อ่าน 25 นาที
ชั้นไขมันสองชั้น
ชั้นไขมันสองชั้น (หรือชั้นฟอสโฟลิปิด สองชั้น ) เป็นเยื่อบาง ๆ ที่มีขั้ว ประกอบด้วยโมเลกุลไขมัน สองชั้น เยื่อเหล่านี้สร้างเป็นกำแพงต่อเนื่องรอบเซลล์ ทั้งหมด เยื่อหุ้มเซลล์ ของ...
ชั้นไขมันสองชั้น

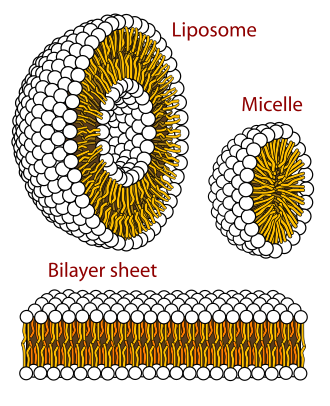
ชั้นไขมันสองชั้น (หรือชั้นฟอสโฟลิปิด สองชั้น ) เป็นเยื่อบาง ๆ ที่มีขั้ว ประกอบด้วยโมเลกุลไขมัน สองชั้น เยื่อเหล่านี้สร้างเป็นกำแพงต่อเนื่องรอบเซลล์ ทั้งหมด เยื่อหุ้มเซลล์ ของ สิ่งมีชีวิตเกือบทั้งหมดและไวรัส หลายชนิด ประกอบด้วยชั้นไขมันสองชั้น เช่นเดียวกับเยื่อหุ้มนิวเคลียสที่ล้อมรอบนิวเคลียสของเซลล์และเยื่อหุ้มของออร์แกเนลล์ที่มีเยื่อหุ้มในเซลล์ ชั้นไขมันสองชั้นเป็นกำแพงที่กักเก็บไอออนโปรตีน และโมเลกุลอื่นๆ ไว้ในตำแหน่งที่จำเป็น และป้องกันไม่ให้แพร่กระจายไปยังบริเวณที่ไม่ควรอยู่ ชั้นไขมันสองชั้นเหมาะอย่างยิ่งสำหรับบทบาทนี้ แม้ว่าจะมีขนาดความกว้าง เพียงไม่กี่ นาโนเมตร[ 2 ]เนื่องจากไม่สามารถซึมผ่านโมเลกุลที่ละลายน้ำได้ส่วนใหญ่ ( ไฮโดรฟิลิก ) ชั้นไขมันสองชั้นไม่สามารถซึมผ่านไอออนได้เป็นพิเศษ ซึ่งช่วยให้เซลล์สามารถควบคุมความเข้มข้นของเกลือและpHโดยการขนส่งไอออนข้ามเยื่อหุ้มเซลล์โดยใช้โปรตีนที่เรียกว่าปั๊มไอออน
โดยทั่วไปแล้วเยื่อชีวภาพสองชั้นจะประกอบด้วยฟอสโฟลิปิดแอมฟิฟิลิก ที่มีหัวฟอสเฟตที่ชอบน้ำและหาง ที่ไม่ชอบ น้ำซึ่งประกอบด้วยกรดไขมัน สอง สาย ฟอสโฟลิปิดที่มีกลุ่มหัวบางอย่างสามารถเปลี่ยนแปลงเคมีพื้นผิวของเยื่อสองชั้นได้ และตัวอย่างเช่น สามารถทำหน้าที่เป็นสัญญาณรวมถึง "จุดยึด" สำหรับโมเลกุลอื่น ๆ ในเยื่อหุ้มเซลล์[ 3 ]เช่นเดียวกับหัว หางของลิปิดก็สามารถส่งผลต่อคุณสมบัติของเยื่อหุ้มเซลล์ได้เช่นกัน ตัวอย่างเช่น โดยการกำหนดเฟสของเยื่อสองชั้น เยื่อสองชั้นสามารถอยู่ในสถานะเจลแข็งที่อุณหภูมิต่ำ แต่จะเกิดการเปลี่ยนเฟสไปเป็นสถานะของเหลวที่อุณหภูมิสูงขึ้น และคุณสมบัติทางเคมีของหางของลิปิดมีอิทธิพลต่ออุณหภูมิที่เกิดการเปลี่ยนแปลงนี้ การจัดเรียงตัวของลิปิดภายในเยื่อสองชั้นยังส่งผลต่อคุณสมบัติทางกล รวมถึงความต้านทานต่อการยืดและการดัดงอ คุณสมบัติเหล่านี้หลายอย่างได้รับการศึกษาโดยใช้เยื่อสองชั้น "จำลอง" เทียมที่ผลิตในห้องปฏิบัติการถุงเวสิเคิลที่สร้างจากเยื่อสองชั้นจำลองยังถูกนำมาใช้ในทางคลินิกเพื่อส่งยาอีกด้วย
โครงสร้างของเยื่อชีวภาพโดยทั่วไปประกอบด้วยโมเลกุลหลายชนิดนอกเหนือจากฟอสโฟลิปิดที่เป็นส่วนประกอบของชั้นไขมันสองชั้น ตัวอย่างที่สำคัญอย่างยิ่งในเซลล์สัตว์คือคอเลสเตอรอลซึ่งช่วยเสริมความแข็งแรงของชั้นไขมันสองชั้นและลดการซึมผ่าน คอเลสเตอรอลยังช่วยควบคุมการทำงานของโปรตีนเยื่อหุ้มเซลล์ บางชนิด โปรตีนเยื่อหุ้มเซลล์จะทำงานเมื่อรวมเข้ากับชั้นไขมันสองชั้น และยึดติดกับชั้นไขมันสองชั้นอย่างแน่นหนาด้วยความช่วยเหลือของเปลือกไขมันรูปวงแหวนเนื่องจากชั้นไขมันสองชั้นกำหนดขอบเขตของเซลล์และส่วนต่างๆ ภายในเซลล์ โปรตีนเยื่อหุ้มเซลล์เหล่านี้จึงมีส่วนเกี่ยวข้องในกระบวนการส่งสัญญาณภายในและระหว่างเซลล์หลายอย่าง โปรตีนเยื่อหุ้มเซลล์บางชนิดมีส่วนเกี่ยวข้องในกระบวนการหลอมรวมชั้นไขมันสองชั้นเข้าด้วยกัน การหลอมรวมนี้ช่วยให้โครงสร้างที่แตกต่างกันสองโครงสร้างมารวมกันได้ เช่น ในปฏิกิริยาอะโครโซมระหว่างการปฏิสนธิของไข่กับอสุจิหรือการเข้าสู่เซลล์ของไวรัสเนื่องจากชั้นไขมันสองชั้นมีความเปราะบางและมองไม่เห็นด้วยกล้องจุลทรรศน์แบบดั้งเดิม จึงยากต่อการศึกษา การทดลองเกี่ยวกับชั้นไขมันสองชั้นมักต้องใช้เทคนิคขั้นสูง เช่นกล้องจุลทรรศน์อิเล็กตรอนและกล้องจุลทรรศน์ แรงอะตอม
โครงสร้างและองค์กร
เมื่อฟอสโฟลิปิดสัมผัสกับน้ำ พวกมันจะประกอบตัวเองเป็นแผ่นสองชั้นโดยมีหางที่ไม่ชอบน้ำชี้ไปทางตรงกลางของแผ่น การจัดเรียงนี้ส่งผลให้เกิด 'แผ่น' สองแผ่นซึ่งแต่ละแผ่นเป็นชั้นโมเลกุลเดียว ตรงกลางของชั้นสองชั้นนี้แทบไม่มีน้ำและกีดกันโมเลกุลเช่นน้ำตาลหรือเกลือที่ละลายในน้ำ กระบวนการประกอบและการบำรุงรักษาถูกขับเคลื่อนโดยการรวมตัวของโมเลกุลที่ไม่ชอบน้ำ (เรียกอีกอย่างว่าผลกระทบที่ไม่ชอบน้ำ ) กระบวนการที่ซับซ้อนนี้รวมถึงปฏิสัมพันธ์ที่ไม่ใช่พันธะโควา เลนต์ เช่นแรงแวนเดอร์วาลส์แรงไฟฟ้าสถิตและพันธะไฮโดรเจน[ 4 ]
การวิเคราะห์ภาคตัดขวาง

ชั้นไขมันสองชั้นนั้นบางมากเมื่อเทียบกับขนาดด้านข้าง หากเซลล์สัตว์เลี้ยงลูกด้วยนมทั่วไป (เส้นผ่านศูนย์กลางประมาณ 10 ไมโครเมตร) ถูกขยายให้มีขนาดเท่าแตงโม (ประมาณ 1 ฟุต/30 เซนติเมตร) ชั้นไขมันสองชั้นที่ประกอบเป็นเยื่อหุ้มพลาสมาจะมีความหนาประมาณกระดาษสำนักงานแผ่นหนึ่ง แม้ว่าจะมีความหนาเพียงไม่กี่นาโนเมตร แต่ชั้นไขมันสองชั้นนี้ประกอบด้วยบริเวณทางเคมีที่แตกต่างกันหลายแห่งตามหน้าตัด บริเวณเหล่านี้และการโต้ตอบกับน้ำโดยรอบได้รับการศึกษาลักษณะเฉพาะในช่วงหลายทศวรรษที่ผ่านมาด้วยเทคนิคการสะท้อนรังสีเอกซ์[6] การกระเจิงของนิวตรอน [ 7 ] และ เทคนิคการเรโซแนนซ์แม่เหล็กนิวเคลียร์[ 8 ]
บริเวณแรกที่อยู่ด้านใดด้านหนึ่งของไบเลเยอร์คือกลุ่มหัวไฮโดรฟิลิก ส่วนนี้ของเยื่อหุ้มเซลล์จะถูกไฮเดรตอย่างสมบูรณ์และโดยทั่วไปมีความหนาประมาณ 0.8-0.9 นาโนเมตร ในไบเลเยอร์ฟอสโฟลิ ปิด กลุ่ม ฟอสเฟตจะอยู่ภายในบริเวณที่ถูกไฮเดรตนี้ โดยอยู่ห่างจากแกนไฮโดรโฟบิกประมาณ 0.5 นาโนเมตร[ 9 ]ในบางกรณี บริเวณที่ถูกไฮเดรตอาจขยายออกไปไกลกว่านั้นมาก เช่น ในลิปิดที่มีโปรตีนขนาดใหญ่หรือโซ่น้ำตาลยาวที่เชื่อมต่อกับหัว ตัวอย่างทั่วไปของการดัดแปลงดังกล่าวในธรรมชาติคือ ชั้น ลิโปโพลีแซคคาไรด์บนเยื่อหุ้มเซลล์ชั้นนอกของแบคทีเรีย[ 10 ]
ถัดจากบริเวณหัวกลุ่มที่ชอบน้ำคือบริเวณกลางซึ่งมีน้ำอยู่เพียงบางส่วน ชั้นขอบเขตนี้มีความหนาประมาณ 0.3 นาโนเมตร ภายในระยะทางสั้นๆ นี้ ความเข้มข้นของน้ำจะลดลงจาก 2M ที่ด้านหัวกลุ่มไปเกือบเป็นศูนย์ที่ด้านหาง (แกนกลาง) [ 11 ] [ 12 ]แกนกลางที่ไม่ชอบน้ำของไบเลเยอร์โดยทั่วไปมีความหนา 3-4 นาโนเมตร แต่ค่านี้จะแตกต่างกันไปตามความยาวของโซ่และองค์ประกอบทางเคมี[ 6 ] [ 13 ]ความหนาของแกนกลางยังแตกต่างกันอย่างมากตามอุณหภูมิ โดยเฉพาะอย่างยิ่งใกล้กับการเปลี่ยนเฟส[ 14 ]
ความไม่สมมาตร
ในเยื่อหุ้มเซลล์แบบสองชั้นที่เกิดขึ้นตามธรรมชาติหลายชนิด องค์ประกอบของเยื่อหุ้มเซลล์ชั้นในและชั้นนอกจะแตกต่างกัน ในเซลล์เม็ดเลือดแดง ของมนุษย์ เยื่อหุ้มเซลล์ชั้นใน (ไซโตพลาสมิก) ส่วนใหญ่ประกอบด้วยฟอสฟาติ ดิ ลเอ ทานอลามี น ฟอสฟาติดิลเซอรีน และฟอสฟา ติดิลอิโน ซิทอลและอนุพันธ์ฟอสฟอริเลตในทางตรงกันข้าม เยื่อหุ้มเซลล์ชั้นนอก (นอกเซลล์) ประกอบด้วยฟอสฟาติดิล โคลีน สฟิงโกไมอีลิน และไกลโคลิปิดหลายชนิด [ 15 ] [ 16 ] [ 17 ]ในบางกรณี ความไม่สมมาตรนี้ขึ้นอยู่กับตำแหน่งที่สร้างลิปิดในเซลล์และสะท้อนถึงการวางตัวเริ่มต้นของ ลิปิด [ 18 ]หน้าที่ทางชีวภาพของความไม่สมมาตรของลิปิดยังไม่เป็นที่เข้าใจอย่างสมบูรณ์ แม้ว่าจะชัดเจนว่ามีการนำไปใช้ในสถานการณ์ต่างๆ หลายอย่าง ตัวอย่างเช่น เมื่อเซลล์เกิดอะพอพโทซิส ฟอสฟาติดิลเซอรีน ซึ่งปกติจะอยู่ที่ชั้นไซโตพลาสมิก จะถูกถ่ายโอนไปยังพื้นผิวด้านนอก ที่นั่นแมโครฟาจ จะจดจำมันได้ จากนั้น จึงกำจัดเซลล์ที่กำลังจะตายอย่างแข็งขัน[ 19 ]
ความไม่สมมาตรของลิปิดเกิดขึ้นอย่างน้อยบางส่วนจากข้อเท็จจริงที่ว่าฟอสโฟลิปิดส่วนใหญ่ถูกสังเคราะห์และแทรกเข้าไปในชั้นโมโนเลเยอร์ด้านในในตอนแรก ส่วนที่ประกอบเป็นชั้นโมโนเลเยอร์ด้านนอกนั้นจะถูกขนส่งจากชั้นโมโนเลเยอร์ด้านในโดยเอนไซม์กลุ่มหนึ่งที่เรียกว่าฟลิปเพส [ 20 ] [ 21 ] ลิปิดอื่นๆ เช่น สฟิงโกไมอีลิน ดูเหมือนจะถูกสังเคราะห์ที่ชั้นนอก ฟลิปเพสเป็นสมาชิกของกลุ่มโมเลกุลขนส่งลิปิดขนาดใหญ่ ซึ่งรวมถึงฟลอปเพส ซึ่งถ่ายโอนลิปิดในทิศทางตรงกันข้าม และสแครมเบลส ซึ่งทำให้การกระจายตัวของลิปิดในไบเลเยอร์เป็นแบบสุ่ม (เช่นในเซลล์อะพอพโทซิส) ไม่ว่าในกรณีใด เมื่อความไม่สมมาตรของลิปิดเกิดขึ้นแล้ว โดยปกติจะไม่หายไปอย่างรวดเร็วเนื่องจากการพลิกกลับของลิปิดระหว่างชั้นเกิดขึ้นเองช้ามาก[ 22 ]เมื่อเร็วๆ นี้ได้มีการแสดงให้เห็นว่าคอเลสเตอรอลดูเหมือนจะมีบทบาทสำคัญในการสร้างและรักษาความไม่สมมาตรของไบเลเยอร์ด้วย[ 23 ]
เป็นไปได้ที่จะจำลองความไม่สมมาตรนี้ในห้องปฏิบัติการในระบบไบเลเยอร์แบบจำลองเวสิเคิล เทียมขนาดเล็กบางชนิด จะทำให้ตัวเองไม่สมมาตรเล็กน้อยโดยอัตโนมัติ แม้ว่ากลไกที่ทำให้เกิดความไม่สมมาตรนี้จะแตกต่างจากในเซลล์มากก็ตาม[ 24 ]โดยการใช้โมโนเลเยอร์ที่แตกต่างกันสองชั้นในการตกตะกอนแบบLangmuir–Blodgett [ 25 ]หรือการรวมกันของการตกตะกอนแบบ Langmuir–Blodgett และการแตกของเวสิเคิล[ 26 ]ก็สามารถสังเคราะห์ไบเลเยอร์ระนาบที่ไม่สมมาตรได้เช่นกัน ความไม่สมมาตรนี้อาจหายไปเมื่อเวลาผ่านไป เนื่องจากลิปิดในไบเลเยอร์ที่รองรับอาจมีแนวโน้มที่จะพลิกกลับ[ 27 ]อย่างไรก็ตาม มีรายงานว่าการพลิกกลับของลิปิดนั้นช้ากว่าคอเลสเตอรอลและโมเลกุลขนาดเล็กอื่นๆ[ 28 ] [ 29 ]
มีรายงานว่าการจัดระเบียบและพลวัตของชั้นไขมันโมโนเลเยอร์ในไบเลเยอร์นั้นเชื่อมโยงกัน[ 30 ] [ 31 ]ตัวอย่างเช่น การนำสิ่งกีดขวางเข้ามาในชั้นโมโนเลเยอร์หนึ่งสามารถทำให้การแพร่กระจายด้านข้างในทั้งสองชั้นโมโนเลเยอร์ช้าลงได้[ 30 ]นอกจากนี้ การแยกเฟสในชั้นโมโนเลเยอร์หนึ่งยังสามารถชักนำให้เกิดการแยกเฟสในชั้นโมโนเลเยอร์อื่นได้ แม้ว่าชั้นโมโนเลเยอร์อื่นจะไม่สามารถแยกเฟสได้ด้วยตัวเองก็ตาม[ 31 ]
เฟสและการเปลี่ยนเฟส

ที่อุณหภูมิที่กำหนด ชั้นไขมันสองชั้นสามารถอยู่ในสถานะของเหลวหรือสถานะเจล (ของแข็ง) ก็ได้ ไขมันทุกชนิดมีอุณหภูมิเฉพาะที่พวกมันจะเปลี่ยนสถานะ (หลอมเหลว) จากสถานะเจลไปเป็นสถานะของเหลว ในทั้งสองสถานะ โมเลกุลของไขมันจะไม่สามารถพลิกกลับไปมาบนชั้นไขมันสองชั้นได้ แต่ในชั้นไขมันสองชั้นในสถานะของเหลว ไขมันแต่ละโมเลกุลจะแลกเปลี่ยนตำแหน่งกับโมเลกุลข้างเคียงหลายล้านครั้งต่อวินาที การแลกเปลี่ยน แบบสุ่ม นี้ ทำให้ไขมันสามารถแพร่กระจายและเคลื่อนที่ไปทั่วพื้นผิวของเยื่อหุ้มเซลล์ได้ ซึ่งแตกต่างจากชั้นไขมันสองชั้นในสถานะของเหลว ไขมันในชั้นไขมันสองชั้นในสถานะเจลมีความคล่องตัวน้อยกว่า[ 32 ]
พฤติกรรมเฟสของชั้นไขมันสองชั้นถูกกำหนดโดยส่วนใหญ่จากความแข็งแรงของแรงดึงดูดแบบแวนเดอร์วาลส์ระหว่างโมเลกุลไขมันที่อยู่ติดกัน ไขมันที่มีหางยาวกว่าจะมีพื้นที่ในการโต้ตอบมากกว่า ทำให้ความแข็งแรงของแรงโต้ตอบนี้เพิ่มขึ้น และส่งผลให้การเคลื่อนที่ของไขมันลดลง ดังนั้น ที่อุณหภูมิที่กำหนด ไขมันที่มีหางสั้นจะมีความเหลวมากกว่าไขมันที่มีหางยาวที่เหมือนกันทุกประการ[ 13 ]อุณหภูมิการเปลี่ยนผ่านอาจได้รับผลกระทบจากระดับความไม่อิ่มตัวของหางไขมันพันธะคู่ ที่ไม่อิ่มตัว สามารถทำให้เกิดการหักงอใน โซ่ แอลเคนทำให้การจัดเรียงตัวของไขมันหยุดชะงัก การหยุดชะงักนี้สร้างพื้นที่ว่างเพิ่มเติมภายในชั้นไขมันสองชั้น ซึ่งช่วยให้โซ่ที่อยู่ติดกันมีความยืดหยุ่นมากขึ้น[ 13 ]ตัวอย่างของผลกระทบนี้สามารถสังเกตได้ในชีวิตประจำวัน เช่น เนยซึ่งมีไขมันอิ่มตัวในปริมาณมากจะเป็นของแข็งที่อุณหภูมิห้อง ในขณะที่น้ำมันพืชซึ่งส่วนใหญ่ไม่อิ่มตัวจะเป็นของเหลว[ 33 ]
เยื่อหุ้มเซลล์ตามธรรมชาติส่วนใหญ่เป็นส่วนผสมที่ซับซ้อนของโมเลกุลไขมันที่แตกต่างกัน หากส่วนประกอบบางส่วนเป็นของเหลวที่อุณหภูมิที่กำหนด ในขณะที่ส่วนประกอบอื่นๆ อยู่ในเฟสเจล เฟสทั้งสองสามารถอยู่ร่วมกันได้ในบริเวณที่แยกจากกันในเชิงพื้นที่ คล้ายกับภูเขาน้ำแข็งที่ลอยอยู่ในมหาสมุทร การแยกเฟสนี้มีบทบาทสำคัญในปรากฏการณ์ทางชีวเคมี เนื่องจากส่วนประกอบของเยื่อหุ้มเซลล์ เช่น โปรตีน สามารถแบ่งตัวเข้าไปอยู่ในเฟสใดเฟสหนึ่ง และทำให้เกิดความเข้มข้นหรือการทำงานเฉพาะที่[ 34 ]ส่วนประกอบที่สำคัญอย่างยิ่งของระบบเฟสผสมหลายระบบคือคอเลสเตอรอลซึ่งปรับเปลี่ยนการซึมผ่านของเยื่อหุ้มเซลล์ ความแข็งแรงเชิงกล และปฏิกิริยาทางชีวเคมี[ 35 ]
เคมีพื้นผิว
ในขณะที่หางของลิปิดทำหน้าที่ปรับเปลี่ยนพฤติกรรมเฟสของไบเลเยอร์เป็นหลัก แต่ส่วนหัวของไบเลเยอร์เป็นตัวกำหนดเคมีพื้นผิวของไบเลเยอร์ ไบเลเยอร์ตามธรรมชาติส่วนใหญ่ประกอบด้วยฟอสโฟ ลิปิดเป็นหลัก แต่สฟิงโกลิปิดและสเตอรอลเช่นคอเลสเตอรอลก็เป็นส่วนประกอบที่สำคัญเช่นกัน[ 36 ]ในบรรดาฟอสโฟลิปิด ส่วนหัวที่พบได้บ่อยที่สุดคือฟอสฟาติดิลโคลีน (PC) ซึ่งคิดเป็นประมาณครึ่งหนึ่งของฟอสโฟลิปิดในเซลล์ของสัตว์เลี้ยงลูกด้วยนมส่วนใหญ่[ 37 ] PC เป็น ส่วนหัวแบบ ซวิตเตอร์ไอออนิกเนื่องจากมีประจุลบที่หมู่ฟอสเฟตและประจุบวกที่หมู่อะมีน แต่เนื่องจากประจุเฉพาะที่เหล่านี้สมดุลกัน จึงไม่มีประจุสุทธิ[ 38 ]
นอกจากนี้ยังมีกลุ่มหัวอื่นๆ อยู่ด้วยในระดับที่แตกต่างกัน และอาจรวมถึงฟอสฟาติดิลเซอรีน (PS) ฟอ สฟาติดิ ลเอทานอลามีน (PE) และฟอสฟาติดิลกลีเซอรอล( PG) กลุ่มหัวทางเลือกเหล่านี้มักให้ฟังก์ชันทางชีวภาพเฉพาะที่ขึ้นอยู่กับบริบทอย่างมาก ตัวอย่างเช่น การมีอยู่ของ PS บนด้านเยื่อหุ้มเซลล์ภายนอกของเม็ดเลือดแดงเป็นเครื่องหมายของการตาย ของเซลล์ [ 19 ]ในขณะที่ PS ใน เวสิเคิล ของแผ่นการเจริญเติบโตมีความจำเป็นสำหรับการก่อตัวของ ผลึกไฮดรอก ซีอะพาไทต์และการสร้างแร่ธาตุในกระดูกในภายหลัง[ 39 ] [ 40 ]แตกต่างจาก PC กลุ่มหัวอื่นๆ บางกลุ่มมีประจุสุทธิ ซึ่งสามารถเปลี่ยนแปลงปฏิสัมพันธ์ทางไฟฟ้าสถิตของโมเลกุลขนาดเล็กกับไบเลเยอร์ได้[ 41 ]
บทบาททางชีววิทยา
การกักกันและการแยก
บทบาทหลักของชั้นไขมันสองชั้นในทางชีววิทยาคือการแยก ช่อง น้ำออกจากสิ่งแวดล้อม หากไม่มีสิ่งกีดขวางที่แบ่งแยก "ตัวเอง" ออกจาก "ไม่ใช่ตัวเอง" ก็ยากที่จะนิยามแนวคิดของสิ่งมีชีวิตหรือชีวิตได้ สิ่งกีดขวางนี้มีรูปแบบเป็นชั้นไขมันสองชั้นในสิ่งมีชีวิตที่รู้จักทั้งหมด ยกเว้นอาร์เคีย บางชนิด ที่ใช้ชั้นไขมันชั้นเดียวที่ปรับตัวเป็นพิเศษ[ 42 ]มีการเสนอด้วยซ้ำว่าสิ่งมีชีวิตรูปแบบแรกสุดอาจเป็นถุงไขมัน อย่างง่ายที่มีความสามารถ ในการสังเคราะห์ทางชีวภาพเพียงอย่างเดียวคือการผลิตฟอสโฟลิปิด เพิ่มขึ้น [ 43 ]ความสามารถในการแบ่งส่วนของชั้นไขมันสองชั้นนั้นขึ้นอยู่กับข้อเท็จจริงที่ว่า โมเลกุล ที่ชอบน้ำ ไม่สามารถผ่านแกนกลางของชั้นไขมันสองชั้น ที่ไม่ชอบน้ำได้ง่ายดังที่ได้กล่าวไว้ในการขนส่งผ่านชั้นไขมันสองชั้นด้านล่าง นิวเคลียส ไมโตคอนเดรีย และคลอโรพลาสต์มีเยื่อไขมันสองชั้น ในขณะที่โครงสร้างย่อยของเซลล์อื่นๆ ถูกล้อมรอบด้วยเยื่อไขมันชั้นเดียว (เช่น เยื่อหุ้มเซลล์ เอนโดพลาสมิกเรติคูลัม กอลจิแอพพาราตัสและไลโซโซม) ดูออร์แกเนลล์[ 44 ]
โปรคาริโอตมีชั้นไขมันสองชั้นเพียงชั้นเดียว คือเยื่อหุ้มเซลล์ (หรือที่เรียกว่าเยื่อพลาสมา) โปรคาริโอตหลายชนิดยังมีผนังเซลล์แต่ผนังเซลล์ประกอบด้วยโปรตีน หรือ คาร์โบไฮเดรตสายยาวไม่ใช่ไขมัน ในทางตรงกันข้ามยูคาริโอต มี ออร์แกเนลล์หลายชนิดได้แก่นิวเคลียส ไม โตคอนเดรี ย ไลโซโซมและเอนโดพลาสมิกเรติคูลัมส่วนประกอบย่อยของเซลล์เหล่านี้ทั้งหมดถูกล้อมรอบด้วยชั้นไขมันสองชั้นหนึ่งชั้นหรือมากกว่า และโดยทั่วไปแล้วจะประกอบกันเป็นพื้นที่ส่วนใหญ่ของชั้นไขมันสองชั้นที่มีอยู่ในเซลล์ ตัวอย่างเช่น ในเซลล์ตับเยื่อพลาสมาคิดเป็นเพียงร้อยละสองของพื้นที่ชั้นไขมันสองชั้นทั้งหมดของเซลล์ ในขณะที่เอนโดพลาสมิกเรติคูลัมมีมากกว่าร้อยละห้าสิบ และไมโตคอนเดรียมีอีกร้อยละสามสิบ[ 45 ]

การส่งสัญญาณ
รูปแบบการส่งสัญญาณระดับเซลล์ที่คุ้นเคยมากที่สุดน่าจะเป็นการส่งสัญญาณผ่านไซแนปส์ซึ่งกระแสประสาทที่ไปถึงปลายของเซลล์ประสาท หนึ่ง จะถูกส่งไปยังเซลล์ประสาทข้างเคียงผ่านการปล่อยสารสื่อประสาทการส่งสัญญาณนี้เกิดขึ้นได้ด้วยการทำงานของถุงไซแนปส์ซึ่งบรรจุสารสื่อประสาทที่จะถูกปล่อยออกมาในภายหลังภายในเซลล์ ถุงที่บรรจุสารสื่อประสาทเหล่านี้จะรวมตัวกับเยื่อหุ้มเซลล์ที่ปลายก่อนไซแนปส์และสารภายในจะถูกปล่อยออกมาสู่พื้นที่ภายนอกเซลล์ จากนั้นสารภายในจะแพร่กระจายข้ามไซแนปส์ไปยังปลายหลังไซแนปส์[ 46 ]
ชั้นไขมันสองชั้นยังเกี่ยวข้องกับการส่งสัญญาณผ่านบทบาทของมันในฐานะที่อยู่ของโปรตีนเมมเบรนแบบบูร ณา การ นี่เป็นกลุ่มของโมเลกุลชีวภาพ ที่กว้างขวางและสำคัญอย่างยิ่ง มีการประมาณการว่า โปรตีนเมมเบรนคิด เป็นสัดส่วน ถึงหนึ่งในสามของโปรตีโอม ของมนุษย์ [ 47 ]โปรตีนเหล่านี้บางส่วนเชื่อมโยงกับด้านนอกของเยื่อหุ้มเซลล์ ตัวอย่างเช่น โปรตีน CD59ซึ่งระบุเซลล์ว่าเป็น "เซลล์ของตัวเอง" และยับยั้งการทำลายโดยระบบภูมิคุ้มกัน ไวรัส HIVหลบเลี่ยงระบบภูมิคุ้มกันได้บางส่วนโดยการปลูกถ่ายโปรตีนเหล่านี้จากเยื่อหุ้มเซลล์ของโฮสต์ไปยังพื้นผิวของมันเอง[ 45 ]ในทางกลับกัน โปรตีนเมมเบรนบางชนิดแทรกซึมผ่านชั้นไขมันสองชั้นและทำหน้าที่ส่งต่อเหตุการณ์สัญญาณแต่ละเหตุการณ์จากภายนอกไปยังภายในเซลล์ กลุ่มโปรตีนประเภทนี้ที่พบได้บ่อยที่สุดคือตัวรับที่เชื่อมต่อกับโปรตีน G (GPCR) GPCR มีหน้าที่รับผิดชอบความสามารถของเซลล์ในการรับรู้สภาพแวดล้อมโดยรอบ และเนื่องจากบทบาทที่สำคัญนี้ ยาที่ใช้ในปัจจุบันประมาณ 40% จึงมุ่งเป้าไปที่ GPCR [ 48 ]
นอกเหนือจากกระบวนการที่อาศัยโปรตีนและสารละลายแล้ว เยื่อไขมันสองชั้นยังสามารถมีส่วนร่วมโดยตรงในการส่งสัญญาณได้อีกด้วย ตัวอย่างคลาสสิกของเรื่องนี้คือฟาโกไซโท ซิส ที่ถูกกระตุ้นโดย ฟอสฟาติดิลเซอรีน โดยปกติแล้ว ฟอสฟาติดิลเซอรีนจะกระจายตัวอย่างไม่สมมาตรในเยื่อหุ้มเซลล์และมีอยู่เฉพาะด้านในเท่านั้น ในระหว่างการตายของเซลล์ตามโปรแกรม โปรตีนที่เรียกว่าscramblaseจะปรับสมดุลการกระจายตัวนี้ โดยแสดงฟอสฟาติดิลเซอรีนบนด้านเยื่อไขมันสองชั้นภายนอกเซลล์ การมีอยู่ของฟอสฟาติดิลเซอรีนจะกระตุ้นฟาโกไซโทซิสเพื่อกำจัดเซลล์ที่ตายแล้วหรือกำลังจะตาย[ 49 ]
วิธีการกำหนดลักษณะเฉพาะ

ชั้นไขมันสองชั้นเป็นโครงสร้างที่ศึกษาได้ยากเนื่องจากบางและเปราะบางมาก[ 50 ]เพื่อเอาชนะข้อจำกัดเหล่านี้ จึงได้มีการพัฒนาเทคนิคต่างๆ เพื่อให้สามารถศึกษาโครงสร้างและหน้าที่ของมันได้[ 51 ] [ 50 ] [ 52 ]
การวัดทางไฟฟ้า
การวัดทางไฟฟ้าเป็นวิธีที่ตรงไปตรงมาในการกำหนดลักษณะการทำงานที่สำคัญของไบเลเยอร์ นั่นคือ ความสามารถในการแยกและป้องกันการไหลของไอออนในสารละลาย โดยการใช้แรงดันไฟฟ้าคร่อมไบเลเยอร์และวัดกระแสไฟฟ้าที่เกิดขึ้น จะสามารถกำหนด ความต้านทานของไบเลเยอร์ได้ ความต้านทานนี้โดยทั่วไปจะค่อนข้างสูง (10⁸ โอห์ม -ซม. ²หรือมากกว่า) [ 51 ]เนื่องจากแกนไฮโดรโฟบิกไม่สามารถซึมผ่านชนิดที่มีประจุได้ การมีรูขนาดเพียงไม่กี่นาโนเมตรก็ส่งผลให้กระแสไฟฟ้าเพิ่มขึ้นอย่างมาก[ 53 ] ความไวของระบบนี้สูงมากจน สามารถแยกแยะกิจกรรมของช่องไอออน เดี่ยวได้ [ 54 ]
กล้องจุลทรรศน์ฟลูออเรสเซนซ์

ชั้นไขมันสองชั้นไม่สามารถมองเห็นได้ด้วยกล้องจุลทรรศน์แบบดั้งเดิมเนื่องจากบางเกินไป ดังนั้นนักวิจัยจึงมักใช้กล้องจุลทรรศน์ฟลูออเรสเซนต์โดยจะกระตุ้นตัวอย่างด้วยแสงความยาวคลื่นหนึ่งและสังเกตด้วยแสงความยาวคลื่นอื่น เพื่อให้เห็นเฉพาะโมเลกุลเรืองแสงที่มีโปรไฟล์การกระตุ้นและการปล่อยแสงที่ตรงกันเท่านั้น ชั้นไขมันสองชั้นตามธรรมชาติไม่เรืองแสง ดังนั้นจึงจำเป็นต้องมีสีย้อมเรืองแสงอย่างน้อยหนึ่งชนิดติดอยู่กับโมเลกุลบางส่วนในชั้นไขมันสองชั้น ความละเอียดมักจะจำกัดอยู่ที่ไม่กี่ร้อยนาโนเมตร ซึ่งน่าเสียดายที่ใหญ่กว่าความหนาของชั้นไขมันสองชั้นมาก[ 50 ]
กล้องจุลทรรศน์อิเล็กตรอน
กล้องจุลทรรศน์อิเล็กตรอนให้ภาพที่มีความละเอียดสูงกว่า ในกล้องจุลทรรศน์อิเล็กตรอนลำแสงอิเล็กตรอน ที่โฟกัส จะทำปฏิกิริยากับตัวอย่าง แทนที่จะเป็นลำแสงเหมือนในกล้องจุลทรรศน์แบบดั้งเดิม เมื่อใช้ร่วมกับเทคนิคการแช่แข็งอย่างรวดเร็ว กล้องจุลทรรศน์อิเล็กตรอนยังถูกนำมาใช้เพื่อศึกษาถึงกลไกการขนส่งระหว่างเซลล์และภายในเซลล์ ตัวอย่างเช่น ในการแสดงให้เห็นว่า เวสิเคิล เอ็กโซไซโทซิสเป็นวิธีการปล่อยสารเคมีที่ไซแนปส์[ 55 ]
สเปกโทรสโกปีเรโซแนนซ์แม่เหล็กนิวเคลียร์
สเปกโทรสโกปีนิวเคลียร์แมกเนติกเรโซแนนซ์ 31P ถูกนำมาใช้กันอย่างแพร่หลายในการศึกษาชั้นไขมันฟอสโฟลิปิดและเยื่อชีวภาพในสภาวะธรรมชาติ การวิเคราะห์ สเปกตรัม 31P -NMR ของลิปิดสามารถให้ข้อมูลที่หลากหลายเกี่ยวกับการจัดเรียงตัวของชั้นไขมัน การเปลี่ยนเฟส (เฟสเจล เฟสผลึกเหลวทางสรีรวิทยา เฟสระลอกคลื่น เฟสที่ไม่ใช่ชั้นไขมัน) การวางแนว/พลวัตของกลุ่มหัวลิปิด และคุณสมบัติความยืดหยุ่นของชั้นไขมันบริสุทธิ์และเป็นผลมาจากการจับตัวของโปรตีนและโมเลกุลชีวภาพอื่นๆ[ 52 ]
กล้องจุลทรรศน์แรงอะตอม


วิธีการใหม่ในการศึกษาชั้นไขมันสองชั้นคือกล้องจุลทรรศน์แรงอะตอม (AFM) แทนที่จะใช้ลำแสงหรืออนุภาค ปลายแหลมขนาดเล็กมากจะสแกนพื้นผิวโดยการสัมผัสทางกายภาพกับชั้นไขมันสองชั้นและเคลื่อนที่ไปมาเหมือนเข็มเครื่องเล่นแผ่นเสียง AFM เป็นเทคนิคที่น่าสนใจเพราะมีศักยภาพในการสร้างภาพด้วยความละเอียดระดับนาโนเมตรที่อุณหภูมิห้องและแม้กระทั่งใต้น้ำหรือบัฟเฟอร์ทางสรีรวิทยา ซึ่งเป็นสภาวะที่จำเป็นสำหรับพฤติกรรมของชั้นไขมันสองชั้นตามธรรมชาติ การใช้ความสามารถนี้ AFM ได้ถูกนำมาใช้เพื่อตรวจสอบพฤติกรรมของชั้นไขมันสองชั้นแบบไดนามิก รวมถึงการก่อตัวของรูพรุนข้ามเยื่อหุ้มเซลล์ (รู) [ 56 ]และการเปลี่ยนเฟสในชั้นไขมันสองชั้นที่รองรับ[ 57 ]ข้อดีอีกประการหนึ่งคือ AFM ไม่จำเป็นต้องติดฉลากเรืองแสงหรือไอโซโทปของไขมัน เนื่องจากปลายโพรบมีปฏิสัมพันธ์เชิงกลกับพื้นผิวของชั้นไขมันสองชั้น ด้วยเหตุนี้ การสแกนเดียวกันจึงสามารถสร้างภาพทั้งไขมันและโปรตีนที่เกี่ยวข้องได้ บางครั้งอาจมีความละเอียดระดับโมเลกุลเดี่ยวด้วยซ้ำ[ 56 ] [ 58 ] AFM ยังสามารถตรวจสอบลักษณะเชิงกลของชั้นไขมันสองชั้นได้อีกด้วย[ 59 ]
อินเตอร์เฟอโรเมตรีแบบโพลาไรเซชันคู่
ชั้นไขมันสองชั้นแสดงการหักเหของแสง ในระดับสูง โดยดัชนีหักเหในระนาบของชั้นไขมันสองชั้นจะแตกต่างจากดัชนีหักเหในแนวตั้งฉากมากถึง 0.1 หน่วย ดัชนีหักเหสิ่งนี้ถูกนำมาใช้เพื่อกำหนดลักษณะระดับความเป็นระเบียบและการหยุดชะงักในชั้นไขมันสองชั้นโดยใช้การแทรกสอดแบบโพลาไรซ์คู่เพื่อทำความเข้าใจกลไกการโต้ตอบของโปรตีน[ 60 ]
การคำนวณทางเคมีควอนตัม
ชั้นไขมันสองชั้นเป็นระบบโมเลกุลที่ซับซ้อนซึ่งมีระดับความเป็นอิสระหลายระดับ ดังนั้นการจำลองอะตอมของเยื่อหุ้มเซลล์และโดยเฉพาะอย่างยิ่ง การคำนวณคุณสมบัติของเยื่อหุ้มเซลล์ แบบ ab initioจึงทำได้ยากและต้องใช้การคำนวณสูง การคำนวณทางเคมีควอนตัมเพิ่งประสบความสำเร็จในการประมาณ ค่า โมเมนต์ไดโพลและควอดรูโพลของเยื่อหุ้มไขมัน[ 61 ]
การขนส่งผ่านชั้นสองชั้น
การแพร่แบบพาสซีฟ
โมเลกุล ขั้วส่วนใหญ่มีความละลายต่ำใน แกน ไฮโดรคาร์บอนของชั้นไขมันสองชั้น และด้วยเหตุนี้จึงมีค่าสัมประสิทธิ์การซึมผ่านต่ำผ่านชั้นไขมันสองชั้น ผลกระทบนี้เด่นชัดเป็นพิเศษสำหรับชนิดที่มีประจุ ซึ่งมีค่าสัมประสิทธิ์การซึมผ่านต่ำกว่าโมเลกุลขั้วที่เป็นกลางเสียอีก[ 62 ] โดยทั่วไปแล้ว แอนไอออนจะมีอัตราการแพร่ผ่านชั้นไขมันสองชั้นสูงกว่า แค ตไอออน[ 63 ] [ 64 ]เมื่อเทียบกับไอออน โมเลกุลของน้ำมีการซึมผ่านชั้นไขมันสองชั้นค่อนข้างมาก ดังที่เห็นได้จากการบวมตัวแบบออสโมซิสเมื่อเซลล์หรือถุงที่มีความเข้มข้นของเกลือภายในสูงถูกวางไว้ในสารละลายที่มีความเข้มข้นของเกลือต่ำ มันจะบวมและแตกในที่สุด ผลลัพธ์ดังกล่าวจะไม่เกิดขึ้นเว้นแต่ว่าน้ำจะสามารถผ่านชั้นไขมันสองชั้นได้ค่อนข้างง่าย การซึมผ่านของน้ำที่สูงผิดปกติผ่านชั้นไขมันสองชั้นยังไม่เป็นที่เข้าใจอย่างสมบูรณ์และยังคงเป็นหัวข้อของการถกเถียงอย่างต่อเนื่อง[ 65 ]โมเลกุลขนาดเล็กที่ไม่มีประจุและไม่มีขั้วจะแพร่กระจายผ่านเยื่อไขมันได้เร็วกว่าไอออนหรือน้ำหลายเท่าตัว ซึ่งใช้ได้ทั้งกับไขมันและตัวทำละลายอินทรีย์ เช่นคลอโรฟอร์มและอีเทอร์โดยไม่คำนึงถึงลักษณะขั้ว โมเลกุลขนาดใหญ่จะแพร่กระจายผ่านเยื่อไขมันได้ช้ากว่าโมเลกุลขนาดเล็ก[ 66 ]

ปั๊มไอออนและช่องทางไอออน
โปรตีนสองกลุ่มพิเศษที่เกี่ยวข้องกับความชันของไอออนที่พบในเยื่อหุ้มเซลล์และเยื่อหุ้มย่อยของเซลล์ในธรรมชาติ ได้แก่ช่องไอออนและปั๊มไอออนทั้งปั๊มและช่องเป็นโปรตีนเมมเบรนแบบฝังตัวที่ผ่านชั้นไบเลเยอร์ แต่บทบาทของพวกมันค่อนข้างแตกต่างกัน ปั๊มไอออนเป็นโปรตีนที่สร้างและรักษาความชันทางเคมีโดยใช้แหล่งพลังงานภายนอกเพื่อเคลื่อนย้ายไอออนสวนทางกับความชันของความเข้มข้นไปยังบริเวณที่มีศักยภาพทางเคมี สูงกว่า แหล่งพลังงานอาจเป็นATPเช่นเดียวกับกรณีของNa + -K + ATPaseหรืออาจเป็นความชันทางเคมีอื่นที่มีอยู่แล้ว เช่นในCa2 + /Na + antiporterเซลล์สามารถควบคุมpHผ่านการปั๊มโปรตอนได้ โดยอาศัยการทำงานของปั๊มไอออน [ 67 ] [ 68 ]
ตรงกันข้ามกับปั๊มไอออน ช่องไอออนไม่ได้สร้างการไล่ระดับทางเคมี แต่จะสลายการไล่ระดับเหล่านั้นเพื่อทำงานหรือส่งสัญญาณ ตัวอย่างที่คุ้นเคยและได้รับการศึกษามากที่สุดน่าจะเป็นช่อง Na +ที่ควบคุมด้วยแรงดันไฟฟ้าซึ่งช่วยให้การนำไฟฟ้าของศักยภาพการกระทำเกิดขึ้นตามเซลล์ประสาทปั๊มไอออนทั้งหมดมีกลไกการกระตุ้นหรือ "การเปิดปิด" บางอย่าง ในตัวอย่างก่อนหน้านี้คืออคติทางไฟฟ้า แต่ช่องอื่นๆ สามารถถูกกระตุ้นได้โดยการจับกับตัวกระตุ้นโมเลกุลหรือผ่านการเปลี่ยนแปลงโครงสร้างในโปรตีนใกล้เคียงอื่น[ 69 ]

เอนโดไซโทซิสและเอ็กโซไซโทซิส
โมเลกุลหรืออนุภาคบางชนิดมีขนาดใหญ่เกินไปหรือชอบน้ำมากเกินไปที่จะผ่านเยื่อไขมันสองชั้นได้ ส่วนโมเลกุลอื่นๆ อาจผ่านเยื่อไขมันสองชั้นได้ แต่ต้องขนส่งอย่างรวดเร็วในปริมาณมากจนการขนส่งแบบช่องทางทำได้ยาก ในทั้งสองกรณี สารเหล่านี้สามารถเคลื่อนย้ายผ่านเยื่อหุ้มเซลล์ได้โดยการรวมตัวหรือการแตกหน่อของเวสิเคิลเมื่อเวสิเคิลถูกสร้างขึ้นภายในเซลล์และรวมตัวกับเยื่อหุ้มพลาสมาเพื่อปล่อยสารภายในออกสู่ช่องว่างภายนอกเซลล์ กระบวนการนี้เรียกว่าเอ็กโซไซโทซิส ในกระบวนการย้อนกลับ บริเวณของเยื่อหุ้มเซลล์จะบุ๋มเข้าไปด้านในและในที่สุดก็จะแยกตัวออก ห่อหุ้มส่วนหนึ่งของของเหลวภายนอก เซลล์ เพื่อขนส่งเข้าไปในเซลล์เอนโดไซโทซิสและเอ็กโซไซโทซิสอาศัยกลไกโมเลกุลที่แตกต่างกันมากในการทำงาน แต่ทั้งสองกระบวนการมีความเชื่อมโยงกันอย่างใกล้ชิดและไม่สามารถทำงานได้หากขาดซึ่งกันและกัน กลไกหลักของการพึ่งพาซึ่งกันและกันนี้คือปริมาณไขมันจำนวนมากที่เกี่ยวข้อง[ 70 ]ในเซลล์ทั่วไป พื้นที่ของเยื่อไขมันสองชั้นที่เทียบเท่ากับเยื่อหุ้มพลาสมาทั้งหมดจะผ่านวงจรเอนโดไซโทซิส/เอ็กโซไซโทซิสในเวลาประมาณครึ่งชั่วโมง[ 71 ]

เอ็กโซไซโทซิสในโปรคาริโอต : เอ็กโซไซ โทซิสของถุง เมม เบรน ซึ่ง เป็นที่รู้จักกันทั่วไปใน ชื่อการขนส่งถุงเมมเบรน ซึ่งเป็นกระบวนการที่ได้รับรางวัลโนเบล (ปี 2013) ถือเป็นลักษณะเฉพาะของเซลล์ยูคาริโอต[ 72 ] อย่างไรก็ตาม ความเชื่อนี้ถูกทำลายลงด้วยการเปิดเผยว่านาโนเวสิเคิล ซึ่งเป็นที่รู้จักกันทั่วไปในชื่อเวสิเคิลเมมเบรนชั้นนอกของแบคทีเรียที่ปล่อยออกมาจากจุลินทรีย์แกรมลบ จะเคลื่อนย้ายโมเลกุลสัญญาณของแบคทีเรียไปยังเซลล์เจ้าบ้านหรือเซลล์เป้าหมาย[ 73 ]เพื่อดำเนินกระบวนการต่างๆ ที่เป็นประโยชน์ต่อจุลินทรีย์ที่หลั่งสาร เช่น ในการบุกรุกเซลล์เจ้าบ้าน[ 74 ]และปฏิสัมพันธ์ระหว่างจุลินทรีย์กับสิ่งแวดล้อมโดยทั่วไป[ 75 ]
อิเล็กโทรพอเรชั่น
อิเล็กโตรพอเรชันคือการเพิ่มขึ้นอย่างรวดเร็วของความสามารถในการซึมผ่านของเยื่อหุ้มเซลล์ที่เกิดจากการใช้สนามไฟฟ้าเทียมขนาดใหญ่ข้ามเยื่อหุ้มเซลล์ ในทางทดลอง อิเล็กโตรพอเรชันถูกใช้เพื่อนำโมเลกุลที่ชอบน้ำเข้าสู่เซลล์ เป็นเทคนิคที่มีประโยชน์อย่างยิ่งสำหรับโมเลกุลขนาดใหญ่ที่มีประจุสูง เช่นDNAซึ่งจะไม่สามารถแพร่กระจายผ่านแกนกลางของเยื่อหุ้มเซลล์ที่ไม่ชอบน้ำได้โดย ธรรมชาติ [ 76 ]ด้วยเหตุนี้ อิเล็กโตรพอเรชันจึงเป็นหนึ่งในวิธีการสำคัญของการถ่ายทอดยีนและการเปลี่ยนแปลง ของแบคทีเรีย มีการเสนอว่าอิเล็กโตรพอเรชันที่เกิดจากฟ้าผ่าอาจเป็นกลไกของการถ่ายทอดยีนแนวนอน ตาม ธรรมชาติ[ 77 ]
กลศาสตร์

ชั้นไขมันสองชั้นเป็นโครงสร้างที่มีขนาดใหญ่พอที่จะมีคุณสมบัติทางกลบางอย่างของของเหลวหรือของแข็งได้ โมดูลัสการบีบอัดพื้นที่ K aโมดูลัสการดัดงอ K bและพลังงานขอบสามารถใช้เพื่ออธิบายคุณสมบัติเหล่านี้ได้ ชั้นไขมันสองชั้นที่เป็นของแข็งยังมีโมดูลัสเฉือน ด้วย แต่เช่นเดียวกับของเหลวใดๆ โมดูลัสเฉือนจะเป็นศูนย์สำหรับชั้นไขมันสองชั้นที่เป็นของเหลว คุณสมบัติทางกลเหล่านี้ส่งผลต่อการทำงานของเยื่อหุ้มเซลล์ K aและ K bส่งผลต่อความสามารถของโปรตีนและโมเลกุลขนาดเล็กในการแทรกเข้าไปในชั้นไขมันสองชั้น[ 78 ] [ 79 ]และคุณสมบัติทางกลของชั้นไขมันสองชั้นได้รับการแสดงให้เห็นว่าสามารถเปลี่ยนแปลงการทำงานของช่องไอออนที่ถูกกระตุ้นด้วยกลไกได้[ 80 ]คุณสมบัติทางกลของชั้นไขมันสองชั้นยังควบคุมประเภทของความเครียดที่เซลล์สามารถทนได้โดยไม่ฉีกขาด แม้ว่าชั้นไขมันสองชั้นจะงอได้ง่าย แต่ส่วนใหญ่ไม่สามารถยืดได้มากกว่าไม่กี่เปอร์เซ็นต์ก่อนที่จะแตก[ 81 ]

ดังที่ได้กล่าวไว้ในส่วนโครงสร้างและการจัดระเบียบ แรงดึงดูดแบบไฮโดรโฟบิกของหางลิปิดในน้ำเป็นแรงหลักที่ยึดลิปิดไบเลเยอร์เข้าด้วยกัน ดังนั้น โมดูลัสความยืดหยุ่นของไบเลเยอร์จึงถูกกำหนดโดยพื้นที่ส่วนเกินที่สัมผัสกับน้ำเมื่อโมเลกุลลิปิดถูกยืดออกจากกันเป็นหลัก[ 82 ]ด้วยความเข้าใจเกี่ยวกับแรงที่เกี่ยวข้องนี้ จึงไม่น่าแปลกใจที่การศึกษาต่างๆ แสดงให้เห็นว่า K aแปรผันอย่างมากกับความดันออสโมติก[ 83 ]แต่แปรผันเพียงเล็กน้อยกับความยาวของหางและความไม่อิ่มตัว[ 13 ]เนื่องจากแรงที่เกี่ยวข้องมีขนาดเล็กมาก จึงเป็นการยากที่จะหาค่า K a ได้จากการทดลอง เทคนิคส่วนใหญ่ต้องใช้กล้องจุลทรรศน์ที่ซับซ้อนและอุปกรณ์วัดที่มีความไวสูงมาก[ 59 ] [ 84 ]
ตรงกันข้ามกับ K aซึ่งเป็นการวัดปริมาณพลังงานที่จำเป็นในการยืดไบเลเยอร์ K bเป็นการวัดปริมาณพลังงานที่จำเป็นในการดัดหรือโค้งงอไบเลเยอร์ ตามหลักการแล้ว โมดูลัสการดัดงอถูกกำหนดให้เป็นพลังงานที่จำเป็นในการเปลี่ยนรูปของเยื่อหุ้มเซลล์จากความโค้งภายในไปสู่ความโค้งอื่น ความโค้งภายในถูกกำหนดโดยอัตราส่วนของเส้นผ่านศูนย์กลางของกลุ่มหัวต่อกลุ่มหาง สำหรับลิปิด PC สองหาง อัตราส่วนนี้เกือบเป็นหนึ่ง ดังนั้นความโค้งภายในจึงเกือบเป็นศูนย์ หากลิปิดชนิดใดชนิดหนึ่งมีความเบี่ยงเบนจากความโค้งภายในเป็นศูนย์มากเกินไป มันจะไม่ก่อตัวเป็นไบเลเยอร์และจะก่อตัวเป็นเฟสอื่น เช่นไมเซลล์หรือไมเซลล์กลับหัว การเติมโมเลกุลไฮโดรฟิลิกขนาดเล็ก เช่นซูโครสลงในไลโปโซมลามิลลาร์ลิปิดผสมที่ทำจากเยื่อไทลาคอยด์ที่อุดมไปด้วยกาแลคโตลิปิด จะทำให้ไบเลเยอร์ไม่เสถียรและกลายเป็นเฟสไมเซลล์[ 85 ]
เป็นการวัดปริมาณพลังงานที่ต้องใช้ในการทำให้ขอบของไบเลเยอร์สัมผัสกับน้ำโดยการฉีกไบเลเยอร์หรือสร้างรูในนั้น ที่มาของพลังงานนี้คือข้อเท็จจริงที่ว่าการสร้างอินเทอร์เฟซดังกล่าวทำให้หางของลิปิดบางส่วนสัมผัสกับน้ำ แต่ทิศทางที่แน่นอนของลิปิดขอบเหล่านี้ยังไม่เป็นที่ทราบแน่ชัด มีหลักฐานบางอย่างที่แสดงว่ารูพรุนทั้งแบบไฮโดรโฟบิก (หางตรง) และไฮโดรฟิลิก (หัวโค้งงอ) สามารถอยู่ร่วมกันได้[ 86 ] [ 87 ]
ฟิวชั่น

การหลอมรวมเป็นกระบวนการที่เยื่อไขมันสองชั้นรวมกัน ส่งผลให้เกิดโครงสร้างที่เชื่อมต่อกันหนึ่งเดียว[ 88 ]หากการหลอมรวมนี้เกิดขึ้นอย่างสมบูรณ์ผ่านทั้งสองชั้นของเยื่อไขมันทั้งสองชั้น จะเกิดสะพานที่เต็มไปด้วยน้ำ และสารละลายที่บรรจุอยู่ภายในเยื่อไขมันสามารถผสมกันได้ ในทางกลับกัน หากมีเพียงชั้นเดียวจากแต่ละเยื่อไขมันที่เกี่ยวข้องกับกระบวนการหลอมรวม เยื่อไขมันเหล่านั้นจะถูกเรียกว่าหลอมรวมครึ่งหนึ่ง การหลอมรวมเกี่ยวข้องกับกระบวนการของเซลล์หลายอย่าง โดยเฉพาะอย่างยิ่งในยูคาริโอตเนื่องจากเซลล์ยูคาริโอตถูกแบ่งย่อยอย่างกว้างขวางโดยเยื่อไขมันสองชั้นการปล่อยสารออกนอก เซลล์ การปฏิสนธิ ของไข่โดยการกระตุ้นอสุจิและการขนส่งของเสียไปยังไลโซโซม เป็นเพียงตัวอย่างบางส่วนของกระบวนการยูคาริโอตจำนวนมากที่อาศัยการหลอมรวมในรูปแบบใดรูปแบบหนึ่ง แม้แต่การเข้าสู่เซลล์ของเชื้อโรคก็สามารถควบคุมได้ด้วยการหลอมรวม เนื่องจาก ไวรัสที่เคลือบด้วยเยื่อไขมันหลายชนิดมีโปรตีนหลอมรวมเฉพาะเพื่อเข้าสู่เซลล์เจ้าบ้าน[ 89 ]
กระบวนการหลอมรวมมีสี่ขั้นตอนพื้นฐาน[ 37 ]ขั้นแรก เยื่อหุ้มที่เกี่ยวข้องต้องรวมตัวกัน เข้าใกล้กันจนเหลือระยะห่างเพียงไม่กี่นาโนเมตร ขั้นที่สอง เยื่อหุ้มสองชั้นต้องสัมผัสกันอย่างใกล้ชิด (ภายในไม่กี่อังสตรอม) เพื่อให้เกิดการสัมผัสอย่างใกล้ชิดนี้ พื้นผิวทั้งสองต้องสูญเสียน้ำอย่างน้อยบางส่วน เนื่องจากน้ำที่เกาะอยู่บนพื้นผิวตามปกติจะทำให้เยื่อหุ้มสองชั้นผลักกันอย่างรุนแรง การมีอยู่ของไอออน โดยเฉพาะไอออนบวกสองวาเลนต์ เช่น แมกนีเซียมและแคลเซียม มีผลอย่างมากต่อขั้นตอนนี้[ 90 ] [ 91 ]หนึ่งในบทบาทสำคัญของแคลเซียมในร่างกายคือการควบคุมการหลอมรวมของเยื่อหุ้ม ขั้นที่สาม ความไม่เสถียรต้องเกิดขึ้น ณ จุดหนึ่งระหว่างเยื่อหุ้มสองชั้น ทำให้โครงสร้างของเยื่อหุ้มบิดเบี้ยวในบริเวณนั้น ลักษณะที่แท้จริงของการบิดเบี้ยวนี้ยังไม่เป็นที่ทราบแน่ชัด ทฤษฎีหนึ่งคือ "ก้าน" ที่โค้งงอมากต้องก่อตัวขึ้นระหว่างเยื่อหุ้มสองชั้น[ 92 ]ผู้สนับสนุนทฤษฎีนี้เชื่อว่ามันอธิบายได้ว่าทำไมฟอสฟาติดิลเอทานอลามีน ซึ่งเป็นลิปิดที่มีความโค้งสูง จึงส่งเสริมการหลอมรวม ในที่สุด ในขั้นตอนสุดท้ายของการหลอมรวม จุดบกพร่องนี้จะเติบโตขึ้น และส่วนประกอบของไบเลเยอร์ทั้งสองจะผสมกันและแพร่กระจายออกไปจากบริเวณที่สัมผัสกัน[ 88 ]


สถานการณ์จะซับซ้อนยิ่งขึ้นเมื่อพิจารณาการหลอมรวมในร่างกายเนื่องจากกระบวนการหลอมรวมทางชีวภาพเกือบทุกครั้งจะถูกควบคุมโดยการทำงานของโปรตีนที่เกี่ยวข้องกับเยื่อหุ้มเซลล์โปรตีนกลุ่มแรกที่ได้รับการศึกษาคือโปรตีนหลอมรวมของไวรัส ซึ่งช่วยให้ไวรัส ที่มีเปลือกหุ้ม สามารถแทรกสารพันธุกรรมเข้าไปในเซลล์เจ้าบ้านได้ (ไวรัสที่มีเปลือกหุ้มคือไวรัสที่ล้อมรอบด้วยเยื่อไขมันสองชั้น บางชนิดมีเพียงเปลือกโปรตีน) เซลล์ ยูคาริโอตก็ใช้โปรตีนหลอมรวมเช่นกัน ซึ่งโปรตีนที่ได้รับการศึกษามากที่สุดคือSNAREโปรตีน SNARE ใช้ในการควบคุม การขนส่ง เวสิเคิลภายในเซลล์ทั้งหมด แม้ว่าจะมีการศึกษามาหลายปีแล้ว แต่ก็ยังมีสิ่งที่ไม่ทราบอีกมากเกี่ยวกับหน้าที่ของโปรตีนกลุ่มนี้ อันที่จริง ยังคงมีการถกเถียงกันอย่างต่อเนื่องว่า SNARE เชื่อมโยงกับการเชื่อมต่อในระยะแรกหรือมีส่วนร่วมในกระบวนการหลอมรวมในภายหลังโดยการอำนวยความสะดวกในการหลอมรวมครึ่งหนึ่ง[ 94 ]
ในการศึกษาชีววิทยาระดับโมเลกุลและเซลล์ มักเป็นที่ต้องการที่จะเหนี่ยวนำให้เกิดการหลอมรวมกันโดยเทียม การเติมโพลีเอทิลีนไกลคอล (PEG) ทำให้เกิดการหลอมรวมกันโดยไม่มีการรวมกลุ่มหรือการรบกวนทางชีวเคมีอย่างมีนัยสำคัญ ปัจจุบันมีการใช้กระบวนการนี้อย่างแพร่หลาย เช่น การหลอมรวมเซลล์ Bกับเซลล์มะเร็งไขกระดูก[ 95 ] " ไฮบริดโดมา " ที่ได้จากการรวมกันนี้จะแสดงแอนติบอดี ที่ต้องการ ตามที่กำหนดโดยเซลล์ B ที่เกี่ยวข้อง แต่จะกลายเป็นอมตะเนื่องจากส่วนประกอบของมะเร็งผิวหนัง การหลอมรวมยังสามารถเหนี่ยวนำได้โดยเทียมผ่าน กระบวนการ อิเล็กโทรโพเรชัน ซึ่งเรียกว่าอิเล็กโทรฟิวชั่น เชื่อกันว่าปรากฏการณ์นี้เกิดจากขอบที่มีพลังงานสูงที่เกิดขึ้นระหว่างอิเล็กโทรโพเรชัน ซึ่งสามารถทำหน้าที่เป็นจุดบกพร่องเฉพาะที่เพื่อกระตุ้นการเจริญเติบโตของก้านระหว่างสองชั้น[ 96 ]
ระบบจำลอง
สามารถสร้างเยื่อไขมันสองชั้นเทียมขึ้นในห้องปฏิบัติการเพื่อให้ผู้วิจัยสามารถทำการทดลองที่ไม่สามารถทำได้กับเยื่อไขมันสองชั้นตามธรรมชาติ นอกจากนี้ยังสามารถใช้ในสาขาชีววิทยาสังเคราะห์เพื่อกำหนดขอบเขตของเซลล์เทียมระบบสังเคราะห์เหล่านี้เรียกว่าเยื่อไขมันสองชั้นแบบจำลอง มีเยื่อไขมันสองชั้นแบบจำลองหลายประเภท แต่ละประเภทมีข้อดีและข้อเสียในการทดลอง สามารถสร้างได้จากไขมันสังเคราะห์หรือไขมันธรรมชาติ ระบบแบบจำลองที่พบได้บ่อยที่สุด ได้แก่: [ 97 ]
การใช้งานเชิงพาณิชย์
จนถึงปัจจุบัน การประยุกต์ใช้เชิงพาณิชย์ที่ประสบความสำเร็จมากที่สุดของชั้นไขมันสองชั้นคือการใช้ไลโปโซมสำหรับการส่งยา โดยเฉพาะอย่างยิ่งสำหรับการรักษามะเร็ง (หมายเหตุ - คำว่า "ไลโปโซม" โดยพื้นฐานแล้วมีความหมายเหมือนกับ " เวสิเคิล " ยกเว้นว่าเวสิเคิลเป็นคำทั่วไปสำหรับโครงสร้าง ในขณะที่ไลโปโซมหมายถึงเฉพาะเวสิเคิลเทียม ไม่ใช่เวสิเคิลตามธรรมชาติ) แนวคิดพื้นฐานของการส่งยาด้วยไลโปโซมคือ ยาจะถูกห่อหุ้มในสารละลายภายในไลโปโซม จากนั้นฉีดเข้าไปในผู้ป่วย ไลโปโซมที่บรรจุยาเหล่านี้จะเดินทางผ่านระบบจนกว่าจะจับกับบริเวณเป้าหมายและแตกออก ปล่อยยาออกมา ในทางทฤษฎี ไลโปโซมควรเป็นระบบส่งยาที่เหมาะสม เนื่องจากสามารถแยกยาที่ชอบน้ำได้เกือบทุกชนิด สามารถเชื่อมต่อกับโมเลกุลเพื่อกำหนดเป้าหมายเนื้อเยื่อเฉพาะ และค่อนข้างไม่เป็นพิษ เนื่องจากร่างกายมีวิถีทางชีวเคมีในการย่อยสลายไขมัน[ 98 ]
ไลโปโซมนำส่งยารุ่นแรกมีองค์ประกอบของไขมันที่เรียบง่ายและมีข้อจำกัดหลายประการ การไหลเวียนในกระแสเลือดมีจำกัดอย่างมากเนื่องจาก การกำจัด โดยไตและ การกลืนกินโดยฟา โกไซต์การปรับปรุงองค์ประกอบของไขมันเพื่อปรับความลื่นไหล ความหนาแน่นของประจุบนพื้นผิว และความชุ่มชื้นของพื้นผิว ส่งผลให้เวสิเคิลดูดซับโปรตีนจากซีรั่ม น้อยลง จึงทำให้ระบบภูมิคุ้มกัน จดจำได้ยาก ขึ้น[ 99 ]ความก้าวหน้าที่สำคัญที่สุดในด้านนี้คือการปลูกถ่ายโพลีเอทิลีนไกลคอล (PEG) ลงบนพื้นผิวของไลโปโซมเพื่อสร้างเวสิเคิล "ล่องหน" ซึ่งไหลเวียนได้เป็นเวลานานโดยไม่ถูกกำจัดโดยระบบภูมิคุ้มกันหรือไต[ 100 ]
ไลโปโซมล่องหนรุ่นแรกถูกกำหนดเป้าหมายไปยัง เนื้อเยื่อ เนื้องอก โดยทางอ้อม เนื่องจากเนื้องอกกระตุ้นการสร้างหลอดเลือด ใหม่อย่างรวดเร็วและควบคุมไม่ได้ ทำให้ไลโปโซม "รั่ว" เป็นพิเศษและยอมให้ไลโปโซมออกจากกระแสเลือดได้ในอัตราที่สูงกว่าเนื้อเยื่อปกติมาก[ 101 ]เมื่อไม่นานมานี้ มีการดำเนินการวิจัยเพื่อปลูกถ่ายแอนติบอดีหรือเครื่องหมายโมเลกุลอื่นๆ ลงบนพื้นผิวของไลโปโซม โดยหวังว่าจะสามารถจับกับเซลล์หรือเนื้อเยื่อชนิดใดชนิดหนึ่งได้อย่างมีประสิทธิภาพ[ 102 ]ตัวอย่างบางส่วนของแนวทางนี้อยู่ในขั้นตอนการทดลองทางคลินิกแล้ว[ 103 ]
การประยุกต์ใช้ที่เป็นไปได้อีกอย่างหนึ่งของชั้นไขมันสองชั้นคือสาขาไบโอเซนเซอร์เนื่องจากชั้นไขมันสองชั้นเป็นกำแพงกั้นระหว่างภายในและภายนอกเซลล์ จึงเป็นแหล่งของการส่งสัญญาณอย่างกว้างขวาง นักวิจัยได้พยายามใช้ประโยชน์จากศักยภาพนี้มาหลายปีเพื่อพัฒนาอุปกรณ์ที่ใช้ชั้นไขมันสองชั้นสำหรับการวินิจฉัยทางคลินิกหรือการตรวจจับการก่อการร้ายทางชีวภาพ ความคืบหน้าในด้านนี้ค่อนข้างช้า และถึงแม้ว่าจะมีบริษัทไม่กี่แห่งที่พัฒนาระบบตรวจจับอัตโนมัติที่ใช้ไขมัน แต่ก็ยังคงมุ่งเป้าไปที่ชุมชนวิจัย ซึ่งรวมถึง Biacore (ปัจจุบันคือ GE Healthcare Life Sciences) ซึ่งมีชิปแบบใช้แล้วทิ้งสำหรับใช้ชั้นไขมันสองชั้นในการศึกษาจลนศาสตร์การจับ[ 104 ]และ Nanion Inc. ซึ่งได้พัฒนาระบบจับยึดแบบอัตโนมัติ[ 105 ]
เยื่อไขมันสองชั้นที่รองรับ (SLB) ตามที่อธิบายไว้ข้างต้นประสบความสำเร็จในเชิงพาณิชย์ในฐานะเทคนิคการคัดกรองเพื่อวัดการซึมผ่านของยา เทคนิคการทดสอบการซึมผ่านของเยื่อเทียมแบบขนาน ( PAMPA ) นี้วัดการซึมผ่าน ของค็อกเทลไขมันที่กำหนดสูตรเฉพาะซึ่งพบว่ามีความสัมพันธ์สูงกับวัฒนธรรมCaco-2 [ 106 ] [ 107 ]ทางเดินอาหาร [ 108 ]อุปสรรคเลือด-สมอง[ 109 ]และผิวหนัง[ 110 ]
ประวัติศาสตร์
ในช่วงต้นศตวรรษที่ 20 นักวิทยาศาสตร์เชื่อว่าเซลล์ถูกล้อมรอบด้วยเยื่อบางๆ คล้ายน้ำมัน[ 111 ]แต่ลักษณะโครงสร้างของเยื่อหุ้มนี้ยังไม่เป็นที่ทราบแน่ชัด การทดลองสองครั้งในปี 1925 ได้วางรากฐานเพื่อเติมเต็มช่องว่างนี้ โดยการวัดค่าความจุของ สารละลาย เม็ดเลือดแดงฮิวโก้ ฟริค ได้กำหนดว่าเยื่อหุ้มเซลล์มีความหนา 3.3 นาโนเมตร[ 112 ]
แม้ว่าผลลัพธ์ของการทดลองนี้จะถูกต้อง แต่ Fricke ตีความข้อมูลผิดพลาดโดยคิดว่าเยื่อหุ้มเซลล์เป็นชั้นโมเลกุลเดียว ศาสตราจารย์ ดร. Evert Gorter [ 113 ] (1881–1954) และ F. Grendel จากมหาวิทยาลัย Leiden ได้เข้าถึงปัญหาจากมุมมองที่แตกต่างออกไป โดยกระจายไขมันของเม็ดเลือดแดงเป็นชั้นเดียวบนราง Langmuir–Blodgettเมื่อพวกเขาเปรียบเทียบพื้นที่ของชั้นเดียวกับพื้นที่ผิวของเซลล์ พวกเขาพบอัตราส่วนสองต่อหนึ่ง[ 114 ]การวิเคราะห์ในภายหลังแสดงให้เห็นข้อผิดพลาดและข้อสมมติฐานที่ไม่ถูกต้องหลายประการในการทดลองนี้ แต่โดยบังเอิญ ข้อผิดพลาดเหล่านี้ได้หักล้างกัน และจากข้อมูลที่ผิดพลาดนี้ Gorter และ Grendel ได้ข้อสรุปที่ถูกต้องว่าเยื่อหุ้มเซลล์เป็นชั้นไขมันสองชั้น[ 37 ]
ทฤษฎีนี้ได้รับการยืนยันผ่านการใช้กล้องจุลทรรศน์อิเล็กตรอนในช่วงปลายทศวรรษ 1950 แม้ว่าเขาจะไม่ได้ตีพิมพ์งานวิจัยกล้องจุลทรรศน์อิเล็กตรอนชิ้นแรกของเยื่อไขมัน สองชั้น [ 115 ]แต่ J. David Robertson เป็นคนแรกที่ยืนยันว่าแถบสีเข้มที่มีความหนาแน่นของอิเล็กตรอนสองแถบนั้นคือกลุ่มหัวและโปรตีนที่เกี่ยวข้องของเยื่อไขมันสองชั้นที่อยู่ตรงข้ามกัน ในงานชุดนี้ Robertson ได้เสนอแนวคิดของ "หน่วยเยื่อหุ้มเซลล์" ซึ่งเป็นครั้งแรกที่โครงสร้างเยื่อสองชั้นได้รับการกำหนดให้กับเยื่อหุ้มเซลล์ทั้งหมดรวมถึงเยื่อหุ้มออร์แกเนลล์ด้วย[ 116 ] [ 117 ]
ในเวลาเดียวกัน การพัฒนาเมมเบรนจำลองยืนยันว่าชั้นไขมันสองชั้นเป็นโครงสร้างที่เสถียรซึ่งสามารถดำรงอยู่ได้โดยอิสระจากโปรตีน โดยการ "ทาสี" สารละลายไขมันในตัวทำละลายอินทรีย์ลงบนช่องเปิด Mueller และ Rudin สามารถสร้างชั้นไขมันสองชั้นเทียมและพบว่ามีคุณสมบัติการไหลในแนวด้านข้าง ความต้านทานไฟฟ้าสูง และการซ่อมแซมตัวเองเมื่อถูกเจาะ[ 118 ]ซึ่งทั้งหมดนี้เป็นคุณสมบัติของเยื่อหุ้มเซลล์ตามธรรมชาติ ไม่กี่ปีต่อมาAlec Banghamแสดงให้เห็นว่าชั้นไขมันสองชั้นในรูปของถุงไขมันสามารถเกิดขึ้นได้ง่ายๆ โดยการนำตัวอย่างไขมันแห้งไปสัมผัสกับน้ำ ซึ่งแสดงให้เห็นว่าชั้นไขมันสองชั้นเกิดขึ้นเองโดยธรรมชาติผ่านการประกอบตัวเองและไม่จำเป็นต้องมีโครงสร้างรองรับที่มีรูปแบบ[ 119 ]ในปี 1977 Kunitake และ Okahata ได้เตรียมเมมเบรนสองชั้นสังเคราะห์ทั้งหมดจากสารประกอบอินทรีย์เพียงชนิดเดียว คือ didodecyldimethylammonium bromide ซึ่งแสดงให้เห็นว่าเมมเบรนสองชั้นประกอบขึ้นโดยแรงระหว่างโมเลกุล[ 120 ]
ดูเพิ่มเติม
ลิงก์ภายนอก
- LIPIDATฐานข้อมูลที่ครอบคลุมเกี่ยวกับคุณสมบัติทางกายภาพของไขมัน
- โครงสร้างของเยื่อไขมันสองชั้นแบบของเหลว เก็บถาวรเมื่อวันที่ 11 เมษายน 2554 ที่Wayback Machineการจำลองและลิงก์สิ่งพิมพ์ที่เกี่ยวข้องกับโครงสร้างภาคตัดขวางของเยื่อไขมันสองชั้น
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ชั้นไขมันสองชั้น
ชั้นไขมันสองชั้น (หรือชั้นฟอสโฟลิปิด สองชั้น ) เป็นเยื่อบาง ๆ ที่มีขั้ว ประกอบด้วยโมเลกุลไขมัน สองชั้น เยื่อเหล่านี้สร้างเป็นกำแพงต่อเนื่องรอบเซลล์ ทั้งหมด เยื่อหุ้มเซลล์ ของ...
โครงสร้างและองค์กร
เมื่อฟอสโฟลิปิดสัมผัสกับน้ำ พวกมัน จะประกอบตัวเอง เป็นแผ่นสองชั้นโดยมีหางที่ไม่ชอบน้ำชี้ไปทางตรงกลางของแผ่น การจัดเรียงนี้ส่งผลให้เกิด 'แผ่น' สองแผ่นซึ่งแต่ละแผ่นเป็นชั้นโมเลกุลเดียว ตรงกลางของชั้นสองชั้นนี้แทบไม่มีน้ำและกีดกันโมเลกุลเช่น น้ำตาล...
การวิเคราะห์ภาคตัดขวาง
ชั้นไขมันสองชั้นนั้นบางมากเมื่อเทียบกับขนาดด้านข้าง หากเซลล์สัตว์เลี้ยงลูกด้วยนมทั่วไป (เส้นผ่านศูนย์กลางประมาณ 10 ไมโครเมตร) ถูกขยายให้มีขนาดเท่าแตงโม (ประมาณ 1 ฟุต/30 เซนติเมตร) ชั้นไขมันสองชั้นที่ประกอบเป็นเยื่อหุ้มพลาสมา จะ...
ความไม่สมมาตร
ในเยื่อหุ้มเซลล์แบบสองชั้นที่เกิดขึ้นตามธรรมชาติหลายชนิด องค์ประกอบของเยื่อหุ้มเซลล์ชั้นในและชั้นนอกจะแตกต่างกัน ใน เซลล์เม็ดเลือดแดง ของมนุษย์ เยื่อหุ้มเซลล์ชั้นใน (ไซโตพลาสมิก) ส่วนใหญ่ประกอบด้วย ฟอสฟาติ ดิ ล เอ ทานอลามี น ฟอสฟาติดิลเซอรีน และ ฟอสฟา...