อ่าน 19 นาที
ชีววิทยาการพัฒนาเชิงวิวัฒนาการ
ชีววิทยาการพัฒนาเชิงวิวัฒนาการ หรือที่เรียกกันอย่างไม่เป็นทางการว่า อีโว-เดโว (evo-devo ) เป็นสาขา การวิจัยทางชีววิทยา ที่เปรียบเทียบ กระบวนการพัฒนา ของ สิ่งมีชีวิต ต่าง ๆ...
ชีววิทยาการพัฒนาเชิงวิวัฒนาการ

ชีววิทยาการพัฒนาเชิงวิวัฒนาการหรือที่เรียกกันอย่างไม่เป็นทางการว่าอีโว-เดโว (evo-devo ) เป็นสาขาการวิจัยทางชีววิทยาที่เปรียบเทียบกระบวนการพัฒนาของสิ่งมีชีวิต ต่าง ๆ เพื่ออนุมานว่ากระบวนการพัฒนาเหล่านั้นวิวัฒนาการมาได้อย่างไร
สาขาวิชานี้เติบโตขึ้นจากจุดเริ่มต้นในศตวรรษที่ 19 ซึ่งในขณะนั้นวิทยาเอ็มบริโอเผชิญกับปริศนา: นักสัตววิทยาไม่ทราบว่าการพัฒนาของตัวอ่อนถูกควบคุมในระดับโมเลกุล อย่างไร ชาร์ลส์ ดาร์วินสังเกตว่าการมีตัวอ่อนที่คล้ายคลึงกันบ่งชี้ถึงบรรพบุรุษร่วมกัน แต่ความคืบหน้ามีน้อยจนกระทั่งถึงทศวรรษ 1970 จากนั้น เทคโนโลยี ดีเอ็นเอลูกผสม ได้นำวิทยาเอ็มบริโอมาผสานรวมกับ พันธุศาสตร์ระดับโมเลกุลในที่สุดการค้นพบที่สำคัญในช่วงแรกคือการค้นพบยีนโฮมีโอติก ซึ่งควบคุมการพัฒนาใน ยูคาริโอตหลากหลาย ชนิด
สาขาวิชานี้ประกอบด้วยแนวคิดวิวัฒนาการหลักหลายประการ ประการหนึ่งคือ ความเหมือนกันอย่างลึกซึ้ง ( deep homology ) ซึ่งเป็นการค้นพบว่าอวัยวะที่แตกต่างกัน เช่น ดวงตาของแมลง สัตว์มีกระดูกสันหลังและ หอย เซฟาโลพอด ซึ่งเคยเชื่อกันว่าวิวัฒนาการแยกจากกันนั้น ถูกควบคุมโดยยีนที่คล้ายคลึงกัน เช่นpax-6จากชุดเครื่องมือยีน evo-devoยีนเหล่านี้มีอายุเก่าแก่และได้รับการอนุรักษ์ไว้อย่างสูงแม้กระทั่งในไฟลัมต่างๆพวกมันสร้างรูปแบบในเวลาและพื้นที่ซึ่งกำหนดรูปร่างของตัวอ่อน และในที่สุดก็ก่อให้เกิดโครงสร้างร่างกายของสิ่งมีชีวิต อีกประการหนึ่งคือ สปีชีส์ต่างๆ ไม่แตกต่างกันมากนักในยีนโครงสร้าง เช่น ยีนที่เข้ารหัสเอนไซม์สิ่งที่แตกต่างกันคือวิธีการควบคุมการแสดงออกของยีนโดยยีนในชุดเครื่องมือยีนเหล่านี้ถูกนำมาใช้ซ้ำโดยไม่เปลี่ยนแปลงหลายครั้งในส่วนต่างๆ ของตัวอ่อนและในระยะต่างๆ ของการพัฒนา ก่อให้เกิดลำดับการควบคุมที่ซับซ้อน สลับยีนควบคุมอื่นๆ รวมถึงยีนโครงสร้างเปิดและปิดในรูปแบบที่แม่นยำ การนำยีน ที่มีหน้าที่หลากหลายมาใช้ซ้ำนี้อธิบายได้ว่าทำไมยีนเหล่านี้จึงได้รับการอนุรักษ์ไว้อย่างสูง เพราะการเปลี่ยนแปลงใดๆ ก็ตามจะส่งผลเสียหลายประการ ซึ่งการคัดเลือกโดยธรรมชาติจะต่อต้าน
ลักษณะ ทางสัณฐานวิทยาใหม่ๆและท้ายที่สุดคือสายพันธุ์ใหม่ๆ เกิดขึ้นจากการเปลี่ยนแปลงในชุดเครื่องมือ ไม่ว่าจะเป็นเมื่อยีนถูกแสดงออกในรูปแบบใหม่ หรือเมื่อยีนในชุดเครื่องมือได้รับหน้าที่เพิ่มเติม อีกความเป็นไปได้หนึ่งคือทฤษฎีแบบนีโอ-ลามาร์ค ที่ว่า การเปลี่ยนแปลงทางเอพิเจเนติกส์ จะ ถูกรวมเข้าด้วยกันในระดับยีนในภายหลังซึ่งอาจมีความสำคัญในช่วงต้นประวัติศาสตร์ของสิ่งมีชีวิตหลายเซลล์
ประวัติศาสตร์
ทฤษฎีในยุคแรก
นักปรัชญาเริ่มคิดเกี่ยวกับวิธีที่สัตว์ได้รับรูปร่างในครรภ์ตั้งแต่สมัยโบราณอริสโตเติลกล่าวใน ตำรา ฟิสิกส์ ของเขา ว่า ตามที่เอมเปโดคลีสกล่าวไว้ ความเป็นระเบียบ "เกิดขึ้นเอง" ในตัวอ่อนที่กำลังพัฒนา ใน ตำรา ส่วนต่างๆ ของสัตว์เขาโต้แย้งว่าทฤษฎีของเอมเปโดคลีสนั้นผิด ในคำอธิบายของอริสโตเติล เอมเปโดคลีสกล่าวว่ากระดูกสันหลังแบ่งออกเป็นกระดูกสันหลังเพราะตัวอ่อนบิดตัวและทำให้กระดูกสันหลังแตกเป็นชิ้นๆ อริสโตเติลโต้แย้งว่ากระบวนการนี้มีเป้าหมายที่กำหนดไว้ล่วงหน้า นั่นคือ "เมล็ดพันธุ์" ที่พัฒนาเป็นตัวอ่อนเริ่มต้นด้วย "ศักยภาพ" ที่มีอยู่แล้วที่จะกลายเป็นส่วนต่างๆ ของร่างกาย เช่น กระดูกสันหลัง นอกจากนี้ สัตว์แต่ละชนิดจะให้กำเนิดสัตว์ในชนิดเดียวกัน มนุษย์มีเพียงทารกที่เป็นมนุษย์เท่านั้น[ 2 ]
สรุป

ทฤษฎีการย้อนรอยวิวัฒนาการถูกเสนอโดยเอเตียน แซร์เรสในปี 1824–26 ซึ่งสะท้อนแนวคิดของโยฮันน์ ฟรีดริช เมคเคล ในปี 1808 พวกเขาโต้แย้งว่าตัวอ่อนของสัตว์ 'ชั้นสูง' ผ่านหรือย้อนรอยขั้นตอนต่างๆ ซึ่งแต่ละขั้นตอนจะคล้ายกับสัตว์ที่อยู่ต่ำกว่าในห่วงโซ่แห่งสิ่งมีชีวิตตัวอย่างเช่น สมองของตัวอ่อนมนุษย์ในตอนแรกดูเหมือนสมองของปลาจากนั้นก็เหมือนสมองของสัตว์เลื้อยคลานนกและสัตว์เลี้ยงลูกด้วยนมก่อนที่จะกลายเป็นมนุษย์ อย่างชัดเจน นักคัพภวิทยา คา ร์ล เอิร์นสต์ ฟอน แบร์คัดค้านเรื่องนี้ โดยโต้แย้งในปี 1828 ว่าไม่มีลำดับเชิงเส้นเหมือนในห่วงโซ่แห่งสิ่งมีชีวิตที่อิงตามแผนผังร่างกาย เดียว แต่เป็นกระบวนการ เอพิเจเน ซิสที่โครงสร้างต่างๆ เกิดการเปลี่ยนแปลง ฟอน แบร์ กลับจำแนกแผนผังร่างกาย ของสัตว์ออกเป็นสี่แบบที่แตกต่างกัน ได้แก่ แบบแผ่รัศมี เช่นดาวทะเลแบบหอย เช่นหอย กาบ แบบข้อต่อ เช่นกุ้งมังกรและสัตว์มีกระดูกสันหลัง เช่น ปลา นักสัตววิทยาส่วนใหญ่จึงเลิกใช้การสรุปผลย้อนหลัง แม้ว่าErnst Haeckelจะฟื้นฟูมันขึ้นมาอีกครั้งในปี พ.ศ. 2409 [ 4 ] [ 5 ] [ 6 ] [ 7 ] [ 8 ]
สัณฐานวิทยาเชิงวิวัฒนาการ

ตั้งแต่ต้นศตวรรษที่ 19 จนถึงเกือบตลอดศตวรรษที่ 20 วิชาคัพภวิทยาต้องเผชิญกับปริศนา สัตว์ต่างๆ ถูกพบว่าพัฒนาไปเป็นตัวเต็มวัยที่มีโครงสร้างร่างกาย แตกต่างกันอย่างมาก โดยมักจะผ่านขั้นตอนที่คล้ายคลึงกันจากไข่ แต่เหล่านักสัตววิทยาแทบไม่รู้อะไรเลยเกี่ยวกับวิธีการควบคุมการพัฒนาของตัวอ่อน ใน ระดับโมเลกุลและด้วยเหตุนี้จึงรู้เพียงเล็กน้อยเกี่ยวกับวิวัฒนาการของกระบวนการพัฒนา[ 9 ]ชาร์ลส์ ดาร์วินแย้งว่าโครงสร้างตัวอ่อนที่เหมือนกันบ่งชี้ถึงบรรพบุรุษร่วมกัน ตัวอย่างเช่น ดาร์วินอ้างถึงตัวอ่อนของเพรียงที่มีลักษณะคล้ายกุ้ง ในหนังสือ On the Origin of Species ปี 1859 ของเขา ซึ่ง ตัวเต็มวัย ที่เกาะติดอยู่กับ ที่นั้นดูไม่เหมือน สัตว์ขาปล้องอื่นๆ เลยลินเนียสและคูเวียร์ได้จัดจำแนกพวกมันเป็นหอย[ 10 ] [ 11 ]ดาร์วินยังสังเกตเห็นการค้นพบของอเล็กซานเดอร์ โควาเลฟสกี ว่า ทูนิเคทก็ไม่ใช่หอยเช่นกัน แต่ในระยะตัวอ่อนมีโนโตคอร์ดและช่องคอหอยซึ่งพัฒนามาจากชั้นเนื้อเยื่อต้นกำเนิดเดียวกันกับโครงสร้างที่เทียบเท่ากันในสัตว์มีกระดูกสันหลังและจึงควรจัดกลุ่มร่วมกับพวกมันเป็นคอร์เดต[ 10 ] [ 12 ]
ดังนั้น สัตววิทยาในศตวรรษที่ 19 จึงเปลี่ยนวิทยาเอ็มบริโอให้เป็นวิทยาศาสตร์เชิงวิวัฒนาการ โดยเชื่อมโยงวิวัฒนาการทางสายพันธุ์เข้ากับความเหมือนกันระหว่างชั้นเนื้อเยื่อของเอ็มบริโอ นักสัตววิทยาหลายคน รวมถึงฟริตซ์ มุลเลอร์เสนอให้ใช้วิทยาเอ็มบริโอเพื่อค้นหาความสัมพันธ์ทางสายพันธุ์ระหว่างกลุ่มสิ่งมีชีวิต มุลเลอร์แสดงให้เห็นว่าสัตว์จำพวกครัสเตเชียน มีตัวอ่อนระยะ นอพลิอุสร่วมกันและระบุชนิดของปรสิตหลายชนิดที่ก่อนหน้านี้ไม่ได้รับการยอมรับว่าเป็นสัตว์จำพวกครัสเตเชียน มุลเลอร์ยังตระหนักว่าการคัดเลือกโดยธรรมชาติจะต้องเกิดขึ้นกับตัวอ่อนเช่นเดียวกับที่เกิดขึ้นกับตัวเต็มวัย ซึ่งเป็นการหักล้างทฤษฎีการย้อนรอยวิวัฒนาการ (recapitulation) ที่จะต้องปกป้องตัวอ่อนจากการคัดเลือกโดยธรรมชาติ[ 10 ]แนวคิดอื่น ๆ ของ Haeckel เกี่ยวกับการวิวัฒนาการของการพัฒนาได้รับการพิสูจน์ได้ดีกว่าการทบทวน: เขาโต้แย้งในช่วงทศวรรษ 1870 ว่าการเปลี่ยนแปลงในจังหวะเวลา ( heterochrony ) และการเปลี่ยนแปลงในตำแหน่งภายในร่างกาย ( heterotopy ) ของลักษณะต่าง ๆ ของการพัฒนาตัวอ่อนจะขับเคลื่อนวิวัฒนาการโดยการเปลี่ยนรูปร่างของร่างกายของลูกหลานเมื่อเทียบกับบรรพบุรุษ ต้องใช้เวลาหนึ่งศตวรรษกว่าที่แนวคิดเหล่านี้จะได้รับการพิสูจน์ว่าถูกต้อง[ 13 ] [ 14 ] [ 15 ]

ในปี พ.ศ. 2460 D'Arcy Thompsonได้เขียนหนังสือเกี่ยวกับรูปร่างของสัตว์โดยแสดงให้เห็นด้วยคณิตศาสตร์ อย่างง่าย ว่าการเปลี่ยนแปลงเล็กน้อยของพารามิเตอร์เช่น มุมของเปลือกเกลียวของหอยทาก สามารถเปลี่ยนแปลง รูปร่างของสัตว์ ได้อย่างมาก แม้ว่าเขาจะชอบคำอธิบายเชิงกลไกมากกว่าเชิงวิวัฒนาการก็ตาม[ 17 ] [ 18 ]แต่หากปราศจากหลักฐานระดับโมเลกุล ความก้าวหน้าก็หยุดชะงัก[ 10 ]
ในปี พ.ศ. 2495 อลัน ทัวริงได้ตีพิมพ์บทความเรื่อง " พื้นฐานทางเคมีของการเกิดรูปร่าง " เกี่ยวกับการพัฒนารูปแบบในร่างกายของสัตว์ เขาเสนอว่าการเกิดรูปร่างสามารถอธิบายได้ด้วยระบบปฏิกิริยา-การแพร่กระจายซึ่งเป็นระบบของสารเคมีที่ทำปฏิกิริยาและสามารถแพร่กระจายไปทั่วร่างกายได้[ 16 ]เขาสร้างแบบจำลองปฏิกิริยาเคมีที่เร่งปฏิกิริยาโดยใช้สมการเชิงอนุพันธ์ย่อยแสดงให้เห็นว่ารูปแบบต่างๆ เกิดขึ้นเมื่อปฏิกิริยาเคมีผลิตทั้งตัวเร่งปฏิกิริยา (A) และตัวยับยั้ง (B) ที่ทำให้การผลิต A ช้าลง หาก A และ B แพร่กระจายในอัตราที่ต่างกัน A จะมีอิทธิพลในบางที่ และ B จะมีอิทธิพลในที่อื่นๆ นักชีวเคมีชาวรัสเซียบอริส เบลูซอฟได้ทำการทดลองที่มีผลลัพธ์คล้ายกัน แต่ไม่สามารถตีพิมพ์ได้เนื่องจากนักวิทยาศาสตร์ในเวลานั้นคิดว่าการสร้างระเบียบที่มองเห็นได้เป็นการละเมิดกฎข้อที่สองของอุณหพลศาสตร์[ 19 ]
การสังเคราะห์สมัยใหม่ของช่วงต้นศตวรรษที่ 20
ใน การสังเคราะห์สมัยใหม่ที่เรียกว่าในช่วงต้นศตวรรษที่ 20 ระหว่างปี 1918 ถึง 1930 โรนัลด์ ฟิชเชอร์ได้นำทฤษฎีวิวัฒนาการ ของดาร์วิน ซึ่งเน้นย้ำถึงการคัดเลือกโดยธรรมชาติพันธุกรรมและความแปรผันเข้า กับ กฎพันธุศาสตร์ของเกรกอร์ เมนเดล มาเป็นโครงสร้างที่สอดคล้องกันสำหรับชีววิทยาเชิงวิวัฒนาการนักชีววิทยาสันนิษฐานว่าสิ่งมีชีวิตเป็นภาพสะท้อนโดยตรงของยีนที่เป็นส่วนประกอบ: ยีนเหล่านี้เข้ารหัสโปรตีน ซึ่งสร้างร่างกายของสิ่งมีชีวิต วิถีทางชีวเคมี (และพวกเขาสันนิษฐานว่ารวมถึงสายพันธุ์ใหม่ด้วย) วิวัฒนาการผ่านการกลายพันธุ์ในยีนเหล่านี้ มันเป็นภาพที่เรียบง่าย ชัดเจน และครอบคลุมเกือบทั้งหมด แต่ก็ไม่ได้อธิบายถึงวิทยาเอ็มบริโอ[ 10 ] [ 20 ]ฌอน บี. แคร์โรลล์ได้แสดงความคิดเห็นว่าหากมีข้อมูลเชิงลึกของ evo-devo วิทยาเอ็มบริโอคงจะมีบทบาทสำคัญในการสังเคราะห์นี้อย่างแน่นอน[ 1 ]
นักคัพภวิทยาเชิงวิวัฒนาการGavin de Beer ได้ คาดการณ์ชีววิทยาการพัฒนาเชิงวิวัฒนาการ ไว้ในหนังสือEmbryos and Ancestors ในปี 1930 ของเขา [ 21 ]โดยแสดงให้เห็นว่าวิวัฒนาการสามารถเกิดขึ้นได้โดยheterochrony [ 22 ]เช่นการคงลักษณะของวัยเยาว์ไว้ในวัยผู้ใหญ่ [ 13 ] de Beer โต้แย้งว่าสิ่งนี้สามารถทำให้เกิดการเปลี่ยนแปลงที่ดูเหมือนฉับพลันในบันทึกฟอสซิลได้ เนื่องจากตัวอ่อนกลายเป็นฟอสซิลได้ไม่ดี เนื่องจากช่องว่างในบันทึกฟอสซิลถูกใช้เป็นข้อโต้แย้งต่อวิวัฒนาการแบบค่อยเป็นค่อยไปของดาร์วิน คำอธิบายของ de Beer จึงสนับสนุนจุดยืนของดาร์วิน[ 23 ]อย่างไรก็ตาม แม้จะมี de Beer การสังเคราะห์สมัยใหม่ส่วนใหญ่ก็ละเลยการพัฒนาของตัวอ่อนเพื่ออธิบายรูปร่างของสิ่งมีชีวิต เนื่องจากพันธุศาสตร์ประชากรดูเหมือนจะเป็นคำอธิบายที่เพียงพอว่ารูปร่างต่างๆ วิวัฒนาการมาได้อย่างไร[ 24 ] [ 25 ] [ a ]
โอเปรอนแลค

ในปี พ.ศ. 2504 Jacques Monod , Jean-Pierre ChangeuxและFrançois Jacobค้นพบlac operonในแบคทีเรียEscherichia coliมันเป็นกลุ่มยีนที่จัดเรียงอยู่ในวงจรควบคุม แบบป้อนกลับ เพื่อให้ผลิตภัณฑ์ของมันถูกสร้างขึ้นก็ต่อเมื่อ "ถูกเปิดใช้งาน" โดยสิ่งกระตุ้นจากสิ่งแวดล้อมเท่านั้น หนึ่งในผลิตภัณฑ์เหล่านี้คือเอนไซม์ที่ย่อยน้ำตาลแลคโตส และแลคโตสเองก็เป็นสิ่งกระตุ้นที่เปิดใช้งานยีน นี่เป็นการค้นพบครั้งสำคัญ เนื่องจากแสดงให้เห็นเป็นครั้งแรกว่ายีน แม้แต่ในสิ่งมีชีวิตขนาดเล็กอย่างแบคทีเรีย ก็อยู่ภายใต้การควบคุมที่แม่นยำ นัยยะก็คือยีนอื่นๆ อีกมากมายก็ได้รับการควบคุมอย่างซับซ้อนเช่นกัน[ 27 ]
การกำเนิดของวิวัฒนาการและการพัฒนา และการสังเคราะห์ครั้งที่สอง
ในปี พ.ศ. 2520 การปฏิวัติทางความคิดเกี่ยวกับการวิวัฒนาการและชีววิทยาการพัฒนาได้เริ่มต้นขึ้น ด้วยการมาถึงของ เทคโนโลยี ดีเอ็นเอลูกผสมในพันธุศาสตร์หนังสือOntogeny and PhylogenyโดยStephen J. Gouldและบทความ"Evolution and Tinkering" [ 28 ]โดยFrançois Jacob Gould ได้ลบล้างการตีความของ Haeckel เกี่ยวกับคัพภวิทยาเชิงวิวัฒนาการ ในขณะที่ Jacob ได้เสนอทฤษฎีทางเลือก[ 10 ] ซึ่งนำไปสู่การสังเคราะห์ครั้งที่สอง [ 29 ] [ 30 ]ในที่สุดก็รวมถึงคัพภวิทยา ตลอดจนพันธุศาสตร์โมเลกุลวิวัฒนาการชาติพันธุ์ และชีววิทยาเชิงวิวัฒนาการ เพื่อสร้าง evo-devo [ 31 ] [ 32 ]ในปี พ.ศ. 2521 Edward B. Lewisได้ค้นพบยีนโฮมีโอติก ที่ควบคุมการพัฒนาของตัวอ่อนในแมลงวันผลไม้ Drosophilaซึ่งเช่นเดียวกับแมลงทั้งหมดเป็นอา ร์โทรพอด ซึ่งเป็นหนึ่งใน ไฟลัมหลักของสัตว์ไม่มีกระดูกสันหลัง[ 33 ]บิล แม็กกินนิสค้นพบลำดับยีนโฮมีโอติก หรือโฮมีโอโบกซ์ในสัตว์ในไฟลัมอื่นๆ อย่างรวดเร็ว ในสัตว์มีกระดูกสันหลังเช่นกบ นกและสัตว์ เลี้ยง ลูกด้วยนมต่อมายังพบในเชื้อราเช่นยีสต์และในพืชอีกด้วย[ 34 ] [ 35 ]เห็นได้ชัดว่ามีความคล้ายคลึงกันอย่างมากในยีนที่ควบคุมการพัฒนาในยูคาริโอตทั้งหมด[ 36 ] ในปี 1980 คริสเตียน นุสส์ไลน์-โวลฮาร์ดและเอริค วีสเชาส์ได้อธิบายถึงยีนช่องว่างซึ่งช่วยสร้างรูปแบบการแบ่งส่วนในตัวอ่อนแมลงวันผลไม้ [ 37 ] [ 38 ]พวกเขาและลูอิสได้รับรางวัลโนเบลจากผลงานของพวกเขาในปี1995 [ 34 ] [ 39 ]
ต่อมามีการค้นพบความคล้ายคลึงที่เฉพาะเจาะจงมากขึ้น เช่น ยีน distal-lessถูกพบในปี 1989 ว่ามีส่วนเกี่ยวข้องกับการพัฒนาของระยางค์หรือแขนขาในแมลงวันผลไม้[ 40 ]ครีบของปลา ปีกของไก่พาราโพเดียของหนอนทะเล แอนเน ลิดแอมพูลลาและไซฟอนของทูนิเคต และเท้าท่อของเม่นทะเลเป็นที่ชัดเจนว่ายีนนี้ต้องมีอายุเก่าแก่ ย้อนกลับไปถึงบรรพบุรุษร่วมสุดท้ายของสัตว์ที่มีสมมาตรสองด้าน (ก่อน ยุค เอเดียคารันซึ่งเริ่มต้นเมื่อประมาณ 635 ล้านปีก่อน) วิวัฒนาการและการพัฒนาได้เริ่มเปิดเผยวิธีการสร้างร่างกายของสัตว์ทุกชนิดในระหว่างการพัฒนา[ 41 ] [ 42 ]
การควบคุมโครงสร้างร่างกาย
ความเหมือนกันเชิงลึก
ไข่ทรงกลมโดยประมาณของสัตว์ต่าง ๆ ก่อให้เกิดรูปร่างที่แตกต่างกัน ตั้งแต่แมงกะพรุนไปจนถึงกุ้งมังกร ผีเสื้อไปจนถึงช้าง สิ่งมีชีวิตเหล่านี้หลายชนิดมียีนโครงสร้างเดียวกันสำหรับโปรตีนที่สร้างร่างกาย เช่น คอลลาเจนและเอนไซม์ แต่เหล่านักชีววิทยาคาดหวังว่าสัตว์แต่ละกลุ่มจะมีกฎการพัฒนาของตัวเอง ความประหลาดใจของวิวัฒนาการและการพัฒนาคือการสร้างรูปร่างของร่างกายถูกควบคุมโดยยีนเพียงไม่กี่เปอร์เซ็นต์ และยีนควบคุมเหล่านี้เป็นยีนโบราณที่สัตว์ทุกชนิดมีร่วมกันยีราฟไม่มียีนสำหรับคอยาว เช่นเดียวกับที่ช้างไม่มียีนสำหรับลำตัวใหญ่ ร่างกายของพวกมันถูกกำหนดรูปแบบโดยระบบการสลับซึ่งทำให้การพัฒนาลักษณะต่าง ๆ เริ่มต้นเร็วขึ้นหรือช้าลง เกิดขึ้นในส่วนนี้หรือส่วนนั้นของตัวอ่อน และดำเนินต่อไปเป็นระยะเวลามากหรือน้อย[ 9 ]
ปริศนาเกี่ยวกับการควบคุมการพัฒนาของตัวอ่อนเริ่มได้รับการแก้ไขโดยใช้แมลงวันผลไม้Drosophila melanogasterเป็นแบบจำลองสิ่งมีชีวิต การควบคุม การเกิดตัวอ่อนทีละขั้นตอนได้รับการแสดงให้เห็นโดยการติด สี ย้อมเรืองแสงที่มีสีต่างกันเข้ากับโปรตีนชนิดเฉพาะที่สร้างขึ้นโดยยีนที่แสดงออกในตัวอ่อน[ 9 ] โดยทั่วไปแล้ว สีย้อมเช่นโปรตีนเรืองแสงสีเขียวซึ่งเดิมมาจากแมงกะพรุนจะถูกติดเข้ากับแอนติบอดีที่จำเพาะต่อโปรตีนของแมลงวันผลไม้ ทำให้เกิดตัวบ่งชี้ที่แม่นยำว่าโปรตีนนั้นปรากฏขึ้นที่ใดและเมื่อใดในตัวอ่อนที่มีชีวิต[ 43 ]

ด้วยเทคนิคดังกล่าว ในปี 1994 Walter Gehringพบว่า ยีน pax-6ซึ่งมีความสำคัญต่อการสร้างดวงตาของแมลงวันผลไม้ ตรงกับยีนสร้างดวงตาในหนูและมนุษย์อย่างแม่นยำ ยีนเดียวกันนี้ถูกพบอย่างรวดเร็วในสัตว์กลุ่มอื่นๆ อีกมากมาย เช่นปลาหมึกซึ่ง เป็นหอยในกลุ่ม เซฟาโลพอดนักชีววิทยาหลายคน รวมถึงErnst Mayrเชื่อว่าดวงตาเกิดขึ้นในอาณาจักรสัตว์อย่างน้อย 40 ครั้ง เนื่องจากกายวิภาคของดวงตาประเภทต่างๆ แตกต่างกันอย่างมาก[ 9 ]ตัวอย่างเช่นดวงตาประกอบ ของแมลงวันผลไม้ ประกอบด้วยโครงสร้างเลนส์ขนาดเล็กหลายร้อยอัน ( ommatidia ) ดวงตาของมนุษย์มีจุดบอดตรงที่เส้นประสาทตาเข้าสู่ดวงตา และเส้นใยประสาทวิ่งผ่านพื้นผิวของเรตินาดังนั้นแสงจึงต้องผ่านชั้นของเส้นใยประสาทก่อนที่จะไปถึงเซลล์ตรวจจับในเรตินา ดังนั้นโครงสร้างจึงมีลักษณะ "กลับหัว" ในทางตรงกันข้าม ดวงตาของเซฟาโลพอดมีเรตินา จากนั้นเป็นชั้นของเส้นใยประสาท จากนั้นเป็นผนังของดวงตา "ในทิศทางที่ถูกต้อง" [ 44 ] อย่างไรก็ตาม หลักฐานของpax-6แสดงให้เห็นว่ายีนเดียวกันควบคุมการพัฒนาของดวงตาของสัตว์เหล่านี้ทั้งหมด ซึ่งบ่งชี้ว่าพวกมันทั้งหมดวิวัฒนาการมาจากบรรพบุรุษร่วมกัน[ 9 ]ยีนโบราณได้รับการอนุรักษ์ไว้ตลอดวิวัฒนาการหลายล้านปีเพื่อสร้างโครงสร้างที่แตกต่างกันสำหรับฟังก์ชันที่คล้ายคลึงกัน แสดงให้เห็นถึงความเหมือนกันอย่างลึกซึ้งระหว่างโครงสร้างที่เคยคิดว่าเป็นเพียงความคล้ายคลึงกันเท่านั้น[ 45 ] [ 46 ]แนวคิดนี้ได้รับการขยายไปสู่วิวัฒนาการของการเกิดตัวอ่อน ในภายหลัง [ 47 ]และทำให้เกิดการแก้ไขความหมายของความเหมือนกันในชีววิทยาเชิงวิวัฒนาการอย่างรุนแรง[ 45 ] [ 46 ] [ 1 ]
ชุดเครื่องมือยีน

ยีนเพียงส่วนน้อยในจีโนมของสิ่งมีชีวิตควบคุมการพัฒนาของสิ่งมีชีวิตนั้น ยีนเหล่านี้เรียกว่าชุดเครื่องมือทางพันธุกรรมเพื่อการพัฒนา (developmental-genetic toolkit) ยีนเหล่านี้มีการอนุรักษ์ไว้อย่างสูงในกลุ่มไฟลัมต่างๆหมายความว่าพวกมันมีมาแต่โบราณและคล้ายคลึงกันมากในกลุ่มสัตว์ที่อยู่ห่างไกลกัน ความแตกต่างในการใช้งานของยีนในชุดเครื่องมือส่งผลต่อโครงสร้างร่างกาย จำนวน ชนิด และรูปแบบของส่วนต่างๆ ของร่างกาย ยีนในชุดเครื่องมือส่วนใหญ่เป็นส่วนหนึ่งของวิถีการส่งสัญญาณ : พวกมันเข้ารหัสปัจจัยการ ถอดรหัส โปรตีนยึดเกาะเซลล์ โปรตีน ตัวรับบนพื้นผิวเซลล์และลิแกนด์ ส่งสัญญาณ ที่จับกับพวกมัน รวมถึงมอร์โฟเจนที่หลั่งออกมาและแพร่กระจายไปทั่วตัวอ่อน ทั้งหมดนี้ช่วยกำหนดชะตากรรมของเซลล์ที่ยังไม่แตกต่างในตัวอ่อน ร่วมกันพวกมันสร้างรูปแบบในเวลาและพื้นที่ซึ่งกำหนดรูปร่างของตัวอ่อน และในที่สุดก็ก่อให้เกิดโครงสร้างร่างกายของสิ่งมีชีวิต ในบรรดายีนในชุดเครื่องมือที่สำคัญที่สุดได้แก่ยีนHoxปัจจัยการถอดรหัสเหล่านี้มี โมทีฟ DNA ที่จับกับโปรตีน โฮมีโอโบกซ์ซึ่งพบได้ในยีนชุดเครื่องมืออื่นๆ ด้วย และสร้างรูปแบบพื้นฐานของร่างกายตามแกนหน้า-หลัง[ 1 ] ยีน Hox กำหนดตำแหน่งที่ส่วนที่ซ้ำกัน เช่นกระดูกสันหลัง จำนวนมาก ของงูจะเติบโตในเอ็มบริโอหรือตัวอ่อนที่กำลังพัฒนา[ 9 ] Pax-6ที่กล่าวถึงไปแล้ว เป็นยีนชุดเครื่องมือแบบคลาสสิก[ 48 ]แม้ว่ายีนชุดเครื่องมืออื่นๆ จะมีส่วนเกี่ยวข้องในการสร้างโครงสร้างร่างกาย ของพืช [ 49 ] แต่ยีน โฮมีโอโบก ซ์ก็พบได้ในพืชเช่นกัน ซึ่งหมายความ ว่า ยีนเหล่านี้มีอยู่ในยูคาริโอต ทั้งหมด [ 50 ] [ 51 ] [ 52 ]
เครือข่ายควบคุมของตัวอ่อน

ผลิตภัณฑ์โปรตีนของชุดเครื่องมือควบคุมไม่ได้ถูกนำกลับมาใช้ใหม่โดยการทำซ้ำและการดัดแปลง แต่โดยโมเสกที่ซับซ้อนของพลีโอโทรปี กล่าวคือ ถูกนำไปใช้โดยไม่เปลี่ยนแปลงในกระบวนการพัฒนาอิสระหลายอย่าง ทำให้เกิดรูปแบบในโครงสร้างร่างกายที่แตกต่างกันหลายอย่าง[ 1 ]ตำแหน่งของยีนชุดเครื่องมือพลีโอโทรปีเหล่านี้มีองค์ประกอบควบคุมซิส ขนาดใหญ่ ซับซ้อน และเป็นแบบโมดูลาร์ ตัวอย่างเช่น ในขณะที่ยีน โรดอปซินที่ไม่พลีโอโทรปีในแมลงวันผลไม้มีองค์ประกอบควบคุมซิสที่มีความยาว เพียงไม่กี่ร้อย คู่เบส แต่ บริเวณควบคุมซิสของยีนeyeless ที่พลีโอโทรปีนั้นมีองค์ประกอบควบคุมซิส 6 ตัวในความยาวมากกว่า 7000 คู่เบส [ 1 ]เครือข่ายควบคุมที่เกี่ยวข้องมักมีขนาดใหญ่มาก โปรตีนควบคุมแต่ละตัวควบคุมองค์ประกอบควบคุมซิส "หลายสิบถึงหลายร้อย" ตัว ตัวอย่างเช่น ปัจจัยการถอดรหัสของแมลงวันผลไม้ 67 ตัวควบคุมยีนเป้าหมายโดยเฉลี่ย 124 ยีนต่อตัว[ 1 ]ความซับซ้อนทั้งหมดนี้ทำให้ยีนที่เกี่ยวข้องกับการพัฒนาของตัวอ่อนสามารถเปิดและปิดได้ในเวลาที่เหมาะสมและในสถานที่ที่เหมาะสม ยีนบางส่วนเหล่านี้เป็นยีนโครงสร้างที่สร้างเอนไซม์ เนื้อเยื่อ และอวัยวะของตัวอ่อนโดยตรง แต่ยีนอื่นๆ อีกมากมายเป็นยีนควบคุม ดังนั้นสิ่งที่ถูกเปิดใช้งานมักจะเป็นลำดับการเปิดปิดที่กำหนดเวลาอย่างแม่นยำ ซึ่งเกี่ยวข้องกับการเปิดกระบวนการพัฒนาทีละอย่างในตัวอ่อนที่กำลังพัฒนา[ 1 ]

เครือข่ายควบคุมแบบเรียงลำดับดังกล่าวได้รับการศึกษาอย่างละเอียดในการพัฒนาตัวอ่อนของแมลงวันผลไม้ตัวอ่อนที่ยังเล็กมีรูปร่างเป็นรูปไข่คล้ายลูกรักบี้ยีนจำนวนเล็กน้อยสร้างอาร์เอ็นเอส่งสารที่สร้างความเข้มข้นไล่ระดับตามแกนยาวของตัวอ่อน ในตัวอ่อนระยะแรก ยีน bicoidและhunchbackมีความเข้มข้นสูงใกล้กับส่วนหน้า และให้รูปแบบแก่ส่วนหัวและอกในอนาคต ยีน caudalและnanosมีความเข้มข้นสูงใกล้กับส่วนท้าย และให้รูปแบบแก่ส่วนท้องส่วนท้ายสุด ผลกระทบของยีนเหล่านี้มีปฏิสัมพันธ์กัน ตัวอย่างเช่น โปรตีน Bicoid จะปิดกั้นการแปล อาร์เอ็นเอส่งสาร ของcaudalดังนั้นความเข้มข้นของโปรตีน Caudal จึงต่ำที่ส่วนหน้า ต่อมา Caudal จะเปิดใช้งานยีนที่สร้างส่วนท้ายสุดของแมลงวัน แต่เฉพาะที่ส่วนท้ายซึ่งมีความเข้มข้นสูงสุด[ 53 ] [ 54 ]
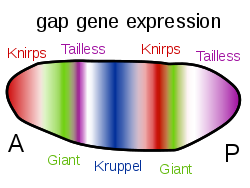
โปรตีน Bicoid, Hunchback และ Caudal จะควบคุมการถอดรหัสของยีน gapเช่นgiant , knirps , Krüppelและtaillessในรูปแบบลายทาง ทำให้เกิดโครงสร้างระดับแรกที่จะกลายเป็นส่วนต่างๆ[ 37 ]โปรตีนเหล่านี้จะควบคุมยีน pair-ruleซึ่งในขั้นตอนต่อไปจะสร้างแถบ 7 แถบตามแกนยาวของตัวอ่อน สุดท้าย ยีน segment polarity เช่นengrailedจะแบ่งแต่ละแถบทั้ง 7 แถบออกเป็นสองส่วน ทำให้เกิดส่วนต่างๆ ในอนาคต 14 ส่วน[ 53 ] [ 54 ]
กระบวนการนี้อธิบายถึงการอนุรักษ์ลำดับยีนชุดเครื่องมืออย่างแม่นยำ ซึ่งส่งผลให้เกิดความเหมือนกันอย่างลึกซึ้งและความเท่าเทียมกันในการทำงานของโปรตีนชุดเครื่องมือในสัตว์ที่แตกต่างกัน (ตัวอย่างเช่น เมื่อโปรตีนของหนูควบคุมการพัฒนาของแมลงวันผลไม้) ปฏิสัมพันธ์ของปัจจัยการถอดรหัสและองค์ประกอบควบคุมซิส หรือของโปรตีนส่งสัญญาณและตัวรับ จะถูกล็อกไว้ผ่านการใช้งานหลายครั้ง ทำให้การกลายพันธุ์เกือบทุกชนิดเป็นอันตรายและถูกกำจัดโดยการคัดเลือกโดยธรรมชาติ[ 1 ]
กลไกที่กำหนด แกนหน้า-หลังของ สัตว์ ทุกชนิดนั้น เหมือนกัน ซึ่งบ่งชี้ถึงบรรพบุรุษร่วมกัน มีกลไกที่คล้ายกันสำหรับแกนหลัง-ท้องสำหรับ สัตว์ ที่มีสมมาตรสองด้านแต่จะกลับกันระหว่างสัตว์ขาปล้องและสัตว์มีกระดูกสันหลัง [ 55 ] กระบวนการอีกอย่างหนึ่งคือการเกิดแกสตรูเลชันของตัวอ่อน ซึ่งขับเคลื่อนโดย มอเตอร์โมเลกุล ไมโอซิน IIซึ่งไม่ได้รับการอนุรักษ์ไว้ในสปีชีส์ต่างๆ กระบวนการนี้อาจเริ่มต้นจากการเคลื่อนที่ของน้ำทะเลในสิ่งแวดล้อม ต่อมาถูกแทนที่ด้วยวิวัฒนาการของการเคลื่อนที่ของเนื้อเยื่อในตัวอ่อน[ 56 ] [ 57 ] [ 58 ]
จุดเริ่มต้นของความแปลกใหม่
ผลลัพธ์ ที่น่าประหลาดใจและอาจขัดแย้งกับสามัญสำนึก (จาก มุมมองของ นีโอ-ดาร์วิน ) จากการวิจัยล่าสุดในชีววิทยาการพัฒนาเชิงวิวัฒนาการคือ ความหลากหลายของแผนผังร่างกายและสัณฐานวิทยาในสิ่งมีชีวิตในไฟลัม ต่างๆ มากมาย นั้น ไม่จำเป็นต้องสะท้อนถึงความหลากหลายในระดับลำดับของยีน รวมถึงยีนของชุดเครื่องมือทางพันธุกรรมในการพัฒนาและยีนอื่นๆ ที่เกี่ยวข้องกับการพัฒนา อันที่จริง ดังที่ John Gerhart และ Marc Kirschner ได้กล่าวไว้ มีความขัดแย้งที่เห็นได้ชัด: "ในที่ที่เราคาดหวังว่าจะพบความแปรผันมากที่สุด เรากลับพบการอนุรักษ์ การขาดการเปลี่ยนแปลง" [ 59 ]ดังนั้น หากความแปลกใหม่ทางสัณฐานวิทยาที่สังเกตได้ระหว่างกลุ่ม ต่างๆ ไม่ได้มาจากการเปลี่ยนแปลงในลำดับยีน (เช่น โดยการกลายพันธุ์ ) แล้วมันมาจากไหน? ความแปลกใหม่อาจเกิดขึ้นจากการเปลี่ยนแปลงที่ขับเคลื่อนโดยการกลายพันธุ์ใน การ ควบคุมยีน[ 1 ] [ 60 ] [ 61 ] [ 62 ]
ความหลากหลายในชุดเครื่องมือ


ความแปรผันในชุดเครื่องมืออาจเป็นส่วนสำคัญของการวิวัฒนาการทางสัณฐานวิทยาของสัตว์ ชุดเครื่องมือสามารถขับเคลื่อนวิวัฒนาการได้สองวิธี ยีนชุดเครื่องมือสามารถแสดงออกในรูปแบบที่แตกต่างกัน เช่น เมื่อจงอยปากของนกฟินช์พื้นดินขนาดใหญ่ ของดาร์วิน ขยายใหญ่ขึ้นโดยยีนBMP [ 63 ]หรือเมื่องูสูญเสียขาเนื่องจาก ยีน distal-lessแสดงออกน้อยลงหรือไม่แสดงออกเลยในบริเวณที่สัตว์เลื้อยคลานชนิดอื่นยังคงสร้างแขนขา[ 64 ]หรือยีนชุดเครื่องมือสามารถได้รับหน้าที่ใหม่ ดังที่เห็นได้จากหน้าที่มากมายของยีนdistal-less เดียวกัน ซึ่งควบคุมโครงสร้างที่หลากหลาย เช่น ขากรรไกรในสัตว์มีกระดูกสันหลัง[ 65 ] [ 66 ]ขาและหนวดในแมลงวันผลไม้[ 67 ]และรูปแบบจุดตาบนปีกผีเสื้อ [ 68 ] เนื่องจากการเปลี่ยนแปลงเล็กน้อยในยีนชุดเครื่องมือสามารถทำให้เกิดการเปลี่ยนแปลงอย่างมีนัยสำคัญใน โครงสร้างร่างกาย จึงมักทำให้เกิดหน้าที่เดียวกันแบบบรรจบกันหรือแบบขนานdistal-less สร้างรูปแบบปีกในผีเสื้อHeliconius eratoและHeliconius melpomeneซึ่งเป็นผีเสื้อเลียนแบบ Müllerianในสิ่งที่เรียกว่าการแปรผันที่อำนวยความสะดวก [ 69 ]รูปแบบปีกของพวกมันเกิดขึ้นในเหตุการณ์วิวัฒนาการที่แตกต่างกัน แต่ถูกควบคุมโดยยีนเดียวกัน[ 70 ]การเปลี่ยนแปลงการพัฒนาสามารถมีส่วนช่วยโดยตรงต่อการเกิดสปีชีส์[ 71 ]
การรวมตัวของการเปลี่ยนแปลงทางเอพิเจเนติกส์
นวัตกรรมเชิงวิวัฒนาการบางครั้งอาจเริ่มต้นในรูปแบบลามาร์คด้วย การเปลี่ยนแปลง ทางพันธุกรรมของการควบคุมยีนหรือการสร้างฟีโนไทป์ซึ่งต่อมาได้รับการเสริมความแข็งแกร่งด้วยการเปลี่ยนแปลงในระดับยีนการเปลี่ยนแปลงทางพันธุกรรมรวมถึงการดัดแปลง DNA โดยการเติมหมู่เมทิลแบบย้อนกลับได้[ 72 ]ตลอดจนการปรับเปลี่ยนรูปร่างของสิ่งมีชีวิตโดยไม่เป็นไปตามโปรแกรมโดยผลกระทบทางกายภาพและสิ่งแวดล้อมอื่นๆ อันเนื่องมาจากความยืดหยุ่น โดยธรรมชาติ ของกลไกการพัฒนา[ 73 ]นักชีววิทยาStuart A. NewmanและGerd B. Müllerได้เสนอแนะว่าสิ่งมีชีวิตในช่วงต้นของประวัติศาสตร์ของสิ่งมีชีวิตหลายเซลล์มีความอ่อนไหวต่อการกำหนดทางพันธุกรรมประเภทที่สองนี้มากกว่าสิ่งมีชีวิตในปัจจุบัน ซึ่งเป็นพื้นฐานสำหรับการเปลี่ยนแปลงวิวัฒนาการระดับมหภาค ในยุคแรก [ 74 ]
อคติในการพัฒนา

การพัฒนาในสายพันธุ์เฉพาะอาจมีอคติไปในทางบวก มุ่งไปสู่เส้นทางหรือฟีโนไทป์ที่กำหนด[ b ]หรือในทางลบ หลีกเลี่ยงการเปลี่ยนแปลงบางประเภท อาจเป็นแบบสัมบูรณ์ (การเปลี่ยนแปลงเกิดขึ้นเสมอหรือไม่เคยเกิดขึ้นเลย) หรือแบบสัมพัทธ์ อย่างไรก็ตาม หลักฐานสำหรับทิศทางดังกล่าวในการวิวัฒนาการนั้นหาได้ยาก และอาจเป็นผลมาจากข้อจำกัดในการพัฒนาที่จำกัดความหลากหลาย[ 47 ]ตัวอย่างเช่น ในหอยทากเปลือกแบบหอยทากมักสร้างเป็นท่อที่เติบโตทั้งความยาวและเส้นผ่านศูนย์กลาง การคัดเลือกได้สร้างรูปทรงเปลือกที่หลากหลาย เช่น เกลียวแบน เปลือกหอยเบี้ยและเกลียวหอคอยสูง ภายใต้ข้อจำกัดเหล่านี้ ในกลุ่มตะขาบ Lithobiomorpha จะมีปล้องลำตัว 15 ปล้องเมื่อโตเต็มวัย ซึ่งอาจเป็นผลมาจากอคติ ในการพัฒนาไปสู่จำนวนปล้องลำตัวที่เป็นเลขคี่ ตะขาบอีกอันดับหนึ่งคือGeophilomorphaจำนวนปล้องแตกต่างกันไปในแต่ละชนิด โดยอยู่ระหว่าง 27 ถึง 191 แต่จำนวนปล้องจะเป็นเลขคี่เสมอ ทำให้เป็นข้อจำกัดที่แน่นอน เกือบทุกจำนวนคี่ในช่วงนั้นถูกครอบครองโดยชนิดใดชนิดหนึ่ง[ 75 ] [ 76 ] [ 77 ]
ชีววิทยาการพัฒนาเชิงวิวัฒนาการเชิงนิเวศวิทยา
ชีววิทยาการพัฒนาเชิงวิวัฒนาการเชิงนิเวศวิทยา หรือที่เรียกกันอย่างไม่เป็นทางการว่า eco-evo-devo เป็นการบูรณาการงานวิจัยจากชีววิทยาการพัฒนาและนิเวศวิทยาเพื่อตรวจสอบความสัมพันธ์กับทฤษฎีวิวัฒนาการ[ 78 ]นักวิจัยศึกษาแนวคิดและกลไกต่างๆ เช่นความยืดหยุ่นในการพัฒนาการถ่ายทอดทางพันธุกรรมแบบเอพิเจเนติกการหลอมรวมทางพันธุกรรม การสร้างนิชและการ อยู่ ร่วมกันแบบพึ่งพาอาศัย กัน [ 79 ] [ 80 ]
ดูเพิ่มเติม
- ปัญหาเกี่ยวกับหัวของสัตว์ขาปล้อง
- การส่งสัญญาณของเซลล์
- วิวัฒนาการและการพัฒนา (วารสาร)
- ชีววิทยาการพัฒนาเชิงวิวัฒนาการของมนุษย์
- นิทานเรื่อง Just So (ตามมุมมองของนักชีววิทยาด้านพัฒนาการเชิงวิวัฒนาการ)
- ชีววิทยาการพัฒนาเชิงวิวัฒนาการของพืช
- ทฤษฎีการสรุป
หมายเหตุ
- ^แม้ว่า CH Waddingtonจะเรียกร้องให้เพิ่มวิชาคัพภวิทยาเข้าไปในการสังเคราะห์ในบทความ "Epigenetics and Evolution" ของเขาในปี พ.ศ. 2496 ก็ตาม [ 26 ]
- ^อคติเชิงบวกบางครั้งเรียกว่าแรงขับในการพัฒนา [ 75 ]
ลิงก์ภายนอก
 สื่อที่เกี่ยวข้องกับชีววิทยาการพัฒนาเชิงวิวัฒนาการในวิกิมีเดียคอมมอนส์
สื่อที่เกี่ยวข้องกับชีววิทยาการพัฒนาเชิงวิวัฒนาการในวิกิมีเดียคอมมอนส์
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ชีววิทยาการพัฒนาเชิงวิวัฒนาการ
ชีววิทยาการพัฒนาเชิงวิวัฒนาการ หรือที่เรียกกันอย่างไม่เป็นทางการว่า อีโว-เดโว (evo-devo ) เป็นสาขา การวิจัยทางชีววิทยา ที่เปรียบเทียบ กระบวนการพัฒนา ของ สิ่งมีชีวิต ต่าง ๆ...
ทฤษฎีในยุคแรก
นักปรัชญาเริ่มคิดเกี่ยวกับวิธีที่สัตว์ได้รับรูปร่างใน ครรภ์ ตั้งแต่สมัย โบราณ อริสโตเติล กล่าวใน ตำรา ฟิสิกส์ ของเขา ว่า ตามที่ เอมเปโดคลีส กล่าวไว้ ความเป็นระเบียบ "เกิดขึ้นเอง" ในตัวอ่อนที่กำลังพัฒนา ใน ตำรา ส่วนต่างๆ ของสัตว์...
สรุป
ทฤษฎี การย้อนรอย วิวัฒนาการถูกเสนอโดย เอเตียน แซร์เรส ในปี 1824–26 ซึ่งสะท้อนแนวคิดของ โยฮันน์ ฟรีดริช เมคเคล ในปี 1808 พวกเขาโต้แย้งว่าตัวอ่อนของสัตว์ 'ชั้นสูง' ผ่านหรือย้อนรอยขั้นตอนต่างๆ ซึ่งแต่ละขั้นตอนจะคล้ายกับสัตว์ที่อยู่ต่ำกว่าใน...
สัณฐานวิทยาเชิงวิวัฒนาการ
ตั้งแต่ต้นศตวรรษที่ 19 จนถึงเกือบตลอดศตวรรษที่ 20 วิชาคัพภวิทยา ต้องเผชิญกับปริศนา สัตว์ต่างๆ ถูกพบว่าพัฒนาไปเป็นตัวเต็มวัยที่มี โครงสร้างร่างกาย แตกต่างกันอย่างมาก โดยมักจะผ่านขั้นตอนที่คล้ายคลึงกันจากไข่...