อ่าน 22 นาที
เซกโนซอรัส
เซกโนซอรัส (Segnosaurus)เป็นสกุลของไดโนเสาร์เทอริซิโนซอริเดีย(Therizinosauridae ) ที่อาศัยอยู่ในบริเวณที่เป็นประเทศมองโกเลียตะวันออกเฉียงใต้ในปัจจุบัน...
เซกโนซอรัส
| เซกโนซอรัส ช่วงเวลา: ปลายยุคครีเทเชียส | |
|---|---|
 | |
| แผนภาพแสดงซากที่ทราบแล้ว | |
| การจำแนกทางวิทยาศาสตร์ | |
| อาณาจักร: | แอนิมอลเลีย |
| ไฟลัม: | คอร์ดาต้า |
| ระดับ: | สัตว์เลื้อยคลาน |
| กลุ่มสายพันธุ์ : | ไดโนเสาร์ |
| กลุ่มสายพันธุ์ : | ซอริสเชีย |
| กลุ่มสายพันธุ์ : | เทโรโปดา |
| ตระกูล: | † เทริซิโนซอริเด |
| ประเภท: | † เซกโนซอรัสเพิร์ล , 1979 |
| สายพันธุ์: | † S. galbinensis |
| ชื่อทวินาม | |
| † เซกโนซอรัส กัลบินเนนซิส เพิร์ล, 1979 | |
เซกโนซอรัส (Segnosaurus)เป็นสกุลของไดโนเสาร์เทอริซิโนซอริเดีย(Therizinosauridae ) ที่อาศัยอยู่ในบริเวณที่เป็นประเทศมองโกเลียตะวันออกเฉียงใต้ในปัจจุบัน ในช่วงปลายยุคครีเทเชียสเมื่อประมาณ 102–86 ล้านปีก่อน มีการค้นพบซากดึกดำบรรพ์ที่ไม่สมบูรณ์แต่ได้รับการอนุรักษ์ไว้อย่างดีหลายชิ้นในทะเลทรายโกบีในช่วงทศวรรษ 1970 และในปี 1979 สกุลและชนิด Segnosaurus galbinensisได้รับการตั้งชื่อ ชื่อสกุล Segnosaurusหมายถึง "กิ้งก่าเคลื่อนไหวช้า" และชื่อชนิด galbinensisหมายถึงภูมิภาคกัลบิน (Galbin) ซากดึกดำบรรพ์ที่ค้นพบของไดโนเสาร์ชนิดนี้ ได้แก่ ขากรรล่าง กระดูกสันหลังส่วนคอและหาง กระดูกเชิงกราน กระดูกหัวไหล่ และกระดูกแขนขา บางส่วนของซากดึกดำบรรพ์ได้หายไปหรือเสียหายไปนับตั้งแต่มีการเก็บรวบรวม ซากดึกดำบรรพ์จากประเทศญี่ปุ่นที่เดิมจัดอยู่ในสกุล Allosaurusได้ถูกจัดใหม่ให้อยู่ในสกุล Segnosaurus sp. (ชนิดที่ไม่ระบุ) ในปี 2025
เซกโนซอรัส เป็น เทอริซิโนซอร์ขนาดใหญ่คาดว่ามีความยาวประมาณ 6–7 เมตร (20–23 ฟุต) และหนักประมาณ 1.3 ตัน (1.4 ตันสั้น) มันเดินสองขา โดยลำตัวเอียงขึ้นด้านบน หัวเล็ก มีจงอยปากอยู่ที่ปลายขากรรไกร และคอยาวและเรียว ขากรรไกรล่างง้างลงด้านหน้า และฟันมีลักษณะเด่นคือมีเดนติเคิล เพิ่มเติม รวมถึงคมตัดที่สามในฟันซี่หลังสุดบางซี่ แขนขาหน้าแข็งแรงและมีสามนิ้วซึ่งมีกรงเล็บขนาดใหญ่ และเท้ามีสี่นิ้วเพื่อรองรับเท้า—ยกเว้นเทอริซิโนซอร์แล้วเทอโรพอด ทั้งหมด มีเท้าสามนิ้ว กระดูกเชิงกรานส่วนหน้าปรับตัวเพื่อรองรับท้องที่ขยายใหญ่ขึ้นกระดูกหัวหน่าวหันไปด้านหลัง ซึ่งเป็นลักษณะที่พบได้เฉพาะในนกและไดโนเสาร์ที่ใกล้เคียงกับนกมากที่สุด
เดิมที ความสัมพันธ์ของSegnosaurusนั้นไม่ชัดเจน และมันได้รับวงศ์เทอโรพอดของตัวเอง คือ Segnosauridae และต่อมาเมื่อมีการระบุสกุลที่เกี่ยวข้องได้ จึงได้จัดเป็นอันดับย่อยคือ Segnosauria มีการเสนอแผนการจัดจำแนกทางเลือกอื่นๆ จนกระทั่งมีการอธิบายญาติที่สมบูรณ์มากขึ้นในทศวรรษ 1990 ซึ่งยืนยันว่าพวกมันเป็นเทอโรพอด ฟอสซิลใหม่ยังแสดงให้เห็นว่า Segnosauridae เป็นชื่อพ้องรองของวงศ์ Therizinosauridae ที่ตั้งชื่อไว้ก่อนหน้านี้ เชื่อกันว่า Segnosaurusและญาติของมันเป็นสัตว์ที่เคลื่อนไหวช้า ซึ่งจากลักษณะที่ผิดปกติของพวกมัน บ่งชี้ว่าพวกมันกินพืชเป็นหลัก ในขณะที่กลุ่มเทอโรพอดอื่นๆ ส่วนใหญ่เป็นสัตว์กินเนื้อ เทอริซิโนซอร์น่าจะใช้แขนหน้ายาว คอยาว และจะงอยปากในการหากินและใช้ลำไส้ขนาดใหญ่ในการย่อยอาหารSegnosaurusพบในแหล่ง ฟอสซิล Bayan Shireh Formationซึ่งมันอาศัยอยู่ร่วมกับเทอริซิโนซอร์อื่นๆ เช่นErlikosaurusและEnigmosaurusสกุลที่เกี่ยวข้องเหล่านี้อาจมีการแบ่งส่วนนิเวศวิทยา
ประวัติการค้นพบ

ในปี 1973 คณะสำรวจร่วมระหว่างโซเวียตและมองโกเลียที่สำรวจชั้นหินบายันชีเรห์ณ บริเวณอัมต์กาย ในทะเลทรายโกบี ทาง ตะวันออกเฉียงใต้ของมองโกเลีย ได้ค้นพบฟอสซิลซึ่งรวมถึงโครงกระดูกบางส่วนของไดโนเสาร์ ที่ไม่รู้จัก ในช่วงปี 1974 และ 1975 มีการค้นพบซากเพิ่มเติมในบริเวณอัมต์กายและคารา-คูทูล แม้ว่าโครงกระดูกจะไม่สมบูรณ์ แต่กระดูกที่พบนั้นได้รับการอนุรักษ์ไว้อย่างดี บริเวณอื่น ๆ ที่ระบุไว้ในเอกสาร ได้แก่ บายชิน-ซาว และอูริลเบ-คูดุก ฟอสซิลเหล่านี้ได้รับการอธิบาย ทางวิทยาศาสตร์ ในปี 1979 โดยนักบรรพชีวินวิทยาอัลตังเกอเรล เพอร์เลซึ่งตั้งชื่อสกุลและชนิด ใหม่ ว่า เซกโนซอรัส กัลเบียนซิสชื่อสกุลมาจากคำภาษาละตินว่าsegnis ("ช้า") และคำภาษากรีกโบราณว่าsauros ("กิ้งก่า") ชื่อชนิดหมายถึงภูมิภาคกัลบินในทะเลทรายโกบี[ 1 ] [ 2 ] [ 3 ]
ตัวอย่างต้นแบบ (holotype specimen)จากแหล่ง Amtgay ถูกเก็บรักษาไว้ที่สถาบันวิทยาศาสตร์แห่งมองโกเลียภายใต้หมายเลขตัวอย่าง IGM 100/80 (สถาบันธรณีวิทยาแห่งมองโกเลีย เดิมคือ GIN) ตัวอย่างนี้ประกอบด้วยขากรรไกรล่าง ( mandible ), กระดูกต้นแขน ( humerus) ที่ไม่สมบูรณ์, กระดูกปลายแขน (radius และ ulna) ที่สมบูรณ์, กระดูกนิ้วมือ , กระดูกเล็บ (ungual) ของแขนขาหน้า, กระดูกเชิงกรานที่เกือบสมบูรณ์ , กระดูกต้นขาขวาที่ไม่สมบูรณ์ , กระดูกสันหลังส่วนกระเบนเหน็บ 6 ชิ้น, กระดูกสันหลังส่วนหางด้านหน้า 10 ชิ้น, กระดูกสันหลังส่วนหางด้านหลัง 15 ชิ้น, กระดูกซี่โครงส่วน ท้องชิ้นแรก และชิ้นส่วนของกระดูกซี่โครงส่วนหลัง นอกจากนี้ยังมีตัวอย่างอีก 2 ชิ้นที่ถูกกำหนดให้เป็นตัวอย่างรอง (paratype specimens ) ตัวอย่าง IGM 100/82 จากแหล่ง Khara Khutul ประกอบด้วยกระดูกต้นขา กระดูกหน้าแข้งและกระดูกน่อง (กระดูกขา) กระดูกข้อเท้าและ กระดูก ฝ่าเท้ากระดูกนิ้วเท้า 5 ข้อ รวมถึงเล็บเท้า กระดูกซี่โครงกระดูกเชิงกราน ที่สมบูรณ์ ส่วนบนของกระดูกสะโพกและส่วนล่างของกระดูกหัวหน่าวตัวอย่าง IGM 100/83 ประกอบด้วยกระดูกสะบักและกระดูกหัวหน่าวซ้าย กระดูกเรเดียส กระดูกอัลนา เล็บเท้า และชิ้นส่วนของกระดูกสันหลังส่วนคอ[ 1 ] [ 4 ] ในปี 1980 Perle และนักบรรพชีวินวิทยาRinchen Barsboldได้จัดตัวอย่างอีกชิ้นหนึ่งให้กับSegnosaurus ; IGM 100/81 จากแหล่ง Amtgay ประกอบด้วยกระดูกหน้าแข้งและกระดูกน่องซ้าย[ 5 ] [ 4 ]

ในปี 1983 บาร์สโบลด์ได้ระบุตัวอย่างเพิ่มเติมคือ GIN 100/87 และ 100/88 อย่างไรก็ตาม ในปี 2010 นักบรรพชีวินวิทยา ลินด์เซย์ อี. แซนโนได้เสนอว่าตัวอย่างเหล่านี้อาจหมายถึงพาราไทป์ IGM 100/82 และ IGM 100/83 (ซึ่งได้ระบุไว้แล้วในปี 1979) เนื่องจากบทความของบาร์สโบลด์ที่แปลจากภาษารัสเซียเป็นภาษาอังกฤษมีข้อผิดพลาดทางด้านการพิมพ์หลายจุดเกี่ยวกับหมายเลขตัวอย่าง แซนโนยังตั้งข้อสังเกตอีกว่า ในช่วงเวลาที่เธอทำการศึกษา มีปัญหามากมายเกี่ยวกับ ตัวอย่าง Segnosaurus IGM รวมถึงความเสียหายที่เกิดขึ้นหลังจากเก็บรวบรวม การหายไปของชิ้นส่วนของโฮโลไทป์ การระบุชิ้นส่วนที่กำหนดไม่ถูกต้อง และมีมากกว่าหนึ่งตัวอย่างที่มีหมายเลขตัวอย่างเดียวกัน องค์ประกอบโฮโลไทป์ที่ Zanno สามารถเข้าถึงได้ในปี 2010 ได้แก่ กระดูกเชิงกรานที่เสียหายอย่างรุนแรง กระดูกศักรัมที่ขาดซี่โครงศักรัมด้านซ้ายเนื่องจากความเสียหายทำให้ไม่สามารถเชื่อมต่อกับกระดูกเชิงกรานส่วนที่เหลือได้ดี และกระดูกหัวหน่าวและกระดูกอิสเคียมที่ขาดส่วนบน กระดูกเพิ่มเติมที่มีหมายเลขตัวอย่าง IGM 100/82 ถูกค้นพบแต่ไม่ได้กล่าวถึงในคำอธิบายของ Perle ในขณะที่ตำแหน่งขององค์ประกอบพาราไทป์บางส่วนไม่เป็นที่ทราบ[ 3 ] [ 4 ]ในการบรรยายลักษณะใหม่ของขากรรไกรล่างโฮโลไทป์ในปี 2016 ซึ่งแทบไม่ได้รับการศึกษาเลยตั้งแต่ปี 1979 Zanno และเพื่อนร่วมงานรายงานว่าส่วนยอดฟันส่วนใหญ่เสียหายหลังจากการเก็บรวบรวม และส่วนใหญ่ขาดปลายฟัน ในบรรดาขากรรไกรล่างสองซีก (ครึ่งของขากรรไกรล่าง) ซีกขวาเกือบสมบูรณ์ มี เพียงส่วนท้ายสุดและส่วนหน้าด้านบนของกระดูกขากรรไกรล่าง (บริเวณที่กระดูกขากรรไกรล่างทั้งสองข้างมาบรรจบกัน) เท่านั้นที่หายไป กระดูกขากรรไกรล่างซีกซ้ายแตกเป็นชิ้นเล็กชิ้นน้อยและยังคงรักษาส่วนหน้าไว้โดยมีการเคลื่อนที่ของกระดูกบางส่วนเนื่องจากการบดอัด[ 6 ]
ในปี 1991 นักบรรพชีวินวิทยา Minoru Tamura และเพื่อนร่วมงานได้รายงานการค้นพบกระดูกสันหลัง กระดูกแขนขา และฟันจากชั้นหินJobu Formation ในยุคครีเทเชียสตอนปลาย ของคุมาโมโตะประเทศญี่ปุ่น ซึ่งพวกเขาได้จัดให้เป็นAllosaurus sp. (ชนิดที่ไม่ระบุ) ซึ่งจะทำให้เป็นตัวแทนที่อายุน้อยที่สุดของสกุลนั้น ต่อมาในปี 2025 นักบรรพชีวินวิทยา André Burigo และOctávio Mateusได้ค้นพบกระดูกต้นขาที่คล้ายกับของSegnosaurusและสังเกตว่า มีซากของ เทริซิโนซอร์ อื่นๆ ที่พบในบริเวณนั้น ดังนั้นพวกเขาจึงจัดซากเหล่านี้ใหม่เป็นSegnosaurus sp. [ 7 ] [ 8 ]
คำอธิบาย

เซกโนซอรัสเป็นเทอริซิโนซอร์ขนาดใหญ่ที่คาดว่ามีความยาวประมาณ 6–7 เมตร (20–23 ฟุต) และมีน้ำหนักประมาณ 1.3 ตัน (1.4 ตันสั้น) [ 9 ] [ 10 ] [ 6 ]อย่างไรก็ตาม Campione & Evans ในปี 2020 คำนวณมวลร่างกายของมันที่ 4.17 เมตริกตัน (4.60 ตันสั้น) [ 11 ]เซกโนซอรัสยังไม่เป็นที่รู้จักอย่างสมบูรณ์ แต่ในฐานะเทอริซิโนซอริเดมันน่าจะเป็นสัตว์สองขาและมีโครงสร้างที่แข็งแรง โดยลำตัวจะเอียงขึ้นเมื่อเทียบกับเทอโรพอดชนิด อื่นๆ หัวจะมีขนาดเล็ก มีแรมโฟเทกา (จงอยปากแข็ง) ที่ปลายขากรรไกร และคอยาวเรียว นิ้วไม่ยาวมากนัก แต่มีกรงเล็บขนาดใหญ่ ด้านหน้าของกระดูกเชิงกรานถูกปรับให้รองรับท้องที่ขยายใหญ่ขึ้น[ 9 ] [ 6 ] [ 1 ]เป็นที่ทราบกันว่าเทริซิโนซอร์มีขนที่เรียบง่ายและดั้งเดิมดังที่เห็นได้จากฟอสซิลของสกุลพื้นฐาน (หรือ "ดั้งเดิม") อย่างเบเปียโอซอรัสซึ่งเป็นไดโนเสาร์ที่ไม่ใช่นกตัวที่สองที่รู้จักกันว่ามีผิวหนัง แบบนี้ หลังจากซิโนซอโร พเทอริกซ์ และเจียนชางโกซอรัส [ 12 ] [ 13 ] เนื่องจากเทริซิโนซอร์ส่วนใหญ่ยังไม่เป็นที่รู้จักอย่างสมบูรณ์ จึงไม่แน่ใจว่าลักษณะทางกายวิภาคที่ใช้ในการจำแนกเซกโนซอรัสแพร่หลายในกลุ่มนี้มากน้อยเพียงใด หลายสกุลไม่สามารถเปรียบเทียบกันได้โดยตรงเพราะกระดูกที่เทียบเท่ากันไม่ได้รับการเก็บรักษาไว้[ 4 ] [ 6 ]
ขากรรไกรล่างและฟันล่าง
ขากรรไกรล่างของเซกโนซอรัสมีลักษณะต่ำและยาว แต่ค่อนข้างแข็งแรงและไม่มีรูปทรงที่แน่นอนเมื่อเทียบกับของเออร์ลิโคซอรัสซึ่งมีลักษณะเพรียวบางกว่า ขากรรไกรล่างซีกขวาที่เกือบสมบูรณ์ (ครึ่งหนึ่งของขากรรไกรล่าง) มีความยาว 379 มม. (14.9 นิ้ว) จากด้านหน้าไปด้านหลัง จุดสูงสุดอยู่ที่ 55.5 มม. (2.19 นิ้ว) และจุดต่ำสุดอยู่ที่ 24.5 มม. (0.96 นิ้ว) กระดูกเดนทารีซึ่งเป็นกระดูกที่รองรับฟันและเป็นส่วนใหญ่ของส่วนหน้าของขากรรไกรล่าง มีรูปร่างซับซ้อนกว่าของเทอริซิโนซอรัสในยุคแรกๆ ส่วนที่รองรับฟันมีลักษณะเกือบเป็นสี่เหลี่ยมผืนผ้าและลาดลงเมื่อมองจากด้านข้าง โดยมีส่วนโค้งที่เด่นชัดตลอดความยาวส่วนบนของส่วนหน้า ซึ่งมีความสุดขั้วมากกว่าที่พบในเทอริซิโนซอรัสชนิดอื่นๆ ส่วนหน้าสุดของกระดูกเดนทารีเบี่ยงเบนลงอย่างมากในมุมประมาณ 30 องศา ซึ่งเป็นลักษณะเฉพาะของสกุลนี้ เมื่อขากรรไกรล่างแต่ละข้างประกบกัน จะเกิดเป็นส่วนเชื่อมต่อขากรรไกรล่างรูปตัวยูที่ไม่มีฟัน ยื่นขึ้นไปทางด้านหน้า ดังเช่นในErlikosaurusและNeimongosaurusบริเวณด้านหน้าของกระดูกขากรรไกรล่างที่ไม่มีฟันนั้นมีความยาว 25.5 มม. (1.00 นิ้ว) ในขากรรไกรล่างด้านขวาของตัวอย่างต้นแบบ เมื่อเทียบสัดส่วนแล้ว ส่วนที่ไม่มีฟันของกระดูกขากรรไกรล่างคิดเป็น 20% ของแถวฟัน ซึ่งมีความยาว 150.3 มม. (5.92 นิ้ว) เมื่อเปรียบเทียบกันแล้ว บริเวณที่ไม่มีฟันของErlikosaurusคิดเป็นประมาณ 12% ของความยาวแถวฟัน และแทบไม่มีเลยในJianchangosaurusความสูงของกระดูกขากรรไกรล่างลดลงไปทางส่วนท้ายสุดของแถวฟัน จากนั้นจึงแผ่กว้างออกไปสัมผัสกับ กระดูก surangular ที่อยู่ด้านหลัง ในทางตรงกันข้าม ส่วนท้ายของกระดูกขากรรไกรล่างในErlikosaurusค่อยๆ เข้าหากระดูก surangular ในลักษณะโค้งอย่างนุ่มนวล[ 6 ] [ 1 ]

เซกโนซอรัสมีความโดดเด่นในกลุ่มเทอริซิโนซอรัสตรงที่ส่วนท้ายสุดของกระดูกขากรรไกรล่างไม่มีฟัน ฟันมีอยู่เฉพาะในส่วนหน้าสองในสามของกระดูกขากรรไกรล่าง ซึ่งมีเบ้าฟัน 24 เบ้าในลักษณะคล้ายกับเจียนชางโกซอรัสแต่แตกต่างจากเออร์ลิโคซอรัสซึ่งกระดูกขากรรไกรล่างเกือบทั้งหมดมีฟัน โดยมีเบ้าฟัน 31 เบ้า แถวฟันของเซกโนซอรัสฝังลึกและมีขอบยื่นออกมาทางด้านนอกเช่นเดียวกับเทอริซิโนซอรัสที่วิวัฒนาการแล้ว (หรือ "ขั้นสูง") แต่ต่างจากในกลุ่มอนุกรมวิธานอื่นๆ ที่เกี่ยวข้อง เบ้าฟันนี้มีอยู่เฉพาะในส่วนท้ายของกระดูกขากรรไกรล่าง และขอบที่ยกขึ้นซึ่งเป็นตัวกำหนดเบ้าฟันนั้นไม่เด่นชัดนักเซกโนซอรัสมีความพิเศษตรงที่มีสันเตี้ยๆ ยื่นขึ้นมาระหว่างเบ้าฟันที่ห้าถึงสิบสี่ ซึ่งแบ่งกระดูกขากรรไกรล่างออกเป็นสองส่วนที่มีขนาดเกือบเท่ากัน คือส่วนหน้าและส่วนหลัง เหนือสันนี้ขึ้นไปเล็กน้อย กระดูกขากรรไกรล่างถูกเจาะด้วยรูเรียง เป็นแถว เหมือนในJianchangosaurusและAlxasaurusซึ่งแถวรูเหล่านี้ไม่สม่ำเสมอมากขึ้นในบริเวณรอบๆ รอยต่อของขากรรไกรล่าง ซึ่งเป็นจุดที่ขากรรไกรล่างสองซีกมาบรรจบกันที่ด้านหน้า แต่ในErlikosaurus แถวรูเหล่านี้กลับอยู่ในแนวเดียวกันและอยู่ด้านข้างของ สัน ร่อง Meckelianที่วิ่งไปตามด้านในของขากรรไกรล่างนั้นอยู่ต่ำลงไปกว่าในErlikosaurusและมีความลึกคงที่จนถึงตำแหน่งฟันซี่ที่สิบสาม หลังจากนั้นจึงกว้างขึ้น ส่วนประกอบของขากรรไกรล่างด้านหลังกระดูกขากรรไกรล่าง ( กระดูก splenial , surangular, angularและprearticular ) มีลักษณะแตกต่างจากเทอริซิโนซอร์อื่นๆ โดยมีลักษณะเรียวและเป็นเส้นตรง และมีส่วนทำให้ส่วนท้ายของขากรรไกรล่างครึ่งซีกนั้นยาวและเกือบเป็นรูปสี่เหลี่ยมผืนผ้า[ 6 ]กระดูกซูแรนกูลาร์ยาวและมีรูปร่างคล้ายดาบ กระดูกแองกูลาร์มีรูปร่างคล้ายปีก กระดูกพรีอาร์ติคูลาร์แคบและโค้ง และกระดูกสเปลเนียลบางและมีรูปร่างเป็นสามเหลี่ยม ช่องเปิดขากรรไกรด้านนอก ซึ่งเป็นช่องเปิดที่ด้านนอกของขากรรไกร มีขนาดใหญ่กว่าของเออร์ลิโคซอรัสเนื่องจากกระดูกซูแรนกูลาร์ตื้นจากบนลงล่าง[ 1 ]

เซกโนซอรัสมีฟันในขากรรไกรล่างน้อยที่สุด โดยมี 24 ซี่ในแต่ละครึ่ง ซึ่งกำหนดจากจำนวนเบ้าฟัน และยังมีฟันขนาดใหญ่ที่สุดเท่าที่รู้จักในกลุ่มเทอริซิโนซอรัส ฟันในขากรรไกรล่างมีลักษณะเป็นรูปทรงใบไม้ (foliodont) และมีส่วนยอดฟันที่ขยายใหญ่ขึ้น ค่อนข้างสูง และแบนด้านข้าง โดยมีส่วนโค้งเล็กน้อยที่ขอบบนของปลายฟัน เมื่อเปรียบเทียบกันแล้ว ฟันของเออร์ลิโคซอรัสมีขนาดเล็กกว่า สมมาตร และเรียบง่ายกว่า ฐานของส่วนยอดฟันมีขนาดใหญ่ขึ้นเล็กน้อยไปทางด้านหลังตามแนวฟัน ซึ่งสะท้อนให้เห็นถึงการลดลงของการบีบอัดด้านข้าง พื้นผิวด้านหน้าของส่วนยอดฟันและด้านข้างที่หันออกด้านนอกมีลักษณะนูน ในขณะที่ด้านข้างที่หันเข้าด้านในมีลักษณะเว้า มีสันหนาพาดตามแนวยาวของด้านข้างที่หันเข้าด้านในใกล้กับครึ่งบนของส่วนยอดฟัน ซึ่งมีร่องเล็กๆ ขนาบข้างอยู่ใกล้กับขอบด้านหน้าและด้านหลังของฟัน และทอดยาวเกือบถึงส่วนคอ (cervix; ส่วนเปลี่ยนผ่านระหว่างส่วนยอดและราก) ของฟัน โดยทั่วไป ฟัน 18 ซี่แรกที่อยู่ด้านหน้าสุดมีลักษณะค่อนข้างเหมือนกัน (ประเภทเดียวกัน) แม้ว่าส่วนยอดของฟันซี่ที่สองจะสั้นกว่าและเรียวกว่าก็ตาม ซึ่งอาจเป็นจริงสำหรับฟันซี่แรกด้วย แต่ฟันซี่แรกไม่ได้รับการเก็บรักษาไว้ ฟันที่อยู่ด้านหลังในแถวก็ลดความสูงลงไปทางด้านหลังเช่นกัน เมื่อเปรียบเทียบกันแล้ว ฟันกรามล่างด้านหน้า 4-5 ซี่ของErlikosaurusมีลักษณะเป็นรูปทรงกรวย (conidont) โดยมีการเปลี่ยนผ่านอย่างค่อยเป็นค่อยไปเป็นฟันแบบ foliodont [ 6 ] [ 1 ]
ฟันกรามล่างเรียงตัวแน่น แต่ไม่ชิดกันจนเกินไป โดยส่วนยอดของฟันจะเข้าใกล้กันที่กึ่งกลางความยาว ฟันย่อย (รอยหยัก) มีขนาดใหญ่และโป่งออกเล็กน้อย ค่อยๆ ลดขนาดลงไปทางปลายฟัน โดยมีฟันย่อยประมาณ 5-6 ซี่ต่อ 3 มิลลิเมตร (0.12 นิ้ว) สัน ด้านหน้า (ขอบตัด) พับขึ้นทับซ้อนกับพื้นผิวด้านในของส่วนยอดฟันในฟันซี่ที่สามถึงสิบแปด แต่ไม่มีรอยพับดังกล่าวในฟันซี่ที่สองและอาจรวมถึงฟันซี่แรกด้วย ฟันย่อยตั้งฉากกับปลายส่วนยอดฟันโดยประมาณ แต่ขนานกับความสูงของส่วนยอดฟันทางด้านหน้า และขนานกับด้านสามเหลี่ยมทางด้านหลัง มีฟันย่อยเสริม (นอกเหนือจากที่อยู่บนสัน) ยื่นออกมาจากพื้นผิวด้านหน้าของรอยพับสัน ทำให้ขอบด้านหน้าของส่วนยอดฟันขรุขระมากขึ้น สันนูนของขอบด้านหลังก็มีการเปลี่ยนแปลงอย่างมากและแยกออกเป็นสองส่วนใกล้กับส่วนคอ โดยมีลักษณะเป็นรูปสามเหลี่ยมแบนและนูนขึ้นมา ซึ่งยื่นออกมาจากส่วนยอดของฟันและสัมผัสหรือเข้าใกล้สันนูนที่พับอยู่บนขอบด้านหน้าของส่วนยอดฟันด้านหลัง (การจัดเรียงแบบนี้พบในฟันซี่ที่ 2–12) สันนูนที่แยกออกเป็นสองส่วนเช่นนี้พบได้ใน เทโรพอดเท ทานูรันชนิด อื่น ๆ ซึ่งถือว่าเป็นความผิดปกติที่เกิดจากการบาดเจ็บการเปลี่ยนฟัน ที่ผิดปกติ หรือปัจจัยทางพันธุกรรม แม้ว่าสภาพในเซกโนซอรัสจะคล้ายกัน แต่ก็แสดงออกอย่างสม่ำเสมอทั่วทั้งฟันของขากรรไกรล่างทั้งสองข้าง และดูเหมือนจะไม่ใช่ความผิดปกติ แต่ทำหน้าที่ทำให้การสัมผัสระหว่างฐานฟันหยาบขึ้น[ 6 ]
ฟันกรามซี่ที่ 22 และ 23 ของSegnosaurusมีขนาดเล็กกว่าฟันซี่อื่นๆ อย่างเห็นได้ชัด เกือบจะเป็นฟันกรามแบน และมีสันนูนที่สามเพิ่มขึ้นมา โดยมีฟันเล็กๆ อยู่ด้านใน ฟันซี่หลังสุดส่วนใหญ่เสียหาย จึงไม่ทราบลักษณะที่สมบูรณ์ สันนูนที่เพิ่มขึ้นบนฟันซี่ที่ 23 ดูเหมือนจะมีฟันเล็กๆ ขึ้นเต็มซี่ ในขณะที่ฟันเล็กๆ บนฟันซี่ที่ 22 มีเฉพาะด้านฐานของตัวฟันเท่านั้นSegnosaurusเป็นไดโนเสาร์เทอโรพอดที่มีเอกลักษณ์เฉพาะตัวในบรรดาไดโนเสาร์เทอโรพอดที่รู้จักกันทั้งหมด เนื่องจากมีสันนูนสามชั้น กระดูกเบ้าฟันที่ 14 บนขากรรไกรล่างด้านขวาของตัวอย่างต้นแบบถูกปิดทับด้วยการเจริญเติบโตของกระดูกที่ดูเหมือนจะผิดปกติ (เนื่องจากบาดเจ็บหรือโรค) แต่ฟันในส่วนนั้นของขากรรไกรล่างเสียหาย จึงไม่สามารถระบุได้ว่าฟันได้รับผลกระทบจากสิ่งนี้อย่างไร ฟันในบริเวณเดียวกันของขากรรไกรล่างด้านซ้ายมีสันสามอัน แม้ว่าขากรรไกรล่างนี้จะไม่มีร่องรอยภายนอกของพยาธิสภาพที่อาจนำไปสู่สภาวะนี้ ดังนั้นจึงไม่สามารถสรุปหรือตัดความเป็นไปได้ที่ลักษณะนี้เป็นผลมาจากพยาธิสภาพได้เซกโนซอรัสเปลี่ยนฟันเป็นคลื่นที่วิ่งจากด้านหลังไปด้านหน้าของขากรรไกร ซึ่งครอบคลุมยอดฟันที่งอกออกมาสองถึงสามซี่ ฟันที่งอกออกมาเต็มที่บางซี่มีการสึกหรอที่สันด้านหลัง ซึ่งแตกต่างจากที่พบในเทอริซิโนซอร์อื่นๆ เนื้อสัมผัสของเคลือบฟันดูเหมือนจะไม่สม่ำเสมอโดยทั่วไป และรากฟันมีลักษณะเกือบเป็นวงกลม[ 6 ]
โครงกระดูกส่วนลำตัว

กระดูกสะบัก (กระดูกหัวไหล่) ของSegnosaurusมีลักษณะตรงและแบนที่ปลายด้านบน และเชื่อมติดกับกระดูกโคราคอยด์ ทำให้เกิดเป็นกระดูกสะบักโคราคอยด์ กระดูกโคราคอยด์มีความกว้างมาก มีรูปร่างเป็นสี่เหลี่ยมผืนผ้าและหนาตรงกลาง กระดูกต้นแขนขนาดใหญ่มีความยาว 560 มม. (22 นิ้ว) มีแกนเกือบเป็นทรงกระบอกและมีปุ่มกระดูกที่ชัดเจนสำหรับการเชื่อมต่อกับกระดูกเรเดียสและกระดูกอัลนาของแขนส่วนล่าง สันเดลโตเพคทอรัล ซึ่งเป็นบริเวณที่กล้ามเนื้อเดลตอยด์ยึดติดกับด้านหน้าส่วนบนของกระดูกต้นแขนนั้นได้รับการพัฒนาอย่างดี[ 1 ]กระดูกต้นแขนมีลักษณะแตกต่างจากของเทอริซิโนซอร์อื่นๆ โดยมีลักษณะตรงมากกว่ารูปทรงโค้งงอและไม่ขยายหรือโค้งไปข้างหน้าที่ปลายด้านบน กระดูกต้นแขนยังไม่ขยายออกตรงกลาง และเอ็นทีปิคอนไดล์ก็ไม่ได้รับการพัฒนาอย่างดี การขาดคุณลักษณะเหล่านี้มีความคล้ายคลึงกับออร์นิโทมิโมซอร์และโทรโอโดนทิดมากกว่าเทอริซิโนซอร์อื่นๆ[ 4 ]กระดูกเรเดียสมีขนาดใหญ่มาก—ประมาณ 60 เปอร์เซ็นต์ของกระดูกฮิวเมอรัส—โดยมีแกนตรง กระดูกอัลนาหนากว่ากระดูกเรเดียสและยาวกว่าเล็กน้อย—ประมาณ 70 เปอร์เซ็นต์ของกระดูกฮิวเมอรัส—และบิดเล็กน้อยตามแกนกลาง มือมีสามนิ้ว กระดูกนิ้วมีลักษณะแบนจากบนลงล่าง และร่องข้อต่อด้านข้างไม่พัฒนามากนัก กระดูกนิ้วแรกของนิ้วแรกยาวและบาง ในขณะที่กระดูกนิ้วแรกและนิ้วที่สองของนิ้วที่สองสั้น เล็บของนิ้วที่สามยาวกว่ากระดูกนิ้วที่สองเล็กน้อยและค่อนข้างแบนจากบนลงล่าง ซึ่งอาจเป็นคุณลักษณะเฉพาะของเซกโนซอรัสเล็บนี้โค้งแหลมมาก แหลมคม และแบนจากด้านหนึ่งไปอีกด้านหนึ่ง ปุ่มล่างซึ่งเป็นจุดที่เอ็นงอเกาะติดกับเล็บนั้นหนาและแข็งแรง[ 1 ] [ 4 ] [ 5 ] [ 14 ]

กระดูกเชิงกรานของSegnosaurusแข็งแรงและมีส่วนยื่นที่ชี้ไปด้านข้างอย่างชัดเจนที่ด้านหน้า กระดูกเชิงกรานสั้นลงที่ด้านหน้า ซึ่งเป็นลักษณะที่พบในเทโรพอดที่คล้ายนก แต่ไม่พบทั่วไปในเทโรพอดโดยรวม[ 1 ]กระดูกหัวหน่าวชี้ไปด้านหลังและลงขนานกับกระดูกเชิงกรานส่วนล่าง การวางตัวไปด้านหลังของกระดูกหัวหน่าวนี้เรียกว่า ภาวะ โอพิสโทพิ วบิก ลักษณะนี้พบเฉพาะในนกและญาติสนิทที่สุดของพวกมันคือโคเอลูโรซอเรียนในขณะที่ไดโนเสาร์เทโรพอดอื่นๆ มีกระดูกหัวหน่าวชี้ไปด้านหน้า[ 15 ] [ 14 ]กระดูกหัวหน่าวมีลักษณะยาว แบนไปด้านข้าง และมีส่วนยื่นรูปทรงรีหรือ "รองเท้าบูท" ที่ด้านหน้าของปลายล่าง[ 5 ] [ 14 ]กระดูกเชิงกรานของ Segnosaurus แตกต่างจากของเทอริซิโนซอรัสชนิดอื่นตรงที่ขอบบนของกระดูกเชิงกรานส่วนล่างยื่นออกมาอย่างเห็นได้ชัด และส่วนที่ยื่นไปด้านหลังของกระดูกเชิงกรานส่วนล่างนั้นยาวมาก เกือบ 50 เปอร์เซ็นต์ของความยาวจากด้านหน้าไปด้านหลังของกระดูกเชิงกรานส่วนล่าง ลักษณะบางอย่างของกระดูกเชิงกรานคล้ายกับของNothronychusโดยเฉพาะกระดูกเชิงกรานส่วนล่าง แต่ไม่แน่ใจว่าความคล้ายคลึงกันเหล่านี้เกิดจากการมีบรรพบุรุษร่วมกันโดยไม่รวมเทอริซิโนซอริเดชนิดอื่นที่วิวัฒนาการมา หรือเพราะพวกมันยังคงรักษาลักษณะพื้นฐานที่สูญหายไปในญาติชนิดอื่น กระดูกเชิงกรานส่วนล่างของSegnosaurusแตกต่างจากของNothronychusตรงที่มีกระดูกเชิงกรานส่วนล่างเกือบเป็นรูปสี่เหลี่ยมผืนผ้าและรูเปิดของกระดูกเชิงกรานเกือบเป็นรูปวงกลม กระดูกเชิงกรานของ Segnosaurus แตกต่างจากของEnigmosaurusตรงที่กระบวนการ obturator ที่ลึกไม่เชื่อมกับส่วนที่ตรงกันตรงกลาง กระดูกหัวหน่าวไม่เชื่อมติดกัน และเนื่องจากส่วนล่างของกระดูกหัวหน่าวมีความกว้างจากด้านหน้าไปด้านหลังSegnosaurusแตกต่างจากทั้งNothronychusและEnigmosaurusตรงที่มีร่อง brevis ที่ลึก (ร่องที่กล้ามเนื้อ caudofemoralis brevisของหางเริ่มต้น) และเนื่องจากกระดูกหัวหน่าวขาดการยื่นออกไปด้านหลังที่พัฒนาอย่างดี[ 4 ]
กระดูกต้นขา (femur) ตรง มีหน้าตัดรูปไข่ และยาว 840 มม. (33 นิ้ว) หัวของกระดูกต้นขาวางอยู่บน "คอ" ที่ยาว และปุ่มกระดูกด้านล่างมีความชัดเจน กระดูกหน้าแข้ง (tibia) ตรง สั้นกว่ากระดูกต้นขาเล็กน้อย และบิดไปตามแกน กระดูกน่อง (fibula) ยาวและแคบลงไปทางปลายด้านล่าง กระดูกฝ่าเท้า (metatarsus) สั้น ใหญ่ และประกอบด้วยกระดูกห้าชิ้น โดยสี่ชิ้นทำหน้าที่เป็นส่วนรองรับและมีนิ้วเท้าสี่นิ้ว เท้าที่มีสี่นิ้ว (tetradactyl) เป็นลักษณะเฉพาะของเทอริซิโนซอร์ที่วิวัฒนาการแล้ว เทอริซิโนซอร์พื้นฐานและเทอโรพอดอื่นๆ ทั้งหมดมีเท้าที่มีสามนิ้ว (tridactyl) ซึ่งนิ้วแรกสั้นและไม่ถึงพื้น ภายนอกแล้ว กระดูกฝ่าเท้ามีลักษณะคล้ายกับ แต่มีสัดส่วนใหญ่กว่า กระดูกฝ่าเท้าของ โปรซอโรพอด ซึ่งเป็น กลุ่มวิวัฒนาการช่วงแรกของซอโรโพโดมอร์ฟ(sauropodomorphs ) กระดูกส่วนปลายของกระดูกฝ่าเท้าส่วนบนมีขนาดใหญ่ขึ้น (ขยายใหญ่) ซึ่งเป็นลักษณะเด่นของสกุลนี้ นิ้วเท้าแรกสั้นกว่านิ้วอื่นๆ แต่มีความสำคัญในการใช้งานเท่าเทียมกัน นิ้วเท้าที่สองและสามมีความยาวเท่ากัน ในขณะที่นิ้วเท้าที่สี่บางที่สุด เล็บนิ้วเท้าแข็งแรง โค้งงออย่างชัดเจน แบนด้านข้าง และแหลมกว่าของโปรซอโรพอด ปุ่มกระดูกด้านล่างที่เอ็นงอเกาะอยู่นั้นแข็งแรง แม้ว่าการขาดการบีบอัดที่แข็งแรงของเล็บนิ้วเท้าจะทำให้Segnosaurus แตกต่าง จากErlikosaurus จากแหล่งกำเนิดเดียวกัน แต่การขาดการบีบอัดเป็นเรื่องปกติในกลุ่มเทอริซิ โนซอร์ ดังนั้นจึงไม่ใช่ลักษณะเฉพาะของSegnosaurus [ 1 ] [ 4 ] [ 5 ]กระดูกสันหลังส่วนคอมีลักษณะแบนและมีแกนกลาง (ลำตัว) ขนาดใหญ่และส่วนโค้งประสาทต่ำ กระดูกเชิงกรานประกอบด้วยกระดูกสันหลังหกชิ้นที่เชื่อมติดกันอย่างแน่นหนา กระดูกสันหลังส่วนกลางของกระดูกสันหลังเหล่านี้กว้างและค่อนข้างยาว และกระดูกสันหลังส่วนกลางแต่ละชิ้นยาวกว่าความกว้างเล็กน้อย กระดูกสันหลังส่วนประสาทที่นี่ไม่ยาวมากนัก แต่ยาวเกินระดับของกระดูกเชิงกราน กระดูกสันหลังส่วนหางที่อยู่ใกล้ลำตัวมากที่สุดมีขนาดใหญ่ สูง และค่อนข้างแบนจากด้านหนึ่งไปอีกด้านหนึ่ง ส่วนโค้งประสาทต่ำและมีช่องประสาทขนาดเล็ก กระดูกสันหลังส่วนหางที่อยู่ใกล้ปลายหางมีลักษณะแบนและมีกระดูกสันหลังส่วนกลางที่สั้นและใหญ่ กระบวนการตามขวางของกระดูกสันหลังส่วนหางและซี่โครงแข็งแรงและยาว[ 1 ] [ 5 ]
การจำแนกประเภท
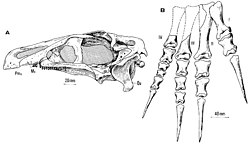
เซกโนซอรัสและญาติของมัน ซึ่งปัจจุบันได้รับการยอมรับว่าเป็นเทอริซิโนซอรัส ("สัตว์เลื้อยคลานเคียว") เคยถูกมองว่าเป็นกลุ่มที่ลึกลับมานานแล้ว ลักษณะที่ผสมผสานกันคล้ายกับไดโนเสาร์กลุ่มต่างๆ และความหายากของฟอสซิลทำให้เกิดข้อถกเถียงเกี่ยวกับความสัมพันธ์ทางวิวัฒนาการของพวกมันเป็นเวลาหลายทศวรรษหลังจากการค้นพบครั้งแรก (ชิ้นส่วนแขนขาหน้าของเทอริซิโนซอรัสเองนั้น เดิมทีถูกระบุว่าเป็นของเต่ายักษ์เมื่อมีการอธิบายในปี 1954) [ 16 ] [ 17 ] [ 14 ]ในปี 1979 เพิร์ลตั้งข้อสังเกตว่า ฟอสซิล ของเซกโนซอรัสอาจเป็นตัวแทนของวงศ์ไดโนเสาร์ใหม่ ซึ่งเขาตั้งชื่อว่าเซกโนซอริเด โดยเซกโนซอรัสเป็นสกุลต้นแบบและสมาชิกเพียงตัวเดียว เขาจัดประเภทเซกโนซอริเดอย่างคร่าวๆ ว่าเป็นเทอโรพอด ซึ่งโดยทั่วไปแล้วถือว่าเป็นไดโนเสาร์ "กินเนื้อ" โดยชี้ให้เห็นถึงความคล้ายคลึงกันในขากรรไกรล่างและฟันหน้า โดยใช้ลักษณะของกระดูกต้นแขนและกรงเล็บมือ เขาได้จำแนก Segnosauridae ออกจากวงศ์เทโรพอดDeinocheiridaeและ Therizinosauridae ซึ่งในขณะนั้นรู้จักกันเฉพาะจากสกุลDeinocheirusและTherizinosaurusตามลำดับ ซึ่งส่วนใหญ่มีแขนขาหน้าขนาดใหญ่ที่พบในมองโกเลีย[ 1 ]ต่อมาในปี 1979 Barsbold และ Perle พบว่าลักษณะเชิงกรานของเซกโนซอริเดและโดรเมโอซอริเดนั้นแตกต่างจากเทโรพอด "แท้" มากจนควรแยกออกเป็นสามกลุ่มอนุกรมวิธานที่มีลำดับชั้นเดียวกัน อาจอยู่ในระดับอันดับย่อยภายในSaurischia ซึ่ง เป็นหนึ่งในสองกลุ่มหลักของไดโนเสาร์ อีกกลุ่มหนึ่งคือOrnithischia [ 18 ]
ในปี 1980 บาร์สโบลด์และเพิร์ลได้ตั้งชื่อกลุ่มไดโนเสาร์เทอโรพอดกลุ่มใหม่ว่า เซกโนซอเรีย (Segnosauria) ซึ่งประกอบด้วยเฉพาะวงศ์เซกโนซอริดี (Segnosauridae) เท่านั้น ในบทความเดียวกัน พวกเขายังตั้งชื่อสกุลใหม่ว่าเออร์ ลิโคซอรัส (Erlikosaurus ) (ซึ่งรู้จักจากกะโหลกและโครงกระดูกบางส่วนที่ได้รับการอนุรักษ์ไว้อย่างดี) ซึ่งพวกเขาสันนิษฐานว่าเป็นไดโนเสาร์ในวงศ์เซกโนซอริดี และรายงานการค้นพบกระดูกเชิงกรานบางส่วนของไดโนเสาร์เซกโนซอเรียนที่ไม่สามารถระบุชนิดได้ โดยทั้งสองตัวอย่างมาจากชั้นหินเดียวกันกับเซกโนซอรัส (Segnosaurus ) ซากดึกดำบรรพ์เหล่านี้ให้ข้อมูลที่ค่อนข้างสมบูรณ์เกี่ยวกับกลุ่มนี้ พวกมันมีลักษณะร่วมกันคือ กระดูกเชิงกรานแบบโอพิสโทพิวบิก (opisthopubic pelvis) ขากรรไกรล่างเรียว และขากรรไกรส่วนหน้าไม่มีฟัน บาร์สโบลด์และเพิร์ลระบุว่า แม้ว่าลักษณะบางอย่างของพวกมันจะคล้ายกับไดโนเสาร์กลุ่มออร์นิธิสเชียน (Ornithischians) และซอโรพอด (Sauropods) แต่ความคล้ายคลึงเหล่านี้เป็นเพียงผิวเผินและแตกต่างกันอย่างชัดเจนเมื่อพิจารณาในรายละเอียด แม้ว่าโดยพื้นฐานแล้วพวกมันจะแตกต่างจากเทโรพอดอื่นๆ—อาจเป็นเพราะการแยกตัวออกจากพวกมันค่อนข้างเร็ว—และสมควรได้รับการจัดอยู่ในอันดับย่อยใหม่ แต่พวกมันก็แสดงความคล้ายคลึงกับเทโรพอดอยู่บ้าง เนื่องจาก ตัวอย่าง Erlikosaurusขาดกระดูกเชิงกราน ผู้เขียนจึงไม่แน่ใจว่าเซกโนซอเรียนที่ยังไม่ได้รับการระบุชนิดนั้นจะอยู่ในสกุลเดียวกันหรือไม่ ซึ่งในกรณีนั้นพวกเขาจะพิจารณาว่าเป็นส่วนหนึ่งของวงศ์ที่แยกต่างหาก[ 5 ]แม้ว่าErlikosaurusจะเปรียบเทียบโดยตรงกับSegnosaurus ได้ยาก เนื่องจากซากของมันไม่สมบูรณ์ แต่ Perle ระบุในปี 1981 ว่าไม่มีเหตุผลใดที่จะแยกมันออกเป็นวงศ์อื่น[ 19 ]

ในปี พ.ศ. 2525 Perle รายงานการค้นพบชิ้นส่วนขาหลังที่คล้ายกับของSegnosaurusและจัดให้อยู่ในกลุ่มTherizinosaurusซึ่งพบชิ้นส่วนขาหน้าในตำแหน่งเกือบเดียวกัน เขาสรุปว่า Therizinosauridae, Deinocheiridae และ Segnosauridae ซึ่งทั้งหมดมีขาหน้าที่ขยายใหญ่ขึ้น เป็นกลุ่มอนุกรมวิธานเดียวกันSegnosaurusและTherizinosaurusมีความคล้ายคลึงกันเป็นพิเศษ ทำให้ Perle เสนอว่าพวกมันควรอยู่ในวงศ์เดียวกันโดยไม่รวม Deinocheiridae (ปัจจุบันDeinocheirusได้รับการยอมรับว่าเป็นornithomimosaur ) [ 20 ] [ 21 ] Barsbold ยังคงจัดSegnosaurusและErlikosaurus ไว้ ในวงศ์ Segnosauridae ในปี พ.ศ. 2526 และตั้งชื่อสกุลใหม่ว่าEnigmosaurusโดยอิงจากกระดูกเชิงกรานของเซกโนซอเรียนที่ยังไม่ได้รับการระบุมาก่อน โครงสร้างของกระดูกเชิงกรานของErlikosaurusยังไม่เป็นที่รู้จัก แต่ Barsbold พิจารณาว่าไม่น่าเป็นไปได้ ที่กระดูกเชิงกราน ของ Enigmosaurusจะเป็นของ Erlikosaurus เนื่องจากErlikosaurusและSegnosaurusมีความคล้ายคลึงกันในด้านอื่นๆ ในขณะที่กระดูกเชิงกรานของEnigmosaurusแตกต่างจากของSegnosaurus มาก Barsbold พบว่าเซกโนซอริเดมีลักษณะแปลกประหลาดเมื่อเทียบกับเทโรพอดทั่วไป ซึ่งอาจเป็นความเบี่ยงเบนที่สำคัญมากในวิวัฒนาการของเทโรพอด หรืออาจอยู่นอกกลุ่ม อย่างไรก็ตาม เขายังคงจัดพวกมันไว้ในกลุ่มเทโรพอด[ 3 ]ต่อมาในปี 1983 Barsbold ระบุว่ากระดูกเชิงกรานของเซกโนซอเรียนเบี่ยงเบนอย่างมีนัยสำคัญจากมาตรฐานของเทโรพอด และพบว่าการจัดเรียงของกระดูกเชิงกรานของพวกมันโดยทั่วไปคล้ายกับของซอโรพอด[ 22 ]

ในปี 1984 เกรกอรี เอส. พอลสรุปว่าเซกโนซอร์ไม่มีลักษณะของเทอโรโพดา แต่เป็นโปรซอโรพอดที่วิวัฒนาการมา จากยุคครี เทเชียส ตอนปลาย โดยมีการปรับตัวคล้ายกับออร์นิธิสเชียน เขาพบว่าเซกโนซอร์มีลักษณะคล้ายกับโปรซอโรพอดในส่วนของจมูก ขากรรไกร และเท้าหลัง คล้ายกับออร์นิธิสเชียนในส่วนของแก้ม เพดานปาก กระดูกเชิงกราน และข้อเท้า และคล้ายกับไดโนเสาร์ยุคแรกในด้านอื่นๆ เขาเสนอว่าออร์นิธิสเชียนสืบเชื้อสายมาจากโปรซอโรพอด และเซกโนซอร์เป็นซากดึกดำบรรพ์ขั้นกลางของการเปลี่ยนแปลงนี้ ซึ่งคาดว่าเกิดขึ้นในยุคไทรแอสสิกด้วยวิธีนี้ เขาจึงพิจารณาว่าเซกโนซอร์มีสถานะเทียบเคียงได้กับไดโนเสาร์กินพืชโดยทั่วไป เช่นเดียวกับที่โมโนทรีมมีสถานะเทียบเคียงได้กับสัตว์เลี้ยงลูกด้วยนม เขาพบว่าไม่น่าเป็นไปได้ แต่ก็ไม่ได้ตัดความเป็นไปได้ที่เซกโนซอร์อาจมีต้นกำเนิดมาจากเทโรพอด หรือว่าเซกโนซอร์ โปรซอโรพอด และออร์นิธิสเชียน อาจมีต้นกำเนิดแยกกันจากไดโนเสาร์ยุคแรก[ 23 ]เดวิด บี. นอร์แมนพิจารณาว่าแนวคิดของพอลเป็นที่ถกเถียงและ "น่าจะก่อให้เกิดการโต้เถียงกันมาก" ในปี 1985 [ 24 ]ในปี 1988 พอลยืนยันว่าเซกโนซอร์เป็นโปรซอโรพอดที่รอดชีวิตมาได้ในยุคหลัง มีลักษณะคล้ายออร์นิธิสเชียน และเสนอเอกลักษณ์ของเซกโนซอเรียนสำหรับเทริซิโนซอรัสเขายังจัดเซกโนซอเรียไว้ในไฟโตไดโนซอเรียซึ่งเป็นอันดับใหญ่ ที่ โรเบิร์ต บักเกอร์สร้างขึ้นในปี 1985 เพื่อบรรจุไดโนเสาร์กินพืชทั้งหมด[ 25 ]ในการศึกษาความสัมพันธ์ระหว่างไดโนเสาร์ซอริสเชียนในปี 1986 ฌาคส์ โกติเยร์สรุปว่าเซกโนซอร์เป็นโปรซอโรพอด แม้ว่าเขาจะยอมรับว่าพวกมันมีความคล้ายคลึงกับออร์นิธิสเชียนและเทโรพอด แต่เขาก็เสนอว่าลักษณะเหล่านี้วิวัฒนาการขึ้นอย่างอิสระ[ 26 ]ในบทคัดย่อการประชุมปี 1989 เกี่ยวกับความสัมพันธ์ระหว่างซอโรโพโดมอร์ฟ พอล เซเรโนยังพิจารณาว่าเซกโนซอร์เป็นโปรซอโรพอด โดยพิจารณาจากลักษณะของกะโหลกศีรษะ[ 27 ]

ในบทความวิจารณ์ ปี 1990 Barsbold และTeresa Maryańskaพบว่า Segnosauria เป็นกลุ่มซอริสเชียนที่หายากและผิดปกติ ซึ่งอยู่ในตำแหน่งที่ไม่ชัดเจนระหว่างซอโรโพโดมอร์ฟและเทโรพอด และน่าจะใกล้เคียงกับกลุ่มแรกมากกว่า ดังนั้น พวกเขาจึงจัดกลุ่มพวกมันเป็น Saurischia sedis mutabilis (ตำแหน่งอาจมีการเปลี่ยนแปลง) พวกเขายอมรับว่าขาหลังที่กำหนดให้กับTherizinosaurusในปี 1982 นั้นเป็นเซกโนซอเรียน แต่ไม่ถือว่านี่เป็นเหตุผลที่เพียงพอสำหรับTherizinosaurusเองที่จะเป็นเซกโนซอร์ เนื่องจากรู้จักมันจากขาหน้าเท่านั้น[ 28 ]ในปี 1993 Dale A. RussellและDong Zhi-Mingได้อธิบายสกุลใหม่Alxasaurusจากประเทศจีน ในขณะนั้น นี่คือเทโรพอดขนาดใหญ่ที่สมบูรณ์ที่สุดจากยุคและสถานที่นั้น แม้ว่าAlxasaurusจะมีความคล้ายคลึงกับโปรซอโรพอดในบางแง่มุม แต่ลักษณะทางสัณฐานวิทยาโดยละเอียดของแขนขาของมันเชื่อมโยงมันกับTherizinosaurusและเซกโนซอร์ เนื่องจากแขนขาหน้าและหลังของAlxasaurus ได้รับการอนุรักษ์ไว้ จึงแสดงให้เห็นว่าการจัดกลุ่มแขนขาหลังของเซกโนซอร์เข้ากับTherizinosaurus ของ Perle นั้นน่าจะถูกต้อง ดังนั้น Russell และ Dong จึงเสนอว่า Segnosauridae เป็นชื่อพ้องรองของชื่อเดิม Therizinosauridae และAlxasaurusเป็นตัวแทนที่รู้จักอย่างสมบูรณ์ที่สุดในปัจจุบัน พวกเขายังตั้งชื่อลำดับชั้นทางอนุกรมวิธานที่สูงขึ้นใหม่ว่าTherizinosauroideaเพื่อรวมAlxasaurusและ Therizinosauridae ไว้ด้วยกัน เนื่องจากสกุลใหม่นี้มีความแตกต่างจากญาติของมันอยู่บ้าง พวกเขาสรุปว่าเทอริซิโนซอร์เป็นเทโรพอดเททานูรัน ซึ่งมีความสัมพันธ์ใกล้ชิดที่สุดกับออร์นิโทมิมิด โทรโอโดนทิด และโอวิแรปทอริด ซึ่งพวกเขารวมไว้ในกลุ่มโอวิแรปทอโรซอเรีย (เนื่องจากพวกเขาพบว่ามานิแรปทอราซึ่งเป็นการจัดกลุ่มแบบดั้งเดิมของพวกมันนั้นไม่ถูกต้อง และอนุกรมวิธานระดับสูงของเทโรพอดกำลังเปลี่ยนแปลง) [ 29 ] [ 30 ]

Perle และผู้ร่วมเขียนคำอธิบายใหม่ของกะโหลก Erlikosaurus ในปี 1994 ยอมรับการใช้ชื่อพ้องของ Segnosauridae กับ Therizinosauridae และพวกเขาพิจารณาว่า therizinosaurs เป็นเทโรพอดกลุ่ม maniraptoran ซึ่งเป็นกลุ่มที่รวมถึงนกในปัจจุบันด้วย (เนื่องจากพวกเขาพบว่า Maniraptora เป็นกลุ่มที่ถูกต้องจากการวิเคราะห์ของพวกเขา) พวกเขายังได้หารือเกี่ยวกับสมมติฐานทางเลือกก่อนหน้านี้เกี่ยวกับความสัมพันธ์ของ therizinosaur และแสดงให้เห็นถึงข้อบกพร่องของสมมติฐานเหล่านั้น[ 31 ]ในปี 1995 Lev A. Nessov ปฏิเสธความคิดที่ว่า therizinosaurs เป็นเทโรพอด เขาพิจารณาว่าพวกมันเป็นกลุ่มที่แตกต่างกันภายใน Saurischia [ 32 ]ในปี 1996 Thomas R. Holtz Jr.พบว่า therizinosaurs จัดอยู่ในกลุ่มเดียวกับ oviraptorosaurs ในการวิเคราะห์ทางวิวัฒนาการของเทโรพอดกลุ่ม coelurosaurian [ 33 ]รัสเซลล์ตั้งชื่อกลุ่มนี้ว่าTherizinosauria ในปี 1997 [ 30 ]ในปี 1999 ซิงซูและเพื่อนร่วมงานได้บรรยายลักษณะของ Beipiaosaurusซึ่งเป็นเทริซิโนซอร์ขนาดเล็กดั้งเดิมจากประเทศจีน ซึ่งยืนยันว่ากลุ่มนี้อยู่ในกลุ่มเทโรพอดโคเอลูโรซอเรียน และความคล้ายคลึงกับโปรซอโรพอดนั้นวิวัฒนาการขึ้นอย่างอิสระ พวกเขาได้ตีพิมพ์แผนภูมิวิวัฒนาการ ฉบับแรก ที่แสดงความสัมพันธ์ทางวิวัฒนาการของ Therizinosauria และแสดงให้เห็นว่าBeipiaosaurusยังคงมีลักษณะของเทโรพอดและโคเอลูโรซอร์ดั้งเดิม ซึ่งเชื่อมโยงพวกมันกับเทริซิโนซอร์ การคงสภาพของโครงสร้างคล้ายขนนกในBeipiaosaurusยังชี้ให้เห็นว่าลักษณะนี้มีการกระจายตัวอย่างกว้างขวางในกลุ่มเทโรพอดมากกว่าที่เคยคิดไว้[ 12 ]
ในช่วงต้นศตวรรษที่ 21 มีการค้นพบกลุ่มเทริซิโนซอร์เพิ่มขึ้นอีกมากมาย รวมถึงบางกลุ่มที่อยู่นอกทวีปเอเชีย โดยกลุ่มแรกคือNothronychusจากทวีปอเมริกาเหนือในปี 2001 นอกจากนี้ยังมีการค้นพบกลุ่มพื้นฐานที่ช่วยให้เข้าใจวิวัฒนาการในช่วงแรกของกลุ่ม เช่นFalcariusในปี 2005 เทริซิโนซอร์ไม่ได้ถูกมองว่าเป็นสัตว์หายากหรือผิดปกติอีกต่อไป แต่มีความหลากหลายในด้านคุณลักษณะต่างๆ รวมถึงขนาด มากกว่าที่เคยคิดไว้ และการจัดประเภทเป็นเทโรพอดกลุ่มมานิแรปทอแรนก็ได้รับการยอมรับโดยทั่วไป[ 34 ] [ 35 ] [ 36 ] การจัดวาง Therizinosauria ภายใน Maniraptora ยังคงไม่ชัดเจน ในปี 2017 Alan H. Turner และเพื่อนร่วมงานพบว่าพวกมันจัดกลุ่มร่วมกับโอวิแรปทอโรซอร์ ในขณะที่ในปี 2009 Zanno และเพื่อนร่วมงานพบ ว่าพวกมันเป็นกลุ่มพื้นฐานที่สุดภายใน Maniraptora ซึ่งอยู่ระหว่าง Ornithomimosauria และAlvarezsauridae [ 37 ] [ 38 ]แม้จะมีฟอสซิลเพิ่มเติม แต่ความสัมพันธ์ภายในกลุ่มก็ยังไม่แน่นอนจนถึงปี 2010 เมื่อ Zanno ได้ทำการวิเคราะห์ทางวิวัฒนาการของ Therizinosauria อย่างละเอียดที่สุดในเวลานั้น เธออ้างถึงความยากลำบากในการเข้าถึง ความเสียหาย การสูญหายของตัวอย่างต้นแบบ ความหายากของซากกะโหลก และตัวอย่างที่แตกหักซึ่งมีองค์ประกอบที่ทับซ้อนกันน้อย เป็นอุปสรรคสำคัญที่สุดในการแก้ไขความสัมพันธ์ทางวิวัฒนาการภายในกลุ่ม ตำแหน่งของSegnosaurusและ Therizinosaurids อื่นๆ ในเอเชียได้รับผลกระทบจากปัจจัยเหล่านี้ Zanno ระบุว่าจำเป็นต้องมีตัวอย่างที่ได้รับการอนุรักษ์ไว้อย่างดีมากขึ้นและการค้นพบองค์ประกอบที่หายไปอีกครั้ง Zanno ยังได้แก้ไข Therizinosauroidea เพื่อไม่รวมFalcariusและคงไว้ในกลุ่ม Therizinosauria ที่กว้างกว่า ซึ่งกลายเป็นชื่อพ้องหลักของ Segnosauria [ 4 ]ในปี 2015 Segnosaurusยังคงเป็นหนึ่งในเทอริซิโนซอร์ที่รู้จักกันดีที่สุด ตามที่ Christophe Hendrickx และเพื่อนร่วมงานระบุ[ 13 ]
แผนภูมิวิวัฒนาการต่อไปนี้แสดงความสัมพันธ์ภายใน Therizinosauria ตามการศึกษาในปี 2013 โดย Hanyong Pu และเพื่อนร่วมงาน ซึ่งอิงตามการวิเคราะห์ของ Zanno ในปี 2010 โดยมีการเพิ่มสกุลพื้นฐานJianchangosaurus เข้าไปด้วย : [ 39 ]
ภูมิศาสตร์ชีวภาพโบราณ

ไดโนเสาร์เทอริซิโนซอร์ที่เก่าแก่ที่สุดคือFalcariusจากยุคครีเทเชียสตอนต้นของอเมริกาเหนือ ซึ่งแสดงให้เห็นว่ากระดูกเชิงกรานและฟันเป็นลักษณะแรกที่ถูกดัดแปลงไปจากรูปแบบทั่วไปของแมนิแรปทอแรนในเทอริซิโนซอร์ ซึ่งอาจสะท้อนถึงการเปลี่ยนผ่านจากการกินเนื้อเป็นการกินพืช[ 36 ] [ 40 ]เทอริซิโนซอร์ส่วนใหญ่พบในยุคครีเทเชียสของเอเชียและอเมริกาเหนือ และซากที่อาจพบในยุคและสถานที่อื่นๆ ยังเป็นที่ถกเถียงกันอยู่ เนื่องจากเทอริซิโนซอร์เป็นที่ทราบกันว่าอาศัยอยู่ทั่วทวีปใหญ่ลอราเซีย (ซึ่งประกอบด้วยสิ่งที่ปัจจุบันคืออเมริกาเหนือ ยุโรป และเอเชีย) Zanno จึงเสนอสองสถานการณ์สำหรับ การกระจาย ทางชีวภูมิศาสตร์ ของพวกมัน ในปี 2010 ความเป็นไปได้หนึ่งคือพวกมันแพร่กระจายผ่านการแยกตัวโดยที่เทอริซิโนซอร์มีอยู่ในพื้นที่ที่กลายเป็นเอเชียและอเมริกาเหนือก่อนการแยกตัวที่แบ่งพื้นที่เหล่านี้ในปลายยุคไทรแอสสิก ความเป็นไปได้อีกประการหนึ่งคือ เทอริซิโนซอร์กลุ่มพื้นฐานอาจแพร่กระจายระหว่างเอเชียและอเมริกาเหนือผ่านทางยุโรปหลังจากเหตุการณ์การแยกตัวของแผ่นดิน แต่ก่อนช่วงกลางยุคบาร์เรเมียนระหว่าง 132 ถึง 138 ล้านปีก่อนสะพานแผ่นดิน ชั่วคราว ได้เชื่อมต่ออเมริกาเหนือและยุโรป หลังจากนั้นแผ่นดินทั้งสองก็แยกออกจากกันอีกครั้ง ซึ่งอธิบายได้ว่าทำไมเทอริซิโนซอร์กลุ่มพื้นฐานอย่างBeipiaosaurusจากเอเชียและFalcariusจากอเมริกาเหนือจึงมีความแตกต่างทางสัณฐานวิทยาอย่างมาก แม้ว่าจะมีอายุร่วมสมัยกันก็ตาม การปรากฏตัวของNothronychus ซึ่งเป็นเทอริซิโนซอริเดที่พัฒนาแล้ว ซึ่งมีความสัมพันธ์ใกล้ชิดกับสกุลในเอเชียมากที่สุด ในอเมริกาเหนือในช่วง ยุค ทูโรเนียนของยุคครีเทเชียสตอนปลาย ตอนต้น ยังแสดงให้เห็นว่าน่าจะมีการแลกเปลี่ยนสัตว์ระหว่างอเมริกาเหนือและเอเชียผ่านทางสะพานแผ่นดินในช่วงปลายยุคครีเทเชียสตอนต้นก่อนหน้านั้น (ในช่วงยุคแอปเทียน / อัลเบียน ) ซึ่งพบเห็นได้ในกลุ่มไดโนเสาร์อื่นๆ อีกด้วย[ 4 ]
บรรพชีววิทยา

ในปี พ.ศ. 2522 และ พ.ศ. 2524 บาร์สโบลด์และเพิร์ลกล่าวว่ากระดูกฝ่าเท้าที่สั้นและใหญ่โตผิดปกติ และนิ้วเท้าที่กางออกขนาดใหญ่ บ่งชี้ว่าเซกโนซอรัสและญาติของมันไม่ได้ปรับตัวให้เคลื่อนที่ได้อย่างรวดเร็ว อาจเป็นเพราะวิถีชีวิตของพวกมันไม่ได้ต้องการเช่นนั้น บาร์สโบลด์และเพิร์ลแนะนำว่าพวกมันอาจเป็นสัตว์สะเทินน้ำสะเทินบก[ 18 ] [ 19 ]บาร์สโบลด์และมารียานสกาเห็นพ้องกันในปี พ.ศ. 2533 ว่าเท้าที่สั้นและกว้าง และลำตัวที่ใหญ่โตของกลุ่มนี้ บ่งชี้ว่าพวกมันเป็นสัตว์ที่เคลื่อนที่ช้า[ 28 ]ในปี 1988 พอลได้วาดภาพโครงกระดูก "เซกโนซอร์" ที่คล้ายกับโปรซอโรพอด (ซึ่งเป็นการผสมผสานของหลายสกุล) ในท่าทางสี่ขา[ 25 ]จากซากดึกดำบรรพ์ที่สมบูรณ์กว่าของอัลซาซอรัสและการเชื่อมต่อของกระดูกสันหลัง รัสเซลล์จึงสรุปในปี 1993 ว่าการฟื้นฟูโครงกระดูกของพอลนั้นไม่ถูกต้อง และแขนของเทอริซิโนซอร์นั้นยกขึ้นจากพื้นอย่างชัดเจน[ 41 ]ในปี 1995 เนสซอฟเสนอว่ากรงเล็บที่ยาวของเทอริซิโนซอร์นั้นใช้สำหรับการป้องกันตัวจากผู้ล่า และลูกอ่อนของพวกมันอาจใช้กรงเล็บในการเคลื่อนที่บนต้นไม้ตามลำต้นและยอดไม้ในลักษณะเดียวกับสลอธ[ 32 ]
ในการศึกษา กายวิภาค ภายในกะโหลกของErlikosaurusและเทอริซิโนซอร์อื่นๆ ที่มีกะโหลกสมอง ที่เก็บรักษาไว้ในปี 2012 Stephan Lautenschlager และเพื่อนร่วมงานพบว่าไดโนเสาร์เหล่านี้มีประสาทสัมผัสในการดมกลิ่น การได้ยิน และการ ทรงตัวที่พัฒนามาเป็นอย่าง ดี ประสาทสัมผัสสองอย่างแรกอาจมีบทบาทในการหาอาหาร การหลบหนีจากผู้ล่า และพฤติกรรมทางสังคม ประสาทสัมผัสเหล่านี้ยังได้รับการพัฒนามาเป็นอย่างดีในโคเอลูโรซอร์รุ่นก่อนๆ ดังนั้นเทอริซิโนซอร์อาจสืบทอดลักษณะเหล่านี้มาจากบรรพบุรุษที่กินเนื้อเป็นอาหารและใช้พวกมันเพื่อวัตถุประสงค์ในการหาอาหารที่แตกต่างกัน[ 42 ]ในการศึกษาการทำงานของกรงเล็บมือของเทอริซิโนซอร์ในปี 2014 Lautenschlager พบว่ากรงเล็บเหล่านี้ไม่ได้ถูกใช้สำหรับการขุด ซึ่งจะทำด้วยกรงเล็บเท้า เนื่องจากเช่นเดียวกับในมานิแรปเตอร์อื่นๆ ขนบนแขนขาหน้าจะขัดขวางการทำงานนี้ เขาไม่สามารถยืนยันหรือปฏิเสธได้ว่ากรงเล็บมืออาจถูกใช้เพื่อการป้องกัน การต่อสู้ การทรงตัวโดยการจับลำต้นของต้นไม้ระหว่างการกินใบไม้ สูง การแสดงออกทางเพศ หรือการจับคู่ระหว่างการผสมพันธุ์ เขาตัดความเป็นไปได้ที่พวกมันจะขุดโพรงออกไปเป็นส่วนใหญ่เนื่องจากขนาดของพวกมัน[ 43 ]

ไข่ไดโนเสาร์ที่มีตัวอ่อน ชนิด Dendroolithidaeจากชั้นหิน Nanchaoของจีนถูกระบุว่าเป็นของเทอริซิโนซอร์และได้รับการอธิบายโดย Martin Kundrát และเพื่อนร่วมงานในปี 2007 การพัฒนาของตัวอ่อนและข้อเท็จจริงที่ว่าไม่พบตัวเต็มวัยที่เกี่ยวข้องกับรังบ่งชี้ว่าลูกเทอริซิโนซอร์ที่ฟักออกมาเป็นสัตว์ที่สามารถเคลื่อนที่ได้ ตั้งแต่เกิดและสามารถออกจากรังเพื่อหาอาหารได้ด้วยตนเองโดยไม่ต้องพึ่งพาพ่อแม่ [ 44 ] [ 14 ]ในบทคัดย่อการประชุมปี 2013 Yoshitsugu Kobayashi และเพื่อนร่วมงานรายงานแหล่งทำรังของไดโนเสาร์เทอโรพอดจากชั้นหิน Javkhlant ของมองโกเลียซึ่งมี กลุ่มไข่อย่างน้อย 17 กลุ่มภายในพื้นที่ 22 x 52 เมตร (72 ฟุต x 171 ฟุต) แต่ละกลุ่มมีไข่ทรงกลม 8 ฟองที่มีพื้นผิวขรุขระ ไข่สัมผัสกันและเรียงตัวเป็นโครงสร้างวงกลมโดยไม่มีช่องเปิดตรงกลาง นักวิจัยระบุว่าไข่เป็นเดนโดรลิธิด ดังนั้นจึงเป็นเทอริซิโนซอเรียน แม้ว่าจะไม่พบเทอริซิโนซอรัสในชั้นหินนี้ แต่ชั้นหินนี้อยู่เหนือชั้นหินบายัน ชีเรห์ ซึ่งเป็นที่พบเซกโนซอรัส เออร์ลิโคซอรัส และเอนิกโมซอรัส การวางไข่หลายครอกแสดงให้เห็นว่าเทอริซิโนซอรัสบางชนิดทำรังเป็นกลุ่มเหมือนฮาโดรซอรัสโปรซอโร พอด ไททาโนซอรัสและนก ไข่ถูกพบในชั้นหินชั้น เดียว ซึ่งบ่งชี้ว่าไดโนเสาร์ทำรังในบริเวณนี้เพียงครั้งเดียวและไม่ได้แสดงความภักดีต่อสถานที่ (โดยกลับไปยังสถานที่เดิมเพื่อผสมพันธุ์เสมอ) [ 45 ]
อาหารและการให้อาหาร

ลักษณะที่ผิดปกติของเทอริซิโนซอรัสทำให้เกิดการตีความพฤติกรรมการกินของพวกมันได้หลายแบบ ไม่มีหลักฐานโดยตรงเกี่ยวกับอาหารของพวกมัน เช่น เนื้อหาในกระเพาะอาหารและร่องรอยการกิน ในปี 1970 อนาโตลี เค. โรจเดสต์เวนสกีเสนอว่าเทอริซิโนซอรัสซึ่งเป็นสมาชิกเพียงตัวเดียวของกลุ่มที่รู้จักในขณะนั้น ใช้กรงเล็บขนาดใหญ่ของมันในการเปิดรังปลวกหรือเก็บผลไม้จากต้นไม้[ 14 ]บาร์สโบลด์และเพิร์ลชี้ให้เห็นในปี 1979 และ 1980 ว่าลักษณะเฉพาะของพวกมันน่าจะสะท้อนถึงทิศทางการวิวัฒนาการที่แตกต่างจากเทอโรพอดทั่วไป ซึ่งหลายชนิดถือว่าเป็นนักล่าที่มีประสิทธิภาพและว่องไว ขากรรไกรที่บอบบาง ฟันและจะงอยปากที่เล็กและอ่อนแอ และเท้าที่สั้นและกะทัดรัด บ่งชี้ว่าพวกมันจะไม่ใช้อาวุธของเทอโรพอดตัวอื่นในการหาอาหาร แต่อาจล่าปลาเป็นอาหาร[ 18 ] [ 5 ]ในปี 1983 บาร์สโบลด์กล่าวว่าจะงอยปากแข็งที่ด้านหน้าของขากรรไกรและฟันที่อ่อนแอที่ด้านหลังเป็นลักษณะทั่วไปในไดโนเสาร์กินพืช แต่ไม่ใช่ในเทโรพอดกินเนื้อ และคาดการณ์ว่านี่อาจบ่งชี้ว่าเซกโนซอร์เปลี่ยนมากินพืช[ 3 ]ในปี 1984 พอลเสนอว่าพวกมันกินพืชเนื่องจากความคล้ายคลึงกันของกะโหลกกับไดโนเสาร์โปรซอโรพอดและออร์นิธิสเชียน ซึ่งรวมถึงจะงอยปากแข็ง แถวฟันที่ฝังอยู่ และชั้นวางที่ด้านข้างของขากรรไกรที่บ่งบอกถึงการมีแก้ม เช่นเดียวกับออร์นิธิสเชียน พวกมันจึงสามารถเก็บเกี่ยว จัดการ และเคี้ยวพืชได้อย่างซับซ้อน เขายังเสนอว่ากระดูกเชิงกรานมีใบมีดที่กางออกไปด้านข้างที่ด้านหน้าคล้ายกับของซอโรพอดเพื่อรองรับลำไส้ขนาดใหญ่ที่ใช้ในการหมักและแปรรูปอาหาร[ 23 ]นอร์แมนกล่าวในปี 1985 ว่าความเป็นไปได้ที่เซกโนซอรัสจะเป็นสัตว์น้ำกินปลาอาจอธิบายฟันที่เล็กและแหลมคม รวมถึงเท้าที่กว้างและอาจมีพังผืดได้ แต่เขาพบว่าเป็นเรื่องลึกลับว่าทำไมมันถึงมีจงอยปากที่เป็นเขา[ 24 ]

ในปี 1993 รัสเซลล์และดงพิจารณาว่าขนาดหัวที่เล็ก ปากทู่ และน้ำหนักตัวมากของเทอริซิโนซอร์นั้นสอดคล้องกับการกินพืช[ 29 ]ในปี 1993 และ 1997 รัสเซลล์เสนอว่าเทอริซิโนซอร์จะ "นั่ง" บนกระดูกเชิงกรานและพยุงร่างกายด้วยขาหลัง ในขณะที่ใช้แขนยาว กรงเล็บ และคอที่ยืดหยุ่นเพื่อเอื้อมไปถึงใบไม้จากต้นไม้และพุ่มไม้ด้วยปากของพวกมัน พวกมันสามารถเอื้อมไปได้สูงกว่านั้นในขณะที่ยืนและกินพืชด้วยสองขา ซึ่งคล้ายคลึงกับวิธีที่สัตว์เลี้ยงลูกด้วยนมกินพืชบางชนิดใช้แขนขาหน้าในการจัดการพืช รัสเซลล์พิจารณาว่าชาลิโคเทอเรสและสลอธพื้นดิน ที่สูญพันธุ์ไปแล้ว รวมถึงกอริลลามีการปรับตัวแบบบรรจบกับเทอริซิโนซอร์ เนื่องจากซากของเทอริซิโนซอร์มักพบในตะกอนที่สะสมในสภาพแวดล้อมของแม่น้ำและทะเลสาบ รัสเซลล์กล่าวว่าพวกมันอาจกินพุ่มไม้และต้นไม้ริมน้ำ[ 30 ] [ 41 ]จากการรวบรวมฟอสซิลในแหล่งฟอสซิลบิสเซกตีของอุซเบกิสถาน เนสซอฟเสนอในปี 1995 ว่าเทอริซิโนซอร์อาจเป็นส่วนหนึ่งของระบบนิเวศทางน้ำที่อุดมไปด้วยสารอาหาร แม้ว่าอาจจะเป็นทางอ้อม โดยการกินตัวต่อซึ่งกินซากสัตว์มีกระดูกสันหลังในน้ำเป็นอาหาร เขาพบว่าสิ่งนี้สอดคล้องกับข้อเสนอของโรจเดสต์เวนสกีที่ว่าเทอริซิโนซอร์อาจกินแมลงสังคม เป็นอาหาร [ 32 ]ในบทคัดย่อการประชุมปี 2006 ซารา เบิร์ชได้นำเสนอช่วงการเคลื่อนไหวที่อนุมานได้ในแขนของเทอริซิโนซอร์นีมองโก ซอรัส และสรุปว่าการเคลื่อนไหวโดยรวมที่ข้อต่อกลีนอยด์ -ฮิวเมอรัลที่ไหล่นั้นเป็นวงกลมโดยประมาณ และมีทิศทางไปด้านข้างและลงเล็กน้อย ซึ่งแตกต่างจากช่วงการเคลื่อนไหวที่เป็นรูปวงรีและมีทิศทางไปด้านหลังและลงของเทอโรพอดอื่นๆ ความสามารถในการยืดแขนไปข้างหน้าอย่างมากนี้อาจช่วยให้เทอริซิโนซาร์เอื้อมมือไปคว้าใบไม้ได้[ 46 ]
ในปี 2009 แซนโนและเพื่อนร่วมงานระบุว่าเทอริซิโนซอร์เป็นไดโนเสาร์เทอโรพอดที่ได้รับการยอมรับมากที่สุดว่าเป็นสัตว์กินพืช และได้ระบุลักษณะที่เกี่ยวข้องกับอาหารชนิดนี้ ซึ่งได้แก่ ฟันที่มีรอยหยักเล็ก หนาแน่น และหยาบ ฟันรูปหอกที่มีอัตราการเปลี่ยนฟันต่ำ จะงอยปากที่ด้านหน้าของขากรรไกร แถวฟันที่เว้าเข้าไปด้านในซึ่งบ่งบอกถึงแก้มที่อวบอิ่ม คอยาว กะโหลกศีรษะขนาดเล็ก ความจุของลำไส้ที่ใหญ่มาก ดังที่แสดงโดยเส้นรอบวงของซี่โครงที่ลำตัวและส่วนที่บานออกด้านนอกของกระดูกเชิงกราน และการสูญเสีย การปรับ ตัวที่เกี่ยวข้องกับการวิ่งในขาหลัง รวมถึงการพัฒนาของเท้าที่มีสี่นิ้วที่ใช้งานได้จริง Zanno และเพื่อนร่วมงานพบว่ากลุ่มสายพันธุ์ที่ฐานของ Maniraptora ได้แก่ Ornithomimosauria, Therizinosauria และ Oviraptorosauria มีหลักฐานโดยตรงหรือทางสัณฐานวิทยาเกี่ยวกับการกินพืช ซึ่งหมายความว่าอาหารประเภทนี้วิวัฒนาการขึ้นอย่างอิสระหลายครั้งในเทโรพอดโคเอลูโรซอเรียน หรือว่าสภาพดั้งเดิมของกลุ่มอย่างน้อยก็เป็นการกินพืชแบบเลือกได้ โดยการกินเนื้อจะเกิดขึ้นเฉพาะในมานิแรปทอแรนที่มีวิวัฒนาการสูงกว่า[ 38 ] Zanno และ Peter J. Makovicky พบในปี 2011 ว่าเทริซิโนซอร์และไดโนเสาร์กินพืชกลุ่มอื่นๆ ที่มีจะงอยปากและฟันที่ยังคงอยู่ ไม่สามารถสูญเสียฟันได้อย่างสมบูรณ์เนื่องจากพวกมันไม่มีกระเพาะบด (กึ๋น) และจำเป็นต้องใช้ฟันในการย่อยอาหาร และอาหาร ที่มีเส้นใยสูง แบบกินใบไม้ของเทริซิโนซอร์และ อาร์โคซอร์อื่นๆอาจขัดขวางวิวัฒนาการของจะงอยปากที่สมบูรณ์ด้วย[ 47 ] Lautenschlager พบในปี 2014 ว่ามือของเทอริซิโนซอร์จะต้องสามารถขยายขอบเขตของสัตว์ไปยังจุดที่หัวไม่สามารถเข้าถึงได้ หากใช้มือในการเล็มและดึงพืชลงมา อย่างไรก็ตาม ในสกุลที่มีการอนุรักษ์ทั้งส่วนคอและส่วนแขนขาหน้าไว้ คอมีความยาวเท่ากันหรือยาวกว่าแขนขาหน้า ดังนั้นการดึงพืชลงมาจึงสมเหตุสมผลก็ต่อเมื่อดึงส่วนล่างของกิ่งยาวลงมาเพื่อเข้าถึงส่วนต่างๆ ของต้นไม้ที่อยู่สูงเกินเอื้อม[ 43 ]

ในปี 2016 Zanno และเพื่อนร่วมงานระบุว่าโดยทั่วไปแล้วไดโนเสาร์กลุ่มเทอริซิโนซอร์นั้นจัดอยู่ในกลุ่มที่กินทั้งพืชและสัตว์ หรือกินพืชเป็นหลัก โดยมีแนวโน้มไปทางกินพืชมากกว่า แม้ว่าจะมีการใช้ลักษณะทางกายวิภาคต่างๆ มาสนับสนุนแนวคิดนี้ แต่รูปร่างของฟันนั้นถือว่าค่อนข้างเรียบง่ายและมีลักษณะพิเศษเฉพาะน้อยเมื่อเทียบกับไดโนเสาร์กินพืชชนิดอื่นๆ การเปลี่ยนแปลงเพียงเล็กน้อยที่พบ ได้แก่ ความสมมาตรที่เพิ่มขึ้นในฟันของErlikosaurusและการขยายตัวของเดนติเคิลในSegnosaurus Zanno และเพื่อนร่วมงานได้ระบุลักษณะที่ซับซ้อนและแปลกใหม่ในฟันกรามล่างของSegnosaurusรวมถึงการมีสันเพิ่มเติมและสันที่พับงอพร้อมขอบด้านหน้าเป็นเดนติเคิล ซึ่งบ่งชี้ว่าSegnosaurusมีกระบวนการย่อยอาหารในช่องปากที่สูงกว่าเทอริซิโนซอร์ชนิดอื่นๆ ลักษณะเหล่านี้รวมกันทำให้เกิดพื้นผิวที่ขรุขระและใช้สำหรับฉีกอาหารใกล้กับฐานของส่วนยอดฟัน ซึ่งเป็นเอกลักษณ์เฉพาะของSegnosaurusและชี้ให้เห็นว่ามันบริโภคแหล่งอาหารที่ไม่เหมือนใคร หรือใช้กลยุทธ์การกินอาหารที่เชี่ยวชาญเป็นพิเศษ เนื่องจากหลักฐานทางธรณีวิทยาหลายแหล่งแสดงให้เห็นถึง ส ปีชีส์เทริซิโนซอร์ที่อาศัยอยู่ในพื้นที่เดียวกันในเวลาเดียวกัน จึงเป็นไปได้ว่าการแบ่งส่วนนิเวศวิทยาระหว่างพวกมันอาจมีบทบาทต่อความสำเร็จทางวิวัฒนาการของกลุ่มนี้ ซึ่งได้รับการสนับสนุนจากSegnosaurusที่มีฟันที่เชี่ยวชาญเป็นพิเศษซึ่งมีอายุร่วมสมัยกับErlikosaurusที่มีฟันค่อนข้างไม่ชัดเจน แสดงให้เห็นถึงการแบ่งส่วนในการหาอาหาร การแปรรูป หรือทรัพยากร ข้อสรุปนี้ยังได้รับการเสริมความแข็งแกร่งด้วยความแตกต่างอย่างมากในมวลร่างกายโดยประมาณของเทริซิโนซอร์ที่อาศัยอยู่ในพื้นที่เดียวกันใน Bayan Shireh Formation ซึ่งสูงถึง 500% [ 6 ]
ในการศึกษาเกี่ยวกับการแบ่งส่วนนิเวศวิทยาในเทอริซิโนซอร์ผ่านการจำลองดิจิทัลในปี 2017 เลาเทนชลาเกอร์พบว่าขากรรไกรล่างของเซกโนซอรัสมีความเครียดต่ำที่สุดในระหว่างสถานการณ์การกินอาหารจากภายนอกเซกโนซอรัสและเออร์ลิโคซอรัสได้รับความช่วยเหลือจากปลายขากรรไกรล่างที่โค้งลงและบริเวณรอยต่อ และอาจรวมถึงจะงอยปากซึ่งเป็นที่ทราบกันดีว่าช่วยลดความเครียดและความตึง ในทางตรงกันข้าม ขากรรไกรล่างที่ตรงและยาวกว่าของเทอริซิโนซอร์ดั้งเดิม ซึ่งเป็นลักษณะเฉพาะของบรรพบุรุษโคเอลูโรซอเรียน มีความเครียดและความตึงสูงสุด การเคลื่อนไหวแบบดึงลงของหัวขณะจับพืชมีแนวโน้มมากกว่าการเคลื่อนไหวไปด้านข้างหรือขึ้นด้านบน แม้ว่าพฤติกรรมดังกล่าวจะมีแนวโน้มมากกว่าในเซกโนซอรัสและเออร์ลิโคซอรัสที่มีขากรรไกรที่ช่วยลดความเครียด ความแตกต่างของแรงกัดสัมพัทธ์ระหว่างSegnosaurusและErlikosaurus ที่อาศัย อยู่ในพื้นที่เดียวกัน แสดงให้เห็นว่า Segnosaurus น่าจะสามารถกินพืชที่แข็งกว่าได้ ในขณะที่ความแข็งแกร่งโดยรวมของ Erlikosaurus บ่งชี้ถึงความยืดหยุ่นที่มากกว่าในวิธีการกินอาหาร เนื่องจากระดับความเครียดอยู่ในระดับต่ำตลอดการจำลองการกินอาหาร Lautenschlager เห็นด้วยว่าสัตว์ทั้งสองชนิดปรับตัวให้เข้ากับวิธีการกินอาหารและการเลือกอาหารที่แตกต่างกันSegnosaurusปรับตัวให้ใช้ฟันในการหาหรือแปรรูปอาหารได้ดีกว่า ในขณะที่Erlikosaurusส่วนใหญ่ใช้จะงอยปากในการกัดกินและใช้กล้ามเนื้อคอในการหาอาหาร ความแตกต่างของขนาดระหว่างSegnosaurusและErlikosaurus (โดยคาดว่า Segnosaurus มีน้ำหนักมากกว่า Erlikosaurus) บ่งชี้ว่าผลกระทบเหล่านี้เพิ่มขึ้น และมีกลไกเพิ่มเติมในการแบ่งปันทรัพยากร เช่น ความสูงที่แตกต่างกัน เนื่องจากสัตว์ในกลุ่ม Therizinosaur อื่นๆ มีการแบ่งแยกกันมากขึ้นทั้งในด้านเวลาและพื้นที่ ปัจจัยอื่นๆ นอกเหนือจากการแข่งขันภายในกลุ่มอาจมีส่วนทำให้เกิดความแปรผัน เช่น การปรับตัวให้เข้ากับพืชพรรณที่แตกต่างกัน และการแข่งขันกับสัตว์กินพืชชนิดอื่นๆ[ 48 ]
ในปี 2018 Loredana Macaluso และเพื่อนร่วมงานชี้ให้เห็นว่าสะโพกของเทอริซิโนซอร์มีความแปลกประหลาด เนื่องจากแกนของกระดูกหัวหน่าวหมุนไปด้านหลัง ในขณะที่ส่วนปลายของกระดูกหัวหน่าวยื่นออกมาด้านหน้าอย่างมาก แม้ว่าลำไส้ขนาดใหญ่ที่เกี่ยวข้องกับการกินพืชจะสามารถดันแกนไปด้านหลังได้ แต่พวกเขาเสนอว่าส่วนปลายของกระดูกหัวหน่าวถูกยึดไว้ด้วยกล้ามเนื้อหายใจซึ่งมีความสำคัญต่อการหายใจแบบ cuirassal ซึ่งเป็นการหายใจด้วยถุงลม พิเศษ ซึ่งแสดงให้เห็นถึงความสำคัญของโหมดการหายใจนี้[ 49 ]ในการศึกษาเกี่ยวกับกล้ามเนื้อขากรรไกรในปี 2019 Ali Nabavizadeh สรุปว่าเทอริซิโนซอร์ส่วนใหญ่กินอาหารแบบ orthal คือการขยับขากรรไกรขึ้นลง และยกขากรรไกรขึ้นแบบ isognathously ซึ่งทำให้ฟันบนและฟันล่างของแต่ละข้างสบกัน (สัมผัสกัน) พร้อมกัน ตำแหน่งต้นกำเนิดและจุดสิ้นสุดของกล้ามเนื้อขากรรไกรยังเพิ่มความแข็งแรงให้กับการปิดขากรรไกรอีกด้วย[ 50 ] David J. Button และ Zanno พบในปี 2019 ว่าไดโนเสาร์กินพืชส่วนใหญ่มีวิธีการกินอาหารสองแบบที่แตกต่างกัน คือ การย่อยอาหารในลำไส้ ซึ่งมีลักษณะเด่นคือกะโหลกที่บอบบางและแรงกัดต่ำ หรือการย่อยอาหารในปาก ซึ่งมีลักษณะที่เกี่ยวข้องกับการย่อยอาหารอย่างละเอียดSegnosaurusพร้อมกับ ซอโรพอดกลุ่ม diplodocoidและ titanosaur, ออร์นิโทมิโมซอร์กลุ่ม deinocheirid และ ornithomimid และcaenagnathidsพบว่าอยู่ในกลุ่มแรก ในขณะที่Erlikosaurusมีความคล้ายคลึงกับซอโรโพโดมอร์ฟและออร์นิธิสเชียนบางกลุ่มมากกว่า ซึ่งบ่งชี้ว่าเทอริซิโนซอร์ทั้งสองชนิดนี้แยกจากกันในเชิงหน้าที่และครอบครองนิเวศวิทยาที่แตกต่างกัน[ 51 ]
สภาพแวดล้อมโบราณ

ฟอสซิลของSegnosaurusถูกค้นพบจากชั้นหิน Bayan Shireh ในมองโกเลีย ซึ่งมีอายุประมาณ 102–86 ล้านปีก่อน ในช่วง ยุค Cenomanianถึง Turonian ของยุคครีเทเชียสตอนปลาย โดยอาศัย การวิเคราะห์ ทางธรณีแม่เหล็กและการวัด U–Pbของแคลไซต์[ 52 ] [ 53 ]ซากดังกล่าวถูกพบในทรายสีเทาที่ยึดเกาะกันไม่ดี ประกอบด้วยหินกรวดหินกรวด และหินดินเหนียวสีเทา[ 28 ]ชั้นหิน Bayan Shireh อยู่เหนือชั้นหิน Baruunbayanและอยู่ใต้ชั้นหิน Javkhlant ตะกอนของชั้นหินเหล่านี้ถูกสะสมโดยแม่น้ำและทะเลสาบที่คดเคี้ยวบนที่ราบลุ่ม (ที่ราบที่ประกอบด้วยตะกอนที่สะสมโดยแม่น้ำบนที่สูง) ที่มีสภาพภูมิอากาศกึ่งแห้งแล้ง[ 54 ] [ 55 ]
เทอริซิโนซอร์เป็นเทอโรพอดที่อุดมสมบูรณ์ที่สุดในชั้นหินบายัน ชีเรห์ ในแง่ของความหลากหลายทางชีวภาพนอกจากเซกโนซอรัสแล้ว สมาชิกในกลุ่มยังรวมถึงเออร์ลิโค ซอรัส เอ นิ กโม ซอรัสและดูโอนิคัส [ 55 ] [ 56 ] เทอโรพอดอื่นๆ ได้แก่ ไทแรนโนซอรัส คานคูลู ออร์นิโทมิโมซอรัส การูดิ มิมัส และดรอเมโอซอรัสอะคิลโลเบเตอร์ [ 57 ] [ 58 ] [ 59 ] ไดโนเสาร์อื่นๆ ได้แก่ แอนคิโลซอรัสทาลารูรัส[ 52 ]ฮาโดรซอรัสโกบิฮาดรอส [ 60 ] ซอโรพอดเออร์เคตู [ 61 ] และเซราทอปเซียนกราซิลิเซราทอปส์[ 62 ]ไข่ไดโนเสาร์บางส่วนถูกระบุว่าเป็น Dendroolithidae รวมถึงรอยเท้าไดโนเสาร์และจระเข้ก็ถูกพบเช่นกัน ชั้นหินนี้มีความโดดเด่นในด้านความหลากหลายและความอุดมสมบูรณ์ของเต่า และสัตว์ไม่มีกระดูกสันหลัง ได้แก่ออสทราคอดและหอยน้ำจืด[ 52 ] [ 28 ] [ 55 ]ชั้นหิน Bayan Shireh อาจมีอายุร่วมสมัยกับชั้นหิน Iren Dabasuใน ภูมิภาค มองโกเลีย ในของจีน ซึ่ง พบฟอสซิลเทอริซิโนซอร์ที่คล้ายกับSegnosaurusและErlikosaurus ด้วย [ 63 ]
ดูเพิ่มเติม
ลิงก์ภายนอก
 สื่อที่เกี่ยวข้องกับSegnosaurusใน Wikimedia Commons
สื่อที่เกี่ยวข้องกับSegnosaurusใน Wikimedia Commons ข้อมูลที่เกี่ยวข้องกับSegnosaurusใน Wikispecies
ข้อมูลที่เกี่ยวข้องกับSegnosaurusใน Wikispecies
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เซกโนซอรัส
เซกโนซอรัส (Segnosaurus)เป็นสกุลของไดโนเสาร์เทอริซิโนซอริเดีย(Therizinosauridae ) ที่อาศัยอยู่ในบริเวณที่เป็นประเทศมองโกเลียตะวันออกเฉียงใต้ในปัจจุบัน...
ประวัติการค้นพบ
ในปี 1973 คณะสำรวจร่วมระหว่าง โซเวียต และมองโกเลียที่สำรวจ ชั้นหินบายันชีเรห์ ณ บริเวณอัมต์กาย ใน ทะเลทรายโกบี ทาง ตะวันออกเฉียงใต้ของมองโกเลีย ได้ค้นพบฟอสซิลซึ่งรวมถึงโครงกระดูกบางส่วนของ ไดโนเสาร์ ที่ไม่รู้จัก ในช่วงปี 1974 และ 1975...
คำอธิบาย
เซกโนซอรัส เป็นเทอริซิโนซอร์ขนาดใหญ่ที่คาดว่ามีความยาวประมาณ 6–7 เมตร (20–23 ฟุต) และมีน้ำหนักประมาณ 1.3 ตัน (1.4 ตันสั้น) [ 9 ] [ 10 ] [ 6 ] อย่างไรก็ตาม Campione & Evans ในปี 2020 คำนวณมวลร่างกายของมันที่ 4.17 เมตริกตัน (4.
ขากรรไกรล่างและฟันล่าง
ขากรรไกรล่างของ เซกโนซอรัส มีลักษณะต่ำและยาว แต่ค่อนข้างแข็งแรงและไม่มีรูปทรงที่แน่นอนเมื่อเทียบกับของ เออร์ลิโคซอรัส ซึ่งมี ลักษณะเพรียว บางกว่า ขากรรไกรล่างซีกขวาที่เกือบสมบูรณ์ (ครึ่งหนึ่งของขากรรไกรล่าง) มีความยาว 379 มม. (14.