อ่าน 21 นาที
ไวรัสโคโรนาที่เกี่ยวข้องกับ SARS
ไวรัสโคโรนาที่เกี่ยวข้องกับโรคทางเดินหายใจเฉียบพลันรุนแรง ( SARSr-CoVหรือSARS-CoV , Betacoronavirus pandemicum ) เป็นไวรัสชนิด หนึ่งที่ประกอบด้วยสายพันธุ์ที่รู้จักกันหลาย...
ไวรัสโคโรนาที่เกี่ยวข้องกับ SARS
| ไวรัสโคโรนาที่เกี่ยวข้องกับ SARS | |
|---|---|
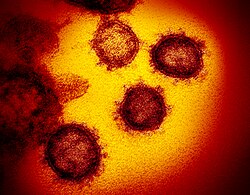 | |
| ภาพถ่ายอิเล็กตรอนไมโครสโคปแบบส่งผ่านของไวรัสโคโรนาที่เกี่ยวข้องกับ SARS ที่เกิดขึ้นจากเซลล์เจ้าบ้านที่เพาะเลี้ยงในห้องปฏิบัติการ | |
| การจำแนกประเภทไวรัส | |
| (ไม่จัดอันดับ): | ไวรัส |
| อาณาจักร: | ไรโบวิเรีย |
| อาณาจักร: | ออร์ธอร์นาไวเร |
| ไฟลัม: | พิสุวิริโคตา |
| ระดับ: | พิโซนิวิริเซเตส |
| คำสั่ง: | นิโดไวรัลส์ |
| ตระกูล: | ไวรัสโคโรนา |
| ประเภท: | เบตาโคโรนาไวรัส |
| สกุลย่อย: | ไวรัสซาร์เบโค |
| สายพันธุ์: | การระบาดใหญ่ของไวรัสโคโรนาเบตา |
| สายพันธุ์ | |
| |
| คำพ้องความหมาย | |
| |
ไวรัสโคโรนาที่เกี่ยวข้องกับโรคทางเดินหายใจเฉียบพลันรุนแรง ( SARSr-CoVหรือSARS-CoV , Betacoronavirus pandemicum ) [ 2 ] [หมายเหตุ 1 ]เป็นไวรัสชนิด หนึ่งที่ประกอบด้วยสายพันธุ์ที่รู้จักกันหลาย สายพันธุ์ไวรัสสองสายพันธุ์นี้ก่อให้เกิดการระบาด ของ โรคทางเดินหายใจรุนแรงในมนุษย์ ได้แก่ไวรัสโคโรนาที่ก่อให้เกิดโรคทางเดินหายใจเฉียบพลันรุนแรง 1 (SARS-CoV หรือ SARS-CoV-1) ซึ่งเป็นสาเหตุของ การระบาด ของโรคทางเดินหายใจเฉียบพลันรุนแรง (SARS) ในปี 2545-2547และไวรัสโคโรนาที่ก่อให้เกิดโรคทางเดินหายใจเฉียบพลันรุนแรง 2 (SARS-CoV-2) ซึ่งเป็นสาเหตุของการระบาดใหญ่ของCOVID-19 [ 3 ] [ 4 ] นอกจากนี้ยังมีไวรัส SARSr-CoV อีกหลายร้อยสายพันธุ์ ซึ่งทราบกันว่าติดเชื้อเฉพาะในสัตว์เลี้ยงลูกด้วยนมที่ไม่ใช่มนุษย์เท่านั้น เช่นค้างคาวเป็นแหล่งสะสมหลักของไวรัส SARSr-CoV หลายสายพันธุ์ มีการระบุสายพันธุ์ต่างๆ ในชะมดหิมาลัยซึ่งน่าจะเป็นบรรพบุรุษของ SARS-CoV-1 [ 3 ] [ 5 ] [ 6 ] [ 7 ]
ไวรัส RNA สายเดี่ยวที่มีเปลือกหุ้มและ มีทิศทางบวก เหล่านี้จะเข้าสู่เซลล์โฮสต์โดยการจับกับ ตัวรับ เอนไซม์แองจิโอเทนซินคอนเวอร์ติง 2 (ACE2) [ 8 ]สายพันธุ์ SARSr-CoV เป็นสมาชิกของสกุลBetacoronavirusและเป็นสายพันธุ์เดียวของสกุลย่อยSarbecovirus ( SARS Betacoronavirus ) [ 9 ] [ 10 ]
ไวรัสโคโรนาที่เกี่ยวข้องกับ SARS เป็นหนึ่งในไวรัสหลายชนิดที่องค์การอนามัยโลก (WHO) ระบุไว้ในปี 2016 ว่าเป็นสาเหตุที่น่าจะเป็นไปได้ของการระบาด ในอนาคต ในแผนใหม่ที่พัฒนาขึ้นหลังจากการระบาดของอีโบลาเพื่อการวิจัยและพัฒนาอย่างเร่งด่วนก่อนและระหว่างการระบาด ไปสู่การทดสอบวินิจฉัยวัคซีนและยาการคาดการณ์นี้เป็นจริงกับ การระบาดใหญ่ ของCOVID-19 [ 11 ] [ 12 ]
การจำแนกประเภท
ไวรัสโคโรนาที่เกี่ยวข้องกับ SARS เป็นสมาชิกของสกุลBetacoronavirus (กลุ่ม 2) และเป็นโมโนไทป์ของสกุลย่อยSarbecovirus (กลุ่มย่อย B) [ 13 ] Sarbecoviruses แตกต่างจากembecovirusesหรือalphacoronaviruses ตรงที่มี โปรตีเอสคล้ายปาเปน (PLpro) เพียงตัวเดียวแทนที่จะมีสองตัวในกรอบการอ่านแบบเปิดORF1ab [ 14 ] SARSr-CoV ถูกกำหนดให้เป็นกลุ่มที่แยกตัวออกมาในช่วงแรกจาก betacoronaviruses โดยพิจารณาจากชุดโดเมนที่อนุรักษ์ไว้ซึ่งมีร่วมกับกลุ่มนี้[ 15 ] [ 16 ]
ค้างคาวเป็นแหล่งกักเก็บหลักของไวรัสโคโรนาที่เกี่ยวข้องกับ SARS เช่น SARS-CoV-1 และ SARS-CoV-2 ไวรัสได้วิวัฒนาการร่วมกันในแหล่งกักเก็บค้างคาวในช่วงระยะเวลานาน[ 17 ]เพิ่งไม่นานมานี้เองที่พบว่าสายพันธุ์ของไวรัสโคโรนาที่เกี่ยวข้องกับ SARS ได้วิวัฒนาการจนสามารถแพร่ข้ามสายพันธุ์จากค้างคาวสู่มนุษย์ได้ ดังเช่นในกรณีของสายพันธุ์SARS-CoV-1และSARS-CoV-2 [ 18 ] [ 8 ] ทั้งสองสายพันธุ์นี้สืบเชื้อสายมาจากบรรพบุรุษเดียวกัน แต่แพร่ข้ามสายพันธุ์มาสู่มนุษย์แยกกัน SARS-CoV-2 ไม่ใช่ลูกหลานโดยตรงของ SARS-CoV-1 [ 3 ]
จีโนม

ไวรัสโคโรนาที่เกี่ยวข้องกับ SARS เป็นไวรัส RNA สายเดี่ยวที่มีเปลือกหุ้มและมีทิศทางบวกจีโนมของมันมีขนาดประมาณ 30 กิโลเบสซึ่งเป็นหนึ่งในจีโนมที่ใหญ่ที่สุดในบรรดาไวรัส RNA ไวรัสนี้มีกรอบการอ่านแบบเปิด 14 เฟรม ซึ่งทับซ้อนกันในบางกรณี[ 19 ]จีโนมมีหมวกเมทิลเลต 5′ ตามปกติ และหางโพลีอะดีนิเลต 3′ [ 20 ] มีนิ วคลีโอ ไทด์ 265 ตัว ใน5'UTRและนิวคลีโอไทด์ 342 ตัวใน3'UTR [ 19 ]
หมวกเมทิลเลต 5' และหางโพลีอะดีนิเลต 3' ช่วยให้จีโนม RNA ที่มีทิศทางบวกสามารถแปล โดยตรงโดย ไรโบโซมของเซลล์เจ้าบ้านเมื่อไวรัสเข้าสู่เซลล์[ 21 ] SARSr-CoV คล้ายกับโคโรนาไวรัสอื่นๆ ตรงที่การแสดงออกของจีโนมเริ่มต้นด้วยการแปลโดยไรโบโซมของเซลล์เจ้าบ้านของกรอบการอ่านแบบเปิด (ORF) ขนาดใหญ่สองอันแรกที่ทับซ้อนกัน คือ 1a และ 1b ซึ่งทั้งสองอันสร้างโพลีโปรตีน[ 19 ]
| หน้าที่ของโปรตีนในจีโนมของไวรัส SARSr-CoV | |
|---|---|
| โปรตีน | ฟังก์ชัน[ 22 ] [ 23 ] [ 24 ] [ 25 ] |
| ORF1ab P0C6X7 | โพลีโปรตีนรีพลิเคส/ทรานสคริปเทส (pp1ab) ( โปรตีนที่ไม่ใช่โครงสร้าง ) |
| ORF2 P59594 | โปรตีน สไปค์ (S) ทำหน้าที่จับและนำไวรัสเข้าสู่เซลล์( โปรตีนโครงสร้าง ) |
| ORF3a P59632 | มีปฏิสัมพันธ์กับโปรตีนโครงสร้าง S, E, M; กระตุ้นการทำงาน ของช่องไอออน ; เพิ่มการสร้างไซโตไคน์และเคโมไคน์เช่นIL-8และRANTES ; เพิ่มการสร้างNF-κBและJNK ; ชักนำให้เกิดอะพอพโทซิสและการหยุดวงจรเซลล์ผ่านทางCaspase 8และ-9และโดยBax , p53และp38 MAP kinase |
| ORF3b P59633 | เพิ่มการ สร้าง ไซโตไคน์และเคโมไคน์โดยผ่านRUNX1b ; ยับยั้ง การผลิตและการส่งสัญญาณของ IFN ชนิดที่ 1 ; ชักนำให้เกิดอะพอพโทซิสและการหยุดชะงักของวงจรเซลล์ ; |
| ORF3c P0DTG1 | ไม่ทราบสาเหตุ; พบครั้งแรกใน SARS-CoV-2 แต่ก็พบใน SARS-CoV ด้วยเช่นกัน |
| ORF3d P0DTG0 | ยีนใหม่ในไวรัส SARS-CoV-2 ซึ่งยังไม่ทราบหน้าที่ |
| ORF4 P59637 | โปรตีน เปลือก (E) การประกอบและการแตกหน่อของไวรัส( โปรตีนโครงสร้าง ) |
| ORF5 P59596 | โปรตีน เยื่อหุ้มเซลล์ (M) การประกอบและการแตกหน่อของไวรัส( โปรตีนโครงสร้าง ) |
| ORF6 P59634 | ช่วยเพิ่มการสังเคราะห์ DNA ในเซลล์; ยับยั้งการผลิตและการส่งสัญญาณของ IFN ชนิดที่ 1 |
| ORF7a P59635 | ยับยั้งการสังเคราะห์โปรตีนในเซลล์; กระตุ้นการอักเสบโดยผ่าน ตัวกระตุ้น NF-kappaBและIL-8 ; เพิ่มระดับสารเคมีชีวภาพ เช่น IL-8 และ RANTES; เพิ่มระดับ JNK และ p38 MAP kinase; กระตุ้นการตายของเซลล์และการหยุดวงจรเซลล์ |
| ORF7b Q7TFA1 | ไม่ทราบ |
| ORF8a Q7TFA0 | กระตุ้นการเกิดอะพอพโทซิสผ่านทางวิถี ไมโทคอนเดรีย |
| ORF8b Q80H93 | ช่วยเพิ่มการสังเคราะห์ DNA ในเซลล์ หรือที่รู้จักกันในชื่อ X5 |
| ORF9a P59595 | โปรตีน นิวคลีโอแคปซิด (N) โปรตีนที่บรรจุ RNA ของไวรัส( โปรตีนโครงสร้าง ) |
| ORF9b P59636 | กระตุ้นให้เกิดอะพอพโทซิส |
| ORF9c Q7TLC7 | รู้จักกันในชื่อ ORF14; หน้าที่ยังไม่ทราบแน่ชัดและอาจไม่ใช่ส่วนที่เข้ารหัสโปรตีน |
| ORF10 A0A663DJA2 | ยีนใหม่ใน SARS-CoV-2 ซึ่งมีหน้าที่ไม่ทราบแน่ชัด อาจไม่ใช่ยีนที่สร้างโปรตีน |
| รหัส UniProtที่แสดงเป็นรหัสสำหรับ โปรตีนของ SARS-CoVยกเว้นรหัสที่เฉพาะเจาะจงสำหรับ SARS-CoV-2 | |
หน้าที่ของโปรตีนไวรัสหลายชนิดเป็นที่ทราบกันดีอยู่แล้ว[ 26 ] ORF 1a และ 1b เข้ารหัสโพลีโปรตีนรีพลิเคส/ทรานสคริปเทส และ ORF 2, 4, 5 และ 9a ในภายหลังเข้ารหัสโปรตีนโครงสร้างหลักสี่ชนิดตามลำดับ ได้แก่สไปค์ (S), เอนเวลอป (E), เมม เบรน (M) และนิวคลีโอแคปซิด (N) [ 27 ] ORF ในภายหลังยังเข้ารหัสโปรตีนที่ไม่ซ้ำกันแปดชนิด (orf3a ถึง orf9b) ซึ่งรู้จักกันในชื่อโปรตีนเสริมหลายชนิดไม่มีโฮโมล็อกที่รู้จัก หน้าที่ต่างๆ ของโปรตีนเสริมยังไม่เป็นที่เข้าใจดีนัก[ 26 ]
ไวรัสโคโรนา SARS ได้รับการดัดแปลงพันธุกรรมในห้องปฏิบัติการหลายแห่ง[ 28 ]
วิวัฒนาการทางสายพันธุ์

การวิเคราะห์ทางวิวัฒนาการแสดงให้เห็นว่าสาขาวิวัฒนาการที่ประกอบด้วยไวรัสโคโรนาค้างคาว BtKY72 และ BM48-31 เป็นกลุ่มฐานของต้นไม้วิวัฒนาการของ CoV ที่เกี่ยวข้องกับ SARS ซึ่งแยกออกจาก CoV ที่เกี่ยวข้องกับ SARS อื่นๆ ก่อน SARS-CoV-1 และ SARS-CoV-2 [ 29 ] [ 3 ]
| SARSr-CoV |
| ||||||||||||||||||||||||||||||||||||||||||
ที่เกี่ยวข้องกับ SARS-CoV-1
แผนภูมิวิวัฒนาการทางสายพันธุ์ที่สร้างขึ้นจากลำดับจีโนมทั้งหมดของ SARS-CoV-1 และไวรัสโคโรนาที่เกี่ยวข้อง มีดังนี้:
ที่เกี่ยวข้องกับ SARS-CoV-2
แผนภูมิวิวัฒนาการตามลำดับจีโนมทั้งหมดของ SARS-CoV-2 และไวรัสโคโรนาที่เกี่ยวข้องคือ: [ 39 ] [ 40 ]
| ไวรัสโคโรนาที่เกี่ยวข้องกับ SARS-CoV-2 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
SARS-CoV-1 , 79% ถึง SARS-CoV-2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
สัณฐานวิทยา
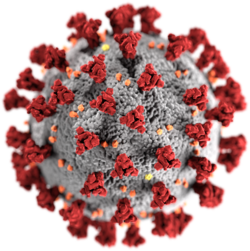
ลักษณะทางสัณฐานวิทยาของไวรัสโคโรนาที่เกี่ยวข้องกับ SARS มีลักษณะเฉพาะของตระกูลไวรัสโคโรนาโดยรวม ไวรัสเป็นอนุภาคทรงกลมขนาดใหญ่ที่มีรูปร่างหลากหลายมีส่วนยื่นบนพื้นผิวที่โป่งพองซึ่งก่อตัวเป็นมงกุฎรอบอนุภาคในภาพถ่ายอิเล็กตรอนไมโครสโคป[ 50 ]ขนาดของอนุภาคไวรัสอยู่ในช่วง 80–90 นาโนเมตร เปลือกของไวรัสในภาพถ่ายอิเล็กตรอนไมโครสโคปปรากฏเป็นเปลือกที่มีความหนาแน่นของอิเล็กตรอนคู่หนึ่งที่ชัดเจน[ 51 ]
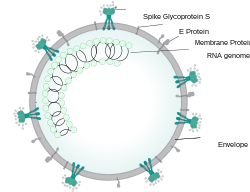
เปลือกหุ้มไวรัสประกอบด้วยชั้นไขมันสองชั้นซึ่งโปรตีนเมมเบรน (M) โปรตีน เปลือกหุ้ม (E) และ โปรตีน หนาม (S) ยึดติดอยู่[ 52 ]โปรตีนหนามทำให้ไวรัสมีส่วนยื่นบนพื้นผิวที่โป่งพอง ซึ่งเรียกว่าเปปโลเมอ ร์ การโต้ตอบของโปรตีนหนามกับตัวรับเซลล์โฮสต์ คอมพลีเมนต์ มีความสำคัญในการกำหนดความชอบเนื้อเยื่อ ความสามารถในการติดเชื้อและช่วงสายพันธุ์ของไวรัส[ 53 ] [ 54 ]
ภายในซองหุ้มจะมีนิวคลีโอแคปซิดซึ่งเกิดจาก โปรตีน นิวคลีโอแคปซิด (N) หลายสำเนาที่ยึดติดกับจีโนม RNA สายเดี่ยวแบบบวก (~30 kb ) ใน รูปแบบลูกปัด เรียงต่อกันเป็นเส้น [ 55 ] [ 56 ]ซองหุ้มไขมันสองชั้น โปรตีนเยื่อหุ้มเซลล์ และนิวคลีโอแคปซิดจะปกป้องไวรัสเมื่ออยู่นอกร่างกายโฮสต์[ 57 ]
วงจรชีวิต
ไวรัสโคโรนาที่เกี่ยวข้องกับ SARS ใช้กลยุทธ์การจำลองแบบทั่วไปของไวรัสโคโรนาทั้งหมด[ 20 ] [ 58 ]
เอกสารแนบและรายการ

การยึดเกาะของไวรัสโคโรนาที่เกี่ยวข้องกับ SARS กับเซลล์โฮสต์นั้นเกิดขึ้นโดยอาศัยโปรตีนหนามและตัวรับของมัน[ 59 ]โดเมนการจับตัวรับของโปรตีนหนาม (RBD) จะจดจำและยึดเกาะกับ ตัวรับ เอนไซม์แองจิโอเทนซินคอนเวอร์ติง 2 (ACE2) [ 8 ]หลังจากการยึดเกาะ ไวรัสสามารถเข้าสู่เซลล์โฮสต์ได้สองเส้นทาง เส้นทางที่ไวรัสใช้ขึ้นอยู่กับโปรตีเอส ของโฮสต์ ที่มีอยู่เพื่อตัดและกระตุ้นโปรตีนหนามที่ยึดติดกับตัวรับ[ 60 ]ความแตกต่างที่สำคัญระหว่าง SARS-CoV-1 และ SARS-CoV-2 คือ SARS-CoV-2 ถูกตัดล่วงหน้าแล้วเนื่องจากมีไซต์การตัดฟูริน[ 61 ]
การยึดเกาะของซาร์เบโคไวรัสกับ ACE2 ได้รับการพิสูจน์แล้วว่าเป็น ลักษณะ ที่ได้รับการอนุรักษ์ทางวิวัฒนาการซึ่งมีอยู่ในหลายสปีชีส์ (แต่ไม่ใช่ทั้งหมด) ของกลุ่มอนุกรมวิธานที่มี ACE2 โดยใช้ตัวแทนในแอฟริกา เอเชีย และยุโรป[ 62 ]
เส้นทางแรกที่ไวรัสโคโรนา SARS สามารถใช้ในการเข้าสู่เซลล์โฮสต์คือโดยเอนโดไซโทซิสและการดูดซึมไวรัสในเอนโดโซม จากนั้นโปรตีนหนามที่ติดอยู่กับตัวรับจะถูกกระตุ้นโดย เอนไซม์โปรตีเอสซิสทีน ที่ขึ้นอยู่กับ pH ของโฮสต์ที่เรียกว่าแคทเทปซิน Lการกระตุ้นโปรตีนหนามที่ติดอยู่กับตัวรับทำให้เกิดการเปลี่ยนแปลงโครงสร้างและตามมาด้วยการหลอมรวมของซองไวรัสกับผนังเอนโดโซม[ 60 ]
อีกทางเลือกหนึ่ง ไวรัสสามารถเข้าสู่เซลล์โฮสต์โดยตรงโดย การ ตัดโปรตีนหนามที่ติดอยู่กับตัวรับด้วยเอนไซม์โปรตีเอสเซรินTMPRSS2หรือTMPRSS11D ของโฮสต์ ที่ผิวเซลล์[ 63 ] [ 64 ]ในไวรัสโคโรนา SARS การกระตุ้นส่วนปลาย Cของโปรตีนหนามจะกระตุ้นการหลอมรวมของซองไวรัสกับเยื่อหุ้มเซลล์โฮสต์โดยการเหนี่ยวนำให้เกิดการเปลี่ยนแปลงโครงสร้างซึ่งยังไม่เป็นที่เข้าใจอย่างสมบูรณ์[ 65 ]
การแปลจีโนม
| หน้าที่ของโปรตีนที่ไม่ใช่โครงสร้างของไวรัสโคโรนา (nsps) [ 66 ] | |
|---|---|
| โปรตีน | การทำงาน |
| เอ็นเอสพี1 | ส่งเสริม การย่อยสลาย mRNA ของโฮสต์ ขัดขวางการแปลของโฮสต์[ 67 ]ขัดขวางการ ตอบสนองภูมิคุ้มกันโดยกำเนิด |
| เอ็นเอสพี2 | จับกับ โปรตีนโปร ฮิบิติน ; หน้าที่ยังไม่ทราบแน่ชัด |
| เอ็นเอสพี3 | โปรตีนทรานส์เมมเบรนแบบหลายโดเมน ; ทำปฏิกิริยากับโปรตีน N ; ส่งเสริม การแสดงออก ของไซโตไคน์ ; โดเมนPLPro ตัดโพลี โปรตีน pp1ab และปิดกั้นการตอบสนองภูมิคุ้มกันโดยกำเนิดของโฮสต์; โดเมนอื่นๆ ยังไม่ทราบหน้าที่ |
| เอ็นเอสพี4 | โปรตีนโครงสร้างทรานส์เมมเบรน; ช่วยให้ เวสิเคิลแบบเยื่อหุ้มสองชั้น (DMVs) มีโครงสร้างที่เหมาะสม |
| เอ็นเอสพี5 | 3CLProทำหน้าที่ตัดโพลีโปรตีน pp1ab |
| เอ็นเอสพี6 | โปรตีนโครงสร้างทรานส์เมมเบรน; หน้าที่ยังไม่ทราบแน่ชัด |
| เอ็นเอสพี7 | สร้าง คอมเพล็กซ์ เฮกซาเดคาเมอริกกับ nsp8; ตัวควบคุมกระบวนการทำงานของRdRp (nsp12) |
| เอ็นเอสพี8 | สร้างคอมเพล็กซ์เฮกซาเดคาเมอริกกับ nsp7; ทำหน้าที่เป็นแคลมป์ควบคุมกระบวนการทำงานของ RdRp (nsp12); และทำหน้าที่เป็นไพรเมส |
| เอ็นเอสพี9 | โปรตีนจับอาร์เอ็นเอ (RBP) |
| เอ็นเอสพี10 | โคแฟคเตอร์ของ nsp16 และ nsp14 ; สร้างเฮเทอโรไดเมอร์กับทั้งสองตัว; กระตุ้น การทำงานของ 2-O-MT (nsp16) และExoN (nsp14) |
| เอ็นเอสพี11 | ฟังก์ชันที่ไม่รู้จัก |
| เอ็นเอสพี12 | เอนไซม์ RNA-dependent RNA polymerase (RdRp) |
| เอ็นเอสพี13 | อาร์เอ็นเอ เฮลิเคส , 5′ ไตรฟอสเฟต |
| เอ็นเอสพี14 | N7 เมทิลทรานสเฟอเรส , 3′-5′ เอ็กโซไรโบนิวคลีเอส (ExoN); N7 MTase เพิ่มหมวก 5′ , ExoN ตรวจสอบความถูกต้องของจีโนม |
| เอ็นเอสพี15 | เอนโดไรโบนิวคลีเอส (NendoU) |
| เอ็นเอสพี16 | 2′-O-เมทิลทรานสเฟอเรส (2-O-MT); ปกป้อง RNA ของไวรัสจากMDA5 |
หลังจากเกิดการหลอมรวม นิวคลีโอแคปซิดจะผ่านเข้าไปในไซโตพลาสซึมซึ่งจีโนมของไวรัสจะถูกปล่อยออกมา[ 59 ]จีโนมทำหน้าที่เป็นอาร์เอ็นเอส่งสารและไรโบโซมของเซลล์จะแปลจีโนมสองในสามส่วน ซึ่งสอดคล้องกับกรอบการอ่านแบบเปิดORF1aและORF1bให้เป็นโพลีโปรตีนขนาดใหญ่สองชนิดที่ทับซ้อนกัน คือ pp1a และ pp1ab
โพลีโปรตีนขนาดใหญ่ pp1ab เป็นผลมาจากการเลื่อนเฟรมของไรโบโซม -1ที่เกิดจากลำดับลื่น (UUUAAAC) และปมเทียม RNA ที่ อยู่ปลายสุดของเฟรมการอ่านแบบเปิด ORF1a [ 68 ]การเลื่อนเฟรมของไรโบโซมทำให้การแปล ORF1a ต่อเนื่องตามด้วย ORF1b [ 69 ]
โพ ลี โปรตีนประกอบด้วย โปรตีเอสของตัวเองPLpro และ3CLproซึ่งตัดโพลีโปรตีนที่ไซต์เฉพาะต่างๆ การตัดโพลีโปรตีน pp1ab ทำให้เกิดโปรตีนที่ไม่ใช่โครงสร้าง 16 ชนิด (nsp1 ถึง nsp16) โปรตีนที่เป็นผลิตภัณฑ์ประกอบด้วยโปรตีนการจำลองแบบต่างๆ เช่นRNA-dependent RNA polymerase (RdRp), RNA helicaseและexoribonuclease (ExoN) [ 69 ]
โปรตีเอส SARS-CoV-2 สองตัว (PLpro และ 3CLpro) ยังรบกวนการตอบสนองของระบบภูมิคุ้มกันต่อการติดเชื้อไวรัสโดยการตัดโปรตีนของระบบภูมิคุ้มกันสามตัว PLpro ตัดIRF3และ 3CLpro ตัดทั้งNLRP12และTAB1 "การตัด IRF3 โดยตรงโดย NSP3 อาจอธิบายการตอบสนองของ IFN ชนิดที่ 1 ที่ลดลงซึ่งพบในระหว่างการติดเชื้อ SARS-CoV-2 ในขณะที่การตัด NLRP12 และ TAB1 โดย NSP5 ชี้ให้เห็นถึงกลไกระดับโมเลกุลสำหรับการผลิต IL-6 ที่เพิ่มขึ้นและการตอบสนองการอักเสบที่พบในผู้ป่วย COVID-19" [ 70 ]
การจำลองแบบและการถอดรหัส
โปรตีนการจำลองแบบที่ไม่ใช่โครงสร้างจำนวนหนึ่งรวมตัวกันเพื่อสร้าง คอมเพล็กซ์ โปรตีนจำลองแบบ -ถอดรหัส (RTC) [ 69 ]โปรตีนจำลองแบบ-ถอดรหัสหลักคือRNA-dependent RNA polymerase (RdRp) ซึ่งเกี่ยวข้องโดยตรงกับการจำลองแบบและการถอดรหัสของ RNA จากสาย RNA โปรตีนที่ไม่ใช่โครงสร้างอื่นๆ ในคอมเพล็กซ์จะช่วยในกระบวนการจำลองแบบและการถอดรหัส[ 66 ]
โปรตีน nsp14 เป็นเอ็กโซไรโบนิวคลีเอส 3'-5'ซึ่งให้ความแม่นยำเป็นพิเศษแก่กระบวนการจำลองแบบ เอ็กโซไรโบนิวคลีเอสทำ หน้าที่ ตรวจสอบความถูกต้องให้กับคอมเพล็กซ์ ซึ่งเอนไซม์ RNA-dependent RNA polymerase ขาดไป ในทำนองเดียวกัน โปรตีน nsp7 และ nsp8 ก่อตัวเป็นแคลมป์เลื่อนเฮกซาเดคาเมอริกซึ่งเป็นส่วนหนึ่งของคอมเพล็กซ์ ซึ่งช่วยเพิ่มประสิทธิภาพของเอนไซม์ RNA-dependent RNA polymerase อย่างมาก [ 66 ]ไวรัสโคโรนาต้องการความแม่นยำและประสิทธิภาพที่เพิ่มขึ้นในระหว่างการสังเคราะห์ RNA เนื่องจากขนาดจีโนมที่ค่อนข้างใหญ่เมื่อเทียบกับไวรัส RNA อื่นๆ[ 71 ]
หนึ่งในหน้าที่หลักของคอมเพล็กซ์รีพลิเคส-ทรานสคริปเทสคือการถอดรหัสจีโนมของไวรัส RdRp ทำหน้าที่สังเคราะห์ โมเลกุล RNA ย่อยจีโนมแบบลบ จาก RNA จีโนมแบบบวกโดยตรง จากนั้นจึงทำการถอดรหัสโมเลกุล RNA ย่อยจีโนมแบบลบเหล่านี้ไป เป็นmRNAแบบบวกที่สอดคล้องกัน[ 72 ]
หน้าที่สำคัญอีกประการหนึ่งของคอมเพล็กซ์รีพลิเคส-ทรานสคริปเทสคือการจำลองจีโนมของไวรัส RdRp ทำหน้าที่สังเคราะห์ RNA จีโนมแบบลบจาก RNA จีโนมแบบบวกโดยตรง ตามด้วยการจำลอง RNA จีโนมแบบบวกจาก RNA จีโนมแบบลบ[ 72 ] การจำลองและการถอดรหัสของ SARS-CoV ส่วนใหญ่เกิดขึ้นในเวสิเคิลเยื่อหุ้มสองชั้น (DMVs) ที่เกิดจากไวรัส ซึ่งสร้างจาก เอนโดพลาสมิกเรติคูลัมของโฮสต์ที่เปลี่ยนแปลงไป[ 73 ]
RNA จีโนมแบบบวกที่จำลองแบบจะกลายเป็นจีโนมของไวรัสรุ่นลูก mRNA ขนาดเล็กต่างๆ เป็นการถอดรหัสจากหนึ่งในสามส่วนสุดท้ายของจีโนมไวรัสซึ่งเป็นไปตามเฟรมการอ่าน ORF1a และ ORF1b mRNA เหล่านี้จะถูกแปลเป็นโปรตีนโครงสร้างสี่ชนิด (S, E, M และ N) ซึ่งจะกลายเป็นส่วนหนึ่งของอนุภาคไวรัสรุ่นลูก และโปรตีนเสริมอื่นๆ อีกแปดชนิด (orf3 ถึง orf9b) ซึ่งช่วยไวรัส[ 74 ]
การรวมตัวใหม่
เมื่อ มี จีโนม SARS-CoV สองตัว อยู่ในเซลล์โฮสต์ พวกมันอาจโต้ตอบกันเพื่อสร้างจีโนมลูกผสมที่สามารถส่งต่อไปยังไวรัสรุ่นลูกได้ การรวมตัวใหม่น่าจะเกิดขึ้นระหว่างการจำลองจีโนมเมื่อRNA polymeraseเปลี่ยนจากแม่แบบหนึ่งไปเป็นอีกแม่แบบหนึ่ง (การรวมตัวใหม่แบบเลือกสำเนา) [ 75 ] ดูเหมือนว่า SARS-CoV ในมนุษย์จะมีประวัติการรวมตัว ใหม่ที่ซับซ้อน ระหว่างโคโรนาไวรัส บรรพบุรุษ ที่อาศัยอยู่ในกลุ่มสัตว์หลายกลุ่มที่แตกต่างกัน[ 75 ] [ 76 ]
การประกอบและการปล่อย
การแปล RNAเกิดขึ้นภายในเอนโดพลาสมิกเรติคูลัมโปรตีนโครงสร้างของไวรัส S, E และ M เคลื่อนที่ไปตามเส้นทางการหลั่งเข้าสู่ช่องกลางของกอลจิที่นั่น โปรตีน M จะควบคุมปฏิสัมพันธ์ระหว่างโปรตีนส่วนใหญ่ที่จำเป็นสำหรับการประกอบไวรัสหลังจากที่มันจับกับนิวคลีโอแคปซิด[ 77 ]ไวรัสลูกหลานจะถูกปล่อยออกจากเซลล์โฮสต์โดยการหลั่งออกนอกเซลล์ผ่านถุงหลั่ง[ 77 ]
ดูเพิ่มเติม
- โคโรนาไวรัสคล้ายค้างคาว SARS WIV1 (SL-CoV-WIV1)
- ไวรัสโคโรนาสายพันธุ์คล้าย SARS จากค้างคาว RSSHC014
- ไวรัสโคโรนาสายพันธุ์ค้างคาว RaTG13
- ไวรัส SARS-CoV ในชะมด
หมายเหตุ
- ^ บางครั้งมีการใช้ คำว่า SARS-CoVและ SARS-CoVสลับกัน โดยเฉพาะอย่างยิ่งก่อนการค้นพบ SARS-CoV-2 ซึ่งอาจทำให้เกิดความสับสนเมื่อบางสิ่งพิมพ์อ้างถึง SARS-CoV-1 ว่าเป็น SARS- CoV
อ่านเพิ่มเติม
- Peiris JS, Lai ST, Poon LL, Guan Y, Yam LY, Lim W และคณะ (เมษายน 2546). "ไวรัสโคโรนาเป็นสาเหตุที่เป็นไปได้ของโรคทางเดินหายใจเฉียบพลันรุนแรง" . Lancet . 361 (9366): 1319– 25. Bibcode : 2003Lanc..361.1319P . doi : 10.1016/s0140-6736(03)13077-2 . PMC 7112372 . PMID 12711465 .
- Rota PA, Oberste MS, Monroe SS, Nix WA, Campagnoli R, Icenogle JP และคณะ (พฤษภาคม 2546). "การจำแนกลักษณะของไวรัสโคโรนาสายพันธุ์ใหม่ที่เกี่ยวข้องกับโรคทางเดินหายใจเฉียบพลันรุนแรง" Science . 300 ( 5624): 1394– 9. Bibcode : 2003Sci...300.1394R . doi : 10.1126/science.1085952 . hdl : 1765/3917 . PMID 12730500 .
- Marra MA, Jones SJ, Astell CR, Holt RA, Brooks-Wilson A, Butterfield YS และคณะ (พฤษภาคม 2546). "ลำดับจีโนมของไวรัสโคโรนาที่เกี่ยวข้องกับ SARS" Science . 300 ( 5624): 1399– 404. Bibcode : 2003Sci...300.1399M . doi : 10.1126/science.1085953 . PMID 12730501 .
- Snijder EJ, Bredenbeek PJ, Dobbe JC, Thiel V, Ziebuhr J, Poon LL และคณะ (สิงหาคม 2546) "ลักษณะเฉพาะและอนุรักษ์ไว้ของจีโนมและโปรตีโอมของ SARS-coronavirus ซึ่งแยกตัวออกมาในช่วงต้นจากสายพันธุ์โคโรนาไวรัสกลุ่ม 2"วารสารชีววิทยาโมเลกุล 331 ( 5): 991– 1004. CiteSeerX 10.1.1.319.7007 . doi : 10.1016/ S0022-2836 (03) 00865-9 . PMC 7159028. PMID 12927536. S2CID 14974326 .
- Yount B, Roberts RS, Lindesmith L, Baric RS (สิงหาคม 2549). "การปรับวงจรการถอดรหัสของไวรัสโคโรนาที่ก่อให้เกิดโรคทางเดินหายใจเฉียบพลันรุนแรง (SARS-CoV): การออกแบบจีโนมที่ต้านทานการรวมตัวใหม่" Proceedings of the National Academy of Sciences of the United States of America . 103 (33): 12546– 51. Bibcode : 2006PNAS..10312546Y . doi : 10.1073/pnas.0605438103 . PMC 1531645. PMID 16891412 .
- Thiel V, บรรณาธิการ (2007). ไวรัสโคโรนา: ชีววิทยาระดับโมเลกุลและเซลล์ (ฉบับพิมพ์ครั้งที่ 1). สำนักพิมพ์ Caister Academic Press. ISBN 978-1-904455-16-5.
- Enjuanes L, Sola I, Zúñiga S, Almazán F (2008). "การจำลองแบบและการโต้ตอบของไวรัสโคโรนากับโฮสต์" ใน Mettenleiter TC, Sobrino F (บรรณาธิการ). ไวรัสในสัตว์: ชีววิทยาระดับโมเลกุล . สำนักพิมพ์ Caister Academic Press. ISBN 978-1-904455-22-6.
ลิงก์ภายนอก
 สื่อที่เกี่ยวข้องกับการระบาดใหญ่ของไวรัสโคโรนาเบตาในวิกิมีเดียคอมมอนส์
สื่อที่เกี่ยวข้องกับการระบาดใหญ่ของไวรัสโคโรนาเบตาในวิกิมีเดียคอมมอนส์ ข้อมูลที่เกี่ยวข้องกับไวรัสโคโรนาสายพันธุ์ SARSใน Wikispecies
ข้อมูลที่เกี่ยวข้องกับไวรัสโคโรนาสายพันธุ์ SARSใน Wikispecies- แถลงการณ์ขององค์การอนามัยโลกเกี่ยวกับการระบุและตั้งชื่อไวรัส SARS (เก็บถาวรเมื่อวันที่ 23 เมษายน 2546)
- แผนที่พันธุกรรมของไวรัส SARS ถูกเก็บถาวรไว้เมื่อวันที่ 18 สิงหาคม 2549 ที่Wayback Machine
- บทความพิเศษด้าน วิทยาศาสตร์เกี่ยวกับไวรัส SARS (เนื้อหาฟรี: ไม่ต้องลงทะเบียน)
- แหล่งข้อมูลเกี่ยวกับโรคซาร์สของมหาวิทยาลัยแมคกิลล์ในWayback Machine (เก็บถาวรเมื่อวันที่ 1 มีนาคม 2548)
- ศูนย์ควบคุมและป้องกันโรคแห่งสหรัฐอเมริกา (CDC) หน้าหลักเกี่ยวกับโรคซาร์ส (เก็บถาวรเมื่อ 12 เมษายน 2559)
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ไวรัสโคโรนาที่เกี่ยวข้องกับ SARS
ไวรัสโคโรนาที่เกี่ยวข้องกับโรคทางเดินหายใจเฉียบพลันรุนแรง ( SARSr-CoVหรือSARS-CoV , Betacoronavirus pandemicum ) เป็นไวรัสชนิด หนึ่งที่ประกอบด้วยสายพันธุ์ที่รู้จักกันหลาย...
การจำแนกประเภท
ไวรัสโคโรนาที่เกี่ยวข้องกับ SARS เป็นสมาชิกของสกุล Betacoronavirus (กลุ่ม 2) และ เป็นโมโนไทป์ ของสกุลย่อย Sarbecovirus (กลุ่มย่อย B) [ 13 ] Sarbecoviruses แตกต่างจาก embecoviruses หรือ alphacoronaviruses ตรงที่มี โปรตีเอสคล้ายปาเปน (PLpro)...
จีโนม
ไวรัสโคโรนา ที่เกี่ยวข้องกับ SARS เป็น ไวรัส RNA สายเดี่ยวที่มีเปลือกหุ้มและมีทิศทางบวก จีโนมของมันมีขนาดประมาณ 30 กิโลเบส ซึ่งเป็นหนึ่งในจีโนมที่ใหญ่ที่สุดในบรรดาไวรัส RNA ไวรัสนี้มี กรอบการอ่านแบบเปิด 14 เฟรม ซึ่งทับซ้อนกันในบางกรณี [ 19 ] จีโนมมี...
วิวัฒนาการทางสายพันธุ์
การวิเคราะห์ทางวิวัฒนาการ แสดงให้เห็นว่าสาขาวิวัฒนาการที่ประกอบด้วยไวรัสโคโรนาค้างคาว BtKY72 และ BM48-31 เป็นกลุ่มฐานของต้นไม้วิวัฒนาการของ CoV ที่เกี่ยวข้องกับ SARS ซึ่งแยกออกจาก CoV ที่เกี่ยวข้องกับ SARS อื่นๆ ก่อน SARS-CoV-1 และ SARS-CoV-2 [ 29 ] [ 3 ]