้จะไปถึงเซลล์เป้าหมาย ซึ่งสามารถจดจำและตอบสนองต่อฮอร์โมนและก่อให้เกิดผลลัพธ์ สิ่งนี้เรียกอีกอย่างว่าการส่งสัญญาณทางต่อมไร้ท่อสารควบคุมการเจริญเติบโตของพืช หรือฮอร์โมนพืชเคลื่อนที่ผ่านเซลล์หรือแพร่กระจายผ่านอากาศในรูปของก๊าซเพื่อไปถึงเป้าหมาย[ 11 ]ไฮโดรเจนซัลไฟด์ ถูกผลิตขึ้นในปริมาณเล็กน้อยโดยเซลล์บางส่วนของร่างกายมนุษย์และมีหน้าที่ในการส่งสัญญาณทางชีวภาพหลายประการ ปัจจุบันมีก๊าซอื่นเพียงสองชนิดเท่านั้นที่ทราบว่าทำ หน้าที่เป็นโมเลกุลส่งสัญญาณในร่างกายมนุษย์ ได้แก่ไนตริกออกไซด์และคาร์บอนมอนอกไซด์[ 12 ]
เอ็กโซไซโทซิส
เอ็กโซไซโทซิสเป็นกระบวนการที่เซลล์ขนส่งโมเลกุลเช่นสารสื่อประสาทและโปรตีนออกจากเซลล์ เนื่องจากเป็น กลไก การขนส่งแบบแอคทีฟ เอ็กโซไซโทซิสจึงต้องใช้พลังงานในการขนส่งสาร เซลล์ทุกเซลล์ใช้เอ็กโซไซโทซิสและเอนโดไซโทซิสซึ่งเป็นกระบวนการที่นำสารเข้าสู่เซลล์ เนื่องจากสารเคมี ส่วนใหญ่ ที่สำคัญต่อเซลล์เป็น โมเลกุล ขั้วขนาด ใหญ่ ที่ไม่สามารถผ่าน ส่วนที่ ไม่ชอบน้ำของเยื่อหุ้มเซลล์ได้ด้วยการขนส่งแบบพาสซีฟ เอ็กโซไซโทซิสเป็นกระบวนการที่ปล่อยโมเลกุลจำนวนมากออกมา ดังนั้นจึงเป็นรูปแบบหนึ่งของการขนส่งแบบกลุ่ม เอ็กโซไซโทซิสเกิดขึ้นผ่านทางช่องทางหลั่งที่เยื่อหุ้มพลาสมาของเซลล์ที่เรียกว่าพอโรโซมพอโรโซมเป็นโครงสร้างไลโปโปรตีนรูปถ้วยถาวรที่เยื่อหุ้มพลาสมาของเซลล์ ซึ่งถุงหลั่งจะเชื่อมต่อและหลอมรวมกันชั่วคราวเพื่อปล่อยสารภายในถุงออกจากเซลล์[ 13 ]
ในบริบทของการส่งสัญญาณประสาทสารสื่อประสาทมักจะถูกปล่อยออกมาจากถุงไซแนปส์เข้าไปในช่องว่างไซแนปส์ผ่านกระบวนการเอ็กโซไซโทซิส อย่างไรก็ตาม สารสื่อประสาทยังสามารถถูกปล่อยออกมาผ่านการขนส่งย้อนกลับโดยผ่านโปรตีนขนส่งของเยื่อหุ้มเซลล์ ได้อีก ด้วย[ 14 ]
ประเภท

ออโตครีน

การส่งสัญญาณแบบออโตครีนเกี่ยวข้องกับเซลล์ที่หลั่งฮอร์โมนหรือสารสื่อสารทางเคมี (เรียกว่าตัวแทนออโตครีน) ที่จับกับตัวรับออโตครีนบนเซลล์เดียวกัน ทำให้เกิดการเปลี่ยนแปลงในเซลล์นั้นเอง[ 15 ]ซึ่งแตกต่างจากการส่งสัญญาณแบบพาราครีน การส่งสัญญาณแบบอินทราครีน หรือการส่งสัญญาณแบบเอนโดครีนแบบดั้งเดิม
อินทราครีน
ในการส่งสัญญาณภายในเซลล์ สารเคมีส่งสัญญาณจะถูกผลิตขึ้นภายในเซลล์และจับกับตัวรับไซโตโซลหรือนิวเคลียร์ภายในเซลล์ การส่งสัญญาณประเภทนี้ทำให้แตกต่างจากกลไกการส่งสัญญาณของเซลล์อื่นๆ เช่น การส่งสัญญาณแบบออโตครีน ในการส่งสัญญาณทั้งแบบออโตครีนและภายในเซลล์ สัญญาณจะมีผลต่อเซลล์ที่ผลิตสัญญาณนั้น[ 16 ]
จัคสทาครีน
การส่งสัญญาณแบบจุกซ์ตาครีน (Juxtacrine signaling)เป็นรูปแบบหนึ่งของ การส่งสัญญาณระหว่าง เซลล์กับเซลล์ หรือเซลล์กับเมทริกซ์นอกเซลล์ในสิ่งมีชีวิตหลายเซลล์ซึ่งต้องอาศัยการสัมผัสใกล้ชิด มีอยู่สามประเภท:
- สาร ที่จับกับเยื่อหุ้มเซลล์( โปรตีนโอ ลิโกแซ็ กคาไรด์ลิปิด ) และโปรตีนในเยื่อหุ้มเซลล์ของเซลล์ที่อยู่ติดกันสองเซลล์จะทำปฏิกิริยาต่อกัน
- ช่องว่าง เชื่อมต่อ ระหว่าง เซลล์ (gap junction)ทำหน้าที่เชื่อมโยงช่องว่างภายในเซลล์ของเซลล์ที่อยู่ติดกันสองเซลล์ ทำให้โมเลกุลขนาดเล็กสามารถผ่านเข้าไปได้
- ไกลโคโปรตีนในเมทริกซ์นอกเซลล์และโปรตีนในเยื่อหุ้มเซลล์มีปฏิสัมพันธ์กัน
นอกจากนี้ ในสิ่งมีชีวิตเซลล์เดียวเช่นแบคทีเรียการส่งสัญญาณแบบจุกซ์ตาครีนหมายถึงการโต้ตอบผ่านการสัมผัสของเยื่อหุ้มเซลล์ มีการสังเกตการส่งสัญญาณแบบจุกซ์ตาครีนสำหรับปัจจัยการเจริญเติบโต บางชนิด สัญญาณ เซลล์ไซโตไคน์และเคโมไคน์ซึ่งมีบทบาทสำคัญในการตอบสนองทางภูมิคุ้มกันการส่งสัญญาณแบบจุกซ์ตาครีนผ่านการสัมผัสโดยตรงของเยื่อหุ้มเซลล์ยังเกิดขึ้นระหว่างตัวเซลล์ประสาทและกระบวนการเคลื่อนที่ของไมโครเกลียทั้งในระหว่างการพัฒนา[ 17 ]และในสมองของผู้ใหญ่[ 18 ]
พาราครีน

ในการส่งสัญญาณแบบพาราครีนเซลล์จะสร้างสัญญาณเพื่อกระตุ้นให้เกิดการเปลี่ยนแปลงในเซลล์ใกล้เคียง ส่งผลให้พฤติกรรมของเซลล์เหล่านั้นเปลี่ยนแปลงไป โมเลกุลส่งสัญญาณที่เรียกว่าปัจจัยพาราครีนจะแพร่กระจายในระยะทางค่อนข้างสั้น (การออกฤทธิ์เฉพาะที่) ซึ่งแตกต่างจากการส่งสัญญาณของเซลล์โดยปัจจัยเอนโดครีน ฮอร์โมน ซึ่งเดินทางในระยะทางที่ไกลกว่ามากผ่านระบบไหลเวียนโลหิตการปฏิสัมพันธ์แบบจุกซ์ตาครีน และการส่งสัญญาณแบบออโตครีน เซลล์ที่สร้างปัจจัยพาราครีนจะหลั่งปัจจัยเหล่านั้นออกสู่ สิ่งแวดล้อม ภายนอกเซลล์โดยตรง จากนั้นปัจจัยจะเดินทางไปยังเซลล์ใกล้เคียง ซึ่งความเข้มข้นของปัจจัยที่ได้รับจะเป็นตัวกำหนดผลลัพธ์ อย่างไรก็ตาม ระยะทางที่แน่นอนที่ปัจจัยพาราครีนสามารถเดินทางได้นั้นยังไม่เป็นที่แน่ชัด
สัญญาณพาราครีน เช่นกรดเรติโนอิกจะกำหนดเป้าหมายเฉพาะเซลล์ที่อยู่ใกล้เคียงกับเซลล์ที่ปล่อยสัญญาณเท่านั้น[ 19 ]สารสื่อประสาทถือเป็นอีกตัวอย่างหนึ่งของสัญญาณพาราครีน
โมเลกุลส่งสัญญาณบางชนิดสามารถทำหน้าที่ได้ทั้งเป็นฮอร์โมนและสารสื่อประสาท ตัวอย่างเช่นเอพิเนฟรินและ นอร์ เอพิเนฟรินสามารถทำหน้าที่เป็นฮอร์โมนได้เมื่อถูกปล่อยออกมาจากต่อมหมวกไตและถูกลำเลียงไปยังหัวใจผ่านทางกระแสเลือด นอร์เอพิเนฟรินยังสามารถผลิตโดยเซลล์ประสาทเพื่อทำหน้าที่เป็นสารสื่อประสาทภายในสมองได้อีก ด้วย [ 20 ]เอสโตรเจนสามารถถูกปล่อยออกมาจากรังไข่และทำหน้าที่เป็นฮอร์โมนหรือออกฤทธิ์เฉพาะที่ผ่านการส่งสัญญาณแบบพาราครีนหรือออโตครีน[ 21 ]
แม้ว่าการส่งสัญญาณแบบพาราครีนจะก่อให้เกิดการตอบสนองที่หลากหลายในเซลล์ที่ถูกเหนี่ยวนำ แต่ปัจจัยพาราครีนส่วนใหญ่ใช้ชุดตัวรับและเส้นทางที่ค่อนข้างเรียบง่าย ในความเป็นจริงอวัยวะ ต่างๆ ในร่างกาย แม้กระทั่งระหว่างสายพันธุ์ต่างๆ ก็เป็นที่ทราบกันดีว่าใช้ชุดปัจจัยพาราครีนที่คล้ายคลึงกันในการพัฒนาที่แตกต่างกัน[ 22 ]ตัว รับและเส้นทาง ที่มีการอนุรักษ์สูงสามารถจัดกลุ่มเป็นสี่ตระกูลหลักตามโครงสร้างที่คล้ายคลึงกัน ได้แก่ ตระกูลปัจจัยการเจริญเติบโตของไฟโบรบลาสต์ (FGF) ตระกูลเฮดจ์ฮ็อก ตระกูล Wntและตระกูล TGF-βการจับกันของปัจจัยพาราครีนกับตัวรับที่เกี่ยวข้องจะเริ่มต้น ลำดับ การส่งสัญญาณทำให้เกิดการตอบสนองที่แตกต่างกัน
ต่อมไร้ท่อ

สัญญาณจากระบบ ต่อมไร้ท่อเรียกว่าฮอร์โมนฮอร์โมนผลิตโดยเซลล์ต่อมไร้ท่อและเดินทางผ่านทางเลือดไปยังทุกส่วนของร่างกาย ความจำเพาะของการส่งสัญญาณสามารถควบคุมได้หากเฉพาะบางเซลล์เท่านั้นที่สามารถตอบสนองต่อฮอร์โมนชนิดใดชนิดหนึ่งได้ การส่งสัญญาณจากระบบต่อมไร้ท่อเกี่ยวข้องกับการปล่อยฮอร์โมนจากต่อม ภายใน ของสิ่งมีชีวิตโดยตรงเข้าสู่ระบบไหลเวียนโลหิตเพื่อควบคุมอวัยวะเป้าหมายที่อยู่ห่างไกล ในสัตว์มีกระดูกสันหลังไฮโปทาลามัสเป็นศูนย์ควบคุมประสาทสำหรับระบบต่อมไร้ท่อทั้งหมด ในมนุษย์ต่อมไร้ท่อที่สำคัญคือต่อมไทรอยด์และต่อมหมวกไตการศึกษาเกี่ยวกับระบบต่อมไร้ท่อและความผิดปกติของระบบนี้เรียกว่าวิทยาต่อมไร้ท่อ
ตัวรับ

เซลล์รับข้อมูลจากเซลล์ข้างเคียงผ่านโปรตีนกลุ่มหนึ่งที่เรียกว่าตัวรับ (receptors ) ตัวรับอาจจับกับโมเลกุลบางชนิด ( ligands ) หรืออาจมีปฏิสัมพันธ์กับปัจจัยทางกายภาพ เช่น แสง แรงกล อุณหภูมิ ความดัน เป็นต้น การรับสัญญาณเกิดขึ้นเมื่อเซลล์เป้าหมาย (เซลล์ใดๆ ที่มีโปรตีนตัวรับจำเพาะต่อโมเลกุลสัญญาณ) ตรวจจับสัญญาณ ซึ่งโดยปกติอยู่ในรูปของโมเลกุลขนาดเล็กที่ละลายน้ำได้ ผ่านการจับกับโปรตีนตัวรับบนพื้นผิวเซลล์ หรือเมื่ออยู่ภายในเซลล์แล้ว โมเลกุลสัญญาณสามารถจับกับตัวรับภายในเซลล์องค์ประกอบอื่นๆ หรือกระตุ้น การทำงาน ของเอนไซม์ (เช่น ก๊าซ) ดังเช่นในการส่งสัญญาณภายในเซลล์ (intracrine signaling)
โมเลกุลส่งสัญญาณจะโต้ตอบกับเซลล์เป้าหมายโดยทำหน้าที่เป็นลิแกนด์ให้กับตัวรับบนพื้นผิวเซลล์และ/หรือโดยการเข้าสู่เซลล์ผ่านเยื่อหุ้มเซลล์หรือเอนโดไซโทซิสเพื่อการส่งสัญญาณภายในเซลล์ โดยทั่วไปแล้วจะส่งผลให้เกิดการกระตุ้นตัวส่งสัญญาณรองซึ่งนำไปสู่ผลกระทบทางสรีรวิทยาต่างๆ ในสัตว์เลี้ยงลูกด้วยนมหลายชนิด เซลล์ ตัวอ่อน ในระยะแรก จะแลกเปลี่ยนสัญญาณกับเซลล์ของมดลูก[ 23 ]ในระบบทางเดินอาหารของ มนุษย์ แบคทีเรียจะแลกเปลี่ยนสัญญาณซึ่งกันและกันและกับเซลล์เยื่อบุผิวและระบบภูมิคุ้มกัน ของมนุษย์ [ 24 ]สำหรับยีสต์Saccharomyces cerevisiaeในระหว่างการผสมพันธุ์เซลล์บางเซลล์จะส่งสัญญาณเปปไทด์ ( ฟีโรโมน ปัจจัยการผสมพันธุ์ ) เข้าสู่สิ่งแวดล้อม เปปไทด์ปัจจัยการผสมพันธุ์อาจจับกับตัวรับบนพื้นผิวเซลล์ของเซลล์ยีสต์อื่นๆ และกระตุ้นให้เซลล์เหล่านั้นเตรียมพร้อมสำหรับการผสมพันธุ์[ 25 ]
ตัวรับบนพื้นผิวเซลล์
ตัวรับบนพื้นผิวเซลล์มีบทบาทสำคัญในระบบชีวภาพของสิ่งมีชีวิตเซลล์เดียวและหลายเซลล์ และการทำงานผิดปกติหรือความเสียหายของโปรตีนเหล่านี้เกี่ยวข้องกับโรคมะเร็ง โรคหัวใจ และโรคหอบหืด[ 26 ] ตัวรับ แบบทรานส์เมม เบรน เหล่านี้สามารถส่งข้อมูลจากภายนอกเซลล์ไปยังภายในได้ เนื่องจากมีการเปลี่ยนแปลงโครงสร้างเมื่อลิแกนด์เฉพาะจับกับมัน มีสามประเภทหลัก ได้แก่ ตัวรับที่เชื่อมโยง กับช่องไอออนตัวรับที่เชื่อมโยงกับโปรตีน Gและ ตัว รับ ที่เชื่อมโยงกับเอนไซม์
ตัวรับที่เชื่อมโยงกับช่องไอออน

ตัวรับที่เชื่อมโยงกับช่องไอออนเป็นกลุ่มของ โปรตีน ช่องไอออนแบบทรานส์เมมเบรน ซึ่งเปิดเพื่อให้ ไอออนเช่นNa + , K + , Ca2 +และ/หรือCl−ผ่านเยื่อหุ้มเซลล์ได้เมื่อมีการจับกับสารสื่อสารทางเคมี (เช่นลิแกนด์ ) เช่นสารสื่อประสาท[ 27 ] [ 28 ] [ 29 ]
เมื่อเซลล์ประสาทก่อนซิแนปส์ถูกกระตุ้น มันจะปล่อยสารสื่อประสาทจากถุงบรรจุสารสื่อประสาทเข้าไปในช่องว่างซิแนปส์จากนั้นสารสื่อประสาทจะจับกับตัวรับที่อยู่บนเซลล์ประสาทหลังซิแนปส์หากตัวรับเหล่านี้เป็นช่องไอออนที่ควบคุมด้วยลิแกนด์ (LICs) การเปลี่ยนแปลงโครงสร้างที่เกิดขึ้นจะเปิดช่องไอออน ซึ่งนำไปสู่การไหลของไอออนผ่านเยื่อหุ้มเซลล์ ซึ่งจะส่งผลให้เกิดการลดศักย์ไฟฟ้า (depolarization ) สำหรับการตอบสนองของตัวรับแบบกระตุ้น หรือการเพิ่มศักย์ไฟฟ้า (hyperpolarization ) สำหรับการตอบสนองแบบยับยั้ง
โปรตีนตัวรับเหล่านี้โดยทั่วไปประกอบด้วยโดเมนอย่างน้อยสองโดเมนที่แตกต่างกัน ได้แก่ โดเมนที่อยู่ภายในเยื่อหุ้มเซลล์ซึ่งรวมถึงรูพรุนของไอออน และโดเมนภายนอกเซลล์ซึ่งรวมถึงตำแหน่งการจับกับลิแกนด์ ( ตำแหน่งการจับแบบ อัลโลสเตอริก ) ความเป็นโมดูลาร์นี้ทำให้สามารถใช้วิธี "แบ่งและพิชิต" ในการค้นหาโครงสร้างของโปรตีน (โดยการตกผลึกแต่ละโดเมนแยกกัน) หน้าที่ของตัวรับดังกล่าวที่อยู่บริเวณไซแนปส์คือการแปลงสัญญาณเคมีของ สารสื่อประสาทที่ปล่อยออกมาจากเซลล์ประสาท ก่อน ไซแนปส์ โดยตรงและรวดเร็วมากให้กลายเป็น สัญญาณไฟฟ้าในเซลล์ ประสาทหลังไซ แนปส์ LIC หลายชนิดยังถูกปรับเปลี่ยนเพิ่มเติมโดยลิแกนด์แบบอัลโลสเตอริก ตัวบล็อกช่องไอออนหรือศักย์ไฟฟ้าของเยื่อหุ้มเซลล์ LIC ถูกจัดกลุ่มออกเป็นสามซูเปอร์แฟมิลีที่ไม่มีความสัมพันธ์ทางวิวัฒนาการ ได้แก่ตัวรับแบบไซโคลสปอนซีสตัวรับกลูตาเมตแบบไอโอโนโทรปิกและ ช่องที่ควบคุมด้วย ATP
ตัวรับที่เชื่อมต่อกับโปรตีนจี

ตัวรับที่เชื่อมต่อกับโปรตีน Gเป็นกลุ่มโปรตีนขนาดใหญ่ที่มีความสัมพันธ์กันในเชิงวิวัฒนาการซึ่งเป็นตัวรับบนพื้นผิวเซลล์ที่ตรวจจับโมเลกุลภายนอกเซลล์และกระตุ้นการตอบสนองของเซลล์ เมื่อเชื่อมต่อกับโปรตีน Gแล้ว จึงเรียกอีกอย่างว่าตัวรับเจ็ดทรานส์เมมเบรน เนื่องจากพวกมันผ่านเยื่อหุ้มเซลล์เจ็ดครั้ง โปรตีน G ทำหน้าที่เป็น "ตัวกลาง" ในการถ่ายทอดสัญญาณจากตัวรับที่ถูกกระตุ้นไปยังเป้าหมาย และด้วยเหตุนี้จึงควบคุมโปรตีนเป้าหมายนั้นโดยอ้อม[ 30 ] ลิแกนด์สามารถจับกับปลาย N-terminus และลูปภายนอกเซลล์ (เช่น ตัวรับกลูตาเมต) หรือกับตำแหน่งการจับภายในเกลียวทรานส์เมมเบรน (ตระกูลโรดอปซิน) พวกมันทั้งหมดถูกกระตุ้นโดยสารกระตุ้นแม้ว่าจะสามารถสังเกตการกระตุ้นตัวเองโดยธรรมชาติของตัวรับที่ว่างเปล่าได้เช่นกัน[ 30 ]
ตัวรับที่เชื่อมต่อกับโปรตีน G พบได้เฉพาะในยูคาริโอตรวมถึงยีสต์โคอาโนแฟลเจลเลต [ 31 ]และสัตว์ลิแกนด์ที่จับและกระตุ้นตัวรับเหล่านี้ ได้แก่ สารประกอบที่ไวต่อแสงกลิ่นฟีโรโมนฮอร์โมนและสารสื่อประสาทและมีขนาดแตกต่างกันไปตั้งแต่โมเลกุลขนาดเล็กไปจนถึงเปปไทด์และโปรตีน ขนาดใหญ่ ตัวรับที่เชื่อมต่อกับโปรตีน G มีส่วนเกี่ยวข้องกับโรคต่างๆ มากมาย
มีเส้นทางการส่งสัญญาณหลักสองเส้นทางที่เกี่ยวข้องกับตัวรับที่เชื่อมต่อกับโปรตีน G ได้แก่เส้นทางการส่งสัญญาณ cAMPและเส้นทางการส่งสัญญาณฟอส ฟา ติดิลอิโนซิ ทอล [ 32 ]เมื่อลิแกนด์จับกับ GPCR จะทำให้เกิดการเปลี่ยนแปลงโครงสร้างใน GPCR ซึ่งทำให้สามารถทำหน้าที่เป็นตัวแลกเปลี่ยนนิวคลีโอไทด์กัวนีน (GEF) จากนั้น GPCR สามารถกระตุ้นโปรตีน G ที่เกี่ยวข้องได้ โดยการแลกเปลี่ยนGDPที่จับกับโปรตีน G กับGTPหน่วยย่อย α ของโปรตีน G พร้อมกับ GTP ที่จับอยู่ สามารถแยกตัวออกจากหน่วยย่อย β และ γ เพื่อส่งผลต่อโปรตีนส่งสัญญาณภายในเซลล์หรือโปรตีนเป้าหมายโดยตรง ขึ้นอยู่กับชนิดของหน่วยย่อย α ( G αs , G αi/o , G αq/11 , G α12/13 ) [ 33 ] : 1160
ตัวรับที่เชื่อมโยงกับโปรตีน G เป็นเป้าหมายยา ที่สำคัญ และประมาณ 34% [ 34 ]ของยาทั้งหมดที่ได้รับการอนุมัติจากสำนักงานคณะกรรมการอาหารและยา (FDA) มีเป้าหมายที่สมาชิก 108 ตัวในกลุ่มนี้ ปริมาณการขายทั่วโลกของยาเหล่านี้คาดว่าจะอยู่ที่ 180 พันล้านดอลลาร์สหรัฐในปี 2018 [ 34 ]คาดว่า GPCRs เป็นเป้าหมายของยาประมาณ 50% ที่วางจำหน่ายอยู่ในปัจจุบัน ส่วนใหญ่เป็นเพราะเกี่ยวข้องกับเส้นทางการส่งสัญญาณที่เกี่ยวข้องกับโรคต่างๆ มากมาย เช่น โรคทางจิต โรคเมตาบอลิซึมรวมถึงความผิดปกติของต่อมไร้ท่อ โรคภูมิคุ้มกันรวมถึงการติดเชื้อไวรัส โรคหัวใจและหลอดเลือด โรคอักเสบ ความผิดปกติของระบบประสาทสัมผัส และมะเร็ง ความสัมพันธ์ที่ค้นพบมานานแล้วระหว่าง GPCRs กับสารภายในและภายนอกร่างกายหลายชนิด ซึ่งส่งผลให้เกิดการบรรเทาอาการปวด เป็นอีกหนึ่งสาขาการวิจัยทางเภสัชกรรมที่กำลังพัฒนาอย่างรวดเร็ว[ 30 ]
ตัวรับที่เชื่อมโยงกับเอนไซม์
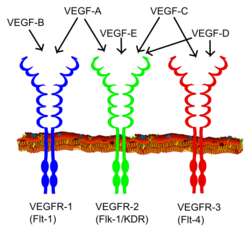
ตัวรับที่เชื่อมโยงกับเอนไซม์ (หรือตัวรับเร่งปฏิกิริยา) เป็นตัวรับแบบทรานส์เมมเบรนซึ่งเมื่อถูกกระตุ้นโดยลิแกนด์ ภายนอกเซลล์ จะทำให้เกิด กิจกรรม ของเอนไซม์ที่ด้านภายในเซลล์[ 35 ]ดังนั้น ตัวรับเร่งปฏิกิริยาจึงเป็นโปรตีนเมมเบรนแบบบูรณาการที่มีทั้งหน้าที่ของเอนไซม์เร่งปฏิกิริยาและตัวรับ[ 36 ]
พวกมันมีโดเมนสำคัญสองโดเมน ได้แก่ โดเมนจับลิแกนด์ภายนอกเซลล์และโดเมนภายในเซลล์ ซึ่งมีหน้าที่เร่งปฏิกิริยา และมีเกลียวทรานส์เมมเบรนเดี่ยว[ 10 ]โมเลกุลส่งสัญญาณจะจับกับตัวรับที่ด้านนอกของเซลล์และทำให้เกิดการเปลี่ยนแปลงโครงสร้างบนหน้าที่เร่งปฏิกิริยาที่อยู่บนตัวรับภายในเซลล์[ 37 ] [ 38 ]ตัวอย่างของกิจกรรมเอนไซม์ ได้แก่:
- ตัวรับไทโรซีนไคเนสเช่นตัวรับปัจจัยการเจริญเติบโตของไฟโบรบลาสต์ตัวรับที่เชื่อมโยงกับเอนไซม์ส่วนใหญ่เป็นประเภทนี้[ 39 ]
- โปรตีนตัวรับเซริน/ทรีโอนีนไคเนสเช่นในโปรตีนสร้างกระดูก[ 40 ]
- กัวนิเลตไซเคลสเช่นเดียวกับตัวรับปัจจัยนาทริยูเรติกในหัวใจห้องบน[ 41 ]
ตัวรับภายในเซลล์
ตัวรับภายในเซลล์มีอยู่ทั่วไปในไซโตพลาสซึม นิวเคลียส หรืออาจจับกับออร์แกเนลล์หรือเยื่อหุ้มเซลล์ ตัวอย่างเช่น การมีอยู่ของ ตัวรับใน นิวเคลียสและไมโตคอนเดรียได้รับการบันทึกไว้อย่างดี[ 42 ]การจับกันของลิแกนด์กับตัวรับภายในเซลล์มักจะกระตุ้นให้เกิดการตอบสนองในเซลล์ ตัวรับภายในเซลล์มักมีความจำเพาะในระดับหนึ่ง ซึ่งช่วยให้ตัวรับสามารถเริ่มต้นการตอบสนองบางอย่างเมื่อจับกับลิแกนด์ที่สอดคล้องกัน[ 43 ]ตัวรับภายในเซลล์มักจะทำงานกับโมเลกุลที่ละลายในไขมัน ตัวรับจะจับกับกลุ่มของโปรตีนที่จับกับ DNAเมื่อจับกันแล้ว คอมเพล็กซ์ตัวรับ-ลิแกนด์จะเคลื่อนย้ายไปยังนิวเคลียสซึ่งสามารถเปลี่ยนแปลงรูปแบบการแสดงออกของยีนได้[ 10 ]
ตัวรับฮอร์โมนสเตียรอยด์พบได้ในนิวเคลียส ไซ โตซอลและเยื่อหุ้มพลาสมาของเซลล์เป้าหมาย โดยทั่วไปแล้วจะเป็นตัวรับภายใน เซลล์ (โดยทั่วไปอยู่ในไซโตพลาสซึมหรือนิวเคลียส) และเริ่มต้นการส่งสัญญาณสำหรับฮอร์โมนสเตียรอยด์ซึ่งนำไปสู่การเปลี่ยนแปลงการแสดงออกของยีนในช่วงเวลาหลายชั่วโมงถึงหลายวันตัวรับ ฮอร์โมนสเตียรอยด์ที่ได้รับการศึกษามากที่สุด คือสมาชิกของ กลุ่ม ย่อยตัวรับนิวเคลียร์ 3 (NR3) ซึ่งรวมถึงตัวรับสำหรับเอสโตรเจน (กลุ่ม NR3A) [ 44 ]และ 3-คีโตสเตียรอยด์ (กลุ่ม NR3C) [ 45 ] นอกจากตัวรับนิวเคลียร์แล้วตัวรับที่เชื่อมต่อกับโปรตีน Gและช่องไอออน หลายตัว ยังทำหน้าที่เป็นตัวรับบนพื้นผิวเซลล์สำหรับฮอร์โมนสเตียรอยด์บางชนิด
กลไกการลดระดับตัวรับ
เอนโดไซโทซิสที่อาศัยตัวรับเป็นวิธีทั่วไปในการ "ปิด" ตัวรับการควบคุมเอนโดไซโทซิสแบบลดลงถือเป็นวิธีการลดการส่งสัญญาณของตัวรับ[ 46 ]กระบวนการนี้เกี่ยวข้องกับการจับตัวของลิแกนด์กับตัวรับ ซึ่งจะกระตุ้นให้เกิดการก่อตัวของหลุมเคลือบ หลุมเคลือบจะเปลี่ยนเป็นเวสิเคิลเคลือบและถูกขนส่งไปยังเอนโดโซม
การฟอสโฟรีเลชันของตัวรับเป็นการลดระดับตัวรับอีกประเภทหนึ่ง การเปลี่ยนแปลงทางชีวเคมีสามารถลดความสัมพันธ์ของตัวรับกับลิแกนด์ได้[ 47 ]
การลดความไวของตัวรับเป็นผลมาจากการที่ตัวรับถูกครอบครองเป็นเวลานาน ส่งผลให้เกิดการปรับตัวของตัวรับซึ่งตัวรับจะไม่ตอบสนองต่อโมเลกุลส่งสัญญาณอีกต่อไป ตัวรับหลายตัวมีความสามารถในการเปลี่ยนแปลงเพื่อตอบสนองต่อความเข้มข้นของลิแกนด์[ 48 ]
เส้นทางการส่งสัญญาณ
เมื่อจับกับโมเลกุลส่งสัญญาณ โปรตีนตัวรับจะเปลี่ยนแปลงไปในบางลักษณะและเริ่มต้นกระบวนการส่งสัญญาณ ซึ่งอาจเกิดขึ้นในขั้นตอนเดียวหรือเป็นการเปลี่ยนแปลงต่อเนื่องในลำดับของโมเลกุลที่แตกต่างกัน (เรียกว่าเส้นทางการส่งสัญญาณ) โมเลกุลที่ประกอบเป็นเส้นทางเหล่านี้เรียกว่าโมเลกุลถ่ายทอด กระบวนการหลายขั้นตอนของขั้นตอนการส่งสัญญาณมักประกอบด้วยการกระตุ้นโปรตีนโดยการเพิ่มหรือการกำจัดกลุ่มฟอสเฟต หรือแม้แต่การปล่อยโมเลกุลขนาดเล็กหรือไอออนอื่นๆ ที่สามารถทำหน้าที่เป็นผู้ส่งสารได้ การขยายสัญญาณเป็นหนึ่งในประโยชน์ของลำดับหลายขั้นตอนนี้ ประโยชน์อื่นๆ ได้แก่ โอกาสในการควบคุมที่มากกว่าระบบที่ง่ายกว่า และการปรับแต่งการตอบสนองอย่างละเอียด ทั้งในสิ่งมีชีวิตเซลล์เดียวและหลายเซลล์[ 11 ]
ในบางกรณี การกระตุ้นตัวรับที่เกิดจากการจับกันของลิแกนด์กับตัวรับนั้นเชื่อมโยงโดยตรงกับการตอบสนองของเซลล์ต่อลิแกนด์ ตัวอย่างเช่น สารสื่อประสาทGABAสามารถกระตุ้นตัวรับบนพื้นผิวเซลล์ซึ่งเป็นส่วนหนึ่งของช่องไอออนการจับกันของ GABA กับตัวรับGABA Aบนเซลล์ประสาทจะเปิดช่องไอออนที่เลือกเฉพาะคลอไรด์ซึ่งเป็นส่วนหนึ่งของตัวรับ การกระตุ้นตัวรับ GABA Aทำให้ไอออนคลอไรด์ที่มีประจุลบเคลื่อนที่เข้าไปในเซลล์ประสาท ซึ่งยับยั้งความสามารถของเซลล์ประสาทในการสร้างศักย์ไฟฟ้าอย่างไรก็ตาม สำหรับตัวรับบนพื้นผิวเซลล์จำนวนมาก ปฏิสัมพันธ์ระหว่างลิแกนด์กับตัวรับไม่ได้เชื่อมโยงโดยตรงกับการตอบสนองของเซลล์ ตัวรับที่ถูกกระตุ้นจะต้องมีปฏิสัมพันธ์กับโปรตีนอื่นๆ ภายในเซลล์ก่อนที่ จะเกิดผล ทางสรีรวิทยา ขั้นสุดท้าย ของลิแกนด์ต่อพฤติกรรมของเซลล์ บ่อยครั้งที่พฤติกรรมของโปรตีนในเซลล์หลายตัวที่มีปฏิสัมพันธ์กันจะเปลี่ยนแปลงไปหลังจากการกระตุ้นตัวรับ การเปลี่ยนแปลงของเซลล์ทั้งหมดที่เกิดจากการกระตุ้นตัวรับเรียกว่ากลไกหรือเส้นทางการส่งสัญญาณ[ 49 ]

เส้นทางการส่งสัญญาณที่ซับซ้อนกว่าคือเส้นทาง MAPK/ERK ซึ่งเกี่ยวข้องกับการเปลี่ยนแปลงปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีนภายในเซลล์ที่ถูกกระตุ้นโดยสัญญาณภายนอก[ 10 ]ปัจจัยการเจริญเติบโตหลายชนิดจับกับตัวรับที่พื้นผิวเซลล์และกระตุ้นให้เซลล์ดำเนินไปตามวงจรเซลล์และแบ่งตัว ตัวรับเหล่านี้หลายตัวเป็นไคเนสที่เริ่มฟอสโฟรีเลตตัวเองและโปรตีนอื่นๆ เมื่อจับกับลิแกนด์การฟอสโฟรีเลต นี้ สามารถสร้างตำแหน่งการจับสำหรับโปรตีนที่แตกต่างกันและทำให้เกิดปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีน ในกรณีนี้ ลิแกนด์ (เรียกว่าปัจจัยการเจริญเติบโตของผิวหนังหรือ EGF) จับกับตัวรับ (เรียกว่าEGFR ) [ 10 ]ซึ่งจะกระตุ้นให้ตัวรับฟอสโฟรีเลตตัวเอง ตัวรับที่ถูกฟอสโฟรีเลตจะจับกับโปรตีนอะแดปเตอร์ ( GRB2 ) ซึ่งเชื่อมโยงสัญญาณกับกระบวนการส่งสัญญาณปลายน้ำต่อไป[ 10 ]ตัวอย่างเช่น หนึ่งในเส้นทางการส่งสัญญาณที่ถูกกระตุ้นเรียกว่า เส้นทาง มิทโทเจนแอคติเวตโปรตีนไคเนส (MAPK) [ 10 ]ส่วนประกอบการส่งสัญญาณที่ติดป้ายว่า "MAPK" ในเส้นทางนี้เดิมเรียกว่า "ERK" ดังนั้นเส้นทางนี้จึงเรียกว่าเส้นทาง MAPK/ERKโปรตีน MAPK เป็นเอนไซม์โปรตีนไคเนสที่สามารถเชื่อมฟอสเฟตเข้ากับโปรตีนเป้าหมาย เช่นปัจจัยการถอดรหัสMYCและด้วยเหตุนี้จึงเปลี่ยนแปลงการถอดรหัสยีนและในที่สุดก็ส่งผลต่อความก้าวหน้าของวงจรเซลล์ โปรตีนในเซลล์จำนวนมากถูกกระตุ้นที่ปลายน้ำของตัวรับปัจจัยการเจริญเติบโต (เช่น EGFR) ที่เริ่มต้นเส้นทางการส่งสัญญาณนี้[ 50 ] [ 10 ]
เส้นทางการส่งสัญญาณบางเส้นทางตอบสนองแตกต่างกัน ขึ้นอยู่กับปริมาณสัญญาณที่เซลล์ได้รับ ตัวอย่างเช่นโปรตีนเฮดจ์ฮ็อกจะกระตุ้นยีนที่แตกต่างกัน ขึ้นอยู่กับปริมาณของโปรตีนเฮดจ์ฮ็อกที่มีอยู่[ 51 ]
เส้นทางการส่งสัญญาณแบบหลายองค์ประกอบที่ซับซ้อนเปิดโอกาสให้เกิดการป้อนกลับ การขยายสัญญาณ และการโต้ตอบภายในเซลล์เดียวระหว่างสัญญาณและเส้นทางการส่งสัญญาณหลายเส้นทาง[ 10 ]
การตอบสนองของเซลล์ที่เฉพาะเจาะจงเป็นผลมาจากสัญญาณที่ส่งผ่านในขั้นตอนสุดท้ายของการส่งสัญญาณของเซลล์ การตอบสนองนี้โดยพื้นฐานแล้วอาจเป็นกิจกรรมของเซลล์ใดๆ ที่มีอยู่ในร่างกาย มันสามารถกระตุ้นการจัดเรียงใหม่ของโครงร่างเซลล์หรือแม้กระทั่งการเร่งปฏิกิริยาโดยเอนไซม์ ขั้นตอนทั้งสามของการส่งสัญญาณของเซลล์ทั้งหมดนี้ทำให้มั่นใจได้ว่าเซลล์ที่ถูกต้องจะทำงานตามที่ได้รับคำสั่ง ในเวลาที่เหมาะสม และประสานกับเซลล์อื่นๆ และหน้าที่ของพวกมันเองภายในสิ่งมีชีวิต[ 10 ]ในท้ายที่สุด จุดสิ้นสุดของเส้นทางสัญญาณนำไปสู่การควบคุมกิจกรรมของเซลล์ การตอบสนองนี้สามารถเกิดขึ้นได้ในนิวเคลียสหรือในไซโตพลาสซึมของเซลล์ เส้นทางสัญญาณส่วนใหญ่ควบคุมการสังเคราะห์โปรตีนโดยการเปิดและปิดยีนบางตัวในนิวเคลียส[ 52 ]
ในสิ่งมีชีวิตเซลล์เดียว เช่น แบคทีเรีย การส่งสัญญาณสามารถใช้เพื่อ 'กระตุ้น' เพื่อนร่วมเซลล์จากสภาวะพักตัวเพิ่มความรุนแรงป้องกันแบคทีริโอเฟจเป็นต้น[ 53 ]ในการรับรู้จำนวนประชากร ซึ่งพบได้ในแมลงสังคมเช่นกัน ความหลากหลายของสัญญาณแต่ละสัญญาณมีศักยภาพที่จะสร้างวงจรป้อนกลับเชิงบวก ทำให้เกิดการตอบสนองที่ประสานกัน ในบริบทนี้ โมเลกุลส่งสัญญาณเรียกว่าออโตอินดิวเซอร์ [ 54 ] [ 55 ] [ 56 ] กลไกการส่งสัญญาณนี้อาจมีส่วนเกี่ยวข้องกับวิวัฒนาการจากสิ่งมีชีวิตเซลล์เดียวไปสู่สิ่งมีชีวิตหลายเซลล์[ 54 ] [ 57 ]แบคทีเรียยังใช้การส่งสัญญาณแบบพึ่งพาการสัมผัส โดยเฉพาะอย่างยิ่งเพื่อจำกัดการเจริญเติบโตของพวกมัน[ 58 ]
โมเลกุลส่งสัญญาณที่ใช้โดยสิ่งมีชีวิตหลายเซลล์มักเรียกว่าฟีโรโมนพวกมันมีจุดประสงค์ต่างๆ เช่น การเตือนภัยอันตราย การบ่งชี้แหล่งอาหาร หรือช่วยในการสืบพันธุ์[ 59 ]
การตอบสนองของเซลล์ในระยะสั้น
| ตระกูลตัวรับ | ตัวอย่างของลิแกนด์/ตัวกระตุ้น (วงเล็บ: ตัวรับสำหรับลิแกนด์/ตัวกระตุ้น) | ตัวอย่างของตัวกระตุ้น | ผลกระทบต่อเนื่องอื่นๆ |
|---|---|---|---|
| ช่องไอออนที่ควบคุมด้วยลิแกนด์ | อะเซทิลโคลีน (เช่นตัวรับอะเซทิลโคลีนชนิดนิโคตินิก ) | การเปลี่ยนแปลงการซึมผ่านของเยื่อหุ้มเซลล์ | การเปลี่ยนแปลงศักย์เยื่อหุ้มเซลล์ |
| ตัวรับเจ็ดเกลียว | แสง ( โรดอปซิน ), โดปามีน ( ตัวรับโดปามีน ), GABA ( ตัวรับ GABA ), โปรสตาแกลนดิน ( ตัวรับโปรสตาแกลนดิน ) เป็นต้น | โปรตีนจีแบบไตรเมอร์ | อะดีนิเลตไซเคลส , ซีจีเอ็มพีฟอสโฟไดเอสเทอเรส , ช่องไอออนที่ควบคุมโดยโปรตีนจีเป็นต้น |
| สององค์ประกอบ | ตัวกระตุ้นที่หลากหลาย | ฮิสติดีนไคเนส | ตัวควบคุมการตอบสนอง - การเคลื่อนที่ของแฟลเจลลา การแสดงออกของยีน |
| เมมเบรน กัวนิลลิล ไซเคลส | เปปไทด์เอเทรียลนาทริยูเรติก , เปปไทด์จากไข่เม่นทะเล เป็นต้น | ซีจีเอ็มพี | การควบคุมไคเนสและช่องสัญญาณ - การทำงานที่หลากหลาย |
| ไซโตพลาสมิก กัวนิลไซเคลส | ไนตริกออกไซด์ ( ตัวรับไนตริกออกไซด์ ) | ซีจีเอ็มพี | การควบคุมช่องสัญญาณ cGMP และไคเนส |
| อินทิกริน | ไฟโบรเนคตินโปรตีนเมทริกซ์นอกเซลล์ชนิดอื่น | ไทโรซีนไคเนสที่ไม่ใช่ตัวรับ | การตอบสนองที่หลากหลาย |
การควบคุมกิจกรรมของยีน
| ผมหยิกฟู (ตัวรับ 7Helix ชนิดพิเศษ) | วนท์ | ผมยุ่งเหยิง , แอ็กซิน - เอพีซี, จีเอสเค3-เบต้า - เบต้าแคทเทนิน | การแสดงออกของยีน |
| สององค์ประกอบ | ตัวกระตุ้นที่หลากหลาย | ฮิสติดีนไคเนส | ตัวควบคุมการตอบสนอง - การเคลื่อนที่ของแฟลเจลลา การแสดงออกของยีน |
| ตัวรับไทโรซีนไคเนส | อินซูลิน ( ตัวรับอินซูลิน ), EGF ( ตัวรับ EGF ), FGF-อัลฟา, FGF-เบตา ฯลฯ ( ตัวรับ FGF ) | Ras , MAP-kinases , PLC , PI3-Kinase | การเปลี่ยนแปลงการแสดงออกของยีน |
| ตัวรับไซโตไคน์ | อีริโทรโปเอติน, ฮอร์โมนการเจริญเติบโต ( ตัวรับฮอร์โมนการเจริญเติบโต ), ไอเอฟเอ็น-แกมมา ( ตัวรับไอเอฟเอ็น-แกมมา ) เป็นต้น | เจเอเค ไคเนส | ปัจจัยถอดรหัส STAT - การแสดงออกของยีน |
| ตัวรับที่เชื่อมโยงกับไทโรซีนไคเนส | คอมเพล็กซ์ MHC-peptide - TCR, แอนติเจน - BCR | ไทโรซีนไคเนสในไซโตพลาสซึม | การแสดงออกของยีน |
| ตัวรับเซริน/ทรีโอนีนไคเนส | แอคติวิน ( ตัวรับแอคติวิน ), อินฮิบิน , โปรตีนสร้างกระดูก ( ตัวรับ BMP ), TGF-เบต้า | ปัจจัยการถอดรหัส Smad | การควบคุมการแสดงออกของยีน |
| ตัวรับที่เชื่อมโยงกับสฟิงโกไมเอลินเนส | IL-1 ( ตัวรับ IL-1 ), TNF ( ตัวรับ TNF ) | เซราไมด์กระตุ้นไคเนส | การแสดงออกของยีน |
| ตัวรับสเตียรอยด์ในไซโตพลาสซึม | ฮอร์โมนสเตียรอยด์ , ฮอร์โมนไทรอยด์ , กรดเรติโนอิกเป็นต้น | ทำงาน/มีปฏิสัมพันธ์กับปัจจัยการถอดรหัส | การแสดงออกของยีน |
วิถีการส่งสัญญาณน็อตช์

Notchเป็นโปรตีนบนพื้นผิวเซลล์ที่ทำหน้าที่เป็นตัวรับ สัตว์มีชุดยีน ขนาดเล็ก ที่เข้ารหัสโปรตีนส่งสัญญาณที่โต้ตอบกับตัวรับ Notch โดยเฉพาะและกระตุ้นการตอบสนองในเซลล์ที่แสดง Notch บนพื้นผิว โมเลกุลที่กระตุ้น (หรือในบางกรณี ยับยั้ง) ตัวรับสามารถจำแนกได้เป็นฮอร์โมนสารสื่อประสาทไซโตไคน์และปัจจัยการเจริญเติบโตซึ่งโดยทั่วไปเรียกว่าลิแกนด์ตัวรับ ปฏิสัมพันธ์ระหว่างลิแกนด์และตัวรับเช่น ปฏิสัมพันธ์ของตัวรับ Notch เป็นที่ทราบกันว่าเป็นปฏิสัมพันธ์หลักที่รับผิดชอบต่อกลไกการส่งสัญญาณและการสื่อสารของเซลล์[ 62 ] Notch ทำหน้าที่เป็นตัวรับสำหรับลิแกนด์ที่แสดงออกบนเซลล์ข้างเคียง ในขณะที่ตัวรับบางตัวเป็นโปรตีนบนพื้นผิวเซลล์ แต่บางตัวก็พบอยู่ภายในเซลล์ ตัวอย่างเช่นเอสโตรเจนเป็น โมเลกุล ที่ไม่ชอบน้ำซึ่งสามารถผ่านชั้นไขมันของเยื่อหุ้มเซลล์ได้ ในฐานะส่วนหนึ่งของระบบต่อมไร้ท่อ ตัว รับเอสโตรเจนภายในเซลล์ จาก เซลล์หลายประเภท(เช่น รังไข่ ต่อมลูกหมาก ปอด ระบบหัวใจและหลอดเลือด และระบบประสาทส่วนกลาง) สามารถถูกกระตุ้นโดยเอสโตรเจนที่ผลิตในรังไข่ได้[ 63 ]
ในกรณีของการส่งสัญญาณผ่าน Notch กลไกการส่งสัญญาณอาจค่อนข้างง่าย ดังแสดงในรูปที่ 2 การกระตุ้น Notch อาจทำให้โปรตีน Notch ถูกเปลี่ยนแปลงโดยโปรตีเอสโปรตีน Notch บางส่วนจะถูกปล่อยออกมาจากเยื่อหุ้มเซลล์และมีส่วนร่วมในการควบคุมยีนการวิจัยการส่งสัญญาณของเซลล์เกี่ยวข้องกับการศึกษาพลวัตเชิงพื้นที่และเวลาของทั้งตัวรับและส่วนประกอบของเส้นทางการส่งสัญญาณที่ถูกกระตุ้นโดยตัวรับในเซลล์ประเภทต่างๆ[ 64 ] [ 65 ]วิธีการใหม่ๆ สำหรับการวิเคราะห์มวลสเปกโทรเมตรีของเซลล์เดี่ยวมีแนวโน้มที่จะช่วยให้สามารถศึกษาการส่งสัญญาณด้วยความละเอียดระดับเซลล์เดี่ยวได้[ 66 ]
ในการส่งสัญญาณ Notchการสัมผัสโดยตรงระหว่างเซลล์ทำให้สามารถควบคุมการแบ่งตัว ของเซลล์ได้อย่างแม่นยำ ในระหว่างการพัฒนาของตัวอ่อนในหนอนCaenorhabditis elegansเซลล์สองเซลล์ของอวัยวะสืบพันธุ์ ที่กำลังพัฒนา แต่ละเซลล์มีโอกาสเท่ากันที่จะแบ่งตัวอย่างสมบูรณ์หรือกลายเป็นเซลล์ต้นกำเนิดมดลูกที่แบ่งตัวต่อไป การเลือกเซลล์ใดที่จะแบ่งตัวต่อไปนั้นถูกควบคุมโดยการแข่งขันของสัญญาณบนพื้นผิวเซลล์ เซลล์หนึ่งจะผลิตโปรตีนบนพื้นผิวเซลล์มากกว่า ซึ่งจะกระตุ้นตัวรับ Notch บนเซลล์ข้างเคียง สิ่งนี้จะกระตุ้นวงจรป้อนกลับหรือระบบที่ลดการแสดงออกของ Notch ในเซลล์ที่จะแบ่งตัวและเพิ่ม Notch บนพื้นผิวของเซลล์ที่ยังคงเป็นเซลล์ต้นกำเนิด[ 67 ]
ดูเพิ่มเติม
- โปรตีนโครงสร้าง
- ชีวสัญศาสตร์
- การรับรู้ระดับเซลล์โมเลกุล
- การสื่อสารข้ามเซลล์ (ชีววิทยา)
- ถุงเยื่อหุ้มชั้นนอกของแบคทีเรีย
- การลำเลียงเวสิเคิลเยื่อหุ้มเซลล์
- ปฏิสัมพันธ์ระหว่างโฮสต์และเชื้อโรค
- กรดเรติโนอิก
- วิถีการส่งสัญญาณ JAK-STAT
- เส้นทางอิมเอ็มดี
- สัญญาณระบุตำแหน่ง
- การสั่น
- พลวัตของโปรตีน
- ชีววิทยาระบบ
- การส่งสัญญาณของไขมัน
- การส่งสัญญาณรีดอกซ์
- การส่งสัญญาณแบบต่อเนื่อง
- เทคโนโลยีการส่งสัญญาณเซลล์ – บริษัทพัฒนาและผลิตแอนติบอดี
- Netpath – แหล่งข้อมูลที่คัดสรรมาอย่างดีเกี่ยวกับเส้นทางการส่งสัญญาณในมนุษย์
- ภาษาเปิดชีววิทยาเชิงสังเคราะห์
- การสร้างเครือข่ายระดับนาโน – การใช้ประโยชน์จากการส่งสัญญาณทางชีวภาพเพื่อสร้างเครือข่ายการสื่อสารเฉพาะกิจภายในร่างกาย
- แบบจำลองโซลิตอนในประสาทวิทยาศาสตร์ – การสื่อสารทางกายภาพผ่านคลื่นเสียงในเยื่อหุ้มเซลล์
- การตอบสนองเชิงเวลา
อ่านเพิ่มเติม
- "เรื่องราวเบื้องลึกของการสื่อสารระหว่างเซลล์" learn.genetics.utah.eduสืบค้นเมื่อ 2018-10-20
- "เมื่อการสื่อสารระหว่างเซลล์ผิดพลาด" learn.genetics.utah.edu . สืบค้นเมื่อ 2018-10-24
ลิงก์ภายนอก
- ฐานข้อมูลปฏิสัมพันธ์ของเส้นทางส่งสัญญาณ NCI-Nature : ข้อมูลที่เชื่อถือได้เกี่ยวกับเส้นทางส่งสัญญาณในเซลล์มนุษย์
- Intercellular+Signaling+Peptides+and+Proteinsที่ US National Library of Medicine Medical Subject Headings (MeSH)
- การสื่อสารระหว่างเซลล์ ใน ระบบพิกัด ทางการแพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
- โครงการเส้นทางการส่งสัญญาณ : ฐานความรู้สำหรับการสร้างสมมติฐานเกี่ยวกับการส่งสัญญาณในเซลล์ สร้างขึ้นโดยใช้ชุดข้อมูลทรานสคริปโตมิกและ ChIP-Seq ที่ได้รับการจัดเก็บและคัดกรองทางชีวภาพ
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การส่งสัญญาณของเซลล์
การส่งสัญญาณของเซลล์ ( cell signalling ใน ภาษาอังกฤษแบบ บริติช ) คือ กระบวนการทางชีววิทยา ที่ เซลล์ มีปฏิสัมพันธ์กับตัวเอง กับเซลล์อื่น ๆ และกับสิ่งแวดล้อม...
ขอบเขตทางอนุกรมวิธาน
ในสิ่งมีชีวิตขนาดเล็กหลาย ชนิดเช่น แบคทีเรีย การรับรู้จำนวนประชากร ช่วยให้แต่ละบุคคลสามารถเริ่มกิจกรรมได้ก็ต่อเมื่อประชากรมีขนาดใหญ่เพียงพอ การส่งสัญญาณระหว่างเซลล์นี้ถูกสังเกตครั้งแรกในแบคทีเรียในทะเล Aliivibrio fischeri ซึ่ง ผลิตแสง...
การสังเคราะห์และการปล่อย
สัญญาณเซลล์จำนวนมากถูกส่งผ่านโมเลกุลที่ถูกปล่อยออกมาจากเซลล์หนึ่งและเคลื่อนที่ไปสัมผัสกับเซลล์อื่น โมเลกุลส่งสัญญาณสามารถอยู่ในกลุ่มสารเคมีหลายประเภท ได้แก่ลิ ปิด ฟอ สโฟลิปิด กรดอะมิโน โมโนอะมีน โปรตีน ไกลโคโปรตีน หรือ ก๊าซ โมเลกุล ส่งสัญญาณ ที่ จับ กับ ตัว...
ประเภท
การส่งสัญญาณแบบออโตครีน เกี่ยวข้องกับเซลล์ที่หลั่งฮอร์โมนหรือสารสื่อสารทางเคมี (เรียกว่าตัวแทนออโตครีน) ที่จับกับตัวรับออโตครีนบนเซลล์เดียวกัน ทำให้เกิดการเปลี่ยนแปลงในเซลล์นั้นเอง [ 15 ] ซึ่งแตกต่างจากการส่งสัญญาณแบบพาราครีน การส่งสัญญาณแบบอินทราครีน...