อ่าน 10 นาที
จุดเชื่อมต่อประสาทกล้ามเนื้อ
จุด เชื่อมต่อประสาทกล้ามเนื้อ (หรือ จุดเชื่อมต่อกล้ามเนื้อและเส้นประสาท ) คือ ไซแนปส์ทางเคมี ระหว่าง เซลล์ประสาทสั่งการ และ เส้นใยกล้าม เนื้อ [ 1 ]
จุดเชื่อมต่อประสาทกล้ามเนื้อ
| จุดเชื่อมต่อประสาทกล้ามเนื้อของมนุษย์ | |
|---|---|
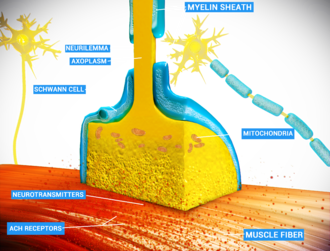 ที่จุดเชื่อมต่อระหว่างเส้นประสาทและกล้ามเนื้อ เส้นใยประสาทสามารถส่งสัญญาณไปยังเส้นใยกล้ามเนื้อได้โดยการปล่อย ACh (และสารอื่นๆ) ซึ่งทำให้กล้ามเนื้อหดตัว | |
 ภาพแสดงแผนผังของจุดเชื่อมต่อประสาทกล้ามเนื้อ | |
| รายละเอียด | |
| ตัวระบุ | |
| ละติน | ไซแนปซิส นิวโรมัสคูลาริส; จังก์ติโอ นิวโรมัสคูลาริส |
| เมช | D009469 |
| ไทย | H2.00.06.1.02001 |
| เอฟเอ็มเอ | 61803 |
| ศัพท์ทางกายวิภาคศาสตร์ | |

จุดเชื่อมต่อประสาทกล้ามเนื้อ (หรือจุดเชื่อมต่อกล้ามเนื้อและเส้นประสาท ) คือไซแนปส์ทางเคมีระหว่างเซลล์ประสาทสั่งการและเส้นใยกล้ามเนื้อ[ 1 ]
ช่วยให้เซลล์ประสาทสั่งการส่งสัญญาณไปยังเส้นใยกล้ามเนื้อ ทำให้เกิดการหดตัวของกล้ามเนื้อ[ 2 ]
กล้ามเนื้อต้องการการควบคุมโดยเส้นประสาทเพื่อการทำงาน และแม้กระทั่งเพื่อรักษาสภาพกล้ามเนื้อป้องกันการฝ่อลีบในระบบประสาทและกล้ามเนื้อ เส้นประสาทจากระบบประสาทส่วนกลางและระบบประสาทส่วนปลายจะเชื่อมโยงกันและทำงานร่วมกับกล้ามเนื้อ[ 3 ]การส่งสัญญาณประสาทที่จุดเชื่อมต่อประสาทและกล้ามเนื้อเริ่มต้นเมื่อศักยภาพการกระทำไปถึงปลายประสาทก่อนซินแนปส์ของเซลล์ประสาท สั่งการ ซึ่งจะกระตุ้นช่องแคลเซียมที่ควบคุมด้วยแรงดันไฟฟ้าเพื่อให้ไอออนแคลเซียมเข้าสู่เซลล์ประสาท ไอออนแคลเซียมจะจับกับโปรตีนเซนเซอร์ ( ซินแนปโทแท็กมิน ) บนถุงซินแนปส์ กระตุ้นให้ถุงซินแนปส์รวมตัวกับเยื่อหุ้มเซลล์และ ปล่อย สารสื่อประสาท จากเซลล์ประสาทสั่งการ เข้าไปในช่องว่างซินแนปส์ในสัตว์มีกระดูกสันหลังเซลล์ประสาทสั่งการจะปล่อยอะเซทิลโคลีน (ACh) ซึ่งเป็นสารสื่อประสาทโมเลกุลเล็ก ซึ่งแพร่กระจายข้ามช่องว่างซินแนปส์และจับกับตัวรับอะเซทิลโคลีนนิโคตินิก (nAChRs) บนเยื่อหุ้มเซลล์ของเส้นใยกล้ามเนื้อ หรือที่เรียกว่าซาร์โคเลมมา nAChR เป็น ตัวรับ แบบไอโอโนโทรปิกซึ่งหมายความว่ามันทำหน้าที่เป็นช่องไอออนที่ควบคุมโดยลิแกนด์การจับกันของ ACh กับตัวรับสามารถทำให้เส้นใยกล้ามเนื้อเกิดการเปลี่ยนแปลงศักย์ไฟฟ้า ก่อให้เกิดปฏิกิริยาลูกโซ่ที่ในที่สุดจะนำไปสู่การหดตัวของกล้ามเนื้อ
โรคที่เกี่ยวข้องกับจุดเชื่อมต่อประสาทกล้ามเนื้ออาจมีสาเหตุมาจากพันธุกรรมและระบบภูมิคุ้มกันผิดปกติ ความผิดปกติ ทางพันธุกรรม เช่น กลุ่มอาการกล้ามเนื้ออ่อนแรง แต่กำเนิด (Congenital myasthenic syndrome ) เกิดจากการกลายพันธุ์ของโปรตีนโครงสร้างที่ประกอบเป็นจุดเชื่อมต่อประสาทกล้ามเนื้อ ในขณะที่โรคที่เกิดจากระบบภูมิคุ้มกันผิดปกติ เช่น โรคกล้ามเนื้ออ่อนแรงชนิด ไมแอสทีเนียกราวิส ( Myasthenia gravis ) เกิดขึ้นเมื่อร่างกายสร้างแอนติบอดีต่อต้านตัวรับอะเซทิลโคลีนชนิดนิโคตินิกบนซาร์โคเลมมา
โครงสร้างและหน้าที่
การส่งผ่านควอนตัม
ที่จุดเชื่อมต่อประสาทกล้ามเนื้อ แอกซอนมอเตอร์ก่อนซิแนปส์จะสิ้นสุดลงที่ระยะ 30 นาโนเมตรจากเยื่อหุ้มเซลล์หรือซาร์โคเลมมาของเส้นใยกล้ามเนื้อ ซาร์โคเลมมาที่จุดเชื่อมต่อมีส่วนเว้าที่เรียกว่ารอยพับหลังจุดเชื่อมต่อซึ่งเพิ่มพื้นที่ผิวที่หันเข้าหาช่องว่างซิแนปส์[ 4 ] รอยพับหลังจุดเชื่อมต่อเหล่านี้ก่อตัวเป็นแผ่นปลายมอเตอร์ ซึ่งมีตัวรับอะเซทิลโคลีนนิโคตินิก (nAChRs) กระจายอยู่หนาแน่นที่ความหนาแน่น 10,000 ตัวรับ/μm² [ 5 ] แอก ซอนก่อนซิแนป ส์ จะสิ้นสุดลงที่ส่วนนูนที่เรียกว่าปุ่มปลาย (หรือปลายก่อนซิแนปส์) ซึ่งยื่นออกไปทางรอยพับหลังจุดเชื่อมต่อของซาร์โคเลมมา ในกบ ปลายประสาทมอเตอร์แต่ละอันจะมีเวสิเคิล ประมาณ 500,000 อัน [ 6 ]โดยมีเส้นผ่านศูนย์กลางเฉลี่ย 0.05 ไมโครเมตร เวสิเคิลเหล่านี้บรรจุอะเซทิลโคลีน เวสิเคิลบางส่วนเหล่านี้รวมตัวกันเป็นกลุ่มละห้าสิบเวสิเคิล วางอยู่ที่บริเวณแอคทีฟโซนใกล้กับเยื่อหุ้มประสาท บริเวณแอคทีฟโซนอยู่ห่างกันประมาณ 1 ไมโครเมตร ช่องว่างขนาด 30 นาโนเมตรระหว่างปลายประสาทและแผ่นปลายประสาทมีโครงข่ายของอะเซทิลโคลีนเอสเทอเรส (AChE) ที่ความหนาแน่น 2,600 โมเลกุลของเอนไซม์/μm² ซึ่งยึดไว้ด้วยโปรตีนโครงสร้างไดสโทรฟินและแรปซินนอกจากนี้ยังมีโปรตีนตัวรับไทโรซีนไคเนสMuSKซึ่งเป็นโปรตีนส่งสัญญาณที่เกี่ยวข้องกับการพัฒนาของจุดเชื่อมต่อประสาทกล้ามเนื้อ ซึ่งยึดไว้ด้วยแรปซินเช่นกัน[ 4 ]
ประมาณทุกๆ วินาที ในจุดเชื่อมต่อที่พักตัว ถุงไซแนปส์หนึ่งถุงจะหลอมรวมกับเยื่อหุ้ม เซลล์ของเซลล์ประสาทก่อนไซแนปส์โดยสุ่ม ในกระบวนการที่ควบคุมโดย โปรตีน SNAREการหลอมรวมส่งผลให้โมเลกุลอะเซทิลโคลีน 7,000–10,000 โมเลกุลใน ถุง ไซแนปส์ถูกปล่อยออกสู่ช่องว่าง ไซแนปส์ ซึ่ง เป็นกระบวนการที่เรียกว่าเอ็กโซไซโทซิส [ 7 ] ดังนั้นเอ็กโซไซโทซิสจึงปล่อยอะเซทิลโคลีนออกมาเป็นกลุ่มๆ ที่เรียกว่าควอนตา ควอนตัมอะเซทิลโคลีนจะแพร่กระจายผ่านโครงข่ายของอะเซทิลโคลีนเอสเตอเรส ซึ่งความเข้มข้นของสารส่งสัญญาณในบริเวณนั้นจะสูง ทำให้อะเซทิลโคลีนเข้าไปจับกับตำแหน่งการจับทั้งหมดบนเอนไซม์ในเส้นทางของมัน อะเซทิลโคลีนที่ไปถึงแผ่นปลายประสาทจะกระตุ้นตัวรับอะเซทิลโคลีนประมาณ 2,000 ตัว ทำให้ช่องไอออนเปิดออก ซึ่งอนุญาตให้ไอออนโซเดียมเคลื่อนที่เข้าไปในแผ่นปลายประสาท ทำให้เกิดการลดขั้วประมาณ 0.5 mV ซึ่งเรียกว่าศักย์แผ่นปลายประสาทขนาดเล็ก (MEPP) เมื่ออะเซทิลโคลีนถูกปล่อยออกจากตัวรับ อะเซทิลโคลีนเอสเทอเรสจะทำลาย ACh ที่จับอยู่กับตัวรับนั้นแล้ว ซึ่งใช้เวลาประมาณ ~0.16 มิลลิวินาที ดังนั้นจึงพร้อมที่จะทำลาย ACh ที่ถูกปล่อยออกจากตัวรับ
เมื่อเส้นประสาทสั่งการถูกกระตุ้น จะมีความล่าช้าเพียง 0.5 ถึง 0.8 มิลลิวินาที ระหว่างการมาถึงของกระแสประสาทที่ปลายประสาทสั่งการและการตอบสนองครั้งแรกของแผ่นปลายประสาท[ 8 ] การมาถึงของ ศักยภาพการกระทำ ของเส้นประสาท สั่งการ ที่ปลายประสาทก่อนซินแนปส์จะเปิดช่องแคลเซียมที่ขึ้นอยู่กับแรงดันไฟฟ้าและไอออน Ca 2+ จะไหลจากของเหลวนอกเซลล์เข้าสู่ไซโตโซล ของเซลล์ประสาทก่อนซินแนปส์ การไหลเข้าของ Ca 2+ นี้ทำให้ เวสิเคิลที่มีสารสื่อประสาทหลายร้อย เวสิเคิล รวมตัวกับเยื่อหุ้มเซลล์ของเซลล์ประสาทก่อนซินแนปส์ผ่าน โปรตีน SNAREเพื่อปล่อยควอนตัมอะเซทิลโคลีนโดยเอ็กโซไซโทซิส การลดขั้วของแผ่นปลายประสาทโดยอะเซทิลโคลีนที่ปล่อยออกมาเรียกว่าศักยภาพแผ่นปลายประสาท (EPP) EPP เกิดขึ้นเมื่อ ACh จับกับตัวรับอะเซทิลโคลีนนิโคตินิก (nAChR) ที่แผ่นปลายประสาทสั่งการ และทำให้เกิดการไหลเข้าของไอออนโซเดียม การไหลเข้าของไอออนโซเดียมนี้ก่อให้เกิด EPP (การลดขั้ว) และกระตุ้นศักยภาพการกระทำที่เดินทางไปตามซาร์โคเลมมาและเข้าไปในเส้นใยกล้ามเนื้อผ่านท่อ T (ท่อตามขวาง) โดยอาศัยช่องโซเดียมที่ควบคุมด้วยแรงดันไฟฟ้า[ 9 ]การนำศักยภาพการกระทำไปตามท่อ T กระตุ้นการเปิดช่อง Ca 2+ ที่ควบคุมด้วยแรงดันไฟฟ้า ซึ่งเชื่อมต่อทางกลกับช่องปล่อย Ca 2+ในซาร์โคพลาสมิกเรติคูลัม[ 10 ] จากนั้น Ca 2+จะแพร่กระจายออกจากซาร์โคพลาสมิกเรติคูลัมไปยังไมโอไฟบริลเพื่อกระตุ้นการหดตัว ดังนั้นศักยภาพของแผ่นปลายจึงมีหน้าที่ในการสร้างศักยภาพการกระทำในเส้นใยกล้ามเนื้อซึ่งกระตุ้นการหดตัวของกล้ามเนื้อ การส่งผ่านจากเส้นประสาทไปยังกล้ามเนื้อนั้นรวดเร็วมากเพราะอะเซทิลโคลีนแต่ละควอนตัมไปถึงแผ่นปลายในความเข้มข้นระดับมิลลิโมลาร์ ซึ่งสูงพอที่จะรวมกับตัวรับที่มีความสัมพันธ์ต่ำ จากนั้นจึงปล่อยตัวส่งสัญญาณที่จับอยู่อย่างรวดเร็ว
ตัวรับอะเซทิลโคลีน

- ตัวรับที่เชื่อมโยงกับช่องไอออน
- ไอออน
- สารยึดเกาะ (เช่นอะเซทิลโคลีน )
อะเซทิลโคลีนเป็นสารสื่อประสาทที่สังเคราะห์จากโคลีน ในอาหาร และอะเซทิล-โคเอ (ACoA) และมีส่วนเกี่ยวข้องกับการกระตุ้นเนื้อเยื่อกล้ามเนื้อในสัตว์มีกระดูกสันหลังและ สัตว์ ไม่มีกระดูกสันหลัง บางชนิด ในสัตว์มีกระดูกสันหลัง ตัวรับอะเซทิลโคลีนชนิดย่อยที่พบที่รอยต่อประสาทกล้ามเนื้อของกล้ามเนื้อโครง ร่าง คือตัวรับอะเซทิลโคลีนนิโคตินิก (nAChR) ซึ่งเป็นช่องไอออนที่ควบคุมโดยลิแกนด์แต่ละหน่วยย่อยของตัวรับนี้มี "cys-loop" ที่เป็นลักษณะเฉพาะ ซึ่งประกอบด้วย กรด อะมิโนซิสทีน หนึ่ง ตัว ตามด้วย กรด อะมิโน 13 ตัว และกรดอะมิโนซิสทีนอีกหนึ่งตัว กรดอะมิโนซิสทีนสองตัวนี้สร้างพันธะไดซัลไฟด์ทำให้เกิดตัวรับ "cys-loop" ที่สามารถจับกับอะเซทิลโคลีนและลิแกนด์อื่นๆ ได้ ตัวรับ cys-loop เหล่านี้พบเฉพาะในยูคาริโอต เท่านั้น แต่โปรคาริโอตก็มีตัวรับ ACh ที่มีคุณสมบัติคล้ายกัน[ 5 ] ไม่ใช่ทุกสายพันธุ์ที่ใช้ จุดเชื่อมต่อประสาทกล้ามเนื้อ โคลินเนอร์จิกเช่นกุ้งเครย์ฟิชและแมลงวันผลไม้มีจุดเชื่อมต่อประสาทกล้ามเนื้อกลูตาเมอร์ จิก [ 4 ]
AChR ที่จุดเชื่อมต่อประสาทกล้ามเนื้อโครงร่าง ก่อตัวเป็น เฮเทอโรเพนตาเมอร์ที่ประกอบด้วยซับยูนิต α สองตัว β หนึ่งตัว ɛ หนึ่งตัว และ δ หนึ่งตัว[ 11 ] เมื่อลิแกนด์ ACh เพียงตัวเดียวจับกับซับยูนิต α ตัวใดตัวหนึ่งของตัวรับ ACh มันจะเหนี่ยวนำให้เกิดการเปลี่ยนแปลงโครงสร้างที่ส่วนต่อประสานกับซับยูนิต α ตัวที่สองของ AChR การเปลี่ยนแปลงโครงสร้างนี้ส่งผลให้ความสัมพันธ์ของซับยูนิต α ตัวที่สองกับลิแกนด์ ACh ตัวที่สองเพิ่มขึ้น ดังนั้น AChR จึงแสดงเส้นโค้งการแยกตัวแบบซิกมอยด์เนื่องจากการจับแบบร่วมมือกันนี้[ 5 ] การมีอยู่ของโครงสร้างตัวรับระดับกลางที่ไม่ทำงานซึ่งมีลิแกนด์ที่จับเพียงตัวเดียวทำให้ ACh อยู่ในไซแนปส์ซึ่งอาจสูญหายไปจาก การไฮโดรไลซิสของ โคลีนเอสเตอเรสหรือการแพร่กระจาย การคงอยู่ของลิแกนด์ ACh เหล่านี้ในไซแนปส์สามารถทำให้เกิดการตอบสนองหลังไซแนปส์ที่ยาวนานขึ้น[ 12 ]
การพัฒนา
การพัฒนาจุดเชื่อมต่อประสาทกล้ามเนื้อต้องอาศัยการส่งสัญญาณจากทั้งปลายประสาทสั่งการและบริเวณส่วนกลางของเซลล์กล้ามเนื้อ ในระหว่างการพัฒนา เซลล์กล้ามเนื้อจะสร้างตัวรับอะเซทิลโคลีน (AChRs) และแสดงออกในบริเวณส่วนกลางในกระบวนการที่เรียกว่าการสร้างรูปแบบเบื้องต้น อะกรินซึ่งเป็นเฮพารินโปรตีโอไกลแคนและไคเนส MuSK เชื่อว่าช่วยทำให้การสะสมของ AChR ในบริเวณส่วนกลางของไมโอไซต์มีเสถียรภาพ MuSK เป็นตัวรับไทโรซีนไคเนสซึ่งหมายความว่ามันกระตุ้นการส่งสัญญาณของเซลล์โดยการจับ โมเลกุล ฟอสเฟตกับบริเวณของตัวเอง เช่นไทโรซีนและเป้าหมายอื่นๆ ในไซโตพลาสซึม [ 13 ] เมื่อถูกกระตุ้นโดยลิแกนด์อะกริน MuSK จะส่งสัญญาณผ่านโปรตีนสองชนิดที่เรียกว่า " Dok-7 " และ " rapsyn " เพื่อกระตุ้นให้เกิด "การรวมกลุ่ม" ของตัวรับอะเซทิลโคลีน[ 14 ]การปล่อย ACh โดยเซลล์ประสาทสั่งการที่กำลังพัฒนาทำให้เกิดศักยภาพหลังไซแนปส์ในเซลล์กล้ามเนื้อซึ่งเสริมแรงเชิงบวกต่อการกำหนดตำแหน่งและเสถียรภาพของจุดเชื่อมต่อประสาทกล้ามเนื้อที่กำลังพัฒนา[ 15 ]
ผลการค้นพบเหล่านี้ได้รับการพิสูจน์บางส่วนโดยการศึกษา " น็อคเอาต์ " ของหนู ในหนูที่ขาดอะกรินหรือมิวเอสเค การเชื่อมต่อประสาทกล้ามเนื้อจะไม่เกิดขึ้น นอกจากนี้ หนูที่ขาดโดก-7ก็ไม่สร้างคลัสเตอร์ตัวรับอะเซทิลโคลีนหรือไซแนปส์ประสาทกล้ามเนื้อ[ 16 ]
การพัฒนาจุดเชื่อมต่อประสาทกล้ามเนื้อส่วนใหญ่ศึกษาในสิ่งมีชีวิตจำลอง เช่น สัตว์ฟันแทะ นอกจากนี้ ในปี 2558 ยังมีการสร้างจุดเชื่อมต่อประสาทกล้ามเนื้อของมนุษย์ขึ้นในหลอดทดลองโดยใช้เซลล์ต้นกำเนิดตัวอ่อน ของมนุษย์ และเซลล์ต้นกำเนิดกล้ามเนื้อร่างกาย[ 17 ] ในแบบจำลองนี้เซลล์ประสาท สั่งการก่อนซินแนปส์ จะถูกกระตุ้นด้วยออปโตเจเนติกส์และเส้นใยกล้ามเนื้อที่เชื่อมต่อกันทางซินแนปส์จะกระตุกเมื่อได้รับการกระตุ้นด้วยแสง
วิธีการวิจัย
โฮเซ่ เดล กัสติลโล และเบอร์นาร์ด แคทซ์ใช้เทคนิคไอโอโนโฟเรซิสเพื่อกำหนดตำแหน่งและความหนาแน่นของตัวรับอะเซทิลโคลีนชนิดนิโคตินิก (nAChRs) ที่บริเวณรอยต่อประสาทกล้ามเนื้อ โดยใช้เทคนิคนี้ ไมโครอิเล็กโทรดจะถูกวางไว้ภายในปลายประสาทสั่งการของเส้นใยกล้ามเนื้อ และไมโครปิเปตที่บรรจุอะเซทิลโคลีน (ACh) จะถูกวางไว้ตรงหน้าปลายประสาทสั่งการในช่องว่างไซแนปส์ จากนั้นจะใช้แรงดันไฟฟ้าบวกกับปลายไมโครปิเปต ซึ่งจะทำให้โมเลกุล ACh ที่มีประจุบวกถูกปล่อยออกมาจากปิเปตอย่างรวดเร็ว โมเลกุลเหล่านี้จะไหลเข้าไปในช่องว่างไซแนปส์และจับกับ AChRs ไมโครอิเล็กโทรดภายในเซลล์จะตรวจสอบขนาดของการเปลี่ยนแปลง ศักย์ไฟฟ้า ของปลายประสาทสั่งการเพื่อตอบสนองต่อการจับกันของ ACh กับตัวรับนิโคตินิก (ไอโอโนโทรปิก) Katz และ del Castillo แสดงให้เห็นว่าแอมพลิจูดของการลดขั้ว ( ศักยภาพหลังไซแนปส์กระตุ้น ) ขึ้นอยู่กับความใกล้ชิดของไมโครปิเปตที่ปล่อยไอออน ACh ไปยังแผ่นปลายประสาท ยิ่งไมโครปิเปตอยู่ห่างจากแผ่นปลายประสาทสั่งการมากเท่าใด การลดขั้วในเส้นใยกล้ามเนื้อก็จะยิ่งน้อยลงเท่านั้น ซึ่งทำให้ผู้วิจัยสามารถระบุได้ว่าตัวรับนิโคตินิกอยู่บริเวณแผ่นปลายประสาทสั่งการในความหนาแน่นสูง[ 4 ] [ 5 ]
สารพิษยังใช้เพื่อกำหนดตำแหน่งของตัวรับอะเซทิลโคลีนที่จุดเชื่อมต่อประสาทกล้ามเนื้อα-Bungarotoxinเป็นสารพิษที่พบในงูสายพันธุ์Bungarus multicinctusซึ่งทำหน้าที่เป็นตัวต้าน ACh และจับกับ AChRs อย่างถาวร โดยการเชื่อมต่อเอนไซม์ที่สามารถวิเคราะห์ได้ เช่นฮอร์สแรดิชเปอร์ออกซิเดส (HRP) หรือโปรตีนเรืองแสง เช่นโปรตีนเรืองแสงสีเขียว (GFP) เข้ากับ α-bungarotoxin ทำให้สามารถมองเห็นและวัดปริมาณ AChRs ได้[ 4 ]
สารพิษที่ส่งผลต่อจุดเชื่อมต่อระหว่างเส้นประสาทและกล้ามเนื้อ
แก๊สพิษทำลายประสาท
ก๊าซพิษต่อระบบประสาทจะจับกับและฟอสโฟรีเลตเอนไซม์ AChE ทำให้เอนไซม์นี้ไม่ทำงาน การสะสมของ ACh ภายในช่องว่างไซแนปส์ทำให้เซลล์กล้ามเนื้อหดตัวอย่างต่อเนื่อง นำไปสู่ภาวะแทรกซ้อนร้ายแรง เช่น อัมพาตและเสียชีวิตภายในไม่กี่นาทีหลังได้รับสารพิษ

สารพิษโบทูลินัม
สารพิษโบทูลินัม (หรือที่รู้จักกันในชื่อ สาร พิษโบ ทูลินัม ต่อระบบประสาท และจำหน่ายในเชิงพาณิชย์ภายใต้ชื่อทางการค้า Botox) ยับยั้งการปล่อยอะเซทิลโคลีนที่จุดเชื่อมต่อประสาทกล้ามเนื้อโดยการรบกวนโปรตีน SNARE [ 4 ]สารพิษนี้จะผ่านเข้าไปในปลายประสาทโดยกระบวนการเอนโดไซโทซิสและจากนั้นจะตัดโปรตีน SNARE ป้องกันไม่ให้ถุง ACh รวมตัวกับเยื่อหุ้มเซลล์ภายใน ซึ่งทำให้เกิดอัมพาตอ่อนแรง ชั่วคราว และการทำลายเส้นประสาททางเคมีเฉพาะที่ในกล้ามเนื้อลายที่ได้รับผลกระทบ การยับยั้งการปล่อย ACh จะไม่เกิดขึ้นจนกว่าจะผ่านไปประมาณสองสัปดาห์หลังจากการฉีด สามเดือนหลังจากที่การยับยั้งเกิดขึ้น กิจกรรมของเซลล์ประสาทจะเริ่มฟื้นคืนบางส่วน และหกเดือนหลังจากนั้น การทำงานของเซลล์ประสาทจะฟื้นคืนอย่างสมบูรณ์[ 18 ]
สารพิษบาดทะยัก
สารพิษบาดทะยัก หรือที่รู้จักกันในชื่อเททาโนสปาสมินเป็นสารพิษต่อระบบประสาทที่มีฤทธิ์รุนแรง ผลิตโดยแบคทีเรีย Clostridium tetaniและเป็นสาเหตุของโรคบาดทะยัก ค่า LD50 ของสารพิษนี้วัดได้ประมาณ 1 นาโนกรัมต่อกิโลกรัม ทำให้เป็นสารพิษที่อันตรายถึงชีวิตเป็นอันดับสองรองจากสารพิษโบทูลินัมชนิดดี มันทำงานคล้ายกับสารพิษโบทูลินัมโดยการเกาะติดและดูดซึมเข้าสู่ปลายประสาทก่อนซิแนปส์และรบกวนโปรตีน SNARE มันแตกต่างจากสารพิษโบทูลินัมในหลายๆ ด้าน ที่เห็นได้ชัดที่สุดคือผลลัพธ์สุดท้าย ซึ่งเททาโนสปาสมินทำให้เกิดอัมพาตเกร็งในขณะที่สารพิษโบทูลินัมทำให้เกิดอัมพาตอ่อนแรง
ลาโทรท็อกซิน
ลาโทรทอกซิน (α-Latrotoxin) ที่พบในพิษของแมงมุมแม่ม่ายดำ มีผลต่อจุดเชื่อมต่อระหว่างเส้นประสาทและกล้ามเนื้อโดยทำให้เกิดการปล่อยอะเซทิลโคลีนจากเซลล์ก่อนซินแนปส์ กลไกการออกฤทธิ์รวมถึงการจับกับตัวรับบนเซลล์ก่อนซินแนปส์ กระตุ้นวิถี IP3/DAGและการปล่อยแคลเซียมจากแหล่งเก็บภายในเซลล์และการสร้างรูพรุนส่งผลให้ไอออนแคลเซียมไหลเข้าโดยตรง กลไกใดกลไกหนึ่งทำให้แคลเซียมในเซลล์ก่อนซินแนปส์เพิ่มขึ้น ซึ่งนำไปสู่การปล่อยถุงซินแนปส์ของอะเซทิลโคลีน ลาโทรทอกซินทำให้เกิดอาการปวด กล้ามเนื้อหดตัว และหากไม่ได้รับการรักษาอาจทำให้เป็นอัมพาตและเสียชีวิตได้
พิษงู
พิษงูทำหน้าที่เป็นสารพิษที่จุดเชื่อมต่อประสาทกล้ามเนื้อและสามารถทำให้เกิดอาการอ่อนแรงและเป็นอัมพาตได้พิษสามารถทำหน้าที่เป็นสารพิษต่อระบบประสาททั้งก่อนและหลังไซแนปส์ได้[ 19 ]
สารพิษต่อระบบประสาทก่อนไซแนปส์ ซึ่งโดยทั่วไปเรียกว่า β-neurotoxins มีผลต่อบริเวณก่อนไซแนปส์ของจุดเชื่อมต่อประสาทกล้ามเนื้อ สารพิษต่อระบบประสาทเหล่านี้ส่วนใหญ่ออกฤทธิ์โดยการยับยั้งการปล่อยสารสื่อประสาท เช่น อะเซทิลโคลีน เข้าสู่ไซแนปส์ระหว่างเซลล์ประสาท อย่างไรก็ตาม สารพิษบางชนิดก็เป็นที่ทราบกันดีว่าช่วยเพิ่มการปล่อยสารสื่อประสาท สารพิษที่ยับยั้งการปล่อยสารสื่อประสาทจะทำให้เกิดการปิดกั้นการทำงานของระบบประสาทกล้ามเนื้อซึ่งป้องกันไม่ให้โมเลกุลส่งสัญญาณไปถึงตัวรับเป้าหมายหลังไซแนปส์ ส่งผลให้ผู้ที่ถูกงูกัดมีอาการอ่อนแรงอย่างรุนแรง สารพิษต่อระบบประสาทเหล่านี้ไม่ตอบสนองต่อเซรุ่มแก้พิษงูได้ดี หลังจากฉีดสารพิษเหล่านี้เข้าไปหนึ่งชั่วโมง รวมถึงโนเท็กซินและไทพอกซินปลายประสาทที่ได้รับผลกระทบจำนวนมากจะแสดงสัญญาณของความเสียหายทางกายภาพที่ไม่สามารถย้อนกลับได้ ทำให้ไม่มีถุงไซแนปส์เหลือ อยู่ [ 19 ]
สารพิษต่อระบบประสาทหลังไซแนปส์ หรือที่รู้จักกันในชื่อ α-neurotoxins ออกฤทธิ์ตรงข้ามกับสารพิษต่อระบบประสาทก่อนไซแนปส์ โดยการจับกับตัวรับอะเซทิลโคลีนที่เซลล์หลังไซแนปส์ ซึ่งจะป้องกันการปฏิสัมพันธ์ระหว่างอะเซทิลโคลีนที่ปล่อยออกมาจากปลายประสาทก่อนไซแนปส์กับตัวรับบนเซลล์หลังไซแนปส์ โดยมีผลทำให้การเปิดช่องโซเดียมที่เกี่ยวข้องกับตัวรับอะเซทิลโคลีนเหล่านี้ถูกยับยั้ง ส่งผลให้เกิดการปิดกั้นการทำงานของระบบประสาทและกล้ามเนื้อ คล้ายกับผลที่พบจากสารพิษต่อระบบประสาทก่อนไซแนปส์ ซึ่งทำให้กล้ามเนื้อที่เกี่ยวข้องกับจุดเชื่อมต่อที่ได้รับผลกระทบเป็นอัมพาต แต่ต่างจากสารพิษต่อระบบประสาทก่อนไซแนปส์ สารพิษหลังไซแนปส์จะได้รับผลกระทบจากเซรุ่มแก้พิษได้ง่ายกว่า เซรุ่มแก้พิษจะเร่งการแยกตัวของสารพิษออกจากตัวรับ ทำให้ในที่สุดอาการอัมพาตกลับคืนสู่ปกติ สารพิษต่อระบบประสาทเหล่านี้ช่วยในการศึกษาความหนาแน่นและการหมุนเวียน ของตัวรับอะเซทิลโคลีนทั้งในเชิงทดลองและเชิงคุณภาพ รวมถึงการศึกษาที่สังเกตทิศทางของแอนติบอดีไปยังตัวรับอะเซทิลโคลีนที่ได้รับผลกระทบในผู้ป่วยที่ได้รับการวินิจฉัยว่าเป็นโรคกล้ามเนื้ออ่อนแรง[ 19 ]
โรคต่างๆ
โรคใด ๆที่ส่งผลกระทบต่อการส่งสัญญาณประสาทระหว่างเซลล์ประสาทสั่งการและเซลล์กล้ามเนื้อ จะถูกจัดอยู่ในกลุ่มโรคระบบประสาทและกล้ามเนื้อโรคเหล่านี้อาจเป็นกรรมพันธุ์หรือเกิดขึ้นภายหลัง และมีความรุนแรงและอัตราการเสียชีวิตแตกต่างกันไป โดยทั่วไปแล้ว โรคเหล่านี้ส่วนใหญ่มักเกิดจากการกลายพันธุ์หรือความผิดปกติของระบบภูมิคุ้มกัน ความผิดปกติของระบบภูมิคุ้มกันในกรณีของโรคระบบประสาทและกล้ามเนื้อ มักเกิดจากกลไกที่เกี่ยวข้อง กับสารน้ำใน ร่างกาย โดยเฉพาะ เซลล์บีและส่งผลให้มี การสร้าง แอนติบอดีที่ไม่เหมาะสมต่อโปรตีนในเซลล์ประสาทสั่งการหรือเส้นใยกล้ามเนื้อ ซึ่งจะไปรบกวนการส่งสัญญาณประสาทหรือการส่งสัญญาณผ่านไซแนปส์
ภูมิคุ้มกันตนเอง
โรคกล้ามเนื้ออ่อนแรง
โรคกล้ามเนื้ออ่อนแรงชนิด ไมแอสทีเนียกราวิสเป็นโรคภูมิต้านตนเองที่ร่างกายสร้างแอนติบอดีต่อตัวรับอะเซทิลโคลีน (AchR) (ใน 80% ของกรณี) หรือต่อไคเนสเฉพาะกล้ามเนื้อหลังไซแนปส์ (MuSK) (0–10% ของกรณี) ในโรคกล้ามเนื้ออ่อนแรงชนิดไมแอสทีเนียกราวิสที่ไม่มีแอนติบอดี โปรตีนที่เกี่ยวข้องกับตัวรับไล โปโปรตีนความหนาแน่นต่ำ 4 จะถูกกำหนดเป้าหมายโดยIgG1ซึ่งทำหน้าที่เป็นตัวยับยั้งการแข่งขันของลิแกนด์ ป้องกันไม่ให้ลิแกนด์จับกับตัวรับ ไม่ทราบว่าโรคกล้ามเนื้ออ่อนแรงชนิดไมแอสทีเนียกราวิสที่ไม่มีแอนติบอดีจะตอบสนองต่อการรักษามาตรฐานหรือไม่[ 20 ]
MG ในทารกแรกเกิด
โรค MG ในทารกแรกเกิดเป็นความผิดปกติของระบบภูมิคุ้มกันที่ส่งผลกระทบต่อเด็ก 1 ใน 8 คนที่เกิดจากมารดาที่ได้รับการวินิจฉัยว่าเป็นโรคกล้ามเนื้ออ่อนแรง (MG) โรค MG สามารถถ่ายทอดจากมารดาไปยังทารกในครรภ์ได้โดยการเคลื่อนตัวของแอนติบอดี AChR ผ่านทางรกอาการของโรคนี้เมื่อแรกเกิด ได้แก่ อาการอ่อนแรง ซึ่งตอบสนองต่อยาต้านเอนไซม์โคลีนเอสเตอเรส รวมถึงภาวะทารกในครรภ์เคลื่อนไหวช้า หรือไม่มีการเคลื่อนไหว รูปแบบของโรคนี้เป็นแบบชั่วคราว โดยมีระยะเวลาประมาณสามเดือน อย่างไรก็ตาม ในบางกรณี โรค MG ในทารกแรกเกิดอาจนำไปสู่ผลกระทบต่อสุขภาพอื่นๆ เช่น ภาวะข้อติดแข็ง และอาจถึงขั้นเสียชีวิตของทารกในครรภ์ได้ เชื่อกันว่าภาวะเหล่านี้เริ่มต้นขึ้นเมื่อแอนติบอดี AChR ของมารดาไปจับกับ AChR ของ ทารกในครรภ์ และอาจคงอยู่จนถึงสัปดาห์ที่ 33 ของการตั้งครรภ์เมื่อหน่วยย่อย γ ของ AChR ถูกแทนที่ด้วยหน่วยย่อย ε [ 21 ] [ 22 ]
กลุ่มอาการกล้ามเนื้ออ่อนแรงแลมเบิร์ต-อีตัน
กลุ่มอาการกล้ามเนื้ออ่อนแรงแลมเบิร์ต-อีตัน (LEMS) เป็นความผิดปกติของระบบภูมิคุ้มกันที่ส่งผลต่อส่วนก่อนไซแนปส์ของจุดเชื่อมต่อประสาทกล้ามเนื้อ โรคหายากนี้มีลักษณะเฉพาะคืออาการสามอย่าง ได้แก่ กล้ามเนื้ออ่อนแรงส่วน ต้น ความผิดปกติของระบบประสาทอัตโนมัติและภาวะไม่มีรีเฟล็กซ์[ 23 ]กล้ามเนื้ออ่อนแรงส่วนต้นเป็นผลมาจาก แอนติบอดีก่อ โรคที่มุ่งเป้าไปที่ช่องแคลเซียมแบบ P/Q ที่ควบคุมด้วยแรงดันไฟฟ้า ซึ่งส่งผลให้การปล่อยอะเซทิลโคลีนจากปลายประสาทสั่งการบนเซลล์ก่อนไซแนปส์ลดลง ตัวอย่างของความผิดปกติของระบบประสาทอัตโนมัติที่เกิดจาก LEMS ได้แก่ภาวะหย่อนสมรรถภาพทางเพศในผู้ชายท้องผูกและที่พบบ่อยที่สุดคือปากแห้งความผิดปกติที่พบน้อยกว่า ได้แก่ตาแห้งและเหงื่อ ออก ผิด ปกติ ภาวะไม่มีรีเฟล็กซ์คือภาวะที่รีเฟล็กซ์ของเอ็นลดลง และอาจบรรเทาลงชั่วคราวหลังจากออกกำลังกายเป็นระยะเวลาหนึ่ง[ 24 ]
50–60% ของผู้ป่วยที่ได้รับการวินิจฉัยว่าเป็น LEMS มักมีเนื้องอก ร่วมด้วย ซึ่งโดยทั่วไปคือมะเร็งปอดชนิดเซลล์เล็ก (SCLC) เนื้องอกชนิดนี้ยังแสดงออกถึงช่องแคลเซียมที่ควบคุมด้วยแรงดันไฟฟ้าด้วย[ 24 ]บ่อยครั้งที่ LEMS เกิดขึ้นร่วมกับโรคกล้ามเนื้ออ่อนแรง[ 23 ]
การรักษา LEMS ประกอบด้วยการใช้ 3,4-ไดอะมิโนไพริดีนเป็นมาตรการแรก ซึ่งช่วยเพิ่มศักยภาพการทำงานของกล้ามเนื้อโดยรวมและเพิ่มความแข็งแรงของกล้ามเนื้อโดยการยืดระยะเวลาที่ช่องแคลเซียมที่ควบคุมด้วยแรงดันไฟฟ้ายังคงเปิดอยู่หลังจากปิดกั้นช่องโพแทสเซียมที่ควบคุมด้วยแรงดันไฟฟ้า ในสหรัฐอเมริกา การรักษาด้วย 3,4-ไดอะมิโนไพริดีนสำหรับผู้ป่วย LEMS ที่มีคุณสมบัติเหมาะสมนั้นไม่มีค่าใช้จ่ายภายใต้โครงการขยายการเข้าถึง[ 25 ] [ 26 ]การรักษาเพิ่มเติมรวมถึงการใช้เพรดนิโซนและอะซาไธโอพรีนในกรณีที่ 3,4-ไดอะมิโนไพริดีนไม่ได้ช่วยในการรักษา[ 24 ]
นิวโรไมโอโทเนีย
โรคกล้ามเนื้อกระตุก ( Neuromyotoniaหรือ NMT) หรือที่รู้จักกันในชื่อกลุ่มอาการไอแซค (Isaac's syndrome) แตกต่างจากโรคอื่นๆ ที่เกิดขึ้นที่จุดเชื่อมต่อระหว่างเส้นประสาทและกล้ามเนื้อ แทนที่จะทำให้กล้ามเนื้ออ่อนแรง NMT กลับทำให้เกิดการกระตุ้นมากเกินไปของเส้นประสาทสั่งการ NMT ทำให้เกิดการกระตุ้นมากเกินไปนี้โดยการสร้างการโพลาไรเซชันที่ยาวนานขึ้นโดยการลดการทำงานของช่องโพแทสเซียมที่ควบคุมด้วยแรงดันไฟฟ้าซึ่งทำให้มีการปล่อยสารสื่อประสาทมากขึ้นและการยิงซ้ำๆ อัตราการยิงที่เพิ่มขึ้นนี้ทำให้เกิดการส่งสัญญาณที่มากขึ้น และส่งผลให้กล้ามเนื้อของผู้ป่วยมีการทำงานมากขึ้น นอกจากนี้ยังเชื่อว่า NMT มี ต้นกำเนิด จากภูมิคุ้มกันบกพร่องเนื่องจากมีความเกี่ยวข้องกับอาการภูมิคุ้มกันบกพร่องในผู้ป่วย[ 21 ]
พันธุกรรม
กลุ่มอาการกล้ามเนื้ออ่อนแรงแต่กำเนิด
กลุ่มอาการกล้ามเนื้ออ่อนแรงแต่กำเนิด (CMS) มีลักษณะการทำงานคล้ายคลึงกับทั้ง MG และ LEMS มาก แต่ความแตกต่างหลักระหว่าง CMS กับโรคเหล่านั้นคือ CMS มีต้นกำเนิดทางพันธุกรรม โดยเฉพาะอย่างยิ่ง กลุ่มอาการเหล่านี้เป็นโรคที่เกิดจากการกลายพันธุ์ ซึ่งโดยทั่วไปแล้วเป็นการกลายพันธุ์แบบด้อย ใน ยีนอย่างน้อย 1 ใน 10 ยีนที่ส่งผลต่อโปรตีนก่อนไซแนปส์ ไซแนปส์ และหลังไซแนปส์ในจุดเชื่อมต่อประสาทกล้ามเนื้อ การกลายพันธุ์ดังกล่าวมักเกิดขึ้นในหน่วยย่อย ε ของ AChR [ 21 ]ซึ่งส่งผลต่อจลนศาสตร์และการแสดงออกของตัวรับเอง การแทนที่หรือการลบของนิวคลีโอไทด์เดี่ยวอาจทำให้สูญเสียการทำงานในหน่วยย่อย การกลายพันธุ์ อื่นๆ เช่น การกลายพันธุ์ที่ส่งผลต่ออะเซทิลโคลีนเอส เทอเรส และอะเซทิลทรานสเฟอเรสก็สามารถทำให้เกิดการแสดงออกของ CMS ได้เช่นกัน โดยเฉพาะอย่างยิ่งการกลายพันธุ์หลังเกี่ยวข้องกับภาวะหยุด หายใจ เป็น ช่วงๆ [ 27 ]กลุ่มอาการเหล่านี้สามารถแสดงอาการได้ในช่วงเวลาต่างๆ ในชีวิตของแต่ละบุคคล อาการเหล่านี้อาจเกิดขึ้นในช่วงระยะทารกในครรภ์ ทำให้เกิดภาวะกล้ามเนื้ออ่อนแรง ในทารกใน ครรภ์ หรือในช่วงปริกำเนิด ซึ่งอาจพบอาการบางอย่าง เช่นข้อติดแข็งเปลือกตาตก กล้ามเนื้ออ่อน แรงอัมพาตของกล้ามเนื้อตา และปัญหาในการกินหรือหายใจ นอกจากนี้ยังอาจเกิดขึ้นในช่วงวัยรุ่นหรือวัยผู้ใหญ่ ทำให้บุคคลนั้นเกิดอาการกลุ่มอาการช่องช้า[ 21 ]
การรักษา CMS ชนิดย่อยเฉพาะ (CMS ช่องสัญญาณเร็วหลังไซแนปส์) [ 28 ] [ 29 ]คล้ายกับการรักษาความผิดปกติของระบบประสาทและกล้ามเนื้ออื่นๆ3,4-ไดอะมิโนไพริดีนซึ่งเป็นการรักษาหลักสำหรับ LEMS กำลังอยู่ระหว่างการพัฒนาเป็นยารักษาโรคหายากสำหรับ CMS [ 30 ]ในสหรัฐอเมริกา และมีให้สำหรับผู้ป่วยที่มีคุณสมบัติเหมาะสมภายใต้โครงการขยายการเข้าถึงโดยไม่มีค่าใช้จ่าย[ 25 ] [ 26 ]
ดูเพิ่มเติม
อ่านเพิ่มเติม
- คันเดล ER , ชวาร์ตษ์ เจเอช, เจสเซล TM (2000) หลักวิทยาศาสตร์ประสาท (ฉบับที่ 4) นิวยอร์ก: แมคกรอว์-ฮิลล์ไอเอสบีเอ็น 0-8385-7701-6.
- Nicholls JG, Martin AR, Wallace BG, Fuchs PA (2001). จากเซลล์ประสาทสู่สมอง (ฉบับที่ 4). ซันเดอร์แลนด์, แมสซาชูเซตส์: Sinauer Associates. ISBN 0-87893-439-1.
- Engel AG (2004). Myology (ฉบับที่ 3). นิวยอร์ก: McGraw Hill Professional. ISBN 0-07-137180-X.
ลิงก์ภายนอก
- ภาพทางจุลพยาธิวิทยา: 21501lca – ระบบการเรียนรู้จุลพยาธิวิทยา มหาวิทยาลัยบอสตัน
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ จุดเชื่อมต่อประสาทกล้ามเนื้อ
จุด เชื่อมต่อประสาทกล้ามเนื้อ (หรือ จุดเชื่อมต่อกล้ามเนื้อและเส้นประสาท ) คือ ไซแนปส์ทางเคมี ระหว่าง เซลล์ประสาทสั่งการ และ เส้นใยกล้าม เนื้อ [ 1 ]
การส่งผ่านควอนตัม
ที่ จุดเชื่อมต่อประสาทกล้าม เนื้อ แอกซอนมอเตอร์ก่อนซิแนปส์จะสิ้นสุดลงที่ระยะ 30 นาโนเมตรจากเยื่อหุ้มเซลล์หรือ ซาร์โคเลมมา ของเส้นใยกล้ามเนื้อ ซาร์โคเลมมาที่จุดเชื่อมต่อมี ส่วนเว้า ที่เรียกว่า รอยพับหลังจุดเชื่อมต่อ...
ตัวรับอะเซทิลโคลีน
อะเซทิลโคลีน เป็น สารสื่อประสาท ที่สังเคราะห์จาก โคลีน ในอาหาร และ อะเซทิล-โคเอ (ACoA) และมีส่วนเกี่ยวข้องกับการกระตุ้น เนื้อเยื่อกล้ามเนื้อ ใน สัตว์มีกระดูกสันหลัง และ สัตว์ ไม่มีกระดูกสันหลัง บางชนิด ในสัตว์มีกระดูกสันหลัง...
การพัฒนา
การพัฒนาจุดเชื่อมต่อประสาทกล้ามเนื้อต้องอาศัยการส่งสัญญาณจากทั้งปลายประสาทสั่งการและบริเวณส่วนกลางของเซลล์กล้ามเนื้อ ในระหว่างการพัฒนา เซลล์กล้ามเนื้อจะสร้างตัวรับอะเซทิลโคลีน (AChRs) และแสดงออกในบริเวณส่วนกลางในกระบวนการที่เรียกว่าการสร้างรูปแบบเบื้องต้น อะ...