อ่าน 26 นาที
สมองน้อย
สมองน้อย ( พหูพจน์ : cerebellaหรือcerebellums ; ภาษาละตินแปลว่า 'สมองเล็ก') เป็นส่วนประกอบหลักของสมองส่วนท้ายของสัตว์มีกระดูกสันหลัง ทุกชนิด...
สมองน้อย
| สมองน้อย | |
|---|---|
 ตำแหน่งของสมองส่วนซีรีเบลลัมของมนุษย์ (ภาพตัดขวางแนวตั้ง) | |
 ภาพเคลื่อนไหวของสมองส่วนซีรีเบลลัม | |
| รายละเอียด | |
| การออกเสียง | / ˌ s ɛ r ə ˈ b ɛ l ə m / |
| ส่วนหนึ่งของ | สมองส่วนท้าย |
| หลอดเลือดแดง | SCA , AICA , PICA |
| เส้นเลือด | เหนือกว่า , ต่ำกว่า |
| ตัวระบุ | |
| ละติน | สมองน้อย |
| เมช | D002531 |
| นิวโรเนมส์ | 643 |
| รหัสNeuroLex | เบิร์นเล็กซ์_1489 |
| TA98 | A14.1.07.001 |
| ทีเอ2 | 5788 |
| เอฟเอ็มเอ | 67944 |
| คำศัพท์ทางกายวิภาคศาสตร์ของระบบประสาท | |
สมองน้อย ( พหูพจน์ : cerebellaหรือcerebellums ; ภาษาละตินแปลว่า 'สมองเล็ก') เป็นส่วนประกอบหลักของสมองส่วนท้ายของสัตว์มีกระดูกสันหลัง ทุกชนิด แม้ว่าโดยปกติจะมีขนาดเล็กกว่าสมองใหญ่แต่ในสัตว์บางชนิด เช่น ปลา ในวงศ์ Mormyridaeอาจมีขนาดใหญ่เท่าหรือใหญ่กว่าสมองใหญ่ก็ได้[ 1 ]ในมนุษย์ สมองน้อยมีบทบาทสำคัญในการควบคุมการเคลื่อนไหวและหน้าที่การรับรู้เช่นความสนใจและภาษาตลอดจนการควบคุมอารมณ์เช่น การควบคุมความกลัวและความสุข[ 2 ] [ 3 ] [ 4 ]แต่หน้าที่ที่เกี่ยวข้องกับการเคลื่อนไหวนั้นได้รับการยืนยันอย่างมั่นคงที่สุด สมองน้อยของมนุษย์ไม่ได้เริ่มต้นการเคลื่อนไหว แต่มีส่วนช่วยในการประสานงานการเคลื่อนไหวความแม่นยำ และจังหวะเวลาที่ถูกต้อง โดยจะรับข้อมูลจากระบบประสาทสัมผัสของไขสันหลังและจากส่วนอื่นๆ ของสมองและบูรณาการข้อมูลเหล่านี้เพื่อปรับแต่งกิจกรรมการเคลื่อนไหว[ 5 ]ความเสียหายของสมองน้อยทำให้เกิดความผิดปกติใน การ เคลื่อนไหวละเอียดการทรงตัวท่าทางและการเรียนรู้การเคลื่อนไหวในมนุษย์[ 5 ]
ในทางกายวิภาคศาสตร์ สมองน้อยของมนุษย์มีลักษณะเป็นโครงสร้างแยกต่างหากที่ติดอยู่กับส่วนล่างของสมอง ซ่อนอยู่ใต้ซีก สมองใหญ่ ผิวเปลือกสมองถูกปกคลุมด้วยร่องขนานที่เว้นระยะห่างกันอย่างละเอียด ซึ่งแตกต่างอย่างเห็นได้ชัดจากรอยหยักที่กว้างและไม่สม่ำเสมอของเปลือกสมองใหญ่ร่องขนานเหล่านี้ปกปิดความจริงที่ว่าเปลือกสมองน้อยเป็นชั้นเนื้อเยื่อ บางๆ ที่ต่อเนื่องกัน และพับแน่นในลักษณะคล้ายหีบเพลง ภายในชั้นบางๆ นี้มี เซลล์ประสาทหลายชนิดที่มีการจัดเรียงอย่างเป็นระเบียบสูง เซลล์ที่สำคัญที่สุดคือเซลล์ Purkinjeและเซลล์เม็ดเล็กการจัดระเบียบประสาทที่ซับซ้อนนี้ทำให้เกิดความสามารถในการประมวลผลสัญญาณจำนวนมาก แต่เกือบทั้งหมดของสัญญาณที่ออกจากเปลือกสมองน้อยจะผ่านนิวเคลียสขนาด เล็ก ที่อยู่ลึก ภายใน เนื้อขาวของสมองน้อย[ 6 ]
นอกจากบทบาทโดยตรงในการควบคุมการเคลื่อนไหวแล้ว สมองน้อยยังมีความจำเป็นต่อการเรียนรู้การเคลื่อนไหว หลายประเภท โดยเฉพาะอย่างยิ่งการเรียนรู้ที่จะปรับตัวให้เข้ากับการเปลี่ยนแปลงในความสัมพันธ์ระหว่างประสาท สัมผัสและการเคลื่อนไหว มีการพัฒนารูปแบบทฤษฎีหลายแบบเพื่ออธิบายการปรับเทียบประสาทสัมผัสและการเคลื่อนไหวในแง่ของความยืดหยุ่นของไซแนปส์ภายในสมองน้อย รูปแบบเหล่านี้มาจากรูปแบบที่คิดค้นโดยDavid MarrและJames Albusโดยอิงจากการสังเกตว่าเซลล์ Purkinje ในสมองน้อยแต่ละเซลล์ได้รับอินพุตสองประเภทที่แตกต่างกันอย่างมาก: ประเภทหนึ่งประกอบด้วยอินพุตที่อ่อนแอหลายพันตัวจากเส้นใยขนานของเซลล์เม็ดเล็ก อีกประเภทหนึ่งคืออินพุตที่แข็งแกร่งมากจากเส้นใยปีนป่ายเส้น เดียว [ 7 ]แนวคิดพื้นฐานของทฤษฎี Marr–Albusคือเส้นใยปีนป่ายทำหน้าที่เป็น "สัญญาณการสอน" ซึ่งกระตุ้นให้เกิดการเปลี่ยนแปลงที่ยั่งยืนในความแข็งแรงของอินพุตเส้นใยขนาน การสังเกตภาวะซึมเศร้าระยะยาวในอินพุตเส้นใยขนานได้ให้การสนับสนุนทฤษฎีประเภทนี้บ้าง แต่ความถูกต้องของทฤษฎีเหล่านี้ยังคงเป็นที่ถกเถียงกันอยู่[ 8 ]
โครงสร้าง
ในระดับกายวิภาคศาสตร์โดยรวมสมองน้อยประกอบด้วยชั้นคอร์เทกซ์ ที่พับซ้อนกันอย่างแน่นหนา โดยมีเนื้อขาว อยู่ด้านล่างและ โพรงที่เต็มไปด้วยของเหลวที่ฐานนิวเคลียสของสมองน้อยสี่อันที่อยู่ลึกฝังอยู่ในเนื้อขาว[ 9 ]แต่ละส่วนของคอร์เทกซ์ประกอบด้วยชุดของเซลล์ประสาทขนาดเล็กชุดเดียวกัน ซึ่งวางเรียงกันในรูปทรงเรขาคณิตที่เป็นแบบแผนอย่างมาก ในระดับกลาง สมองน้อยและโครงสร้างเสริมสามารถแยกออกเป็นโมดูลที่ทำงานได้อย่างอิสระหลายร้อยหรือหลายพันโมดูล เรียกว่า "ไมโครโซน" หรือ "ไมโครคอมพาร์ทเมนต์"
กายวิภาคศาสตร์ระดับมหภาค

สมองน้อยตั้งอยู่ในโพรงกะโหลกส่วนหลังโพรงสมองที่สี่พอนส์และเมดุลลาอยู่ด้านหน้าของสมองน้อย[ 10 ]มันถูกแยกออกจากสมอง ใหญ่ โดยเยื่อดูรามาเตอร์ที่ มีลักษณะคล้ายหนัง เรียกว่า cerebellar tentoriumการเชื่อมต่อทั้งหมดกับส่วนอื่นๆ ของสมองจะผ่านทางพอนส์ นักกายวิภาคศาสตร์จัดให้สมองน้อยเป็นส่วนหนึ่งของmetencephalonซึ่งรวมถึงพอนส์ด้วย metencephalon เป็นส่วนบนของrhombencephalonหรือ "สมองส่วนหลัง" เช่นเดียวกับเปลือกสมอง สมองน้อยถูกแบ่งออกเป็นสองซีกสมองน้อยนอกจากนี้ยังมีโซนกลางที่แคบ ( vermis ) ตามธรรมเนียมแล้ว จะใช้รอยพับขนาดใหญ่เพื่อแบ่งโครงสร้างโดยรวมออกเป็น 10 "กลีบ" เล็กๆ เนื่องจากมีเซลล์เม็ด เล็กจำนวนมาก สมองส่วนซีรีเบลลัมจึงมีเซลล์ประสาท มากกว่าเซลล์ ประสาททั้งหมดในสมองส่วนอื่น ๆ แต่มีปริมาตรเพียง 10% ของปริมาตรสมองทั้งหมด[ 11 ]จำนวนเซลล์ประสาทในซีรีเบลลัมมีความสัมพันธ์กับจำนวนเซลล์ประสาทในนีโอคอร์เทกซ์โดยมีเซลล์ประสาทในซีรีเบลลัมมากกว่าในนีโอคอร์เทกซ์ประมาณ 3.6 เท่า ซึ่งเป็นอัตราส่วนที่พบได้ในสัตว์เลี้ยงลูกด้วยนมหลายชนิด[ 12 ]
ลักษณะพื้นผิวที่ผิดปกติของสมองน้อยนั้นซ่อนความจริงที่ว่าปริมาตรส่วนใหญ่ของมันประกอบด้วยชั้นของเนื้อเยื่อสีเทา ที่พับซ้อนกันอย่างแน่นหนา นั่น คือ เปลือกสมองน้อย (cerebellar cortex ) สันหรือร่องแต่ละอันในชั้นนี้เรียกว่าโฟเลียม (folium ) การตรวจ MRI ความละเอียดสูงพบว่าเปลือกสมองน้อยของมนุษย์วัยผู้ใหญ่มีพื้นที่ 730 ตารางเซนติเมตร[ 13 ]บรรจุอยู่ภายในปริมาตรที่มีขนาด 6 ซม. × 5 ซม. × 10 ซม. [ 11 ]ใต้เนื้อเยื่อสีเทาของเปลือกสมองน้อยคือเนื้อเยื่อสีขาวซึ่งส่วนใหญ่ประกอบด้วย เส้นใยประสาท ที่มีปลอกไมอีลินหุ้มวิ่งเข้าและออกจากเปลือกสมองน้อย ภายในเนื้อเยื่อสีขาว—ซึ่งบางครั้งเรียกว่า อาร์บอร์ วิเท (ต้นไม้แห่งชีวิต) เนื่องจากมีลักษณะเป็นกิ่งก้านสาขาคล้ายต้นไม้เมื่อมองจากภาคตัดขวาง—มีนิวเคลียสของสมองน้อยสี่อันที่อยู่ลึกเข้าไปซึ่งประกอบด้วยเนื้อเยื่อสีเทา[ 14 ]
สมองน้อยเชื่อมต่อกับส่วนต่างๆ ของระบบประสาทด้วยก้านสมองน้อย สามคู่ ได้แก่ก้านสมองน้อยส่วนบนก้านสมองน้อยส่วนกลางและก้านสมองน้อยส่วนล่างซึ่งตั้งชื่อตามตำแหน่งที่สัมพันธ์กับเวอร์มิส ก้านสมองน้อยส่วนบนส่วนใหญ่ทำหน้าที่ส่งสัญญาณออกไปยังเปลือกสมอง โดยนำเส้นใยประสาทส่งออกผ่านนิวเคลียสของทาลามัสไปยังเซลล์ประสาทสั่งการส่วนบนในเปลือกสมอง เส้นใยเหล่านี้เกิดขึ้นจากนิวเคลียสของสมองน้อยส่วนลึก ก้านสมองน้อยส่วนกลางเชื่อมต่อกับพอนส์และรับสัญญาณเข้าทั้งหมดจากพอนส์ โดยส่วนใหญ่มาจากนิวเคลียสของพอนส์สัญญาณเข้าสู่พอนส์มาจากเปลือกสมองและถูกส่งต่อจากนิวเคลียสของพอนส์ผ่านเส้นใยประสาทตามขวางของพอนส์ไปยังสมองน้อย ก้านสมองส่วนกลางมีขนาดใหญ่ที่สุดในบรรดาก้านสมองทั้งสาม และเส้นใยประสาทนำเข้าของก้านสมองส่วนกลางจะถูกจัดกลุ่มเป็นมัดแยกกันสามมัด โดยนำข้อมูลนำเข้าไปยังส่วนต่างๆ ของสมองน้อย ก้านสมองน้อยส่วนล่างได้รับข้อมูลนำเข้าจากเส้นใยประสาทนำเข้าจากนิวเคลียสเวสติบูลาร์ ไขสันหลัง และเทกเมนตัม การส่งออกของก้านสมองน้อยส่วนล่างเกิดขึ้นผ่านเส้นใยประสาทนำออกไปยังนิวเคลียสเวสติบูลาร์และเรติคูลาร์ฟอร์เมชันสมองน้อยทั้งหมดได้รับข้อมูลนำเข้าแบบปรับเปลี่ยนจากนิวเคลียสโอลิวารีส่วนล่างผ่านทางก้านสมองน้อยส่วนล่าง[ 6 ]
การแบ่งย่อย
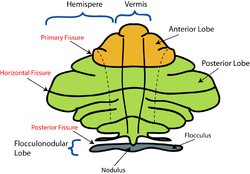
จากลักษณะภายนอกสามารถแบ่งซีรีเบลลัม ออกเป็น 3 ส่วน ได้แก่ ส่วนหน้า (เหนือร่องหลัก ) ส่วน หลัง (ใต้ร่องหลัก) และส่วนฟลอคคูโลโนดูลาร์ (ใต้ร่องหลัง) ส่วนเหล่านี้แบ่งซีรีเบลลัมจากด้านหน้าไปด้านหลัง (ในมนุษย์ จากบนลงล่าง) อย่างไรก็ตาม ในแง่ของหน้าที่การทำงาน มีความแตกต่างที่สำคัญกว่าในแนวกลางไปด้านข้าง หากไม่นับส่วนฟลอคคูโลโนดูลาร์ ซึ่งมีการเชื่อมต่อและหน้าที่ที่แตกต่างกัน ซีรีเบลลัมสามารถแบ่งตามหน้าที่ได้เป็นส่วนตรงกลางที่เรียกว่าสไปโนซีรี เบลลัม และส่วนด้านข้างที่ใหญ่กว่าที่เรียกว่า ซีรีโบรซีรีเบลลัม [ 14 ] เนื้อเยื่อที่ยื่นออกมาเป็นแถบแคบๆ ตามแนวกลางเรียกว่าซีรีเบลลาร์เวอร์มิส ( เวอร์มิสเป็นภาษาละตินแปลว่า "หนอน") [ 14 ]
บริเวณที่เล็กที่สุดคือกลีบฟลอคคูโลโนดูลาร์ ซึ่งมักเรียกว่าเวสติบูโลซีรีเบลลัมเป็นส่วนที่เก่าแก่ที่สุดในแง่ของวิวัฒนาการ (อาร์คิซีรีเบลลัม) และมีส่วนร่วมหลักในการทรงตัวและการวางแนวเชิงพื้นที่ การเชื่อมต่อหลักของมันคือกับนิวเคลียสเวสติบูลาร์แม้ว่าจะได้รับข้อมูลภาพและข้อมูลประสาทสัมผัสอื่นๆ ด้วยก็ตาม ความเสียหายต่อบริเวณนี้ทำให้เกิดความผิดปกติของการทรงตัวและการเดิน[ 14 ]
บริเวณตรงกลางของกลีบหน้าและกลีบหลังประกอบเป็นสไปโนเซเรเบลลัม หรือที่รู้จักกันในชื่อพาเลโอเซเรเบลลัม ส่วนนี้ของเซเรเบลลัมทำหน้าที่หลักในการปรับแต่งการเคลื่อนไหวของร่างกายและแขนขา มันรับ ข้อมูล การรับรู้ตำแหน่งจากคอลัมน์ด้านหลังของไขสันหลัง (รวมถึงเส้นทางสไปโนเซเรเบลลัม ) และจากเส้นประสาทไตรเจมินัล ของสมอง รวมถึงจากระบบ การมองเห็นและ การได้ยิน[ 15 ]มันส่งเส้นใยไปยังนิวเคลียสเซเรเบลลัมส่วนลึก ซึ่งจะฉายไปยังทั้งเปลือกสมองและก้านสมอง จึงทำให้เกิดการปรับเปลี่ยนระบบการเคลื่อนไหวจากส่วนบนลงล่าง[ 14 ]
บริเวณด้านข้าง ซึ่งในมนุษย์ถือเป็นส่วนที่ใหญ่ที่สุด ประกอบด้วยซีรีบรอซีรีเบลลัม หรือที่รู้จักกันในชื่อนีโอซีรีเบลลัม มันรับข้อมูลเข้าจากเปลือกสมอง (โดยเฉพาะกลีบข้างขมับ ) ผ่านนิวเคลียสพอนไทน์ (สร้างเส้นทางคอร์ติโค-พอนโต-ซีรีเบลลาร์) และส่งข้อมูลออกส่วนใหญ่ไปยังทาลามัสเวนโทร ลาเทอรัล (ซึ่งเชื่อมต่อกับบริเวณมอเตอร์ของเปลือกสมองพรีมอเตอร์และบริเวณมอเตอร์หลักของเปลือกสมอง) และไปยังนิวเคลียสสีแดง[ 14 ]มีความเห็นไม่ตรงกันเกี่ยวกับวิธีที่ดีที่สุดในการอธิบายหน้าที่ของซีรีเบลลัมด้านข้าง: เชื่อกันว่ามันเกี่ยวข้องกับการวางแผนการเคลื่อนไหวที่กำลังจะเกิดขึ้น[ 16 ]ในการประเมินข้อมูลทางประสาทสัมผัสเพื่อการกระทำ[ 14 ]และในหน้าที่ทางปัญญาล้วนๆ หลายอย่าง เช่น การกำหนดคำกริยาที่เหมาะสมที่สุดกับคำนามบางคำ (เช่น "นั่ง" สำหรับ "เก้าอี้") [ 17 ] [ 18 ] [ 19 ] [ 20 ]
จุลกายวิภาคศาสตร์
เซลล์ประสาทสองประเภทมีบทบาทสำคัญในวงจรซีรีเบลลัม ได้แก่เซลล์ Purkinjeและเซลล์แกรนูล นอกจากนี้ แอกซอนสามประเภทก็มีบทบาทสำคัญเช่นกัน ได้แก่เส้นใยมอสซีและเส้นใยปีนป่าย (ซึ่งเข้าสู่ซีรีเบลลัมจากภายนอก) และเส้นใยขนาน (ซึ่งเป็นแอกซอนของเซลล์แกรนูล) มีเส้นทางหลักสองเส้นทางผ่านวงจรซีรีเบลลัม โดยมีต้นกำเนิดจากเส้นใยมอสซีและเส้นใยปีนป่าย ซึ่งทั้งสองเส้นทางจะสิ้นสุดในนิวเคลียสซีรีเบลลัมส่วนลึกในที่สุด[ 11 ]
เส้นใยมอสซีส่งตรงไปยังนิวเคลียสส่วนลึก แต่ยังก่อให้เกิดเส้นทางต่อไปนี้ด้วย: เส้นใยมอสซี → เซลล์แกรนูล → เส้นใยขนาน → เซลล์พูร์คินเจ → นิวเคลียสส่วนลึก เส้นใยไต่ส่งไปยังเซลล์พูร์คินเจและยังส่งแขนงไปยังนิวเคลียสส่วนลึกโดยตรง[ 11 ]อินพุตของเส้นใยมอสซีและเส้นใยไต่แต่ละเส้นจะส่งข้อมูลเฉพาะของเส้นใยนั้นๆ สมองน้อยยังได้รับ อินพุต โดปามีน เซโรโทนิน นอร์อะดรีนาลินและโคลีนซึ่งคาดว่าจะทำหน้าที่ปรับเปลี่ยนโดยรวม[ 21 ]
ชั้นคอร์เทกซ์
เปลือกสมองน้อยแบ่งออกเป็นสามชั้น ชั้นล่างสุดคือชั้นเม็ดหนา ซึ่งอัดแน่นไปด้วยเซลล์เม็ดพร้อมกับเซลล์ประสาทระหว่างเซลล์ส่วนใหญ่เป็นเซลล์กอลจิแต่ยังรวมถึงเซลล์ลูกาโรและเซลล์แปรงขั้วเดียวด้วย ชั้นกลางคือชั้นพูร์คินเจ ซึ่งเป็นโซนแคบๆ ที่มีตัวเซลล์ของเซลล์พูร์คินเจและเซลล์เกลียเบิร์กมันน์ ชั้นบนสุดคือชั้นโมเลกุล ซึ่งมี ต้น เดนไดรต์ แบนราบ ของเซลล์พูร์คินเจ พร้อมกับเส้นใยขนานจำนวนมากที่แทรกผ่านต้นเดนไดรต์ของเซลล์พูร์คินเจในมุมฉาก ชั้นนอกสุดของเปลือกสมองน้อยนี้ยังประกอบด้วยเซลล์ประสาทระหว่างเซลล์ยับยั้งสองประเภท ได้แก่เซลล์รูปดาวและเซลล์รูปตะกร้าทั้งเซลล์รูปดาวและเซลล์รูปตะกร้าสร้าง ไซแนปส์ GABAergicบนเดนไดรต์ของเซลล์พูร์คินเจ[ 11 ]


ชั้นโมเลกุล
ชั้นบนสุดด้านนอกสุดของเปลือกสมองน้อยคือชั้นโมเลกุล ชั้นนี้ประกอบด้วยเดนไดรต์ ที่แบนราบ ของเซลล์ Purkinje และเส้นใยขนานจำนวนมากจากชั้นเม็ดเล็กที่แทรกเข้าไปในเดนไดรต์ของเซลล์ Purkinje ในมุมฉาก ชั้นโมเลกุลยังประกอบด้วยอินเตอร์นิวรอนยับยั้งสองประเภท ได้แก่เซลล์รูปดาวและเซลล์รูปตะกร้าทั้งเซลล์รูปดาวและเซลล์รูปตะกร้าสร้าง ไซแนปส์ GABAergicบนเดนไดรต์ของเซลล์ Purkinje [ 11 ]
ชั้นพูร์คินเจ
เซลล์ Purkinjeเป็นหนึ่งในเซลล์ประสาทที่โดดเด่นที่สุดในสมอง และเป็นหนึ่งในประเภทแรกๆ ที่ได้รับการยอมรับ โดยนักกายวิภาคชาวเช็กJan Evangelista Purkyně เป็นผู้บรรยาย ลักษณะเซลล์นี้เป็นครั้งแรกในปี 1837 เซลล์เหล่านี้มีลักษณะเด่นที่รูปร่างของเดนไดรต์ คือ เดนไดรต์แตกแขนงอย่างมากมาย แต่แบนราบอย่างมากในระนาบที่ตั้งฉากกับรอยพับของสมองน้อย ดังนั้น เดนไดรต์ของเซลล์ Purkinje จึงก่อตัวเป็นโครงข่ายระนาบที่หนาแน่น ซึ่งเส้นใยขนานจะผ่านในมุมฉาก[ 11 ]เดนไดรต์ถูกปกคลุมด้วยหนามเดนไดรต์ซึ่งแต่ละหนามจะรับสัญญาณประสาทจากเส้นใยขนาน เซลล์ Purkinje ได้รับสัญญาณประสาทมากกว่าเซลล์ประเภทอื่นๆ ในสมอง โดยมีการประมาณจำนวนหนามบนเซลล์ Purkinje ของมนุษย์เพียงเซลล์เดียวสูงถึง 200,000 อัน[ 11 ]เซลล์ Purkinje ซึ่งมีรูปร่างทรงกลมขนาดใหญ่ จะเรียงตัวกันเป็นชั้นแคบๆ (หนาเพียงเซลล์เดียว) ในเปลือกสมองส่วนซีรีเบลลัม เรียกว่าชั้น Purkinjeหลังจากปล่อยแขนงที่ส่งผลต่อส่วนต่างๆ ของเปลือกสมองที่อยู่ใกล้เคียงแล้ว แอกซอนของเซลล์เหล่านี้จะเดินทางเข้าไปในนิวเคลียสของซีรีเบลลัมส่วนลึกซึ่งจะสร้างการติดต่อประมาณ 1,000 ครั้งกับเซลล์นิวเคลียสหลายประเภท ทั้งหมดนี้อยู่ในบริเวณเล็กๆ เซลล์ Purkinje ใช้GABAเป็นสารสื่อประสาท ดังนั้นจึงออกฤทธิ์ยับยั้งเป้าหมายของพวกมัน[ 11 ]
เซลล์ Purkinje เป็นหัวใจของวงจรสมองน้อย และขนาดที่ใหญ่และรูปแบบการทำงานที่โดดเด่นทำให้การศึกษาแบบแผนการตอบสนองในสัตว์ที่กำลังแสดงพฤติกรรมโดยใช้เทคนิคการบันทึกนอกเซลล์ ทำได้ค่อนข้างง่าย เซลล์ Purkinje โดยปกติจะปล่อย ศักยภาพการกระทำในอัตราสูงแม้ว่าจะไม่มีการป้อนข้อมูลจากไซแนปส์ก็ตาม ในสัตว์ที่ตื่นตัวและแสดงพฤติกรรม อัตราเฉลี่ยประมาณ 40 Hz ถือเป็นเรื่องปกติ ชุดสไปค์แสดงให้เห็นส่วนผสมของสิ่งที่เรียกว่าสไปค์แบบง่ายและแบบซับซ้อน สไปค์แบบง่ายคือศักยภาพการกระทำเพียงครั้งเดียวตามด้วยช่วงเวลาพักประมาณ 10 มิลลิวินาที สไปค์แบบซับซ้อนคือลำดับของศักยภาพการกระทำที่มีรูปแบบตายตัวโดยมีช่วงเวลาระหว่างสไปค์สั้นมากและแอมพลิจูดลดลง[ 23 ]การศึกษาทางสรีรวิทยาแสดงให้เห็นว่าสไปค์แบบซับซ้อน (ซึ่งเกิดขึ้นที่อัตราพื้นฐานประมาณ 1 Hz และไม่เคยเกิดขึ้นที่อัตราสูงกว่า 10 Hz มากนัก) มีความสัมพันธ์อย่างน่าเชื่อถือกับการกระตุ้นเส้นใยปีนป่าย ในขณะที่สไปค์แบบง่ายเกิดจากการรวมกันของกิจกรรมพื้นฐานและการป้อนข้อมูลจากเส้นใยขนาน สไปค์ที่ซับซ้อนมักจะตามมาด้วยช่วงหยุดชั่วคราวหลายร้อยมิลลิวินาที ซึ่งในระหว่างนั้นกิจกรรมสไปค์แบบง่ายจะถูกระงับ[ 24 ]
ลักษณะเฉพาะที่สามารถระบุได้ของเซลล์ประสาท Purkinje คือการแสดงออกของแคลบินดิน [ 25 ] การย้อมสีแคลบินดินในสมองหนูหลังจากได้รับบาดเจ็บที่เส้นประสาทไซแอติกเรื้อรังข้างเดียว แสดงให้เห็นว่าเซลล์ประสาท Purkinje อาจถูกสร้างขึ้นใหม่ในสมองของผู้ใหญ่ ซึ่งเป็นการเริ่มต้นการจัดระเบียบของกลีบสมองน้อยใหม่[ 26 ]

ชั้นเม็ด

เซลล์แกรนูลของสมองน้อยนั้น แตกต่างจากเซลล์พูร์คินเจตรงที่เป็นเซลล์ประสาทที่เล็กที่สุดในสมอง นอกจากนี้ยังเป็นเซลล์ประสาทที่มีจำนวนมากที่สุดในสมองด้วย โดยในมนุษย์นั้น มีการประมาณจำนวนรวมเฉลี่ยอยู่ที่ประมาณ 50 พันล้านเซลล์ ซึ่งหมายความว่าประมาณ 3/4 ของเซลล์ประสาทในสมองเป็นเซลล์แกรนูลของสมองน้อย[ 11 ]ตัวเซลล์ของพวกมันถูกอัดแน่นเป็นชั้นหนาที่ด้านล่างของเปลือกสมองน้อย เซลล์แกรนูลจะปล่อยเดนไดรต์ออกมาเพียง 4-5 เส้น โดยแต่ละเส้นจะสิ้นสุดที่ส่วนขยายที่เรียกว่ากรงเล็บเดนไดรต์ [ 11 ] ส่วนขยายเหล่านี้เป็นตำแหน่งของอินพุตกระตุ้นจากเส้นใยมอสซีและอินพุตยับยั้งจากเซลล์กอลจิ[ 11 ]
แอกซอนที่บางและไม่มีปลอกไมอีลินของเซลล์แกรนูลจะตั้งตรงขึ้นไปยังชั้นบน (ระดับโมเลกุล) ของคอร์เทกซ์ ซึ่งแอกซอนเหล่านี้จะแยกออกเป็นสองส่วน โดยแต่ละส่วนจะเคลื่อนที่ในแนวนอนเพื่อสร้างเส้นใยขนานการแยกของส่วนที่ตั้งตรงออกเป็นสองส่วนในแนวนอนทำให้เกิดรูปร่าง "T" ที่โดดเด่น เส้นใยขนานของมนุษย์มีความยาวเฉลี่ย 3 มม. ในแต่ละทิศทางจากจุดแยก โดยมีความยาวรวมประมาณ 6 มม. (ประมาณ 1/10 ของความกว้างทั้งหมดของชั้นคอร์เทกซ์) [ 11 ]ขณะที่วิ่งไป เส้นใยขนานจะผ่านต้นเดนไดรต์ของเซลล์พูร์คินเจ โดยจะสัมผัสกับเซลล์พูร์คินเจ 1 ใน 3-5 เซลล์ที่ผ่าน ทำให้เกิดการเชื่อมต่อไซแนปส์ทั้งหมด 80-100 จุดกับหนามเดนไดรต์ของเซลล์พูร์คินเจ[ 11 ]เซลล์แกรนูลใช้กลูตาเมตเป็นสารสื่อประสาท ดังนั้นจึงออกฤทธิ์กระตุ้นต่อเป้าหมาย[ 11 ]

เซลล์แกรนูลได้รับข้อมูลทั้งหมดจากเส้นใยมอสซี แต่มีจำนวนมากกว่าเส้นใยมอสซีถึง 200 ต่อ 1 (ในมนุษย์) ดังนั้น ข้อมูลในสถานะกิจกรรมของประชากรเซลล์แกรนูลจึงเหมือนกับข้อมูลในเส้นใยมอสซี แต่ถูกเข้ารหัสใหม่ในรูปแบบที่กว้างขวางกว่ามาก เนื่องจากเซลล์แกรนูลมีขนาดเล็กและหนาแน่นมาก จึงยากที่จะบันทึกกิจกรรมการส่งสัญญาณของพวกมันในสัตว์ที่กำลังแสดงพฤติกรรม ดังนั้นจึงมีข้อมูลน้อยมากที่จะใช้เป็นพื้นฐานสำหรับการสร้างทฤษฎี แนวคิดที่เป็นที่นิยมที่สุดเกี่ยวกับหน้าที่ของพวกมันถูกเสนอในปี 1969 โดยเดวิด มาร์ผู้ซึ่งแนะนำว่าพวกมันสามารถเข้ารหัสการรวมกันของข้อมูลนำเข้าจากเส้นใยมอสซีได้ แนวคิดก็คือ เซลล์แกรนูลแต่ละเซลล์ได้รับข้อมูลจากเส้นใยมอสซีเพียง 4-5 เส้น เซลล์แกรนูลจะไม่ตอบสนองหากมีเพียงข้อมูลนำเข้าเดียวที่ทำงานอยู่ แต่จะตอบสนองหากมีข้อมูลนำเข้ามากกว่าหนึ่งรายการที่ทำงานอยู่ แผนการเข้ารหัสแบบผสมผสานนี้อาจทำให้สมองน้อยสามารถแยกแยะความแตกต่างระหว่างรูปแบบข้อมูลนำเข้าได้ละเอียดกว่าที่เส้นใยมอสซีเพียงอย่างเดียวจะทำได้[ 27 ]
เส้นใยคล้ายมอส
เส้นใยมอสซีเข้าสู่ชั้นแกรนูลาร์จากจุดกำเนิด ซึ่งหลายเส้นเกิดขึ้นจากนิวเคลียสพอนไทน์ บางเส้นมาจากไขสันหลังนิวเคลียสเวสติบูลาร์เป็นต้น ในสมองน้อยของมนุษย์ มีการประมาณจำนวนเส้นใยมอสซีทั้งหมดไว้ที่ 200 ล้านเส้น[ 11 ]เส้นใยเหล่านี้สร้างไซแนปส์กระตุ้นกับเซลล์แกรนูลและเซลล์ของนิวเคลียสสมองน้อยส่วนลึก ภายในชั้นแกรนูลาร์ เส้นใยมอสซีจะสร้างส่วนขยายหลายชุดที่เรียกว่าโรเซ็ตการสัมผัสระหว่างเส้นใยมอสซีและเดนไดรต์ของเซลล์แกรนูลเกิดขึ้นภายในโครงสร้างที่เรียกว่าโกลเมอ รูลัส โกลเมอ รูลัสแต่ละอันมีโรเซ็ตเส้นใยมอสซีอยู่ตรงกลาง และมีกรงเล็บเดนไดรต์ของเซลล์แกรนูลมากถึง 20 อันที่สัมผัสกับมัน ปลายประสาทจากเซลล์กอลจิแทรกซึมเข้าไปในโครงสร้างและสร้างไซแนปส์ยับยั้งบนเดนไดรต์ของเซลล์แกรนูล กลุ่มทั้งหมดถูกล้อมรอบด้วยปลอกของเซลล์เกลีย[ 11 ]เส้นใยมอสซีแต่ละเส้นส่งกิ่งก้านสาขาไปยังโฟเลียของสมองน้อยหลายแห่ง ทำให้เกิดโรเซ็ตทั้งหมด 20–30 โรเซ็ต ดังนั้นเส้นใยมอสซีเส้นเดียวจึงสัมผัสกับเซลล์แกรนูลประมาณ 400–600 เซลล์[ 11 ]
เส้นใยปีนป่าย
เซลล์ Purkinje ยังได้รับอินพุตจากนิวเคลียส inferior olivaryที่อยู่ด้านตรงข้ามของก้านสมองผ่านเส้นใย climbing fiberแม้ว่า inferior olive จะอยู่ในmedulla oblongataและได้รับอินพุตจากไขสันหลัง ก้านสมอง และเปลือกสมอง แต่เอาต์พุตทั้งหมดจะส่งไปยังสมองน้อย เส้นใย climbing fiber จะส่งแขนงไปยังนิวเคลียสของสมองน้อยส่วนลึกก่อนที่จะเข้าสู่เปลือกสมองน้อย ซึ่งจะแยกออกเป็นแขนงปลายทางประมาณ 10 แขนง โดยแต่ละแขนงจะส่งสัญญาณไปยังเซลล์ Purkinje เพียงเซลล์เดียว[ 11 ]ในทางตรงกันข้ามกับอินพุตมากกว่า 100,000 รายการจากเส้นใยขนาน เซลล์ Purkinje แต่ละเซลล์จะได้รับอินพุตจากเส้นใย climbing fiber เพียงเส้นเดียว แต่เส้นใยเส้นเดียวนี้จะ "ปีน" ไปตามเดนไดรต์ของเซลล์ Purkinje พันรอบพวกมันและสร้างไซแนปส์ได้มากถึง 300 จุด[ 11 ]อินพุตสุทธิมีความแรงมากจนศักยภาพการกระทำเพียงครั้งเดียวจากเส้นใยปีนป่ายสามารถสร้างสไปค์ที่ซับซ้อนแบบขยายในเซลล์ Purkinje ได้: การระเบิดของสไปค์หลายตัวติดต่อกัน โดยมีแอมพลิจูดลดลง ตามด้วยช่วงหยุดชั่วคราวซึ่งกิจกรรมจะถูกระงับ ซินแนปส์ของเส้นใยปีนป่ายครอบคลุมตัวเซลล์และเดนไดรต์ส่วนต้น บริเวณนี้ไม่มีอินพุตจากเส้นใยขนาน[ 11 ]
เส้นใยปีนป่าย (Climbing fibers) ส่งสัญญาณในอัตราต่ำ แต่ศักยภาพการกระทำของเส้นใยปีนป่ายเพียงเส้นเดียวสามารถกระตุ้นให้เกิดศักยภาพการกระทำหลายเส้นในเซลล์ Purkinje เป้าหมาย (สไปค์ที่ซับซ้อน) ความแตกต่างระหว่างการรับสัญญาณจากเส้นใยขนาน (Parallel fiber) และเส้นใยปีนป่าย (Climbing fiber) ไปยังเซลล์ Purkinje (มากกว่า 100,000 เส้นของชนิดหนึ่ง เทียบกับเพียงหนึ่งเส้นของอีกชนิดหนึ่ง) อาจเป็นลักษณะที่น่าสนใจที่สุดของกายวิภาคของสมองส่วนซีรีเบลลัม และเป็นแรงบันดาลใจให้เกิดทฤษฎีมากมาย อันที่จริง หน้าที่ของเส้นใยปีนป่ายเป็นหัวข้อที่ถกเถียงกันมากที่สุดเกี่ยวกับสมองส่วนซีรีเบลลัม มีสองสำนักคิด สำนักคิดหนึ่งยึดตาม Marr และ Albus ที่กล่าวว่าการรับสัญญาณจากเส้นใยปีนป่ายทำหน้าที่เป็นสัญญาณการสอนเป็นหลัก อีกสำนักคิดหนึ่งกล่าวว่าหน้าที่ของมันคือการกำหนดรูปร่างของผลลัพธ์ของซีรีเบลลัมโดยตรง ทั้งสองมุมมองได้รับการปกป้องอย่างละเอียดในสิ่งพิมพ์จำนวนมาก ตามคำกล่าวในบทวิจารณ์หนึ่ง “ในการพยายามสังเคราะห์สมมติฐานต่างๆ เกี่ยวกับหน้าที่ของเส้นใยปีนป่ายนั้น ให้ความรู้สึกเหมือนกำลังมองภาพวาดของเอสเชอร์ มุมมองแต่ละจุดดูเหมือนจะอธิบายถึงการค้นพบชุดหนึ่ง แต่เมื่อพยายามนำมุมมองต่างๆ มารวมกัน ภาพรวมที่สอดคล้องกันของสิ่งที่เส้นใยปีนป่ายกำลังทำนั้นกลับไม่ปรากฏ สำหรับนักวิจัยส่วนใหญ่ เส้นใยปีนป่ายส่งสัญญาณข้อผิดพลาดในการทำงานของมอเตอร์ ไม่ว่าจะในลักษณะปกติของการปรับความถี่การปล่อยประจุ หรือเป็นการประกาศเพียงครั้งเดียวของ 'เหตุการณ์ที่ไม่คาดคิด' สำหรับนักวิจัยคนอื่นๆ ข้อความนั้นอยู่ที่ระดับของการประสานกันและจังหวะของกลุ่มเส้นใยปีนป่าย” [ 24 ]
นิวเคลียสส่วนลึก

นิวเคลียสส่วนลึกของสม cerebellum คือกลุ่มของเนื้อเยื่อสีเทาที่อยู่ภายในเนื้อเยื่อสีขาวบริเวณแกนกลางของสม cerebellum โดยส่วนใหญ่แล้ว (ยกเว้นนิวเคลียสเวสติบูลาร์ที่อยู่ใกล้เคียง) เป็นแหล่งส่งสัญญาณออกเพียงแหล่งเดียวจากสม cerebellum นิวเคลียส เหล่านี้ รับสัญญาณจากเส้นใย mossy และเส้นใย climbing รวมถึงสัญญาณยับยั้งจากเซลล์ Purkinje ของเปลือกสม cerebellum นิวเคลียสทั้งสี่ ( dentate , globose , emboliformและfastigial ) แต่ละนิวเคลียสสื่อสารกับส่วนต่างๆ ของสมองและเปลือกสม cerebellum (นิวเคลียส globose และ emboliform เรียกอีกอย่างว่านิวเคลียสรวมในนิวเคลียสแทรก ) ส่วนนิวเคลียส fastigial และ interposed เป็นส่วนหนึ่งของ spinocerebellum นิวเคลียสเดนเทต ซึ่งในสัตว์เลี้ยงลูกด้วยนมมีขนาดใหญ่กว่านิวเคลียสอื่นๆ มาก ก่อตัวเป็นชั้นเนื้อสีเทาที่บางและคดเคี้ยว และสื่อสารเฉพาะกับส่วนด้านข้างของเปลือกสมองน้อยเท่านั้น ฟลอคคูลัสของกลีบฟลอคคูโลโนดูลาร์เป็นส่วนเดียวของเปลือกสมองน้อยที่ไม่ส่งสัญญาณไปยังนิวเคลียสส่วนลึก แต่ส่งสัญญาณไปยังนิวเคลียสเวสติบูลาร์แทน[ 11 ]
เซลล์ประสาทส่วนใหญ่ในนิวเคลียสส่วนลึกมีตัวเซลล์ขนาดใหญ่และเดนไดรต์รูปทรงกลมที่มีรัศมีประมาณ 400 ไมโครเมตร และใช้กลูตาเมตเป็นสารสื่อประสาท เซลล์เหล่านี้ส่งสัญญาณไปยังเป้าหมายต่างๆ นอกสมองน้อย นอกจากนี้ยังมีเซลล์ขนาดเล็กจำนวนน้อยกว่าซึ่งใช้GABAเป็นสารสื่อประสาทและส่งสัญญาณไปยังนิวเคลียสโอลิวารีส่วนล่าง เท่านั้น ซึ่งเป็นแหล่งกำเนิดของเส้นใย ปีนป่าย ดังนั้น การส่งสัญญาณจากนิวเคลียสไปยังโอลิวารีจึงให้ผลตอบ รับยับยั้ง เพื่อให้ตรงกับการส่งสัญญาณกระตุ้นของเส้นใยปีนป่ายไปยังนิวเคลียส มีหลักฐานว่ากลุ่มเซลล์นิวเคลียสขนาดเล็กแต่ละกลุ่มส่งสัญญาณไปยังกลุ่มเซลล์โอลิวารีกลุ่มเดียวกันกับที่ส่งเส้นใยปีนป่ายไปยังกลุ่มนั้น มีโครงสร้างทางภูมิศาสตร์ที่แข็งแกร่งและตรงกันในทั้งสองทิศทาง[ 11 ]
เมื่อแอกซอนของเซลล์ Purkinje เข้าสู่หนึ่งในนิวเคลียสลึก มันจะแตกแขนงเพื่อติดต่อกับเซลล์นิวเคลียสทั้งขนาดใหญ่และขนาดเล็ก แต่จำนวนเซลล์ทั้งหมดที่ติดต่อมีเพียงประมาณ 35 เซลล์ (ในแมว) ในทางกลับกัน เซลล์นิวเคลียสลึกเพียงเซลล์เดียวจะได้รับอินพุตจากเซลล์ Purkinje ประมาณ 860 เซลล์ (ในแมวเช่นกัน) [ 11 ]
ช่องต่างๆ

จากมุมมองของกายวิภาคศาสตร์โดยรวม เปลือกสมองน้อยดูเหมือนจะเป็นแผ่นเนื้อเยื่อที่เป็นเนื้อเดียวกัน และจากมุมมองของกายวิภาคศาสตร์ระดับจุลภาค ทุกส่วนของแผ่นนี้ดูเหมือนจะมีโครงสร้างภายในที่เหมือนกัน อย่างไรก็ตาม โครงสร้างของสมองน้อยนั้นมีการแบ่งเป็นส่วนๆ อยู่หลายประการ มีส่วนขนาดใหญ่ที่โดยทั่วไปเรียกว่าโซนซึ่งสามารถแบ่งออกเป็นส่วนเล็กๆ ที่เรียกว่าไมโครโซนได้[ 28 ]
ข้อบ่งชี้แรกของโครงสร้างแบบแบ่งส่วนมาจากการศึกษาขอบเขตการรับรู้ของเซลล์ในส่วนต่างๆ ของเปลือกสมองน้อย[ 28 ]แต่ละส่วนของร่างกายจะแมปไปยังจุดเฉพาะในสมองน้อย แต่มีการทำซ้ำแผนที่พื้นฐานจำนวนมาก ทำให้เกิดการจัดเรียงที่เรียกว่า "somatotopy ที่แตกหัก" [ 29 ]ข้อบ่งชี้ที่ชัดเจนยิ่งขึ้นของการแบ่งส่วนได้มาจากการย้อมสีภูมิคุ้มกันของสมองน้อยสำหรับโปรตีนบางชนิด เครื่องหมายที่รู้จักกันดีที่สุดเหล่านี้เรียกว่า "zebrins" เนื่องจากเมื่อย้อมสีแล้วจะเกิดรูปแบบที่ซับซ้อนคล้ายกับลายบนตัวม้าลาย ลายที่เกิดจาก zebrins และเครื่องหมายการแบ่งส่วนอื่นๆ จะวางตัวตั้งฉากกับรอยพับของสมองน้อย นั่นคือ จะแคบในทิศทางด้านข้าง แต่กว้างกว่ามากในทิศทางตามยาว เครื่องหมายที่แตกต่างกันจะสร้างแถบที่แตกต่างกัน ความกว้างและความยาวจะแตกต่างกันไปตามตำแหน่ง แต่โดยทั่วไปแล้วจะมีรูปร่างเหมือนกัน[ 28 ]
ในช่วงปลายทศวรรษ 1970 Oscarsson เสนอว่าโซนคอร์เทกซ์เหล่านี้สามารถแบ่งออกเป็นหน่วยย่อยที่เรียกว่าไมโครโซนได้[ 30 ]ไมโครโซนถูกกำหนดให้เป็นกลุ่มของเซลล์ Purkinje ที่มีสนามรับสัญญาณ somatotopic เดียวกันทั้งหมด พบว่าไมโครโซนแต่ละอันมีเซลล์ Purkinje ประมาณ 1,000 เซลล์ เรียงตัวเป็นแถบยาวแคบๆ วางตัวตั้งฉากกับรอยพับของคอร์เทกซ์[ 28 ]ดังนั้น ดังที่แผนภาพที่อยู่ติดกันแสดงให้เห็น เดนไดรต์ของเซลล์ Purkinje จะแบนราบไปในทิศทางเดียวกับที่ไมโครโซนขยายออกไป ในขณะที่เส้นใยขนานจะตัดผ่านพวกมันเป็นมุมฉาก[ 11 ]
โครงสร้างไมโครโซนไม่ได้ถูกกำหนดโดยสนามรับสัญญาณเพียงอย่างเดียว แต่ การป้อนข้อมูลจากเส้นใย ปีนป่ายจากนิวเคลียสโอลิวารีส่วนล่างก็มีความสำคัญไม่แพ้กัน กิ่งก้านของเส้นใยปีนป่าย (โดยปกติจะมีประมาณ 10 กิ่ง) มักจะกระตุ้นเซลล์ Purkinje ที่อยู่ในไมโครโซนเดียวกัน ยิ่งไปกว่านั้น เซลล์ประสาทโอลิวารีที่ส่งเส้นใยปีนป่ายไปยังไมโครโซนเดียวกันมักจะเชื่อมต่อกันด้วยช่องว่างเชื่อมต่อ ซึ่งจะทำให้กิจกรรมของพวกมันประสานกัน ส่งผลให้เซลล์ Purkinje ภายในไมโครโซนแสดงกิจกรรมสไปค์ที่ซับซ้อนสัมพันธ์กันในระดับเวลาเป็นมิลลิวินาที[ 28 ]นอกจากนี้ เซลล์ Purkinje ที่อยู่ในไมโครโซนทั้งหมดจะส่งแอกซอนไปยังกลุ่มเซลล์เอาต์พุตขนาดเล็กกลุ่มเดียวกันภายในนิวเคลียสสมองน้อยส่วนลึก [ 28 ] สุดท้ายแอกซอนของเซลล์ตะกร้าจะยาวกว่าในทิศทางตามยาวมากกว่าในทิศทางด้านข้าง ทำให้พวกมันถูกจำกัดอยู่ในไมโครโซนเดียวเป็นส่วนใหญ่[ 28 ]ผลที่ตามมาของโครงสร้างทั้งหมดนี้คือ ปฏิสัมพันธ์ของเซลล์ภายในไมโครโซนจะแข็งแกร่งกว่าปฏิสัมพันธ์ระหว่างไมโครโซนที่แตกต่างกันมาก[ 28 ]
ในปี พ.ศ. 2548 Richard Apps และ Martin Garwicz ได้สรุปหลักฐานว่าไมโครโซนนั้นเป็นส่วนหนึ่งของหน่วยที่ใหญ่กว่าที่เรียกว่าไมโครคอมเพล็กซ์แบบหลายโซน ไมโครคอมเพล็กซ์ดังกล่าวประกอบด้วยไมโครโซนของเปลือกสมองที่แยกจากกันในเชิงพื้นที่หลายแห่ง ซึ่งทั้งหมดส่งไปยังกลุ่มเซลล์ประสาทซีรีเบลลัมส่วนลึกกลุ่มเดียวกัน รวมทั้งกลุ่มเซลล์ประสาทโอลิวารีที่เชื่อมโยงกันซึ่งส่งไปยังไมโครโซนทั้งหมดที่รวมอยู่ ตลอดจนไปยังบริเวณนิวเคลียสส่วนลึกด้วย[ 28 ]
การไหลเวียนของเลือด
สมองส่วนซีรีเบลลัมได้รับเลือดจากหลอดเลือดแดงหลักสามคู่ ได้แก่หลอดเลือดแดงซีรีเบลลัมส่วนบน (SCA) หลอดเลือดแดงซีรีเบลลัมส่วนล่างด้านหน้า (AICA) และหลอดเลือดแดงซีรีเบลลัมส่วนล่างด้านหลัง (PICA) SCA ทำหน้าที่ส่งเลือดไปยังบริเวณส่วนบนของซีรีเบลลัม โดยจะแตกแขนงที่ผิวด้านบนและเข้าสู่เยื่อหุ้มสมองชั้นใน (pia mater) ซึ่งแขนงเหล่านี้จะเชื่อมต่อกับแขนงของหลอดเลือดแดงซีรีเบลลัมส่วนล่างด้านหน้าและด้านหลัง AICA ทำหน้าที่ส่งเลือดไปยังส่วนหน้าของด้านล่างของซีรีเบลลัม PICA มาถึงด้านล่างและแตกแขนงออกเป็นแขนงตรงกลางและแขนงด้านข้าง แขนงตรงกลางจะทอดยาวไปด้านหลังจนถึงรอยเว้าซีรีเบลลัมระหว่างซีกสมองทั้งสองข้าง ในขณะที่แขนงด้านข้างจะส่งเลือดไปยังด้านล่างของซีรีเบลลัมจนถึงขอบด้านข้าง ซึ่งจะเชื่อมต่อกับ AICA และ SCA
การทำงาน
เบาะแสที่ชัดเจนที่สุดเกี่ยวกับหน้าที่ของซีรีเบลลัมมาจากการตรวจสอบผลที่ตามมาจากการได้รับความเสียหาย สัตว์และมนุษย์ที่มีความผิดปกติของซีรีเบลลัมจะแสดงปัญหาด้านการควบคุมการเคลื่อนไหวเป็นหลัก โดยเฉพาะด้านเดียวกับส่วนของซีรีเบลลัมที่ได้รับความเสียหาย พวกเขายังคงสามารถสร้างการเคลื่อนไหวได้ แต่สูญเสียความแม่นยำ ทำให้เกิดการเคลื่อนไหวที่ผิดปกติ ไม่ประสานกัน หรือผิดจังหวะ การทดสอบมาตรฐานของการทำงานของซีรีเบลลัมคือการเอื้อมปลายนิ้วไปแตะเป้าหมายที่อยู่ห่างออกไปในระยะแขน: คนที่มีสุขภาพดีจะขยับปลายนิ้วเป็นเส้นตรงอย่างรวดเร็ว ในขณะที่คนที่มีความเสียหายของซีรีเบลลัมจะเอื้อมไปอย่างช้าๆ และผิดปกติ โดยมีการแก้ไขระหว่างทางหลายครั้ง ความบกพร่องในหน้าที่ที่ไม่เกี่ยวกับการเคลื่อนไหวจะตรวจจับได้ยากกว่า ดังนั้นข้อสรุปทั่วไปที่ได้มาเมื่อหลายสิบปีก่อนคือ หน้าที่พื้นฐานของซีรีเบลลัมคือการปรับเทียบรูปแบบโดยละเอียดของการเคลื่อนไหว ไม่ใช่การเริ่มต้นการเคลื่อนไหวหรือการตัดสินใจว่าจะทำการเคลื่อนไหวใด[ 14 ]
ก่อนปี 1990 หน้าที่ของซีรีเบลลัมส่วนใหญ่เชื่อกันว่าเกี่ยวข้องกับการเคลื่อนไหวเพียงอย่างเดียว แต่การวิจัยในภายหลังชี้ให้เห็นถึงบทบาทที่ขยายออกไปของการเชื่อมต่อของซีรีเบลลัมที่นอกเหนือไปจากหน้าที่การเคลื่อนไหวพื้นฐาน[ 31 ] การศึกษา ภาพการทำงานแสดงให้เห็นการกระตุ้นของซีรีเบลลัมที่เกี่ยวข้องกับภาษา ความสนใจ และภาพในจิตใจการศึกษาความสัมพันธ์แสดงให้เห็นปฏิสัมพันธ์ระหว่างซีรีเบลลัมและบริเวณที่ไม่เกี่ยวกับการเคลื่อนไหวของเปลือกสมอง และอาการที่ไม่เกี่ยวกับการเคลื่อนไหวต่างๆ ได้รับการยอมรับในผู้ที่มีความเสียหายที่ดูเหมือนจะจำกัดอยู่เฉพาะในซีรีเบลลัม[ 32 ] [ 33 ]โดยเฉพาะอย่างยิ่งกลุ่มอาการทางอารมณ์และการรับรู้ของซีรีเบลลัมหรือกลุ่มอาการของ Schmahmann [ 34 ]ได้รับการอธิบายในผู้ใหญ่[ 35 ]และเด็ก[ 36 ]การประมาณค่าโดยอาศัยการทำแผนที่การทำงานของสมองน้อยโดยใช้MRI เชิงฟังก์ชันบ่งชี้ว่ามากกว่าครึ่งหนึ่งของเปลือกสมองน้อยเชื่อมต่อกับโซนเชื่อมโยงของเปลือกสมอง[ 37 ]
เคนจิ โดะยะ ได้โต้แย้งว่าการทำงานของสมองน้อยนั้นเข้าใจได้ดีที่สุดไม่ใช่ในแง่ของพฤติกรรมที่มันส่งผลกระทบ แต่เป็นการคำนวณทางประสาทที่มันดำเนินการ สมองน้อยประกอบด้วยโมดูลจำนวนมากที่ค่อนข้างเป็นอิสระ โดยทั้งหมดมีโครงสร้างภายในที่เป็นระเบียบทางเรขาคณิตเหมือนกัน และด้วยเหตุนี้จึงสันนิษฐานได้ว่าทั้งหมดดำเนินการคำนวณเดียวกัน หากการเชื่อมต่อขาเข้าและขาออกของโมดูลนั้นเชื่อมต่อกับบริเวณมอเตอร์ (ซึ่งมีหลายโมดูลเป็นเช่นนั้น) โมดูลนั้นก็จะเกี่ยวข้องกับพฤติกรรมมอเตอร์ แต่ถ้าการเชื่อมต่อนั้นเชื่อมต่อกับบริเวณที่เกี่ยวข้องกับการรับรู้ที่ไม่เกี่ยวกับมอเตอร์ โมดูลนั้นก็จะแสดงความสัมพันธ์ทางพฤติกรรมประเภทอื่น ดังนั้นสมองน้อยจึงมีส่วนเกี่ยวข้องกับการควบคุมลักษณะการทำงานที่แตกต่างกันมากมาย เช่น ความรัก อารมณ์ รวมถึงการรับรู้ภาษากายทางอารมณ์[ 38 ]และพฤติกรรม[ 39 ] [ 40 ] Doya เสนอว่า สมองน้อยนั้นเข้าใจได้ดีที่สุดว่าเป็นการเลือกการกระทำเชิงทำนายโดยอาศัย "แบบจำลองภายใน" ของสภาพแวดล้อมหรือเป็นอุปกรณ์สำหรับการเรียนรู้แบบมีผู้กำกับดูแลตรงกันข้ามกับปมประสาทฐานซึ่งทำการเรียนรู้แบบเสริมแรงและเปลือกสมองซึ่งทำการเรียนรู้แบบไม่มีผู้กำกับดูแล[ 33 ] [ 41 ]การวิจัยสมองกว่าสามทศวรรษนำไปสู่ข้อเสนอที่ว่าสมองน้อยสร้างแบบจำลองทางจิตที่เหมาะสมที่สุดและมีปฏิสัมพันธ์อย่างใกล้ชิดกับเปลือกสมอง ซึ่งแบบจำลองภายในที่ได้รับการอัปเดตจะถูกรับรู้ว่าเป็นสัญชาตญาณเชิงสร้างสรรค์ ("a ha") ในหน่วยความจำใช้งาน[ 42 ]
หลักการ
ความเรียบง่ายและความสม่ำเสมอของกายวิภาคของสมองน้อยทำให้เกิดความหวังในระยะแรกว่าอาจหมายถึงความเรียบง่ายของฟังก์ชันการคำนวณที่คล้ายคลึงกัน ดังที่แสดงไว้ในหนังสือเล่มแรกๆ เกี่ยวกับสรีรวิทยาไฟฟ้าของสมองน้อยThe Cerebellum as a Neuronal MachineโดยJohn C. Eccles , Masao ItoและJános Szentágothai [ 43 ] แม้ว่าความเข้าใจอย่างถ่องแท้เกี่ยวกับการทำงานของสมองน้อยจะยังคงเป็นเรื่องที่เข้าใจยาก แต่ก็มีการระบุหลักการที่สำคัญอย่างน้อยสี่ประการ ได้แก่ (1) การประมวลผลแบบส่งต่อ (2) การแยกและการรวม (3) ความเป็นโมดูลาร์ และ (4) ความยืดหยุ่น
- การประมวลผลแบบส่งต่อ (Feedforward processing ): สมองส่วนซีรีเบลลัมแตกต่างจากส่วนอื่นๆ ของสมองส่วนใหญ่ (โดยเฉพาะสมองส่วนเปลือก) ตรงที่การประมวลผลสัญญาณเกือบทั้งหมดเป็นแบบส่งต่อนั่นคือ สัญญาณเคลื่อนที่ไปในทิศทางเดียวผ่านระบบจากอินพุตไปยังเอาต์พุต โดยมีการส่งผ่านภายในแบบย้อนกลับน้อยมาก การย้อนกลับเพียงเล็กน้อยที่มีอยู่ประกอบด้วยการยับยั้งซึ่งกันและกัน ไม่มีวงจรที่กระตุ้นซึ่งกันและกัน โหมดการทำงานแบบส่งต่อนี้หมายความว่าซีรีเบลลัม ซึ่งแตกต่างจากสมองส่วนเปลือก ไม่สามารถสร้างรูปแบบกิจกรรมประสาทที่ยั่งยืนได้ด้วยตนเอง สัญญาณเข้าสู่วงจร ถูกประมวลผลโดยแต่ละขั้นตอนตามลำดับ และจากนั้นก็ออกจากวงจร ตามที่ Eccles, Ito และ Szentágothai เขียนไว้ว่า "การกำจัดความเป็นไปได้ทั้งหมดของห่วงโซ่การสะท้อนกลับของการกระตุ้นเซลล์ประสาทในการออกแบบนี้ถือเป็นข้อได้เปรียบอย่างมากในการทำงานของสมองน้อยในฐานะคอมพิวเตอร์ เพราะสิ่งที่ระบบประสาทส่วนที่เหลือต้องการจากสมองน้อยนั้นไม่ใช่ผลลัพธ์ที่แสดงการทำงานของวงจรสะท้อนกลับที่ซับซ้อนในสมองน้อย แต่เป็นการตอบสนองที่รวดเร็วและชัดเจนต่อข้อมูลป้อนเข้าชุดใดชุดหนึ่งโดยเฉพาะ" [ 44 ]
- การแยกและการรวม : ในสมองน้อยของมนุษย์ ข้อมูลจากเส้นใย มอสซีไฟเบอร์ 200 ล้านเส้นจะถูกขยายไปยังเซลล์แกรนูล 40 พันล้านเซลล์ ซึ่งเส้นใยขนานที่ส่งออกมาจะรวมกันที่เซลล์พูร์คินเจ 15 ล้านเซลล์ [ 11 ]เนื่องจากลักษณะการเรียงตัวตามแนวยาว เซลล์พูร์คินเจประมาณ 1,000 เซลล์ที่อยู่ในไมโครโซนอาจได้รับข้อมูลจากเส้นใยขนานมากถึง 100 ล้านเส้น และโฟกัสการส่งออกของตนเองไปยังกลุ่มเซลล์นิวเคลียร์ที่อยู่ลึกลงไป อีกจำนวนหนึ่ง [ 28 ]ดังนั้น เครือข่ายสมองน้อยจึงได้รับข้อมูลเข้าจำนวนไม่มาก ประมวลผลอย่างละเอียดถี่ถ้วนผ่านเครือข่ายภายในที่มีโครงสร้างอย่างเข้มงวด และส่งผลลัพธ์ออกไปทางเซลล์ส่งออกจำนวนจำกัดมาก
- ความเป็นโมดูลาร์ : ระบบสมองน้อยถูกแบ่งออกเป็นโมดูลที่ทำงานแยกกันโดยประมาณ ซึ่งอาจมีจำนวนหลายร้อยถึงหลายพันโมดูล โมดูลทั้งหมดมีโครงสร้างภายในที่คล้ายคลึงกัน แต่มีอินพุตและเอาต์พุตที่แตกต่างกัน โมดูล (ไมโครคอมพาร์ทเมนต์แบบหลายโซนในศัพท์ของ Apps และ Garwicz) ประกอบด้วยกลุ่มเซลล์ประสาทขนาดเล็กในนิวเคลียสโอลิวารีส่วนล่าง ชุดของเซลล์ Purkinje ที่เป็นแถบยาวและแคบในเปลือกสมองน้อย (ไมโครโซน) และกลุ่มเซลล์ประสาทขนาดเล็กในนิวเคลียสสมองน้อยส่วนลึก โมดูลต่างๆ ใช้ร่วมกันอินพุตจากเส้นใย mossy และเส้นใย parallel แต่ในด้านอื่นๆ ดูเหมือนว่าโมดูลเหล่านี้จะทำงานแยกจากกัน กล่าวคือ เอาต์พุตของโมดูลหนึ่งดูเหมือนจะไม่ส่งผลต่อกิจกรรมของโมดูลอื่นๆ อย่างมีนัยสำคัญ[ 28 ]
- ความยืดหยุ่น : ไซแนปส์ระหว่างเส้นใยขนานและเซลล์ Purkinje และไซแนปส์ระหว่างเส้นใย mossy และเซลล์นิวเคลียร์ส่วนลึก ต่างก็ไวต่อการปรับเปลี่ยนความแข็งแรง ในโมดูลสมองน้อยเพียงโมดูลเดียว อินพุตจากเส้นใยขนานมากถึงหนึ่งพันล้านเส้นจะมาบรรจบกันที่กลุ่มเซลล์นิวเคลียร์ส่วนลึกที่มีจำนวนน้อยกว่า 50 เซลล์ และอิทธิพลของเส้นใยขนานแต่ละเส้นที่มีต่อเซลล์นิวเคลียร์เหล่านั้นสามารถปรับได้ การจัดเรียงนี้ให้ความยืดหยุ่นอย่างมากในการปรับแต่งความสัมพันธ์ระหว่างอินพุตและเอาต์พุตของสมองน้อย[ 45 ]
การเรียนรู้
มีหลักฐานมากมายที่แสดงว่าสมองน้อยมีบทบาทสำคัญในการเรียนรู้การเคลื่อนไหวบางประเภท งานที่สมองน้อยมีบทบาทชัดเจนที่สุดคืองานที่จำเป็นต้องปรับเปลี่ยนวิธีการกระทำอย่างละเอียด อย่างไรก็ตาม มีข้อโต้แย้งมากมายเกี่ยวกับว่าการเรียนรู้เกิดขึ้นภายในสมองน้อยเองหรือไม่ หรือว่ามันทำหน้าที่เพียงแค่ส่งสัญญาณที่ส่งเสริมการเรียนรู้ในโครงสร้างสมองอื่นๆ[ 45 ]ทฤษฎีส่วนใหญ่ที่กำหนดให้การเรียนรู้เกิดขึ้นกับวงจรของสมองน้อยนั้นมาจากแนวคิดของDavid Marr [ 27 ]และJames Albus [ 7 ] ซึ่งตั้งสมมติฐานว่าเส้นใยปีนป่ายให้สัญญาณการสอนที่กระตุ้นการปรับเปลี่ยนไซแนปส์ใน ไซแน ปส์ ของเส้นใย ขนานและเซลล์ Purkinje [ 46 ] Marr สันนิษฐานว่าอินพุตของเส้นใยปีนป่ายจะทำให้อินพุตของเส้นใยขนานที่เปิดใช้งานพร้อมกันมีความแข็งแกร่งขึ้น อย่างไรก็ตาม โมเดลการเรียนรู้ของสมองน้อยส่วนใหญ่ในเวลาต่อมาได้ปฏิบัติตามแนวคิดของ Albus โดยถือว่ากิจกรรมของเส้นใยปีนป่ายจะเป็นสัญญาณผิดพลาด และจะทำให้สัญญาณอินพุตของเส้นใยขนานที่เปิดใช้งานพร้อมกันอ่อนลง โมเดลในภายหลังบางรุ่น เช่น โมเดล ตัวกรองแบบปรับได้ของ Fujita [ 47 ]พยายามทำความเข้าใจการทำงานของสมองน้อยในแง่ของทฤษฎี การควบคุมที่เหมาะสมที่สุด
แนวคิดที่ว่ากิจกรรมของเส้นใยปีนป่ายทำหน้าที่เป็นสัญญาณข้อผิดพลาดได้รับการตรวจสอบในการศึกษาทดลองหลายครั้ง โดยบางการศึกษาสนับสนุนแนวคิดนี้ แต่บางการศึกษาก็ตั้งข้อสงสัย[ 24 ]ในการศึกษาบุกเบิกโดย Gilbert และ Thach ในปี 1977 เซลล์ Purkinje จากลิงที่กำลังเรียนรู้งานการเอื้อมมือแสดงให้เห็นกิจกรรมสไปค์ที่ซับซ้อนเพิ่มขึ้น ซึ่งเป็นที่ทราบกันดีว่าบ่งชี้ถึงกิจกรรมของอินพุตเส้นใยปีนป่ายของเซลล์ได้อย่างน่าเชื่อถือ ในช่วงเวลาที่ประสิทธิภาพการทำงานไม่ดี[ 48 ]การศึกษาการเรียนรู้การเคลื่อนไหวในแมวหลายครั้งพบกิจกรรมสไปค์ที่ซับซ้อนเมื่อมีความไม่ตรงกันระหว่างการเคลื่อนไหวที่ตั้งใจไว้กับการเคลื่อนไหวที่ดำเนินการจริง การศึกษาเกี่ยวกับรีเฟล็กซ์เวสติบูโล-โอคูลาร์ (ซึ่งทำให้ภาพบนเรตินาคงที่เมื่อศีรษะหัน) พบว่ากิจกรรมของเส้นใยปีนป่ายบ่งชี้ถึง "การเลื่อนของเรตินา" แม้ว่าจะไม่ใช่วิธีที่ตรงไปตรงมานัก[ 24 ]
หนึ่งในงานวิจัยเกี่ยวกับการเรียนรู้ของสมองส่วนซีรีเบลลัมที่ได้รับการศึกษาอย่างกว้างขวางที่สุดคือ แบบจำลอง การปรับเงื่อนไขการกระพริบตาซึ่งสิ่งเร้าที่ถูกปรับเงื่อนไข (CS) ที่เป็นกลาง เช่น เสียงหรือแสง จะถูกจับคู่ซ้ำๆ กับสิ่งเร้าที่ไม่ถูกปรับเงื่อนไข (US) เช่น ลมเป่า ที่กระตุ้นให้เกิดการกระพริบตา หลังจากนำเสนอ CS และ US ซ้ำๆ เช่นนี้ ในที่สุด CS จะกระตุ้นให้เกิดการกระพริบตาก่อน US ซึ่งเป็นการตอบสนองที่ถูกปรับเงื่อนไขหรือ CR การทดลองแสดงให้เห็นว่า การทำลายเนื้อเยื่อเฉพาะส่วนใดส่วนหนึ่งของนิวเคลียสที่อยู่ตรงกลาง (หนึ่งในนิวเคลียสของซีรีเบลลัมส่วนลึก) หรือเฉพาะบางจุดในเปลือกสมองส่วนซีรีเบลลัม จะทำให้การเรียนรู้การตอบสนองการกระพริบตาตามเวลาที่กำหนดนั้นหายไป หากเอาต์พุตของซีรีเบลลัมถูกปิดใช้งานทางเภสัชวิทยาในขณะที่อินพุตและวงจรภายในเซลล์ยังคงอยู่ การเรียนรู้จะเกิดขึ้นแม้ว่าสัตว์จะไม่แสดงการตอบสนองใด ๆ ในขณะที่หากวงจรภายในซีรีเบลลัมถูกรบกวน การเรียนรู้จะไม่เกิดขึ้น—ข้อเท็จจริงเหล่านี้รวมกันทำให้เกิดข้อโต้แย้งที่แข็งแกร่งว่าการเรียนรู้เกิดขึ้นภายในซีรีเบลลัมจริง ๆ[ 49 ]
ทฤษฎีและแบบจำลองเชิงคำนวณ

ฐานความรู้ขนาดใหญ่เกี่ยวกับโครงสร้างทางกายวิภาคและหน้าที่การทำงานของสมองส่วนซีรีเบลลัม ทำให้มันเป็นแหล่งกำเนิดทฤษฎีที่อุดมสมบูรณ์—อาจมีทฤษฎีเกี่ยวกับการทำงานของซีรีเบลลัมมากกว่าส่วนอื่นๆ ของสมอง ความแตกต่างพื้นฐานที่สุดระหว่างทฤษฎีเหล่านี้คือระหว่าง "ทฤษฎีการเรียนรู้" และ "ทฤษฎีประสิทธิภาพ"—นั่นคือ ทฤษฎีที่ใช้ความยืดหยุ่นของไซแนปส์ภายในซีรีเบลลัมเพื่ออธิบายบทบาทของมันในการเรียนรู้ เทียบกับทฤษฎีที่อธิบายแง่มุมของพฤติกรรมที่เกิดขึ้นอย่างต่อเนื่องบนพื้นฐานของการประมวลผลสัญญาณของซีรีเบลลัม ทฤษฎีทั้งสองประเภทหลายทฤษฎีได้รับการกำหนดเป็นแบบจำลองทางคณิตศาสตร์และจำลองโดยใช้คอมพิวเตอร์[ 46 ] มีการเสนอ แบบจำลองตัวกรอง Kalmanที่สอดคล้องกับข้อกำหนดหลักสองประการ: ซีรีเบลลัมมีส่วนเกี่ยวข้องในการคาดการณ์และในการจัดลำดับ[ 50 ]
บางทีทฤษฎี "ประสิทธิภาพ" ที่เก่าแก่ที่สุดอาจเป็นสมมติฐาน "สายหน่วงเวลา" ของValentino Braitenbergทฤษฎีดั้งเดิมที่ Braitenberg และ Roger Atwood เสนอในปี 1958 ระบุว่าการแพร่กระจายของสัญญาณอย่างช้าๆ ตามเส้นใยขนานทำให้เกิดความล่าช้าที่คาดการณ์ได้ ซึ่งช่วยให้สมองน้อยสามารถตรวจจับความสัมพันธ์ของเวลาภายในช่วงเวลาที่กำหนดได้[ 51 ]ข้อมูลจากการทดลองไม่สนับสนุนรูปแบบดั้งเดิมของทฤษฎี แต่ Braitenberg ยังคงโต้แย้งเพื่อเวอร์ชันที่ปรับปรุงแล้ว[ 52 ]สมมติฐานที่ว่าสมองน้อยทำงานโดยพื้นฐานแล้วเป็นระบบกำหนดเวลาได้รับการสนับสนุนโดยRichard Ivryเช่น กัน [ 53 ]ทฤษฎี "ประสิทธิภาพ" ที่มีอิทธิพลอีกทฤษฎีหนึ่งคือทฤษฎีเครือข่ายเทนเซอร์ของ Pellionisz และLlinásซึ่งให้สูตรทางคณิตศาสตร์ขั้นสูงของแนวคิดที่ว่าการคำนวณพื้นฐานที่ดำเนินการโดยสมองน้อยคือการแปลงพิกัดประสาทสัมผัสเป็นพิกัดการเคลื่อนไหว[ 54 ]
ทฤษฎีในหมวด "การเรียนรู้" เกือบทั้งหมดมาจากงานตีพิมพ์ของ Marr และ Albus บทความของ Marr ในปี 1969 เสนอว่าสมองน้อยเป็นอุปกรณ์สำหรับการเรียนรู้ที่จะเชื่อมโยงการเคลื่อนไหวพื้นฐานที่เข้ารหัสโดยเส้นใยปีนป่ายกับอินพุตของเส้นใยมอสซีที่เข้ารหัสบริบททางประสาทสัมผัส[ 27 ] Albus เสนอในปี 1971 ว่าเซลล์ Purkinje ในสมองน้อยทำหน้าที่เป็นเพอร์เซปตรอนซึ่งเป็นอุปกรณ์การเรียนรู้เชิงนามธรรมที่ได้รับแรงบันดาลใจจาก ระบบประสาท [ 7 ]ความแตกต่างพื้นฐานที่สุดระหว่างทฤษฎีของ Marr และ Albus คือ Marr สันนิษฐานว่ากิจกรรมของเส้นใยปีนป่ายจะทำให้ไซแนปส์ของเส้นใยขนานแข็งแรงขึ้น ในขณะที่ Albus เสนอว่ามันจะอ่อนแอลง Albus ยังได้กำหนดเวอร์ชันของเขาเป็นอัลกอริทึมซอฟต์แวร์ที่เขาเรียกว่า CMAC (Cerebellar Model Articulation Controller) ซึ่งได้รับการทดสอบในแอปพลิเคชันจำนวนมาก[ 55 ]
ความสำคัญทางคลินิก
ความเสียหายต่อสมองน้อยมักทำให้เกิดอาการที่เกี่ยวข้องกับการเคลื่อนไหว ซึ่งรายละเอียดจะขึ้นอยู่กับส่วนของสมองน้อยที่ได้รับผลกระทบและลักษณะความเสียหาย ความเสียหายต่อกลีบฟลอคคูโลโนดูลาร์อาจแสดงออกมาในรูปของการสูญเสียสมดุล โดยเฉพาะอย่างยิ่งการเดินที่เปลี่ยนแปลงไปและไม่สม่ำเสมอ มีท่าทางการยืนที่กว้างเนื่องจากความยากลำบากในการทรงตัว[ 14 ]ความเสียหายต่อโซนด้านข้างมักทำให้เกิดปัญหาในการเคลื่อนไหวโดยสมัครใจและวางแผน ซึ่งอาจทำให้เกิดข้อผิดพลาดในแรง ทิศทาง ความเร็ว และขนาดของการเคลื่อนไหว อาการอื่นๆ ได้แก่ ภาวะ กล้ามเนื้ออ่อนแรง ( hypotonia ) ภาวะพูดไม่ชัด ( dysarthria ) ภาวะการ กะระยะไม่แม่นยำ ( dysmetria ) ภาวะการเคลื่อนไหวสลับอย่างรวดเร็วไม่แม่นยำ ( dysdiadochokinesia ) (ไม่สามารถทำการเคลื่อนไหวสลับอย่างรวดเร็ว เช่น การเดิน) การตอบสนองแบบเช็ครีเฟล็กซ์บกพร่องหรือปรากฏการณ์รีบาวด์ และอาการสั่น แบบตั้งใจ ( intention tremor ) (การเคลื่อนไหวโดยไม่สมัครใจที่เกิดจากการหดตัวสลับกันของกลุ่มกล้ามเนื้อที่ตรงข้ามกัน) [ 56 ] [ 57 ]ความเสียหายต่อส่วนกลางอาจรบกวนการเคลื่อนไหวของร่างกายทั้งหมด ในขณะที่ความเสียหายที่จำกัดอยู่ด้านข้างมากกว่ามีแนวโน้มที่จะรบกวนการเคลื่อนไหวที่ละเอียดอ่อนของมือหรือแขนขา ความเสียหายต่อส่วนบนของสมองน้อยมีแนวโน้มที่จะทำให้เกิดความบกพร่องในการเดินและปัญหาอื่นๆ เกี่ยวกับการประสานงานของขา ความเสียหายต่อส่วนล่างมีแนวโน้มที่จะทำให้เกิดการเคลื่อนไหวของแขนและมือที่ไม่ประสานกันหรือไม่แม่นยำ รวมถึงความยากลำบากในความเร็ว[ 14 ]กลุ่มอาการทางมอเตอร์นี้เรียกว่าอะแท็กเซีย
เพื่อระบุปัญหาของสมองน้อยการตรวจทางระบบประสาทจะรวมถึงการประเมินการเดิน (การเดินแบบกว้างๆ บ่งชี้ถึงภาวะเสียการทรงตัว) การทดสอบการชี้นิ้ว และการประเมินท่าทาง[ 5 ]หากพบความผิดปกติของสมองน้อย สามารถใช้การสแกนภาพ ด้วยคลื่นแม่เหล็กไฟฟ้าเพื่อให้ได้ภาพโดยละเอียดของการเปลี่ยนแปลงโครงสร้างใดๆ ที่อาจเกิดขึ้นได้[ 58 ]
รายการปัญหาทางการแพทย์ที่อาจทำให้เกิดความเสียหายต่อสมองน้อยนั้นมีมากมาย รวมถึงโรคหลอดเลือดสมองเลือดออกในสมอง สมองบวม ( ภาวะสมองบวม ) เนื้องอก โรค พิษสุราเรื้อรังการบาดเจ็บทางกายภาพเช่น บาดแผลจากกระสุนปืนหรือวัตถุระเบิด และ ภาวะ เสื่อม เรื้อรัง เช่นภาวะสมองน้อยฝ่อ [ 59 ] [ 60 ] อาการ ปวดศีรษะ ไมเกรนบางรูปแบบอาจทำให้เกิดความผิดปกติชั่วคราวของสมองน้อย ซึ่งมีความรุนแรงแตกต่างกันไป[ 61 ]การติดเชื้ออาจทำให้เกิดความเสียหายต่อสมองน้อยในภาวะต่างๆ เช่นโรคพรีออน[ 62 ]และกลุ่มอาการมิลเลอร์ ฟิชเชอร์ซึ่ง เป็นรูปแบบหนึ่งของกลุ่มอาการกิลเลน-บาร์เร
ความชรา
สมองส่วนซีรีเบลลัมของมนุษย์เปลี่ยนแปลงไปตามอายุ การเปลี่ยนแปลงเหล่านี้อาจแตกต่างจากส่วนอื่นๆ ของสมอง ซีรีเบลลัมเป็นบริเวณสมอง (และส่วนของร่างกาย) ที่อายุน้อยที่สุดตามตัวบ่งชี้ทางชีวภาพของเนื้อเยื่อที่เรียกว่านาฬิกาเอพิเจเนติกส์โดยมีอายุน้อยกว่าที่คาดไว้ในผู้ที่มีอายุ 100 ปีประมาณ 15 ปี[ 63 ]นอกจากนี้รูปแบบการแสดงออกของยีน ในซีรีเบลลัมของมนุษย์แสดงให้เห็น การเปลี่ยนแปลงที่เกี่ยวข้องกับอายุ น้อยกว่าใน สมองส่วนคอร์เทกซ์ [ 64 ] บาง การศึกษาได้รายงานการลดลงของจำนวนเซลล์หรือปริมาตรของเนื้อเยื่อ แต่ข้อมูลที่เกี่ยวข้องกับคำถามนี้มีไม่มากนัก[ 65 ] [ 66 ]
ความผิดปกติทางพัฒนาการและการเสื่อมถอย

ความผิดปกติแต่กำเนิด โรคทางพันธุกรรม และภาวะที่เกิดขึ้นภายหลัง อาจส่งผลต่อโครงสร้างของสมองน้อย และส่งผลต่อการทำงานของสมองน้อยด้วย เว้นแต่ว่าภาวะที่เป็นสาเหตุจะสามารถแก้ไขได้ การรักษาที่เป็นไปได้เพียงอย่างเดียวคือการช่วยให้ผู้ป่วยใช้ชีวิตอยู่กับปัญหาของตนเอง[ 67 ]การมองเห็นสมองน้อยของทารกในครรภ์ด้วยการสแกนอัลตราซาวนด์ในช่วง 18 ถึง 20 สัปดาห์ของการตั้งครรภ์ สามารถใช้ในการคัดกรองความผิดปกติของท่อประสาทของทารกใน ครรภ์ได้ โดยมี อัตรา ความไวสูงถึง 99% [ 68 ]
ในการพัฒนาตามปกติการส่งสัญญาณโซนิคเฮดจ์ฮ็อก ภายในร่างกาย จะกระตุ้นการแพร่กระจายอย่างรวดเร็วของเซลล์ต้นกำเนิดเซลล์ประสาทแกรนูลของสมองน้อย (CGNPs) ในชั้นแกรนูลภายนอก (EGL) การพัฒนาของสมองน้อยเกิดขึ้นในช่วงปลายของการเกิดตัวอ่อนและช่วงต้นหลังคลอด โดยการแพร่กระจายของ CGNP ใน EGL จะสูงสุดในช่วงต้นของการพัฒนา (วันที่ 7 หลังคลอดในหนู) [ 69 ]เมื่อ CGNPs แตกต่างไปเป็นเซลล์แกรนูลของสมองน้อย (เรียกอีกอย่างว่าเซลล์ประสาทแกรนูลของสมองน้อย, CGNs) พวกมันจะอพยพไปยังชั้นแกรนูลภายใน (IGL) ก่อตัวเป็นสมองน้อยที่เจริญเต็มที่ (ภายในวันที่ 20 หลังคลอดในหนู) [ 69 ]การกลายพันธุ์ที่กระตุ้นการส่งสัญญาณโซนิคเฮดจ์ฮ็อกอย่างผิดปกติทำให้เกิดความเสี่ยงต่อมะเร็งของสมองน้อย ( เม ดุลโลบลาสโตมา ) ในมนุษย์ที่เป็นโรคกอร์ลินและในแบบจำลองหนู ที่ได้รับการดัดแปลงพันธุกรรม [ 70 ] [ 71 ]
ความผิดปกติแต่กำเนิดหรือการพัฒนาไม่เต็มที่ ( ภาวะ hypoplasia ) ของเวอร์มิสของสมองน้อยเป็นลักษณะเฉพาะของทั้งกลุ่มอาการDandy–Walkerและกลุ่มอาการ Joubert [ 72 ] [ 73 ]ในกรณีที่หายากมาก สมองน้อยทั้งหมดอาจหายไป [ 74 ] โรคทางระบบประสาทที่ถ่ายทอดทางพันธุกรรม เช่นโรค Machado–Joseph , ataxia telangiectasiaและFriedreich's ataxiaทำให้เกิดการเสื่อมของ ระบบประสาทอย่างต่อเนื่อง ที่เชื่อมโยงกับการสูญเสียสมองน้อย[ 59 ] [ 67 ]ความผิดปกติของสมองแต่กำเนิดที่อยู่นอกสมองน้อยสามารถทำให้เกิดการเคลื่อนตัวของเนื้อเยื่อสมองน้อย ได้ ดังที่เห็นในความผิดปกติของ Arnold–Chiariบาง รูปแบบ [ 75 ]
ภาวะอื่นๆ ที่เชื่อมโยงอย่างใกล้ชิดกับการเสื่อมของสมอง น้อยได้แก่ โรคระบบประสาทเสื่อมแบบก้าวหน้าที่ไม่ทราบสาเหตุ เช่นโรคฝ่อของระบบหลายระบบและกลุ่มอาการแรมเซย์ ฮันท์ ประเภทที่ 1 [ 76 ] [ 77 ]และการเสื่อมของสมองน้อยจาก เนื้องอก ซึ่ง เป็นโรคภูมิต้านตนเองโดยเนื้องอกในส่วนอื่นๆ ของร่างกายจะกระตุ้นให้เกิดการตอบสนองของระบบภูมิคุ้มกันที่ทำให้เกิดการสูญเสียเซลล์ประสาทในสมองน้อย[ 78 ]การฝ่อของสมองน้อยอาจเกิดจากการขาดไทอามีน (วิตามินบี 1) เรื้อรัง ดังที่พบในโรคเหน็บชา แห้ง และใน กลุ่มอาการเวิร์นิ ก -คอร์ซาคอฟ [ 79 ]หรือจากการขาดวิตามินอี[ 67 ]
ภาวะสมองน้อยฝ่อพบได้ในความผิดปกติทางระบบประสาทอื่นๆ หลายอย่าง รวมถึงโรคฮันติงตัน โรคปลอกประสาทเสื่อมแข็ง[ 62 ]อาการสั่นที่ เกิดจากสาเหตุที่ไม่ทราบแน่ชัด โรคลมชักแบบไมโอโคลนั สที่ลุกลามและโรคนีแมนน์-พิก ภาวะสมองน้อยฝ่ออาจเกิดขึ้นได้จากการสัมผัสกับสารพิษ รวมถึงโลหะหนักหรือยาหรือสารเสพติด[ 67 ]
ความเจ็บปวด
โดยทั่วไปมีความเห็นพ้องกันว่าสมองน้อยมีส่วนเกี่ยวข้องกับการประมวลผลความเจ็บปวด[ 80 ] [ 81 ]สมองน้อยได้รับข้อมูลความเจ็บปวดจากทั้งเส้นทางคอร์ติโค-ซีรีเบลลาร์ที่ลงมา และเส้นทางสไปโน-ซีรีเบลลาร์ที่ขึ้นไป ผ่านนิวเคลียสพอนไทน์และโอลิฟอินเฟอเรียร์ ข้อมูลบางส่วนนี้จะถูกถ่ายโอนไปยังระบบการเคลื่อนไหว ทำให้เกิดการหลีกเลี่ยงความเจ็บปวดโดยการเคลื่อนไหวอย่างมีสติ โดยแบ่งระดับตามความรุนแรงของความเจ็บปวด
เชื่อกันว่าสัญญาณความเจ็บปวดโดยตรงเหล่านี้ รวมถึงสัญญาณทางอ้อม จะกระตุ้นพฤติกรรมการหลีกเลี่ยงความเจ็บปวดในระยะยาว ซึ่งส่งผลให้เกิดการเปลี่ยนแปลงท่าทางเรื้อรัง และส่งผลให้เกิดการปรับโครงสร้างการทำงานและกายวิภาคของนิวเคลียสเวสติบูลาร์และโพรปริโอเซปทีฟ ส่งผลให้ความเจ็บปวดจากระบบประสาทเรื้อรังสามารถกระตุ้นการปรับโครงสร้างทางกายวิภาคในระดับมหภาคของสมองส่วนท้าย รวมถึงซีรีเบลลัม[ 26 ]ขนาดของการปรับโครงสร้างนี้และการเหนี่ยวนำเครื่องหมายของเซลล์ต้นกำเนิดประสาทบ่งชี้ถึงการมีส่วนร่วมของการสร้างเซลล์ประสาทในผู้ใหญ่ต่อการเปลี่ยนแปลงเหล่านี้
กายวิภาคเปรียบเทียบและวิวัฒนาการ

วงจรในซีรีเบลลัมมีความคล้ายคลึงกันในสัตว์มีกระดูกสันหลังทุกชั้นรวมถึงปลา สัตว์เลื้อยคลาน นก และสัตว์เลี้ยงลูกด้วยนม[ 82 ]นอกจากนี้ยังมีโครงสร้างสมองที่คล้ายคลึงกันในเซฟาโลพอด ที่มี สมองที่พัฒนาแล้ว เช่นปลาหมึกยักษ์ [ 83 ] สิ่งนี้ถือเป็นหลักฐานว่าซีรีเบลลัมทำหน้าที่สำคัญต่อสัตว์ทุกชนิดที่มีสมอง
ขนาดและรูปร่างของซีรีเบลลัมมีความแตกต่างกันอย่างมากในสัตว์มีกระดูกสันหลังชนิดต่างๆ ในสัตว์สะเทินน้ำสะเทินบก ซีรีเบลลัม มีการพัฒนาเพียงเล็กน้อย และในปลาแลมเพรย์และปลาแฮกฟิชซีรีเบลลัมแทบจะแยกไม่ออกจากก้านสมอง แม้ว่าจะมีสปิโนซีรีเบลลัมอยู่ในกลุ่มเหล่านี้ แต่โครงสร้างหลักมีขนาดเล็ก เป็นนิวเคลียสคู่ที่สอดคล้องกับเวสติบูโลซีรีเบลลัม[ 84 ]ซีรีเบลลัมมีขนาดใหญ่ขึ้นเล็กน้อยในสัตว์เลื้อยคลาน ใหญ่กว่ามากในนก และใหญ่กว่ามากในสัตว์เลี้ยงลูกด้วยนม กลีบคู่ขนาดใหญ่และคดเคี้ยวที่พบในมนุษย์เป็นลักษณะทั่วไปของสัตว์เลี้ยงลูกด้วยนม แต่โดยทั่วไปแล้วซีรีเบลลัมในกลุ่มอื่นๆ จะเป็นกลีบตรงกลางเพียงกลีบเดียว และมีผิวเรียบหรือมีร่องเล็กน้อยเท่านั้น ในสัตว์เลี้ยงลูกด้วยนม นีโอซีรีเบลลัมเป็นส่วนใหญ่ของซีรีเบลลัมเมื่อพิจารณาจากมวล แต่ในสัตว์มีกระดูกสันหลังอื่นๆ มักจะเป็นสปิโนซีรีเบลลัม[ 84 ]
สมองน้อยของ ปลา กระดูกอ่อนและปลากระดูกแข็งนั้นมีขนาดใหญ่และซับซ้อนเป็นพิเศษ อย่างน้อยในประเด็นสำคัญประการหนึ่ง โครงสร้างภายในของมันแตกต่างจากสมองน้อยของสัตว์เลี้ยงลูกด้วยนม: สมองน้อยของปลาไม่มีนิวเคลียสสมองน้อยที่ แยก จากกันอย่างลึกซึ้ง แต่เป้าหมายหลักของเซลล์ Purkinje คือเซลล์ชนิดหนึ่งที่แตกต่างกันซึ่งกระจายอยู่ทั่วเปลือกสมองน้อย ซึ่งเป็นเซลล์ชนิดที่ไม่พบในสัตว์เลี้ยงลูกด้วยนม ในปลาวงศ์ Mormyridae (วงศ์ปลาน้ำจืดที่มีความไวต่อไฟฟ้าต่ำ) สมองน้อยมีขนาดใหญ่กว่าส่วนอื่นๆ ของสมองอย่างมาก ส่วนที่ใหญ่ที่สุดของมันคือโครงสร้างพิเศษที่เรียกว่าvalvulaซึ่งมีสถาปัตยกรรมที่เป็นระเบียบอย่างผิดปกติและได้รับข้อมูลส่วนใหญ่จากระบบรับรู้ไฟฟ้า[ 85 ]
ลักษณะเด่นของสมองน้อยในสัตว์เลี้ยงลูกด้วยนมคือการขยายตัวของกลีบด้านข้าง ซึ่งมีปฏิสัมพันธ์หลักกับนีโอคอร์เทกซ์ เมื่อลิงวิวัฒนาการไปเป็นลิงใหญ่ การขยายตัวของกลีบด้านข้างยังคงดำเนินต่อไป ควบคู่ไปกับการขยายตัวของกลีบด้านหน้าของนีโอคอร์เทกซ์ ในบรรพบุรุษของโฮมินิด และในโฮโมเซเปียนส์จนถึงช่วงกลาง ยุค ไพลสโตซีนสมองน้อยยังคงขยายตัวต่อไป แต่กลีบด้านหน้าขยายตัวได้เร็วกว่า อย่างไรก็ตาม ช่วงเวลาล่าสุดของการวิวัฒนาการของมนุษย์อาจเกี่ยวข้องกับการเพิ่มขึ้นของขนาดสัมพัทธ์ของสมองน้อย เนื่องจากนีโอคอร์เทกซ์ลดขนาดลงบ้างในขณะที่สมองน้อยขยายตัว[ 86 ]ขนาดของสมองน้อยของมนุษย์เมื่อเทียบกับส่วนอื่นๆ ของสมอง มีขนาดใหญ่ขึ้นในขณะที่สมองใหญ่มีขนาดลดลง[ 87 ]เนื่องจากการพัฒนาและการดำเนินการของงานด้านการเคลื่อนไหว ทักษะการมองเห็นเชิงพื้นที่ และการเรียนรู้เกิดขึ้นในซีรีเบลลัม จึงเชื่อว่าการเติบโตของซีรีเบลลัมมีความสัมพันธ์บางอย่างกับความสามารถทางปัญญาของมนุษย์ที่สูงขึ้น[ 88 ]ปัจจุบันซีกสมองด้านข้างของซีรีเบลลัมมีขนาดใหญ่กว่าในมนุษย์และลิงถึง 2.7 เท่า เมื่อเทียบกับลิง[ 87 ]การเปลี่ยนแปลงขนาดของซีรีเบลลัมเหล่านี้ไม่สามารถอธิบายได้ด้วยมวลกล้ามเนื้อที่มากขึ้น แสดงให้เห็นว่าการพัฒนาของซีรีเบลลัมมีความเชื่อมโยงอย่างแน่นหนากับการพัฒนาของสมองส่วนอื่นๆ หรือกิจกรรมทางประสาทที่เกิดขึ้นในซีรีเบลลัมมีความสำคัญในระหว่าง วิวัฒนาการ ของโฮมินิดีเนื่องจากบทบาทของซีรีเบลลัมในหน้าที่ทางปัญญา การเพิ่มขนาดของมันอาจมีบทบาทในการขยายขีดความสามารถทางปัญญา[ 87 ]
โครงสร้างคล้ายสมองน้อย
สัตว์มีกระดูกสันหลังส่วนใหญ่มีสมองส่วนซีรีเบลลัมและโครงสร้างคล้ายซีรีเบลลัมอย่างน้อยหนึ่งแห่ง ซึ่งเป็นบริเวณสมองที่มีลักษณะคล้ายซีรีเบลลัมในแง่ของโครงสร้างเซลล์และเคมีประสาท [ 82 ]โครงสร้างคล้ายซีรีเบลลัมเพียงอย่างเดียวที่พบในสัตว์เลี้ยงลูกด้วยนมคือนิวเคลียสดอร์ซัลโคเคลียร์ (DCN) ซึ่งเป็นหนึ่งในสองนิวเคลียสรับความรู้สึกหลักที่ได้รับข้อมูลโดยตรงจากเส้นประสาทการได้ยิน DCN เป็นโครงสร้างแบบหลายชั้น โดยชั้นล่างสุดมีเซลล์แกรนูลคล้ายกับเซลล์ของซีรีเบลลัม ทำให้เกิดเส้นใยขนานที่ขึ้นไปยังชั้นผิวและเคลื่อนที่ไปตามแนวนอน ชั้นผิวประกอบด้วยเซลล์ประสาท GABAergic ชุดหนึ่งที่เรียกว่าเซลล์คาร์ทวีลซึ่งมีลักษณะคล้ายเซลล์ Purkinje ทั้งทางกายวิภาคและทางเคมี พวกมันได้รับข้อมูลจากเส้นใยขนาน แต่ไม่มีข้อมูลใดๆ ที่คล้ายกับเส้นใยปีนป่ายเซลล์ประสาทส่งออกของ DCN คือเซลล์พีระมิด เซลล์เหล่านี้เป็นเซลล์กลูตาเมต แต่ก็มีลักษณะคล้ายเซลล์ Purkinje ในบางแง่มุม กล่าวคือ มีเดนไดรต์ผิวเผินที่มีลักษณะเป็นหนามและแบนราบซึ่งรับอินพุตจากเส้นใยขนาน แต่ยังมีเดนไดรต์ฐานที่รับอินพุตจากเส้นใยประสาทการได้ยิน ซึ่งเดินทางผ่าน DCN ในทิศทางตั้งฉากกับเส้นใยขนาน DCN มีการพัฒนาสูงสุดในสัตว์ฟันแทะและสัตว์ขนาดเล็กอื่นๆ และลดลงอย่างมากในไพรเมต หน้าที่ของมันยังไม่เป็นที่เข้าใจดีนัก การคาดเดาที่เป็นที่นิยมมากที่สุดเกี่ยวข้องกับระบบการได้ยินเชิงพื้นที่ในรูปแบบใดรูปแบบหนึ่ง[ 89 ]
ปลาและสัตว์สะเทินน้ำสะเทินบกส่วนใหญ่มี ระบบ เส้นข้างลำตัวที่รับรู้คลื่นความดันในน้ำ บริเวณสมองส่วนหนึ่งที่รับข้อมูลหลักจากอวัยวะเส้นข้างลำตัว คือ นิวเคลียสมีเดียลอ็อกตาโวลาเทอรัล ซึ่งมีโครงสร้างคล้ายสมองน้อย โดยมีเซลล์เม็ดเล็กและเส้นใยขนาน ในปลาที่ไวต่อกระแสไฟฟ้า ข้อมูลจากระบบรับรู้กระแสไฟฟ้าจะส่งไปยังนิวเคลียสดอร์ซัลอ็อกตาโวลาเทอรัล ซึ่งมีโครงสร้างคล้ายสมองน้อยเช่นกัน ในปลาครีบแข็ง (ซึ่งเป็นกลุ่มที่ใหญ่ที่สุด) เทคตัมรับภาพมีชั้นหนึ่ง คือ ชั้นขอบ ซึ่งมีโครงสร้างคล้ายสมองน้อย[ 82 ]
โครงสร้างคล้ายสมองน้อยเหล่านี้ทั้งหมดดูเหมือนจะเกี่ยวข้องกับการรับรู้ทางประสาทสัมผัสมากกว่าการเคลื่อนไหว ทั้งหมดมีเซลล์เม็ดเล็กที่สร้างเส้นใยขนานที่เชื่อมต่อกับเซลล์ประสาทคล้าย Purkinje ด้วยไซแนปส์ที่ปรับเปลี่ยนได้แต่ไม่มีเส้นใยปีนป่ายที่เทียบได้กับของสมองน้อย—แต่พวกมันรับอินพุตโดยตรงจากอวัยวะรับความรู้สึกส่วนปลาย ไม่มีโครงสร้างใดที่มีหน้าที่ที่ได้รับการพิสูจน์แล้ว แต่การคาดการณ์ที่มีอิทธิพลมากที่สุดคือพวกมันทำหน้าที่แปลงอินพุตทางประสาทสัมผัสในรูปแบบที่ซับซ้อนบางอย่าง บางทีเพื่อชดเชยการเปลี่ยนแปลงท่าทางของร่างกาย[ 82 ]ในความเป็นจริงJames M. Bowerและคนอื่นๆ ได้โต้แย้ง โดยส่วนหนึ่งจากโครงสร้างเหล่านี้และส่วนหนึ่งจากการศึกษาเกี่ยวกับสมองน้อย ว่าสมองน้อยเองเป็นโครงสร้างรับความรู้สึกโดยพื้นฐาน และมีส่วนช่วยในการควบคุมการเคลื่อนไหวโดยการเคลื่อนไหวของร่างกายในลักษณะที่ควบคุมสัญญาณรับความรู้สึกที่เกิดขึ้น[ 90 ]แม้ว่าโบเวอร์จะมีมุมมองเช่นนั้น แต่ก็มีหลักฐานที่แน่ชัดว่าสมองน้อยมีอิทธิพลโดยตรงต่อการทำงานของมอเตอร์ในสัตว์เลี้ยงลูกด้วยนม[ 91 ] [ 92 ]
ประวัติศาสตร์

คำอธิบาย
แม้แต่นักกายวิภาคศาสตร์ยุคแรกๆ ก็สามารถจำแนกซีรีเบลลัมได้จากลักษณะที่โดดเด่นอริสโตเติลและเฮโรฟิลัส (อ้างอิงในกาเลน ) เรียกมันว่า παρεγκεφαλίς ( parenkephalis ) ซึ่งตรงข้ามกับ ἐγκέφαλος ( enkephalos ) หรือสมองส่วนหลัก คำอธิบายอย่างละเอียดของกาเลนเป็นคำอธิบายที่เก่าแก่ที่สุดที่ยังหลงเหลืออยู่ เขาตั้งข้อสันนิษฐานว่าซีรีเบลลัมเป็นแหล่งกำเนิดของเส้นประสาทสั่งการ[ 93 ]
การพัฒนาที่สำคัญเพิ่มเติมเกิดขึ้นใน ยุคเร เนสซองส์ เวซาลิอุสได้กล่าวถึงซีรีเบลลัมโดยสังเขป และโทมัส วิลลิส ได้อธิบายกายวิภาคศาสตร์อย่างละเอียดมากขึ้น ในปี 1664 มีการศึกษากายวิภาคศาสตร์เพิ่มเติมในช่วงศตวรรษที่ 18 แต่จนกระทั่งต้นศตวรรษที่ 19 จึงได้มีการค้นพบข้อมูลเชิงลึกเกี่ยวกับหน้าที่ของซีรีเบลลัมเป็นครั้งแรกลุยจิ โรลันโดในปี 1809 ได้ค้นพบข้อสำคัญที่ว่าความเสียหายต่อซีรีเบลลัมส่งผลให้เกิดความผิดปกติ ของการเคลื่อนไหว ฌอง ปิแอร์ ฟลอเรนส์ในช่วงครึ่งแรกของศตวรรษที่ 19 ได้ทำการทดลองอย่างละเอียด ซึ่งเผยให้เห็นว่าสัตว์ที่มีความเสียหายต่อซีรีเบลลัมยังคงสามารถเคลื่อนไหวได้ แต่สูญเสียการประสานงาน (การเคลื่อนไหวที่แปลกประหลาด การเดินที่งุ่มง่าม และกล้ามเนื้ออ่อนแรง) และการฟื้นตัวหลังจากการบาดเจ็บสามารถเกือบสมบูรณ์ได้เว้นแต่ว่าการบาดเจ็บจะกว้างขวางมาก[ 94 ]เมื่อถึงต้นศตวรรษที่ 20 เป็นที่ยอมรับกันอย่างกว้างขวางว่าหน้าที่หลักของซีรีเบลลัมเกี่ยวข้องกับการควบคุมการเคลื่อนไหว ช่วงครึ่งแรกของศตวรรษที่ 20 มีคำอธิบายโดยละเอียดเกี่ยวกับอาการทางคลินิกที่เกี่ยวข้องกับโรคของสมองน้อยในมนุษย์[ 5 ]
นิรุกติศาสตร์
ชื่อซีรีเบลลัมเป็นคำย่อของซีรีบรัม (สมอง) [ 95 ]สามารถแปลตรงตัวได้ว่าสมองเล็กชื่อภาษาละตินเป็นการแปลโดยตรงจากภาษากรีกโบราณ παρεγκεφαλίς ( parenkephalis ) ซึ่งใช้ในงานเขียนของอริสโตเติล นักเขียนคนแรกที่ทราบกันว่าอธิบายโครงสร้างนี้[ 96 ] ไม่มีการใช้ชื่ออื่นในวรรณกรรมภาษาอังกฤษ แต่ ใน ทางประวัติศาสตร์มีการใช้ ชื่อ ที่มาจาก ภาษากรีกหรือละตินหลากหลายชื่อ รวมถึงcerebrum parvum [ 97 ] encephalion [ 98 ] encranion [ 97 ] cerebrum posterius [ 99 ]และparencephalis [ 97 ]
ลิงก์ภายนอก
 สื่อที่เกี่ยวข้องกับสมองส่วนซีรีเบลลัมในวิกิมีเดียคอมมอนส์
สื่อที่เกี่ยวข้องกับสมองส่วนซีรีเบลลัมในวิกิมีเดียคอมมอนส์- Llinas R, Negrello MN (2015). "สมองน้อย" . Scholarpedia . 10 (1): 4606. Bibcode : 2015SchpJ..10.4606L . doi : 10.4249/scholarpedia.4606 .
- ฐานข้อมูลที่เน้นเซลล์ในสมองส่วนซีรีเบลลัม
- คู่มือเกี่ยวกับสมองส่วนซีรีเบลลัมและความผิดปกติของสมองส่วนซีรีเบลลัม – Manto, M., Gruol, DL., Schmahmann, J., Koibuchi, N., Rossi, F. (บรรณาธิการ) – Springer – นิวยอร์ก
- ภาพตัดขวางสมองที่ย้อมสี ซึ่งรวมถึง "สมองส่วนซีรีเบลลัม"ในโครงการ BrainMaps
- สมองที่ไม่สมบูรณ์ของชายคนหนึ่งเผยให้เห็นบทบาทของสม cerebellum ในด้านความคิดและอารมณ์
- ผู้หญิงที่ใช้ชีวิตโดยปราศจากสมองส่วนซีรีเบลลัม
- สาระสำคัญของสมองส่วนซีรีเบลลัมและความผิดปกติของสมองส่วนซีรีเบลลัม Gruol, DL, Koibuchi, N., Manto, M., Molinari, M., Schmahmann, JD, Shen, Y. (บรรณาธิการ). Springer, นิวยอร์ก, 2016
- ภาพเนื้อเยื่อวิทยาของสม cerebellum
- สมองส่วนซีรีเบลลัม – วารสาร (Springer Nature)
- สมองส่วนซีรีเบลลัมและโรคอะแท็กเซีย – วารสาร (BioMed Central)
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ สมองน้อย
สมองน้อย ( พหูพจน์ : cerebellaหรือcerebellums ; ภาษาละตินแปลว่า 'สมองเล็ก') เป็นส่วนประกอบหลักของสมองส่วนท้ายของสัตว์มีกระดูกสันหลัง ทุกชนิด...
โครงสร้าง
ในระดับ กายวิภาคศาสตร์โดยรวม สมองน้อยประกอบด้วยชั้น คอร์เทกซ์ ที่พับซ้อนกันอย่างแน่นหนา โดยมี เนื้อขาว อยู่ด้านล่างและ โพรง ที่เต็มไปด้วยของเหลวที่ฐาน นิวเคลียสของสมองน้อยสี่อันที่อยู่ลึก ฝังอยู่ในเนื้อขาว [ 9 ]...
กายวิภาคศาสตร์ระดับมหภาค
สมองน้อยตั้งอยู่ใน โพรงกะโหลกส่วนหลัง โพรงสมองที่สี่ พอน ส์ และ เมดุลลา อยู่ด้านหน้าของสมองน้อย [ 10 ] มันถูกแยกออกจาก สมอง ใหญ่ โดยเยื่อ ดูรามาเตอร์ที่ มีลักษณะคล้ายหนัง เรียกว่า cerebellar tentorium การเชื่อมต่อทั้งหมดกับส่วนอื่นๆ ของสมองจะผ่านทางพอนส์...
จุลกายวิภาคศาสตร์
เซลล์ประสาทสองประเภทมีบทบาทสำคัญในวงจรซีรีเบลลัม ได้แก่ เซลล์ Purkinje และ เซลล์แกรนูล นอกจากนี้ แอกซอน สามประเภทก็มีบทบาทสำคัญเช่นกัน ได้แก่ เส้นใยมอสซี และ เส้นใยปีนป่าย (ซึ่งเข้าสู่ซีรีเบลลัมจากภายนอก) และ เส้นใยขนาน (ซึ่งเป็นแอกซอนของเซลล์แกรนูล)...