อ่าน 38 นาที
คามาราซอรัส
คามาราซอรัส ( Camarasaurus )เป็นสกุลของไดโนเสาร์ซอโรพอดที่อาศัยอยู่ในช่วงปลายยุคจูราสสิกระหว่างประมาณ 155 ถึง 145 ล้านปีก่อน ใน ทวีปอเมริกาเหนือ...
คามาราซอรัส
| คามาราซอรัส | |
|---|---|
 | |
| CM 11338 โครงกระดูกที่เกือบสมบูรณ์ของCamarasaurus lentus | |
| การจำแนกทางวิทยาศาสตร์ | |
| อาณาจักร: | แอนิมอลเลีย |
| ไฟลัม: | คอร์ดาต้า |
| ระดับ: | สัตว์เลื้อยคลาน |
| กลุ่มสายพันธุ์ : | ไดโนเสาร์ |
| กลุ่มสายพันธุ์ : | ซอริสเชีย |
| กลุ่มสายพันธุ์ : | † ซอโรโพโดมอร์ฟา |
| กลุ่มสายพันธุ์ : | † ซอโรโปดา |
| กลุ่มสายพันธุ์ : | † มาโครนาเรีย |
| ตระกูล: | † Camarasauridae |
| อนุวงศ์: | † Camarasaurinae Cope , 1878 |
| ประเภท: | † Camarasaurus Cope, 1877 |
| ชนิดต้นแบบ | |
| † คามาราซอรัส ซูเปรมัส โคป, 1877 | |
| สายพันธุ์อื่นๆ | |
| |
| คำพ้องความหมาย | |
รายการ
| |
คามาราซอรัส ( Camarasaurus )เป็นสกุลของไดโนเสาร์ซอโรพอดที่อาศัยอยู่ในช่วงปลายยุคจูราสสิกระหว่างประมาณ 155 ถึง 145 ล้านปีก่อน ใน ทวีปอเมริกาเหนือ ได้รับการตั้งชื่อโดยเอ็ดเวิร์ดดริงเกอร์โคปในปี 1877 ประกอบด้วย 4 ส ปีชีส์ได้แก่ C. supremus , C. grandis , C. lentusและ C. lewisiชื่อคามาราซอรัสมีความหมายว่า ' กิ้งก่ามีโพรง'ซึ่งหมายถึงโพรงกลวงที่เรียกว่าเพลอโรซีล (pleurocoels ) ในกระดูกสันหลัง ส่วนคอ มีการขุดค้นพบซากดึกดำบรรพ์มากกว่า 530 ชิ้นในหินตะกอนของชั้นหินมอร์ริสัน (Morrison Formation)รวมถึงโครงกระดูกที่สมบูรณ์หลายชิ้น มันเป็นหนึ่งในไดโนเสาร์ที่ได้รับการศึกษาอย่างละเอียดที่สุด
Camarasaurus supremusเป็นซอโรพอดขนาดกลางถึงใหญ่คาดว่ามีความยาว 23 เมตร (75 ฟุต) และหนัก 42.3 ตัน ในขณะที่C. lentusมีความยาวเพียงประมาณ 15 เมตร (49 ฟุต) มันเป็นซอโรพอดที่ตัวใหญ่และมีโครงกระดูกซี่โครงที่กว้างเมื่อเทียบกับขนาดตัว กะโหลกศีรษะมีขนาดใหญ่และแข็งแรงกว่าซอโรพอดชนิดอื่น ๆ มีขากรรไกรที่ทรงพลังทำให้มีแรงกัดสูง ฟันมีรูปร่างคล้ายช้อนและเป็นคมตัดต่อเนื่อง มีฟัน 13-14 ซี่ในแต่ละด้านของขากรรไกรบนและ 13 ซี่ในแต่ละด้านของขากรรไกรล่างซึ่งจะถูกแทนที่อย่างรวดเร็วCamarasaurusได้รับการตั้งชื่อในช่วงสงครามกระดูก ซึ่ง เป็นการแข่งขันระหว่างนักบรรพชีวินวิทยา Edward Drinker Cope และOthniel Charles Marsh Marsh ได้จัดให้หลายชนิดอยู่ในสกุลMorosaurus ของเขา รวมถึงM. lentusและM. grandisซึ่งปัจจุบันถูกจัดให้อยู่ในสกุล Camarasaurus
คามาราซอรัสเป็นสมาชิกของกลุ่มแมคโครนาเรียนักบรรพชีวินวิทยาบางคนจัดกลุ่มคามาราซอรัสไว้กับกลุ่มอื่นๆ เช่นเทฮูเอลเชซอรัสและลูรินฮาซอรัสในวงศ์คามาราซอริเดในขณะที่บางคนถือว่ามันเป็นสมาชิกเพียงชนิดเดียวของวงศ์คามาราซอริเด คา มารา ซอ รัส อาจเป็นไดโนเสาร์ที่เลือกกินอาหาร โดยหากินที่ความสูง 2 ถึง 5 เมตร (6 ฟุต 7 นิ้ว ถึง 16 ฟุต 5 นิ้ว) ตัวอย่างที่พบมีตั้งแต่ตัวอ่อนไปจนถึงตัวที่อาจมีอายุถึง 40 ปี ซึ่งเป็นหนึ่งในอายุที่สูงที่สุดที่เคยรายงานสำหรับไดโนเสาร์ตัวหนึ่งคามาราซอรัสยังอาศัยอยู่ร่วมกับไดโนเสาร์อื่นๆ เช่นสเตโกซอรัสอัลโลซอรัสและบราคิโอซอรัส
ประวัติศาสตร์
การค้นพบและการตั้งชื่อ

คามาราซอรัสถูกค้นพบในช่วงสงครามกระดูกซึ่งเป็นความขัดแย้งระหว่างนักบรรพชีวินวิทยาชาวอเมริกันสองคน คือ โอ ธเนียล ชาร์ลส์ มาร์ชและเอ็ดเวิร์ด ดริงเกอร์ โคปซึ่งนำไปสู่การค้นพบฟอสซิลจำนวนมากในภาคตะวันตกของสหรัฐอเมริกาในช่วงปลายศตวรรษที่ 19 [ 1 ] กระดูกคา มาราซอรัส ชิ้น แรกถูกค้นพบโดยเดวิด บอลด์วิน หนึ่งในนักสะสมของมาร์ช ในช่วงต้นปี 1877 ในการ์เดนพาร์ ค รัฐโคโลราโดใกล้กับยอดเขาที่ปัจจุบันรู้จักกันในชื่อ 'Cope's Nipple' บอลด์วินพบกระดูกขณะไปเยี่ยมชมสถานที่ค้นพบฟอสซิลที่ชาวบ้านระบุว่าเป็นนกฟอสซิล และเขาตั้งใจที่จะเก็บรักษาไว้ให้มาร์ช (ซึ่งต่อมาได้อธิบายว่าเป็นฮัลโลปัสซึ่งเป็นจระเข้ ) ทั้งบอลด์วินและมาร์ชไม่ได้สนใจที่จะเก็บกระดูกไดโนเสาร์ ไม่นานหลังจากนั้น กระดูกเดียวกันนี้ก็ถูกค้นพบโดยครูท้องถิ่นชื่อโอราเมล ดับเบิลยู. ลูคัส ระหว่างการเดินทางล่าสัตว์ ลูคัสจึงเขียนจดหมายถึงโคป ซึ่งเสนอเงินให้เขาสำหรับการขุดฟอสซิล[ 1 ]ฟอสซิลชิ้นแรกที่โคปได้รับจากลูคัสคือขากรรไกรของเทอโรพอด ซึ่งโคปตั้งชื่อว่าLaelaps trihedrodonในสิ่งพิมพ์สั้นๆ[ 1 ]การจัดส่งครั้งที่สองจากลูคัสประกอบด้วยกระดูกสันหลังส่วนคอ 1 ชิ้น กระดูกสันหลังส่วนอก 3 ชิ้น และกระดูกสันหลังส่วนหาง 4 ชิ้น ของสัตว์ที่มีขนาดใหญ่กว่ามาก ซึ่งโคปอธิบายว่าเป็น "สัตว์ที่ใหญ่ที่สุดหรือมีขนาดใหญ่ที่สุดที่สามารถเคลื่อนที่บนบกได้ ซึ่งเรามีบันทึกอยู่" [ 2 ]


Cope ตั้งชื่อฟอสซิลเหล่านี้ว่าCamarasaurus supremusในสิ่งพิมพ์ที่เขียนขึ้นอย่างรวดเร็วในปี 1877 [ 2 ]ชื่อCamarasaurusหมายถึง "กิ้งก่าห้อง" มาจากภาษากรีกkamara ' ห้อง'และsauros ' กิ้งก่า'ซึ่งหมายถึงห้องภายในของกระดูกสันหลังที่ Cope กล่าวว่า "มีน้ำหนักเบากว่าเมื่อเทียบกับขนาดตัวของมันเมื่อเทียบกับสัตว์มีกระดูกสันหลังที่หายใจด้วยอากาศ" [ 3 ] Lucas และ Ira H. Lucas น้องชายของเขายังคงส่งกระดูกเพิ่มเติมของไดโนเสาร์ตัวใหม่นี้ให้ Cope จนถึงปี 1884 ซึ่งรวมกันแล้วเป็นโครงกระดูกส่วนใหญ่[ 4 ] : 484 [ 5 ] : 33 เป็นเวลาหลายทศวรรษหลังจากการค้นพบCamarasaurus supremusยังคงเป็นอนุกรมวิธานที่ค่อนข้างคลุมเครือเมื่อเทียบกับซอโรพอดที่สมบูรณ์และมีภาพประกอบมากกว่าที่ค้นพบโดยOthniel Marsh คู่แข่งของ Cope [ 6 ] : 251 คอลเลกชันของ Cope ได้รับมาโดยพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติอเมริกันและต่อมาได้รับการอธิบายโดยละเอียดมากขึ้นโดยHenry Fairfield Osbornและ Charles Craig Mook ในปี 1921 [ 6 ]กระดูกสันหลังแปดชิ้นที่ Cope อธิบายไว้ในตอนแรก ( วัสดุต้นแบบของC. supremusซึ่งเป็นพื้นฐานของสายพันธุ์) น่าจะมาจากสองตัว พวกมันถูกจัดทำเป็นแคตตาล็อกภายใต้หมายเลขตัวอย่างAMNH 5760 ซึ่งรวมถึง ฟอสซิล Camarasaurus อื่นๆ ที่ค้นพบโดย OW Lucas ด้วย[ 6 ] : 260 [ 4 ]
น่าจะเป็นช่วงต้นปี พ.ศ. 2422 จอห์น เอ. ไรเดอร์ได้สร้างภาพวาดโครงกระดูกที่ประกอบขึ้นใหม่ขนาดเท่าของจริงภายใต้การกำกับดูแลของโคป[ 7 ] [ 1 ]ภาพวาดมีความยาว 68 ฟุต (21 เมตร) และอิงจาก กระดูก ของคามาราซอรัส ที่ทราบแล้ว รวมถึงแขนขาของโมโรซอรัส แกรนดิส ที่ประกอบขึ้นใหม่ ซึ่งตีพิมพ์โดยมาร์ชเมื่อไม่กี่เดือนก่อนหน้านั้น[ 7 ]ในขณะนั้นยังไม่ค่อยมีใครรู้จักกะโหลกศีรษะ และเนื่องจากสัตว์เลื้อยคลานในปัจจุบันส่วนใหญ่เป็นสัตว์กินเนื้อจึงสันนิษฐานว่าคามาราซอรัส ก็เป็นเช่นเดียวกัน ดังนั้น กะโหลกศีรษะในสมมติฐานจึงมีฟันยาวสำหรับกินเนื้อ[ 1 ]ภาพวาดนี้เป็นการสร้างโครงกระดูกของซอโรพอดขึ้นใหม่ครั้งแรกเท่าที่เคยมีมา แม้ว่ามุกจะตีพิมพ์ในปี พ.ศ. 2457 ในขนาดที่เล็กกว่ามาก คือ 1/100 ของขนาดจริง[ 7 ] [ 8 ]ในปี พ.ศ. 2428 นักธรรมชาติวิทยาชาร์ลส์ เฟรเดอริค โฮลเดอร์ได้ตีพิมพ์ภาพวาดของคามาราซอรัสที่ มีชีวิต ซึ่งอาจเป็นการฟื้นฟูสภาพมีชีวิต ครั้งแรก ของซอโรพอด คอ หาง และแขนขาถูกวาดให้เรียวมาก และจมูกมีลักษณะงวงสั้น[ 9 ] [ 10 ]
โมโรซอรัสและสงครามกระดูก



เมื่อวันที่ 7 มีนาคม พ.ศ. 2420 วิลเลียม ฮาร์โลว์ รีดพนักงานรถไฟได้พบกระดูกฟอสซิลทางใต้ของสถานีโคโม รัฐไวโอมิง บนเนินเขาซึ่งต่อมากลายเป็นหนึ่งในแหล่งไดโนเสาร์ที่สำคัญแห่งแรกๆ นั่นคือโคโมบลัฟฟ์[ 11 ]รีดและวิลเลียม เอ็ดเวิร์ด คาร์ลิน เพื่อนร่วมงานของเขา ได้ส่งกระดูกบางส่วนไปให้มาร์ช ซึ่งรวมถึง กระดูกสันหลังส่วนหาง ที่เชื่อมต่อกันกระดูกสันหลังส่วนหลัง และกระดูกแขนขา 2 ชิ้น แม้ว่ารีดและคาร์ลินจะระบุว่ากระดูกเหล่านั้นเป็นของเมกาเธอ เรียม สัตว์จำพวกสลอธบนพื้นดิน แต่ มาร์ชก็ตระหนักถึงลักษณะของไดโนเสาร์และส่งซามูเอล เวนเดลล์ วิลลิสตัน ผู้ช่วยของเขา ไปดูแลและช่วยเหลือในการเก็บรวบรวม[ 5 ] : 34 [ 11 ] : 34 ในเดือนธันวาคม พ.ศ. 2420 มาร์ชได้อธิบายกระดูกเหล่านั้น (YPM 1901) ว่าเป็นสายพันธุ์ใหม่ของสกุลอะพาโทซอรัส ที่เขาตั้งขึ้นก่อนหน้านี้ คือA. grandis [ 12 ] [ 5 ] : 34
วิลลิสตันตกตะลึงเมื่อเห็นบริเวณนั้น โดยรายงานว่าพบกระดูกไดโนเสาร์ที่ "ได้รับการอนุรักษ์ไว้อย่างงดงาม" "กระจัดกระจายเป็นระยะทาง 6 หรือ 7 ไมล์" [ 11 ] : 28 ในช่วงปลายปี 1877 และต้นปี 1878 มาร์ชได้รับกระดูกหลายชุดจากเหมืองเดียวกัน ซึ่งปัจจุบันรู้จักกันในชื่อเหมือง YPM-Marsh 1 นอกจากกระดูกเพิ่มเติมจำนวนมากของA. grandisตัวแรกแล้ว ยังมีโครงกระดูกบางส่วนของตัวที่สองที่มีขนาดใกล้เคียงกัน (YPM 1905) ชิ้นส่วนของโครงกระดูกเหล่านี้ปะปนกัน และกระดูกบางชิ้นไม่สามารถระบุได้อย่างน่าเชื่อถือว่าเป็นของตัวใด[ 5 ] : 34 การจัดส่งยังประกอบด้วยกระดูกสันหลังส่วนล่าง (YPM 1900) ซึ่งมาร์ชได้อธิบายว่าเป็นสกุลและสายพันธุ์ใหม่Morosaurus imparในปี พ.ศ. 2421 [ 5 ] : 34 [ 13 ]ชื่อMorosaurusมาจากภาษากรีก Μόρος ( morosแปลว่า "โง่") ซึ่งหมายถึงขนาดสมองที่เล็กเมื่อเทียบกับขนาดตัว และσαυρος ( saurosแปลว่า "กิ้งก่า") [ 14 ]ต่อมาในปี พ.ศ. 2421 มาร์ชเขียนว่าMorosaurus ของเขา "ต้องเคลื่อนไหวเชื่องช้ามาก" และ "สมองของมันมีขนาดเล็กกว่าเมื่อเทียบกับสัตว์มีกระดูกสันหลังชนิดใดๆ ที่รู้จัก" [ 15 ] : 514 เขายังย้ายA. grandisไปอยู่ในสกุล Morosaurusโดยใช้ชื่อว่าM. grandisและตั้งชื่อMorosaurus อีกชนิดหนึ่งที่มีขนาดใหญ่กว่า ว่าM. robustusโดยอิงจากกระดูกเชิงกรานจากแหล่งขุดเดียวกัน[ 5 ] : 34 [ 15 ]ต่อมา Marsh ตระหนักว่าM. grandisและM. imparเป็นชนิดเดียวกัน แต่ไม่ได้เผยแพร่การค้นพบนี้[ 15 ]
Morosaurus lentusได้รับการตั้งชื่อโดย Marsh ในปี 1889 โดยอิงจากโครงกระดูกซอโรพอดวัยเยาว์ที่พบที่ Como Bluff [ 16 ] [ 17 ]ตัวอย่างต้นแบบ YPM 1910 ถูกจัดแสดงที่ ห้องฟอสซิล ของพิพิธภัณฑ์ Yale Peabodyในปี 1930 และเป็นหนึ่งในโครงกระดูกซอโรพอดที่เกือบสมบูรณ์ชุดแรกที่ถูกค้นพบ[ 18 ]ซึ่งประกอบด้วย: กระดูกสันหลังตั้งแต่กระดูกคอชิ้นที่สองถึงกระดูกหางชิ้นที่แปด กระดูกซี่โครงคอ กระดูกสะบักซ้าย กระดูกโคราคอยด์ขวา กระดูกต้นแขน กระดูกเรเดียสซ้าย กระดูกเชิงกราน กระดูกอิสเคียมซ้าย กระดูกหัวหน่าวขวา ขาหลังซ้ายและเท้าซ้าย และกระดูกหน้าแข้งขวา[ 19 ] [ 18 ]

หลังจากสงครามกระดูก นักบรรพชีวินวิทยายังพยายามจัดการกับอนุกรมวิธานของสกุลไดโนเสาร์จำนวนมากที่ตั้งชื่อโดย Marsh และ Cope [ 6 ] [ 16 ]ในปี 1898 Williston ได้รวมM. impar เข้า กับM. grandis [ 20 ]ซึ่งเป็นข้อเสนอแนะที่ได้รับการสนับสนุนจากผู้เขียนในภายหลัง[ 21 ] [ 6 ] [ 22 ]ในปี 1901 นักวิจัยชาวอเมริกันElmer Riggs สรุปว่าจาก Morosaurusห้าสายพันธุ์ที่ตั้งชื่อในขณะนั้น มีเพียงสามสายพันธุ์ที่ถูกต้อง ได้แก่M. grandis, M. lentusและM. agilis [ 23 ] Riggsได้รวมM. imparซึ่งเป็นสายพันธุ์ต้นแบบ เข้ากับM. grandisและยังเสนอแนะเพิ่มเติมว่าMorosaurusและCamarasaurusเป็นชื่อพ้อง[ 21 ]อย่างไรก็ตาม ข้อเสนอแนะที่ว่าMorosaurusและCamarasaurusเป็นชื่อพ้องของสกุลนั้นยังไม่ได้รับการยืนยันอย่างเป็นทางการจนกระทั่งมีการวิจัยในภายหลัง[ 16 ]ในงานวิจัยเรื่องCamarasaurus ในปี 1921 Osborn และ Mook ได้จัดให้Morosaurus เป็นชื่อพ้อง กับCamarasaurus [ 6 ]ซึ่งเป็นแนวคิดที่ได้รับการยอมรับตั้งแต่นั้นเป็นต้นมา[ 19 ] [ 24 ]ในปี 1925 นักวิจัยชาวอเมริกันCharles W. Gilmoreได้จัดให้C. grandis เป็นชื่อพ้อง กับC. impar โดยไม่ได้ตั้งใจ ทั้งๆ ที่ชื่อแรกถูกตั้งขึ้นก่อน[ 24 ] ปัจจุบัน M. imparและM. robustusถือเป็นชื่อพ้องกับC. grandis [ 25 ] [ 16 ]และM. agilisได้ถูกย้ายไปอยู่ในสกุลของตัวเองคือSmitanosaurus [ 16 ]
ไดโนเสาร์รัชครั้งที่สอง

หลังจากสิ้นสุดสงครามกระดูก สถาบันสำคัญหลายแห่งในภาคตะวันออกของสหรัฐอเมริกาได้รับแรงบันดาลใจจากภาพวาดและการค้นพบของ Marsh และ Cope เพื่อรวบรวมคอลเลกชันฟอสซิลไดโนเสาร์ของตนเอง[ 26 ] : 64–65, 105 การแข่งขันเพื่อจัดแสดงโครงกระดูกซอโรพอดตัวแรกนั้นเข้มข้นที่สุด โดยพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติอเมริกัน (AMNH) พิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติคาร์เนกี (CM) และพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติฟิลด์ (FMNH) ต่างส่งคณะสำรวจไปยังทางตะวันตกเพื่อค้นหาตัวอย่างซอโรพอดที่สมบูรณ์ที่สุด[ 26 ] : 1, 5 นำกลับมายังสถาบันของตน และจัดแสดงในห้องฟอสซิล[ 26 ] : 1, 5, 247 ในช่วงกลางถึงปลายทศวรรษ 1890 AMNH และ FMNH เป็นแห่งแรกที่ส่งคณะสำรวจไปค้นหาวัสดุซอโรพอดที่ Como Bluff และFruita รัฐโคโลราโดตามลำดับ[ 26 ] : 38–39, 117–118 วัสดุนี้ซึ่งส่วนใหญ่ประกอบด้วยกระดูกแขนขา ถูกอ้างถึงว่าเป็นMorosaurusและนำไปสู่การสร้างโครงสร้างมือและเท้า ของซอโรพอดขึ้นใหม่ [ 22 ] [ 21 ]ในช่วงทศวรรษ 1890 Osborn ยังเสนอแนะว่าMorosaurusเป็นคำพ้องความหมายของCamarasaurus [ 6 ] [ 27 ]ในปี 1899 Walter Grangerและ Peter Kaisen คนงานภาคสนามของ AMNH ได้ขุดพบกะโหลกศีรษะ ขากรรไกร และกระดูกสันหลังส่วนคอของ Camarasaurus ที่สมบูรณ์ในชั้นหินที่Bone Cabin Quarry รัฐไวโอมิงซึ่งเป็นการค้นพบกะโหลกศีรษะของ Camarasaurus ที่เกือบสมบูรณ์เป็นครั้งแรก [ 28 ] [ 29 ]ในปี พ.ศ. 2448 กะโหลกปูนปลาสเตอร์ที่สร้างขึ้นโดยอิง จากกะโหลก ของ Camarasaurusที่พบใน Bone Cabin Quarry และอิงจากกะโหลกที่ต่อมาถูกเรียกว่าBrachiosaurus [ 30 ] [ 31 ]ถูกนำมาติดตั้งบนโครงกระดูกของBrontosaurus [ 32 ] [ 29 ]ซึ่งนำไปสู่แนวโน้มของการติดตั้ง กะโหลกที่คล้ายกับ Camarasaurusบนร่างกายของ apatosaurus อย่างผิดพลาด นี่เป็นโครงกระดูกของซอโรพอดที่ถูกติดตั้งเป็นครั้งแรก[ 29 ]
ในช่วงเวลาเดียวกันกับ AMNH ทาง CM ก็ได้ดำเนินการสำรวจฟอสซิลของตนเองไปยังเหมืองหินในไวโอมิง ซึ่งได้ค้นพบ ตัวอย่าง Camarasaurus หลายชิ้น อย่างไรก็ตาม นักบรรพชีวินวิทยาในสมัยนั้นได้จัดฟอสซิลเหล่านี้ให้เป็นMorosaurusแทนที่จะเป็นCamarasaurus [ 33 ] [ 34 ] ในปี 1909 เอิร์ล ดักลาสนักล่าฟอสซิลชาวอเมริกันจาก CM ได้ค้นพบแหล่งสะสมฟอสซิลไดโนเสาร์ขนาดใหญ่ ณ สถานที่ซึ่งปัจจุบันคืออนุสรณ์สถานแห่งชาติไดโนเสาร์อันโด่งดังใกล้กับเมืองเจนเซน รัฐยูทาห์ [ 35 ] ในปี 1914 ระหว่างการขุดค้นที่อนุสรณ์สถาน ดักลาสได้ขุดพบกะโหลกและโครงกระดูกที่เกือบสมบูรณ์ของC. lentus วัยเยาว์ ที่ยังคงอยู่ในสภาพสมบูรณ์ ในปี 1925 โครงกระดูกนี้ได้รับการอธิบายโดยชาร์ลส์ ดับเบิลยู กิลมอร์ ซึ่งสนับสนุนข้ออ้างที่ว่าMorosaurusเป็นชื่อพ้องของCamarasaurusซึ่งเป็นตำแหน่งที่ได้รับการสนับสนุนจากผู้เขียนคนอื่นๆ[ 25 ] [ 19 ]โครงกระดูกนี้เป็นหนึ่งในตัวอย่างซอโรพอดที่ดีที่สุดที่วิทยาศาสตร์รู้จัก โดยเกือบทุกส่วนได้รับการอนุรักษ์ไว้ในข้อต่อ ก่อนปี 1922 ดักลาสพบ โครงกระดูก คามาราซอรัส อีกชิ้นหนึ่ง ที่อุทยานแห่งชาติไดโนเสาร์ แต่โครงกระดูกนั้นยังคงอยู่ในปลอกปูนปลาสเตอร์เป็นเวลาหลายปีพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติแห่งชาติได้รับโครงกระดูกและฟอสซิลอื่นๆ อีกหลายชิ้นเพื่อแลกกับฟอสซิลหลายชิ้น เงิน และเวลาในการทำงานร่วมกับกิลมอร์ จากนั้นตัวอย่างนี้ได้รับการจัดเตรียมต่อสาธารณะที่งานนิทรรศการครบรอบร้อยปีของเท็กซัสในปี 1936ก่อนที่จะถูกจัดแสดงในท่าทางเหมือนตายในห้องฟอสซิลของพิพิธภัณฑ์ในปี 1947 โครงกระดูกนี้กลายเป็นโครงกระดูกคามาราซอรัส ที่สมบูรณ์ที่สุดเป็นอันดับสอง ที่รู้จัก[ 36 ] [ 25 ]นักบรรพชีวินวิทยาเรียกโครงกระดูกนี้ว่าC. lentus เช่นกัน [ 19 ] [ 25 ]
ความสนใจที่กลับมาอีกครั้ง

ในปี พ.ศ. 2510 นักวิจัยชาวอเมริกันJames Jensenได้รวบรวมโครงกระดูกส่วนลำตัวของCamarasaurus ที่ได้รับการอนุรักษ์ไว้อย่างดีและสมบูรณ์ จากUncompahgre Hillในโคโลราโดตะวันตกซึ่งต่อมาได้ถูกนำไปเก็บไว้ที่มหาวิทยาลัย Brigham Youngภายใต้หมายเลขตัวอย่าง BYU 9740 [ 37 ] [ 38 ]โครงกระดูกนี้ยังคงไม่ได้รับการเตรียมการเป็นเวลาหลายปีและไม่ได้ถูกอธิบายจนกระทั่งปี พ.ศ. 2531 ซึ่ง Jensen ได้อธิบายว่ามันเป็นของสกุลและชนิดใหม่ของซอโรพอด คือCathetosaurus lewisi [ 38 ] [ 37 ] [ 39 ]ชื่อสกุลCathetosaurus lewisiมาจากภาษากรีก κάθετος ( kathetosหมายถึง "แนวตั้ง") ซึ่งหมายถึงความสามารถในการยืนตัวตรง และσαυρος ( saurosหมายถึง "กิ้งก่า") ชื่อเฉพาะนี้ตั้งขึ้นเพื่อเป็นเกียรติแก่ Arnold D. Lewis ผู้เตรียมฟอสซิลและนักทำงานภาคสนาม[ 14 ] [ 38 ]คำอธิบายดั้งเดิมในปี 1988 นี้สั้นมาก[ 38 ]แต่ John McIntosh และเพื่อนร่วมงานได้อธิบาย BYU 9740 อย่างละเอียดในปี 1996 โดยพวกเขาได้ระบุว่าCathetosaurus lewisiเป็นสายพันธุ์หนึ่งของCamarasaurus [ 37 ]ซึ่งเป็นความคิดเห็นที่ได้รับการสนับสนุนจากผู้เขียนคนอื่นๆ อีกมากมาย[ 25 ] [ 40 ] [ 41 ] ในปี2013นักบรรพชีวินวิทยาOctavio Mateus และ Emanuel Tschopp ได้โต้แย้งในบทคัดย่อการประชุมว่าCathetosaurusเป็นสกุลที่แตกต่างกัน[ 39 ]แต่สิ่งนี้ไม่ได้รับการสนับสนุนจากการวิจัยและการวิเคราะห์ทางวิวัฒนาการ ในภายหลัง (การศึกษาความสัมพันธ์ระหว่างสิ่งมีชีวิต) [ 41 ] [ 42 ] [ 40 ]
ในปี พ.ศ. 2535 โครงกระดูกของC. grandis [ 43 ] ที่เกือบสมบูรณ์และเชื่อมต่อกัน [ 44 ] ถูกเก็บรวบรวมโดยนักล่าฟอสซิล Jeffrie Parker และเพื่อนร่วมงานจากแหล่งใกล้กับเหมือง Bone Cabin [ 44 ]ตัวอย่างนี้ (GMNH-PV 101) ปัจจุบันอยู่ที่พิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติ GunmaในGunma ประเทศญี่ปุ่น [ 25 ] [ 45 ] ตัวอย่าง Camarasaurusที่ได้รับการอนุรักษ์ไว้อย่างดีอีกตัวอย่างหนึ่งถูกพบในปี พ.ศ. 2535 ที่เหมือง Howeในไวโอมิงโดยคนงานภาคสนามชาวสวิสที่ทำงานให้กับSauriermuseum Aathalในซูริก ประเทศสวิตเซอร์แลนด์โครงกระดูกนี้เป็นหนึ่งในโครงกระดูกที่รู้จักกันดีที่สุด โดยมีองค์ประกอบเกือบทุกส่วนเชื่อมต่อกันและมีรอยประทับของผิวหนังจากกะโหลกศีรษะและขาหลัง[ 46 ] [ 40 ]ตัวอย่าง SMA 002 ยังไม่ได้รับการระบุอย่างครบถ้วน แต่ได้รับการเสนอแนะว่าเป็นตัวอย่างของC. lewisi [ 40 ]ในปี พ.ศ. 2539 มีการบรรยายซากดึกดำบรรพ์ของCamarasaurus ที่แตกหักหลายชิ้น จากทางตะวันตกของเซาท์ดาโคตา[ 47 ]และนิวเม็กซิโก [ 48 ]ซึ่งขยายขอบเขตทางตะวันออกเฉียงเหนือและทางใต้ของสกุลนี้ ซากดึกดำบรรพ์จากนิวเม็กซิโกพบในSummerville Formationซึ่งบ่งชี้ว่าCamarasaurusอาศัยอยู่นอก Morrison Formation [ 48 ]ในปี พ.ศ. 2548 มีการค้นพบตัวอย่างที่อยู่เหนือสุด ซึ่งเป็นกะโหลกศีรษะและโครงกระดูกส่วนลำตัวที่ไม่สมบูรณ์ของCamarasaurusในเทือกเขา Little Snowy Mountainsของมอนแทนา[ 49 ]
คำอธิบาย

Camarasaurusเป็นซอโรพอดที่มีลำตัวคล้ายช้าง มีคอยาวที่ปลายสุดเป็นกะโหลกขนาดเล็กเมื่อเทียบกับขนาดตัว และกระดูกสันหลังของลำตัวและคอที่เบามาก ซึ่งแตกต่างอย่างสิ้นเชิงกับแขนขาที่ใหญ่โตและเป็นทรงกระบอก[ 50 ] : 11 เมื่อเทียบกับซอโรพอดชนิดอื่น ๆ มันค่อนข้างใหญ่โต มีซี่โครงกว้าง คอและหางค่อนข้างสั้น และกะโหลกมีขนาดใหญ่ เนื่องจากแขนขาหน้าค่อนข้างยาว มันจึงสูงกว่าที่ไหล่เล็กน้อยเมื่อเทียบกับที่สะโพก[ 51 ] [ 52 ]
Camarasaurusเป็นซอโรพอดขนาดกลางถึงขนาดใหญ่[ 25 ] [ 51 ]สายพันธุ์ที่พบมากที่สุดคือC. lentusมีความยาวประมาณ 15 เมตร (49 ฟุต) และC. grandisและC. lewisi น่าจะ มีขนาดใกล้เคียงกัน[ 53 ] [ 51 ]น้ำหนักตัวของC. grandis ที่โตเต็มวัยนั้น คาดว่าเฉลี่ยอยู่ที่ประมาณ 12.6 ตัน โดยมีน้ำหนักตั้งแต่ 8.3 ตันในตัวที่เล็กที่สุดไปจนถึง 16.6 ตันในตัวที่ใหญ่ที่สุด[ 51 ]ในปี 2020 John Foster ระบุว่าสายพันธุ์ล่าสุดC. supremusมีกระดูกยาวกว่าสายพันธุ์ที่เล็กกว่าประมาณ 50% [ 51 ]แม้ว่าC. supremusจะยังไม่เป็นที่รู้จักอย่างสมบูรณ์มากพอที่จะประมาณขนาดได้อย่างแม่นยำ แต่หากสัดส่วนร่างกายของมันเหมือนกับสายพันธุ์ที่เล็กกว่า มันจะมีความยาวเกือบ 23 เมตร (75 ฟุต) และหนัก 42.3 ตัน[ 51 ]นักบรรพชีวินวิทยาชาวอเมริกันGregory S. Paulในปี 2024 ได้ประมาณความยาวไว้ที่ 18 เมตร (59 ฟุต) และประมาณน้ำหนักไว้ที่ 24 ตันสำหรับสายพันธุ์นี้[ 52 ]
กะโหลก

กะโหลกมีขนาดใหญ่และแข็งแรงกว่าซอโรพอดชนิดอื่น ๆ โดยมีขากรรไกรบนและล่างขนาดใหญ่[ 54 ] [ 25 ] [ 51 ]ความยาวของกะโหลกแตกต่างกันไปในแต่ละตัว โดยมีความยาวตั้งแต่ประมาณ 46% ถึง 58% ของความยาวของกระดูกต้นขา[ 55 ]เช่นเดียวกับบราคิโอซอรัส ในยุคเดียวกัน รู จมูก ภายนอกมีขนาดใหญ่ขึ้น ส่งผลให้หน้าผากโค้ง[ 56 ] [ 24 ] : 353 ทั้งสองสกุลมีจมูกที่ชัดเจน[ 57 ]แต่กะโหลกของคามาราซอรัสมีลักษณะกลมและหน้าสั้นกว่า[ 51 ] [ 58 ] : 194 เมื่อมองจากด้านบน จมูกจะกลมและเรียวเล็กน้อย ซึ่งแตกต่างจากจมูกรูปสี่เหลี่ยมผืนผ้าของดิโพลโดคัสกะโหลกเกือบจะเป็นรูปสี่เหลี่ยมผืนผ้าเมื่อมองจากด้านหลังและสูงกว่ากว้าง[ 24 ] : 354
ช่องเปิดที่ใหญ่ที่สุดที่ทะลุผ่านกะโหลกศีรษะ ได้แก่ รูจมูกภายนอกเบ้าตา (ช่องเปิดตา) และช่องอินฟราเทมโพรัลซึ่งอยู่ด้านหลังและใต้เบ้าตา ช่องเปิดทั้งสามนี้มีขนาดใกล้เคียงกัน รูจมูกภายนอกมีรูปร่างเป็นวงรี และเบ้าตามีรูปร่างคล้ายหยดน้ำตา เรียวแหลมที่ปลายด้านล่าง เบ้าตามีวงแหวนสเคลอรัล ซึ่งเป็น วงแหวนของกระดูกแผ่นเล็กๆ รอบรูม่านตาช่องอินฟราเทมโพรัลมีรูปร่างคล้ายสามเหลี่ยมและเอียงไปด้านหลังเป็นมุม 30° มีช่องแอนทอร์บิทัลขนาดเล็กรูปร่างคล้ายลูกแพร์อยู่ระหว่างรูจมูกภายนอกและเบ้าตา ช่อง ซูพราเท มโพรัล มีขนาดเล็กกว่ามาก อยู่ในมุมด้านหลังของหลังคากะโหลกศีรษะและส่วนใหญ่หันขึ้นด้านบน ช่องเปิดนี้มีรูปร่างเป็นวงรีและวางตัวในแนวเฉียง[ 5 ] : 6 [ 24 ] : 364–365 [ 54 ]
กะโหลกศีรษะแสดงลักษณะ เฉพาะหลายประการ (ลักษณะที่ไม่พบในสกุลที่เกี่ยวข้อง) จากการตรวจสอบในปี 2547 พบว่ารวมถึงกระดูกน้ำตา (กระดูกที่ประกอบเป็นขอบด้านหน้าของเบ้าตา) ซึ่งเอียงไปข้างหน้า กระดูกควอดเรต ซึ่งเป็นกระดูกทรงกระบอกที่ด้านหลังของกะโหลกศีรษะ ไม่ถึงช่องอินฟราเทมโพรัล เนื่องจากส่วนที่หันขึ้นด้านบนนั้นสั้นกระดูกควอดราโตจูกัลซึ่งเป็นมุมด้านล่างด้านหลังของกะโหลกศีรษะ ยื่นขึ้นไปสัมผัสกับกระดูกสควาโมซัลพื้นผิวด้านข้างของขากรรไกรล่างมีร่องเฉียงวิ่งจากกระดูกซูแองกูลาร์ไปข้างหน้าและลงไปที่ขอบล่างของกระดูกเดนทารี [ 56 ] : 306
ตัวอย่างที่ทราบมีฟันสี่ซี่ในแต่ละพรีแม็กซิลลา (กระดูกด้านหน้าของขากรรไกรบน) ฟัน 9 ถึง 10 ซี่ในแต่ละแม็กซิลลา (กระดูกหลักของขากรรไกรบน) และฟัน 13 ซี่ในแต่ละขากรรไกรล่าง[ 5 ]ฟันมีรูปร่างคล้ายช้อนและมีขอบคมต่อเนื่อง ฟันมีความยาวเพิ่มขึ้นและสมมาตรมากขึ้นไปทางปลายจมูก ฟันเอียงไปข้างหน้าและโค้งงอเข้าด้านในเล็กน้อย ขอบด้านหน้าของฟันโค้งมากกว่าขอบด้านหลัง และฟันในขากรรไกรล่างตรงกว่าและแข็งแรงน้อยกว่าฟันในขากรรไกรบนเล็กน้อย ทำให้สามารถระบุได้ว่าฟันที่แยกออกมานั้นมาจากด้านซ้ายหรือด้านขวาของขากรรไกร และมาจากขากรรไกรบนหรือล่าง[ 19 ] : 490 [ 24 ] : 365–366 [ 40 ]
กระดูกส่วนหลังกะโหลก

กระดูกสันหลังประกอบด้วยกระดูกสันหลังส่วนคอ 12 ชิ้น กระดูกสันหลังส่วนหลัง 12 ชิ้น และกระดูกสันหลังส่วนสะโพก 5 ชิ้น[ 51 ] [ 59 ] : 81 หางประกอบด้วยกระดูกสันหลังส่วนหาง 53 ชิ้น โดยตัวอย่างสองชิ้นที่ยังคงมีหางที่สมบูรณ์ (CM 11338 และ GMNH 101) [ 60 ] : 5 ปริมาตรส่วนใหญ่ของกระดูกสันหลังส่วนหลังและส่วนคอประกอบด้วยถุงลมซึ่งเชื่อมต่อกับปอด ถุงลมเหล่านี้เติมเต็มโพรงขนาดใหญ่ในกระดูกสันหลังซึ่งเป็นที่มาของชื่อCamarasaurus ( ' กิ้งก่ามีห้อง' ) [ 61 ] : 14–15 [ 62 ]กระดูกสันหลังส่วนคอและส่วนอกมีลักษณะเว้า ( เว้าที่ด้านหลังและนูนที่ด้านหน้า) และมีโพรงขนาดใหญ่ที่ด้านข้างเรียกว่าเพลูโรซีล [ 24 ] : 370 [ 59 ] : 76 กระดูกสันหลังส่วนปลาย (ส่วนบนของกระดูกสันหลัง) ของบริเวณไหล่แยกออก และครึ่งซ้ายและขวาจะรวมกันเป็นรูปตัวยูเมื่อมองจากด้านหน้าหรือด้านหลัง ในบริเวณสะโพก กระดูกสันหลังส่วนปลายจะไม่แยกออก สั้น และมีรูปร่างคล้ายพัดเมื่อมองจากด้านหน้าหรือด้านหลัง[ 51 ] [ 5 ] : 37 กระดูกสันหลังส่วนปลายของกระดูกศักระข้อที่สองถึงข้อที่ห้ามักจะเชื่อมติดกัน[ 5 ] : 37 กระดูกสันหลังส่วนคอมีซี่โครงคอ ที่เรียวและยาวมาก ซึ่งทับซ้อนกับกระดูกสันหลังก่อนหน้าหลายชิ้น[ 24 ] : 374 [ 51 ]

กระดูกหัวไหล่ด้านซ้ายและขวาน่าจะเชื่อมต่อกันที่ด้านหน้าของลำตัวโดยกระดูกโคราคอยด์ [ 63 ] : 40 ปลายด้านบนของกระดูกสะบักขยายออก ในบริเวณสะโพก กระดูกหัวหน่าวมีขนาดใหญ่ ในขณะที่กระดูกอิสเคียมมีลักษณะเรียว โค้ง และปลายไม่ขยายออก พื้นผิวข้อต่อระหว่างกระดูกหัวหน่าวและกระดูกอิสเคียมมีความยาว[ 51 ] [ 5 ] : 37 แขนส่วนหน้าเรียว และกระดูกต้นแขนมีความยาวประมาณ 77% ของกระดูกต้นขาในทางตรงกันข้าม แขนส่วนหลังมีขนาดใหญ่ และกระดูกหน้าแข้งมีความยาวประมาณ 60% ของกระดูกต้นขาCamarasaurusเป็นหนึ่งในซอโรพอดไม่กี่ชนิดที่ยังคงรักษากระดูกข้อมือไว้ ซึ่งในสกุลนี้ประกอบด้วยกระดูกเพียงสองชิ้น คือ กระดูกอัลนาและกระดูกเรเดียล[ 25 ] [ 56 ] : 290 เช่นเดียวกับแมโครนาเรียนอื่นๆกระดูกฝ่ามือ ทั้งห้าชิ้น ยาว โดยกระดูกฝ่ามือชิ้นที่สามยาวถึงหนึ่งในสามของความยาวกระดูกต้นแขน[ 5 ] : 37 [ 56 ] : 290 ตามแบบฉบับของซอโรพอด กระดูกฝ่ามือตั้งตรงและเรียงตัวเป็นท่อ นิ้วมือลดขนาดลงอย่างมาก โดยนิ้วหัวแม่มือประกอบด้วยกระดูกนิ้ว สองชิ้น รวมถึงกรงเล็บที่เอียงไปด้านข้าง นิ้วที่เหลืออาจประกอบด้วยกระดูกนิ้วหนึ่งชิ้นและไม่มีกรงเล็บ[ 24 ] : 379–380 [ 56 ] : 290 ที่ข้อเท้าของเท้าหลังกระดูกส้นเท้ามีขนาดเล็กและกลม[ 5 ] : 37 เช่นเดียวกับซอโรพอดอื่นๆ เท้าหลังมีห้านิ้ว ประกอบด้วยกระดูกนิ้ว 2, 3, 4, 2 และ 1 ตามลำดับ นิ้วสามนิ้วแรกมีกรงเล็บโค้งงอและแบนราบจากด้านหนึ่งไปอีกด้านหนึ่ง[ 56 ] : 295
เนื้อเยื่ออ่อนและรอยเท้า

รอยเท้าแสดงให้เห็นว่าเท้าหลังของซอโรพอดได้รับการรองรับด้วยแผ่นรองเนื้อขนาดใหญ่คล้ายกับของช้าง[ 64 ] : 146 แผ่นรองเนื้อดังกล่าวไม่มีอยู่ในเท้าหน้า ดังที่แสดงโดยรอยประทับของผิวหนังที่เก็บรักษาไว้ใกล้กับ พื้น ผิวด้านฝ่ามือ (ด้านหลัง) ของกระดูกฝ่ามือของ ตัวอย่าง Camarasaurus SMA 0002 ("ET") รอยประทับของผิวหนังยังบ่งชี้ว่านิ้วที่สองถึงสี่ของเท้าหน้าถูกห่อหุ้มด้วยเนื้อเยื่อ รอยประทับของผิวหนังบางส่วนยังคงได้รับการเก็บรักษาไว้บนขาหลังของ "ET" ซึ่งส่วนใหญ่แสดงเกล็ดรูปหกเหลี่ยมที่มีเส้นผ่านศูนย์กลางระหว่าง 6 ถึง 18 มม. (0.24 ถึง 0.71 นิ้ว) [ 46 ] : 42, 54 ในปี 2015 Tschopp และเพื่อนร่วมงานได้สร้างแบบจำลองของเท้าหน้าและเท้าหลังโดยอิงจาก "ET" เพื่อสร้างรอยเท้าสมมุติ โดยแสดงให้เห็นว่ารอยเท้าหลังมีขนาดใหญ่กว่ารอยเท้าหน้าเกือบสี่เท่า รอยเท้าฟอสซิลของซอโรพอดอาจเป็น "รอยเท้าแคบ" หรือ "รอยเท้ากว้าง" ขึ้นอยู่กับว่ารอยเท้าอยู่ใกล้กับเส้นกลางของรอยเท้ามากแค่ไหนCamarasaurusอาจมีรอยเท้าขนาดกลาง จนถึงขณะนี้ยังไม่มีการระบุรอยเท้าที่แน่ชัดว่าเป็นของCamarasaurus [ 46 ] : 54–55
การจำแนกประเภทและชนิด

ในปี พ.ศ. 2420 Cope ได้อธิบายCamarasaurusว่าเป็นสกุลไดโนเสาร์ชนิดใหม่ และเปรียบเทียบกับสกุลต่างๆ ที่ต่อมาจะถูกจัดอยู่ในกลุ่มซอโรพอด ซึ่งรวมถึงCetiosaurus , BothriospondylusและOrnithopsis [ 65 ]หนึ่งปีต่อมา ในการอธิบายAmphicoelias Cope ได้ใช้ตระกูลCamarasauridaeและAmphicoelidae โดยตระกูลแรกมีกระดูกสันหลัง แบบopisthocoelous (กระดูกสันหลังที่เว้าด้านหลัง) และตระกูลหลังมีกระดูกสันหลังแบบ amphicoelous (กระดูกสันหลังที่เว้าทั้งสองด้าน) [ 66 ]ในปี พ.ศ. 2420 เช่นกัน Marsh ได้ตั้งชื่อสกุลMorosaurusซึ่งเขาจัดไว้ในAtlantosauridaeร่วมกับสกุลAtlantosaurusและApatosaurus ของ เขา[ 13 ]ต่อมาในปีเดียวกันนั้น มาร์ชได้ตั้งชื่อกลุ่มย่อยใหม่ว่า Sauropoda โดยมี Atlantosauridae เป็นวงศ์เดียว เนื่องจาก "พวกมันแตกต่างจาก Dinosauria ทั่วไปอย่างมาก" [ 15 ]ต่อมามาร์ชได้จัดจำแนกMorosaurusไว้ในวงศ์ซอโรพอดของตัวเอง คือMorosauridae [ 67 ] ตั้งแต่ นั้น มา Morosaurusและ Morosauridae ได้ถูกจัดให้เป็นชื่อพ้องกับCamarasaurusและ Camarasauridae ตามลำดับ เนื่องจากชื่อหลังได้รับการตีพิมพ์ก่อนหน้านี้[ 68 ]
Camarasauridae ถูกนำมาใช้กันอย่างแพร่หลายในอดีต[ 69 ] : 1555 ตัวอย่างเช่น ในการทบทวนในปี 1990 John Stanton McIntoshได้จำแนกวงศ์ย่อย 2 วงศ์ภายในวงศ์นี้ ได้แก่Camarasaurinaeซึ่งประกอบด้วยCamarasaurus , Aragosaurus , EuhelopusและTienshanosaurusและOpisthocoelicaudiinaeซึ่งประกอบด้วยOpisthocoelicaudiaและChondrosteosaurus [ 7 ]อย่างไรก็ตาม ปัจจุบันวงศ์นี้มักถูกจำกัดไว้เฉพาะCamarasaurusเท่านั้น ทำให้ผู้วิจัยบางคนปฏิเสธว่าวงศ์นี้ซ้ำซ้อน[ 69 ] [ 70 ]
ตั้งแต่ปี 1998 Camarasaurusโดยทั่วไปจัดอยู่ใน กลุ่มสมาชิก พื้นฐาน (แยกตัวออกมาในช่วงต้น) ของกลุ่มซอโรพอดMacronariaซึ่งตรงข้ามกับกลุ่มซอโรพอดหลักอีกกลุ่มหนึ่งคือDiplodocoideaภายใน Macronaria นั้นCamarasaurusถูกจัดไว้นอกกลุ่มTitanosauriformesซึ่งประกอบด้วยสกุลส่วนใหญ่[ 71 ] [ 57 ] : 461 หรืออีกทางหนึ่ง ชื่อCamarasauromorphaถูกใช้เพื่อรวมCamarasaurusและ Titanosauriformes เข้าด้วยกัน [ 72 ] : 9 การศึกษาหลายชิ้นใช้ทั้งสองชื่อ โดย Camarasauromorpha ไม่รวมสมาชิกพื้นฐานบางส่วนของ Macronaria [ 57 ] : 461 [ 73 ] Camarasauromorpha ยังไม่ได้รับการยอมรับอย่างเป็นสากล[ 73 ]
ด้านล่างนี้คือแผนภูมิวิวัฒนาการแบบง่ายจาก Pedro Mocho และเพื่อนร่วมงาน (2014) ซึ่งพบว่า Camarasauridae ประกอบด้วยCamarasaurus , TehuelchesaurusและLourinhasaurusซึ่งเป็นซอโรพอดสามสกุลในยุคจูราสสิกตอนปลาย: [ 74 ]

ชนิดที่ได้รับการยอมรับในปัจจุบัน
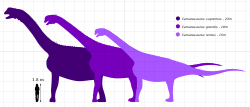
โดยทั่วไปมีการจำแนกออกเป็น 4 ชนิด ได้แก่C. supremus , C. grandis , C. lentusและC. lewisi [ 42 ] C. supremusซึ่งตั้งชื่อโดย Cope ในปี 1877 เป็นชนิดต้นแบบ (ชนิดที่ใช้เป็นพื้นฐานของสกุล) C. grandisได้รับการตั้งชื่อในปี 1877 และC. lentusในปี 1889 C. lewisiเดิมทีถูกอธิบายว่าเป็นสกุลที่แตกต่างกันCathetosaurusในปี 1988 แต่ถูกจัดประเภทใหม่เป็นชนิดของCamarasaurusในปี 1996 [ 38 ] [ 59 ]นักวิจัยบางคนเสนอแนะว่าCathetosaurusอาจได้รับการฟื้นฟูให้เป็นสกุลที่แตกต่างกัน[ 75 ] [ 42 ]ในขณะที่คนอื่นๆ เสนอแนะว่าC. lewisiควรถูกจัดให้อยู่ในC. grandisหรือC. sp. (ของชนิดที่ไม่แน่นอน) [ 25 ] [ 76 ]
ตัวอย่าง Camarasaurusส่วนใหญ่ไม่สามารถระบุชนิดได้[ 77 ]การระบุชนิดมีความซับซ้อนเนื่องจากความแปรปรวนภายในชนิดและการกำหนดอายุสัมพัทธ์ของสถานที่ที่ไม่แม่นยำ[ 25 ] : 370–371 สองชนิดที่พบได้บ่อยที่สุด คือC. grandisและC. lentusสามารถแยกแยะได้จากส่วนโค้งประสาทของกระดูกสันหลังส่วนอกด้านหน้าสุด ซึ่งกว้างกว่าในC. lentusและปลายของหนามประสาทของกระดูกสันหลังส่วนหางด้านหน้าสุด ซึ่งกว้างกว่าในC. grandis C. supremusมีส่วนโค้งประสาทกว้างเหมือนC. lentusแต่มีปลายหนามประสาทกว้างเหมือนC. grandisชนิดนี้มีขนาดใหญ่กว่ามาก ในC. lewisiการแตกแขนงของหนามประสาทไม่ได้จำกัดอยู่เฉพาะบริเวณไหล่ แต่เกิดขึ้นตั้งแต่กระดูกสันหลังส่วนคอที่สามถึงกระดูกสันหลังส่วนอกที่สิบสอง[ 25 ] [ 77 ]
ในปี 2548 Takehito Ikejiri ได้โต้แย้งว่าสายพันธุ์ทั้งสี่ที่จัดตั้งขึ้นนั้นแยกจากกันตามช่วงเวลา[ 25 ]สายพันธุ์ที่เก่าแก่ที่สุดC. grandisน่าจะเกิดขึ้นในช่วง Kimmeridgian และตามมาด้วยC. lentusในช่วงปลาย Kimmeridgian ส่วนC. supremusน่าจะเกิดขึ้นเป็นหลักในช่วง Tithonian ที่ส่วนบนสุดของ Morrison Formation [ 25 ]ในปี 2560 Cary Woodruff และ Foster ได้เตือนว่าการกำหนดอายุสัมพัทธ์ของสถานที่ต่างๆ ภายใน Morrison Formation ยังไม่แน่นอน ซึ่งอาจบั่นทอนลำดับของสายพันธุ์ที่เสนอไว้[ 78 ]นอกจากนี้ อาจมีการแยกทางภูมิศาสตร์ระหว่างสายพันธุ์ และC. supremusดูเหมือนจะถูกจำกัดไว้เฉพาะส่วนตะวันออกของ Morrison Formation ในโคโลราโดตอนใต้และโอคลาโฮมาตะวันตก[ 25 ] [ 78 ]อย่างไรก็ตาม Foster ตั้งข้อสังเกตในปี 2563 ว่าไม่มีรูปแบบการกระจายตัวที่ชัดเจนของสายพันธุ์ต่างๆ[ 51 ] : 274
ชนิดพันธุ์ที่ได้รับการยอมรับก่อนหน้านี้และชื่อพ้อง



อีกสี่สายพันธุ์ที่ได้รับการตั้งชื่อโดย Cope ระหว่างปี พ.ศ. 2420 ถึง พ.ศ. 2422 ปัจจุบันถือว่าเป็นคำพ้องของCamarasaurus supremus : Amphicoelias latus , Caulodon Diversidens , Caulodon leptoganusและCamarasaurus leptodirus [ 25 ] [ 79 ] [ 6 ]ในทำนองเดียวกัน สองสายพันธุ์ที่ตั้งชื่อโดยบึงในปี พ.ศ. 2421 และ พ.ศ. 2439 ถือเป็นคำพ้องความหมายของCamarasaurus grandis : Morosaurus imparและPleurocoelus montanus [ 25 ] [ 80 ] : 275 สองสายพันธุ์ที่อธิบายมาจากกระดูกสันหลังที่พบในอนุสาวรีย์แห่งชาติไดโนเสาร์Uintasaurus douglassiและCamarasaurus annaeปัจจุบันถือว่าเป็นคำพ้องของCamarasaurus lentus [ 81 ] [ 25 ] [ 82 ]
สายพันธุ์อื่นๆ ที่เคยถูกจัดอยู่ในสกุลCamarasaurusได้ถูกย้ายไปยังสกุลอื่นๆ แล้วMorosaurus agilisได้รับการตั้งชื่อในปี พ.ศ. 2432 โดย Marsh โดยอาศัยกะโหลกศีรษะบางส่วนและกระดูกสันหลัง 3 ชิ้น[ 83 ]แต่ได้รับการอธิบายว่าเป็นสกุลใหม่ของไดเครโอซอริเดภายใต้ชื่อSmitanosaurusในปี พ.ศ. 2563 [ 16 ] Morosaurus marcheiได้รับการตั้งชื่อในปี พ.ศ. 2440-2491 โดยHenri Sauvageโดยอาศัยฟันและกระดูกสันหลังส่วนหางที่ไม่สมบูรณ์ที่พบในAlcobaça Formationของลิสบอน ประเทศโปรตุเกส [ 84 ] ต่อมาพบว่ากระดูกสันหลังนั้นเป็นของเทโรพอด ในขณะที่ฟันนั้นมาจากซอโรพอดแมคโครนาเรียนที่ไม่สามารถระบุชนิดได้[ 85 ] : 259–260 Apatosaurus alenquerensisได้รับการตั้งชื่อในปี พ.ศ. 2490 โดย Lapparent และ Zbyszewski โดยอิงจากโครงกระดูกส่วนลำตัวบางส่วนจากLourinha Formationใน Lourinha ประเทศโปรตุเกส[ 86 ] McIntosh ได้จัดให้อยู่ในสกุล Camarasaurus อย่างไม่เป็นทางการ ในปี พ.ศ. 2533 [ 7 ]แต่ได้รับสกุลใหม่ในปี พ.ศ. 2541 คือLourinhasaurus [ 74 ]
บรรพชีววิทยา
การให้อาหาร

Camarasaurusเป็นสัตว์กินพืชที่น่าจะหากินในระดับความสูงปานกลาง 2 ถึง 5 เมตร (6 ฟุต 7 นิ้ว ถึง 16 ฟุต 5 นิ้ว) [ 51 ] : 274 มันอาจสามารถยืนด้วยขาหลังเพื่อเอื้อมถึงพืชพรรณที่สูงขึ้นได้ ดังที่ระบุโดยลักษณะทางกายวิภาค เช่น กระดูกสันหลังส่วนหางที่สั้น[ 87 ]ในปี 1998 Anthony Fiorilloได้วิเคราะห์ หลุมและรอยขีดข่วน ขนาด เล็ก บนพื้นผิวฟันและสรุปว่าCamarasaurus ตัวเต็มวัย กินอาหารที่หยาบกว่าDiplodocus ในยุคเดียวกัน ในทางตรงกันข้าม Camarasaurusวัยเยาว์ดูเหมือนจะกินอาหารอ่อนชนิดเดียวกับDiplodocusตัว เต็มวัย [ 88 ]สัตว์เลี้ยงลูกด้วยนมกินพืชสมัยใหม่ที่มีจมูกกลมมักเลือกกินอาหาร โดยกินเฉพาะพืชบางชนิดที่มีปริมาณน้อยแต่มีคุณค่าทางโภชนาการสูง ในขณะที่สายพันธุ์ที่มีจมูกกว้างมักไม่เลือกกินอาหาร โดยกินอาหารที่มีคุณค่าทางโภชนาการต่ำแต่มีปริมาณมากในปริมาณมาก จากการสังเกตนี้ จอห์น วิทล็อค ได้โต้แย้งในปี 2011 ว่าCamarasaurusและBrachiosaurus ที่มีจมูกกลม เป็นผู้เลือกกินอาหาร ในขณะที่ diplodocids และrebbachisaurids ที่มีจมูกเหลี่ยม เป็นผู้กินอาหารปริมาณมาก[ 89 ] [ 87 ]อาหารของCamarasaurus ตัวเต็มวัย อาจประกอบด้วยใบของต้นสน เช่น ใบของCheirolepidiaceae ที่สูญพันธุ์ไปแล้ว และใบแปะก๊วย [ 87 ] ในปี 2016 มาร์ค ฮัลเลตต์ และแมตต์ เวเดล แนะนำว่ากรวยตัวเมียของaraucariansรวมถึงเรซินอาจเป็นส่วนเสริมในอาหารของมัน[ 87 ]

Camarasaurusน่าจะมีแรงกัดที่ทรงพลังกว่าซอโรพอดชนิดอื่นๆ เนื่องจากกระบวนการโคโรนอยด์ ขนาดใหญ่มาก (ส่วนที่ยื่นขึ้นของขากรรไกรล่าง) และช่องเหนือขมับที่ให้พื้นผิวยึดเกาะที่กว้างขวางสำหรับกล้ามเนื้อบดเคี้ยวขนาดใหญ่ (กล้ามเนื้อดึงขากรรไกรล่างภายนอก) [ 54 ]การศึกษาในปี 2016 โดย David Button และเพื่อนร่วมงานประเมินว่าแรงกัดของCamarasaurusสูงกว่าDiplodocus เกือบสี่เท่า แรงกัดสูงสุดอยู่ที่ส่วนหลังของแถวฟัน ซึ่งคาดว่าสูงถึง 1978 นิวตัน [ 90 ] : 900 [ 91 ] : 182 โครงสร้างที่แข็งแรงของกะโหลกศีรษะยังบ่งชี้ว่ามันสามารถทนต่อแรงกดดันที่มากกว่าซอโรพอดชนิดอื่นๆ ในระหว่างการกินอาหารได้[ 54 ] [ 90 ] Per Christiansen ในบทความปี 2000 แนะนำว่าCamarasaurusปรับตัวให้กัดกินพืช แต่ไม่ได้กวาดใบไม้เหมือนDiplodocus หรือ Brachiosaurus [ 54 ] ฟันบนและล่างดูเหมือนจะเข้ากันพอดี ในการศึกษาปี 1994 Jorge Calvoแนะนำว่าCamarasaurusสามารถบดอาหารด้วยฟันของมันโดยการขยับขากรรไกรไปมา ทำให้สามารถแปรรูปอาหารได้ในระดับหนึ่งก่อนที่จะกลืนกิน[ 92 ] [ 54 ]

เช่นเดียวกับไดโนเสาร์ชนิดอื่นๆCamarasaurusเปลี่ยนฟันอย่างต่อเนื่อง และใต้ฟันที่งอกออกมาแต่ละซี่จะมีฟันสำรองมากถึงสามซี่ โดยเฉลี่ยแล้วฟันจะถูกเปลี่ยนหลังจาก 62 วัน ดังที่แสดงโดยวงแหวนการเจริญเติบโตรายวันที่เรียกว่าเส้น von Ebnerซึ่งมองเห็นได้ในภาคตัดขวางของฟัน กระบวนการนี้ช้ากว่าในDiplodocusซึ่งฟันมีอายุเพียงประมาณ 35 วัน แต่เร็วเท่าหรือเร็วกว่าในไดโนเสาร์กลุ่ม Ornithischian [ 93 ]ในการศึกษาปี 2017 Kayleigh Wiersma และ Martin Sander ได้อธิบายถึงเนื้อเยื่ออ่อนที่ปกคลุมบางส่วนของขากรรไกรล่างและฟันของ ตัวอย่าง Camarasaurusที่มีชื่อเล่นว่า "ET" รอยประทับนี้ดูเหมือนจะเป็นเหงือกของสัตว์ ซึ่งบ่งชี้ว่าส่วนยอดของฟันถูกห่อหุ้มบางส่วนด้วยเหงือก เหงือกดังกล่าวอาจอธิบายได้ว่าทำไมแถวฟันของซอโรพอดจึงมักพบในสภาพสมบูรณ์แม้ว่าจะแยกออกจากขากรรไกรแล้วก็ตาม ผู้เขียนเหล่านี้ยังแนะนำว่าเหงือกอาจถูกปกคลุมด้วยจงอยปากที่เป็นเขา ซึ่งอาจช่วยในการตัดพืชพรรณในขณะที่ปกป้องฟัน การมีจงอยปากเช่นนี้สอดคล้องกับการมีรูเล็กๆ (ช่องเปิด) และร่องบนพื้นผิวด้านนอกของขากรรไกรซึ่งจะมีหลอดเลือดอยู่ภายในในขณะที่ยังมีชีวิตอยู่[ 40 ]หรืออีกทางหนึ่ง หลอดเลือดเหล่านี้อาจรองรับ "ริมฝีปาก" เช่นเดียวกับที่พบในกิ้งก่าในปัจจุบัน[ 91 ] : 157
พบซากดึกดำบรรพ์ของ Camarasaurusวัยเยาว์ที่ถูกสัตว์อื่นแทะกินอย่างหนัก จากไวโอมิง ซึ่งมีหินควอตซ์ขัดเงา 14 ก้อน ขนาดเส้นผ่านศูนย์กลางระหว่าง 1 ถึง 13 เซนติเมตร (0.39 ถึง 5.12 นิ้ว) และระบุว่าเป็น แกสโทรลิธ (หินในกระเพาะอาหาร) เดิมทีเชื่อกันว่าซอโรพอดกลืนหินเหล่านี้เข้าไปเพื่อช่วยบดอาหารในกระเพาะ แต่ความหายากของโครงกระดูกที่ยังคงรักษาแกสโทรลิธไว้ได้ และจำนวนที่น้อย บ่งชี้ว่าพวกมันอาจถูกกลืนเข้าไปโดยไม่ได้ตั้งใจ หรือเพื่อรับแร่ธาตุแทน[ 94 ] [ 50 ] : 13
ท่าทางและการทำงานของคอ
คอยาวของซอโรพอดอาจวิวัฒนาการมาเพื่อกินพืชที่อยู่สูงจากพื้นดินหรือเข้าถึงได้ยาก หรือเพื่อเพิ่มปริมาณอาหารที่พวกมันสามารถเข้าถึงได้โดยไม่ต้องขยับร่างกาย จึงเป็นการประหยัดพลังงาน[ 50 ] : 25 ท่าทางของคอที่น่าจะเป็นไปได้นั้นเป็นหัวข้อถกเถียงกัน[ 50 ] : 12 การสร้างโครงกระดูกของCamarasaurus ขึ้นใหม่ในปี 1921 โดย Osborn และ Mook แสดงให้เห็นคอที่ค่อนข้างตรงและอยู่ในแนวนอน[ 95 ] : 219 ในขณะที่ผู้เขียนบางคนในภายหลังสันนิษฐานว่าคอเกือบตั้งตรงเหมือนหงส์[ 95 ] [ 50 ] : 12 โครงกระดูกที่สมบูรณ์บางส่วน เช่น ตัวอย่าง C. lentus วัยเยาว์ CM 11338 ก็แสดงให้เห็นคอที่ตั้งตรงเช่นกัน แต่สิ่งเหล่านี้แสดงถึงท่าทางการตายแบบ opisthotonicซึ่งไม่จำเป็นต้องสะท้อนถึงท่าทางของคอเดิม[ 95 ]ในปี 1998 จอห์น มาร์ตินและเพื่อนร่วมงานได้โต้แย้งว่าคอของซอโรพอดนั้นอยู่ในแนวราบโดยประมาณ เหมือนกับคาน พวกเขาระบุว่าคอของคามาราซอรัสจะต้องแข็งแรงและไม่ยืดหยุ่น และซี่โครงคอที่ยาวจะช่วยค้ำยันไว้ตามด้านล่าง[ 96 ]ในปี 1999 และ 2005 เคนต์ สตีเวนส์และไมเคิล พาร์ริชได้วิเคราะห์ว่ากระดูกสันหลังส่วนคอเชื่อมต่อกันอย่างไรในท่าปกติ และสรุปว่าคอของคามาราซอรัสและซอโรพอดอื่นๆ มักจะตั้งตรงโดยมีความลาดเอียงลงเล็กน้อย[ 97 ] [ 95 ] : 228
แนวคิดเรื่องคอที่อยู่ในแนวนอนนั้นถูกตั้งคำถามโดยการศึกษาวิจัยหลายชิ้นในภายหลัง[ 50 ] : 12 ในปี 2548 David Berman และ Bruce Rothschild ใช้ ข้อมูล จากการตรวจเอกซเรย์คอมพิวเตอร์ (CT) เพื่อเสนอว่ามีกระดูกสันหลังส่วนคอของซอโรพอดอยู่ 2 ประเภท คือ ประเภทที่แข็งแรงและประเภทที่เรียวบางCamarasaurusมีกระดูกสันหลังส่วนคอแบบแข็งแรง ซึ่งบ่งชี้ว่าคอของมันอยู่ในแนวตั้งหรือเกือบแนวตั้ง ในขณะที่แบบเรียวบางบ่งชี้ว่าคออยู่ในแนวนอน[ 98 ]ในปี 2552 Mike Taylor และเพื่อนร่วมงานแสดงให้เห็นว่าในสัตว์สมัยใหม่ คอมักจะยืดออกและโค้งขึ้นด้านบน ซึ่งบ่งชี้ว่าซอโรพอดก็เป็นเช่นนั้นเช่นกัน ในการศึกษาวิจัยในปี 2550 Paul Serenoและเพื่อนร่วมงานเสนอว่าหัวของCamarasaurusมักจะเอียงลงประมาณ 15° โดยพิจารณาจากทิศทางของท่อครึ่งวงกลมในหูชั้นในซึ่งเป็นที่อยู่ของ อวัยวะ รับความรู้สึกสมดุลเทย์เลอร์และเพื่อนร่วมงานโต้แย้งว่าในท่าทางนี้ ปุ่มกระดูกท้ายทอยจะหันลงด้านล่าง ทำให้ส่วนหน้าของคอชันขึ้น และอาจใกล้เคียงกับแนวตั้ง[ 99 ] [ 100 ]


กระดูกสะบักด้านขวาของตัวอย่าง CM 11338 เอียงประมาณ 45° เมื่อเทียบกับแนวราบ กิลมอร์ ในงานเขียนเชิงวิชาการปี 1925 ของเขา ได้โต้แย้งว่าตัวอย่างนี้สะท้อนถึงทิศทางดั้งเดิมของกระดูก และด้วยเหตุนี้ การสร้างโครงกระดูกขึ้นใหม่ของเขาจึงทำให้สะโพกสูงกว่าไหล่เล็กน้อย การค้นพบนี้ขัดแย้งกับการสร้างโครงกระดูกขึ้นใหม่ในปี 1921 ของออสบอร์นและมุก ซึ่งแสดงให้เห็นกระดูกสะบักที่ชันกว่ามาก ส่งผลให้สัตว์มีไหล่สูงกว่าสะโพก และฐานคออยู่สูงกว่าพื้น การตีความของกิลมอร์เกี่ยวกับกระดูกสะบักที่ทำมุมต่ำนั้นต่อมาได้รับการยอมรับอย่างกว้างขวางสำหรับซอโรพอดโดยทั่วไป ในการศึกษาปี 2007 ดาเนียลา ชวาร์ซและเพื่อนร่วมงานได้เปรียบเทียบกายวิภาคของกระดูกหัวไหล่กับสัตว์ในปัจจุบัน และสรุปว่าการตีความดั้งเดิมของออสบอร์นและมุกเกี่ยวกับกระดูกสะบักที่เอียงชัน (60–65°) และไหล่ที่สูงขึ้นนั้นถูกต้อง[ 63 ]เอ็นจะวิ่งไปตามส่วนบนของคอ ซึ่งจะตึงเมื่อคอเอียงลงหรือไปด้านข้าง ช่วยในการยึดคอไว้ ในการศึกษาในปี 2004 Takanobu Tsuihiji ได้สร้างเอ็นของCamarasaurus ขึ้นใหม่ โดยอิงจากเอ็นของGreater Rheaซึ่งกระดูกสันหลังส่วนประสาทแยกออกเป็นสองแฉกเช่นเดียวกันเอ็นท้ายทอยจะวิ่งไปตามด้านบนโดยมีกิ่งก้านเชื่อมต่อกับด้านใดด้านหนึ่งของกระดูกสันหลังส่วนประสาทที่แยกออกเป็นสองแฉก ในขณะที่เอ็นที่สอง Ligamentum elasticum interspinaleจะวิ่งอยู่ระหว่างสองแฉกของกระดูกสันหลังส่วนประสาทที่แยกออกเป็นสองแฉก[ 101 ] [ 91 ] : 184–185
ในอดีต มีการสร้างภาพจำลองของซอโรพอดโดยให้หางลากไปกับพื้น[ 50 ] : 11 ใน การสร้างภาพจำลอง Camarasaurusของ Osborn และ Mook ในปี 1921 หางเริ่มลาดลงทันทีหลังสะโพก ในขณะที่อภิปรายเกี่ยวกับ การสร้างภาพจำลอง Camarasaurus ของเขาเอง ในปี 1925 Gilmore โต้แย้งว่ากระดูกสันหลังส่วนหางสองสามข้อแรกจะต้องอยู่ในแนวนอน และหางจะลาดลงหลังจากส่วนนี้เท่านั้น[ 24 ] : 383–384 ปัจจุบันเชื่อกันว่าซอโรพอดได้ยกหางทั้งหมดขึ้นจากพื้น ดังที่แสดงโดยหลักฐานทางกายวิภาคและร่องรอย[ 50 ] : 11
ความแตกต่างทางเพศ
ในการศึกษาเมื่อปี 1991 บรูซ รอธไชลด์และเดวิด เบอร์แมน สังเกตว่าใน ตัวอย่าง คามาราซอรัส 25% กระดูกสันหลังส่วนหางส่วนหน้าบางส่วนเชื่อมติดกัน ในอะพาโทซอรัสและดิโพลโดคัสการเชื่อมติดกันเช่นนี้เกิดขึ้นในตัวอย่างถึง 50% การเชื่อมติดกันนี้เกิดจากเอ็นที่กลายเป็นกระดูกมากกว่าการเชื่อมติดกันโดยตรงของกระดูกสันหลัง และถูกระบุว่าเป็น ภาวะกระดูกงอกเกินแบบไม่ทราบสาเหตุ ( Diffuse Idiopathic Skeletal Hyperostosisหรือ DISH) [ a ]ดังนั้น การเชื่อมติดกันนี้จึงไม่ใช่พยาธิสภาพแต่อาจเป็นการปรับตัวเพื่อทำให้หางแข็งแรงขึ้น รอธไชลด์และเบอร์แมนโต้แย้งว่าการเชื่อมติดกันนี้เป็น ลักษณะ ทางเพศที่แตกต่างกันซึ่งเกิดขึ้นเฉพาะในตัวผู้หรือเฉพาะในตัวเมียเท่านั้น ในตัวผู้ การเชื่อมติดกันอาจช่วยรองรับการเคลื่อนไหวแบบสะบัดด้วยปลายหางระหว่างการต่อสู้กับตัวผู้ตัวอื่น ในตัวเมีย ความแข็งแรงอาจช่วยในการโค้งหางเพื่อการผสมพันธุ์[ 103 ]ในการศึกษาเมื่อปี 2551 Takehito Ikejiri เสนอว่า ตัวอย่าง ของ Camarasaurusสามารถจำแนกได้เป็นแบบแข็งแรง (มีโครงสร้างที่แข็งแรง) หรือแบบเรียว (ผอมเพรียว) รูปแบบที่แข็งแรงและเรียวเหล่านี้ยังมีความแตกต่างกันเล็กน้อยในขนาดและปรากฏให้เห็นในสามสายพันธุ์ที่พบได้บ่อยที่สุด Ikejiri โต้แย้งว่ารูปแบบทั้งสองสะท้อนถึงความแตกต่างระหว่างเพศ แม้ว่าจะไม่ชัดเจนว่ารูปแบบใดแสดงถึงตัวผู้และรูปแบบใดแสดงถึงตัวเมีย[ 104 ]ในการศึกษาอีกครั้งในปี 2551 Nicole Klein และ Martin Sander พบว่าบุคคลที่มีอายุใกล้เคียงกันมักจะอยู่ในสองกลุ่มขนาดที่อาจแสดงถึงสายพันธุ์ที่แตกต่างกันหรือความแตกต่างทางเพศ[ 105 ]
ประวัติชีวิต
ในปี พ.ศ. 2426 มาร์ชได้รายงานการค้นพบโครงกระดูกที่แตกหักของซอโรพอดขนาดเล็กมากที่โคโมบลัฟฟ์ ซึ่งเขาประมาณความยาวลำตัวไว้ที่ประมาณ 2.1 เมตร (7 ฟุต) จากขนาดที่เล็กและการสร้างกระดูกที่ไม่สมบูรณ์ มาร์ชจึงสันนิษฐานว่ามันน่าจะเป็นของตัวอ่อน [ 67 ] [ b ] ในปี พ.ศ. 2439 มาร์ชได้จัดตัวอย่างนี้ให้อยู่ในสปีชีส์ใหม่Pleurocoelus montanusโดยไม่มีคำอธิบายเพิ่มเติม แต่คาดว่าน่าจะเป็นเพราะช่องว่างขนาดใหญ่ (โพรง) ในกระดูกสันหลัง[ 106 ] [ 80 ]ในปี พ.ศ. 2537 คาร์เพนเตอร์และแมคอินทอชได้จัดตัวอย่างนี้ให้อยู่ในCamarasaurus grandisและตีความว่าช่องว่างขนาดใหญ่เป็นลักษณะของตัวอ่อน[ 80 ]แม้ว่าจะเป็นตัวอ่อนขนาดเล็ก แต่ก็ไม่มีหลักฐานว่าตัวอย่างนี้เป็นตัวอ่อนจริงตามที่มาร์ชเสนอ[ 107 ]ในปี 1994 Brooks Britt และ Bruce Naylor ได้บรรยายถึงกระดูกขากรรไกรบนขนาดเล็กของCamarasaurusที่ค้นพบในDry Mesa Quarryฟันของกระดูกชิ้นนี้ยังไม่ขึ้นแสดงว่าตัวอย่างนี้ยังไม่ฟักออกจากไข่และจึงเป็นตัวอ่อน กระดูกมีความยาว 37 มม. (1.5 นิ้ว) ซึ่งบ่งชี้ว่าความยาวกะโหลกประมาณ 70 มม. (2.8 นิ้ว) ความยาวลำตัวมากกว่า 1 ม. (39 นิ้ว) เล็กน้อย และน้ำหนักตัว 7.5 กก. (17 ปอนด์) จากการประมาณการเหล่านี้ เส้นผ่านศูนย์กลางของไข่ทรงกลมที่คาดการณ์ไว้จะมีขนาดประมาณ 24 ซม. (9.4 นิ้ว) ซึ่งเล็กกว่าไข่นกที่ใหญ่ที่สุดที่รู้จัก ขนาดเล็กของตัวอ่อนเป็นหลักฐานที่แสดงว่าซอโรพอดเป็นสัตว์วางไข่ซึ่งเป็นการตั้งคำถามถึงสมมติฐานที่เสนอโดยRobert T. Bakkerในปี 1980 ที่ว่าซอโรพอดเป็นสัตว์ออกลูกเป็นตัวและให้กำเนิดลูกที่มีขนาดค่อนข้างใหญ่[ 107 ] [ 108 ]ตัวอ่อนและไข่ซอโรพอดตัวแรกที่ระบุได้อย่างชัดเจนนั้นมาจากแหล่งโบราณคดีAuca Mahuevo ในอาร์เจนตินา ในปี 1998 [ 109 ]
เช่นเดียวกับไดโนเสาร์ชนิดอื่นๆ ลูกไดโนเสาร์จะมีหัวที่ใหญ่กว่าเมื่อเทียบกับขนาดตัว คอและหางสั้นกว่า และแขนขาสั้นกว่าตัวเต็มวัย[ 110 ] : 264 เมื่อไดโนเสาร์โตเต็มวัย ส่วนโค้งประสาทของกระดูกสันหลังจะเชื่อมติดกับตัวกระดูกสันหลัง ในCamarasaurusกระดูกสันหลังส่วนกลางและส่วนท้ายของหางจะเชื่อมติดกันก่อนกระดูกสันหลังส่วนกระเบนเหน็บ[ 25 ]การแตกแขนง (แยกเป็นสองแฉก) ของกระดูกสันหลังจะเด่นชัดมากขึ้นเมื่อไดโนเสาร์มีอายุมากขึ้น การเปลี่ยนแปลงอื่นๆ ที่เกี่ยวข้องกับอายุที่พบในตัวเต็มวัย ได้แก่ พื้นผิวข้อต่อที่ขรุขระในแขนขา การปิดของ รู โคราคอยด์ (การเปิด) การเกิดกระดูกแข็งของเอ็นเทซิสของกระดูกสันหลัง และการเชื่อมติดกันของกระดูกสันหลังส่วนกระเบนเหน็บแต่ละชิ้นเข้าเป็นโครงสร้างเดียว[ 111 ] [ 59 ] : 81 [ 110 ] : 264 การเปลี่ยนแปลงระหว่างการเจริญเติบโตนั้นเด่นชัดเป็นพิเศษในกระดูกอก ซึ่งมีรูปทรงกลมใน CM 11338 วัยเยาว์ แต่กลับยาวและแคบลงในตัวเต็มวัย ในทางตรงกันข้าม สัดส่วนของกระดูกแขนขาไม่ได้เปลี่ยนแปลงระหว่างการเจริญเติบโต[ 112 ]แม้ว่าแขนขาโดยรวมจะแข็งแรงกว่าในตัวเต็มวัยก็ตาม[ 110 ] : 264
วงปีการเจริญเติบโตและลักษณะอื่นๆ ที่มองเห็นได้ในส่วนตัดบางๆของกระดูกช่วยให้สามารถสร้างประวัติชีวิตขึ้นใหม่ได้ เช่นเดียวกับซอโรพอดอื่นๆCamarasaurusเติบโตเร็วเท่ากับนกและสัตว์เลี้ยงลูกด้วยนมในปัจจุบัน และถึงวัยเจริญพันธุ์ก่อนที่จะมีขนาดตัวสูงสุด[ 105 ] [ 113 ] : 865 ในปี 2013 Eva Maria Griebeler และเพื่อนร่วมงานได้ตรวจสอบส่วนตัดบางๆ ของกระดูกแขนขาของCamarasaurusตัวใหญ่ (CM 36664) ซึ่งมีน้ำหนักโดยประมาณ 14.3 ตัน คาดว่าตัวนี้มีอัตราการเติบโตสูงสุด 1.5 ตันต่อปี ถึงวัยเจริญพันธุ์เมื่ออายุประมาณ 21 ปี และเสียชีวิตเมื่ออายุประมาณ 26 ปี[ 114 ] : 3 การศึกษาในปี 2017 โดย Woodruff และ Foster ประมาณการว่าตัวอย่าง GPDM 220 น่าจะมีอายุประมาณ 30 ปี และสูงสุด 35 ปี เมื่อเสียชีวิต[ 115 ]ตัวอย่าง "ET" มีอายุมากกว่านั้นอีก ในการศึกษาเมื่อปี 2557 Katja Waskow และ Sander ประเมินว่าตัวอย่างนี้ถึงวัยเจริญพันธุ์เมื่ออายุ 18 หรือ 19 ปี และมีขนาดเต็มที่เมื่ออายุ 40 ปี[ 113 ] : 866 ณ ปี 2567 GPDM 220 และ "ET" เป็นหนึ่งในไดโนเสาร์ที่มีอายุมากที่สุดที่ได้รับการระบุ แม้ว่าจะเป็นตัวแทนของไดโนเสาร์ที่มีขนาดค่อนข้างเล็กก็ตาม[ 116 ] : 684 ไดโนเสาร์อาจเติบโตตลอดช่วงชีวิตส่วนใหญ่ของพวกมัน ในปี 2564 Bruce Rothschild และ Florian Witzmann พบว่าจาก ตัวอย่าง Camarasaurus ที่วิเคราะห์ 13 ตัวอย่าง มี 2 ตัวอย่างที่น่าจะถึงขนาดเต็มที่แล้ว ดังที่ระบุโดยการปิดของช่องหลอดเลือดบนพื้นผิวข้อต่อของกระดูกยาวที่ให้สารอาหารสำหรับการเจริญเติบโตของกระดูก[ 117 ] : 263, 267
การเผาผลาญ
โดยทั่วไปแล้วเชื่อกันว่าไดโนเสาร์เป็น สัตว์เลือดเย็นที่ต้องอาศัยอุณหภูมิจากสิ่งแวดล้อม ตั้งแต่ปี 1960 เป็นต้นมา มีหลักฐานที่แสดงให้เห็นว่าไดโนเสาร์เป็นสัตว์เลือดอุ่น (สามารถรักษาอุณหภูมิร่างกายให้คงที่ได้) หรือแม้แต่สัตว์เลือดอุ่น (มีการเผาผลาญที่เพิ่มขึ้นซึ่งสามารถรักษาอุณหภูมิให้คงที่ได้) [ 50 ] : 15 [ 118 ]ซอโรพอดวัยเยาว์น่าจะเป็นสัตว์เลือดอุ่น ทำให้พวกมันเติบโตได้อย่างรวดเร็ว ในขณะที่ตัวที่โตเต็มวัยอาจมีอัตราการเผาผลาญลดลง เนื่องจากอุณหภูมิร่างกายสามารถรักษาไว้ได้ด้วยมวลร่างกายเพียงอย่างเดียว[ 50 ] : 17 อุณหภูมิร่างกายสามารถประมาณได้โดยตรงจาก องค์ประกอบ ไอโซโทปของกระดูกและฟัน[ 118 ] [ 119 ]ในการศึกษาปี 2002 วิลเลียม โชเวอร์สและเพื่อนร่วมงานใช้ เทอ ร์โมเมตรีไอโซโทปออกซิเจนเพื่อวิเคราะห์อะพาไทต์ ของกระดูก พวกเขาพบว่า ใน ตัวอย่าง Camarasaurus ที่พวกเขาศึกษา อุณหภูมิในลำตัวมีความแปรปรวน แต่ลดลงในขาและสะโพก และเพิ่มขึ้นในคอและหาง เมื่อเปรียบเทียบกับตัวอย่างของเทโรพอดGiganotosaurusความแตกต่างระหว่างส่วนต่างๆ ของร่างกายนี้อาจเกิดจากการแลกเปลี่ยนความร้อนแบบสวนทางกัน เมื่อความร้อนส่วนเกินถูกสูบจากลำตัวไปยังส่วนต่างๆ ของร่างกายรอบนอกเพื่อเป็นกลไกการระบายความร้อน[ 118 ]ในปี 2011 โรเบิร์ต อีเกิลและเพื่อนร่วมงานได้วิเคราะห์เคลือบฟันของซอโรพอดสองชนิด ได้แก่CamarasaurusและGiraffatitanโดยใช้เทอร์โมเมตรีไอโซโทปแบบรวมกลุ่มการวิเคราะห์นี้บ่งชี้ว่าอุณหภูมิร่างกายอยู่ที่ 36–38 °C (97–100 °F) ซึ่งเทียบได้กับอุณหภูมิของสัตว์เลี้ยงลูกด้วยนมในปัจจุบัน[ 119 ]
พยาธิวิทยา
มีการบันทึกกรณีความผิดปกติ (การบาดเจ็บหรือโรค) หลายครั้งใน ตัวอย่าง Camarasaurusในปี 1996 McIntosh และเพื่อนร่วมงานได้อธิบายความผิดปกติในกระดูกสันหลัง 13 ชิ้นของหางที่สมบูรณ์ของ ตัวอย่าง C. grandis GMNH-PV 101 ในกระดูกสันหลังสองชิ้นนี้ ส่วนโค้งประสาทพัฒนาไม่สมบูรณ์ ซึ่งเป็นความผิดปกติในการพัฒนาที่เรียกว่าspina bifidaซึ่งเป็นตัวอย่างแรกที่มีการรายงานเกี่ยวกับภาวะนี้ในไดโนเสาร์ ในกระดูกสันหลังหางชิ้นที่ 40 ส่วนโค้งประสาทก่อตัวขึ้นเพียงครึ่งเดียว ทำให้ไขสันหลังไม่ได้รับการปกป้องบางส่วน กระดูกสันหลังอย่างน้อยห้าชิ้นแสดงให้เห็นการเจริญเติบโตของกระดูกรอบข้อต่อของกระดูกสันหลังส่วนกลาง ซึ่งบ่งชี้ถึง โรค ข้อเสื่อม (โรคข้อเสื่อม) กระดูกสันหลังหางห้าชิ้นติดต่อกัน (49 ถึง 53) เชื่อมติดกันเป็นโครงสร้างเดียวอย่างผิดปกติ[ 60 ] : 17–18 [ 120 ]
ตัวอย่างจาก Bone Cabin Quarry แสดงให้เห็นการสึกกร่อนในข้อ ต่อกระดูกสันหลัง ( zygapophyses ) ในกระดูกสันหลังส่วนหาง 4 ใน 20 ชิ้น ในปี 2545 Rothschild และเพื่อนร่วมงานได้ระบุพยาธิสภาพเหล่านี้ว่าเป็นหลักฐานฟอสซิลที่เก่าแก่ที่สุดของโรคข้ออักเสบ[ 121 ] [ c ]ในปี 2544 Lorrie McWhinney และเพื่อนร่วมงานได้อธิบายถึง ภาวะเยื่อหุ้มกระดูกอักเสบ ( periostitis ) ซึ่งเป็นการบาดเจ็บของเยื่อหุ้มกระดูก (ชั้นนอกของกระดูก) ในกระดูกต้นแขนที่ระบุว่าเป็นC. grandisการบาดเจ็บนี้เกี่ยวข้องกับส่วนของกระดูกที่แตกหักหรือฉีกขาดอาจเนื่องมาจากความเครียด หรือ การออกแรง กล้ามเนื้อ มากเกินไปซ้ำๆกระบวนการรักษาที่ตามมาทำให้เกิดก้อนคล้ายเนื้องอกยื่นออกมาจากผิวกระดูก การบาดเจ็บนี้น่าจะเป็นแบบเรื้อรังและอาจทำให้การเคลื่อนไหวของแขนขาด้านหน้าบกพร่องและทำให้เดินกะเผลก[ 123 ]ในปี 2559 Emanuel Tschopp และเพื่อนร่วมงานได้อธิบายพยาธิสภาพที่แตกต่างกัน 5 ประเภทในกระดูกของเท้าหน้าและเท้าหลังของ SMA 0002 ("ET") การเกิดพยาธิสภาพที่แตกต่างกันในบุคคลเดียวกันนั้นหายากและอาจเกิดจากอายุที่มากของบุคคลนั้น พยาธิสภาพดังกล่าวรวมถึงหลุมลึกที่ตีความว่าเป็นโรคกระดูกอ่อนเสื่อมเช่นเดียวกับการเจริญเติบโตของกระดูกหลายประเภท ซึ่งหนึ่งในนั้นตีความว่าเป็นโรคข้อเสื่อม กระดูกยื่นออกมาเหนือพื้นผิวข้อต่อด้านหน้าของกระดูกนิ้วเท้าของเท้าหลังถูกตีความว่าเป็นเอ็นทีโซไฟต์ที่เกิดจากการแทรกของเอ็น สิ่งเหล่านี้อาจเกิดขึ้นเนื่องจากการใช้กรงเล็บมากเกินไปในระหว่างมีชีวิตอยู่ อาจเนื่องมาจากการขุดดินเพื่อขีดข่วน[ 41 ]
นิเวศวิทยาบรรพกาล

การกระจายตัวและความอุดมสมบูรณ์
Camarasaurusเป็นที่รู้จักจากหินตะกอนของ Morrison Formation ที่มีอายุย้อนไปถึงยุคKimmeridgianและTithonian ( 155 ถึง 143ล้านปีก่อน ) [ 56 ] : 266 Morrison Formation ครอบคลุมพื้นที่ประมาณ 1.2 ล้านตารางกิโลเมตรในอเมริกาเหนือตะวันตก และ พบ Camarasaurusกระจายอยู่ทั่วบริเวณนี้ ตั้งแต่แหล่งที่ตั้งมากกว่า 100 แห่งทางเหนือสุดที่มอนแทนา ไปจนถึงทางใต้สุดที่นิวเม็กซิโก[ 124 ] [ 125 ]กระดูกสันหลังส่วนหางชิ้นเดียวจากSummerville Formationของนิวเม็กซิโกได้รับการจัดให้อยู่ในสกุลนี้โดย Adrian Hunt และSpencer G. Lucasในปี 1993 แต่การค้นพบนี้ไม่ได้รับการยอมรับในการตรวจสอบครั้งต่อมา[ 126 ] [ 48 ] [ 56 ] : 266 ซากดึกดำบรรพ์จากซิมบับเวและ เยอรมนีได้รับการจัดให้อยู่ในCamarasaurus [ 127 ] [ 128 ]ซึ่งถูกตั้งคำถามโดยการศึกษาในภายหลัง[ 129 ] [ 130 ] : 26–27
Camarasaurusเป็นที่รู้จักจากตัวอย่างมากกว่า 530 ชิ้น รวมถึงกระดูกที่แยกออกมาและโครงกระดูกบางส่วนประมาณ 50 ชิ้น[ 131 ]เป็นไดโนเสาร์ที่พบได้บ่อยที่สุดใน Morrison Formation และเนื่องจากมีจำนวนมาก จึงเป็นหนึ่งในซอโรพอดที่เข้าใจได้ดีที่สุด[ 51 ] : 240, 272 ในการสำรวจแหล่งฟอสซิลมากกว่า 200 แห่งในปี 2003 John Foster รายงานตัวอย่างของสกุลนี้ 179 ตัวอย่าง เทียบได้กับApatosaurus (112) และDiplodocus (98) แต่มากกว่าBrachiosaurus (12), Haplocanthosaurus (12) และBarosaurus (13) มาก [ 132 ]ณ ปี 2022 ตัวอย่างซอโรพอดจาก Morrison ที่สามารถระบุสกุลได้ 27% เป็นตัวอย่างของ Camarasaurus [ 110 ]ตัวอย่างCamarasaurus ที่ระบุได้ส่วนใหญ่ เป็นของหนึ่งในสองชนิด คือC. grandisและC. lentus ; C. supremusและโดยเฉพาะอย่างยิ่งC. lewisiนั้นหายากกว่ามาก[ 77 ]แม้ว่าคอที่สมบูรณ์จะพบได้ยากในซอโรพอด แต่ตัวอย่างCamarasaurus จำนวน 5 ตัวอย่าง ยังคงรักษากระดูกสันหลังส่วนคอทั้งหมดหรือเกือบทั้งหมดไว้ได้[ 133 ] ตัวอย่างซอโรพอดวัยเยาว์โดยทั่วไปนั้นหายาก เนื่องจากขนาดที่เล็กกว่าทำให้ศักยภาพในการเก็บรักษาลดลง ณ ปี 2005 ตัวอย่างซอโรพอดที่พบใน Morrison Formation ที่มีขนาดเล็กกว่า 50% ของขนาดตัวเต็มวัย 44 % มาจากCamarasaurus [ 134 ]
สภาพแวดล้อมโบราณและการอพยพ
ชั้นหินมอร์ริสันถูกตีความว่าเป็น สภาพแวดล้อม กึ่งแห้งแล้งที่ มีฤดู ฝนและฤดูแล้งที่ชัดเจน[ 135 ] [ 136 ]ในปี 2011 เฮนรี ฟริกเกและเพื่อนร่วมงานได้วิเคราะห์ความอุดมสมบูรณ์สัมพัทธ์ของไอโซโทปออกซิเจน (ค่า δ 18 O) ของทั้ง ฟัน ของคามาราซอรัสและหินคาร์บอเนตทั่วแอ่งมอร์ริสัน ค่า δ 18 O แตกต่างกันไปตามภูมิศาสตร์ขึ้นอยู่กับปัจจัยต่างๆ เช่น ความแห้งแล้งและระดับความสูง เนื่องจากค่าที่ได้จากฟันแตกต่างจากค่าที่ได้จากหินที่พบฟันนั้น ฟริกเกและเพื่อนร่วมงานจึงสรุปว่าคามาราซอรัสต้องอพยพระหว่างแอ่งมอร์ริสันและพื้นที่สูงทางตะวันตกเพื่อหลีกเลี่ยงฤดูแล้งของแอ่ง การอพยพนี้อาจเป็นไปตามฤดูกาลและครอบคลุมระยะทาง 300 กม. (190 ไมล์) [ 137 ]
ณ ปี 2024 มีการระบุชนิดของซอโรพอดทั้งหมด 25 ชนิดจากชั้นหินมอร์ริสัน[ 125 ]สกุลที่พบได้บ่อยที่สุดคือไดโพลโดซิด ได้แก่ อะพาโทซอรัส ดิโพลโดคัส ซูเปอร์ซอรัสบาโรซอรัสบรอนโทซอ รัส และกาเลียมอปัส ; แมคโครนา เรียน ได้แก่ บราคิโอ ซอรัส และฮาโพลแคนโท ซอรัส ไดเครโอ ซอริเด เช่น สมิทาโนซอ รัส ได ส โล โคซอรัส และซูวาสเซียเป็นส่วนประกอบที่หายากกว่าของสัตว์[ 125 ]ไดโนเสาร์อื่นๆ ที่รู้จักจากชั้นหินมอร์ริสัน ได้แก่เทโรพอด นักล่า ได้แก่ โคพาริ ออน สโตกซอ รัส ออร์ นิโทเลสเตส เซราโทซอรัส อั ลโล ซอรัส และทอร์โวซอรัสรวมถึงออร์ นิธิสเชีย น กินพืช ได้แก่ แคมป์โทซอรัส ไดรโอซอ รัส การ์ กอยเลโอซอรัสและสเตโกซอรัส[ 138 ]อัลโลซอรัสคิดเป็น 70 ถึง 75 เปอร์เซ็นต์ของตัวอย่างเทอโรพอด และอยู่ในระดับสูงสุดของห่วงโซ่อาหารของมอร์ริสัน[ 139 ] สัตว์มีกระดูกสันหลังอื่นๆ ที่อาศัยอยู่ในสภาพแวดล้อมโบราณนี้ ได้แก่ ปลา ครีบแข็งกบ ซาลาแมนเดอร์เต่าเช่นดอร์เซโตเชลิส สเฟโนดอนต์ กิ้งก่าจระเข้บกและจระเข้ น้ำ เช่น ฮอปโล ซูคัส และสัตว์ ปีกดึกดำบรรพ์หลายชนิดเช่น ฮาร์ แพคโตกนา ทัส และเมซาแดคทิลัสเปลือกหอยสองฝาและหอยทาก น้ำก็พบได้ทั่วไป พืช พรรณในยุคนั้นได้รับการเปิดเผยจากฟอสซิลของสาหร่ายสีเขียวเชื้อรามอสหญ้าหางม้าไซแคด แปะก๊วยและสนหลายวงศ์พืชพรรณมีความหลากหลาย ตั้งแต่ป่าริมแม่น้ำในบริเวณที่ปราศจากต้นไม้ ( ป่าริมแม่น้ำ ) ที่มีเฟิร์นต้นไม้และเฟิร์น ชนิดอื่นๆ ไปจนถึงทุ่งหญ้าสะวัน นาที่มีเฟิร์นขึ้นอยู่ประปราย และมีต้นไม้บ้าง เช่น ต้นอะราอุคาเรีย-คล้ายสนBrachyphyllum [ 140 ]
หมายเหตุ
- ^การทบทวนในปี 2012 เตือนว่า DISH ไม่เป็นที่รู้จักในสัตว์เลื้อยคลานและนกสมัยใหม่ และคำจำกัดความทางการแพทย์ของภาวะนี้ต้องการการเชื่อมต่อของกระดูกสันหลังอย่างน้อยสี่ชิ้นที่ต่อเนื่องกัน ในขณะที่ตัวอย่างของซอโรพอดโดยทั่วไปเกี่ยวข้องเพียงสองชิ้น [ 102 ] : 694–695 [ 103 ]
- ^มาร์ชใช้คำว่าทารก ในครรภ์ แทนคำว่าตัวอ่อน
- ^มีการรายงานตัวอย่างโรคข้ออักเสบจากการอักเสบในยุคไทรแอสสิกที่เก่ากว่านั้นอีก [ 122 ]
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ คามาราซอรัส
คามาราซอรัส ( Camarasaurus )เป็นสกุลของไดโนเสาร์ซอโรพอดที่อาศัยอยู่ในช่วงปลายยุคจูราสสิกระหว่างประมาณ 155 ถึง 145 ล้านปีก่อน ใน ทวีปอเมริกาเหนือ...
การค้นพบและการตั้งชื่อ
คามาราซอรัส ถูกค้นพบในช่วง สงครามกระดูก ซึ่งเป็นความขัดแย้งระหว่างนักบรรพชีวินวิทยาชาวอเมริกันสองคน คือ โอ ธเนียล ชาร์ลส์ มาร์ช และ เอ็ดเวิร์ด ดริงเกอร์ โคป ซึ่งนำไปสู่การค้นพบฟอสซิลจำนวนมากในภาคตะวันตกของสหรัฐอเมริกาในช่วงปลายศตวรรษที่ 19 [ 1 ] กระดูกคา...
โมโรซอรัส และสงครามกระดูก
เมื่อวันที่ 7 มีนาคม พ.ศ. 2420 วิลเลียม ฮาร์โลว์ รีด พนักงานรถไฟได้พบกระดูกฟอสซิลทางใต้ของสถานีโคโม รัฐไวโอมิง บนเนินเขาซึ่งต่อมากลายเป็นหนึ่งในแหล่งไดโนเสาร์ที่สำคัญแห่งแรกๆ นั่นคือโค โมบลัฟฟ์ [ 11 ] รีดและวิลเลียม เอ็ดเวิร์ด คาร์ลิน เพื่อนร่วมงานของเขา...
ไดโนเสาร์รัชครั้งที่สอง
หลังจากสิ้นสุดสงครามกระดูก สถาบันสำคัญหลายแห่งในภาคตะวันออกของสหรัฐอเมริกาได้รับแรงบันดาลใจจากภาพวาดและการค้นพบของ Marsh และ Cope เพื่อรวบรวมคอลเลกชันฟอสซิลไดโนเสาร์ของตนเอง [ 26 ] : 64–65, 105 การแข่งขันเพื่อจัดแสดงโครงกระดูกซอโรพอดตัวแรกนั้นเข้มข้นที่สุด...