อ่าน 20 นาที
ดีเอ็นเอพอลิเมอเรส
ดีเอ็นเอพอลิเมอเรสเป็นสมาชิกของกลุ่มเอนไซม์ที่เร่งปฏิกิริยาการสังเคราะห์โมเลกุลดีเอ็นเอ จากนิวคลีโอไซด์ไตรฟอสเฟตซึ่งเป็นสารตั้งต้นโมเลกุลของดีเอ็นเอ
ดีเอ็นเอพอลิเมอเรส
| ดีเอ็นเอพอลิเมอเรสที่ควบคุมโดยดีเอ็นเอ | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 โครงสร้างสามมิติของ โมทีฟ เฮลิกซ์-เทิร์น-เฮลิกซ์ ที่จับกับ DNA ในเอนไซม์ DNA โพลีเมอเรสเบตาของมนุษย์ (อ้างอิงจากไฟล์ PDB 7ICG ) | |||||||||
| ตัวระบุ | |||||||||
| หมายเลข EC | 2.7.7.7 | ||||||||
| หมายเลข CAS | 9012-90-2 | ||||||||
| ฐานข้อมูล | |||||||||
| อินท์เอ็นซ์ | มุมมองของ IntEnz | ||||||||
| เบรนด้า | เบรนด้าเข้าร่วม | ||||||||
| เอ็กซ์แพซี่ | มุมมองของ NiceZyme | ||||||||
| เคกก์ | รายการ KEGG | ||||||||
| เมตาไซค์ | วิถีการเผาผลาญ | ||||||||
| ไพรแอม | ประวัติโดยย่อ | ||||||||
| โครงสร้างPDB | RCSB PDB PDBe PDBsum | ||||||||
| ออนโทโลยีของยีน | อามิโก้ / ควิกโก้ | ||||||||
| |||||||||
ดีเอ็นเอพอลิเมอเรสเป็นสมาชิกของกลุ่มเอนไซม์ที่เร่งปฏิกิริยาการสังเคราะห์โมเลกุลดีเอ็นเอ จากนิวคลีโอไซด์ไตรฟอสเฟตซึ่งเป็นสารตั้งต้นโมเลกุลของดีเอ็นเอ เอนไซม์เหล่านี้มีความสำคัญต่อการจำลองดีเอ็นเอและมักทำงานเป็นกลุ่มเพื่อสร้างดีเอ็นเอคู่สองสายที่เหมือนกันจากดีเอ็นเอคู่สายเดิมเพียงสายเดียว ในระหว่างกระบวนการนี้ ดีเอ็นเอพอลิเมอเรสจะ "อ่าน" สายดีเอ็นเอที่มีอยู่เพื่อสร้างสายใหม่สองสายที่ตรงกับสายที่มีอยู่[ 1 ] [ 2 ] [ 3 ] [ 4 ] [ 5 ] [ 6 ] เอนไซม์เหล่านี้เร่งปฏิกิริยาเคมี
- ดีออกซีนิวคลีโอไซด์ไตรฟอสเฟต + DNA n ⇌ ไพโรฟอสเฟต + DNA n+ 1
เอนไซม์ดีเอ็นเอพอลิเมอเรสจะเพิ่มนิวคลีโอไทด์เข้าไปที่ปลาย 3' (3')ของสายดีเอ็นเอทีละนิวคลีโอไทด์ ทุกครั้งที่เซลล์แบ่งตัวจำเป็นต้องใช้เอนไซม์ดีเอ็นเอพอลิเมอเรสในการจำลองดีเอ็นเอของเซลล์ เพื่อให้สำเนาของโมเลกุลดีเอ็นเอเดิมสามารถส่งต่อไปยังเซลล์ลูกแต่ละเซลล์ได้ ด้วยวิธีนี้ ข้อมูลทางพันธุกรรมจึงถูกส่งต่อจากรุ่นสู่รุ่น
ก่อนที่กระบวนการจำลองแบบจะเกิดขึ้นได้ เอนไซม์ที่เรียกว่าเฮลิเคสจะคลายเกลียวโมเลกุล DNA จากโครงสร้างที่พันกันแน่น โดยในกระบวนการนี้จะทำลายพันธะไฮโดรเจนระหว่างเบสของนิวคลีโอไทด์ซึ่งจะทำให้ DNA สองสายคลายตัวหรือ "แยกออก" กลายเป็น DNA สองสายเดี่ยวที่สามารถใช้เป็นแม่แบบสำหรับการจำลองแบบในปฏิกิริยาข้างต้นได้
ประวัติศาสตร์
ในปี พ.ศ. 2499 อาร์เธอร์ คอร์นเบิร์กและเพื่อนร่วมงานได้ค้นพบดีเอ็นเอพอลิเมอเรส I (Pol I) ในแบคทีเรีย Escherichia coliพวกเขาได้อธิบายกระบวนการจำลองดีเอ็นเอโดยที่ดีเอ็นเอพอลิเมอเรสจะคัดลอกลำดับเบสของสายดีเอ็นเอแม่แบบ คอร์นเบิร์กได้รับรางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ในปี พ.ศ. 2492 จากผลงานนี้[ 7 ]ดีเอ็นเอพอลิเมอเรส IIถูกค้นพบโดยโทมัส คอร์นเบิร์ก (บุตรชายของอาร์เธอร์ คอร์นเบิร์ก ) และมัลคอล์ม อี. เกฟเตอร์ ในปี พ.ศ. 2513 ขณะที่กำลังอธิบายบทบาทของ Pol I ในการจำลองดีเอ็นเอของ E. coli เพิ่มเติม [ 8 ]มีการค้นพบดีเอ็นเอพอลิเมอเรสอีก 3 ชนิดในE. coliได้แก่ดีเอ็นเอพอลิเมอเรส III (ค้นพบในช่วงปี พ.ศ. 2513) และดีเอ็นเอพอลิเมอเรส IVและV (ค้นพบในปี พ.ศ. 2542) [ 9 ]ตั้งแต่ปี พ.ศ. 2526 เป็นต้นมา มีการใช้ DNA polymerase ในปฏิกิริยาลูกโซ่โพลีเมอเรส (PCR) และตั้งแต่ปี พ.ศ. 2531 มีการใช้ DNA polymerase ที่ทนความร้อนแทน เนื่องจากไม่จำเป็นต้องเติมในทุกรอบของ PCR
การทำงาน


หน้าที่หลักของเอนไซม์ดีเอ็นเอพอลิเมอเรสคือการสังเคราะห์ดีเอ็นเอจากดีออกซีไรโบนิวคลีโอไทด์ ซึ่งเป็นหน่วยพื้นฐานของดีเอ็นเอ การสร้างสำเนาดีเอ็นเอเกิดขึ้นจากการจับคู่ของนิวคลีโอไทด์กับเบสที่มีอยู่บนแต่ละสายของโมเลกุลดีเอ็นเอเดิม การจับคู่นี้เกิดขึ้นในรูปแบบเฉพาะเสมอ โดยไซโตซีนจับคู่กับกัวนีนและไทมีน จับ คู่กับอะดีนีนจะเกิดเป็นสองคู่แยกกัน ในทางตรงกันข้าม เอนไซม์อาร์เอ็นเอพอลิเมอเรสจะสังเคราะห์อาร์เอ็นเอจากไรโบนิวคลีโอไทด์จากอาร์เอ็นเอหรือดีเอ็นเอ
เมื่อสังเคราะห์ DNA ใหม่ เอนไซม์ DNA polymerase สามารถเพิ่มนิวคลีโอไทด์อิสระได้เฉพาะที่ปลาย 3'ของสาย DNA ที่กำลังสร้างขึ้นใหม่เท่านั้น ส่งผลให้สาย DNA ที่กำลังสร้างขึ้นใหม่ยาวขึ้นในทิศทาง 5'–3'
สิ่งสำคัญที่ควรทราบคือ ทิศทางของสายดีเอ็นเอที่เกิดขึ้นใหม่ (สายลูก) นั้นตรงข้ามกับทิศทางที่เอนไซม์ดีเอ็นเอพอลิเมอเรสเคลื่อนที่ไปตามสายแม่แบบ เนื่องจากเอนไซม์ดีเอ็นเอพอลิเมอเรสต้องการหมู่ 3' OH ที่เป็นอิสระสำหรับการเริ่มต้นการสังเคราะห์ จึงสามารถสังเคราะห์ได้เพียงทิศทางเดียวโดยการต่อเติมปลาย 3' ของสายนิวคลีโอไทด์ที่มีอยู่เดิม ดังนั้น เอนไซม์ดีเอ็นเอพอลิเมอเรสจึงเคลื่อนที่ไปตามสายแม่แบบในทิศทาง 3'–5' และสายลูกจะเกิดขึ้นในทิศทาง 5'–3' ความแตกต่างนี้ทำให้ดีเอ็นเอสองสายที่เกิดขึ้นนั้นประกอบด้วยสายดีเอ็นเอสองสายที่ ขนาน กัน ในทิศทางตรงกันข้าม
การทำงานของเอนไซม์ดีเอ็นเอพอลิเมอเรสไม่ได้สมบูรณ์แบบเสียทีเดียว โดยเอนไซม์จะทำผิดพลาดประมาณหนึ่งครั้งต่อเบสคู่หนึ่งพันล้านคู่ที่ถูกคัดลอก การแก้ไขข้อผิดพลาดเป็นคุณสมบัติของดีเอ็นเอพอลิเมอเรสบางชนิด แต่ไม่ใช่ทุกชนิด กระบวนการนี้จะแก้ไขข้อผิดพลาดในดีเอ็นเอที่สังเคราะห์ขึ้นใหม่ เมื่อตรวจพบเบสคู่ที่ไม่ถูกต้อง ดีเอ็นเอพอลิเมอเรสจะเคลื่อนถอยหลังไปหนึ่งเบสคู่ การทำงานของเอนไซม์ในส่วน 3'–5' exonucleaseจะช่วยให้สามารถตัดเบสคู่ที่ไม่ถูกต้องออกไปได้ (กระบวนการนี้เรียกว่าการตรวจสอบความถูกต้อง ) หลังจากตัดเบสออกแล้ว พอลิเมอเรสสามารถใส่เบสที่ถูกต้องกลับเข้าไปใหม่ได้ และการจำลองแบบก็จะดำเนินต่อไปได้ ซึ่งจะรักษาความสมบูรณ์ของสายดีเอ็นเอเดิมที่ส่งต่อไปยังเซลล์ลูก
ความแม่นยำมีความสำคัญมากในการจำลองดีเอ็นเอ การจับคู่เบสของดีเอ็นเอที่ไม่ตรงกันอาจส่งผลให้โปรตีน ทำงานผิดปกติ และอาจนำไปสู่โรคมะเร็งได้ ดีเอ็นเอพอลิเมอเรสหลายตัวมีโดเมนเอ็กโซนิวคลีเอส ซึ่งทำหน้าที่ตรวจจับการจับคู่เบสที่ไม่ตรงกันและทำหน้าที่กำจัดนิวคลีโอไทด์ที่ไม่ถูกต้องเพื่อแทนที่ด้วยนิวคลีโอไทด์ที่ถูกต้อง[ 10 ]รูปร่างและการโต้ตอบที่รองรับการจับคู่เบสแบบวัตสันและคริกเป็นสิ่งที่มีส่วนสำคัญในการตรวจจับหรือความผิดพลาด พันธะไฮโดรเจนมีบทบาทสำคัญในการจับและการโต้ตอบของการจับคู่เบส การสูญเสียการโต้ตอบที่เกิดขึ้นที่การจับคู่ที่ไม่ตรงกันจะกระตุ้นให้เกิดการเปลี่ยนแปลงสมดุลสำหรับการจับของแม่แบบ-ไพรเมอร์จากพอลิเมอเรสไปยังโดเมนเอ็กโซนิวคลีเอส นอกจากนี้ การรวมนิวคลีโอไทด์ที่ผิดจะทำให้การพอลิเมอไรเซชันของดีเอ็นเอช้าลง ความล่าช้านี้ทำให้ดีเอ็นเอมีเวลาเปลี่ยนจากตำแหน่งพอลิเมอเรสไปยังตำแหน่งเอ็กโซนิวคลีเอส การเปลี่ยนแปลงโครงสร้างและการสูญเสียการโต้ตอบที่แตกต่างกันเกิดขึ้นที่การจับคู่ที่ไม่ตรงกันที่แตกต่างกัน ในการจับคู่ที่ไม่ตรงกันระหว่างพิวรีนและไพริมิดีน จะมีการเคลื่อนที่ของไพริมิดีนไปทางร่องหลักและพิวรีนไปทางร่องรอง เมื่อเทียบกับรูปร่างของช่องยึดของดีเอ็นเอพอลิเมอเรส จะเกิดการชนกันทางสเตอริกขึ้นระหว่างพิวรีนและสารตกค้างในร่องรอง และไพริมิดีนจะสูญเสียปฏิสัมพันธ์แวนเดอร์วาลส์ และไฟฟ้าสถิตที่สำคัญไป [ 11 ]การจับคู่ที่ไม่ตรงกันระหว่างไพริมิดีนและไพริมิดีนจะทำให้เกิดการเปลี่ยนแปลงที่สังเกตได้น้อยกว่า เนื่องจากเบสจะเคลื่อนที่ไปทางร่องหลัก และมีการกีดขวางทางสเตอริกน้อยกว่า อย่างไรก็ตาม แม้ว่าการจับคู่ที่ไม่ตรงกันที่แตกต่างกันจะส่งผลให้คุณสมบัติทางสเตอริกแตกต่างกัน แต่ดีเอ็นเอพอลิเมอเรสก็ยังสามารถตรวจจับและแยกแยะความแตกต่างได้อย่างสม่ำเสมอและรักษาความถูกต้องแม่นยำในการจำลองดีเอ็นเอ[ 12 ]การพอลิเมอไรเซชันของดีเอ็นเอยังมีความสำคัญต่อกระบวนการกลายพันธุ์หลายอย่างและถูกนำไปใช้อย่างกว้างขวางในเทคโนโลยีชีวภาพ
โครงสร้าง
เอนไซม์ DNA polymerase ที่รู้จักกันนั้นมีโครงสร้างที่อนุรักษ์ไว้สูง ซึ่งหมายความว่าหน่วยย่อย เร่งปฏิกิริยาโดยรวมของพวกมัน จะแตกต่างกันเพียงเล็กน้อยในแต่ละสปีชีส์ โดยไม่ขึ้นอยู่กับโครงสร้างโดเมน โครงสร้างที่อนุรักษ์ไว้มักบ่งชี้ถึงหน้าที่สำคัญที่ไม่สามารถทดแทนได้ของเซลล์ ซึ่งการรักษาโครงสร้างดังกล่าวไว้จะให้ประโยชน์ในเชิงวิวัฒนาการ รูปร่างสามารถอธิบายได้ว่าคล้ายกับมือขวาที่มีโดเมนนิ้วหัวแม่มือ นิ้ว และฝ่ามือ โดเมนฝ่ามือดูเหมือนจะทำหน้าที่เร่งปฏิกิริยาการถ่ายโอนหมู่ฟอสฟอริลในปฏิกิริยาการถ่ายโอนฟอสฟอริล DNA จะจับกับฝ่ามือเมื่อเอนไซม์ทำงาน เชื่อกันว่าปฏิกิริยานี้ถูกเร่งปฏิกิริยาโดยกลไกไอออนโลหะสองตัว โดเมนนิ้วทำหน้าที่จับนิวคลีโอไซด์ไตรฟอสเฟตกับฐานแม่แบบ โดเมนนิ้วหัวแม่มือมีบทบาทที่เป็นไปได้ในกระบวนการ การเคลื่อนย้าย และการจัดตำแหน่งของ DNA [ 13 ]
กระบวนการทำงาน
การเร่งปฏิกิริยาอย่างรวดเร็วของ DNA polymerase เกิดจากลักษณะการทำงานแบบต่อเนื่อง (processive nature) ลักษณะการทำงานแบบต่อเนื่องเป็นลักษณะเฉพาะของเอนไซม์ที่ทำงานบนสารตั้งต้นที่เป็นพอลิเมอร์ ในกรณีของ DNA polymerase ระดับการทำงานแบบต่อเนื่องหมายถึงจำนวนนิวคลีโอไทด์เฉลี่ยที่เพิ่มเข้ามาในแต่ละครั้งที่เอนไซม์จับกับแม่แบบ โดยเฉลี่ยแล้ว DNA polymerase ต้องใช้เวลาประมาณหนึ่งวินาทีในการค้นหาและจับกับจุดเชื่อมต่อไพรเมอร์/แม่แบบ เมื่อจับแล้ว DNA polymerase ที่ทำงานแบบต่อเนื่องจะเพิ่มนิวคลี โอไทด์ ในอัตราหนึ่งนิวคลีโอไทด์ต่อวินาที[ 14 ] : 207–208 อย่างไรก็ตาม DNA polymerase ที่ทำงานแบบต่อเนื่องจะเพิ่มนิวคลีโอไทด์หลายตัวต่อวินาที ทำให้เพิ่มอัตราการสังเคราะห์ DNA อย่างมาก ระดับการทำงานแบบต่อเนื่องเป็นสัดส่วนโดยตรงกับอัตราการสังเคราะห์ DNA อัตราการสังเคราะห์ DNA ในเซลล์ที่มีชีวิตถูกกำหนดครั้งแรกเป็นอัตราการยืดตัวของ DNA ของฟาจ T4 ในE. coli ที่ติดเชื้อฟา จ ในช่วงระยะเวลาที่ DNA เพิ่มขึ้นแบบทวีคูณที่อุณหภูมิ 37 °C อัตราการเพิ่มขึ้นของ DNA คือ 749 นิวคลีโอไทด์ต่อวินาที[ 15 ]
ความสามารถของ DNA polymerase ในการเลื่อนไปตามแม่แบบ DNA ช่วยให้เกิดกระบวนการที่เพิ่มขึ้น มีการเพิ่มขึ้นอย่างมากของกระบวนการที่จุดแยกการจำลองแบบ การเพิ่มขึ้นนี้เกิดขึ้นได้จากการที่ DNA polymerase เชื่อมโยงกับโปรตีนที่เรียกว่า sliding DNA clamp clamps เหล่านี้เป็นหน่วยย่อยของโปรตีนหลายหน่วยที่เชื่อมโยงกันในรูปทรงวงแหวน โดยใช้การไฮโดรไลซิสของ ATP โปรตีนกลุ่มหนึ่งที่เรียกว่าsliding clamp loading proteinsจะเปิดโครงสร้างวงแหวนของ sliding DNA clamps ทำให้สามารถจับและปล่อยจากสาย DNA ได้ปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีนกับ clamp จะป้องกันไม่ให้ DNA polymerase แพร่กระจายออกจากแม่แบบ DNA จึงมั่นใจได้ว่าเอนไซม์จะจับกับจุดเชื่อมต่อของไพรเมอร์/แม่แบบเดียวกันและดำเนินการจำลองแบบต่อไป[ 14 ] : 207–208 DNA polymerase จะเปลี่ยนโครงสร้าง เพิ่มความสัมพันธ์กับ clamp เมื่อเชื่อมโยงกับมัน และลดความสัมพันธ์เมื่อการจำลองแบบของ DNA ส่วนหนึ่งเสร็จสมบูรณ์เพื่อให้ปล่อยออกจาก clamp ได้
กระบวนการทำงานของ DNA polymerase ได้รับการศึกษาด้วยการทดลองโมเลกุลเดี่ยวในหลอดทดลอง (เช่นคีมจับแสงและคีมจับแม่เหล็ก ) ซึ่งเผยให้เห็นการทำงานร่วมกันระหว่าง DNA polymerase และโมเลกุลอื่นๆ ของ replisome ( helicaseและSSB ) และกับส้อมการจำลองแบบ DNA [ 16 ]ผลลัพธ์เหล่านี้นำไปสู่การพัฒนารูปแบบจลนศาสตร์แบบเสริมฤทธิ์สำหรับการจำลองแบบ DNA ซึ่งอธิบายถึงการเพิ่มขึ้นของกระบวนการทำงานของ DNA polymerase ที่เกิดขึ้น[ 16 ]
ความแตกต่างระหว่างสายพันธุ์
| ดีเอ็นเอพอลิเมอเรสแฟมิลี A | |||||||
|---|---|---|---|---|---|---|---|
 คู่ c:o6-เมทิล-กัวนีน ในตำแหน่งเบสคู่ของพอลิเมอเรส-2 | |||||||
| ตัวระบุ | |||||||
| เครื่องหมาย | ดีเอ็นเอ_พอล_เอ | ||||||
| พีแฟม | PF00476 | ||||||
| อินเตอร์โปร | IPR001098 | ||||||
| ปราดเปรื่อง | - | ||||||
| โปรไซต์ | PDOC00412 | ||||||
| สโคป2 | 1dpi / SCOPe / SUPFAM | ||||||
| |||||||
| ดีเอ็นเอพอลิเมอเรสแฟมิลี บี | |||||||
|---|---|---|---|---|---|---|---|
 โครงสร้างผลึกของ rb69 gp43 ที่จับคู่กับ DNA ที่มีไทมีนไกลคอล | |||||||
| ตัวระบุ | |||||||
| เครื่องหมาย | ดีเอ็นเอ_พอล_บี | ||||||
| พีแฟม | PF00136 | ||||||
| ตระกูลพีแฟม | ซีแอล0194 | ||||||
| อินเตอร์โปร | IPR006134 | ||||||
| โปรไซต์ | PDOC00107 | ||||||
| สโคป2 | 1noy / SCOPe / SUPFAM | ||||||
| |||||||
| ดีเอ็นเอพอลิเมอเรสชนิด B, ออร์แกเนลล์ และไวรัส | |||||||
|---|---|---|---|---|---|---|---|
 ดีเอ็นเอพอลิเมอเรส phi29 รูปแบบผลึกออร์โธรอมบิก คอมเพล็กซ์ ssdna | |||||||
| ตัวระบุ | |||||||
| เครื่องหมาย | ดีเอ็นเอ_พอล_บี_2 | ||||||
| พีแฟม | PF03175 | ||||||
| ตระกูลพีแฟม | ซีแอล0194 | ||||||
| อินเตอร์โปร | IPR004868 | ||||||
| |||||||
โดยพิจารณาจากความคล้ายคลึงกันของลำดับ สามารถแบ่งเอนไซม์ DNA polymerase ออกเป็นเจ็ดตระกูลย่อย ได้แก่ A, B, C, D, X, Y และ RT
ไวรัสบางชนิดยังสร้างเอนไซม์ดีเอ็นเอพอลิเมอเรสชนิดพิเศษ เช่นดีเอ็นเอพอลิเมอเรสของไวรัสตับอักเสบ บี เอนไซม์เหล่านี้อาจจำลองดีเอ็นเอของไวรัสอย่างเลือกสรรผ่านกลไกต่างๆ ส่วนไวรัสเรโทรไวรัสสร้างเอนไซม์ดีเอ็นเอพอลิเมอเรสชนิดพิเศษที่เรียกว่ารีเวิร์สทรานสคริปเทสซึ่งเป็นดีเอ็นเอพอลิเมอเรสที่ขึ้นอยู่กับอาร์เอ็นเอ (RdDp) มันสร้างดีเอ็นเอจากแม่แบบอาร์เอ็นเอ
| ครอบครัว[ 17 ] | ประเภทของดีเอ็นเอพอลิเมอเรส | แท็กซา | ตัวอย่าง | คุณสมบัติ |
|---|---|---|---|---|
| เอ | โพลีเมอเรสสำหรับการจำลองแบบและการซ่อมแซม | ยูคาริโอตและโปรคาริโอต | เอนไซม์ T7 DNA polymerase, Pol I, Pol γ, θ และ ν | โดเมนเอ็กโซนิวคลีเอสสองโดเมน (3'-5' และ 5'-3') |
| บี | โพลีเมอเรสสำหรับการจำลองแบบและการซ่อมแซม | ยูคาริโอตและโปรคาริโอต | Pol II, Pol B, Pol ζ, Pol α, δ และ ε | 3'-5 เอ็กโซนิวคลีเอส (ตรวจสอบความถูกต้อง); โพลีเมอเรสของไวรัสบางชนิดใช้โปรตีนเป็นไพรเมอร์ |
| ซี | โพลีเมอเรสจำลองแบบ | โปรคาริโอต | Pol III | 3'-5 เอ็กโซนิวคลีเอส (การตรวจสอบความถูกต้อง) |
| ดี | โพลีเมอเรสจำลองแบบ | ยูริอาร์คีโอตา | PolD (เฮเทอโรไดเมอร์ DP1/DP2) [ 18 ] | ไม่มี "ลักษณะมือ" คล้าย RNA polymerase แบบสองลำกล้อง ; 3'-5 exonuclease (ตรวจสอบความถูกต้อง) |
| X | โพลีเมอเรสสำหรับการจำลองแบบและการซ่อมแซม | ยูคาริโอต | Pol β, Pol σ, Pol แล, Pol μ และเทอร์มินัลดีออกซีนิวคลีโอติดิลทรานสเฟอเรส | แม่แบบเป็นตัวเลือก; ฟอสฟาเทส 5' (เฉพาะ Pol β); คุณลักษณะ "มือ" ที่อ่อนแอ |
| วาย | โพลีเมอเรสสำหรับการจำลองแบบและการซ่อมแซม | ยูคาริโอตและโปรคาริโอต | Pol ι, Pol κ, Pol η, [ 19 ] Pol IV และ Pol V | การสังเคราะห์แบบ Translesion [ 20 ] |
| อาร์ที | โพลีเมอเรสสำหรับการจำลองแบบและการซ่อมแซม | ไวรัส, เรโทรไวรัสและยูคาริโอต | เทโลเมอเรส , ไวรัสตับอักเสบ บี | ขึ้นอยู่กับอาร์เอ็นเอ |
โพลีเมอเรสของโปรคาริโอต
เอนไซม์พอลิเมอเรสในแบคทีเรียมีอยู่สองรูปแบบ คือ พอลิเมอเรสหลักและโฮโลเอนไซม์ พอลิเมอเรสหลักสังเคราะห์ DNA จากแม่แบบ DNA แต่ไม่สามารถเริ่มต้นการสังเคราะห์ได้ด้วยตัวเองหรืออย่างแม่นยำ ในขณะที่โฮโลเอนไซม์สามารถเริ่มต้นการสังเคราะห์ได้อย่างแม่นยำ
พอล ไอ
เอนไซม์ พอลิเมอเรสตระกูล A ของโปรคาริโอต ได้แก่ เอนไซม์ DNA polymerase I (Pol I) ซึ่งถูกเข้ารหัสโดยยีนpolA และพบได้ทั่วไปในโปรคาริโอต พอลิเมอเรสซ่อมแซมนี้เกี่ยวข้องกับการซ่อมแซมแบบตัดออกด้วยกิจกรรมเอ็กโซนิวคลีเอสทั้ง 3'–5' และ 5'–3' และการประมวลผลชิ้นส่วน Okazakiที่สร้างขึ้นระหว่างการสังเคราะห์สายล้าหลัง[ 21 ] Pol I เป็นพอลิเมอเรสที่มีปริมาณมากที่สุด คิดเป็น >95% ของกิจกรรมพอลิเมอเรสใน E. coli อย่างไรก็ตาม พบเซลล์ที่ขาด Pol I ซึ่งบ่งชี้ว่ากิจกรรมของ Pol I สามารถถูกแทนที่ได้ด้วยพอลิเมอเรสอีกสี่ตัว Pol I เพิ่มนิวคลีโอไทด์ประมาณ 15-20 ตัวต่อวินาที ดังนั้นจึงแสดงให้เห็นถึงกระบวนการที่ไม่ดี แทนที่จะเป็นเช่นนั้น Pol I เริ่มเพิ่มนิวคลีโอไทด์ที่จุดเชื่อมต่อไพรเมอร์ RNA:แม่แบบที่เรียกว่าต้นกำเนิดของการจำลองแบบ (ori) เมื่ออยู่ห่างจากจุดเริ่มต้นประมาณ 400 bp โฮโลเอนไซม์ Pol III จะถูกประกอบขึ้นและเข้าควบคุมการจำลองแบบด้วยความเร็วและลักษณะการทำงานที่มีประสิทธิภาพสูง[ 22 ]
Taq polymeraseเป็นเอนไซม์ที่ทนความร้อนในกลุ่มนี้ซึ่งไม่มีความสามารถในการตรวจสอบความถูกต้อง [ 23 ]
Pol II
DNA polymerase IIเป็นพอลิเมอเรสตระกูล B ที่เข้ารหัสโดยยีน polB Pol II มีกิจกรรมเอ็กโซนิวคลีเอส 3'–5' และมีส่วนร่วมในการซ่อมแซม DNAการเริ่มต้นการจำลองใหม่เพื่อหลีกเลี่ยงความเสียหาย และการปรากฏตัวของมันในเซลล์สามารถเพิ่มขึ้นจากประมาณ 30-50 สำเนาต่อเซลล์เป็นประมาณ 200–300 ในระหว่างการเหนี่ยวนำ SOS Pol II ยังถูกคิดว่าเป็นตัวสำรองของ Pol III เนื่องจากสามารถโต้ตอบกับโปรตีนโฮโลเอนไซม์และมีระดับการประมวลผลสูง บทบาทหลักของ Pol II ถูกคิดว่าเป็นความสามารถในการชี้นำกิจกรรมพอลิเมอเรสที่ฟอร์กการจำลองและช่วยให้ Pol III ที่หยุดชะงักหลีกเลี่ยงการจับคู่ที่ไม่ตรงกันที่ปลาย[ 24 ]
เอนไซม์ Pfu DNA polymeraseเป็นเอนไซม์ที่ทนความร้อนในตระกูลนี้ พบในอาร์เคีย ที่ทนความร้อนสูง Pyrococcus furiosus [ 25 ] การจำแนกประเภทโดยละเอียดแบ่งตระกูล B ในอาร์เคียออกเป็น B1, B2, B3 โดยที่ B2 เป็นกลุ่มของเอนไซม์เทียม Pfuอยู่ในตระกูล B3 PolB อื่นๆ ที่พบในอาร์เคียเป็นส่วนหนึ่งของ "Casposons" ซึ่งเป็นทรานสโพซอนที่ขึ้นอยู่กับ Cas1 [ 26 ]ไวรัสบางชนิด (รวมถึง Φ29 DNA polymerase ) และพลาสมิดไมโทคอนเดรียก็มี polB เช่นกัน [ 27 ]
Pol III
เอนไซม์ DNA polymerase III holoenzyme เป็นเอนไซม์หลักที่เกี่ยวข้องกับการจำลองแบบ DNA ในE. coliและจัดอยู่ในกลุ่ม polymerase ตระกูล C ประกอบด้วยส่วนประกอบสามส่วน ได้แก่ แกน pol III, ปัจจัยการทำงานของ แคลมป์เลื่อน เบต้า และคอมเพล็กซ์โหลดแคลมป์ แกนประกอบด้วยหน่วยย่อยสามหน่วย ได้แก่ α ซึ่งเป็นศูนย์กลางกิจกรรมของ polymerase, ɛ ซึ่งเป็นตัวตรวจสอบความถูกต้องของ exonucleolytic และ θ ซึ่งอาจทำหน้าที่เป็นตัวทำให้เสถียรสำหรับ ɛ ปัจจัยการทำงานของแคลมป์เลื่อนเบต้ายังมีอยู่เป็นคู่ โดยมีหนึ่งหน่วยสำหรับแต่ละแกน เพื่อสร้างแคลมป์ที่ล้อมรอบ DNA ทำให้มีการทำงานที่มีประสิทธิภาพสูง[ 28 ] ส่วนประกอบที่สามคือ คอมเพล็กซ์โหลดแคลมป์ เจ็ดหน่วยย่อย (τ2γδδ ′ χψ)
แบบจำลอง "ทรอมโบน" ในตำราเก่าแสดงให้เห็นคอมเพล็กซ์การยืดตัวที่มีเอนไซม์หลักสองตัวเทียบเท่าที่แต่ละจุดแยกการจำลองแบบ (RF) หนึ่งตัวสำหรับแต่ละสาย คือสายที่ล้าหลังและสายที่นำหน้า[ 24 ]อย่างไรก็ตาม หลักฐานล่าสุดจากการศึกษาโมเลกุลเดี่ยวบ่งชี้ว่ามีเอนไซม์หลักเทียบเท่าสัดส่วนเฉลี่ยสามตัวที่แต่ละ RF สำหรับทั้ง Pol III และคู่ของมันในB. subtilis คือ PolC [ 29 ] กล้องจุลทรรศน์ฟลูออเรสเซนต์ในเซลล์เผยให้เห็นว่าการสังเคราะห์สายที่นำหน้าอาจไม่ต่อเนื่องอย่างสมบูรณ์ และ Pol III* (เช่น หน่วยย่อย α, ε, τ, δ และ χ ของโฮโลเอนไซม์โดยไม่มีแคลมป์เลื่อน ß2) มีความถี่สูงของการแยกตัวออกจาก RF ที่ทำงานอยู่[ 30 ] ในการศึกษาเหล่านี้ อัตราการหมุนเวียนของจุดแยกการจำลองแบบอยู่ที่ประมาณ 10 วินาทีสำหรับ Pol III*, 47 วินาทีสำหรับแคลมป์เลื่อน ß2 และ 15 นาทีสำหรับเฮลิเคส DnaB สิ่งนี้ชี้ให้เห็นว่าเฮลิเคส DnaB อาจยังคงเชื่อมโยงอย่างมั่นคงที่ RF และทำหน้าที่เป็นจุดเริ่มต้นสำหรับการสร้างโฮโลเอนไซม์ที่มีความสามารถ การศึกษาโมเลกุลเดี่ยว ในหลอดทดลองแสดงให้เห็นว่า Pol III* มีอัตราการหมุนเวียน RF สูงเมื่อมีมากเกินไป แต่ยังคงเชื่อมโยงอย่างมั่นคงกับฟอร์กการจำลองแบบเมื่อความเข้มข้นมีจำกัด[ 30 ] การศึกษาโมเลกุลเดี่ยวอีกการศึกษาหนึ่งแสดงให้เห็นว่ากิจกรรมเฮลิเคส DnaB และการยืดสายสามารถดำเนินไปได้ด้วยจลนศาสตร์แบบสุ่มที่ไม่เชื่อมโยงกัน[ 30 ]
Pol IV
ในE. coliดีเอ็นเอพอลิเมอเรส IV (Pol IV) เป็นดีเอ็นเอพอลิเมอเรสที่มีข้อผิดพลาดได้ง่ายและเกี่ยวข้องกับการกลายพันธุ์แบบไม่เจาะจงเป้าหมาย[ 31 ] Pol IV เป็นพอลิเมอเรสในตระกูล Y ที่แสดงออกโดย ยีน dinBซึ่งถูกเปิดใช้งานผ่านการเหนี่ยวนำ SOS ที่เกิดจากพอลิเมอเรสที่หยุดชะงักที่จุดแยกการจำลองแบบ ในระหว่างการเหนี่ยวนำ SOS การผลิต Pol IV จะเพิ่มขึ้นสิบเท่า และหนึ่งในหน้าที่ในช่วงเวลานี้คือการรบกวนกระบวนการทำงานของเอนไซม์ Pol III แบบสมบูรณ์ ซึ่งจะสร้างจุดตรวจสอบ หยุดการจำลองแบบ และให้เวลาในการซ่อมแซมความเสียหายของดีเอ็นเอผ่านเส้นทางการซ่อมแซมที่เหมาะสม[ 32 ]อีกหน้าที่หนึ่งของ Pol IV คือการสังเคราะห์แบบข้ามรอยโรคที่จุดแยกการจำลองแบบที่หยุดชะงัก เช่น การข้ามสารประกอบ N2-deoxyguanine ในอัตราที่เร็วกว่าการข้ามดีเอ็นเอที่ไม่เสียหาย เซลล์ที่ขาด ยีน dinBจะมีอัตราการกลายพันธุ์ที่สูงขึ้นซึ่งเกิดจากสารที่ทำให้ดีเอ็นเอเสียหาย[ 33 ]
พอล วี
DNA polymerase V (Pol V) เป็น DNA polymerase ในตระกูล Y ที่เกี่ยวข้องกับการตอบสนอง SOSและกลไกการซ่อมแซม DNA แบบtranslesion synthesis [ 34 ]การถอดรหัสของ Pol V ผ่าน ยีน umuDCถูกควบคุมอย่างเข้มงวดเพื่อให้ผลิต Pol V เฉพาะเมื่อมี DNA ที่เสียหายอยู่ในเซลล์ ทำให้เกิดการตอบสนอง SOS โพลีเมอเรสที่หยุดชะงักทำให้RecAจับกับ ssDNA ซึ่งทำให้ โปรตีน LexAย่อยสลายตัวเอง จากนั้น LexAจะสูญเสียความสามารถในการยับยั้งการถอดรหัสของโอเปรอน umuDC นิวคลีโอโปรตีน RecA-ssDNA ตัวเดียวกันนี้จะปรับเปลี่ยนโปรตีน UmuD หลังการแปลเป็นโปรตีน UmuD' UmuD และ UmuD' สร้างเฮเทอโรไดเมอร์ที่โต้ตอบกับ UmuC ซึ่งจะกระตุ้นกิจกรรมเร่งปฏิกิริยาของโพลีเมอเรสของ umuC บน DNA ที่เสียหาย[ 35 ]ในE. coliมีการเสนอแบบจำลอง "เข็มขัดเครื่องมือ" ของพอลิเมอเรสสำหรับการสลับ pol III กับ pol IV ที่จุดหยุดการจำลองแบบ ซึ่งพอลิเมอเรสทั้งสองจะจับกับ β-clamp พร้อมกัน[ 36 ]อย่างไรก็ตาม ยังไม่มีการแสดงให้เห็นถึงการมีส่วนร่วมของพอลิเมอเรส TLS มากกว่าหนึ่งตัวที่ทำงานต่อเนื่องกันเพื่อหลีกเลี่ยงความเสียหายในE. coliยิ่งไปกว่านั้น Pol IV สามารถเร่งปฏิกิริยาทั้งการแทรกและการขยายตัวได้อย่างมีประสิทธิภาพสูง ในขณะที่ pol V ถือเป็นพอลิเมอเรส TLS SOS หลัก ตัวอย่างหนึ่งคือการหลีกเลี่ยงการเชื่อมโยงข้ามกัวนีน-ไทมีนภายในสาย ซึ่งแสดงให้เห็นบนพื้นฐานของความแตกต่างในลายเซ็นการกลายพันธุ์ของพอลิเมอเรสทั้งสองว่า pol IV และ pol V แข่งขันกันเพื่อ TLS ของการเชื่อมโยงข้ามภายในสาย[ 36 ]
ครอบครัว D
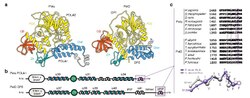
ในปี พ.ศ. 2541 ตระกูล D ของ DNA polymerase ถูกค้นพบในPyrococcus furiosusและMethanococcus jannaschii [ 38 ] คอมเพล็กซ์ PolD เป็นเฮเทอโรไดเมอร์ของสองสาย โดยแต่ละสายถูกเข้ารหัสโดย DP1 (การตรวจสอบความถูกต้องขนาดเล็ก) และ DP2 (ตัวเร่งปฏิกิริยาขนาดใหญ่) แตกต่างจาก DNA polymerase อื่นๆ โครงสร้างและกลไกของแกนเร่งปฏิกิริยา DP2 คล้ายกับของRNA polymerase หลายหน่วยย่อย ส่วนต่อประสาน DP1-DP2 คล้ายกับนิ้วสังกะสีของ polymerase คลาส B ของยูคาริโอตและหน่วยย่อยขนาดเล็ก[ 18 ] DP1 ซึ่ง เป็น เอ็กโซนิวคลีเอสคล้ายMre11 [ 39 ]น่าจะเป็นสารตั้งต้นของหน่วยย่อยขนาดเล็กของ Pol αและεซึ่งให้ความสามารถในการตรวจสอบความถูกต้องที่สูญหายไปในยูคาริโอต[ 26 ]โดเมน HSH ปลาย N ของมันคล้ายกับโปรตีน AAAโดยเฉพาะอย่างยิ่งหน่วยย่อย δ ของ Pol IIIและRuvBในโครงสร้าง[ 40 ] DP2 มีโดเมน KH คลาส II [ 18 ] polD ของ Pyrococcus abyssiมีความเสถียรต่อความร้อนและแม่นยำกว่าพอ ลิเมอเรส Taqแต่ยังไม่ได้รับการนำไปใช้ในเชิงพาณิชย์[ 41 ]มีการเสนอว่าพอลิเมอเรส DNA ตระกูล D เป็นพอลิเมอเรสแรกที่วิวัฒนาการในสิ่งมีชีวิตเซลล์เดียว และพอลิเมอเรสจำลองแบบของบรรพบุรุษเซลล์สากลสุดท้าย (LUCA) จัดอยู่ในตระกูล D [ 42 ]
เอนไซม์ดีเอ็นเอพอลิเมอเรสของยูคาริโอต
โพลีเมอเรส β, แลมบ์ดา, ซิก, μ (เบต้า, แลมบ์ดา, ซิกมา, มู) และ TdT
เอนไซม์พอลิเมอเรสในกลุ่มแฟมิลี X ประกอบด้วยเอนไซม์พอลิเมอเรสยูคาริโอติกที่รู้จักกันดีอย่าง pol β (เบตา) รวมถึงเอนไซม์พอลิเมอเรสยูคาริโอติกอื่นๆ เช่น Pol σ (ซิกมา), Pol λ (แลมบ์ดา) , Pol μ (มิว)และเอนไซม์เทอร์มินัลดีออกซีนิวคลีโอไทด์ทรานสเฟอเรส (TdT)เอนไซม์พอลิเมอเรสในกลุ่มแฟมิลี X พบได้ส่วนใหญ่ในสัตว์มีกระดูกสันหลัง และพบได้บ้างในพืชและเชื้อรา เอนไซม์เหล่านี้มีบริเวณที่มีการอนุรักษ์สูง ซึ่งรวมถึงโมทีฟแบบเฮลิกซ์-แฮร์พิน-เฮลิกซ์สองแบบที่สำคัญต่อปฏิกิริยาระหว่างดีเอ็นเอกับพอลิเมอเรส โมทีฟหนึ่งตั้งอยู่ในโดเมน 8 kDa ที่ทำปฏิกิริยากับดีเอ็นเอส่วนปลาย และอีกโมทีฟหนึ่งตั้งอยู่ในโดเมนแบบนิ้วหัวแม่มือที่ทำปฏิกิริยากับสายไพรเมอร์ Pol β ซึ่งเข้ารหัสโดยยีน POLB จำเป็นสำหรับ การซ่อมแซมการตัดฐานแบบสั้นซึ่งเป็นเส้นทางการซ่อมแซม DNA ที่จำเป็นสำหรับการซ่อมแซมฐานที่ถูกอัลคิเลตหรือออกซิไดซ์ รวมถึงไซต์ที่ไม่มี เบส Pol λ และ Pol μ ซึ่งเข้ารหัสโดย ยีน POLLและPOLMตามลำดับ มีส่วนเกี่ยวข้องกับการเชื่อมต่อปลายที่ไม่เหมือนกันซึ่งเป็นกลไกในการเชื่อมต่อรอยแตกของ DNA สองสายที่เกิดจากไฮโดรเจนเปอร์ออกไซด์และรังสีไอออนไนซ์ ตามลำดับ TdT แสดงออกเฉพาะในเนื้อเยื่อน้ำเหลือง และเพิ่ม "n นิวคลีโอไทด์" ให้กับรอยแตกสองสายที่เกิดขึ้นระหว่างการรวมตัวของ V(D)Jเพื่อส่งเสริมความหลากหลายทางภูมิคุ้มกัน[ 43 ]
เอนไซม์พอลิเมอเรส α, δ และ ε (อัลฟา เดลต้า และเอปซิลอน)
Pol α (อัลฟา) , Pol δ (เดลต้า)และPol ε (เอปไซลอน)เป็นสมาชิกของ Family B Polymerases และเป็นพอลิเมอเรสหลักที่เกี่ยวข้องกับการจำลองแบบ DNA ในนิวเคลียส คอมเพล็กซ์ Pol α (คอมเพล็กซ์ pol α-DNA primase) ประกอบด้วยหน่วยย่อยสี่หน่วย ได้แก่ หน่วยย่อยเร่งปฏิกิริยาPOLA1หน่วยย่อยควบคุมPOLA2และหน่วยย่อยไพรเมสขนาดเล็กและขนาดใหญ่PRIM1และPRIM2ตามลำดับ เมื่อไพรเมสสร้างไพรเมอร์ RNA แล้ว Pol α จะเริ่มการจำลองแบบโดยยืดไพรเมอร์ออกไปประมาณ 20 นิวคลีโอไทด์[ 44 ]เนื่องจากมีกระบวนการทำงานสูง Pol δ จึงเข้ามารับช่วงการสังเคราะห์สายนำและสายตามจาก Pol α [ 14 ] : 218–219 Pol δ ถูกแสดงออกโดยยีนPOLD1ซึ่งสร้างหน่วยย่อยเร่งปฏิกิริยาPOLD2 , POLD3และPOLD4ซึ่งสร้างหน่วยย่อยอื่นๆ ที่มีปฏิสัมพันธ์กับProliferating Cell Nuclear Antigen (PCNA) ซึ่งเป็นแคลมป์ DNAที่ทำให้ Pol δ มีความสามารถในการประมวลผล[ 45 ] Pol ε ถูกเข้ารหัสโดย ยีน POLE1ซึ่งเป็นหน่วยย่อยเร่งปฏิกิริยาPOLE2และPOLE3มีรายงานว่าหน้าที่ของ Pol ε คือการขยายสายนำในระหว่างการจำลองแบบ[ 46 ] [ 47 ]ในขณะที่ Pol δ ส่วนใหญ่จะจำลองแบบสายตามหลัง อย่างไรก็ตาม หลักฐานล่าสุดชี้ให้เห็นว่า Pol δ อาจมีบทบาทในการจำลองแบบสายนำของ DNA ด้วยเช่นกัน[ 48 ]บริเวณ "ซากพอลิเมอเรส" ปลาย C ของ Pol ε แม้ว่าจะไม่จำเป็นต่อกิจกรรมพอลิเมอเรส[ 49 ]ก็เชื่อกันว่ามีความสำคัญต่อความมีชีวิตของเซลล์ บริเวณปลาย C นี้เชื่อกันว่าทำหน้าที่เป็นจุดตรวจสอบก่อนเข้าสู่ระยะแอนาเฟส ให้ความเสถียรแก่โฮโลเอนไซม์ และเพิ่มโปรตีนให้กับโฮโลเอนไซม์ที่จำเป็นสำหรับการเริ่มต้นการจำลองแบบ[ 50 ] Pol ε มีโดเมน "ฝ่ามือ" ที่ใหญ่กว่าซึ่งให้ประสิทธิภาพการทำงานสูงโดยไม่ขึ้นอยู่กับ PCNA [ 49 ]
เมื่อเปรียบเทียบกับพอลิเมอเรสตระกูล B อื่นๆ ตระกูลเอ็กโซนิวคลีเอส DEDD ที่รับผิดชอบการตรวจสอบความถูกต้องจะถูกปิดใช้งานใน Pol α [ 26 ] Pol ε มีเอกลักษณ์ตรงที่มีโดเมนนิ้วสังกะสีสองโดเมนและสำเนาที่ไม่ทำงานของพอลิเมอเรสตระกูล B อีกตัวหนึ่งที่ปลาย C การมีอยู่ของนิ้วสังกะสีนี้มีความเกี่ยวข้องกับต้นกำเนิดของยูคาริโอตา ซึ่งในกรณีนี้จะถูกจัดอยู่ใน กลุ่ม Asgardร่วมกับพอลิเมอเรส B3 ของอาร์เคีย[ 51 ]
โพลีเมอเรส η, ι และ κ (eta, iota และ kappa)
Pol η (eta) , Pol ι (iota)และPol κ (kappa)เป็นเอนไซม์พอลิเมอเรสดีเอ็นเอในแฟมิลี Y ที่เกี่ยวข้องกับการซ่อมแซมดีเอ็นเอโดยการสังเคราะห์แบบแปลรหัส และถูกเข้ารหัสโดยยีน POLH, POLIและPOLKตามลำดับ สมาชิกในแฟมิลี Y มีโมทีฟทั่วไปห้าแบบเพื่อช่วยในการจับกับสารตั้งต้นและปลายไพรเมอร์ และทั้งหมดประกอบด้วยโดเมนทั่วไปของนิ้วหัวแม่มือ ฝ่ามือ และนิ้วมือข้างขวา พร้อมด้วยโดเมนเพิ่มเติม เช่น นิ้วก้อย (LF), โดเมนที่เกี่ยวข้องกับพอลิเมอเรส (PAD) หรือข้อมือ อย่างไรก็ตาม บริเวณออกฤทธิ์จะแตกต่างกันไปในแต่ละสมาชิกในแฟมิลีเนื่องจากความเสียหายที่ได้รับการซ่อมแซมแตกต่างกัน พอลิเมอเรสในแฟมิลี Y เป็นพอลิเมอเรสที่มีความแม่นยำต่ำ แต่ได้รับการพิสูจน์แล้วว่ามีประโยชน์มากกว่าโทษ เนื่องจากกลายพันธุ์ที่ส่งผลต่อพอลิเมอเรสสามารถก่อให้เกิดโรคต่างๆ ได้ เช่นมะเร็งผิวหนังและโรค Xeroderma Pigmentosum Variant (XPS)ความสำคัญของพอลิเมอเรสเหล่านี้ได้รับการพิสูจน์แล้วจากข้อเท็จจริงที่ว่ายีนที่เข้ารหัส DNA พอลิเมอเรส η ถูกเรียกว่า XPV เนื่องจากการสูญเสียยีนนี้ส่งผลให้เกิดโรค Xeroderma Pigmentosum Variant Pol η มีความสำคัญอย่างยิ่งในการช่วยให้การสังเคราะห์แบบทรานส์เลชันของความเสียหายของ DNA ที่เกิดจากรังสีอัลตราไวโอเลต เป็นไปอย่างแม่นยำ การทำงานของ Pol κ ยังไม่เป็นที่เข้าใจอย่างสมบูรณ์ แต่นักวิจัยได้พบหน้าที่ที่เป็นไปได้สองประการ เชื่อกันว่า Pol κ ทำหน้าที่เป็นตัวขยายหรือตัวแทรกเบสเฉพาะที่ความเสียหายของ DNA บางแห่ง พอลิเมอเรสการสังเคราะห์แบบทรานส์เลชันทั้งสามตัวพร้อมกับ Rev1 จะถูกดึงดูดไปยังรอยโรคที่เสียหายผ่าน DNA พอลิเมอเรสการจำลองแบบที่หยุดชะงัก มีสองเส้นทางในการซ่อมแซมความเสียหายซึ่งนำไปสู่การที่นักวิจัยสรุปว่าเส้นทางที่เลือกขึ้นอยู่กับว่าสายใดมีความเสียหาย สายนำหรือสายตาม[ 52 ]
โพลีเมอเรส Rev1 และ ζ (ซีตา)
Pol ζ ซึ่งเป็นพอลิเมอเรสในกลุ่ม B อีกชนิดหนึ่ง ประกอบด้วยสองหน่วยย่อย ได้แก่Rev3ซึ่งเป็นหน่วยย่อยเร่งปฏิกิริยา และ Rev7 ( MAD2L2 ) ซึ่งเพิ่มประสิทธิภาพการเร่งปฏิกิริยาของพอลิเมอเรส และมีส่วนเกี่ยวข้องกับการสังเคราะห์แบบแปลรหัส Pol ζ ขาดกิจกรรมเอ็กโซนิวคลีเอสจาก 3' ไป 5' และมีความพิเศษตรงที่สามารถต่อขยายไพรเมอร์ที่มีปลายไม่ตรงกันได้Rev1มีสามบริเวณที่น่าสนใจ ได้แก่โดเมน BRCTโดเมนจับยูบิควิตินและโดเมนปลาย C และมีความสามารถในการถ่ายโอน dCMP ซึ่งจะเติมดีออกซีไซทิดีนตรงข้ามกับรอยโรคที่จะทำให้พอลิเมอเรสจำลอง แบบ Pol δและPol εหยุดชะงัก พอลิเมอเรสที่หยุดชะงักเหล่านี้จะกระตุ้นคอมเพล็กซ์ยูบิควิติน ซึ่งจะทำให้พอลิเมอเรสจำลองแบบแยกตัวออก และดึงดูด Pol ζ และ Rev1 เข้ามา เมื่อรวมกันแล้ว Pol ζ และ Rev1 จะเติมดีออกซีไซทิดีน และ Pol ζ จะต่อขยายผ่านรอยโรคไปได้ ผ่านกระบวนการที่ยังไม่ทราบแน่ชัด Pol ζ จะแยกตัวออก และพอลิเมอเรสการจำลองจะกลับมารวมตัวกันและดำเนินการจำลองต่อไป Pol ζ และ Rev1 ไม่จำเป็นสำหรับการจำลอง แต่การสูญเสียยีน REV3 ในยีสต์ที่กำลังแตกหน่ออาจทำให้มีความไวต่อสารที่ทำลาย DNA เพิ่มขึ้นเนื่องจากการยุบตัวของส้อมการจำลองที่พอลิเมอเรสการจำลองหยุดชะงัก[ 53 ]
เทโลเมอเรส
เทโลเมอเรสเป็นไรโบนิวคลีโอโปรตีนที่ทำหน้าที่จำลองปลายของโครโมโซมเชิงเส้น เนื่องจากดีเอ็นเอพอลิเมอเรสปกติไม่สามารถจำลองปลายหรือเทโลเมียร์ได้ ส่วนปลาย 3' ของโครโมโซมสองสายที่มีลำดับ 5'-TTAGGG-3' จะดึงดูดเทโลเมอเรส เทโลเมอเรสทำงานเหมือนกับดีเอ็นเอพอลิเมอเรสอื่นๆ โดยการขยายปลาย 3' แต่แตกต่างจากดีเอ็นเอพอลิเมอเรสอื่นๆ ตรงที่เทโลเมอเรสไม่ต้องการแม่แบบ หน่วยย่อย TERT ซึ่งเป็นตัวอย่างของรีเวิร์สทรานสคริปเทส ใช้หน่วยย่อย RNA เพื่อสร้างจุดเชื่อมต่อไพรเมอร์-แม่แบบที่ทำให้เทโลเมอเรสสามารถขยายปลาย 3' ของปลายโครโมโซมได้ การลดขนาดของเทโลเมียร์อย่างค่อยเป็นค่อยไปอันเป็นผลมาจากการจำลองหลายครั้งตลอดช่วงชีวิตนั้นเชื่อว่าเกี่ยวข้องกับผลกระทบของความชรา[ 14 ] : 248–249
โพลีเมอเรส γ, θ และ ν (แกมมา, ทีต้า และ nu)
Pol γ (แกมมา), Pol θ (ธีตา) และ Pol ν (นิว) เป็นพอลิเมอเรสในกลุ่ม A Pol γ ซึ่งถูกเข้ารหัสโดย ยีน POLGเคยถูกคิดว่าเป็นพอ ลิเมอเรส ในไมโท คอนเดรียเพียงชนิดเดียวมานานแล้ว อย่างไรก็ตาม การวิจัยล่าสุดแสดงให้เห็นว่าอย่างน้อยPol β (เบตา)ซึ่งเป็นพอลิเมอเรสในกลุ่ม X ก็มีอยู่ในไมโทคอนเด รียเช่นกัน [ 54 ] [ 55 ]การกลายพันธุ์ใดๆ ที่นำไปสู่การทำงานของ Pol γ ที่จำกัดหรือไม่ทำงานเลยจะมีผลกระทบอย่างมากต่อ mtDNA และเป็นสาเหตุที่พบบ่อยที่สุดของโรคไมโทคอนเดรียที่ถ่ายทอดทางพันธุกรรมแบบออโตโซม[ 56 ] Pol γ ประกอบด้วยโดเมนพอลิเมอเรสที่ปลาย C และโดเมนเอ็กโซนิวคลีเอส 3'–5' ที่ปลาย N ซึ่งเชื่อมต่อกันผ่านบริเวณตัวเชื่อมซึ่งจับกับหน่วยย่อยเสริม หน่วยย่อยเสริมจะจับกับ DNA และจำเป็นสำหรับกระบวนการทำงานของ Pol γ การกลายพันธุ์แบบจุด A467T ในบริเวณตัวเชื่อมเป็นสาเหตุของความผิดปกติของไมโทคอนเดรียที่เกี่ยวข้องกับ Pol γ มากกว่าหนึ่งในสาม[ 57 ] แม้ว่าจะมีโฮโมล็อกของ Pol θ จำนวนมากที่เข้ารหัสโดย ยีน POLQอยู่ในยูคาริโอต แต่หน้าที่ของมันยังไม่เป็นที่เข้าใจอย่างชัดเจน ลำดับของกรดอะมิโนที่ปลาย C เป็นตัวจัดประเภท Pol θ เป็นพอลิเมอเรสตระกูล A แม้ว่าอัตราความผิดพลาดของ Pol θ จะมีความสัมพันธ์ใกล้ชิดกับพอลิเมอเรสตระกูล Y มากกว่า Pol θ ขยายปลายไพรเมอร์ที่ไม่ตรงกันและสามารถข้ามไซต์ที่ไม่มีเบสได้โดยการเพิ่มนิวคลีโอไทด์ นอกจากนี้ยังมีกิจกรรมดีออกซีไรโบฟอสโฟไดเอสเตอเรส (dRPase) ในโดเมนพอลิเมอเรสและสามารถแสดง กิจกรรม ATPaseในบริเวณใกล้เคียงกับ ssDNA [ 58 ] Pol ν (nu) ถือว่าเป็นเอนไซม์พอลิเมอเรสที่มีประสิทธิภาพน้อยที่สุด[ 59 ]อย่างไรก็ตาม DNA polymerase nu มีบทบาทสำคัญในการซ่อมแซมโฮโมโลจีระหว่างการตอบสนองของเซลล์ต่อการเชื่อมโยงข้าม โดยทำหน้าที่ของมันในคอมเพล็กซ์กับเฮลิเคส[ 59 ]
พืชใช้พอลิเมอเรสตระกูล A สองตัวเพื่อคัดลอกทั้งจีโนมไมโตคอนเดรียและพลาสติด พวกมันมีความคล้ายคลึงกับ Pol I ของแบคทีเรียมากกว่า Pol γ ของสัตว์เลี้ยงลูกด้วยนม[ 60 ]
รีเวิร์สทรานสคริปเทส
เรโทรไวรัสเข้ารหัสเอนไซม์ดีเอ็นเอพอลิเมอเรสชนิดพิเศษที่เรียกว่ารีเวิร์สทรานสคริปเทส ซึ่งเป็นเอนไซม์ดีเอ็นเอพอลิเมอเรสที่ขึ้นอยู่กับอาร์เอ็นเอ (RdDp) ที่สังเคราะห์ดีเอ็นเอจากแม่แบบอาร์เอ็นเอ ตระกูลรีเวิร์สทรานสคริปเทสประกอบด้วยทั้งฟังก์ชันการทำงานของดีเอ็นเอพอลิเมอเรสและฟังก์ชันการทำงานของอาร์เอ็นเอสเอช ซึ่งย่อยสลายอาร์เอ็นเอที่จับคู่กับดีเอ็นเอ ตัวอย่างของเรโทรไวรัสคือเอชไอวี [ 14 ] รีเวิร์สทรานสคริปเทสถูกนำมาใช้กันอย่างแพร่หลายในการขยายอาร์เอ็นเอเพื่อวัตถุประสงค์ในการวิจัย โดยใช้แม่แบบอาร์เอ็นเอ PCR สามารถใช้รีเวิร์สทรานสคริปเทสเพื่อสร้างแม่แบบดีเอ็นเอ จากนั้นแม่แบบดีเอ็นเอใหม่นี้สามารถนำไปใช้ในการขยาย PCR ทั่วไปได้ ผลิตภัณฑ์ของการทดลองดังกล่าวจึงเป็นผลิตภัณฑ์ PCR ที่ขยายจากอาร์เอ็นเอ[ 9 ]
อนุภาคไวรัสเรโทรไวรัสเอชไอวีแต่ละตัวมีจีโนมRNA สองตัว แต่หลังจากการติดเชื้อ ไวรัสแต่ละตัวจะสร้างโปรไวรัสเพียง ตัวเดียว [ 61 ] หลังจากการติดเชื้อ การถอดรหัสย้อนกลับจะเกิดขึ้นพร้อมกับการสลับแม่แบบระหว่างสำเนาจีโนมทั้งสอง (การรวมตัวใหม่แบบเลือกสำเนา) [ 61 ] จะมีเหตุการณ์การรวมตัวใหม่เกิดขึ้น 5 ถึง 14 ครั้งต่อจีโนมในแต่ละรอบการจำลอง แบบ [ 62 ] การสลับแม่แบบ (การรวมตัวใหม่) ดูเหมือนจะจำเป็นสำหรับการรักษาความสมบูรณ์ของจีโนมและเป็นกลไกการซ่อมแซมเพื่อกู้คืนจีโนมที่เสียหาย[ 63 ] [ 61 ]
เอนไซม์ดีเอ็นเอพอลิเมอเรสของแบคทีริโอเฟจ T4
แบคทีริโอเฟจ (ฟาจ) T4เข้ารหัสเอนไซม์ดีเอ็นเอพอลิเมอเรสที่เร่งปฏิกิริยาการสังเคราะห์ดีเอ็นเอในทิศทาง 5' ถึง 3' [ 64 ] เอนไซม์ พอลิเมอเรสของฟาจยังมีกิจกรรมเอ็กโซนิวคลีเอสที่ทำงานในทิศทาง 3' ถึง 5' [ 65 ]และกิจกรรมนี้ถูกนำมาใช้ในการตรวจสอบและแก้ไขเบสที่แทรกใหม่[ 66 ] พบว่า ฟาจกลายพันธุ์ที่มีเอนไซม์ดีเอ็นเอพอลิเมอเรสที่ไวต่ออุณหภูมิเมื่อเจริญเติบโตที่อุณหภูมิที่เหมาะสม จะเกิดการรวมตัวใหม่ในความถี่ที่สูงกว่าฟาจชนิดปกติประมาณสองเท่า[ 67 ]
มีการเสนอว่าการเปลี่ยนแปลงการกลายพันธุ์ในพอลิเมอเรส DNA ของฟาจสามารถกระตุ้นการสลับสายแม่แบบ (การรวมตัวเลือกสำเนา) ในระหว่างการจำลองแบบ[ 67 ]
ดูเพิ่มเติม
- เครื่องจักรชีวภาพ
- การจัดลำดับดีเอ็นเอ
- การเร่งปฏิกิริยาโดยเอนไซม์
- การรวมตัวทางพันธุกรรม
- การโคลนนิ่งระดับโมเลกุล
- ปฏิกิริยาลูกโซ่พอลิเมอเรส
- พลวัตของโดเมนโปรตีน
- การถอดรหัสย้อนกลับ
- อาร์เอ็นเอพอลิเมอเรส
- เอนไซม์ Taq DNA polymerase
อ่านเพิ่มเติม
- Burgers PM, Koonin EV, Bruford E, Blanco L, Burtis KC, Christman MF และคณะ (พฤศจิกายน 2544). "เอนไซม์ DNA polymerase ของยูคาริโอต: ข้อเสนอสำหรับระบบการตั้งชื่อที่แก้ไขใหม่"วารสารเคมีชีวภาพ 276 ( 47): 43487– 43490. Bibcode : 2001JBiCh.27643487B . doi : 10.1074/jbc.R100056200 . hdl : 10261/338658 . PMID 11579108 .
ลิงก์ภายนอก
- DNA+polymerases ที่ หัวข้อทางการ แพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
- โมเลกุลประจำเดือนของPDB : ดีเอ็นเอพอลิเมอเรส
- กลไกการซ่อมแซมที่ผิดปกติในดีเอ็นเอพอลิเมอเรสแลมบ์ดามหาวิทยาลัยโอไฮโอสเตท 25 กรกฎาคม 2549
- ภาพเคลื่อนไหวที่ยอดเยี่ยมเกี่ยวกับเอนไซม์ DNA Polymerase จาก WEHI นาทีที่ 1:45 ในไฟล์เก็บถาวรเมื่อวันที่ 5 ธันวาคม 2014 บนWayback Machine
- โครงสร้างระดับมหภาคสามมิติของเอนไซม์ดีเอ็นเอพอลิเมอเรสจากฐานข้อมูล EM (EMDB)
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ดีเอ็นเอพอลิเมอเรส
ดีเอ็นเอพอลิเมอเรสเป็นสมาชิกของกลุ่มเอนไซม์ที่เร่งปฏิกิริยาการสังเคราะห์โมเลกุลดีเอ็นเอ จากนิวคลีโอไซด์ไตรฟอสเฟตซึ่งเป็นสารตั้งต้นโมเลกุลของดีเอ็นเอ
ประวัติศาสตร์
ในปี พ.ศ. 2499 อาร์เธอร์ คอร์นเบิร์ก และเพื่อนร่วมงานได้ค้นพบ ดีเอ็นเอพอลิเมอเรส I (Pol I) ใน แบคทีเรีย Escherichia coli พวกเขาได้อธิบายกระบวนการจำลองดีเอ็นเอโดยที่ดีเอ็นเอพอลิเมอเรสจะคัดลอกลำดับเบสของสายดีเอ็นเอแม่แบบ คอร์นเบิร์กได้รับ...
การทำงาน
หน้าที่หลักของเอนไซม์ดีเอ็นเอพอลิเมอเรสคือการสังเคราะห์ดีเอ็นเอจาก ดีออกซีไร โบนิวคลีโอไทด์ ซึ่งเป็นหน่วยพื้นฐานของดีเอ็นเอ การสร้างสำเนาดีเอ็นเอเกิดขึ้นจากการจับคู่ของนิวคลีโอไทด์กับเบสที่มีอยู่บนแต่ละสายของโมเลกุลดีเอ็นเอเดิม...
โครงสร้าง
เอนไซม์ DNA polymerase ที่รู้จักกันนั้นมีโครงสร้างที่อนุรักษ์ไว้สูง ซึ่งหมายความว่า หน่วยย่อย เร่งปฏิกิริยาโดยรวมของพวกมัน จะแตกต่างกันเพียงเล็กน้อยในแต่ละสปีชีส์ โดยไม่ขึ้นอยู่กับโครงสร้างโดเมน...