อ่าน 29 นาที
เฮโมโกลบิน
ฮีโมโกลบิน ( haemoglobin , HbหรือHgb ) เป็นโปรตีนที่มีธาตุเหล็กซึ่งช่วยในการขนส่งออกซิเจนในเม็ดเลือดแดง สัตว์...
เฮโมโกลบิน
| เฮโมโกลบิน | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (เฮเทอโรเตตราเมอร์, ( α β ) 2 ) | |||||||||||||
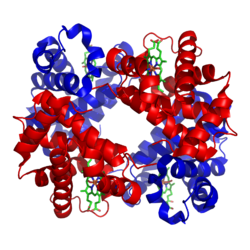 โครงสร้างของฮีโมโกลบินในมนุษย์ หน่วยย่อย โกลบินอัลฟาและเบตา แสดงด้วยสีแดงและสีน้ำเงินตามลำดับ และ กลุ่ม ฮีม ที่มีธาตุเหล็ก แสดงด้วยสีเขียว จากPDB : 1GZX ProteopediaHemoglobin | |||||||||||||
| โปรตีนชนิด | เมทัลโลโปรตีน , โครโมโปรตีน , โกลบูลิน | ||||||||||||
| การทำงาน | การขนส่งออกซิเจน | ||||||||||||
| โคแฟกเตอร์ | ฮีม (4) | ||||||||||||
| |||||||||||||
ฮีโมโกลบิน ( haemoglobin , [ a ] HbหรือHgb ) เป็นโปรตีนที่มีธาตุเหล็กซึ่งช่วยในการขนส่งออกซิเจนในเม็ดเลือดแดง สัตว์ มีกระดูกสันหลังเกือบทั้งหมดมีฮีโมโกลบิน[ 3 ]ยกเว้นเพียงวงศ์ปลาChannichthyidae [ 4 ]ฮีโมโกลบินในเลือดจะนำออกซิเจนจากอวัยวะระบบหายใจ ( ปอดหรือเหงือก ) ไปยังเนื้อเยื่ออื่นๆ ของร่างกาย ซึ่งจะปล่อยออกซิเจนออกมาเพื่อช่วยให้เกิดการหายใจแบบใช้ออกซิเจนซึ่งเป็นพลังงานขับเคลื่อนกระบวนการเผา ผลาญของสัตว์ มนุษย์ที่มีสุขภาพดีจะมีฮีโมโกลบิน 12 ถึง 20 กรัมใน เลือด ทุกๆ 100 มิลลิลิตร ฮีโมโกลบินเป็น เมทัลโลโปรตีนโครโมโปรตีนและโกลบูลิน
ในสัตว์เลี้ยงลูกด้วยนม ฮีโมโกลบินประกอบขึ้นเป็นประมาณ 96% ของ น้ำหนักแห้งของเม็ดเลือดแดง(ไม่รวมน้ำ) และประมาณ 35% ของน้ำหนักรวม (รวมน้ำ) [ 5 ]ฮีโมโกลบินมีความสามารถในการจับออกซิเจนได้ 1.34 มิลลิลิตรของ O 2ต่อกรัม[ 6 ] ซึ่งเพิ่ม ความสามารถในการรับออกซิเจนของเลือดทั้งหมดถึงเจ็ดสิบเท่าเมื่อเทียบกับออกซิเจนที่ละลายอยู่ในพลาสมาของเลือดเพียงอย่างเดียว[ 7 ]โมเลกุลฮีโมโกลบินของสัตว์เลี้ยงลูกด้วยนมสามารถจับและขนส่งโมเลกุลออกซิเจนได้มากถึงสี่โมเลกุล[ 8 ]
ฮีโมโกลบินยังขนส่งก๊าซอื่นๆ ด้วย โดยจะขนส่งคาร์บอนไดออกไซด์ จากการหายใจของร่างกายบางส่วน (ประมาณ 20–25% ของทั้งหมด) [ 9 ]ในรูปของคาร์บามิโนฮีโมโกลบินซึ่ง CO 2จะจับกับโปรตีนฮีมโมเลกุลนี้ยังขนส่งโมเลกุลควบคุมที่สำคัญอย่างไนตริกออกไซด์ที่จับกับ กลุ่ม ไทออลในโปรตีนโกลบิน และปล่อยออกมาพร้อมกับออกซิเจน[ 10 ]
ฮีโมโกลบินยังพบได้ในเซลล์อื่นๆ รวมถึงเซลล์ประสาทโดปามีน A9ของ ซับ สแตนเซียไนกราแมโครฟาจ เซลล์ถุงลม ปอด เยื่อบุผิวเม็ดสีเรตินาเซลล์ตับ เซลล์เมแซงเจียลของไต เซลล์เยื่อบุโพรง มดลูก เซลล์ปากมดลูก และเซลล์เยื่อบุช่องคลอด [ 11 ]ในเนื้อเยื่อเหล่านี้ ฮีโมโกลบินจะดูดซับออกซิเจนที่ไม่จำเป็นในฐานะสารต้านอนุมูลอิสระและควบคุมการเผาผลาญธาตุเหล็ก[ 12 ]กลูโคสที่มากเกินไปในเลือดสามารถจับกับฮีโมโกลบินและเพิ่มระดับฮีโมโกลบิน A1c ได้[ 13 ]
ฮีโมโกลบินและโมเลกุลคล้ายฮีโมโกลบินยังพบได้ในสัตว์ไม่มีกระดูกสันหลัง เชื้อรา และพืชหลายชนิด[ 14 ]ในสิ่งมีชีวิตเหล่านี้ ฮีโมโกลบินอาจขนส่งออกซิเจน หรืออาจขนส่งและควบคุมโมเลกุลและไอออนขนาดเล็กอื่นๆ เช่น คาร์บอนไดออกไซด์ ไนตริกออกไซด์ ไฮโดรเจนซัลไฟด์ และซัลไฟด์ ตัวแปรที่เรียกว่าเลกฮีโมโกลบินทำหน้าที่กำจัดออกซิเจนออกจาก ระบบแบบ ไม่ใช้ออกซิเจนเช่น ปมตรึงไนโตรเจนของ พืช ตระกูลถั่วป้องกันพิษจากออกซิเจน
ภาวะฮีโมโกลบินใน เลือดสูง ซึ่งเป็นรูปแบบหนึ่งของโรคโลหิตจางเกิดจากการแตกตัวของเม็ดเลือดแดงในหลอดเลือดทำให้ฮีโมโกลบินรั่วไหลจากเม็ดเลือดแดงเข้าสู่ พลาสมา ใน เลือด
ประวัติการวิจัย

แม้ว่าเลือดจะทราบกันดีว่าสามารถลำเลียงออกซิเจนได้ตั้งแต่ปี ค.ศ. 1794 เป็นอย่างน้อย[ 16 ] [ 17 ]แต่คุณสมบัติในการลำเลียงออกซิเจนของฮีโมโกลบินนั้นได้รับการอธิบายโดย Hünefeld ในปี ค.ศ. 1840 [ 18 ]ในปี ค.ศ. 1851 นักสรีรวิทยาชาวเยอรมันOtto Funkeได้ตีพิมพ์บทความชุดหนึ่งซึ่งเขาได้อธิบายถึงการปลูกผลึกฮีโมโกลบินโดยการเจือจางเซลล์เม็ดเลือดแดงด้วยตัวทำละลาย เช่น น้ำบริสุทธิ์ แอลกอฮอล์ หรืออีเทอร์อย่างต่อเนื่อง ตามด้วยการระเหยตัวทำละลายออกจากสารละลายโปรตีนที่ได้ช้าๆ[ 19 ] [ 20 ] การเติมออกซิเจนแบบย้อนกลับได้ของฮีโมโกลบินได้รับการอธิบายโดย Felix Hoppe-Seylerในอีกไม่กี่ปีต่อมา[ 21 ]
ในปี ค.ศ. 1825 โยฮันน์ ฟรีดริช เองเกลฮาร์ท ค้นพบว่าอัตราส่วนของเหล็กต่อโปรตีนนั้นเหมือนกันในฮีโมโกลบินของหลายสายพันธุ์[ 22 ] [ 23 ]จากมวลอะตอมของเหล็กที่ทราบ เขาคำนวณมวลโมเลกุลของฮีโมโกลบินได้เป็นn × 16000 ( n = จำนวนอะตอมของเหล็กต่อโมเลกุลของฮีโมโกลบิน ซึ่งปัจจุบันทราบกันดีว่าเท่ากับ 4) ซึ่งเป็นการกำหนดมวลโมเลกุลของโปรตีนครั้งแรก "ข้อสรุปที่รีบร้อน" นี้ทำให้เพื่อนร่วมงานเยาะเย้ย เพราะพวกเขาไม่เชื่อว่าโมเลกุลใดๆ จะมีขนาดใหญ่ได้ขนาดนั้น อย่างไรก็ตามกิลเบิร์ต สมิธสัน แอดแอร์ได้ยืนยันผลลัพธ์ของเองเกลฮาร์ทในปี ค.ศ. 1925 โดยการวัดความดันออสโมติกของสารละลายฮีโมโกลบิน[ 24 ]
ด้วยการพัฒนาของผลึกศาสตร์รังสีเอกซ์ทำให้สามารถไขโครงสร้างโปรตีนได้[ 25 ]ในปี พ.ศ. 2492 แม็กซ์ เพรุตซ์ได้กำหนดโครงสร้างโมเลกุลของฮีโมโกลบิน[ 26 ] [ 27 ]จากผลงานนี้ เขาได้รับรางวัลโนเบลสาขาเคมี ร่วม กับจอห์น เคนดรูว์ ในปี พ.ศ. 2505 ซึ่งเป็นผู้จัดลำดับโปรตีนทรงกลมไมโอโกลบิน[ 25 ] [ 28 ]
บทบาทของฮีโมโกลบินในเลือดได้รับการอธิบายอย่างชัดเจนโดยนักสรีรวิทยา ชาวฝรั่งเศส โคลด แบร์นาร์ด
ชื่อฮีโมโกลบิน (หรือhaemoglobin ) มาจากคำว่าheme (หรือhaem ) และglobinซึ่งสะท้อนให้เห็นว่าแต่ละหน่วยย่อยของฮีโมโกลบินเป็นโปรตีนทรงกลมที่มีกลุ่ม heme ฝังอยู่ แต่ละกลุ่ม heme ประกอบด้วยอะตอมเหล็กหนึ่งอะตอม ซึ่งสามารถจับกับโมเลกุลออกซิเจนหนึ่งโมเลกุลผ่านแรงไดโพลที่เหนี่ยวนำโดยไอออน ฮีโมโกลบินชนิดที่พบมากที่สุดในสัตว์เลี้ยงลูกด้วยนมมีหน่วยย่อยดังกล่าวสี่หน่วย[ 29 ]
พันธุศาสตร์
ฮีโมโกลบินประกอบด้วยหน่วยย่อยของโปรตีน ( โมเลกุล โกลบิน ) ซึ่งเป็นพอลิเปปไทด์ โซ่ยาวที่พับงอของกรดอะมิโน จำเพาะ ซึ่งเป็นตัวกำหนดคุณสมบัติทางเคมีและหน้าที่ของโปรตีน ลำดับกรดอะมิโนของพอลิเปปไทด์ใดๆ นั้นได้มา จาก การถอดรหัสจากส่วนหนึ่งของดีเอ็นเอ ซึ่งก็คือยีนที่ เกี่ยวข้อง
มีมากกว่าหนึ่งยีนฮีโมโกลบิน ในมนุษย์ฮีโมโกลบิน A (ฮีโมโกลบินรูปแบบหลักในผู้ใหญ่) ถูกเข้ารหัสโดยยีนHBA1 , HBA2และHBB [ 30 ] หน่วยย่อยอัลฟา 1 และอัลฟา 2 ถูกเข้ารหัสโดยยีนHBA1และHBA2 ที่อยู่ใกล้กันบนโครโมโซม 16 ตาม ลำดับ ในขณะที่หน่วยย่อยเบตาถูกเข้ารหัสโดยยีนHBBบนโครโมโซม 11 ลำดับกรดอะมิโนของหน่วยย่อยโกลบินมักจะแตกต่างกันระหว่างสายพันธุ์ โดยความแตกต่างจะเพิ่มขึ้นตามระยะทางวิวัฒนาการ ตัวอย่างเช่น ลำดับฮีโมโกลบินที่พบมากที่สุดในมนุษย์ โบโนโบ และชิมแปนซีนั้นเหมือนกันทุกประการ โดยมีสายโปรตีนโกลบินอัลฟาและเบตาที่เหมือนกันทุกประการ[ 31 ] [ 32 ] [ 33 ]ฮีโมโกลบินของมนุษย์และกอริลลาแตกต่างกันในกรดอะมิโนหนึ่งตัวในทั้งสายอัลฟาและเบตา และความแตกต่างเหล่านี้จะมากขึ้นระหว่างสายพันธุ์ที่มีความสัมพันธ์กันน้อยลง
การกลายพันธุ์ในยีนสำหรับฮีโมโกลบินสามารถส่งผลให้เกิดฮีโมโกลบินชนิดต่างๆภายในสายพันธุ์เดียวกันได้ แม้ว่าโดยปกติแล้วจะมีลำดับหนึ่งที่ "พบได้บ่อยที่สุด" ในแต่ละสายพันธุ์ก็ตาม[ 34 ] [ 35 ]การกลายพันธุ์เหล่านี้จำนวนมากไม่ก่อให้เกิดโรค แต่บางส่วนก่อให้เกิดกลุ่มโรคทางพันธุกรรมที่เรียกว่าฮีโมโกลบินโนพาธี โรค ฮีโมโกลบินโนพาธีที่รู้จักกันดีที่สุดคือโรคโลหิตจางเคียวซึ่งเป็นโรคของมนุษย์โรคแรกที่ เข้าใจ กลไกในระดับโมเลกุล กลุ่มโรคที่แยกออกไปอีกกลุ่มหนึ่งที่เรียกว่าธาลัสซีเมียเกี่ยวข้องกับการผลิตฮีโมโกลบินปกติและบางครั้งก็ผิดปกติในปริมาณที่น้อยกว่าปกติ ผ่านปัญหาและการกลายพันธุ์ในการควบคุมยีน โกลบิน โรคเหล่านี้ทั้งหมดทำให้เกิดภาวะโลหิตจาง[ 36 ]
วิวัฒนาการ

โปรตีนโกลบินและยีนของพวกมันจึงมีต้นกำเนิดทางวิวัฒนาการที่เก่าแก่ แม้แต่พืชก็ยังมีโกลบินชนิดหนึ่งที่เรียกว่าเลกฮีโมโกลบินซึ่งมีลำดับร่วมกับฮีโมโกลบินประมาณ 15% แม้ว่าความคล้ายคลึงในระดับนี้อาจเกิดจากความบังเอิญ แต่โครงสร้างสามมิติของโปรตีนทั้งสองชนิดนั้นคล้ายคลึงกันมากจนโดยทั่วไปถือว่าพวกมันมีต้นกำเนิดร่วมกัน ซึ่งได้รับการยืนยันโดยการอนุรักษ์ตำแหน่งของอินทรอนในยีนทั้งสองประเภท[ 37 ]
ความแปรผันในลำดับของฮีโมโกลบิน เช่นเดียวกับโปรตีนอื่นๆ อาจเป็นการปรับตัว ตัวอย่างเช่น พบว่าฮีโมโกลบินสามารถปรับตัวในรูปแบบต่างๆ กับอากาศเบาบางในที่สูง ซึ่งความดันย่อยของออกซิเจนที่ต่ำกว่าจะลดการจับตัวของออกซิเจนกับฮีโมโกลบินเมื่อเทียบกับความดันที่สูงกว่าในระดับน้ำทะเล
การศึกษาล่าสุดเกี่ยวกับหนูเดียร์พบการกลายพันธุ์ในยีนสี่ตัวที่สามารถอธิบายความแตกต่างระหว่างประชากรที่อาศัยอยู่ในพื้นที่สูงและต่ำได้ พบว่ายีนของหนูทั้งสองสายพันธุ์นั้น "แทบจะเหมือนกันทุกประการ ยกเว้นยีนที่ควบคุมความสามารถในการขนส่งออกซิเจนของฮีโมโกลบิน . . . ความแตกต่างทางพันธุกรรมนี้ทำให้หนูที่อาศัยอยู่ในพื้นที่สูงสามารถใช้ประโยชน์จากออกซิเจนได้อย่างมีประสิทธิภาพมากขึ้น" [ 38 ]
วิวัฒนาการของฮีโมโกลบินในสัตว์มีกระดูกสันหลัง
นักวิทยาศาสตร์เห็นพ้องกันว่าเหตุการณ์ที่แยกไมโอโกลบินออกจากฮีโมโกลบินเกิดขึ้นหลังจากปลาแลม เพรย์ แยกตัวออกจากสัตว์มีกระดูกสันหลังที่มีขา กรรไกร [ 39 ]การแยกตัวของไมโอโกลบินและฮีโมโกลบินนี้ทำให้โมเลกุลทั้งสองมีหน้าที่ที่แตกต่างกันและพัฒนาขึ้นได้ โดยไมโอโกลบินเกี่ยวข้องกับการเก็บออกซิเจน ในขณะที่ฮีโมโกลบินมีหน้าที่ในการขนส่งออกซิเจน[ 40 ]ยีนโกลบินแบบ α และ β เข้ารหัสหน่วยย่อยแต่ละหน่วยของโปรตีน[ 30 ]บรรพบุรุษของยีนเหล่านี้เกิดขึ้นจากเหตุการณ์การจำลองแบบอีกครั้งหลังจากบรรพบุรุษร่วมของสัตว์มีขากรรไกรที่สืบเชื้อสายมาจากปลาไม่มีขากรรไกรเมื่อประมาณ 450–500 ล้านปีก่อน[ 39 ]การศึกษาการสร้างบรรพบุรุษใหม่ชี้ให้เห็นว่าบรรพบุรุษก่อนการจำลองแบบของยีน α และ β เป็นไดเมอร์ที่ประกอบด้วยหน่วยย่อยโกลบินที่เหมือนกัน ซึ่งต่อมาได้วิวัฒนาการเพื่อประกอบกันเป็นโครงสร้างเตตระเมอร์หลังจากการจำลองแบบ[ 41 ]การพัฒนาของยีน α และ β ทำให้เกิดศักยภาพที่ฮีโมโกลบินจะประกอบด้วยหน่วยย่อยที่แตกต่างกันหลายหน่วย ซึ่งเป็นองค์ประกอบทางกายภาพที่สำคัญต่อความสามารถของฮีโมโกลบินในการขนส่งออกซิเจน การมีหน่วยย่อยหลายหน่วยช่วยให้ฮีโมโกลบินสามารถจับกับออกซิเจนร่วมกันได้ รวมถึงถูกควบคุมแบบอั ลโลสเตอริกด้วย [ 40 ] [ 41 ]ต่อมา ยีน α ก็ได้เกิดการเพิ่มจำนวนขึ้นเพื่อสร้างยีนHBA1และHBA2 [ 42 ]การเพิ่มจำนวนและการแยกตัวเพิ่มเติมเหล่านี้ได้สร้างยีนโกลบินที่คล้ายกับ α และ β ที่หลากหลาย ซึ่งถูกควบคุมเพื่อให้รูปแบบบางอย่างเกิดขึ้นในระยะต่างๆ ของการพัฒนา[ 40 ]
ปลา
ปลาน้ำแข็งส่วนใหญ่ในวงศ์Channichthyidaeสูญเสียยีนฮีโมโกลบินไปเพื่อปรับตัวให้เข้ากับน้ำเย็น[ 4 ]
นก
ปอดที่มีระบบไหลเวียนโลหิตที่เป็นเอกลักษณ์ของนกยังช่วยส่งเสริมการใช้ออกซิเจนอย่างมีประสิทธิภาพที่ความดันย่อยของ O2 ต่ำการปรับตัวทั้งสองนี้เสริมซึ่งกันและกันและอธิบายถึงประสิทธิภาพที่โดดเด่นของนกในที่สูง[ 43 ]โดยเฉพาะอย่างยิ่งพบในนกฮัมมิ่งเบิร์ดที่อาศัยอยู่ในเทือกเขาแอนดีส นกฮัมมิ่งเบิร์ดใช้พลังงานมากอยู่แล้วจึงมีความต้องการออกซิเจนสูง แต่นกฮัมมิ่งเบิร์ดในเทือกเขาแอนดีสก็สามารถเจริญเติบโตได้ในที่สูง การกลายพันธุ์ที่ไม่ใช่แบบเดียวกันในยีนฮีโมโกลบินของหลายสายพันธุ์ที่อาศัยอยู่ในที่สูง ( Oreotrochilus, A. castelnaudii, C. violifer, P. gigasและA. viridicuada ) ทำให้โปรตีนมีความสัมพันธ์กับอิโนซิทอลเฮกซาฟอสเฟต (IHP) น้อยลง ซึ่งเป็นโมเลกุลที่พบในนกและมีบทบาทคล้ายกับ 2,3-BPG ในมนุษย์ ส่งผลให้สามารถจับออกซิเจนได้ที่ความดันย่อยต่ำลง[ 44 ]
สัตว์เลี้ยงลูกด้วยนม
ฮีโมโกลบิน ของแมมมอธมีการกลายพันธุ์ที่ทำให้สามารถส่งออกซิเจนได้ที่อุณหภูมิต่ำลง ส่งผลให้แมมมอธสามารถอพยพไปยังละติจูดที่สูงขึ้นได้ในช่วงยุคไพลสโตซีน[ 45 ]
การปรับตัวของฮีโมโกลบินยังขยายไปถึงมนุษย์ด้วยเช่นกัน พบว่าอัตราการรอดชีวิตของลูกหลานสูงขึ้นในกลุ่มสตรีชาวทิเบตที่มีจีโนไทป์ที่มีความอิ่มตัวของออกซิเจนสูงซึ่งอาศัยอยู่ที่ระดับความสูง 4,000 เมตร[ 46 ]การคัดเลือกโดยธรรมชาติดูเหมือนจะเป็นแรงผลักดันหลักที่ส่งผลต่อยีนนี้ เนื่องจากอัตราการตายของลูกหลานต่ำกว่าอย่างมีนัยสำคัญในกลุ่มสตรีที่มีความสัมพันธ์ระหว่างฮีโมโกลบินกับออกซิเจนสูงกว่า เมื่อเทียบกับอัตราการตายของลูกหลานจากสตรีที่มีความสัมพันธ์ระหว่างฮีโมโกลบินกับออกซิเจนต่ำ แม้ว่าจีโนไทป์และกลไกที่ทำให้เกิดสิ่งนี้ยังไม่ชัดเจน แต่การคัดเลือกกำลังส่งผลต่อความสามารถของสตรีเหล่านี้ในการจับออกซิเจนในความดันย่อยต่ำ ซึ่งโดยรวมแล้วช่วยให้พวกเธอสามารถรักษากระบวนการเผาผลาญที่สำคัญได้ดียิ่งขึ้น
สังเคราะห์
ฮีโมโกลบิน (Hb) ถูกสังเคราะห์ขึ้นเป็นชุดขั้นตอนที่ซับซ้อน ส่วนของฮีมถูกสังเคราะห์ขึ้นเป็นชุดขั้นตอนในไมโทคอนเดรียและไซโตซอลของเซลล์เม็ดเลือดแดงที่ยังไม่เจริญเต็มที่ ในขณะที่ ส่วนของโปรตีน โกลบินถูกสังเคราะห์โดยไรโบโซมในไซโตซอล[ 47 ]การผลิต Hb ยังคงดำเนินต่อไปในเซลล์ตลอดช่วงการพัฒนาในระยะแรก ตั้งแต่โปรอีริโทรบลาสต์ไปจนถึง เรติ คิวโลไซต์ในไขกระดูกณ จุดนี้นิวเคลียสจะหายไปในเซลล์เม็ดเลือดแดงของสัตว์เลี้ยงลูกด้วยนม แต่ไม่ใช่ในนกและสัตว์ชนิดอื่นๆ อีกหลายชนิด แม้หลังจากที่นิวเคลียสหายไปในสัตว์เลี้ยงลูกด้วยนมแล้วRNA ไรโบโซมที่เหลืออยู่ ก็ยังช่วยให้สามารถสังเคราะห์ Hb ต่อไปได้จนกระทั่งเรติคิวโลไซต์สูญเสีย RNA ไปไม่นานหลังจากเข้าสู่ระบบหลอดเลือด (RNA สังเคราะห์ฮีโมโกลบินนี้เองที่ทำให้เรติคิวโลไซต์มีลักษณะเป็นตาข่ายและเป็นที่มาของชื่อ) [ 48 ]
โครงสร้างของฮีม

ฮีโมโกลบินมีโครงสร้างควอเทอร์นารีที่เป็นลักษณะเฉพาะของโปรตีนทรงกลมหลายหน่วยย่อย[ 49 ]กรดอะมิโนส่วนใหญ่ในฮีโมโกลบินก่อตัวเป็นเกลียวอัลฟาและเกลียวเหล่านี้เชื่อมต่อกันด้วยส่วนที่ไม่เป็นเกลียวสั้นๆ พันธะไฮโดรเจนทำให้ส่วนที่เป็นเกลียวภายในโปรตีนนี้มีเสถียรภาพ ทำให้เกิดแรงดึงดูดภายในโมเลกุล ซึ่งจะทำให้สายโพลีเปปไทด์แต่ละสายพับตัวเป็นรูปร่างเฉพาะ[ 50 ]โครงสร้างควอเทอร์นารีของฮีโมโกลบินมาจากหน่วยย่อยทั้งสี่ของมันในลักษณะการจัดเรียงแบบทรงสี่หน้าโดยประมาณ[ 49 ]
ในสัตว์มีกระดูกสันหลังส่วนใหญ่ โมเลกุลฮีโมโกลบินประกอบด้วย หน่วยย่อย โปรตีนทรงกลมสี่หน่วยแต่ละหน่วยย่อยประกอบด้วยสายโปรตีนที่เชื่อมโยงอย่างแน่นหนากับกลุ่มฮีมที่ไม่ใช่โปรตีนสายโปรตีนแต่ละสายจะเรียงตัวเป็นชุดของ ส่วนโครงสร้าง อัลฟาเฮลิกซ์ที่เชื่อมต่อกันใน รูปแบบการพับ แบบโกลบินชื่อดังกล่าวตั้งขึ้นเนื่องจากการจัดเรียงนี้เป็นรูปแบบการพับแบบเดียวกันกับที่ใช้ในโปรตีนฮีม/โกลบินอื่นๆ เช่นไมโอโกลบิน [ 51 ] [ 52 ] รูปแบบการพับนี้มีช่องที่ยึดกลุ่มฮีมไว้อย่างแน่นหนา
กลุ่มฮีมประกอบด้วยไอออน เหล็ก (Fe) ที่ถูกยึดไว้ใน วงแหวน เฮเทอโรไซคลิกที่เรียกว่าพอร์ ฟิริน วงแหวนพอร์ฟิรินนี้ประกอบด้วยโมเลกุลไพร์โรลสี่โมเลกุลที่เชื่อมต่อกันเป็นวงจร (โดย สะพาน เมทิน ) โดยมีไอออนเหล็กยึดอยู่ตรงกลาง[ 53 ]ไอออนเหล็กซึ่งเป็นตำแหน่งของการจับออกซิเจน จะประสานงานกับ อะตอม ไนโตรเจน สี่ อะตอมที่อยู่ตรงกลางของวงแหวน ซึ่งทั้งหมดอยู่ในระนาบเดียวกัน ฮีมจะยึดติดอย่างแน่นหนา (แบบโคเวเลนต์) กับโปรตีนทรงกลมผ่านอะตอม N ของ วงแหวน อิมิดาโซล ของสารตกค้าง ฮิสติ ดีน F8 (หรือที่เรียกว่าฮิสติดีนใกล้เคียง) ใต้วงแหวนพอร์ฟิริน ตำแหน่งที่หกสามารถจับออกซิเจนได้แบบย้อนกลับได้ด้วยพันธะโคเวเลนต์แบบประสานงาน[ 54 ]ทำให้กลุ่มออกตาเฮดรัลของลิแกนด์หกตัวสมบูรณ์ การจับกับออกซิเจนแบบย้อนกลับได้นี้เป็นเหตุผลว่าทำไมฮีโมโกลบินจึงมีประโยชน์มากในการขนส่งออกซิเจนไปทั่วร่างกาย[ 55 ]ออกซิเจนจะยึดติดในรูปทรงเรขาคณิตแบบ "ปลายงอ" โดยที่อะตอมออกซิเจนหนึ่งตัวจะยึดติดอยู่กับ Fe และอีกตัวจะยื่นออกมาเป็นมุม เมื่อออกซิเจนไม่ยึดติด โมเลกุลน้ำที่ยึดติดอย่างอ่อนมากจะเติมเต็มตำแหน่งนั้น ทำให้เกิดรูปทรงแปด เหลี่ยมที่ บิดเบี้ยว
แม้ว่าคาร์บอนไดออกไซด์จะถูกลำเลียงโดยฮีโมโกลบิน แต่ก็ไม่ได้แย่งตำแหน่งการจับกับธาตุเหล็กกับออกซิเจน แต่จะจับกับหมู่เอมีนของสายโปรตีนที่เชื่อมต่อกับหมู่ฮีมแทน
ไอออนเหล็กอาจอยู่ใน สถานะ เฟอร์รัส Fe 2+หรือเฟอร์ริก Fe 3+ ก็ได้ แต่เฟอร์ริฮีโมโกลบิน (เมทฮีโมโกลบิน ) (Fe 3+ ) ไม่สามารถจับกับออกซิเจนได้[ 56 ]ในการจับกัน ออกซิเจนจะออกซิไดซ์ (Fe 2+ ) เป็น (Fe 3+ ) ชั่วคราวและย้อนกลับได้ ในขณะที่ออกซิเจนจะเปลี่ยนเป็น ไอออน ซูเปอร์ออกไซด์ ชั่วคราว ดังนั้นเหล็กต้องอยู่ในสถานะออกซิเดชัน +2 เพื่อจับกับออกซิเจน หากไอออนซูเปอร์ออกไซด์ที่เกี่ยวข้องกับ Fe 3+ถูกโปรตอน เหล็กในฮีโมโกลบินจะยังคงอยู่ในสถานะออกซิไดซ์และไม่สามารถจับกับออกซิเจนได้ ในกรณีเช่นนี้ เอนไซม์เมทฮีโมโกลบินรีดักเทสจะสามารถกระตุ้นเมทฮีโมโกลบินได้อีกครั้งโดยการลดศูนย์กลางเหล็ก
ในมนุษย์ผู้ใหญ่ ฮีโมโกลบินชนิดที่พบมากที่สุดคือเทตราเมอร์ (ซึ่งประกอบด้วยโปรตีนย่อยสี่หน่วย) เรียกว่าฮีโมโกลบิน Aซึ่งประกอบด้วยหน่วยย่อย α สองหน่วยและ β สองหน่วยที่ยึดติดกันโดยไม่ใช้พันธะโควาเลนต์ โดยแต่ละหน่วยประกอบด้วยกรดอะมิโน 141 และ 146 หน่วยตามลำดับ ซึ่งเขียนแทนด้วย α 2 β 2 หน่วยย่อยเหล่านี้มีโครงสร้างคล้ายกันและ มีขนาดใกล้เคียงกัน แต่ละหน่วยย่อยมีน้ำหนักโมเลกุลประมาณ 16,000 ดาลตัน [ 57 ] ดังนั้น น้ำหนักโมเลกุลรวมของเทตราเมอร์จึงอยู่ที่ประมาณ 64,000 ดาลตัน (64,458 กรัม/โมล) [ 58 ]ดังนั้น 1 กรัม/เดซิลิตร = 0.1551 มิลลิโมล/ลิตร ฮีโมโกลบิน A เป็นโมเลกุลฮีโมโกลบินที่ได้รับการศึกษาอย่างเข้มข้นที่สุด
ในทารกมนุษย์ โมเลกุล ฮีโมโกลบินของทารกในครรภ์ประกอบด้วยโซ่ α 2 โซ่และโซ่ γ 2 โซ่ โซ่ γ จะค่อยๆ ถูกแทนที่ด้วยโซ่ β เมื่อทารกเติบโตขึ้น[ 59 ]
สายโซ่โพลีเปปไทด์ทั้งสี่เชื่อมต่อกันด้วย พันธะ ไอออนิก พันธะไฮโดรเจนและผลกระทบจากแรงดึงดูดระหว่างโมเลกุลที่ไม่ชอบน้ำ (hydrophobic effect )
ความอิ่มตัวของออกซิเจน
โดยทั่วไป ฮีโมโกลบินสามารถอิ่มตัวด้วยโมเลกุลออกซิเจน (ออกซีฮีโมโกลบิน) หรือไม่อิ่มตัวด้วยโมเลกุลออกซิเจน (ดีออกซีฮีโมโกลบิน) [ 60 ]
ออกซีฮีโมโกลบิน
ออก ซีฮีโมโกลบินเกิดขึ้นระหว่างการหายใจทางสรีรวิทยาเมื่อออกซิเจนจับกับส่วนประกอบฮีมของโปรตีนฮีโมโกลบินในเม็ดเลือดแดง กระบวนการนี้เกิดขึ้นในเส้นเลือดฝอยปอดที่อยู่ติดกับถุงลมของปอด จากนั้นออกซิเจนจะเดินทางผ่านกระแสเลือดไปตกที่เซลล์ซึ่งจะถูกนำไปใช้เป็นตัวรับอิเล็กตรอนตัวสุดท้ายในการผลิตATPโดยกระบวนการฟอสโฟรีเลชันแบบออกซิเดชันอย่างไรก็ตาม มันไม่ได้ช่วยต่อต้านการลดลงของค่า pH ในเลือดการระบายอากาศหรือการหายใจอาจย้อนกลับสภาวะนี้ได้โดยการกำจัดคาร์บอนไดออกไซด์ทำให้ค่า pH สูงขึ้น[ 61 ]
ฮีโมโกลบินมีอยู่สองรูปแบบ คือรูปแบบตึง (T) และรูปแบบผ่อนคลาย (R) ปัจจัยต่างๆ เช่น pH ต่ำ CO2 สูงและ 2,3 BPG สูง ในระดับเนื้อเยื่อ จะส่งเสริมรูปแบบตึง ซึ่งมีความสัมพันธ์กับออกซิเจนต่ำและปล่อยออกซิเจนในเนื้อเยื่อ ในทางกลับกัน pH สูง CO2 ต่ำหรือ 2,3 BPG ต่ำ จะส่งเสริมรูปแบบผ่อนคลาย ซึ่งสามารถจับออกซิเจนได้ดีกว่า[ 62 ]ความดันย่อยของระบบยังส่งผลต่อความสัมพันธ์กับ O2 ด้วยโดยที่ความดันย่อยของออกซิเจนสูง (เช่น ที่มีอยู่ในถุงลม) สถานะผ่อนคลาย (ความสัมพันธ์สูง, R) จะเป็นที่ต้องการ ในทางกลับกัน ที่ความดันย่อยต่ำ (เช่น ที่มีอยู่ในเนื้อเยื่อที่หายใจ) สถานะตึง (ความสัมพันธ์ต่ำ, T) จะเป็นที่ต้องการ[ 63 ]นอกจากนี้ การจับตัวของออกซิเจนกับฮีมเหล็ก(II) จะดึงเหล็กเข้าไปในระนาบของวงแหวนพอร์ไฟริน ทำให้เกิดการเปลี่ยนแปลงโครงสร้างเล็กน้อย การเปลี่ยนแปลงนี้กระตุ้นให้ออกซิเจนจับกับหน่วยฮีมที่เหลืออีก 3 หน่วยภายในฮีโมโกลบิน (ดังนั้น การจับตัวของออกซิเจนจึงเป็นแบบร่วมมือกัน)
ตามหลักการแล้ว เหล็กในออกซีฮีโมโกลบินจะถูกมองว่าอยู่ในสถานะออกซิเดชันของเหล็ก(II) อย่างไรก็ตาม สารประกอบของออกซิเจนกับเหล็กฮีมเป็นไดอะ แมกเนติก ในขณะที่ทั้งออกซิเจนและเหล็ก(II) ที่มีสปินสูงเป็นพาราแมกเนติกหลักฐานเชิงทดลองชี้ให้เห็นอย่างชัดเจนว่าเหล็กฮีมอยู่ในสถานะออกซิเดชันของเหล็ก(III) ในออกซีฮีโมโกลบิน โดยออกซิเจนมีอยู่ในรูปของซูเปอร์ออกไซด์แอนไอออน (O 2 •− ) หรือในสารประกอบเชิงซ้อนการถ่ายโอนประจุแบบโควาเลนต์[ 64 ]
ฮีโมโกลบินที่ขาดออกซิเจน
ฮีโมโกลบินที่ไม่มีออกซิเจน (ดีออกซีฮีโมโกลบิน) คือรูปแบบของฮีโมโกลบินที่ไม่มีออกซิเจนจับ อยู่ สเปกตรัมการดูดกลืนแสงของออกซีฮีโมโกลบินและดีออกซีฮีโมโกลบินแตกต่างกัน ออกซีฮีโมโกลบินมีการดูดกลืนแสงที่ความยาวคลื่น 660 นาโนเมตรต่ำกว่า ดีออกซีฮีโมโกลบินอย่างมีนัยสำคัญ ในขณะที่ที่ความยาวคลื่น 940 นาโนเมตร การดูดกลืนแสงจะสูงกว่าเล็กน้อย ความแตกต่างนี้ถูกนำมาใช้ในการวัดปริมาณออกซิเจนในเลือดของผู้ป่วยด้วยเครื่องมือที่เรียกว่า เครื่องวัดออกซิเจนใน เลือดแบบพัลส์ความแตกต่างนี้ยังอธิบายถึงอาการตัวเขียวซึ่งเป็นสีน้ำเงินถึงม่วงที่เนื้อเยื่อพัฒนาขึ้นระหว่างภาวะขาดออกซิเจน[ 65 ]
ฮีโมโกลบินที่ปราศจากออกซิเจนมีคุณสมบัติเป็นพาราแมกเนติกกล่าวคือ มันถูกดึงดูดเข้าหาสนามแม่เหล็ก อย่างอ่อน ๆ[ 66 ] [ 67 ]ในทางตรงกันข้าม ฮีโมโกลบินที่มีออกซิเจนแสดงคุณสมบัติเป็นไดอะแมกเนติกคือมีการผลักออกจากสนามแม่เหล็กอย่างอ่อนๆ[ 67 ]
ความร่วมมือ

เมื่อออกซิเจนจับกับสารประกอบเหล็ก จะทำให้อะตอมเหล็กเคลื่อนที่กลับเข้าหาจุดศูนย์กลางของระนาบวงแหวนพอร์ฟิริน (ดูแผนภาพการเคลื่อนที่) ในขณะเดียวกัน โซ่ ข้าง อิมิดาโซลของหมู่ฮิสติดีนที่ทำปฏิกิริยากับขั้วอีกด้านหนึ่งของเหล็กจะถูกดึงเข้าหาวงแหวนพอร์ฟิริน ปฏิกิริยานี้บังคับให้ระนาบของวงแหวนเคลื่อนไปด้านข้างออกไปด้านนอกของเตตระเมอร์ และยังเหนี่ยวนำให้เกิดความเครียดในเกลียวโปรตีนที่มีฮิสติดีนอยู่ขณะที่มันเคลื่อนเข้าใกล้อะตอมเหล็ก ความเครียดนี้จะถูกส่งต่อไปยังโมโนเมอร์อีกสามตัวในเตตระเมอร์ ซึ่งจะเหนี่ยวนำให้เกิดการเปลี่ยนแปลงโครงสร้างที่คล้ายกันในตำแหน่งฮีมอื่นๆ ทำให้การจับของออกซิเจนกับตำแหน่งเหล่านี้ง่ายขึ้น
เมื่อออกซิเจนจับกับโมโนเมอร์ของฮีโมโกลบินหนึ่งตัว โครงสร้างของเตตระเมอร์จะเปลี่ยนจากสถานะ T (ตึง) ไปเป็นสถานะ R (ผ่อนคลาย) การเปลี่ยนแปลงนี้ส่งเสริมการจับของออกซิเจนกับกลุ่มฮีมของโมโนเมอร์ที่เหลืออีกสามตัว ทำให้โมเลกุลของฮีโมโกลบินอิ่มตัวด้วยออกซิเจน[ 68 ]
ในฮีโมโกลบินแบบเตตระเมอร์ของผู้ใหญ่ปกติ การจับกับออกซิเจนจึงเป็นกระบวนการแบบร่วมมือกันความสามารถในการจับกับออกซิเจนของฮีโมโกลบินจะเพิ่มขึ้นตามความอิ่มตัวของออกซิเจนในโมเลกุล โดยโมเลกุลออกซิเจนกลุ่มแรกที่จับจะส่งผลต่อรูปร่างของตำแหน่งการจับสำหรับโมเลกุลถัดไปในลักษณะที่เอื้อต่อการจับ การจับแบบร่วมมือกันในเชิงบวกนี้เกิดขึ้นได้จาก การเปลี่ยนแปลงโครงสร้าง เชิงพื้นที่ของโปรตีนเชิงซ้อนฮีโมโกลบินดังที่ได้กล่าวไว้ข้างต้น กล่าวคือ เมื่อโปรตีนย่อยหนึ่งในฮีโมโกลบินจับกับออกซิเจน การเปลี่ยนแปลงโครงสร้างหรือรูปร่างในเชิงซ้อนทั้งหมดจะเริ่มต้นขึ้น ทำให้โปรตีนย่อยอื่นๆ มีความสามารถในการจับกับออกซิเจนเพิ่มขึ้น ผลที่ตามมาคือ เส้นโค้งการจับออกซิเจนของฮีโมโกลบินมีลักษณะเป็นรูปตัวSหรือรูปซิกมอยด์ ตรงข้ามกับ เส้นโค้ง ไฮเปอร์โบลา ปกติ ที่เกี่ยวข้องกับการจับแบบไม่ร่วมมือกัน
กลไกไดนามิกของความร่วมมือในฮีโมโกลบินและความสัมพันธ์กับการสั่นพ้อง ความถี่ต่ำ ได้รับการกล่าวถึงแล้ว[ 69 ]
การจับตัวของลิแกนด์อื่นที่ไม่ใช่ออกซิเจน
นอกจากลิแกนด์ ออกซิเจน ซึ่งจับกับฮีโมโกลบินในลักษณะร่วมมือกันแล้ว ลิแกนด์ของฮีโมโกลบินยังรวมถึงสารยับยั้งการแข่งขันเช่นคาร์บอนมอนอกไซด์ (CO) และลิแกนด์อัลโลสเตอริกเช่น คาร์บอนไดออกไซด์ (CO2 )และไนตริกออกไซด์ (NO) คาร์บอนไดออกไซด์จะจับกับหมู่เอมีนของโปรตีนโกลบินเพื่อสร้างคาร์บามิโนฮีโมโกลบิน กลไกนี้เชื่อว่ามีส่วนรับผิดชอบประมาณ 10% ของการขนส่งคาร์บอนไดออกไซด์ในสัตว์เลี้ยงลูกด้วยนม ไนตริกออกไซด์ยังสามารถถูกขนส่งโดยฮีโมโกลบินได้เช่นกัน โดยจะจับกับ หมู่ ไทออล เฉพาะ ในโปรตีนโกลบินเพื่อสร้าง S-ไนโตรโซไทออล ซึ่งจะแตกตัวเป็นไนตริกออกไซด์อิสระและไทออลอีกครั้ง เมื่อฮีโมโกลบินปล่อยออกซิเจนจากตำแหน่งฮีม การขนส่งไนตริกออกไซด์ไปยังเนื้อเยื่อส่วนปลายนี้คาดว่าจะช่วยในการขนส่งออกซิเจนในเนื้อเยื่อ โดยการปล่อย ไนตริกออกไซด์ ที่ทำให้หลอดเลือดขยายตัวไปยังเนื้อเยื่อที่มีระดับออกซิเจนต่ำ[ 70 ]
การแข่งขัน
การจับตัวของออกซิเจนได้รับผลกระทบจากโมเลกุลต่างๆ เช่น คาร์บอนมอนอกไซด์ (ตัวอย่างเช่น จากการสูบบุหรี่ก๊าซไอเสียและการเผาไหม้ที่ไม่สมบูรณ์ในเตาเผา) CO จะแข่งขันกับออกซิเจนที่ตำแหน่งการจับฮีม ความสามารถในการจับของฮีโมโกลบินกับ CO นั้นมากกว่าความสามารถในการจับกับออกซิเจนถึง 250 เท่า[ 71 ] [ 72 ]เนื่องจากคาร์บอนมอนอกไซด์เป็นก๊าซที่ไม่มีสี ไม่มีกลิ่น และไม่มีรส และเป็นอันตรายถึงชีวิตได้ จึง มีการผลิต เครื่องตรวจจับคาร์บอนมอนอกไซด์ออกมาจำหน่ายเพื่อเตือนถึงระดับที่เป็นอันตรายในที่อยู่อาศัย เมื่อฮีโมโกลบินรวมตัวกับ CO จะเกิดเป็นสารประกอบสีแดงสดที่เรียกว่าคาร์บอกซีฮีโมโกลบินซึ่งอาจทำให้ผิวหนังของ ผู้ที่ได้รับ พิษจาก COมีสีชมพูเมื่อเสียชีวิต แทนที่จะเป็นสีขาวหรือสีน้ำเงิน เมื่ออากาศที่สูดดมเข้าไปมีระดับ CO ต่ำถึง 0.02% จะทำให้เกิด อาการปวดหัวและคลื่นไส้หากความเข้มข้นของ CO เพิ่มขึ้นเป็น 0.1% จะทำให้หมดสติ ในผู้ที่สูบบุหรี่จัด บริเวณที่ออกซิเจนทำงานอาจถูกปิดกั้นโดยคาร์บอนมอนอกไซด์ได้มากถึง 20%
ในทำนองเดียวกัน ฮีโมโกลบินยังมีแรงยึดเกาะแบบแข่งขันกับไซยาไนด์ (CN⁻ )ซัลเฟอร์โมโนออกไซด์ (SO) และซัลไฟด์ (S²⁻ )รวมถึงไฮโดรเจนซัลไฟด์ (H₂S )สารเหล่านี้ทั้งหมดจับกับเหล็กในฮีมโดยไม่เปลี่ยนแปลงสถานะออกซิเดชัน แต่ถึงกระนั้นก็ยับยั้งการจับกับออกซิเจน ทำให้เกิดความเป็นพิษร้ายแรง
อะตอมเหล็กในกลุ่มฮีมจะต้องอยู่ใน สถานะออกซิเดชัน เฟอร์รัส (Fe 2+ ) ในตอนเริ่มต้นเพื่อรองรับการจับและการขนส่งออกซิเจนและก๊าซอื่นๆ (มันจะเปลี่ยนเป็นเฟอร์ริกชั่วคราวในระหว่างที่ออกซิเจนจับอยู่ ดังที่ได้อธิบายไว้ข้างต้น) การออกซิเดชันเริ่มต้นไปเป็น สถานะ เฟอร์ริก (Fe 3+ ) โดยปราศจากออกซิเจนจะเปลี่ยนฮีโมโกลบินเป็น "ฮีมไอโกลบิน" หรือเมทฮีโมโกลบินซึ่งไม่สามารถจับออกซิเจนได้ ฮีโมโกลบินในเม็ดเลือดแดงปกติได้รับการปกป้องโดยระบบรีดักชันเพื่อป้องกันไม่ให้เกิดเหตุการณ์นี้ขึ้น ไนตริกออกไซด์สามารถเปลี่ยนฮีโมโกลบินเพียงส่วนน้อยให้เป็นเมทฮีโมโกลบินในเม็ดเลือดแดงได้ ปฏิกิริยาหลังนี้เป็นกิจกรรมที่หลงเหลืออยู่จากหน้าที่ ของ ไนตริกออกไซด์ไดออกซิเจ เนสในโกลบินที่มีมาแต่โบราณ
อัลโลสเตอริก
คาร์บอนไดออกไซด์จะเข้าจับกับตำแหน่งที่แตกต่างกันบนฮีโมโกลบิน ในเนื้อเยื่อที่มีความเข้มข้นของคาร์บอนไดออกไซด์สูงกว่า คาร์บอนไดออกไซด์จะจับกับตำแหน่งอัลโลสเตอริกของฮีโมโกลบิน ทำให้การปลดปล่อยออกซิเจนจากฮีโมโกลบินง่ายขึ้น และในที่สุดก็จะถูกกำจัดออกจากร่างกายหลังจากที่ออกซิเจนถูกปล่อยไปยังเนื้อเยื่อที่กำลังเผาผลาญ ความสามารถในการจับกับคาร์บอนไดออกไซด์ที่เพิ่มขึ้นของเลือดดำนี้เรียกว่าปรากฏการณ์โบห์ร (Bohr effect ) โดยผ่านเอนไซม์คาร์บอนิกแอนไฮดราส คาร์บอนไดออกไซด์จะทำปฏิกิริยากับน้ำเพื่อให้ได้กรดคาร์บอนิกซึ่งจะสลายตัวเป็นไบคาร์บอเนตและโปรตอน
- CO 2 + H 2 O → H 2 CO 3 → HCO 3 − + H +

ดังนั้น เลือดที่มีระดับคาร์บอนไดออกไซด์สูงจึงมีค่า pH ต่ำลง ( เป็นกรด มากขึ้น ) ฮีโมโกลบินสามารถจับกับโปรตอนและคาร์บอนไดออกไซด์ ซึ่งทำให้เกิดการเปลี่ยนแปลงโครงสร้างของโปรตีนและอำนวยความสะดวกในการปล่อยออกซิเจน โปรตอนจะจับกับตำแหน่งต่างๆ บนโปรตีน ในขณะที่คาร์บอนไดออกไซด์จะจับกับกลุ่ม α-อะมิโน[ 73 ]คาร์บอนไดออกไซด์จับกับฮีโมโกลบินและก่อตัวเป็นคาร์บามิโน ฮีโม โกลบิน[ 74 ]การลดลงของความสัมพันธ์ของฮีโมโกลบินกับออกซิเจนเนื่องจากการจับของคาร์บอนไดออกไซด์และกรดนี้เรียกว่าปรากฏการณ์โบร์ปรากฏการณ์โบร์สนับสนุนสถานะ T มากกว่าสถานะ R (เลื่อนเส้นโค้งความอิ่มตัวของ O 2 ไปทาง ขวา ) ในทางกลับกัน เมื่อระดับคาร์บอนไดออกไซด์ในเลือดลดลง (เช่น ในเส้นเลือดฝอยของปอด) คาร์บอนไดออกไซด์และโปรตอนจะถูกปล่อยออกจากฮีโมโกลบิน ทำให้ความสัมพันธ์ของโปรตีนกับออกซิเจนเพิ่มขึ้น การลดลงของความสามารถในการจับออกซิเจนโดยรวมของฮีโมโกลบิน (กล่าวคือ กราฟเลื่อนลง ไม่ใช่แค่เลื่อนไปทางขวา) เนื่องจากการลดลงของค่า pH เรียกว่า ปรากฏการณ์ราก (root effect ) ซึ่งพบได้ในปลาที่มีกระดูก
ฮีโมโกลบินจำเป็นต้องปล่อยออกซิเจนที่มันจับไว้ หากไม่เช่นนั้น การจับไว้ก็ไม่มีประโยชน์ เส้นโค้งรูปตัว S ของฮีโมโกลบินทำให้มีประสิทธิภาพในการจับ (รับ O2 ในปอด) และมีประสิทธิภาพในการปล่อย (ปล่อย O2 ในเนื้อเยื่อ) [ 75 ]
ในคนที่ปรับตัวเข้ากับระดับความสูงแล้ว ความเข้มข้นของ2,3-บิสฟอสโฟกลีเซอเรต (2,3-BPG) ในเลือดจะเพิ่มขึ้น ซึ่งช่วยให้บุคคลเหล่านี้สามารถส่งออกซิเจนไปยังเนื้อเยื่อได้มากขึ้นภายใต้สภาวะที่มีความดันออกซิเจน ต่ำ ปรากฏการณ์นี้ ซึ่งโมเลกุล Y ส่งผลต่อการจับของโมเลกุล X กับโมเลกุลขนส่ง Z เรียกว่า ผลกระทบอัลโลสเตอ ริกแบบเฮเทอโรโทรปิกฮีโมโกลบินในสิ่งมีชีวิตที่ระดับความสูงยังปรับตัวให้มีความสัมพันธ์กับ 2,3-BPG น้อยลง ดังนั้นโปรตีนจะเปลี่ยนไปสู่สถานะ R มากขึ้น ในสถานะ R ฮีโมโกลบินจะจับกับออกซิเจนได้ง่ายขึ้น จึงช่วยให้สิ่งมีชีวิตสามารถดำเนินกระบวนการเผาผลาญที่จำเป็นได้เมื่อมีออกซิเจนอยู่ที่ความดันย่อยต่ำ[ 76 ]
สัตว์อื่นๆ นอกเหนือจากมนุษย์ใช้โมเลกุลที่แตกต่างกันในการจับกับฮีโมโกลบินและเปลี่ยนแปลงความสัมพันธ์กับออกซิเจนภายใต้สภาวะที่ไม่เอื้ออำนวย ปลาใช้ทั้งATPและGTPโมเลกุลเหล่านี้จะจับกับ "ช่อง" ฟอสเฟตบนโมเลกุลฮีโมโกลบินของปลา ซึ่งทำให้สถานะตึงตัวมีเสถียรภาพมากขึ้น และด้วยเหตุนี้จึงลดความสัมพันธ์กับออกซิเจน[ 77 ] GTP ลดความสัมพันธ์กับออกซิเจนของฮีโมโกลบินได้มากกว่า ATP มาก ซึ่งเชื่อว่าเป็นผลมาจากพันธะไฮโดรเจน พิเศษ ที่เกิดขึ้นซึ่งทำให้สถานะตึงตัวมีเสถียรภาพมากขึ้น[ 78 ]ภายใต้สภาวะที่มีออกซิเจนต่ำ ความเข้มข้นของทั้ง ATP และ GTP จะลดลงในเซลล์เม็ดเลือดแดงของปลาเพื่อเพิ่มความสัมพันธ์กับออกซิเจน[ 79 ]
ฮีโมโกลบินชนิดหนึ่งที่เรียกว่า ฮี โมโกลบินของทารกในครรภ์ (HbF, α₂γ₂ )พบในทารก ในครรภ์ที่กำลังพัฒนา และจับกับออกซิเจนได้ดีกว่าฮีโมโกลบินของผู้ใหญ่ หมายความว่า กราฟ การจับออกซิเจนของฮีโมโกลบินของทารกในครรภ์จะเลื่อนไปทางซ้าย (กล่าวคือ ฮีโมโกลบินส่วนใหญ่จะจับกับออกซิเจนได้ในสภาวะความดันออกซิเจนต่ำกว่า) เมื่อเทียบกับฮีโมโกลบินของผู้ใหญ่ ด้วยเหตุนี้ เลือดของทารกในครรภ์ ในรก จึงสามารถรับออกซิเจนจากเลือดของมารดาได้
ฮีโมโกลบินยังขนส่งไนตริกออกไซด์ (NO) ในส่วนของโกลบินของโมเลกุล ซึ่งช่วยปรับปรุงการส่งออกซิเจนในส่วนปลายและมีส่วนช่วยในการควบคุมการหายใจ NO จับกับสารตกค้างซิสเทอีนเฉพาะในโกลบินแบบย้อนกลับได้ การจับขึ้นอยู่กับสถานะ (R หรือ T) ของฮีโมโกลบิน ฮีโมโกลบิน S-ไนโตรซิเลตที่เกิดขึ้นจะมีอิทธิพลต่อกิจกรรมต่างๆ ที่เกี่ยวข้องกับ NO เช่น การควบคุมความต้านทานของหลอดเลือด ความดันโลหิต และการหายใจ NO ไม่ได้ถูกปล่อยออกมาในไซโตพลาสซึมของเซลล์เม็ดเลือดแดง แต่ถูกขนส่งออกจากเซลล์โดยตัวแลกเปลี่ยนแอนไอออนที่เรียกว่า AE1 [ 80 ]
ชนิดของฮีโมโกลบินในมนุษย์
ฮีโมโกลบินชนิดต่างๆเป็นส่วนหนึ่งของ การพัฒนา ตัวอ่อนและทารก ในครรภ์ตามปกติ นอกจากนี้ยังอาจเป็นรูปแบบกลายพันธุ์ของฮีโมโกลบินที่ก่อให้เกิดพยาธิสภาพในประชากรซึ่งเกิดจากการเปลี่ยนแปลงทางพันธุกรรม ฮีโมโกลบินชนิดต่างๆ ที่รู้จักกันดีบางชนิด เช่นโรคโลหิตจางชนิดเคียว เป็นสาเหตุของโรคต่างๆ และถือเป็น โรค ฮีโมโกลบิน ผิดปกติ ฮีโมโกลบินชนิดอื่นๆ ไม่ก่อให้ เกิดพยาธิสภาพที่ตรวจพบได้จึงถือเป็นฮีโมโกลบินชนิดที่ไม่ก่อให้เกิดพยาธิสภาพ[ 34 ] [ 81 ]
ในตัวอ่อน :
- Gower 1 (ζ 2 ε 2 ).
- โกเวอร์ 2 (α 2 ε 2 ) ( PDB : 1A9W ).
- เฮโมโกลบินพอร์ตแลนด์ 1 (ζ 2 γ 2 )
- เฮโมโกลบินพอร์ตแลนด์ II (ζ 2 β 2 )
ในทารกในครรภ์:
- เฮโมโกลบิน เอฟ (α 2 γ 2 ) ( PDB : 1FDH ).
ในทารกแรกเกิด (เด็กแรกเกิดทันทีหลังคลอด):
- ฮีโมโกลบินเอ (ฮีโมโกลบินในผู้ใหญ่) (α 2 β 2 ) ( PDB : 1BZ0 ) – เป็นชนิดที่พบได้บ่อยที่สุด โดยมีปริมาณปกติมากกว่า 95%
- การสังเคราะห์สายโซ่ δ ของฮีโมโกลบิน A2 ( α2δ2 ) เริ่มขึ้นในช่วงปลายไตรมาสที่สาม และในผู้ใหญ่จะมีค่าปกติอยู่ที่ 1.5–3.5%
- ฮีโมโกลบินเอฟ (ฮีโมโกลบินของทารกในครรภ์) (α 2 γ 2 ) – ในผู้ใหญ่ ฮีโมโกลบินเอฟจะจำกัดอยู่เฉพาะในกลุ่มเซลล์เม็ดเลือดแดงกลุ่มเล็กๆ ที่เรียกว่าเซลล์เอฟ อย่างไรก็ตาม ระดับของฮีโมโกลบินเอฟอาจสูงขึ้นได้ในผู้ที่เป็นโรคโลหิตจางชนิดเคียวและโรคธาลัสซีเมียเบต้า

รูปแบบที่ผิดปกติที่พบในโรคต่างๆ:
- เฮโมโกลบิน D – (α 2 β D 2 ) – รูปแบบหนึ่งของฮีโมโกลบิน
- ฮีโมโกลบิน H (β 4 ) – ฮีโมโกลบินรูปแบบหนึ่งที่เกิดจากการรวมตัวของสายโซ่ β สี่สาย ซึ่งอาจพบได้ในโรคธาลัสซีเมียอัลฟาชนิด ต่างๆ
- ฮีโมโกลบินบาร์ตส์ (γ 4 ) – ฮีโมโกลบินรูปแบบหนึ่งที่เกิดจากการรวมตัวเป็นเตตระเมอร์ของโซ่ γ ซึ่งอาจพบได้ในโรคธาลัสซีเมียอัลฟาชนิดต่างๆ
- ฮีโมโกล บินเอส (α₂βS₂ ) –ฮีโมโกลบินรูปแบบหนึ่งที่พบในผู้ป่วยโรคโลหิตจางชนิดเคียว มีความแปรผันในยีนสายเบต้า ทำให้คุณสมบัติของฮีโมโกลบินเปลี่ยนแปลงไป ส่งผลให้เม็ดเลือดแดงมีรูปร่างคล้ายเคียว
- ฮีโมโกลบินซี (α 2 β C 2 ) – อีกหนึ่งรูปแบบที่เกิดจากการเปลี่ยนแปลงในยีนสายเบต้า รูปแบบนี้ทำให้เกิดภาวะโลหิตจาง เรื้อรังชนิดเม็ดเลือดแดงแตก ที่ ไม่รุนแรง
- ฮีโมโกลบินอี (α 2 β E 2 ) – อีกหนึ่งรูปแบบที่เกิดจากการเปลี่ยนแปลงในยีนสายเบต้า รูปแบบนี้ทำให้เกิดภาวะโลหิตจางเรื้อรังชนิดเม็ดเลือดแดงแตกที่ไม่รุนแรง
- ฮีโมโกลบิน AS – รูปแบบเฮเทอโรไซกัสที่ทำให้เกิดลักษณะเซลล์เคียวโดยมีหนึ่งยีนสำหรับผู้ใหญ่และหนึ่งยีนของโรคเซลล์เคียว
- โรคฮีโมโกลบิน SC – เป็นรูปแบบเฮเทโรไซกัสแบบผสมที่มีหนึ่งยีนที่เข้ารหัสโปรตีนเคียว และอีกหนึ่งยีนที่เข้ารหัสฮีโมโกลบิน C
- ฮีโมโกลบินฮอปกินส์-2 – ฮีโมโกลบินรูปแบบหนึ่งที่บางครั้งพบร่วมกับฮีโมโกลบินเอสทำให้เกิดโรคโลหิตจางชนิดเคียว (sickle cell disease)
การเสื่อมสภาพในสัตว์มีกระดูกสันหลัง
เมื่อเซลล์เม็ดเลือดแดงมีอายุขัยสิ้นสุดลงเนื่องจากความชราหรือความบกพร่อง เซลล์เหล่านั้นจะถูกกำจัดออกจากระบบไหลเวียนโลหิตโดยกิจกรรมฟาโกไซติกของแมโครฟาจในม้ามหรือตับ หรือเกิดการแตกตัวเป็นเม็ดเลือดภายในระบบไหลเวียนโลหิต จากนั้น ฮีโมโกลบินอิสระจะถูกกำจัดออกจากระบบไหลเวียนโลหิตโดยตัวขนส่งฮีโมโกลบินCD163ซึ่งแสดงออกเฉพาะในโมโนไซต์หรือแมโครฟาจเท่านั้น ภายในเซลล์เหล่านี้ โมเลกุลของฮีโมโกลบินจะถูกสลาย และธาตุเหล็กจะถูกนำกลับมาใช้ใหม่ กระบวนการนี้ยังผลิตคาร์บอนมอนอกไซด์หนึ่งโมเลกุลต่อฮีมหนึ่งโมเลกุลที่ถูกย่อยสลาย[ 82 ]การย่อยสลายฮีมเป็นแหล่งกำเนิดคาร์บอนมอนอกไซด์ตามธรรมชาติเพียงแหล่งเดียวในร่างกายมนุษย์ และเป็นสาเหตุของระดับคาร์บอนมอนอกไซด์ในเลือดปกติในคนที่หายใจเอาอากาศปกติ[ 83 ]
ผลิตภัณฑ์สุดท้ายที่สำคัญอีกอย่างหนึ่งของการสลายตัวของฮีมคือบิลิรูบินระดับสารเคมีนี้ที่เพิ่มขึ้นจะถูกตรวจพบในเลือดหากเซลล์เม็ดเลือดแดงถูกทำลายเร็วกว่าปกติ โปรตีนฮีโมโกลบินที่สลายตัวไม่เหมาะสมหรือฮีโมโกลบินที่ถูกปล่อยออกจากเซลล์เม็ดเลือดเร็วเกินไปอาจอุดตันหลอดเลือดขนาดเล็ก โดยเฉพาะอย่างยิ่งหลอดเลือดกรองเลือดที่บอบบางของไตทำให้ไตเสียหาย ธาตุเหล็กจะถูกแยกออกจากฮีมและนำกลับมาใช้ใหม่ในภายหลัง โดยจะถูกเก็บไว้ในรูปของเฮโมซิเดอรินหรือเฟอร์ริตินในเนื้อเยื่อและถูกขนส่งในพลาสมาโดยเบต้าโกลบูลินในรูปของทรานสเฟอร์รินเมื่อวงแหวนพอร์ฟิรินแตกออก ชิ้นส่วนเหล่านั้นมักจะถูกขับออกมาเป็นเม็ดสีเหลืองที่เรียกว่าบิลิรูบิน ซึ่งถูกขับออกมาในลำไส้ในรูปของน้ำดี ลำไส้จะเผาผลาญบิลิรูบินเป็นยูโรบิลิโนเจน ยูโรบิลิโนเจนจะถูกขับออกจากร่างกายทางอุจจาระในรูปของเม็ดสีที่เรียกว่าสเตอโคบิลิน โกลบูลินจะถูกเผาผลาญเป็นกรดอะมิโนแล้วปล่อยเข้าสู่ระบบไหลเวียนโลหิต
โรคที่เกี่ยวข้องกับฮีโมโกลบิน
ภาวะขาดฮีโมโกลบินอาจเกิดจากปริมาณโมเลกุลฮีโมโกลบินลดลง เช่นในภาวะโลหิตจางหรือเกิดจากความสามารถของแต่ละโมเลกุลในการจับออกซิเจนลดลงที่ความดันย่อยของออกซิเจนเท่าเดิมโรคฮีโมโกลบินผิดปกติ (ความบกพร่องทางพันธุกรรมที่ส่งผลให้โครงสร้างของโมเลกุลฮีโมโกลบินผิดปกติ) [ 84 ]อาจทำให้เกิดทั้งสองอย่าง ในทุกกรณี ภาวะขาดฮีโมโกลบินจะลดความสามารถในการขนส่งออกซิเจนของเลือดโดยทั่วไป ภาวะขาดฮีโมโกลบินจะแตกต่างอย่างชัดเจนจากภาวะไฮโปซีเมียซึ่งหมายถึงความดันย่อยของออกซิเจนในเลือด ลดลง [ 85 ] [ 86 ] [ 87 ] [ 88 ]แม้ว่าทั้งสองจะเป็นสาเหตุของภาวะไฮโปเซีย (การได้รับออกซิเจนไม่เพียงพอของเนื้อเยื่อ) ก็ตาม
สาเหตุอื่นๆ ที่พบบ่อยของภาวะฮีโมโกลบินต่ำ ได้แก่ การเสียเลือด ภาวะขาดสารอาหาร ปัญหาไขกระดูก การทำเคมีบำบัด ภาวะไตวาย หรือฮีโมโกลบินผิดปกติ (เช่น ฮีโมโกลบินในโรคโลหิตจางชนิดเคียว)
โดยปกติแล้ว ความสามารถของโมเลกุลฮีโมโกลบินแต่ละโมเลกุลในการขนส่งออกซิเจนจะถูกปรับเปลี่ยนโดยค่า pH ของเลือดหรือคาร์บอนไดออกไซด์ (CO2 ) ที่เปลี่ยนแปลง ไป ทำให้เส้นโค้งการแยกตัวของออกซิเจน-ฮีโมโกลบิน เปลี่ยนแปลงไป อย่างไรก็ตาม ความสามารถนี้ก็อาจเปลี่ยนแปลงไปในทางพยาธิสภาพได้เช่นกัน เช่น ในกรณี พิษจากคาร์บอนมอนอกไซด์
การลดลงของฮีโมโกลบิน ไม่ว่าจะมีการลดลงของเม็ดเลือดแดงหรือไม่ก็ตาม จะนำไปสู่อาการของภาวะโลหิตจาง ภาวะโลหิตจางมีสาเหตุหลายประการ แม้ว่าการขาดธาตุเหล็กและภาวะโลหิตจางจากการขาดธาตุเหล็กจะเป็นสาเหตุที่พบได้บ่อยที่สุดในโลกตะวันตก เนื่องจากการขาดธาตุเหล็กทำให้การสังเคราะห์ฮีมลดลง เม็ดเลือดแดงในภาวะโลหิตจางจากการขาดธาตุเหล็กจึงมีสีซีด (ขาดเม็ดสีแดงของฮีโมโกลบิน) และมีขนาดเล็กกว่าปกติ ภาวะโลหิตจางชนิดอื่นพบได้น้อยกว่า ในภาวะ เม็ดเลือดแดงแตก (การแตกตัวของเม็ดเลือดแดงอย่างรวดเร็ว) อาการดีซ่าน ที่เกิดขึ้นร่วมด้วยนั้น เกิดจากสารเมตาโบไลต์ของฮีโมโกลบินคือบิลิรูบิน และฮีโมโกลบินที่ไหลเวียนอยู่ในกระแสเลือดอาจทำให้ไตวายได้
การกลายพันธุ์บางอย่างในสายโกลบินมีความเกี่ยวข้องกับโรคฮีโมโกลบิน ผิดปกติ เช่น โรคโลหิตจางชนิดเคียวและธาลัสซีเมีย ส่วนการกลายพันธุ์อื่นๆ ดังที่ได้กล่าวไว้ในตอนต้นของบทความ นั้น ไม่เป็นอันตรายและเรียกง่ายๆ ว่าเป็นเพียงฮีโมโกลบินชนิดแปรผัน
มีกลุ่มโรคทางพันธุกรรมกลุ่มหนึ่งที่เรียกว่า โรคพอร์ฟิเรียซึ่งมีลักษณะเฉพาะคือความผิดปกติในกระบวนการเมตาบอลิซึมของการสังเคราะห์ฮีม พระเจ้าจอร์จที่ 3 แห่งสหราชอาณาจักรน่าจะเป็นผู้ป่วยโรคพอร์ฟิเรียที่มีชื่อเสียงที่สุด
ฮีโมโกลบินเอจะค่อยๆ รวมตัวกับกลูโคสที่ปลายสุดของกรดอะมิโนวาลีน (อัลฟาอะมิโนแอซิด) ในแต่ละสายเบต้า โมเลกุลที่ได้มักเรียกว่าHb A 1cซึ่งเป็นฮีโมโกลบินที่เกิดปฏิกิริยาไกลเคชั่น การจับตัวของกลูโคสกับกรดอะมิโนในฮีโมโกลบินเกิดขึ้นเองตามธรรมชาติ (โดยไม่ต้องอาศัยเอนไซม์) ในโปรตีนหลายชนิด แต่ยังไม่ทราบแน่ชัดว่ามีประโยชน์หรือไม่ อย่างไรก็ตาม เมื่อความเข้มข้นของกลูโคสในเลือดเพิ่มขึ้น เปอร์เซ็นต์ของ Hb A ที่เปลี่ยนเป็น Hb A 1cก็จะเพิ่มขึ้น ในผู้ป่วยเบาหวานที่มีระดับกลูโคสสูงเป็นประจำ เปอร์เซ็นต์ของ Hb A 1cก็จะสูงเช่นกัน เนื่องจากอัตราการรวมตัวของ Hb A กับกลูโคสเป็นไปอย่างช้าๆ เปอร์เซ็นต์ของ Hb A 1cจึงสะท้อนถึงค่าเฉลี่ยถ่วงน้ำหนักของระดับกลูโคสในเลือดตลอดช่วงอายุของเซลล์เม็ดเลือดแดง ซึ่งประมาณ 120 วัน[ 89 ]ดังนั้นจึงมีการวัดระดับฮีโมโกลบินไกลเคตเพื่อติดตามการควบคุมโรคเบาหวานชนิดที่ 2 (T2DM) ในระยะยาว การควบคุม T2DM ที่ไม่ดีส่งผลให้ระดับฮีโมโกลบินไกลเคตในเม็ดเลือดแดงสูงขึ้น ช่วงค่าอ้างอิงปกติอยู่ที่ประมาณ 4.0–5.9% แม้ว่าจะยากที่จะได้มา แต่แนะนำให้ผู้ป่วย T2DM มีค่าต่ำกว่า 7% ระดับที่สูงกว่า 9% สัมพันธ์กับการควบคุมฮีโมโกลบินไกลเคตที่ไม่ดี และระดับที่สูงกว่า 12% สัมพันธ์กับการควบคุมที่แย่มาก ผู้ป่วยเบาหวานที่รักษาระดับฮีโมโกลบินไกลเคตให้อยู่ใกล้เคียง 7% มีโอกาสที่ดีกว่ามากในการหลีกเลี่ยงภาวะแทรกซ้อนที่อาจเกิดขึ้นกับโรคเบาหวาน (มากกว่าผู้ที่มีระดับ 8% หรือสูงกว่า) [ 90 ]นอกจากนี้ ฮีโมโกลบินไกลเคตที่เพิ่มขึ้นจะเพิ่มความสัมพันธ์กับออกซิเจน ดังนั้นจึงป้องกันการปล่อยออกซิเจนที่เนื้อเยื่อและทำให้เกิดภาวะขาดออกซิเจนในกรณีที่รุนแรง[ 91 ]
ระดับฮีโมโกลบินที่สูงขึ้นสัมพันธ์กับจำนวนหรือขนาดของเซลล์เม็ดเลือดแดงที่เพิ่มขึ้น เรียกว่าภาวะเม็ดเลือด แดง มากเกินไป(polycythemia ) การเพิ่มขึ้นนี้อาจเกิดจากโรคหัวใจพิการแต่กำเนิด โรคหัวใจและปอด โรคปอดพังผืด ฮอร์โมนอิริโทรโปเอตินมากเกินไป หรือ ภาวะเม็ดเลือดแดงมากเกินไป ( polycythemia vera ) [ 92 ]ระดับฮีโมโกลบินที่สูงอาจเกิดจากการสัมผัสกับระดับความสูง การสูบบุหรี่ ภาวะขาดน้ำ (โดยการเพิ่มความเข้มข้นของ Hb) โรคปอดขั้นรุนแรง และเนื้องอกบางชนิด[ 59 ]
การใช้งานเพื่อการวินิจฉัย

การวัดความเข้มข้นของฮีโมโกลบินเป็นหนึ่งในการตรวจเลือด ที่ทำกันบ่อยที่สุด โดยปกติจะเป็นส่วนหนึ่งของการตรวจนับเม็ดเลือดครบถ้วนตัวอย่างเช่น มักจะทำการทดสอบก่อนหรือหลังการบริจาคเลือดผลลัพธ์จะรายงานเป็นg / L , g/ dLหรือmol /L 1 g/dL เท่ากับประมาณ 0.6206 mmol/L แม้ว่าหน่วยหลังจะไม่ค่อยได้ใช้บ่อยนักเนื่องจากความไม่แน่นอนเกี่ยวกับสถานะพอลิเมอร์ของโมเลกุล[ 93 ]ปัจจัยการแปลงนี้ โดยใช้น้ำหนักโมเลกุลของหน่วยโกลบินเดี่ยวที่ 16,000 Daเป็นที่นิยมใช้สำหรับการวัดความเข้มข้นของฮีโมโกลบินในเลือด สำหรับ MCHC (ความเข้มข้นของฮีโมโกลบินในเม็ดเลือดแดงเฉลี่ย) ปัจจัยการแปลง 0.155 ซึ่งใช้น้ำหนักของเตตระเมอร์ที่ 64,500 Da เป็นที่นิยมใช้มากกว่า[ 94 ]ระดับปกติคือ:
- ผู้ชาย: 13.8 ถึง 18.0 กรัม/เดซิลิตร (138 ถึง 180 กรัม/ลิตร หรือ 8.56 ถึง 11.17 มิลลิโมล/ลิตร)
- ผู้หญิง: 12.1 ถึง 15.1 กรัม/เดซิลิตร (121 ถึง 151 กรัม/ลิตร หรือ 7.51 ถึง 9.37 มิลลิโมล/ลิตร)
- เด็ก: 11 ถึง 16 กรัม/เดซิลิตร (110 ถึง 160 กรัม/ลิตร หรือ 6.83 ถึง 9.93 มิลลิโมล/ลิตร)
- หญิงตั้งครรภ์: 11 ถึง 14 กรัม/เดซิลิตร (110 ถึง 140 กรัม/ลิตร หรือ 6.83 ถึง 8.69 มิลลิโมล/ลิตร) (ค่าปกติระหว่างตั้งครรภ์คือ 9.5 ถึง 15) [ 95 ] [ b ]
ค่าปกติของฮีโมโกลบินในไตรมาสที่ 1 และ 3 ของหญิงตั้งครรภ์ต้องมีอย่างน้อย 11 กรัม/เดซิลิตร และอย่างน้อย 10.5 กรัม/เดซิลิตรในไตรมาสที่ 2 [ 97 ]
ภาวะขาดน้ำหรือภาวะน้ำเกินในร่างกายสามารถส่งผลต่อระดับฮีโมโกลบินที่วัดได้เป็นอย่างมาก อัลบูมินสามารถบ่งชี้สถานะความชุ่มชื้นของร่างกายได้
หากความเข้มข้นต่ำกว่าปกติ จะเรียกว่าภาวะโลหิตจาง ภาวะโลหิตจางแบ่งตามขนาดของเซลล์เม็ดเลือดแดง ซึ่งเป็นเซลล์ที่มีฮีโมโกลบินในสัตว์มีกระดูกสันหลัง ภาวะโลหิตจางเรียกว่า "ไมโครไซติก" หากเซลล์เม็ดเลือดแดงมีขนาดเล็ก "แมโครไซติก" หากเซลล์เม็ดเลือดแดงมีขนาดใหญ่ และ "นอร์โมไซติก" หากเป็นกรณีอื่น ๆ
ค่าฮีมาโตคริตซึ่งเป็นสัดส่วนของปริมาตรเลือดที่ประกอบด้วยเม็ดเลือดแดง โดยทั่วไปจะมีค่าประมาณสามเท่าของความเข้มข้นของฮีโมโกลบินที่วัดได้ในหน่วย g/dL ตัวอย่างเช่น หากวัดฮีโมโกลบินได้ 17 g/dL ค่าฮีมาโตคริตจะเท่ากับ 51% [ 98 ]
วิธีการทดสอบฮีโมโกลบินในห้องปฏิบัติการต้องใช้ตัวอย่างเลือด (หลอดเลือดแดง หลอดเลือดดำ หรือหลอดเลือดฝอย) และการวิเคราะห์ด้วยเครื่องวิเคราะห์โลหิตวิทยาและเครื่องวัด CO-oximeter นอกจากนี้ ยังมีวิธีการทดสอบฮีโมโกลบิน (SpHb) แบบใหม่ที่ไม่ต้องผ่าตัดที่เรียกว่า Pulse CO-Oximetry ซึ่งมีความแม่นยำเทียบเท่ากับวิธีการที่ต้องผ่าตัด[ 99 ]
สามารถวัดความเข้มข้นของออกซีฮีโมโกลบินและดีออกซีฮีโมโกลบินได้อย่างต่อเนื่อง ในระดับภูมิภาค และไม่รุกรานโดยใช้NIRS [ 100 ] [ 101 ] [ 102 ] [ 103 ] [ 104 ] NIRSสามารถใช้ได้ทั้งบนศีรษะและกล้ามเนื้อ เทคนิคนี้มักใช้ในการวิจัย เช่น การฝึกกีฬาระดับสูง การยศาสตร์ การฟื้นฟูสมรรถภาพ การติดตามผู้ป่วย การวิจัยทารกแรกเกิด การติดตามการทำงานของสมองอินเทอร์เฟซระหว่างสมองกับคอมพิวเตอร์ ระบบทางเดินปัสสาวะ (การหดตัวของกระเพาะปัสสาวะ) ระบบประสาท (การเชื่อมโยงระหว่างระบบประสาทและหลอดเลือด) และอื่นๆ
สามารถวัดมวลฮีโมโกลบินในมนุษย์ได้โดยใช้เทคนิคการหายใจซ้ำคาร์บอนมอนอกไซด์ (CO) ที่ไม่ใช้สารกัมมันตรังสี ซึ่งใช้มานานกว่า 100 ปีแล้ว ด้วยเทคนิคนี้ จะสูดดมก๊าซ CO บริสุทธิ์ปริมาณเล็กน้อยเข้าไปและหายใจซ้ำเป็นเวลาสองสามนาที ในระหว่างการหายใจซ้ำ CO จะจับกับฮีโมโกลบินที่มีอยู่ในเม็ดเลือดแดง จากการเพิ่มขึ้นของ CO ในเลือดหลังจากช่วงเวลาการหายใจซ้ำ สามารถกำหนดมวลฮีโมโกลบินได้โดยใช้หลักการเจือจาง[ 105 ]
การควบคุมระดับ น้ำตาลในเลือดในระยะยาวสามารถวัดได้จากความเข้มข้นของ Hb A 1cการวัดโดยตรงจะต้องใช้ตัวอย่างจำนวนมากเนื่องจากระดับน้ำตาลในเลือดเปลี่ยนแปลงอย่างมากตลอดทั้งวัน Hb A 1cเป็นผลผลิตจากปฏิกิริยาที่ไม่สามารถย้อนกลับได้ของฮีโมโกลบิน A กับกลูโคสความเข้มข้น ของกลูโคสที่สูงขึ้น จะส่งผลให้มี Hb A 1c มากขึ้น เนื่องจากปฏิกิริยานี้ช้า สัดส่วนของ Hb A 1cจึงแสดงถึงระดับกลูโคสในเลือดโดยเฉลี่ยในช่วงครึ่งชีวิตของเม็ดเลือดแดง ซึ่งโดยทั่วไปประมาณ 120 วัน สัดส่วนของ Hb A 1cที่ 6.0% หรือน้อยกว่าแสดงถึงการควบคุมระดับกลูโคสที่ดีในระยะยาว ในขณะที่ค่าที่สูงกว่า 7.0% ถือว่าสูงขึ้น การทดสอบนี้มีประโยชน์อย่างยิ่งสำหรับผู้ป่วยเบาหวาน[ c ]
เครื่องสร้างภาพด้วยคลื่นแม่เหล็กไฟฟ้าเชิงฟังก์ชัน (fMRI) ใช้สัญญาณจากดีออกซีฮีโมโกลบิน ซึ่งไวต่อสนามแม่เหล็กเนื่องจากเป็นพาราแมกเนติก การวัดแบบรวมกับNIRSแสดงให้เห็นความสัมพันธ์ที่ดีกับทั้งสัญญาณออกซีฮีโมโกลบินและดีออกซีฮีโมโกลบินเมื่อเทียบกับสัญญาณBOLD [ 106 ]
การติดตามกิจกรรมกีฬาและการติดตามตนเอง
สามารถติดตามฮีโมโกลบินได้โดยไม่รุกราน เพื่อสร้างชุดข้อมูลเฉพาะบุคคลที่ติดตามผลกระทบของการเพิ่มความเข้มข้นและการเจือจางของเลือดจากกิจกรรมประจำวัน เพื่อให้เข้าใจประสิทธิภาพการกีฬาและการฝึกฝนได้ดียิ่งขึ้น นักกีฬามักกังวลเกี่ยวกับความอดทนและความเข้มข้นของการออกกำลังกาย เซ็นเซอร์ใช้ไดโอดเปล่งแสงที่ปล่อยแสงสีแดงและอินฟราเรดผ่านเนื้อเยื่อไปยังตัวตรวจจับแสง จากนั้นตัวตรวจจับจะส่งสัญญาณไปยังโปรเซสเซอร์เพื่อคำนวณการดูดซับแสงโดยโปรตีนฮีโมโกลบิน[ 107 ]เซ็นเซอร์นี้คล้ายกับ เครื่องวัดออกซิเจนใน เลือดแบบพัลส์ซึ่งประกอบด้วยอุปกรณ์ตรวจจับขนาดเล็กที่หนีบกับนิ้ว
ลักษณะที่คล้ายคลึงกันในสิ่งมีชีวิตที่ไม่มีกระดูกสันหลัง
สิ่งมีชีวิตในอาณาจักรพืชและสัตว์มีโปรตีนที่ทำหน้าที่ขนส่งและจับออกซิเจนอยู่หลากหลายชนิด สิ่งมีชีวิตต่างๆ รวมถึงแบคทีเรียโปรโตซัวและรา ล้วนมีโปรตีนคล้ายฮีโมโกลบิน ซึ่งมีบทบาทที่ทราบและคาดการณ์ไว้ ได้แก่ การจับกับลิแกนด์ที่ เป็นก๊าซแบบผันกลับได้ เนื่องจากโปรตีนเหล่านี้หลายชนิดมีโกลบินและส่วนประกอบ ฮีม (เหล็กในโครงสร้างพอร์ฟิรินแบบแบน) จึงมักถูกเรียกว่าฮีโมโกลบิน แม้ว่าโครงสร้างสามมิติโดยรวมจะแตกต่างจากฮีโมโกลบินของสัตว์มีกระดูกสันหลังมากก็ตาม โดยเฉพาะอย่างยิ่ง การแยกแยะ "ไมโอโกลบิน" และฮีโมโกลบินในสัตว์ชั้นต่ำมักเป็นไปไม่ได้ เพราะสิ่งมีชีวิตเหล่านี้บางชนิดไม่มีกล้ามเนื้อหรืออาจมีระบบไหลเวียนโลหิต ที่แยกต่างหาก แต่ไม่ใช่ระบบที่เกี่ยวข้องกับการขนส่งออกซิเจน (ตัวอย่างเช่นแมลงและสัตว์ขาปล้อง อื่นๆ หลายชนิด ) ในกลุ่มทั้งหมดนี้ โมเลกุลที่มีฮีม/โกลบิน (แม้แต่โกลบินโมโนเมอร์) ที่เกี่ยวข้องกับการจับก๊าซจะถูกเรียกว่าออกซีฮีโมโกลบิน นอกจากจะเกี่ยวข้องกับการขนส่งและการรับรู้ของออกซิเจนแล้ว พวกมันยังอาจเกี่ยวข้องกับการกำจัด NO, CO2 ,สารประกอบซัลไฟด์ และแม้กระทั่ง O2 ในสภาพแวดล้อมที่ต้องปราศจากออกซิเจน[ 108 ]พวกมันอาจเกี่ยวข้องกับการล้างพิษของสารคลอรีนในลักษณะที่คล้ายคลึงกับเอนไซม์ P450 และเพอร์ออกซิเดสที่มีฮีม

โครงสร้างของฮีโมโกลบินแตกต่างกันไปตามสปีชีส์ ฮีโมโกลบินพบได้ในสิ่งมีชีวิตทุกอาณาจักร แต่ไม่พบในสิ่งมีชีวิตทุกชนิด สปีชีส์ดั้งเดิม เช่น แบคทีเรีย โปรโตซัวสาหร่ายและพืชมักจะมีฮีโมโกลบินแบบโกลบินเดี่ยวหนอนตัวกลมหอยและกุ้ง หลายชนิด มีโมเลกุลหลายหน่วยย่อยขนาดใหญ่มาก ใหญ่กว่าในสัตว์มีกระดูกสันหลังมาก โดยเฉพาะอย่างยิ่ง ฮีโมโกลบินแบบไคเมอริกที่พบในเชื้อราและหนอนปล้อง ยักษ์ อาจมีทั้งโกลบินและโปรตีนชนิดอื่น[ 14 ]
หนึ่งในปรากฏการณ์และการใช้ประโยชน์ของฮีโมโกลบินที่น่าทึ่งที่สุดในสิ่งมีชีวิต คือหนอนท่อยักษ์ ( Riftia pachyptilaหรือ Vestimentifera) ซึ่งมีความยาวได้ถึง 2.4 เมตร และอาศัยอยู่ในปล่องภูเขาไฟ ใต้ทะเล แทนที่จะมีระบบย่อยอาหารหนอนเหล่านี้มีแบคทีเรียจำนวนมากซึ่งมีน้ำหนักครึ่งหนึ่งของตัวหนอน แบคทีเรียจะออกซิไดซ์ H₂S จากปล่องภูเขาไฟด้วย O₂ จากน้ำเพื่อผลิตพลังงานในการสร้างอาหารจากH₂Oและ CO₂ ส่วนบนของหนอนเป็นโครงสร้างรูปพัดสีแดงเข้ม ("กลุ่มขน") ซึ่งยื่นออกไปในน้ำและดูดซับ H₂S และ O₂ สำหรับแบคทีเรีย และดูดซับ CO₂ เพื่อใช้เป็นวัตถุดิบในการสังเคราะห์คล้ายกับพืชที่สังเคราะห์แสง โครงสร้างเหล่านี้มีสีแดงสดเนื่องจากมีฮีโมโกลบินที่ซับซ้อนอย่างยิ่งหลายชนิด ซึ่งมีสายโกลบินมากถึง 144 สาย แต่ละสายมีโครงสร้างฮีมที่เกี่ยวข้องอยู่ด้วย ฮีโมโกลบินเหล่านี้มีความโดดเด่นตรงที่สามารถขนส่งออกซิเจนได้แม้จะมีซัลไฟด์อยู่ และยังสามารถขนส่งซัลไฟด์ได้โดยไม่ถูก "วางยาพิษ" หรือถูกยับยั้งอย่างสมบูรณ์เหมือนฮีโมโกลบินในสายพันธุ์อื่นๆ ส่วนใหญ่[ 109 ] [ 110 ]
โปรตีนจับออกซิเจนชนิดอื่นๆ
- ไมโอโกลบิน
- พบได้ในเนื้อเยื่อกล้ามเนื้อของสัตว์มีกระดูกสันหลังหลายชนิด รวมทั้งมนุษย์ ซึ่งทำให้เนื้อเยื่อกล้ามเนื้อมีสีแดงหรือเทาเข้มที่โดดเด่น โครงสร้างและลำดับของมันคล้ายกับฮีโมโกลบินมาก แต่ไม่ใช่เทตราเมอร์ (หน่วยย่อยสี่หน่วย) แต่เป็นโมโนเมอร์ (หน่วยย่อยเดียว) ที่ขาดการจับกันแบบร่วมมือ (cooperative binding) มันทำหน้าที่เก็บสะสมออกซิเจนมากกว่าขนส่งออกซิเจน
- เฮโมไซยานิน
- โปรตีนชนิดนี้เป็นโปรตีนขนส่งออกซิเจนที่พบได้มากเป็นอันดับสองในธรรมชาติ พบในเลือดของสัตว์ขาปล้องและหอยหลายชนิด ใช้กลุ่มโปรสเตติกทองแดงแทนกลุ่มฮีมเหล็ก และมีสีน้ำเงินเมื่อได้รับออกซิเจน
- เฮเมอรีทริน
- สัตว์ไม่มีกระดูกสันหลังในทะเลบางชนิดและหนอนปล้องบางชนิดใช้โปรตีนที่ไม่ใช่ฮีมที่มีธาตุเหล็กเป็นส่วนประกอบนี้ในการลำเลียงออกซิเจนในเลือด โปรตีนนี้จะมีสีชมพู/ม่วงเมื่อมีออกซิเจน และใสเมื่อไม่มีออกซิเจน
- คลอโรครูโอริน
- พบได้ในหนอนปล้องหลายชนิด มีโครงสร้างคล้ายกับอีริโทรครูโอรินมาก แต่กลุ่มฮีมมีโครงสร้างแตกต่างกันอย่างเห็นได้ชัด มีสีเขียวเมื่อปราศจากออกซิเจน และมีสีแดงเมื่อมีออกซิเจน
- วานาบินส์
- สารเหล่านี้ รู้จักกันในชื่อวาเนเดียมโครมาเจนพบได้ในเลือดของเพรียงทะเลเคยมีการตั้งสมมติฐานว่าสารเหล่านี้ใช้โลหะวาเนเดียมเป็นกลุ่มโปรสเตติกที่จับกับออกซิเจน อย่างไรก็ตาม แม้ว่าสารเหล่านี้จะมีวาเนเดียมเป็นองค์ประกอบหลัก แต่ดูเหมือนว่าจะจับกับออกซิเจนได้น้อยมาก ดังนั้นจึงอาจมีหน้าที่อื่นที่ยังไม่ได้รับการอธิบาย (เพรียงทะเลยังมีฮีโมโกลบินอยู่บ้าง) สารเหล่านี้อาจทำหน้าที่เป็นสารพิษ
- เอริโทรครูโอริน
- พบได้ในหนอนปล้องหลายชนิด รวมทั้งไส้เดือนดินมันเป็นโปรตีนในเลือดขนาดใหญ่ที่ลอยตัวอิสระ ประกอบด้วยหน่วยย่อยของโปรตีนที่มีธาตุเหล็กและฮีมหลายสิบหน่วย หรืออาจถึงหลายร้อยหน่วย ที่เชื่อมต่อกันเป็นโปรตีนเชิงซ้อนเดียวที่มีมวลโมเลกุลมากกว่า 3.5 ล้านดาลตัน
- เลกฮีโมโกลบิน
- ในพืชตระกูลถั่ว เช่น อัลฟัลฟาหรือถั่วเหลือง แบคทีเรียตรึงไนโตรเจนในรากจะได้รับการปกป้องจากออกซิเจนโดยโปรตีนจับออกซิเจนที่มีเหล็กฮีมเป็นองค์ประกอบ เอนไซม์ที่ได้รับการปกป้องโดยเฉพาะคือไนโตรเจเนสซึ่งไม่สามารถลดก๊าซไนโตรเจนได้ในสภาวะที่มีออกซิเจนอิสระ
- โคโบโกลบิน
- โคโบโปรตีนเป็นสารสังเคราะห์ที่มีโคบอลต์เป็นองค์ประกอบหลัก เมื่อสัมผัสกับออกซิเจน โคโบโปรตีนจะไม่มีสี แต่จะมีสีเหลืองเมื่ออยู่ในเส้นเลือด
พบในเซลล์ที่ไม่ใช่เม็ดเลือดแดง
เซลล์ที่ไม่ใช่เม็ดเลือดแดงบางชนิด (เช่น เซลล์อื่นที่ไม่ใช่เซลล์เม็ดเลือดแดง) มีฮีโมโกลบินอยู่ ในสมอง เซลล์เหล่านี้ได้แก่เซลล์ประสาทโดปามีน A9 ใน ซับสแตนเซียไน ก ราเซลล์แอสโทรไซต์ในเปลือกสมองและฮิปโปแคมปัสและในโอลิโกเดนโดรไซต์ ที่โตเต็มที่ทั้งหมด [ 12 ] มีการเสนอแนะว่าฮีโมโกลบินในสมองของเซลล์เหล่านี้อาจช่วยให้ "สามารถเก็บออกซิเจนไว้เพื่อสร้างกลไก การรักษาสมดุลในสภาวะที่ขาดออกซิเจน ซึ่งมีความสำคัญอย่างยิ่งสำหรับเซลล์ประสาทโดปามีน A9 ที่มีการเผาผลาญสูงและมีความต้องการพลังงานสูง" [ 12 ]มีข้อสังเกตเพิ่มเติมว่า "เซลล์ประสาทโดปามีน A9 อาจมีความเสี่ยงเป็นพิเศษต่อการเสื่อมสภาพเนื่องจากขาดออกซิเจน เนื่องจากนอกจากจะมีกิจกรรมของไมโทคอนเดรียสูงแล้ว ยังอยู่ภายใต้ความเครียดออกซิเดชันอย่างรุนแรงที่เกิดจากการผลิตไฮโดรเจนเปอร์ออกไซด์ผ่านการออกซิเดชันด้วยตนเองและ/หรือการดีอะมิเนชันของโดปามีนโดยโมโนอะมีนออกซิเดส (MAO) และปฏิกิริยาของเหล็กเฟอร์รัสที่เข้าถึงได้ในภายหลังเพื่อสร้างอนุมูลไฮดรอกซิลที่เป็นพิษสูง" [ 12 ]นี่อาจอธิบายถึงความเสี่ยงของการเสื่อมสภาพของเซลล์เหล่านี้ในโรคพาร์กินสัน [ 12 ] เหล็กที่ได้จากฮีโมโกลบินในเซลล์เหล่านี้ไม่ใช่สาเหตุของความมืดหลังการตายของเซลล์เหล่านี้ (ที่มาของชื่อภาษาละติน substantia nigra ) แต่เป็นผลมาจากนิวโรเมลานิน
นอกเหนือจากสมองแล้ว ฮีโมโกลบินยังมีหน้าที่ที่ไม่เกี่ยวข้องกับการขนส่งออกซิเจน เช่น เป็นสารต้านอนุมูลอิสระและควบคุมการเผาผลาญธาตุเหล็กในแมโครฟาจ [ 111 ] เซลล์ถุงลม [ 112 ]และเซลล์เมแซงเจียลในไต[ 113 ]
ในประวัติศาสตร์และศิลปะ

ในทางประวัติศาสตร์ การเชื่อมโยงระหว่างสีของเลือดและสนิมเกิดขึ้นในการเชื่อมโยงของดาวอังคารกับเทพเจ้าแห่งสงครามของโรมัน เนื่องจากดาวเคราะห์มีสีส้มแดง ซึ่งทำให้คนโบราณนึกถึงเลือด แม้ว่าสีของดาวเคราะห์จะเกิดจากสารประกอบเหล็กที่รวมกับออกซิเจนในดินของดาวอังคาร แต่ก็เป็นความเข้าใจผิดทั่วไปที่ว่าเหล็กในฮีโมโกลบินและออกไซด์ของมันทำให้เลือดมีสีแดง สีแดงนั้นเกิดจากส่วนประกอบพอร์ฟิริน ของฮีโมโกลบินที่เหล็กจับอยู่ ไม่ใช่เหล็กเอง[ 114 ]แม้ว่าการจับกันและสถานะรีดอกซ์ของเหล็กจะส่งผลต่อการเปลี่ยนผ่านทางอิเล็กตรอนจาก pi เป็น pi* หรือ n เป็น pi* ของพอร์ฟิริน และด้วยเหตุนี้จึงส่งผลต่อลักษณะทางแสงของมัน
ศิลปินJulian Voss-Andreaeสร้างประติมากรรมชื่อHeart of Steel (Hemoglobin)ในปี 2548 โดยอิงจากโครงสร้างหลักของโปรตีน ประติมากรรมนี้ทำจากแก้วและเหล็กที่ผุกร่อนการเกิดสนิมโดยเจตนาของงานศิลปะที่เดิมทีดูเงางามนั้นสะท้อนถึงปฏิกิริยาทางเคมีพื้นฐานของฮีโมโกลบินที่ออกซิเจนจับกับเหล็ก[ 115 ] [ 116 ]
นิโคลัส ไบเออร์ ศิลปินชาวมอนทรีออล ได้สร้างประติมากรรมชื่อLustre (Hémoglobine) ซึ่งทำจากสแตนเลส แสดงโครงสร้างของโมเลกุลฮีโมโกลบิน ประติมากรรมนี้จัดแสดงอยู่ในห้องโถงของศูนย์วิจัยของ ศูนย์สุขภาพมหาวิทยาลัยแมคกิลล์ในมอนทรีออล ประติมากรรมมีขนาดประมาณ 10 เมตร × 10 เมตร × 10 เมตร[ 117 ] [ 118 ]
ดูเพิ่มเติม
- คาร์ บามิโนฮีโมโกลบิน (ฮีโมโกลบินที่จับกับคาร์บอนไดออกไซด์ )
- คาร์บอกซีฮีโมโกลบิน (Hb ที่เกี่ยวข้องกับ CO)
- คลอโรฟิลล์ (แมกนีเซียมฮีม)
- การตรวจนับเม็ดเลือดครบถ้วน
- เดลต้าโกลบิน
- เครื่องวัดฮีโมโกลบิน
- ฮีโมโปรตีน
- เมทฮีโมโกลบิน (เฟอร์ริกฮีโมโกลบิน หรือ เฟอร์ริฮีโมโกลบิน)
- ออกซีฮีโมโกลบิน (มีออกซิเจน โมเลกุล คู่ สีแดงเหมือนเลือด)
- Tegillarca granosa - "หอยสีเลือด"
- สารประกอบของวาสก้า – สารประกอบออร์กาโนเมทัลลิกของอิริเดียม โดดเด่นด้วยความสามารถในการจับกับ O₂ ได้อย่างย้อนกลับได้
อ่านเพิ่มเติม
- แคมป์เบลล์, เอ็มเค (1999). ชีวเคมี (ฉบับที่ 3). ฮาร์คอร์ต. ISBN 978-0-03-024426-1.
- Eshaghian, S; Horwich, TB; Fonarow, GC (2006). "ความสัมพันธ์ผกผันที่ไม่คาดคิดระหว่างระดับ HbA1c และอัตราการเสียชีวิตในผู้ป่วยเบาหวานและภาวะหัวใจล้มเหลวซิสโตลิกขั้นรุนแรง" Am Heart J . 151 (1): 91.e1–91.e6. doi : 10.1016/j.ahj.2005.10.008 . PMID 16368297 .
- Ganong, WF (2003). บทวิจารณ์สรีรวิทยาทางการแพทย์ (ฉบับที่ 21). Lange. ISBN 978-0-07-140236-1.
- Hager, T (1995). พลังแห่งธรรมชาติ: ชีวิตของไลนัส พอลลิง . Simon and Schuster. ISBN 978-0-684-80909-0.
- เฮเซลวูด, ลอเรน (2001) ขาดไม่ได้เลย: เรื่องราวของฮีโมโกลบินในยามเจ็บป่วยและในยามสุขภาพดีสำนักพิมพ์โนวาไซแอนซ์ISBN 1-56072-907-4
- คนีปป์ เจ, บาลาคริชแนน จี, เฉิน อาร์, เชน ทีเจ, ซาฮู เอสซี, โฮ เอ็นที, จิโอวานเนลลี เจแอล, ซิมปลาซานู วี, โฮ ซี, สปิโร ที (2005) "พลวัตของอัลโลสเตอรีในฮีโมโกลบิน: บทบาทของพันธะไทโรซีน เอช สุดท้าย" เจ โมล บิออล . 356 (2): 335– 53. ดอย : 10.1016/j.jmb.2005.11.006 . PMID 16368110 .
- Hardison, Ross C. (2012). "วิวัฒนาการของฮีโมโกลบินและยีนของมัน" . Cold Spring Harbor Perspectives in Medicine . 2 (12) a011627. doi : 10.1101/cshperspect.a011627 . ISSN 2157-1422 . PMC 3543078 . PMID 23209182 .
ลิงก์ภายนอก
- โปรตีโอพีเดียฮีโมโกลบิน
- สภาปฏิบัติการแห่งชาติเพื่อต่อต้านโรคโลหิตจางที่anemia.org
- การค้นพบฮีโมโกลบินชนิดใหม่ทำให้เกิดการวินิจฉัยผิดพลาดด้วยเครื่องวัดออกซิเจนในเลือด (pulse oxymeter) เก็บถาวรเมื่อ 2016-03-09 ที่Wayback Machineที่www.life-of-science.net เก็บถาวรเมื่อ 2017-03-07 ที่Wayback Machine
- บทความเกี่ยวกับฮีโมโกลบิน ที่caloriebite.in
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เฮโมโกลบิน
ฮีโมโกลบิน ( haemoglobin , HbหรือHgb ) เป็นโปรตีนที่มีธาตุเหล็กซึ่งช่วยในการขนส่งออกซิเจนในเม็ดเลือดแดง สัตว์...
ประวัติการวิจัย
แม้ว่าเลือดจะทราบกันดีว่าสามารถลำเลียงออกซิเจนได้ตั้งแต่ปี ค.ศ. 1794 เป็นอย่างน้อย [ 16 ] [ 17 ] แต่คุณสมบัติในการลำเลียงออกซิเจนของฮีโมโกลบินนั้นได้รับการอธิบายโดย Hünefeld ในปี ค.ศ. 1840 [ 18 ] ในปี ค.ศ.
พันธุศาสตร์
ฮีโมโกลบินประกอบด้วย หน่วยย่อยของโปรตีน ( โมเลกุล โกลบิน ) ซึ่งเป็น พอลิเปป ไทด์ โซ่ยาวที่พับงอของ กรดอะมิโน จำเพาะ ซึ่งเป็นตัวกำหนดคุณสมบัติทางเคมีและหน้าที่ของโปรตีน ลำดับกรดอะมิโนของพอลิเปปไทด์ใดๆ นั้นได้มา จาก การถอดรหัส จากส่วนหนึ่งของดีเอ็นเอ ซึ่งก็คือ...
วิวัฒนาการ
โปรตีนโกลบินและยีนของพวกมันจึงมีต้นกำเนิดทางวิวัฒนาการที่เก่าแก่ แม้แต่พืชก็ยังมีโกลบินชนิดหนึ่งที่เรียกว่า เลกฮีโมโกลบิน ซึ่งมีลำดับร่วมกับฮีโมโกลบินประมาณ 15% แม้ว่าความคล้ายคลึงในระดับนี้อาจเกิดจากความบังเอิญ...