อ่าน 13 นาที
วิวัฒนาการของดวงตา
วิวัฒนาการ ของดวงตา คือต้นกำเนิดและการพัฒนาที่มีความหลากหลายของการคัดเลือกโดยธรรมชาติในช่วงเวลาทางธรณีวิทยาของอวัยวะที่ไวต่อแสงและการมองเห็นในสิ่งมีชีวิต...
วิวัฒนาการของดวงตา
วิวัฒนาการของดวงตาคือต้นกำเนิดและการพัฒนาที่มีความหลากหลายของการคัดเลือกโดยธรรมชาติในช่วงเวลาทางธรณีวิทยาของอวัยวะที่ไวต่อแสงและการมองเห็นในสิ่งมีชีวิต นักวิทยาศาสตร์หลายคนพบว่าวิวัฒนาการของดวงตาน่าสนใจที่จะศึกษาเพราะดวงตาเป็นตัวอย่างที่โดดเด่นของอวัยวะที่คล้ายคลึง กันซึ่งพบใน สัตว์หลาย ชนิด การตรวจจับแสงอย่างง่ายพบได้ในแบคทีเรีย สิ่งมีชีวิตเซลล์เดียว พืช และสัตว์ ดวงตาที่ซับซ้อนซึ่งสามารถสร้างภาพได้นั้นวิวัฒนาการขึ้นอย่างอิสระหลายครั้ง[ 1 ]
พบดวงตาที่หลากหลายจากBurgess ShaleในยุคแคมเบรียนตอนกลางและจากEmu Bay Shaleที่ เก่ากว่าเล็กน้อย [ 2 ] ดวงตามีความแตกต่างกันในด้านความคมชัดในการมองเห็นช่วงความยาวคลื่นที่สามารถตรวจจับได้ ความไวต่อแสง ความสามารถในการตรวจจับการเคลื่อนไหวหรือการแยกแยะวัตถุ และขอบเขตที่สามารถแยกแยะสีได้
ประวัติการวิจัย

ในปี ค.ศ. 1802 นักปรัชญาWilliam Paleyเรียกมันว่าปาฏิหาริย์แห่ง " การออกแบบ " ในปี ค.ศ. 1859 Charles Darwinเองได้เขียนไว้ในหนังสือ Origin of Species ของเขา ว่า วิวัฒนาการของดวงตาโดยการคัดเลือกโดยธรรมชาติ ดูเหมือนจะ "ไร้สาระอย่างที่สุด" ในตอนแรก[ 3 ] อย่างไรก็ตาม เขากล่าวต่อไปว่า แม้จะยากที่จะจินตนาการถึงวิวัฒนาการของมัน แต่วิวัฒนาการของมันก็เป็นไปได้โดยสมบูรณ์
...หากสามารถแสดงให้เห็นถึงการเปลี่ยนแปลงหลายระดับตั้งแต่ดวงตาที่เรียบง่ายและไม่สมบูรณ์ไปจนถึงดวงตาที่ซับซ้อนและสมบูรณ์แบบ โดยแต่ละระดับมีประโยชน์ต่อผู้ครอบครอง ดังที่เป็นเช่นนั้นแน่นอน และหากดวงตาเปลี่ยนแปลงอยู่เสมอและการเปลี่ยนแปลงนั้นได้รับการถ่ายทอดทางพันธุกรรม ดังที่เป็นเช่นนั้นแน่นอนเช่นกัน และหากการเปลี่ยนแปลงดังกล่าวมีประโยชน์ต่อสัตว์ใดๆ ภายใต้สภาวะชีวิตที่เปลี่ยนแปลงไป ความยากลำบากในการเชื่อว่าดวงตาที่สมบูรณ์แบบและซับซ้อนสามารถเกิดขึ้นได้จากการคัดเลือกโดยธรรมชาติ แม้ว่าจะเอาชนะไม่ได้ด้วยจินตนาการของเรา ก็ไม่ควรถูกมองว่าเป็นการล้มล้างทฤษฎี[ 3 ]
เขาเสนอวิวัฒนาการทีละขั้นจาก "เส้นประสาทตาที่เคลือบด้วยเม็ดสีเพียงอย่างเดียว และไม่มีกลไกอื่นใด" ไปสู่ "ขั้นความสมบูรณ์แบบที่ค่อนข้างสูง" และยกตัวอย่างขั้นกลางที่มีอยู่[ 3 ]การวิจัยในปัจจุบันกำลังตรวจสอบกลไกทางพันธุกรรมที่อยู่เบื้องหลังการพัฒนาและวิวัฒนาการของดวงตา[ 4 ]
อัตราการวิวัฒนาการ
ฟอสซิลดวงตาที่อาจพบเป็นครั้งแรกจนถึงปัจจุบันมาจาก ยุค เอเดียคารัน (ประมาณ555 ล้านปีก่อน ) [ 5 ]ในขณะที่ฟอสซิลดวงตาที่เก่าแก่ที่สุดที่ยืนยันได้คือ ฟอสซิล Schmidtiellus reetaeจาก 530 ล้านปีก่อน ซึ่งเก็บรวบรวมได้ในซาวิรานนาทางตอนเหนือของเอสโตเนีย โครงสร้างคล้ายกับดวงตารวมของแมลงปอและผึ้งในปัจจุบัน แต่มีออมมาทิเดีย (~100) ที่เว้นระยะห่างกันมากขึ้น และไม่มีเลนส์[ 6 ] [ 7 ] ยุคแคมเบรียนตอนต้นมีการวิวัฒนาการที่รวดเร็วอย่างเห็นได้ชัด เรียกว่า " การระเบิดของแคมเบรียน " หนึ่งในสมมติฐานมากมายเกี่ยวกับ "สาเหตุ" ของการระเบิดของแคมเบรียนคือทฤษฎี "สวิตช์ไฟ" ของแอนดรูว์ พาร์คเกอร์ซึ่งกล่าวว่าวิวัฒนาการของดวงตาขั้นสูงได้เริ่มต้นการแข่งขันที่เร่งวิวัฒนาการ[ 8 ]ก่อนการระเบิดของแคมเบรียน สัตว์อาจรับรู้แสงได้ แต่ไม่ได้ใช้แสงเพื่อการเคลื่อนที่อย่างรวดเร็วหรือการนำทางด้วยการมองเห็น
อัตราการวิวัฒนาการของดวงตานั้นประเมินได้ยาก เนื่องจากบันทึกฟอสซิล โดยเฉพาะอย่างยิ่งของยุคแคมเบรียนตอนต้นนั้นค่อนข้างจำกัด ความเร็วในการวิวัฒนาการของกลุ่มเซลล์รับแสงรูปวงกลมไปเป็นดวงตาของสัตว์มีกระดูกสันหลังที่ทำงานได้อย่างสมบูรณ์นั้น ได้รับการประเมินโดยอาศัยอัตราการกลายพันธุ์ ความได้เปรียบเชิงสัมพัทธ์ของสิ่งมีชีวิต และการคัดเลือกโดยธรรมชาติ อย่างไรก็ตาม เวลาที่จำเป็นสำหรับแต่ละสถานะนั้นถูกประเมินสูงเกินไปอย่างสม่ำเสมอ และเวลาของรุ่นถูกกำหนดไว้ที่หนึ่งปี ซึ่งเป็นเรื่องปกติในสัตว์ขนาดเล็ก แม้จะมีค่าที่มองโลกในแง่ร้ายเหล่านี้ ดวงตาของสัตว์มีกระดูกสันหลังก็ยังสามารถวิวัฒนาการจากกลุ่มเซลล์รับแสงได้ในเวลาไม่ถึง 364,000 ปี[ 9 ]
กำเนิดของดวงตา
ไม่ว่าดวงตาจะวิวัฒนาการเพียงครั้งเดียวหรือหลายครั้งนั้นขึ้นอยู่กับนิยามของดวงตา สัตว์ที่มีดวงตาทั้งหมดมีกลไกทางพันธุกรรมร่วมกันมากมายสำหรับการพัฒนาดวงตา ซึ่งบ่งชี้ว่าบรรพบุรุษของสัตว์ที่มีดวงตามีกลไกที่ไวต่อแสงบางรูปแบบ แม้ว่ามันจะไม่ใช่อวัยวะรับแสงโดยเฉพาะก็ตาม อย่างไรก็ตาม แม้แต่เซลล์รับแสงก็อาจวิวัฒนาการมากกว่าหนึ่งครั้งจากเซลล์รับสารเคมีที่มีโครงสร้างโมเลกุลคล้ายกัน เซลล์รับแสงอาจมีอยู่ก่อนการระเบิดของแคมเบรียนนานแล้ว[ 10 ]ความคล้ายคลึงกันในระดับที่สูงขึ้น เช่น การใช้โปรตีนคริสตัลลินในเลนส์ของเซฟาโลพอดและสัตว์มีกระดูกสันหลังที่แยกจากกัน[ 11 ]สะท้อนให้เห็นถึงการนำโปรตีนพื้นฐานมาใช้ในหน้าที่ใหม่ภายในดวงตา[ 12 ]
ลักษณะร่วมที่พบได้ในอวัยวะที่ไวต่อแสงทั้งหมดคือออปซิน ออปซินเป็นโปรตีนในกลุ่มที่ไวต่อแสงและแบ่งออกเป็นเก้ากลุ่ม ซึ่งมีอยู่แล้วในเออร์บิลาเทเรียนซึ่งเป็นบรรพบุรุษร่วมสุดท้ายของสัตว์ที่มีสมมาตรทวิภาคทั้งหมด[ 13 ]นอกจากนี้ชุดเครื่องมือทางพันธุกรรมสำหรับการกำหนดตำแหน่งของดวงตายังพบได้ในสัตว์ทุกชนิด: ยีน PAX6ควบคุมตำแหน่งที่ดวงตาพัฒนาในสัตว์ตั้งแต่ปลาหมึก[ 14 ]ไปจนถึงหนูและแมลงวันผลไม้[ 15 ] [ 16 ] [ 17 ]ยีนระดับสูงดังกล่าวโดยนัยแล้วมีอายุมากกว่าโครงสร้างหลายอย่างที่พวกมันควบคุมในปัจจุบัน พวกมันต้องมีจุดประสงค์ที่แตกต่างออกไปก่อนที่จะถูกนำมาใช้ในการพัฒนาดวงตา[ 12 ]
ดวงตาและอวัยวะรับสัมผัสอื่นๆ น่าจะวิวัฒนาการมาก่อนสมอง: ไม่จำเป็นต้องมีอวัยวะประมวลผลข้อมูล (สมอง) ก่อนที่จะมีข้อมูลให้ประมวลผล[ 18 ]ตัวอย่างที่ยังมีชีวิตอยู่คือ แมงกะพรุน คิวโบ โซแอน ที่มีดวงตาที่เทียบได้กับดวงตาแบบกล้องของ สัตว์ มีกระดูกสันหลังและเซฟาโลพอด แม้ว่าจะไม่มีสมองก็ตาม[ 19 ]
ขั้นตอนวิวัฒนาการ

บรรพบุรุษที่เก่าแก่ที่สุดของดวงตาคือโปรตีนรับแสงที่รับรู้แสง ซึ่งพบได้แม้ในสิ่งมีชีวิตเซลล์เดียว เรียกว่า " จุดรับแสง " [ 20 ]จุดรับแสงสามารถรับรู้ความสว่างโดยรอบเท่านั้น พวกมันสามารถแยกแยะแสงจากความมืด ซึ่งเพียงพอสำหรับการรับรู้แสงตามช่วงเวลาและการซิงโครไนซ์จังหวะชีวภาพ ในแต่ละวัน พวกมันไม่เพียงพอสำหรับการมองเห็น เนื่องจากไม่สามารถแยกแยะรูปร่างหรือกำหนดทิศทางของแสงได้ จุดรับแสงพบได้ในกลุ่มสัตว์หลักเกือบทั้งหมด และพบได้ทั่วไปในสิ่งมีชีวิตเซลล์เดียว รวมถึงยูเกลนาจุดรับแสงของยูเกลนา เรียกว่าสติ๊กมาตั้งอยู่ที่ปลายด้านหน้า มันเป็นจุดเล็กๆ ของเม็ดสีสีแดงที่บังกลุ่มของผลึกที่ไวต่อแสง เมื่อรวมกับแฟลเจล ลัมนำ จุดรับแสงช่วยให้สิ่งมีชีวิตเคลื่อนที่ตอบสนองต่อแสง มักจะเคลื่อนที่ไปทางแสงเพื่อช่วยในการสังเคราะห์แสง[ 21 ]และเพื่อทำนายกลางวันและกลางคืน ซึ่งเป็นหน้าที่หลักของจังหวะชีวภาพ เม็ดสีรับภาพตั้งอยู่ในสมองของสิ่งมีชีวิตที่ซับซ้อนกว่า และเชื่อกันว่ามีบทบาทในการประสานการวางไข่กับวัฏจักรของดวงจันทร์ โดยการตรวจจับการเปลี่ยนแปลงเล็กน้อยของแสงสว่างในเวลากลางคืน สิ่งมีชีวิตสามารถประสานการปล่อยอสุจิและไข่เพื่อเพิ่มโอกาสในการปฏิสนธิให้สูงสุด[ 22 ]
การมองเห็นนั้นอาศัยชีวเคมีพื้นฐานซึ่งเป็นเรื่องปกติในดวงตาทุกดวง อย่างไรก็ตาม วิธีการใช้ชุดเครื่องมือชีวเคมีนี้ในการตีความสภาพแวดล้อมของสิ่งมีชีวิตนั้นแตกต่างกันอย่างมาก ดวงตามีโครงสร้างและรูปแบบที่หลากหลาย ซึ่งทั้งหมดนี้วิวัฒนาการค่อนข้างช้าเมื่อเทียบกับโปรตีนและโมเลกุลพื้นฐาน[ 21 ]
ในระดับเซลล์ ดูเหมือนจะมีดวงตาสองประเภทหลัก ประเภทหนึ่งมีอยู่ในโปรโตสโตม ( หอย หนอนปล้องและสัตว์ขาปล้อง ) อีกประเภทหนึ่งมีอยู่ในดิวเทอโรสโตม ( สัตว์มีกระดูกสันหลังและสัตว์ทะเลที่มีหนาม ) [ 21 ]
หน่วยการทำงานของดวงตาคือเซลล์รับแสง ซึ่งประกอบด้วยโปรตีนออปซินและตอบสนองต่อแสงโดยการเริ่มต้นกระแสประสาท ออปซินที่ไวต่อแสงจะอยู่บนชั้นที่มีขน เพื่อเพิ่มพื้นที่ผิวให้มากที่สุด ลักษณะของ "ขน" เหล่านี้แตกต่างกัน โดยมีสองรูปแบบพื้นฐานที่อยู่เบื้องหลังโครงสร้างของเซลล์รับแสง ได้แก่ไมโครวิลลีและซีเลีย [ 23 ] ในดวงตาของโปรโตสโตม พวกมันคือไมโครวิลลี ซึ่งเป็นส่วนขยายหรือส่วนที่ยื่นออกมาของเยื่อหุ้มเซลล์ แต่ในดวงตาของดิวเทอโรสโตม พวกมันได้มาจากซีเลีย ซึ่งเป็นโครงสร้างที่แยกต่างหาก[ 21 ]อย่างไรก็ตาม นอกดวงตา สิ่งมีชีวิตอาจใช้เซลล์รับแสงประเภทอื่น ตัวอย่างเช่น หนอนหอยPlatynereis dumeriliiใช้เซลล์ไมโครวิลลีในดวงตา แต่ยังมีเซลล์รับแสงซีเลียในสมองส่วนลึกเพิ่มเติมอีกด้วย[ 24 ] การได้ มาซึ่งความจริงอาจซับซ้อนกว่านั้น เนื่องจากไมโครวิลลีบางส่วนมีร่องรอยของซีเลียอยู่ แต่การสังเกตอื่นๆ ดูเหมือนจะสนับสนุนความแตกต่างพื้นฐานระหว่างโปรโตสโตมและดิวเทอโรสโตม[ 21 ]ข้อพิจารณาเหล่านี้มุ่งเน้นไปที่การตอบสนองของเซลล์ต่อแสง บางชนิดใช้โซเดียมเพื่อสร้างสัญญาณไฟฟ้าที่จะก่อให้เกิดกระแสประสาท และบางชนิดใช้โพแทสเซียม นอกจากนี้ โปรโตสโตมโดยรวมสร้างสัญญาณโดยการปล่อยให้ โซเดียมผ่านผนังเซลล์ ได้มากขึ้นในขณะที่ดิวเทอโรสโตมปล่อยให้ผ่านได้น้อยกว่า[ 21 ]
นี่แสดงให้เห็นว่าเมื่อสายพันธุ์ทั้งสองแยกตัวออกจากกันในยุคพรีแคมเบรียน พวกมันมีตัวรับแสงที่ดั้งเดิมมากเท่านั้น ซึ่งพัฒนาไปเป็นดวงตาที่ซับซ้อนมากขึ้นโดยอิสระ
ดวงตาแรกเริ่ม
หน่วยประมวลผลแสงพื้นฐานของดวงตาคือเซลล์รับแสง ซึ่งเป็นเซลล์เฉพาะที่มีโมเลกุลสองชนิดที่ยึดติดกันและอยู่ในเยื่อหุ้มเซลล์ ได้แก่ออปซิน ซึ่งเป็น โปรตีนที่ไวต่อแสงและโครโมฟอร์ ซึ่ง เป็นเม็ดสีที่ดูดซับแสง กลุ่มของเซลล์ดังกล่าวเรียกว่า "จุดรับแสง" และมีการวิวัฒนาการอย่างอิสระในช่วงระหว่าง 40 ถึง 65 ครั้ง จุดรับแสงเหล่านี้ทำให้สัตว์สามารถรับรู้ทิศทางและความเข้มของแสงได้เพียงระดับพื้นฐาน แต่ไม่เพียงพอที่จะแยกแยะวัตถุออกจากสภาพแวดล้อม[ 21 ]
การพัฒนาระบบออปติกที่สามารถแยกแยะทิศทางของแสงได้ภายในไม่กี่องศาดูเหมือนจะยากกว่ามาก และมีเพียงหกไฟลัมจากสามสิบกว่าไฟลัม[หมายเหตุ 1 ] เท่านั้น ที่มีระบบดังกล่าว อย่างไรก็ตาม ไฟลัมเหล่านี้คิดเป็น 96% ของสิ่งมีชีวิตทั้งหมด[ 21 ]

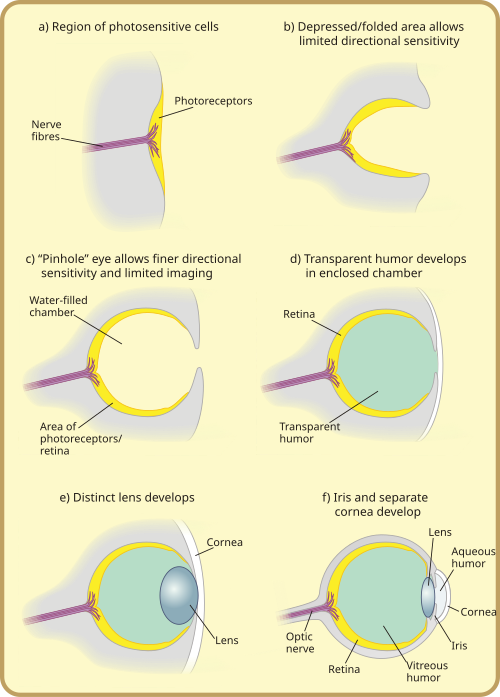
ระบบรับแสงที่ซับซ้อนเหล่านี้เริ่มต้นจากการที่เซลล์รับแสงหลายเซลล์ค่อยๆ ยุบตัวลงเป็นรูปถ้วย ซึ่งในตอนแรกทำให้สามารถแยกแยะความสว่างในทิศทางต่างๆ ได้ จากนั้นก็แยกแยะในทิศทางที่ละเอียดขึ้นเรื่อยๆ เมื่อหลุมลึกขึ้น ในขณะที่เซลล์รับแสงแบบแบนไม่สามารถระบุทิศทางของแสงได้อย่างมีประสิทธิภาพ เนื่องจากลำแสงจะกระตุ้นเซลล์รับแสงกลุ่มเดียวกันไม่ว่าทิศทางจะเป็นอย่างไร รูปทรง "ถ้วย" ของดวงตาแบบหลุมทำให้สามารถแยกแยะทิศทางได้ในระดับจำกัดโดยการเปลี่ยนเซลล์ที่แสงจะตกกระทบขึ้นอยู่กับมุมของแสง ดวงตาแบบหลุมซึ่งเกิดขึ้นใน ยุค แคมเบรียนพบได้ในหอยทาก โบราณ และพบในหอยทากบางชนิดและสัตว์ไม่มีกระดูกสันหลังอื่นๆ ที่มีชีวิตอยู่ในปัจจุบัน เช่นพลานาเรียพลานาเรียสามารถแยกแยะทิศทางและความเข้มของแสงได้เล็กน้อยเนื่องจาก เซลล์ เรตินา ที่มีรูปร่างคล้ายถ้วยและมีเม็ดสีเข้ม ซึ่งปกป้องเซลล์รับแสงจากการสัมผัสกับแสงในทุกทิศทางยกเว้นช่องเปิดเดียวสำหรับแสง อย่างไรก็ตาม ดวงตาแบบดั้งเดิมนี้ยังมีประโยชน์มากกว่าในการตรวจจับการมีอยู่หรือไม่มีอยู่ของแสงมากกว่าทิศทางของแสง สิ่งนี้จะค่อยๆ เปลี่ยนแปลงไปเมื่อหลุมตาลึกขึ้นและจำนวนเซลล์รับแสงเพิ่มขึ้น ทำให้สามารถรับข้อมูลภาพได้แม่นยำยิ่งขึ้น[ 25 ]
เมื่อโฟตอนถูกดูดซับโดยโครโมฟอร์ ปฏิกิริยาเคมีจะทำให้พลังงานของโฟตอนถูกแปลงเป็นพลังงานไฟฟ้าและส่งต่อไปยังระบบประสาท ในสัตว์ชั้นสูง เซลล์รับแสงเหล่านี้เป็นส่วนหนึ่งของเรตินาซึ่งเป็นชั้นเซลล์บาง ๆ ที่ส่งต่อข้อมูลภาพ[ 26 ]รวมถึงข้อมูลแสงและความยาวของวันซึ่งจำเป็นต่อระบบจังหวะชีวภาพไปยังสมอง อย่างไรก็ตามแมงกะพรุน บางชนิด เช่นCladonema ( Cladonematidae ) มีดวงตาที่ซับซ้อนแต่ไม่มีสมอง ดวงตาของพวกมันส่งข้อความไปยังกล้ามเนื้อโดยตรงโดยไม่ต้องผ่านกระบวนการประมวลผลระดับกลางของสมอง[ 18 ]
ในช่วงการระเบิดของแคมเบรียนการพัฒนาของดวงตาเร่งตัวขึ้นอย่างรวดเร็ว พร้อมกับการปรับปรุงอย่างมากในการประมวลผลภาพและการตรวจจับทิศทางของแสง[ 27 ]

หลังจากบริเวณเซลล์รับแสงเกิดการเว้าเข้าไปแล้วถึงจุดหนึ่งการลดความกว้างของช่องรับแสงจะมีประสิทธิภาพมากกว่าในการเพิ่มความละเอียดในการมองเห็นมากกว่าการทำให้ช่องรับแสงลึกขึ้นเรื่อยๆ[ 9 ]ด้วยการลดขนาดของช่องรับแสง สิ่งมีชีวิตจึงสามารถสร้างภาพได้อย่างแท้จริง ทำให้สามารถรับรู้ทิศทางได้อย่างละเอียด และแม้กระทั่งรับรู้รูปร่างได้บางส่วน ปัจจุบันพบดวงตาลักษณะนี้ในนอติลัสแม้ว่าจะไม่มีกระจกตาหรือเลนส์ ทำให้มีความละเอียดต่ำและภาพไม่ชัด แต่ก็ยังถือเป็นการพัฒนาที่สำคัญกว่าแผ่นปิดตาในยุคแรกๆ ในแง่ของการมองเห็น[ 28 ]
การเจริญเติบโตมากเกินไปของเซลล์โปร่งใสช่วยป้องกันการปนเปื้อนและการติดเชื้อปรสิต เนื้อหาภายในช่องซึ่งแยกออกจากกันแล้ว สามารถค่อยๆ พัฒนาไปเป็นของเหลวโปร่งใส เพื่อการปรับปรุงต่างๆ เช่น การกรองสีดัชนีหักเห ที่สูงขึ้น การป้องกัน รังสี อัลตราไวโอเลตหรือความสามารถในการทำงานในและนอกน้ำ ชั้นนี้อาจเกี่ยวข้องกับการลอกคราบของเปลือกหรือผิวหนังของสิ่งมีชีวิตในบางกลุ่ม ตัวอย่างเช่น สามารถสังเกตได้ในOnychophoransซึ่งคิวติคูล่าของเปลือกจะต่อเนื่องไปยังกระจกตา กระจกตาประกอบด้วยชั้นคิวติคูล่าหนึ่งหรือสองชั้น ขึ้นอยู่กับว่าสัตว์นั้นลอกคราบเมื่อเร็วๆ นี้หรือ ไม่ [ 29 ]ร่วมกับเลนส์และของเหลวสองชนิด กระจกตามีหน้าที่ในการรวมแสงและช่วยในการโฟกัสแสงที่ด้านหลังของเรตินา กระจกตาปกป้องลูกตา ในขณะเดียวกันก็มีส่วนในการหักเหแสงประมาณ 2/3 ของกำลังการหักเหแสงทั้งหมดของดวงตา[ 30 ]
เป็นไปได้ว่าเหตุผลสำคัญที่ดวงตาเชี่ยวชาญในการตรวจจับช่วงความยาวคลื่นที่แคบและเฉพาะเจาะจงบนสเปกตรัมแม่เหล็กไฟฟ้า — สเปกตรัมที่มองเห็นได้ —ก็คือสายพันธุ์แรกสุดที่พัฒนาความไวต่อแสงเป็นสิ่งมีชีวิตในน้ำ และน้ำจะกรองรังสีแม่เหล็กไฟฟ้า ออกไป ยกเว้นช่วงความยาวคลื่นหนึ่ง ซึ่งความยาวคลื่นที่สั้นกว่าที่เราเรียกว่าสีน้ำเงิน ไปจนถึงความยาวคลื่นที่ยาวกว่าที่เราเรียกว่าสีแดง คุณสมบัติการกรองแสงของน้ำนี้ยังมีอิทธิพลต่อความไวต่อแสงของพืชด้วย[ 31 ] [ 32 ] [ 33 ]
การก่อตัวและการกระจายตัวของเลนส์
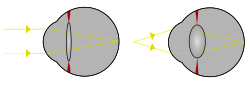
ในดวงตาที่ไม่มีเลนส์ แสงที่มาจากจุดไกลจะกระทบด้านหลังของดวงตาโดยมีขนาดใกล้เคียงกับรูรับแสง ของดวงตา เมื่อมีเลนส์ แสงที่เข้ามาจะถูกรวมไว้บนพื้นที่ผิวที่เล็กลง โดยไม่ลดความเข้มโดยรวมของสิ่งเร้า[ 34 ]ระยะโฟกัสของโลโบพอ ดในยุคแรก ที่มีดวงตาแบบง่ายที่มีเลนส์จะโฟกัสภาพไว้ด้านหลังเรตินา ดังนั้นแม้ว่าจะไม่สามารถโฟกัสส่วนใดส่วนหนึ่งของภาพได้ แต่ความเข้มของแสงทำให้สิ่งมีชีวิตสามารถมองเห็นในน้ำที่ลึกกว่า (และมืดกว่า) ได้[ 29 ] การเพิ่มขึ้นของ ดัชนีหักเหของเลนส์ในภายหลังน่าจะส่งผลให้เกิดภาพที่โฟกัสได้[ 29 ]
โปรดทราบว่าโครงสร้างทางแสงแบบนี้ยังไม่เคยถูกค้นพบ และไม่คาดว่าจะถูกค้นพบด้วย การเกิดฟอสซิลมักไม่สามารถรักษาเนื้อเยื่ออ่อนไว้ได้ และถึงแม้ว่าจะรักษาไว้ได้ เนื้อเยื่อใหม่ก็จะปิดตัวลงอย่างแน่นอนเมื่อซากแห้งลง หรือเมื่อตะกอนทับถมเข้ามาทำให้ชั้นต่างๆ ชิดกัน ทำให้ดวงตาที่กลายเป็นฟอสซิลมีลักษณะคล้ายกับโครงสร้างเดิม
คริสตัลลิน

เลนส์ของสัตว์มีกระดูกสันหลังประกอบด้วย เซลล์ เยื่อบุผิว ที่ปรับตัวแล้ว ซึ่งมีโปรตีนคริสตัลลิน ในปริมาณสูง คริสตัลลินเหล่านี้อยู่ในสองตระกูลหลัก ได้แก่ α-คริสตัลลินและ βγ-คริสตัลลิน โปรตีนทั้งสองประเภทนี้เดิมทีใช้สำหรับหน้าที่อื่นในสิ่งมีชีวิต แต่ในที่สุดก็ปรับตัวเพื่อการมองเห็นในดวงตาของสัตว์[ 35 ]ในตัวอ่อน เลนส์เป็นเนื้อเยื่อที่มีชีวิต แต่กลไกของเซลล์ไม่โปร่งใส ดังนั้นจึงต้องกำจัดออกก่อนที่สิ่งมีชีวิตจะสามารถมองเห็นได้ การกำจัดกลไกหมายความว่าเลนส์ประกอบด้วยเซลล์ที่ตายแล้วซึ่งอัดแน่นไปด้วยคริสตัลลิน คริสตัลลินเหล่านี้มีความพิเศษเนื่องจากมีลักษณะเฉพาะที่จำเป็นต่อความโปร่งใสและการทำงานในเลนส์ เช่น การอัดแน่น ความต้านทานต่อการตกผลึก และอายุยืนยาวมาก เนื่องจากต้องอยู่รอดตลอดช่วงชีวิตของสิ่งมีชีวิต[ 35 ]การไล่ระดับดัชนีหักเห ที่ทำให้เลนส์มีประโยชน์นั้นเกิดจากการเปลี่ยนแปลงเชิงรัศมีของความเข้มข้นของคริสตัลลินในส่วนต่างๆ ของเลนส์ มากกว่าที่จะเกิดจากชนิดของโปรตีนโดยเฉพาะ กล่าวคือ ไม่ใช่การมีอยู่ของคริสตัลลิน แต่เป็นการกระจายตัวสัมพัทธ์ของมันต่างหากที่ทำให้เลนส์มีประโยชน์[ 36 ]
การรักษาชั้นเซลล์ที่โปร่งใสเป็นเรื่องยากทางชีววิทยา[ 37 ] การสะสมของวัสดุโปร่งใสที่ไม่มีชีวิตช่วยลดความจำเป็นในการจัดหาสารอาหารและการกำจัดของเสีย เป็นที่เข้าใจกันโดยทั่วไปว่าไทรโลไบต์ใช้แคลไซต์ซึ่งเป็นแร่ธาตุที่ปัจจุบันทราบกันว่าใช้สำหรับการมองเห็นในดาวเปราะ เพียงชนิดเดียวเท่านั้น [ 38 ]การศึกษาดวงตาจากฟอสซิลแมลงวันขายาวอายุ 55 ล้านปีจากFur Formationบ่งชี้ว่าแคลไซต์ในดวงตาของไทรโลไบต์เป็นผลมาจากกระบวนการทาโฟโนมิกและไดอะเจเนติก ไม่ใช่ลักษณะดั้งเดิม[ 39 ]ในดวงตาประกอบและดวงตากล้องอื่นๆ วัสดุนี้คือคริสตัลลิน [ 40 ] ช่อง ว่างระหว่างชั้นเนื้อเยื่อจะก่อตัวเป็นรูปทรงนูนสองด้านตามธรรมชาติ ซึ่งเหมาะอย่างยิ่งทางแสงและทางกลสำหรับสารที่มีดัชนีหักเหปกติ เลนส์นูนสองด้านไม่เพียงแต่ให้ความละเอียดทางแสงเท่านั้น แต่ยังให้รูรับแสงและความสามารถในการ มองเห็นในที่แสงน้อยด้วย เนื่องจากความละเอียดนั้นแยกออกจากขนาดของรู ซึ่งจะค่อยๆ เพิ่มขึ้นอีกครั้งโดยปราศจากข้อจำกัดของการไหลเวียน
น้ำในช่องหน้าลูกตา ม่านตา และกระจกตา
โดยอิสระ ชั้นโปร่งใสและชั้นทึบแสงอาจแยกตัวออกมาจากเลนส์ด้านหน้า: กระจกตาและม่านตา ที่แยกจากกัน (สิ่งเหล่านี้อาจเกิดขึ้นก่อนหรือหลังการตกผลึก หรืออาจไม่เกิดขึ้นเลยก็ได้) การแยกตัวของชั้นด้านหน้าจะก่อให้เกิดของเหลวขึ้นอีกครั้ง ซึ่งก็คือน้ำหล่อเลี้ยงลูก ตา สิ่งนี้จะเพิ่มกำลังการหักเหของแสงและช่วยบรรเทาปัญหาการไหลเวียนโลหิต การก่อตัวของวงแหวนทึบแสงช่วยให้มีหลอดเลือดมากขึ้น การไหลเวียนโลหิตมากขึ้น และขนาดตาที่ใหญ่ขึ้น แผ่นพับรอบขอบเลนส์นี้ยังช่วยปกปิดความไม่สมบูรณ์ทางแสง ซึ่งมักพบได้บ่อยที่ขอบเลนส์ ความจำเป็นในการปกปิดความไม่สมบูรณ์ของเลนส์จะค่อยๆ เพิ่มขึ้นตามความโค้งและกำลังของเลนส์ ขนาดโดยรวมของเลนส์และดวงตา และความต้องการด้านความละเอียดและรูรับแสงของสิ่งมีชีวิต ซึ่งได้รับแรงผลักดันจากความต้องการในการล่าสัตว์หรือการอยู่รอด ปัจจุบันชนิดนี้มีลักษณะการทำงานเหมือนกับดวงตาของสัตว์มีกระดูกสันหลังส่วนใหญ่ รวมถึงมนุษย์ อันที่จริง "รูปแบบพื้นฐานของดวงตาของสัตว์มีกระดูกสันหลังทั้งหมดมีความคล้ายคลึงกัน" [ 41 ]
ความคืบหน้าอื่นๆ
การมองเห็นสี
พบออปซินสำหรับการมองเห็น 5 ประเภท ในสัตว์มีกระดูกสันหลัง ทั้งหมดนี้ยกเว้นหนึ่งประเภทพัฒนาขึ้นก่อนการแยกสายวิวัฒนาการของ Cyclostomataและปลา[ 42 ]ออปซินทั้ง 5 ประเภทมีการปรับตัวแตกต่างกันไปตามสเปกตรัมของแสงที่พบ เมื่อแสงเดินทางผ่านน้ำคลื่นแสงที่มีความยาวคลื่นยาวกว่า เช่น สีแดงและสีเหลือง จะถูกดูดซับได้เร็วกว่าคลื่น แสงที่มีความยาวคลื่นสั้นกว่า เช่น สีเขียวและสีน้ำเงิน ทำให้เกิดการไล่ระดับความหนาแน่นของพลังงานสเปกตรัมโดยความยาวคลื่นเฉลี่ยจะสั้นลงเมื่อความลึกของน้ำเพิ่มขึ้น[ 43 ]ออปซินสำหรับการมองเห็นในปลามีความไวต่อช่วงของแสงในถิ่นที่อยู่และความลึกของพวกมันมากกว่า อย่างไรก็ตาม สภาพแวดล้อมบนบกไม่มีการเปลี่ยนแปลงในองค์ประกอบของความยาวคลื่น ดังนั้นความไวของออปซินในสัตว์มีกระดูกสันหลังบนบกจึงไม่แตกต่างกันมากนัก ซึ่งส่งผลโดยตรงต่อการมีอยู่ของสีสำหรับการสื่อสารอย่างมีนัยสำคัญ[ 42 ]การมองเห็นสีให้ ข้อได้เปรียบ เชิงเลือก ที่ชัดเจน เช่น การจดจำผู้ล่า อาหาร และคู่ผสมพันธุ์ได้ดีขึ้น อันที่จริง เป็นไปได้ว่ากลไกประสาทรับความรู้สึกแบบง่ายๆ อาจควบคุมรูปแบบพฤติกรรมทั่วไป เช่น การหลบหนี การหาอาหาร และการซ่อนตัว ตัวอย่างมากมายของพฤติกรรมที่เฉพาะเจาะจงกับความยาวคลื่นได้รับการระบุในสองกลุ่มหลัก ได้แก่ ต่ำกว่า 450 นาโนเมตร ซึ่งเกี่ยวข้องกับแสงโดยตรง และสูงกว่า 450 นาโนเมตร ซึ่งเกี่ยวข้องกับแสงสะท้อน[ 44 ]เนื่องจากโมเลกุลออปซินได้รับการปรับแต่งให้ตรวจจับความยาวคลื่นแสงที่แตกต่างกัน ในบางจุดการมองเห็นสีจึงพัฒนาขึ้นเมื่อเซลล์รับแสงใช้ออปซินที่ปรับแต่งแตกต่างกัน[ 26 ]สิ่งนี้อาจเกิดขึ้นในระยะแรกๆ ของวิวัฒนาการของดวงตา และอาจหายไปและวิวัฒนาการขึ้นใหม่เมื่อแรงกดดันในการคัดเลือกที่สัมพันธ์กันในสายพันธุ์แตกต่างกัน
การมองเห็นแบบโพลาไรซ์
การโพลาไรเซชันคือการจัดระเบียบแสงที่ไม่เป็นระเบียบให้เป็นการจัดเรียงเชิงเส้น ซึ่งเกิดขึ้นเมื่อแสงผ่านตัวกรองแบบช่องแคบ รวมถึงเมื่อผ่านเข้าไปในตัวกลางใหม่ ความไวต่อแสงโพลาไรซ์มีประโยชน์อย่างยิ่งสำหรับสิ่งมีชีวิตที่มีถิ่นที่อยู่ใต้น้ำลึกกว่าสองสามเมตร ในสภาพแวดล้อมนี้ การมองเห็นสีมีความน่าเชื่อถือน้อยลง และด้วยเหตุนี้จึงเป็นปัจจัยการคัดเลือกที่อ่อนแอลง ในขณะที่ตัวรับแสงส่วนใหญ่มีความสามารถในการแยกแยะแสงโพลาไรซ์บางส่วน เยื่อหุ้มเซลล์ของสัตว์มีกระดูกสันหลังบนบกมีทิศทางตั้งฉาก ทำให้ไม่ไวต่อแสงโพลาไรซ์[ 45 ]อย่างไรก็ตาม ปลาบางชนิดสามารถแยกแยะแสงโพลาไรซ์ได้ แสดงให้เห็นว่าพวกมันมีตัวรับแสงเชิงเส้นอยู่บ้าง นอกจากนี้ ปลาหมึกยังสามารถรับรู้การโพลาไรซ์ของแสงด้วยความแม่นยำในการมองเห็นสูง แม้ว่าพวกมันดูเหมือนจะขาดความสามารถในการแยกแยะสีอย่างมีนัยสำคัญ[ 46 ]เช่นเดียวกับการมองเห็นสี ความไวต่อการโพลาไรซ์สามารถช่วยให้สิ่งมีชีวิตสามารถแยกแยะวัตถุและบุคคลรอบข้างได้ เนื่องจากการรบกวนการสะท้อนแสงเล็กน้อยของแสงโพลาไรซ์ จึงมักใช้สำหรับการกำหนดทิศทางและการนำทาง รวมถึงการแยกแยะวัตถุที่ซ่อนอยู่ เช่น เหยื่อที่ปลอมตัว[ 45 ]
กลไกการโฟกัส
โดยการใช้ กล้ามเนื้อ หูรูด ม่านตา และร่างกายซิลิ อารี บางชนิดจะขยับเลนส์ไปมา บางชนิดจะยืดเลนส์ให้แบนราบขึ้น กลไกอีกอย่างหนึ่งควบคุมการโฟกัสทางเคมีและเป็นอิสระจากสองกลไกนี้ โดยควบคุมการเจริญเติบโตของดวงตาและรักษาระยะโฟกัส นอกจากนี้ รูปร่างของรูม่านตายังสามารถใช้ในการทำนายระบบโฟกัสที่ใช้ได้ รูม่านตาแบบแคบสามารถบ่งชี้ถึงระบบมัลติโฟกัสทั่วไป ในขณะที่รูม่านตาแบบวงกลมมักจะระบุถึงระบบโมโนโฟกัส เมื่อใช้รูปทรงกลม รูม่านตาจะหดตัวภายใต้แสงสว่าง ทำให้ค่า f เพิ่มขึ้น และจะขยายตัวเมื่อมืดเพื่อลดความชัดลึก[ 47 ]โปรดทราบว่าวิธีการโฟกัสไม่ใช่ข้อกำหนด ช่างภาพทราบดีว่าข้อผิดพลาดในการโฟกัสจะเพิ่มขึ้นเมื่อรูรับแสงเพิ่มขึ้น ดังนั้นสิ่งมีชีวิตจำนวนนับไม่ถ้วนที่มีดวงตาขนาดเล็กจึงเคลื่อนไหวในแสงแดดโดยตรงและอยู่รอดได้โดยไม่มีกลไกการโฟกัสเลย เมื่อสายพันธุ์มีขนาดใหญ่ขึ้นหรือเปลี่ยนไปอยู่ในสภาพแวดล้อมที่มืดลง วิธีการโฟกัสก็ไม่จำเป็นต้องเกิดขึ้นบ่อยนัก
การจัดวาง
โดยทั่วไปแล้ว สัตว์นักล่าจะมีตาอยู่ด้านหน้าของหัวเพื่อการรับรู้เชิงลึก ที่ดีขึ้น ในการโฟกัสไปที่เหยื่อ ส่วนสัตว์เหยื่อมักจะมีตาอยู่ด้านข้างของหัว ทำให้มีมุมมองที่กว้างในการตรวจจับสัตว์นักล่าจากทุกทิศทาง[ 48 ] [ 49 ]ปลาแบน เป็นสัตว์นักล่าที่นอนตะแคงอยู่บนพื้น และมีตาวาง อยู่ ไม่สมมาตรที่ด้านเดียวกันของหัวฟอสซิลที่เป็นตัวเปลี่ยนผ่านจากตำแหน่งสมมาตรทั่วไปไปสู่ตำแหน่งไม่สมมาตรคือAmphistium
สัมภาระวิวัฒนาการ

{kind=link}
ดวงตาของสัตว์หลายชนิดบันทึกประวัติวิวัฒนาการของพวกมันไว้ในกายวิภาคศาสตร์ปัจจุบัน ตัวอย่างเช่น ดวงตาของสัตว์มีกระดูกสันหลังถูกสร้างขึ้น "กลับหัวและคว่ำลง" โดยต้อง "ให้โฟตอนของแสงเดินทางผ่านกระจกตา เลนส์ ของเหลวในช่องหน้าลูกตา หลอดเลือด เซลล์แกงลีออน เซลล์อะมาครีน เซลล์แนวนอน และเซลล์ไบโพลาร์ ก่อนที่จะไปถึงแท่งและกรวยที่ไวต่อแสงซึ่งแปลงสัญญาณแสงเป็นแรงกระตุ้นประสาท จากนั้นจึงส่งไปยังคอร์เทกซ์การมองเห็นที่ด้านหลังของสมองเพื่อประมวลผลเป็นรูปแบบที่มีความหมาย" [ 50 ]แม้ว่าโครงสร้างดังกล่าวจะมีข้อเสียอยู่บ้าง แต่ก็ช่วยให้เรตินาชั้นนอกของสัตว์มีกระดูกสันหลังสามารถรักษาการทำงานของระบบเผาผลาญได้สูงกว่าเมื่อเทียบกับการออกแบบที่ไม่กลับหัว[ 51 ]นอกจากนี้ยังช่วยให้เกิดวิวัฒนาการของชั้นคอรอยด์รวมถึงเซลล์เยื่อบุผิวเม็ดสีเรตินา (RPE) ซึ่งมีบทบาทสำคัญในการปกป้องเซลล์รับแสงจากความเสียหายจากปฏิกิริยาออกซิเดชันของแสง[ 52 ] [ 53 ]
ในทางตรงกันข้าม ดวงตาของสัตว์จำพวกเซฟาโลพอดถูกสร้างขึ้นใน "ทิศทางที่ถูกต้อง" โดยมีเส้นประสาทติดอยู่ที่ด้านหลังของเรตินา ซึ่งหมายความว่าพวกมันไม่มีจุดบอด ความแตกต่างนี้อาจอธิบายได้จากต้นกำเนิดของดวงตา ในเซฟาโลพอด ดวงตาพัฒนามาจากการยุบตัวของพื้นผิวหัว ในขณะที่ในสัตว์มีกระดูกสันหลัง ดวงตาเกิดขึ้นจากส่วนขยายของสมอง[ 54 ]
ดูเพิ่มเติม
เชิงอรรถอธิบาย
- ^ตัวเลขที่แน่นอนอาจแตกต่างกันไปในแต่ละผู้เขียน
อ่านเพิ่มเติม
- Lamb TD, Collin SP, Pugh EN (ธันวาคม 2007). "วิวัฒนาการของดวงตาของสัตว์มีกระดูกสันหลัง: ออปซิน, โฟโตรีเซปเตอร์, เรตินา และถ้วยตา" Nature Reviews Neuroscience 8 ( 12): 960– 76. doi : 10.1038/nrn2283 . PMC 3143066 . PMID 18026166 .ภาพประกอบ. บทวิจารณ์
- Lamb, TD (2011). "วิวัฒนาการของดวงตา" (PDF) . Scientific American . 305 (1): 64– 69. Bibcode : 2011SciAm.305f..64L . doi : 10.1038/scientificamerican0711-64 . เก็บถาวรจากต้นฉบับ(PDF)เมื่อวันที่ 12 ธันวาคม 2013 . สืบค้นเมื่อ28 เมษายน 2013 .
- แลนด์, ไมเคิล เอฟ.; นิลส์สัน, แดน-เอริค (2012). "ต้นกำเนิดของการมองเห็น". ดวงตาของสัตว์ (ฉบับที่ 2). อ็อกซ์ฟอร์ด: สำนักพิมพ์มหาวิทยาลัยอ็อกซ์ฟอร์ด. หน้า 1–22 . ISBN 978-0-19-958114-6.
- ฉบับพิเศษ "วิวัฒนาการของดวงตา"ในวารสาร Evolution: Education and Outreachเล่มที่ 1 ฉบับที่ 4 / ตุลาคม 2551 มีบทความ 26 บทความ สามารถเข้าถึงได้ฟรี
- อีวาน อาร์. ชวาบ (2012). พยานแห่งวิวัฒนาการ: วิวัฒนาการของดวงตา . นิวยอร์ก: สำนักพิมพ์มหาวิทยาลัยออกซ์ฟอร์ด. ISBN 978-0-19-536974-8.
- Hayakawa S, Takaku Y, Hwang JS, Horiguchi T, Suga H, Gehring W และคณะ (2015). "หน้าที่และต้นกำเนิดวิวัฒนาการของโครงสร้างตาแบบกล้องในเซลล์เดียว" PLOS ONE . 10 (3) e0118415. Bibcode : 2015PLoSO..1018415H . doi : 10.1371/journal.pone.0118415 . PMC 4348419 . PMID 25734540 .
- กรอยต์, ซี (1968) "โครงสร้างองค์กรขั้นสูงของ l'ocelle de deux Peridiniens Warnowiidae, Erythropsis papillardi Kofoid และ Swezy และ Warnowia pulchra Schiller" โปรติสโตโลจิกา . 4 : 209– 230.
- Gregory S. Gavelis; Shiho Hayakawa; Richard A. White III; Takashi Gojobori; Curtis A. Suttle; Patrick J. Keeling; Brian S. Leander (2015). " Ocelloids ที่มีลักษณะคล้ายดวงตาถูกสร้างขึ้นจากส่วนประกอบที่ได้รับมาจากการอยู่ร่วมกันแบบพึ่งพาอาศัยกัน" Nature . 523 ( 7559): 204–7 . Bibcode : 2015Natur.523..204G . doi : 10.1038/nature14593 . hdl : 10754/566109 . PMID 26131935. S2CID 4462376.เก็บถาวรจากต้นฉบับเมื่อวันที่ 12 เมษายน 2018. สืบค้นเมื่อ2 กรกฎาคม 2019 .
- Oakley, Todd H.; Speiser, Daniel I. (2015). "How Complexity Originates: The Evolution of Animal Eyes" (PDF) . Annual Review of Ecology, Evolution, and Systematics . 46 : 237–260 . doi : 10.1146/annurev-ecolsys-110512-135907 .
- เอ็ด ยัง; ภาพถ่ายโดย เดวิด ลิตต์ชวาเกอร์ (กุมภาพันธ์ 2016) "ภายในดวงตา: สิ่งสร้างที่งดงามที่สุดของธรรมชาติ"เนชั่นแนลจีโอกราฟิก 229 ( 2): 30– 57 เก็บถาวรจากต้นฉบับเมื่อวันที่ 24 มิถุนายน 2017
ลิงก์ภายนอก
- "วิวัฒนาการของดวงตา"มูลนิธิการศึกษา WGBH และ Clear Blue Sky Productionsสถานีโทรทัศน์ PBS ปี 2001
- ทฤษฎีการสร้างโลกตามคัมภีร์ไบเบิลถูกหักล้างแล้วหรือ?วิดีโอจากศูนย์การศึกษาด้านวิทยาศาสตร์แห่งชาติเกี่ยวกับการวิวัฒนาการของดวงตา
- วิวัฒนาการ:ฉบับพิเศษด้านการศึกษาและการเผยแพร่: วิวัฒนาการและดวงตาเล่ม 1 ฉบับที่ 4 ตุลาคม 2551 หน้า 351–559 ISSN 1936-6426 (ฉบับพิมพ์) 1936–6434 (ฉบับออนไลน์)
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ วิวัฒนาการของดวงตา
วิวัฒนาการ ของดวงตา คือต้นกำเนิดและการพัฒนาที่มีความหลากหลายของการคัดเลือกโดยธรรมชาติในช่วงเวลาทางธรณีวิทยาของอวัยวะที่ไวต่อแสงและการมองเห็นในสิ่งมีชีวิต...
ประวัติการวิจัย
ในปี ค.ศ. 1802 นักปรัชญา William Paley เรียกมันว่าปาฏิหาริย์แห่ง " การออกแบบ " ในปี ค.ศ.
อัตราการวิวัฒนาการ
ฟอสซิลดวงตาที่อาจพบเป็นครั้งแรกจนถึงปัจจุบันมาจาก ยุค เอเดียคารัน (ประมาณ 555 ล้านปีก่อน ) [ 5 ] ในขณะที่ฟอสซิลดวงตาที่เก่าแก่ที่สุดที่ยืนยันได้คือ ฟอสซิล Schmidtiellus reetae จาก 530 ล้านปีก่อน ซึ่งเก็บรวบรวมได้ใน ซาวิรานนา ทางตอนเหนือของเอสโตเนีย...
กำเนิดของดวงตา
ไม่ว่าดวงตาจะวิวัฒนาการเพียงครั้งเดียวหรือหลายครั้งนั้นขึ้นอยู่กับนิยามของดวงตา สัตว์ที่มีดวงตาทั้งหมดมีกลไกทางพันธุกรรมร่วมกันมากมายสำหรับการพัฒนาดวงตา ซึ่งบ่งชี้ว่าบรรพบุรุษของสัตว์ที่มีดวงตามีกลไกที่ไวต่อแสงบางรูปแบบ...