อ่าน 24 นาที
การแก้ไขจีโนม
การแก้ไขจีโนมหรือวิศวกรรมจีโนมหรือการแก้ไขยีน เป็น วิศวกรรมพันธุกรรมประเภทหนึ่งที่แทรก ลบ แก้ไข หรือแทนที่DNA ใน จีโนมของสิ่งมีชีวิต แตกต่างจากเทคนิควิศวกรรมพันธุกรรม ในยุคแรกๆ
การแก้ไขจีโนม

การแก้ไขจีโนมหรือวิศวกรรมจีโนมหรือการแก้ไขยีน เป็น วิศวกรรมพันธุกรรมประเภทหนึ่งที่แทรก ลบ แก้ไข หรือแทนที่DNA ใน จีโนมของสิ่งมีชีวิต แตกต่างจากเทคนิควิศวกรรมพันธุกรรม ในยุคแรกๆ ที่แทรกสารพันธุกรรมเข้าไปในจีโนมของโฮสต์แบบสุ่ม การแก้ไขจีโนมจะกำหนดเป้าหมายการแทรกไปยังตำแหน่งที่เฉพาะเจาะจง กลไกพื้นฐานที่เกี่ยวข้องกับการจัดการทางพันธุกรรมผ่านนิวคลีเอสที่ตั้งโปรแกรมได้ คือการจดจำตำแหน่งจีโนมเป้าหมายและการจับของโดเมนจับ DNA (DBD) ของตัวกระตุ้น การแตกของสายคู่ (DSBs) ใน DNA เป้าหมายโดยเอนโดนิวคลีเอสจำกัด ( FokIและCas ) และการซ่อมแซม DSBs ผ่านการรวมตัวใหม่แบบกำหนดทิศทางตามความเหมือนกัน (HDR) หรือการเชื่อมต่อปลายที่ไม่เหมือนกัน (NHEJ) [ 1 ] [ 2 ]
การพัฒนาการแก้ไขยีน CRISPRในปี 2558 ได้ปรับปรุงประสิทธิภาพ ความจำเพาะ และความเหมาะสมของการแก้ไขจีโนมขนาดใหญ่[ 3 ]
ตั้งแต่ปี 2015 การแก้ไขจีโนมได้รับการศึกษาทดลองใน ตัวอ่อนมนุษย์ที่ไม่สามารถมีชีวิตรอดได้[ 4 ] ในปี 2019 มนุษย์คนแรกถือกำเนิดจากตัวอ่อนที่ได้รับการแก้ไขจีโนมโดยใช้เทคนิค CRISPR อันเป็นผลมาจากกรณีของเหอ เจี้ยนควิที่ เป็นที่ถกเถียง [ 5 ]
ประวัติศาสตร์
การแก้ไขจีโนมได้รับการบุกเบิกในช่วงทศวรรษ 1990 [ 6 ]ก่อนการเกิดขึ้นของแพลตฟอร์มการแก้ไขยีนแบบนิวคลีเอสที่ใช้กันทั่วไปในปัจจุบัน แต่การใช้งานถูกจำกัดด้วยประสิทธิภาพการแก้ไขที่ต่ำ การแก้ไขจีโนมด้วยนิวคลีเอสที่ได้รับการดัดแปลงทางวิศวกรรม กล่าวคือ เอนไซม์ทั้งสามประเภทหลัก ได้แก่ นิวคลีเอสแบบนิ้วสังกะสี (ZFNs) นิวคลีเอสแบบเอฟเฟกเตอร์คล้ายตัวกระตุ้นการถอดรหัส (TALENs) และเมกะนิวคลีเอสที่ได้รับการดัดแปลงทางวิศวกรรม ได้รับเลือกโดยNature Methodsให้เป็นวิธีการแห่งปี 2011 [ 7 ]ระบบ CRISPR-Cas ได้รับเลือกโดยScienceให้เป็นความก้าวหน้าแห่งปี 2015 [ 8 ]
ณ ปี 2015 มีการใช้เอนไซม์นิวคลีเอสที่ได้รับการออกแบบทางวิศวกรรม 4 ตระกูล ได้แก่เมกะนิว คลีเอส , ซิงค์ฟิง เกอร์นิวคลีเอส (ZFNs), TALEN ( transcription activator-like effector-based nucleases ) และ ระบบCRISPR / Cas9 (clustered regularly interspaced short palindromic repeats) [ 9 ] [ 10 ] [ 11 ] [ 12 ]มีตัวแก้ไขจีโนม 9 ตัวที่พร้อมใช้งาน ณ ปี 2017 [ 13 ]
ในปี 2018 วิธีการทั่วไปสำหรับการแก้ไขจีโนมดังกล่าวใช้เอนไซม์นิวคลีเอสที่ถูกดัดแปลง หรือ "กรรไกรโมเลกุล" เอนไซม์นิวคลีเอสเหล่านี้สร้างรอยแตกสองสาย (DSBs) เฉพาะจุดในตำแหน่งที่ต้องการในจีโนม รอยแตกสองสายที่เกิดขึ้นจะได้รับการซ่อมแซมผ่านการเชื่อมต่อปลายที่ไม่เหมือนกัน (NHEJ) หรือการรวมตัวแบบเหมือนกัน (HR) ส่งผลให้เกิดการกลายพันธุ์ เป้าหมาย ('การแก้ไข')
ในเดือนพฤษภาคม 2019 ทนายความในประเทศจีนรายงานว่า จากการที่นักวิทยาศาสตร์ชาวจีนเหอ เจียนควิ อ้างว่าได้สร้าง มนุษย์ที่ได้รับการแก้ไขยีนเป็นครั้งแรก ( ดูข้อโต้แย้งเรื่องลูลู่และนานา ) จึงได้มีการร่างกฎระเบียบที่ระบุว่า ผู้ใดก็ตามที่ทำการดัดแปลงจีโนมของมนุษย์โดยใช้เทคนิคการแก้ไขยีน เช่นCRISPRจะต้องรับผิดชอบต่อผลเสียใดๆ ที่เกี่ยวข้อง[ 5 ]มุมมองเชิงเตือนเกี่ยวกับจุดบอดและความเสี่ยงที่อาจเกิดขึ้นของ CRISPR และเทคโนโลยีชีวภาพที่เกี่ยวข้องได้รับการกล่าวถึงเมื่อเร็วๆ นี้[ 14 ]โดยเน้นที่ลักษณะสุ่มของกระบวนการควบคุมเซลล์
สถาบันRoslin แห่งมหาวิทยาลัยเอดินบะระ ได้ดัดแปลงพันธุกรรมหมูให้ต้านทานไวรัสที่ก่อให้เกิดโรคระบบสืบพันธุ์และระบบทางเดินหายใจในสุกรซึ่งสร้างความเสียหายแก่เกษตรกรผู้เลี้ยงหมูในสหรัฐอเมริกาและยุโรปเป็นมูลค่า 2.6 พันล้านดอลลาร์ต่อปี[ 15 ]
ในเดือนกุมภาพันธ์ พ.ศ. 2563 การทดลองในสหรัฐอเมริกาแสดงให้เห็นการแก้ไขยีน CRISPR อย่างปลอดภัยในผู้ป่วยมะเร็ง 3 ราย[ 16 ]ในปี พ.ศ. 2563 มะเขือเทศ Sicilian Rouge High GABA ซึ่งเป็นมะเขือเทศที่สร้างกรดอะมิโนที่กล่าวกันว่าช่วยส่งเสริมการผ่อนคลาย ได้รับการอนุมัติให้จำหน่ายในญี่ปุ่น[ 15 ]
ในปี 2021 ประเทศอังกฤษ (ไม่ใช่ส่วนอื่นๆ ของสหราชอาณาจักร) วางแผนที่จะยกเลิกข้อจำกัดเกี่ยวกับพืชและสัตว์ที่ได้รับการแก้ไขยีน โดยเปลี่ยนจาก กฎระเบียบที่สอดคล้องกับ สหภาพยุโรปไปเป็นกฎที่ใกล้เคียงกับของสหรัฐอเมริกาและประเทศอื่นๆ รายงานของคณะกรรมาธิการยุโรปในเดือนเมษายน 2021 พบว่ามี "ข้อบ่งชี้ที่ชัดเจน" ว่าระบอบการกำกับดูแลในปัจจุบันไม่เหมาะสมสำหรับการแก้ไขยีน[ 15 ]ต่อมาในปี 2021 นักวิจัยได้ประกาศทางเลือกอื่นแทน CRISPR ซึ่งเรียกว่าโปรตีนที่มีกิจกรรมนำทางโดยองค์ประกอบเคลื่อนที่ที่จำเป็น (OMEGA) รวมถึง IscB, IsrB และ TnpB เป็นเอนโดนิวคลีเอสที่พบในทรานสโพซอนและนำทางโดย ωRNA ขนาดเล็ก[ 17 ] [ 18 ]
พื้นหลัง
วิศวกรรมพันธุกรรมซึ่งเป็นวิธีการนำองค์ประกอบทางพันธุกรรมใหม่เข้าสู่สิ่งมีชีวิต มีมาตั้งแต่ทศวรรษ 1970 ข้อเสียอย่างหนึ่งของเทคโนโลยีนี้คือลักษณะสุ่มของ การแทรก DNA เข้าไปใน จีโนมของโฮสต์ซึ่งอาจทำให้ยีนอื่นๆ ภายในสิ่งมีชีวิตเสียหายหรือเปลี่ยนแปลงได้ อย่างไรก็ตาม มีการค้นพบวิธีการหลายวิธีที่กำหนดเป้าหมายยีนที่แทรกเข้าไปในตำแหน่งเฉพาะภายในจีโนมของสิ่งมีชีวิต[ 6 ]นอกจากนี้ยังทำให้สามารถแก้ไขลำดับเฉพาะภายในจีโนมได้ รวมถึงลดผลกระทบที่ไม่ตรงเป้าหมาย สิ่งนี้สามารถนำไปใช้เพื่อวัตถุประสงค์ในการวิจัย โดยการกำหนดเป้าหมายการกลายพันธุ์ไปยังยีนเฉพาะ และในการบำบัดด้วยยีน โดยการแทรกยีนที่ใช้งาน ได้ เข้าไปในสิ่งมีชีวิต และกำหนดเป้าหมายให้แทนที่ยีนที่บกพร่อง อาจเป็นไปได้ที่จะรักษาโรคทางพันธุกรรม บางชนิด
การกำหนดเป้าหมายยีน
การรวมตัวแบบโฮโมโลกัส
วิธีการในยุคแรกๆ ที่ใช้ในการกำหนดเป้าหมายยีนไปยังตำแหน่งเฉพาะภายในจีโนมของสิ่งมีชีวิต (เรียกว่าการกำหนดเป้าหมายยีน ) อาศัยการรวมตัวกันแบบโฮโมโลจัส (HR) [ 19 ]โดยการสร้างโครงสร้าง DNA ที่มีแม่แบบที่ตรงกับลำดับจีโนมเป้าหมาย ทำให้กระบวนการ HR ภายในเซลล์สามารถแทรกโครงสร้างนั้นในตำแหน่งที่ต้องการได้ การใช้วิธีนี้กับเซลล์ต้นกำเนิดตัวอ่อนนำไปสู่การพัฒนาหนูทรานส์เจนิกที่มียีนเป้าหมายถูกกำจัดออกไปนอกจากนี้ยังสามารถแทรกยีนหรือเปลี่ยนแปลงรูปแบบการแสดงออกของยีน ได้อีกด้วย [ 20 ]เพื่อเป็นการยกย่องการค้นพบวิธีการใช้การรวมตัวกันแบบโฮโมโลจัสเพื่อแนะนำการดัดแปลงทางพันธุกรรมในหนูผ่านเซลล์ต้นกำเนิดตัวอ่อนMario Capecchi , Martin EvansและOliver Smithies ได้รับ รางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ประจำปี2007 [ 21 ]
การกำหนดเป้าหมายแบบมีเงื่อนไข
หากยีนที่สำคัญถูกน็อคเอาท์ อาจเป็นอันตรายถึงชีวิตต่อสิ่งมีชีวิตได้ เพื่อศึกษาหน้าที่ของยีนเหล่านี้ จึง มีการใช้ รีคอมบิเนสเฉพาะตำแหน่ง (SSR) ซึ่งสองประเภทที่พบได้บ่อยที่สุดคือระบบCre-LoxPและFlp-FRT รีคอมบิเนส Creเป็นเอนไซม์ที่กำจัด DNA โดยการรีคอมบิเนชันแบบโฮโมโลจัสระหว่างลำดับการจับที่เรียกว่าไซต์ Lox-P ระบบ Flip-FRT ทำงานในลักษณะเดียวกัน โดยรีคอมบิเนส Flip จะจดจำลำดับ FRT โดยการผสมพันธุ์สิ่งมีชีวิตที่มีไซต์รีคอมบิเนสที่อยู่รอบยีนที่สนใจกับสิ่งมีชีวิตที่แสดง SSR ภายใต้การควบคุมของโปรโมเตอร์เฉพาะเนื้อเยื่อทำให้สามารถน็อคเอาท์หรือเปิดยีนได้เฉพาะในเซลล์บางเซลล์เท่านั้น เทคนิคเหล่านี้ยังถูกนำมาใช้เพื่อกำจัดยีนเครื่องหมายออกจากสัตว์ทรานส์เจนิก การปรับเปลี่ยนระบบเหล่านี้เพิ่มเติมทำให้นักวิจัยสามารถกระตุ้นการรีคอมบิเนชันได้ภายใต้เงื่อนไขบางอย่างเท่านั้น ทำให้สามารถน็อคเอาท์หรือแสดงยีนได้ในเวลาหรือขั้นตอนการพัฒนาที่ ต้องการ [ 20 ]
กระบวนการ
การซ่อมแซมรอยแตกสองเส้น

รูปแบบการแก้ไขจีโนมทั่วไปอาศัยแนวคิดของกลไก การซ่อมแซม การแตกหักของดีเอ็นเอแบบสองสาย (DSB) มีสองเส้นทางหลักที่ซ่อมแซม DSB ได้แก่การเชื่อมต่อปลายที่ไม่เหมือนกัน (NHEJ) และการซ่อมแซมแบบกำหนดทิศทางตามความเหมือนกัน (HDR) NHEJ ใช้เอนไซม์หลายชนิดเพื่อเชื่อมต่อปลายดีเอ็นเอโดยตรง ในขณะที่ HDR ที่แม่นยำกว่าจะใช้ลำดับที่เหมือนกันเป็นแม่แบบสำหรับการสร้างลำดับดีเอ็นเอที่หายไป ณ จุดแตกหักขึ้นใหม่ สามารถใช้ประโยชน์จากสิ่งนี้ได้โดยการสร้างเวกเตอร์ที่มีองค์ประกอบทางพันธุกรรมที่ต้องการภายในลำดับที่เหมือนกับลำดับข้างเคียงของ DSB ซึ่งจะส่งผลให้มีการแทรกการเปลี่ยนแปลงที่ต้องการที่ตำแหน่งของ DSB แม้ว่าการแก้ไขยีนโดยใช้ HDR จะคล้ายกับการกำหนดเป้าหมายยีนโดยใช้การรวมตัวแบบเหมือนกัน แต่อัตราการรวมตัวจะเพิ่มขึ้นอย่างน้อยสามเท่า[ 22 ]
นิวคลีเอสที่ถูกดัดแปลงทางวิศวกรรม

หัวใจสำคัญของการแก้ไขจีโนมคือการสร้าง DSB (การแตกหักของสายดีเอ็นเอสองสาย) ณ จุดเฉพาะภายในจีโนม เอนไซม์ตัดจำเพาะที่ใช้กันทั่วไปนั้นมีประสิทธิภาพในการตัดดีเอ็นเอ แต่โดยทั่วไปแล้วจะจดจำและตัดที่หลายตำแหน่ง เพื่อเอาชนะความท้าทายนี้และสร้าง DSB ที่จำเพาะเจาะจง จึงมีการค้นพบและพัฒนาเอนไซม์นิวคลีเอสสามกลุ่มที่แตกต่างกัน ได้แก่ ซิงค์ฟิงเกอร์นิวคลีเอส ( ZFNs ), ทรานสคริปชันแอคติเวเตอร์ไลค์เอฟเฟคเตอร์นิวคลีเอส ( TALEN ), เมกะนิวคลีเอส และระบบคลัสเตอร์รีเทคทีฟสเปซสั้นพาลินโดรม ( CRISPR /Cas9)
เมกะนิวคลีเอส
เมกะนิวคลีเอสซึ่งถูกค้นพบในช่วงปลายทศวรรษ 1980 เป็นเอนไซม์ใน กลุ่ม เอนโดนิวคลีเอสซึ่งมีลักษณะเฉพาะคือความสามารถในการจดจำและตัดลำดับ DNA ขนาดใหญ่ (ตั้งแต่ 14 ถึง 40 คู่เบส) [ 23 ]เมกะนิวคลีเอสที่แพร่หลายและเป็นที่รู้จักมากที่สุดคือโปรตีนในกลุ่ม LAGLIDADG ซึ่งได้รับชื่อมาจากลำดับกรดอะมิโน ที่อนุรักษ์ ไว้
เมกะนิวคลีเอส ซึ่งพบได้ทั่วไปในจุลินทรีย์ มีคุณสมบัติเฉพาะตัวคือมีลำดับการจดจำที่ยาวมาก (>14bp) ทำให้มีความจำเพาะตามธรรมชาติสูง[ 24 ] [ 25 ]อย่างไรก็ตาม แทบไม่มีโอกาสที่จะพบเมกะนิวคลีเอสที่ต้องการเพื่อทำงานกับลำดับ DNA ที่เลือกไว้โดยเฉพาะ เพื่อเอาชนะความท้าทายนี้ จึงมีการใช้การกลายพันธุ์และการคัดกรองแบบความเร็วสูง เพื่อสร้างเมกะนิวคลีเอสสายพันธุ์ต่างๆ ที่จดจำลำดับเฉพาะ [ 25 ] [ 26 ]บางคนสามารถรวมเมกะนิวคลีเอสต่างๆ เข้าด้วยกันและสร้างเอนไซม์ลูกผสมที่จดจำลำดับใหม่ได้[ 27 ] [ 28 ]บางคนพยายามเปลี่ยนแปลงกรดอะมิโนที่ทำปฏิกิริยากับ DNA ของเมกะนิวคลีเอสเพื่อออกแบบเมกะนิวคลีเอสที่จำเพาะต่อลำดับในวิธีการที่เรียกว่าเมกะนิวคลีเอสที่ออกแบบอย่างมีเหตุผล[ 29 ]อีกแนวทางหนึ่งเกี่ยวข้องกับการใช้แบบจำลองคอมพิวเตอร์เพื่อพยายามทำนายกิจกรรมของเมกะนิวคลีเอสที่ดัดแปลงและความจำเพาะของลำดับนิวคลีอิกที่จดจำได้อย่างแม่นยำที่สุด[ 30 ]
มีการสร้างธนาคารขนาดใหญ่ที่มีหน่วยโปรตีนหลายหมื่นหน่วย หน่วยเหล่านี้สามารถรวมกันเพื่อให้ได้เมกะนิวคลีเอสไคเมอริกที่จดจำไซต์เป้าหมาย ซึ่งจะช่วยให้มีเครื่องมือวิจัยและพัฒนาที่ตอบสนองความต้องการที่หลากหลาย (การวิจัยพื้นฐาน สุขภาพ การเกษตร อุตสาหกรรม พลังงาน ฯลฯ) ซึ่งรวมถึงการผลิตเมกะนิวคลีเอสสองชนิดในระดับอุตสาหกรรมที่สามารถตัดยีน XPC ของมนุษย์ได้ การกลายพันธุ์ในยีนนี้ส่งผลให้เกิด โรค Xeroderma pigmentosum ซึ่งเป็น โรคทางพันธุกรรมชนิดรุนแรงที่ทำให้ผู้ป่วยมีแนวโน้มที่จะเป็นมะเร็งผิวหนังและไหม้เมื่อใดก็ตามที่ผิวหนังสัมผัสกับรังสียูวี[ 31 ]
เมกะนิวคลีเอสมีข้อดีคือทำให้เกิดความเป็นพิษต่อเซลล์น้อยกว่าวิธีการต่างๆ เช่นซิงค์ฟิงเกอร์นิว คลีเอ ส (ZFN) ซึ่งอาจเป็นเพราะการจดจำลำดับ DNA ที่เข้มงวดกว่า[ 25 ]อย่างไรก็ตาม การสร้างเอนไซม์เฉพาะลำดับสำหรับลำดับที่เป็นไปได้ทั้งหมดนั้นมีราคาแพงและใช้เวลานาน เนื่องจากไม่ได้รับประโยชน์จากความเป็นไปได้แบบผสมผสานที่วิธีการต่างๆ เช่น ZFN และฟิวชั่นที่ใช้ TALEN ใช้ประโยชน์
นิวคลีเอสแบบซิงค์ฟิงเกอร์
ตรงกันข้ามกับเมกะนิวคลีเอส แนวคิดเบื้องหลังเทคโนโลยี ZFNs และ TALEN นั้นขึ้นอยู่กับโดเมนเร่งปฏิกิริยาการตัด DNA ที่ไม่จำเพาะ ซึ่งสามารถเชื่อมโยงกับเปปไทด์ที่จดจำลำดับ DNA เฉพาะ เช่น ซิงค์ฟิงเกอร์และตัวกระตุ้นการถอดรหัสคล้ายเอฟเฟกเตอร์ (TALEs) [ 32 ]ขั้นตอนแรกคือการค้นหาเอนโดนิวคลีเอสที่มีไซต์การจดจำ DNA และไซต์การตัดแยกจากกัน ซึ่งเป็นสถานการณ์ที่ไม่ค่อยพบในเอนไซม์จำกัด[ 32 ]เมื่อพบเอนไซม์นี้แล้ว ส่วนการตัดของมันสามารถแยกออกได้ ซึ่งจะไม่จำเพาะมากเพราะไม่มีความสามารถในการจดจำ จากนั้นส่วนนี้สามารถเชื่อมโยงกับเปปไทด์ที่จดจำลำดับซึ่งอาจนำไปสู่ความจำเพาะสูงมาก
โครงสร้างแบบนิ้วสังกะสีพบได้ในปัจจัยการถอดรหัส หลายชนิด ไอออนสังกะสีซึ่งพบในโปรตีนของมนุษย์ 8% มีบทบาทสำคัญในการจัดระเบโครงสร้างสามมิติของโปรตีน ในปัจจัยการถอดรหัส ไอออนสังกะสีมักจะอยู่ที่บริเวณจุดปฏิสัมพันธ์ระหว่างโปรตีนกับดีเอ็นเอ ซึ่งช่วยทำให้โครงสร้างแบบนิ้วสังกะสีมีความเสถียร ส่วนปลายด้านซีของแต่ละนิ้วมีหน้าที่ในการจดจำลำดับดีเอ็นเออย่างจำเพาะเจาะจง
ลำดับที่รู้จักนั้นสั้น ประกอบด้วยเบสคู่ประมาณ 3 คู่ แต่โดยการรวมซิงค์ฟิงเกอร์ 6 ถึง 8 ตัวที่มีไซต์การจดจำที่ได้รับการระบุลักษณะแล้ว สามารถสร้างโปรตีนเฉพาะสำหรับลำดับที่มีเบสคู่ประมาณ 20 คู่ได้ ดังนั้นจึงสามารถควบคุมการแสดงออกของยีนเฉพาะได้ มีการแสดงให้เห็นว่ากลยุทธ์นี้สามารถใช้เพื่อส่งเสริมกระบวนการสร้างหลอดเลือดในสัตว์ได้[ 33 ]นอกจากนี้ยังสามารถรวมโปรตีนที่สร้างขึ้นในลักษณะนี้กับโดเมนเร่งปฏิกิริยาของเอนโดนิวคลีเอสเพื่อเหนี่ยวนำให้เกิดการแตกของ DNA ที่กำหนดเป้าหมาย และด้วยเหตุนี้จึงสามารถใช้โปรตีนเหล่านี้เป็นเครื่องมือทางวิศวกรรมจีโนมได้[ 34 ]
โดยทั่วไปแล้ว วิธีการที่ใช้เกี่ยวข้องกับการเชื่อมโยงโปรตีนที่จับกับ DNA สองตัว – แต่ละตัวประกอบด้วยนิ้วสังกะสีที่เลือกไว้โดยเฉพาะ 3 ถึง 6 นิ้ว – กับโดเมนเร่งปฏิกิริยาของ เอนโดนิวคลีเอ ส FokIซึ่งจำเป็นต้องสร้างไดเมอร์เพื่อตัด DNA สองสาย โปรตีนทั้งสองจะจดจำลำดับ DNA สองลำดับที่ห่างกันเพียงไม่กี่นิวคลีโอไทด์ การเชื่อมโยงโปรตีนนิ้วสังกะสีทั้งสองเข้ากับลำดับที่เกี่ยวข้องจะทำให้โดเมน FokI ทั้งสองอยู่ใกล้กันมากขึ้น FokI ต้องการการสร้างไดเมอร์เพื่อให้มีกิจกรรมนิวคลีเอส และนั่นหมายความว่าความจำเพาะจะเพิ่มขึ้นอย่างมาก เนื่องจากนิวคลีเอสแต่ละตัวจะจดจำลำดับ DNA ที่ไม่ซ้ำกัน เพื่อเพิ่มผลกระทบนี้นิวคลีเอส FokIได้รับการออกแบบให้สามารถทำงานได้เฉพาะในรูปแบบเฮเทอโรไดเมอร์เท่านั้น[ 35 ]
มีการใช้แนวทางหลายวิธีในการออกแบบนิวคลีเอสแบบซิงค์ฟิงเกอร์เฉพาะสำหรับลำดับที่เลือก วิธีที่แพร่หลายที่สุดคือการรวมหน่วยซิงค์ฟิงเกอร์ที่มีความจำเพาะที่ทราบ (การประกอบแบบโมดูลาร์) มีการพัฒนาเทคนิคการคัดเลือกต่างๆ โดยใช้แบคทีเรีย ยีสต์ หรือเซลล์สัตว์เลี้ยงลูกด้วยนม เพื่อระบุชุดค่าผสมที่ให้ความจำเพาะที่ดีที่สุดและความทนทานต่อเซลล์ที่ดีที่สุด แม้ว่าจะยังไม่มีรายงานการจำแนกลักษณะกิจกรรมของนิวคลีเอสแบบซิงค์ฟิงเกอร์ทั่วทั้งจีโนมโดยตรง แต่การทดสอบที่วัดจำนวนรวมของการแตกของดีเอ็นเอแบบสองสายในเซลล์พบว่ามีการแตกดังกล่าวเพียงหนึ่งถึงสองครั้งเหนือระดับพื้นหลังในเซลล์ที่ได้รับการรักษาด้วยนิวคลีเอสแบบซิงค์ฟิงเกอร์ที่มีไซต์การจดจำแบบผสม 24 bp และโดเมนนิวคลีเอสFokI แบบเฮเทอโรไดเมอร์ที่จำเป็น [ 35 ]
นิวคลีเอสที่ทำงานในรูปแบบเฮเทอโรไดเมอร์จะช่วยหลีกเลี่ยงความเป็นไปได้ของการทำงานแบบโฮโมไดเมอร์ที่ไม่พึงประสงค์ และเพิ่มความจำเพาะของ DSB แม้ว่าส่วนของนิวคลีเอสในโครงสร้าง ZFN และ TALEN จะมีคุณสมบัติคล้ายคลึงกัน แต่ความแตกต่างระหว่างนิวคลีเอสที่ถูกสร้างขึ้นเหล่านี้อยู่ที่เปปไทด์ที่ใช้ในการจดจำ DNA ZFN ใช้ซิงค์ฟิงเกอร์ Cys2-His2 และโครงสร้าง TALEN ใช้ TALE โดเมนเปปไทด์ที่ใช้ในการจดจำ DNA ทั้งสองชนิดนี้มีลักษณะเฉพาะคือพบได้ตามธรรมชาติในรูปแบบผสมผสานในโปรตีนของพวกมัน ซิงค์ฟิงเกอร์ Cys2-His2 มักพบในรูปแบบซ้ำๆ ที่ห่างกัน 3 คู่เบส และพบในรูปแบบผสมผสานที่หลากหลายใน โปรตีนที่ทำปฏิกิริยากับ กรดนิวคลีอิก หลายชนิด เช่นปัจจัยการถอดรหัสแต่ละนิ้วของโดเมนซิงค์ฟิงเกอร์มีความเป็นอิสระอย่างสมบูรณ์ และความสามารถในการจับของนิ้วหนึ่งจะได้รับผลกระทบจากนิ้วข้างเคียง ในทางกลับกัน TALE พบในรูปแบบซ้ำๆ โดยมีอัตราส่วนการจดจำแบบหนึ่งต่อหนึ่งระหว่างกรดอะมิโนและคู่ของนิวคลีโอไทด์ที่ถูกจดจำ เนื่องจากทั้งนิ้วสังกะสีและ TALE เกิดขึ้นในรูปแบบซ้ำๆ จึงสามารถลองใช้การผสมผสานที่แตกต่างกันเพื่อสร้างความจำเพาะของลำดับที่หลากหลาย[ 24 ]นิ้วสังกะสีได้รับการยอมรับมากขึ้นในแง่เหล่านี้ และแนวทางต่างๆ เช่น การประกอบแบบโมดูลาร์ (โดยที่นิ้วสังกะสีที่สัมพันธ์กับลำดับสามตัวจะถูกติดเรียงกันเพื่อครอบคลุมลำดับที่ต้องการ) OPEN (การคัดเลือกโดเมนเปปไทด์ที่มีความเข้มงวดต่ำเทียบกับนิวคลีโอไทด์สามตัว ตามด้วยการคัดเลือกชุดเปปไทด์ที่มีความเข้มงวดสูงเทียบกับเป้าหมายสุดท้ายในระบบแบคทีเรีย) และการคัดกรองแบบวันไฮบริดของแบคทีเรียของไลบรารีนิ้วสังกะสี รวมถึงวิธีการอื่นๆ ได้ถูกนำมาใช้เพื่อสร้างนิวคลีเอสที่จำเพาะต่อไซต์
นิวคลีเอสแบบซิงค์ฟิงเกอร์เป็นเครื่องมือวิจัยและพัฒนาที่ถูกนำมาใช้ในการดัดแปลงจีโนมหลายชนิด โดยเฉพาะอย่างยิ่งในห้องปฏิบัติการของกลุ่มซิงค์ฟิงเกอร์คอนซอร์เทียม บริษัทSangamo BioSciences ของสหรัฐอเมริกา ใช้นิวคลีเอสแบบซิงค์ฟิงเกอร์เพื่อทำการวิจัยด้านวิศวกรรมพันธุกรรมของเซลล์ต้นกำเนิดและการดัดแปลงเซลล์ภูมิคุ้มกันเพื่อวัตถุประสงค์ในการรักษา[ 36 ] [ 37 ]ปัจจุบันเซลล์ T ลิมโฟไซต์ที่ได้รับการดัดแปลงกำลังอยู่ระหว่างการทดลองทางคลินิกระยะที่ 1 เพื่อรักษามะเร็งสมองชนิดหนึ่ง ( กลิโอบลาสโตมา ) และในการต่อสู้กับโรคเอดส์[ 35 ]
ทาเลน

เอนไซม์นิวคลีเอสที่กระตุ้นการถอดรหัส (TALENs) เป็นโปรตีนที่จับกับ DNA โดยเฉพาะ ซึ่งมีลำดับกรดอะมิโนซ้ำกัน 33 หรือ 34 ตัว TALENs เป็นเอนไซม์จำกัดเทียมที่ออกแบบโดยการรวมโดเมนตัด DNA ของนิวคลีเอสเข้ากับโดเมน TALE ซึ่งสามารถปรับแต่งให้จดจำลำดับ DNA เฉพาะได้ โปรตีนลูกผสมเหล่านี้ทำหน้าที่เป็น "กรรไกร DNA" ที่สามารถกำหนดเป้าหมายได้ง่ายสำหรับการแก้ไขยีน ซึ่งช่วยให้สามารถทำการดัดแปลงจีโนมแบบกำหนดเป้าหมาย เช่น การแทรก การลบ การซ่อมแซม และการแทนที่ลำดับในเซลล์ที่มีชีวิต[ 38 ]โดเมนที่จับกับ DNA ซึ่งสามารถออกแบบให้จับกับลำดับ DNA ที่ต้องการได้ มาจากTAL effectorsซึ่งเป็นโปรตีนที่จับกับ DNA ที่ถูกขับออกมาโดยXanthomanos app ซึ่งเป็นเชื้อก่อโรคในพืช TAL effectors ประกอบด้วยโดเมนที่ซ้ำกัน โดยแต่ละโดเมนมีลำดับกรดอะมิโน 34 ตัวที่อนุรักษ์ไว้ อย่างสูง และจดจำนิวคลีโอไทด์ DNA เพียงตัวเดียวภายในไซต์เป้าหมาย เอนไซม์นิวคลีเอสสามารถสร้างการแตกของสายดีเอ็นเอสองสายที่ตำแหน่งเป้าหมาย ซึ่งสามารถซ่อมแซมได้ด้วยกระบวนการเชื่อมต่อปลายที่ไม่เหมือนกัน (NHEJ) ที่มีโอกาสเกิดข้อผิดพลาดสูง ส่งผลให้เกิดการรบกวนยีนโดยการแทรกหรือลบส่วนเล็กๆ แต่ละส่วนที่ซ้ำกันจะถูกอนุรักษ์ไว้ ยกเว้นส่วนที่เรียกว่า repeat variable di-residues (RVDs) ที่ตำแหน่งกรดอะมิโนที่ 12 และ 13 RVDs เป็นตัวกำหนดลำดับดีเอ็นเอที่ TALE จะจับ ความสัมพันธ์แบบหนึ่งต่อหนึ่งที่เรียบง่ายระหว่างส่วนที่ซ้ำกันของ TALE กับลำดับดีเอ็นเอที่สอดคล้องกัน ทำให้กระบวนการประกอบอาร์เรย์ส่วนที่ซ้ำกันเพื่อจดจำลำดับดีเอ็นเอใหม่ๆ เป็นไปอย่างตรงไปตรงมา TALE เหล่านี้สามารถเชื่อมต่อกับโดเมนเร่งปฏิกิริยาจากเอนไซม์นิวคลีเอสดีเอ็นเอ FokI เพื่อสร้างเอนไซม์นิวคลีเอสที่ทำหน้าที่คล้ายตัวกระตุ้นการถอดรหัส (TALEN) โครงสร้าง TALEN ที่ได้นั้นรวมความจำเพาะและกิจกรรมเข้าด้วยกัน ทำให้เกิดเอนไซม์นิวคลีเอสที่จำเพาะต่อลำดับซึ่งจับและตัดลำดับดีเอ็นเอเฉพาะที่ตำแหน่งที่เลือกไว้ล่วงหน้าเท่านั้น ระบบการจดจำเป้าหมาย TALEN ใช้รหัสที่คาดเดาได้ง่าย นิวคลีเอส TAL มีความจำเพาะต่อเป้าหมายเนื่องจากความยาวของไซต์การจับที่ มีเบสคู่มากกว่า 30 คู่ TALEN สามารถดำเนินการได้ภายในช่วง 6 เบสคู่ของนิวคลีโอไทด์เดี่ยวใดๆ ในจีโนมทั้งหมด[ 39 ]
โครงสร้าง TALEN ถูกนำมาใช้ในลักษณะเดียวกับนิวคลีเอสซิงค์ฟิงเกอร์ที่ออกแบบไว้ และมีข้อดี 3 ประการในการกลายพันธุ์เป้าหมาย: (1) ความจำเพาะในการจับกับ DNA สูงกว่า (2) ผลกระทบนอกเป้าหมายต่ำกว่า และ (3) การสร้างโดเมนที่จับกับ DNA ทำได้ง่ายกว่า
คริสเปอร์
CRISPRs (Clustered Regularly Interspaced Short Palindromic Repeats) เป็นองค์ประกอบทางพันธุกรรมที่แบคทีเรียใช้เป็น ภูมิคุ้มกันชนิดหนึ่งเพื่อป้องกันไวรัส ประกอบด้วยลำดับสั้นๆ ที่มาจากจีโนมของไวรัสและถูกรวมเข้ากับจีโนมของแบคทีเรีย Cas (โปรตีนที่เกี่ยวข้องกับ CRISPR) จะประมวลผลลำดับเหล่านี้และตัดลำดับ DNA ของไวรัสที่ตรงกัน โดยการนำพลาสมิดที่มียีน Cas และ CRISPRs ที่สร้างขึ้นโดยเฉพาะเข้าไปใน เซลล์ ยูคาริโอต จีโนมของยูคาริโอตสามารถถูกตัดได้ที่ตำแหน่งใดก็ได้ตามต้องการ[ 40 ]
การแทรกจีโนมขนาดใหญ่
การปรับเปลี่ยนขนาดใหญ่ในอดีตนั้นไม่มีประสิทธิภาพและมีแนวโน้มที่จะเกิดข้อผิดพลาด แต่การพัฒนาล่าสุดทำให้การปรับเปลี่ยนขนาดใหญ่มีความน่าเชื่อถือมากขึ้น ตัวอย่างเช่น กลยุทธ์ pegRNA สี่เท่า (QuadPE) ช่วยให้นักวิจัยสามารถแทรกชิ้นส่วน DNA ขนาดใหญ่เข้าไปในเซลล์มนุษย์ได้อย่างเสถียร โดยมีขนาดตั้งแต่ 1.6 ถึง 26 kb ด้วยประสิทธิภาพประมาณ 40% [ 41 ]
การแก้ไขโดยการดัดแปลงนิวคลีโอเบส (การแก้ไขเบส)
หนึ่งในวิธีการแก้ไขกรดนิวคลีอิกที่มีประสิทธิภาพในยุคแรกๆ ใช้เอนไซม์ปรับเปลี่ยนนิวคลีโอเบสที่ควบคุมโดยลำดับนำทางของกรดนิวคลีอิก วิธีนี้ได้รับการอธิบายครั้งแรกในช่วงทศวรรษ 1990 และกลับมาได้รับความนิยมอีกครั้งเมื่อไม่นานมานี้[ 6 ] [ 42 ] [ 43 ] [ 44 ]วิธีนี้มีข้อดีคือไม่จำเป็นต้องทำลายสายดีเอ็นเอจีโนม จึงหลีกเลี่ยงการแทรกและการลบแบบสุ่มที่เกี่ยวข้องกับการทำลายสายดีเอ็นเอ วิธีนี้เหมาะสมสำหรับการแก้ไขที่แม่นยำซึ่งต้องการการเปลี่ยนแปลงนิวคลีโอไทด์เดี่ยวเท่านั้น และพบว่ามีประสิทธิภาพสูงสำหรับการแก้ไขประเภทนี้[ 44 ] [ 45 ]
อาร์คัท
ARCUT ย่อมาจาก artificial restriction DNA cutter ซึ่งเป็นเทคนิคที่พัฒนาโดย Makoto Komiyama วิธีนี้ใช้กรดนิวคลีอิกเปปไทด์แบบเสมือนเสริม (pcPNA) เพื่อระบุตำแหน่งการตัดภายในโครโมโซม เมื่อ pcPNA ระบุตำแหน่งแล้ว การตัดจะดำเนินการโดยใช้ซีเรียม (CE) และ EDTA (ส่วนผสมทางเคมี) ซึ่งทำหน้าที่เชื่อมต่อ[ 46 ]
ความแม่นยำและประสิทธิภาพของนิวคลีเอสที่ได้รับการดัดแปลงทางวิศวกรรม
วิธีการแก้ไขยีนด้วยเมกะนิวคลีเอสเป็นวิธีที่มีประสิทธิภาพน้อยที่สุดในบรรดาวิธีการที่กล่าวมาข้างต้น เนื่องจากลักษณะขององค์ประกอบการจับกับ DNA และองค์ประกอบการตัด ทำให้มีข้อจำกัดในการจดจำเป้าหมายที่เป็นไปได้เพียงหนึ่งเป้าหมายทุกๆ 1,000 นิวคลี โอไทด์ [ 12 ] ZFN ได้รับการพัฒนาขึ้นเพื่อเอาชนะข้อจำกัดของเมกะนิวคลีเอส จำนวนเป้าหมายที่เป็นไปได้ที่ ZFN สามารถจดจำได้เพิ่มขึ้นเป็นหนึ่งเป้าหมายทุกๆ 140 นิวคลีโอไทด์[ 12 ]อย่างไรก็ตาม ทั้งสองวิธีนั้นคาดเดาไม่ได้เนื่องจากองค์ประกอบการจับกับ DNA ของทั้งสองวิธีส่งผลกระทบต่อกันและกัน ส่งผลให้ต้องใช้ความเชี่ยวชาญในระดับสูงและกระบวนการตรวจสอบที่ยาวนานและมีค่าใช้จ่ายสูง
TALE นิวคลีเอส ซึ่งเป็นวิธีการที่แม่นยำและเฉพาะเจาะจงที่สุด ให้ประสิทธิภาพสูงกว่าสองวิธีแรก เนื่องจากองค์ประกอบที่จับกับ DNA ประกอบด้วยซับยูนิต TALE จำนวนมาก แต่ละซับยูนิตมีความสามารถในการจดจำสายนิวคลีโอไทด์ DNA ที่เฉพาะเจาะจงได้อย่างอิสระ ทำให้มีจำนวนไซต์เป้าหมายมากขึ้นด้วยความแม่นยำสูง การสร้าง TALE นิวคลีเอสใหม่ใช้เวลาประมาณหนึ่งสัปดาห์และมีค่าใช้จ่ายไม่กี่ร้อยดอลลาร์ โดยต้องอาศัยความเชี่ยวชาญเฉพาะด้านในชีววิทยาโมเลกุลและวิศวกรรมโปรตีน[ 12 ]
นิวคลีเอส CRISPR มีความแม่นยำต่ำกว่าเล็กน้อยเมื่อเทียบกับนิวคลีเอส TALE เนื่องจากจำเป็นต้องมีนิวคลีโอไทด์เฉพาะที่ปลายด้านหนึ่งเพื่อสร้าง RNA นำทางที่ CRISPR ใช้ในการซ่อมแซมการแตกของสายคู่ที่มันเหนี่ยวนำ ได้รับการพิสูจน์แล้วว่าเป็นวิธีที่เร็วและถูกที่สุด โดยมีค่าใช้จ่ายน้อยกว่าสองร้อยดอลลาร์และใช้เวลาเพียงไม่กี่วัน[ 12 ] CRISPR ยังต้องการความเชี่ยวชาญด้านชีววิทยาโมเลกุลน้อยที่สุด เนื่องจากการออกแบบนั้นอยู่ใน RNA นำทางแทนที่จะเป็นโปรตีน ข้อได้เปรียบที่สำคัญอย่างหนึ่งของ CRISPR เหนือวิธีการ ZFN และ TALEN คือสามารถกำหนดเป้าหมายไปยังลำดับ DNA ที่แตกต่างกันได้โดยใช้ sgRNA ของ CRISPR ประมาณ 80nt ในขณะที่วิธีการ ZFN และ TALEN ทั้งสองวิธีต้องมีการสร้างและทดสอบโปรตีนที่สร้างขึ้นเพื่อกำหนดเป้าหมายไปยังลำดับ DNA แต่ละลำดับ[ 47 ]
เนื่องจากการทำงานนอกเป้าหมายของนิวคลีเอสที่ทำงานอยู่อาจส่งผลเสียร้ายแรงต่อระดับพันธุกรรมและสิ่งมีชีวิต ความแม่นยำของเมกะนิวคลีเอส ZFN CRISPR และฟิวชั่นที่ใช้ TALEN จึงเป็นหัวข้อการวิจัยที่สำคัญ แม้ว่าจะมีการรายงานตัวเลขที่แตกต่างกัน แต่โดยทั่วไปแล้ว ZFN มักมีความเป็นพิษต่อเซลล์มากกว่าวิธีการ TALEN หรือนิวคลีเอสที่นำทางด้วย RNA ในขณะที่ TALEN และวิธีการนำทางด้วย RNA มักมีประสิทธิภาพสูงสุดและมีผลกระทบนอกเป้าหมายน้อยกว่า[ 48 ]จากระยะทางทางทฤษฎีสูงสุดระหว่างการจับกับ DNA และกิจกรรมของนิวคลีเอส วิธีการ TALEN จึงให้ความแม่นยำสูงสุด[ 12 ]
วิศวกรรมจีโนมอัตโนมัติแบบมัลติเพล็กซ์ (MAGE)

วิธีการสำหรับนักวิทยาศาสตร์และนักวิจัยที่ต้องการศึกษาความหลากหลายทางพันธุกรรมและฟีโนไทป์ที่เกี่ยวข้องทั้งหมดนั้นช้า แพง และไม่มีประสิทธิภาพ ก่อนการปฏิวัติครั้งใหม่นี้ นักวิจัยจะต้องทำการจัดการยีนเดี่ยวและปรับแต่งจีโนมทีละส่วนเล็กๆ สังเกตฟีโนไทป์ และเริ่มต้นกระบวนการใหม่ด้วยการจัดการยีนเดี่ยวที่แตกต่างกัน[ 49 ]ดังนั้น นักวิจัยที่สถาบัน Wyss แห่งมหาวิทยาลัยฮาร์วาร์ดจึงออกแบบ MAGE ซึ่งเป็นเทคโนโลยีที่มีประสิทธิภาพที่ช่วยปรับปรุงกระบวนการแก้ไขจีโนมในร่างกาย ช่วยให้สามารถจัดการจีโนมได้อย่างรวดเร็วและมีประสิทธิภาพ โดยทั้งหมดนี้เกิดขึ้นในเครื่องที่มีขนาดเล็กพอที่จะวางบนโต๊ะครัวขนาดเล็กได้ การกลายพันธุ์เหล่านั้นรวมกับความแปรผันที่เกิดขึ้นตามธรรมชาติในระหว่างการแบ่งเซลล์ ทำให้เกิดการกลายพันธุ์ของเซลล์หลายพันล้านครั้ง
ด้วยการผสมผสานทางเคมี ดีเอ็นเอสายเดี่ยวสังเคราะห์ (ssDNA) และกลุ่มของโอลิโกนิวคลีโอไทด์จะถูกนำเข้าสู่บริเวณเป้าหมายของเซลล์ ทำให้เกิดการเปลี่ยนแปลงทางพันธุกรรม กระบวนการแบบวนรอบนี้เกี่ยวข้องกับการเปลี่ยนแปลงของ ssDNA (โดยการใช้กระแสไฟฟ้า ) ตามด้วยการเจริญเติบโต ซึ่งในระหว่างนั้นโปรตีนการรวมตัวแบบโฮโมโลจัสของแบคทีริโอเฟจจะทำหน้าที่เป็นตัวกลางในการเชื่อมต่อ ssDNA กับเป้าหมายในจีโนม การทดลองที่มุ่งเป้าไปที่เครื่องหมายฟีโนไทป์ที่เลือกไว้จะถูกคัดกรองและระบุโดยการเพาะเลี้ยงเซลล์บนอาหารเลี้ยงเชื้อที่แตกต่างกัน แต่ละรอบใช้เวลาดำเนินการ 2.5 ชั่วโมง โดยต้องใช้เวลาเพิ่มเติมในการเพาะเลี้ยงเซลล์ไอโซจีนิกและระบุลักษณะการกลายพันธุ์ ด้วยการนำไลบรารีของ ssDNA ที่ก่อให้เกิดการกลายพันธุ์เข้าสู่หลายตำแหน่งอย่างต่อเนื่อง MAGE สามารถสร้างความหลากหลายทางพันธุกรรมแบบผสมผสานในประชากรเซลล์ได้ สามารถแก้ไขจีโนมได้มากถึง 50 ครั้ง ตั้งแต่คู่เบสของนิวคลีโอไทด์เดี่ยวไปจนถึงจีโนมทั้งหมดหรือเครือข่ายยีนพร้อมกัน โดยได้ผลลัพธ์ภายในไม่กี่วัน[ 49 ]
การทดลอง MAGE สามารถแบ่งออกเป็นสามประเภท โดยมีลักษณะเฉพาะตามระดับขนาดและความซับซ้อนที่แตกต่างกัน: (i) ไซต์เป้าหมายจำนวนมาก การกลายพันธุ์ทางพันธุกรรมเพียงครั้งเดียว; (ii) ไซต์เป้าหมายเดียว การกลายพันธุ์ทางพันธุกรรมหลายครั้ง; และ (iii) ไซต์เป้าหมายจำนวนมาก การกลายพันธุ์ทางพันธุกรรมหลายครั้ง[ 49 ]ตัวอย่างของประเภทที่สามสะท้อนให้เห็นในปี 2552 ซึ่ง Church และเพื่อนร่วมงานสามารถตั้งโปรแกรมEscherichia coliให้ผลิตไลโคปีนได้มากกว่าปกติถึงห้าเท่า ซึ่งเป็นสารต้านอนุมูลอิสระที่พบได้ในเมล็ดมะเขือเทศและเกี่ยวข้องกับคุณสมบัติต้านมะเร็ง พวกเขาใช้ MAGE เพื่อเพิ่มประสิทธิภาพ เส้นทางการเผาผลาญ 1-deoxy- D -xylulose 5-phosphate (DXP) ในEscherichia coliเพื่อผลิตไลโคปีนไอโซพรีนอยด์มากเกินไป พวกเขาใช้เวลาประมาณ 3 วันและวัสดุราคามากกว่า 1,000 ดอลลาร์เล็กน้อย ความง่าย ความรวดเร็ว และความคุ้มค่าในการเปลี่ยนแปลงจีโนมด้วยเทคโนโลยี MAGE สามารถพลิกโฉมวิธีการผลิตสารประกอบสำคัญในอุตสาหกรรมต่างๆ เช่น วิศวกรรมชีวภาพ พลังงานชีวภาพ วิศวกรรมชีวการแพทย์ ชีววิทยาเชิงสังเคราะห์ เภสัชกรรม เกษตรกรรม และเคมี
แอปพลิเคชัน
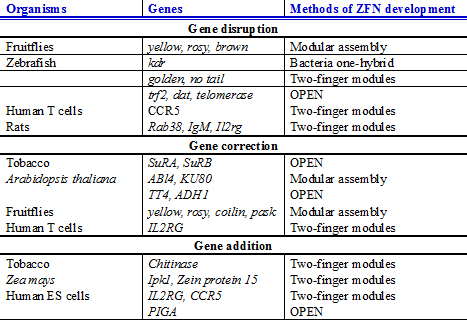
ในปี 2012 การแก้ไขจีโนมที่มีประสิทธิภาพได้รับการพัฒนาสำหรับระบบทดลองที่หลากหลาย ตั้งแต่พืชไปจนถึงสัตว์ ซึ่งมักจะเกินความสนใจทางคลินิก และกำลังกลายเป็นกลยุทธ์การทดลองมาตรฐานในห้องปฏิบัติการวิจัย[ 50 ] การสร้างหนู ปลา ซีบราข้าวโพดและยาสูบกลาย พันธุ์โดยใช้ ZFN รุ่นล่าสุดและการปรับปรุงวิธีการที่ใช้ TALEN เป็นเครื่องยืนยันถึงความสำคัญของวิธีการเหล่านี้ และรายการก็กำลังขยายตัวอย่างรวดเร็ว การแก้ไขจีโนมด้วยนิวคลีเอสที่ได้รับการดัดแปลงทางวิศวกรรมน่าจะช่วยสนับสนุนหลายสาขาของวิทยาศาสตร์ชีวภาพ ตั้งแต่การศึกษาหน้าที่ของยีนในพืชและสัตว์ ไปจนถึงการบำบัดยีนในมนุษย์ ตัวอย่างเช่น สาขาชีววิทยาสังเคราะห์ซึ่งมีเป้าหมายในการดัดแปลงเซลล์และสิ่งมีชีวิตให้ทำงานใหม่ๆ น่าจะได้รับประโยชน์จากความสามารถของนิวคลีเอสที่ได้รับการดัดแปลงทางวิศวกรรมในการเพิ่มหรือลบองค์ประกอบทางจีโนม และสร้างระบบที่ซับซ้อน[ 50 ]นอกจากนี้ ยังสามารถศึกษาหน้าที่ของยีนได้โดยใช้เซลล์ต้นกำเนิดที่มีนิวคลีเอสที่ได้รับการดัดแปลงทางวิศวกรรม
ด้านล่างนี้คือตัวอย่างงานเฉพาะบางอย่างที่วิธีการนี้สามารถดำเนินการได้:
- การกลายพันธุ์ของยีนเป้าหมาย
- การบำบัดด้วยยีน
- การสร้างการจัดเรียงโครโมโซมใหม่
- ศึกษาการทำงานของยีนด้วยเซลล์ต้นกำเนิด
- สัตว์ดัดแปลงพันธุกรรม
- การติดฉลากยีนภายใน
- การเติมทรานส์ยีนเป้าหมาย
การดัดแปลงยีนเป้าหมายในสัตว์
การผสมผสานการค้นพบใหม่ๆ ในด้านวิศวกรรมพันธุกรรม โดยเฉพาะการแก้ไขยีน และการพัฒนาล่าสุดในเทคโนโลยีการสืบพันธุ์ของวัว (เช่น การเพาะเลี้ยงตัวอ่อน ในหลอดทดลอง ) ทำให้สามารถแก้ไขจีโนมได้โดยตรงในเซลล์ไข่ที่ได้รับการปฏิสนธิโดยใช้เอนโดนิวคลีเอสสังเคราะห์ที่มีความจำเพาะสูง เอนโดนิวคลีเอสที่นำทางด้วย RNA: คลัสเตอร์ที่มีการเว้นระยะห่างอย่างสม่ำเสมอของพาลินโดรมสั้นๆ ที่เกี่ยวข้องกับ Cas9 (CRISPR/Cas9) เป็นเครื่องมือใหม่ที่ช่วยเพิ่มขอบเขตของวิธีการที่มีอยู่ให้มากขึ้น โดยเฉพาะอย่าง ยิ่งเอนโดนิวคลีเอสที่ออกแบบโดย CRISPR/Cas9 ช่วยให้สามารถใช้ RNA นำทางหลายตัวสำหรับการกำจัดยีน (KO) พร้อมกันในขั้นตอนเดียวโดยการฉีดโดยตรงเข้าสู่ไซโตพลาสซึม (CDI) ในไซโกตของสัตว์เลี้ยงลูกด้วยนม[ 51 ]
นอกจากนี้ การแก้ไขยีนยังสามารถนำไปใช้กับปลาบางชนิดในการเพาะเลี้ยงสัตว์น้ำ เช่น ปลาแซลมอนแอตแลนติก การแก้ไขยีนในปลายังอยู่ในขั้นตอนการทดลอง แต่ความเป็นไปได้ ได้แก่ การเจริญเติบโต ความต้านทานต่อโรค ความเป็นหมัน การสืบพันธุ์ที่ควบคุมได้ และสี การคัดเลือกคุณลักษณะเหล่านี้จะช่วยให้สภาพแวดล้อมมีความยั่งยืนมากขึ้นและมีสวัสดิภาพที่ดีขึ้นสำหรับปลา[ 52 ]
ปลาแซลมอน AquAdvantageเป็นปลาแซลมอนแอตแลนติกที่ได้รับการดัดแปลงพันธุกรรมโดยAquaBounty Technologiesยีนควบคุมฮอร์โมนการเจริญเติบโตในปลาแซลมอนแอตแลนติกถูกแทนที่ด้วยยีนควบคุมฮอร์โมนการเจริญเติบโตจากปลาแซลมอนชินุกแปซิฟิกและลำดับโปรโมเตอร์จากปลาโอเชียนพาวท์[ 53 ]
ด้วยการพัฒนาควบคู่กันไปของทรานสคริปโตมิกส์ระดับเซลล์เดี่ยวการแก้ไขจีโนม และแบบจำลองเซลล์ต้นกำเนิดใหม่ เรากำลังเข้าสู่ช่วงเวลาที่น่าตื่นเต้นทางวิทยาศาสตร์ ซึ่งพันธุศาสตร์เชิงฟังก์ชันไม่ได้จำกัดอยู่เฉพาะแบบจำลองสัตว์อีกต่อไป แต่สามารถดำเนินการได้โดยตรงในตัวอย่างของมนุษย์ การวิเคราะห์การแสดงออกของยีนระดับเซลล์เดี่ยวได้ไขปริศนาแผนที่การถอดรหัสของพัฒนาการของมนุษย์ ซึ่งกำลังระบุยีนเป้าหมายสำคัญสำหรับการศึกษาเชิงฟังก์ชัน การใช้ข้อมูลทรานสคริปโตมิกส์ทั่วโลกเพื่อเป็นแนวทางในการทดลอง เครื่องมือแก้ไขจีโนมแบบ CRISPR ทำให้สามารถรบกวนหรือกำจัดยีนสำคัญเพื่ออธิบายการทำงานในบริบทของมนุษย์ได้[ 54 ]
การดัดแปลงยีนเป้าหมายในพืช

การแก้ไขจีโนมโดยใช้Meganuclease [ 55 ] ZFNs และ TALEN เป็นกลยุทธ์ใหม่สำหรับการ ดัดแปลงพันธุกรรมในพืช และมีแนวโน้มที่จะช่วยในการสร้างลักษณะพืชที่ต้องการโดยการดัดแปลงยีนภายใน ตัวอย่างเช่น การเพิ่มยีนเฉพาะตำแหน่งในพืชเศรษฐกิจหลักสามารถใช้สำหรับ 'การซ้อนลักษณะ' ซึ่งลักษณะที่ต้องการหลายอย่างจะเชื่อมโยงกันทางกายภาพเพื่อให้แน่ใจว่ามีการแยกตัวร่วมกันในระหว่างกระบวนการผสมพันธุ์[ 35 ]ความคืบหน้าในกรณีดังกล่าวได้รับการรายงานเมื่อเร็ว ๆ นี้ในArabidopsis thaliana [ 56 ] [ 57 ] [ 58 ]และZea maysในArabidopsis thalianaการใช้การกำหนดเป้าหมายยีนโดยใช้ ZFN ช่วยเหลือ ยีนต้านทานสารกำจัดวัชพืชสองยีน (tobacco acetolactate synthase SuRA และ SuRB) ถูกนำเข้าสู่ตำแหน่ง SuR โดยมีเซลล์ที่เปลี่ยนแปลงที่มีการกลายพันธุ์สูงถึง 2% [ 56 ]ในZea maysการรบกวนตำแหน่งเป้าหมายเกิดขึ้นจาก DSB ที่เหนี่ยวนำโดย ZFN และ NHEJ ที่เกิดขึ้น ZFN ยังถูกใช้เพื่อขับเคลื่อนการแสดงออกของยีน ต้านทานสารกำจัดวัชพืช (PAT) เข้าสู่ตำแหน่งเป้าหมายภายใน IPK1 ในกรณีนี้ด้วย[ 59 ]การดัดแปลงจีโนมดังกล่าวที่สังเกตได้ในพืชที่สร้างใหม่ได้รับการพิสูจน์แล้วว่าสามารถถ่ายทอดทางพันธุกรรมและส่งต่อไปยังรุ่นต่อไปได้[ 59 ]ตัวอย่างที่อาจประสบความสำเร็จของการประยุกต์ใช้เทคนิคการแก้ไขจีโนมในการปรับปรุงพันธุ์พืชสามารถพบได้ในกล้วย ซึ่งนักวิทยาศาสตร์ใช้ การแก้ไข CRISPR/Cas9เพื่อทำให้ไวรัสโรคใบด่างกล้วยในจีโนม B ของกล้วย ( Musa spp. ) ไม่ทำงาน เพื่อเอาชนะความท้าทายที่สำคัญในการปรับปรุงพันธุ์กล้วย[ 60 ]
นอกจากนี้ วิศวกรรมจีโนมที่ใช้ TALEN ยังได้รับการทดสอบและปรับให้เหมาะสมอย่างกว้างขวางสำหรับการใช้งานในพืช[ 61 ]การรวม TALEN ยังถูกใช้โดยบริษัทส่วนผสมอาหารของสหรัฐฯ Calyxt [ 62 ]เพื่อปรับปรุงคุณภาพของผลิตภัณฑ์น้ำมันถั่วเหลือง[ 63 ]และเพื่อเพิ่มศักยภาพในการเก็บรักษามันฝรั่ง[ 64 ]
จำเป็นต้องมีการปรับปรุงหลายอย่างเพื่อปรับปรุงการแก้ไขจีโนมพืชโดยใช้การกำหนดเป้าหมายผ่าน ZFN [ 65 ]จำเป็นต้องมีการออกแบบนิวคลีเอสที่เชื่อถือได้และการทดสอบในภายหลัง การปราศจากความเป็นพิษของนิวคลีเอส การเลือกเนื้อเยื่อพืชที่เหมาะสมสำหรับการกำหนดเป้าหมาย เส้นทางการเหนี่ยวนำกิจกรรมของเอนไซม์ การปราศจากการกลายพันธุ์นอกเป้าหมายและการตรวจจับกรณีกลายพันธุ์ที่เชื่อถือได้[ 65 ]
วิธีการส่งมอบ CRISPR/Cas9 ทั่วไปในพืชคือการแปลงสภาพโดยใช้Agrobacterium [ 66 ] T-DNA ถูกนำเข้าสู่จีโนมของพืชโดยตรงด้วยกลไก T4SS คาสเซ็ตต์การแสดงออกที่ใช้ Cas9 และ gRNA จะถูกเปลี่ยนเป็นพลาสมิด Tiซึ่งจะถูกแปลงสภาพในAgrobacteriumเพื่อนำไปใช้กับพืช[ 66 ]เพื่อปรับปรุงการส่งมอบ Cas9 ในพืชที่มีชีวิต ไวรัสจึงถูกนำมาใช้เพื่อการส่งมอบทรานส์ยีนที่มีประสิทธิภาพมากขึ้น[ 66 ]
| ส่วนหนึ่งของบทความชุดเกี่ยวกับ |
| ชีววิทยาเชิงสังเคราะห์ |
|---|
| วงจรชีวภาพสังเคราะห์ |
| การแก้ไขจีโนม |
| เซลล์เทียม |
| ชีววิทยาต่างดาว |
| หัวข้ออื่นๆ |
วิจัย
การบำบัดด้วยยีน
แนวทาง การบำบัดด้วยยีนที่เหมาะสมที่สุดคือการแทนที่ยีนที่บกพร่องด้วยอัลลีลปกติในตำแหน่งตามธรรมชาติ ซึ่งมีข้อดีเหนือกว่าการส่งยีนด้วยไวรัส เนื่องจากไม่จำเป็นต้องรวมลำดับการเข้ารหัสและลำดับควบคุมทั้งหมดเมื่อจำเป็นต้องเปลี่ยนแปลงยีนเพียงบางส่วนเท่านั้น ซึ่งมักจะเป็นเช่นนั้น[ 67 ] [ 68 ]การแสดงออกของยีนที่ถูกแทนที่บางส่วนยังสอดคล้องกับชีววิทยาของเซลล์ปกติมากกว่ายีนทั้งหมดที่ถูกส่งผ่านเวกเตอร์ไวรัส
การใช้การแก้ไขจีโนมโดยใช้ TALEN ในทางคลินิกครั้งแรกเกิดขึ้นในการรักษาโรคมะเร็งเม็ดเลือดขาวชนิดเฉียบพลัน CD19+ ในเด็กอายุ 11 เดือนในปี 2558 เซลล์ T ของผู้บริจาคที่ได้รับการดัดแปลงได้รับการออกแบบให้โจมตีเซลล์มะเร็งเม็ดเลือดขาว ต้านทานต่อAlemtuzumabและหลีกเลี่ยงการตรวจจับโดยระบบภูมิคุ้มกันของโฮสต์หลังจากนำเข้า[ 69 ] [ 70 ]
มีการวิจัยอย่างกว้างขวางในเซลล์และสัตว์โดยใช้ CRISPR-Cas9 เพื่อพยายามแก้ไขการกลายพันธุ์ทางพันธุกรรมที่ทำให้เกิดโรคทางพันธุกรรม เช่น กลุ่มอาการดาวน์ กระดูกสันหลังเปิด ภาวะไม่มีสมอง และกลุ่มอาการเทอร์เนอร์และไคลน์เฟลเตอร์[ 71 ]
ในเดือนกุมภาพันธ์ พ.ศ. 2562 นักวิทยาศาสตร์การแพทย์ที่ทำงานร่วมกับSangamo Therapeuticsซึ่งมีสำนักงานใหญ่อยู่ที่ริชมอนด์ รัฐแคลิฟอร์เนีย ได้ประกาศ การบำบัดแก้ไขยีนของมนุษย์แบบ "ในร่างกาย" ครั้งแรกที่สามารถเปลี่ยนแปลงDNA ได้อย่างถาวร ในผู้ป่วยที่เป็นโรคฮันเตอร์[ 72 ]การทดลองทางคลินิกโดย Sangamo ที่เกี่ยวข้องกับการแก้ไขยีนโดยใช้Zinc Finger Nuclease (ZFN) กำลังดำเนินอยู่[ 73 ]
การกำจัดโรคภัยไข้เจ็บ
นักวิจัยได้ใช้ไดรฟ์ยีน CRISPR-Cas9 เพื่อดัดแปลงยีนที่เกี่ยวข้องกับภาวะเป็นหมันในA. gambiaeซึ่งเป็นพาหะนำโรคมาลาเรีย[ 74 ]เทคนิคนี้ยังมีนัยสำคัญในการกำจัดโรคที่เกิดจากพาหะนำโรคอื่นๆ เช่น ไข้เหลือง ไข้เลือดออก และซิกา[ 75 ]
ระบบ CRISPR-Cas9 สามารถตั้งโปรแกรมเพื่อปรับเปลี่ยนประชากรของแบคทีเรียสายพันธุ์ใดก็ได้โดยการกำหนดเป้าหมายจีโนไทป์ทางคลินิกหรือสายพันธุ์ที่แยกได้จากการระบาดวิทยา ระบบนี้สามารถเพิ่มจำนวนแบคทีเรียที่เป็นประโยชน์เหนือแบคทีเรียที่เป็นอันตรายได้โดยการกำจัดเชื้อก่อโรค ซึ่งทำให้ได้เปรียบยาปฏิชีวนะแบบออกฤทธิ์กว้าง[ 49 ]
การประยุกต์ใช้ยาต้านไวรัสสำหรับการบำบัดที่มุ่งเป้าไปที่ไวรัสในมนุษย์ เช่น HIV, เริม และไวรัสตับอักเสบ B กำลังอยู่ระหว่างการวิจัย CRISPR สามารถใช้เพื่อกำหนดเป้าหมายไวรัสหรือโฮสต์เพื่อขัดขวางยีนที่เข้ารหัสโปรตีนตัวรับบนพื้นผิวเซลล์ของไวรัส[ 47 ]ในเดือนพฤศจิกายน 2018 เหอ เจียนควิประกาศว่าเขาได้แก้ไขตัวอ่อนมนุษย์สองตัว เพื่อพยายามปิดใช้งานยีนCCR5ซึ่งเป็นรหัสสำหรับตัวรับที่HIVใช้ในการเข้าสู่เซลล์ เขากล่าวว่าเด็กหญิงฝาแฝดลูลู่และนานาเกิดเมื่อไม่กี่สัปดาห์ก่อนหน้านี้ เขากล่าวว่าเด็กหญิงทั้งสองยังคงมีสำเนาของ CCR5 ที่ใช้งานได้พร้อมกับ CCR5 ที่ถูกปิดใช้งาน ( โมเสก ) และยังคงมีความเสี่ยงต่อ HIV งานนี้ถูกประณามอย่างกว้างขวางว่าผิดจริยธรรม อันตราย และยังไม่ถึงเวลา[ 76 ]
ในเดือนมกราคม พ.ศ. 2562 นักวิทยาศาสตร์ในประเทศจีนรายงานการสร้างลิงโคลนนิ่งที่ เหมือนกัน 5 ตัวโดยใช้เทคนิคการโคลนนิ่งแบบเดียวกับที่ใช้กับ จงจงและหัวหัวซึ่งเป็นลิงโคลนนิ่งตัวแรก และแกะดอลลี่รวมถึงเทคนิคการตัดต่อยีนCrispr - Cas9 ที่กล่าวอ้างว่า เหอเจี้ยนกุยใช้ในการสร้างทารกมนุษย์ที่ได้รับการดัดแปลงพันธุกรรมเป็นครั้งแรกอย่างลูลู่และนานาลิงโคลนนิ่งเหล่านี้ถูกสร้างขึ้นเพื่อศึกษาโรคทางการแพทย์หลายชนิด[ 77 ] [ 78 ]
โอกาสและข้อจำกัด
ในอนาคต เป้าหมายสำคัญของการวิจัยเกี่ยวกับการแก้ไขจีโนมด้วยนิวคลีเอสที่ได้รับการดัดแปลงทางวิศวกรรมจะต้องเป็นการปรับปรุงความปลอดภัยและความจำเพาะของการทำงานของนิวคลีเอส[ 79 ]ตัวอย่างเช่น การปรับปรุงความสามารถในการตรวจจับเหตุการณ์นอกเป้าหมายสามารถปรับปรุงความสามารถของเราในการเรียนรู้วิธีการป้องกันเหตุการณ์เหล่านั้นได้ นอกจากนี้ นิ้วสังกะสีที่ใช้ใน ZFN มักจะไม่จำเพาะอย่างสมบูรณ์ และบางชนิดอาจทำให้เกิดปฏิกิริยาที่เป็นพิษ อย่างไรก็ตาม มีรายงานว่าความเป็นพิษลดลงได้ด้วยการดัดแปลงที่ทำกับโดเมนการตัดของ ZFN [ 68 ]
นอกจากนี้ การวิจัยของDana Carrollเกี่ยวกับการดัดแปลงจีโนมด้วยนิวคลีเอสที่ได้รับการดัดแปลงทางวิศวกรรมได้แสดงให้เห็นถึงความจำเป็นในการทำความเข้าใจกลไกการรวมตัวและการซ่อมแซมพื้นฐานของ DNA ให้ดียิ่งขึ้น ในอนาคต วิธีการที่เป็นไปได้ในการระบุเป้าหมายรองคือการจับปลายที่แตกหักจากเซลล์ที่แสดง ZFNs และจัดลำดับ DNA ที่อยู่ข้างเคียงโดยใช้การจัดลำดับแบบความเร็วสูง[ 68 ]
เนื่องจาก CRISPR ใช้งานง่ายและประหยัดต้นทุน จึงมีการวิจัยอย่างกว้างขวางเกี่ยวกับ CRISPR ในปัจจุบัน มีสิ่งพิมพ์เกี่ยวกับ CRISPR มากกว่า ZFN และ TALEN แม้ว่าการค้นพบ CRISPR จะเพิ่งเกิดขึ้นไม่นานก็ตาม[ 47 ]ทั้ง CRISPR และ TALEN ได้รับความนิยมในการนำไปใช้ในการผลิตขนาดใหญ่เนื่องจากมีความแม่นยำและมีประสิทธิภาพ
การแก้ไขจีโนมเกิดขึ้นได้เองตามธรรมชาติโดยไม่ต้องอาศัยวิศวกรรมพันธุกรรมเทียม ตัวการที่สามารถแก้ไขรหัสพันธุกรรมได้คือไวรัสหรือสาร RNA ขนาดเล็กที่เป็นส่วนประกอบของไวรัส
แม้ว่า DNA จะมีประสิทธิภาพมากกว่าวิธีการอื่นๆ หลายวิธีในพันธุศาสตร์ย้อนกลับ แต่ก็ยังไม่มีประสิทธิภาพสูงนัก ในหลายกรณี ประชากรที่ได้รับการรักษาเพียงไม่ถึงครึ่งเท่านั้นที่จะได้รับการเปลี่ยนแปลงที่ต้องการ[ 56 ]ตัวอย่างเช่น เมื่อวางแผนที่จะใช้ NHEJ ของเซลล์เพื่อสร้างการกลายพันธุ์ ระบบ HDR ของเซลล์ก็จะทำงานเพื่อแก้ไข DSB ด้วยอัตราการกลายพันธุ์ที่ต่ำกว่า
ตามธรรมเนียมแล้ว หนูเป็นสัตว์ทดลองที่นักวิจัยนิยมใช้มากที่สุดในการสร้างแบบจำลองโรค CRISPR สามารถช่วยเชื่อมช่องว่างระหว่างแบบจำลองนี้กับการทดลองทางคลินิกในมนุษย์ได้ โดยการสร้างแบบจำลองโรคแบบทรานส์เจนิกในสัตว์ขนาดใหญ่ เช่น หมู สุนัข และลิง[ 80 ] [ 81 ]การใช้ระบบ CRISPR-Cas9 โปรตีน Cas9 ที่ตั้งโปรแกรมไว้และ sgRNA สามารถนำเข้าโดยตรงไปยังไซโกตที่ได้รับการปฏิสนธิแล้ว เพื่อให้ได้การดัดแปลงยีนที่ต้องการเมื่อสร้างแบบจำลองทรานส์เจนิกในสัตว์ฟันแทะ วิธีนี้ช่วยให้ข้ามขั้นตอนการกำหนดเป้าหมายเซลล์ตามปกติในการสร้างสายพันธุ์ทรานส์เจนิก และส่งผลให้ลดเวลาในการสร้างลงได้ถึง 90% [ 81 ]
ศักยภาพหนึ่งที่ CRISPR นำมาซึ่งประสิทธิภาพคือการประยุกต์ใช้การปลูกถ่ายอวัยวะข้ามสายพันธุ์ ในการทดลองวิจัยก่อนหน้านี้ CRISPR ได้แสดงให้เห็นถึงความสามารถในการกำหนดเป้าหมายและกำจัดเรโทรไวรัสภายในร่างกาย ซึ่งช่วยลดความเสี่ยงในการแพร่กระจายโรคและลดอุปสรรคทางภูมิคุ้มกัน[ 47 ]การกำจัดปัญหาเหล่านี้ช่วยปรับปรุงการทำงานของอวัยวะผู้บริจาค ซึ่งทำให้การประยุกต์ใช้นี้ใกล้เคียงกับความเป็นจริงมากขึ้น
ในพืช การแก้ไขจีโนมถือเป็นวิธีแก้ปัญหาที่ใช้ได้ผลในการอนุรักษ์ความหลากหลายทางชีวภาพยีนไดรฟ์เป็นเครื่องมือที่มีศักยภาพในการเปลี่ยนแปลงอัตราการสืบพันธุ์ของสายพันธุ์รุกรานแม้ว่าจะมีความเสี่ยงที่เกี่ยวข้องอย่างมากก็ตาม[ 82 ]
การพัฒนาศักยภาพมนุษย์
นักทรานส์ฮิวแมนิสต์หลายคนมองว่าการแก้ไขจีโนมเป็นเครื่องมือที่มีศักยภาพสำหรับการพัฒนาศักยภาพของมนุษย์ [ 83 ] [ 84 ] [ 85 ] เดวิด แอนดรูว์ ซินแคลร์ นักชีววิทยาชาวออสเตรเลียและศาสตราจารย์ด้านพันธุศาสตร์กล่าวว่า " เทคโนโลยีใหม่ในการแก้ไขจีโนมจะทำให้สามารถนำไปใช้กับบุคคล (...) เพื่อให้มี (...) เด็กที่มีสุขภาพดีขึ้น" – เด็กที่ได้รับการออกแบบ [ 86 ] ตามรายงานเดือนกันยายน 2016 โดยสภา Nuffield ด้านชีวจริยธรรม ในอนาคตอาจเป็นไปได้ที่จะพัฒนาศักยภาพของมนุษย์ด้วยยีนจากสิ่งมีชีวิตอื่นหรือยีนสังเคราะห์ทั้งหมด เช่น เพื่อปรับปรุงการมองเห็นในเวลากลางคืนและประสาทสัมผัสในการดมกลิ่น [ 87 ] [ 88 ] จอร์จ เชิร์ชได้รวบรวมรายชื่อการดัดแปลงพันธุกรรมที่มีศักยภาพสำหรับลักษณะที่เป็นประโยชน์ เช่น ความต้องการนอน หลับน้อยลง การเปลี่ยนแปลงที่เกี่ยวข้องกับการรับรู้ที่ป้องกันโรคอัลไซเมอร์ ความต้านทานต่อโรค และ ความสามารถ ในการเรียนรู้ ที่เพิ่มขึ้น พร้อมกับการศึกษาที่เกี่ยวข้องและผลกระทบเชิงลบที่อาจเกิดขึ้น[ 89 ] [ 90 ]
สถาบันวิทยาศาสตร์แห่งชาติและสถาบันการแพทย์แห่งชาติของสหรัฐอเมริกาได้ออกรายงานในเดือนกุมภาพันธ์ พ.ศ. 2560 โดยให้การสนับสนุนอย่างมีเงื่อนไขต่อการแก้ไขจีโนมมนุษย์[ 91 ]พวกเขาแนะนำว่าการทดลองทางคลินิกสำหรับการแก้ไขจีโนมอาจได้รับอนุญาตในอนาคตเมื่อพบคำตอบสำหรับปัญหาด้านความปลอดภัยและประสิทธิภาพ "แต่เฉพาะในกรณีที่ร้ายแรงภายใต้การกำกับดูแลอย่างเข้มงวดเท่านั้น" [ 92 ]
ความเสี่ยง
ในการประเมินภัยคุกคามทั่วโลกประจำปี 2016 ของหน่วยข่าวกรองสหรัฐฯเจมส์ อาร์. แคลปเปอร์ผู้อำนวยการหน่วยข่าวกรองแห่งชาติสหรัฐฯได้ระบุว่าการแก้ไขจีโนมเป็นอาวุธทำลายล้างมวลชนที่มีศักยภาพ โดยระบุว่าการแก้ไขจีโนมที่ดำเนินการโดยประเทศที่มีมาตรฐานด้านกฎระเบียบหรือจริยธรรม "แตกต่างจากประเทศตะวันตก" อาจเพิ่มความเสี่ยงในการสร้างสารชีวภาพหรือผลิตภัณฑ์ที่เป็นอันตราย ตามคำแถลง การกระจายตัวอย่างกว้างขวาง ต้นทุนต่ำ และความเร็วในการพัฒนาเทคโนโลยีนี้ การนำไปใช้ในทางที่ผิดโดยเจตนาหรือไม่เจตนาอาจนำไปสู่ผลกระทบทางเศรษฐกิจและความมั่นคงของชาติในวงกว้าง[ 93 ] [ 94 ] [ 95 ]ตัวอย่างเช่น เทคโนโลยีเช่น CRISPR สามารถนำมาใช้สร้าง "ยุงนักฆ่า" ก่อให้เกิดโรคระบาดที่ทำลายพืชผลหลัก หรือสร้างไวรัสที่มีอันตรายร้ายแรงสูง[ 95 ]
จากรายงานของสภาจริยธรรมชีวภาพนัฟฟิลด์ (Nuffield Council on Bioethics ) เมื่อเดือนกันยายน 2016 ระบุว่า ความเรียบง่ายและต้นทุนต่ำของเครื่องมือในการแก้ไขรหัสพันธุกรรมจะทำให้บุคคลทั่วไป หรือ " นักประดิษฐ์ชีวภาพ " สามารถทำการทดลองของตนเองได้ ซึ่งก่อให้เกิดความเสี่ยงจากการปล่อยจุลินทรีย์ดัดแปลงพันธุกรรมสู่สิ่งแวดล้อม รายงานยังพบว่า ความเสี่ยงและผลประโยชน์ของการดัดแปลงจีโนมของบุคคล และการส่งต่อการเปลี่ยนแปลงเหล่านั้นไปยังรุ่นต่อๆ ไปนั้นมีความซับซ้อนมากจนต้องมีการตรวจสอบด้านจริยธรรมอย่างเร่งด่วน การดัดแปลงดังกล่าวอาจมีผลกระทบที่ไม่คาดคิด ซึ่งอาจเป็นอันตรายไม่เพียงแต่ต่อเด็กเท่านั้น แต่ยังรวมถึงลูกหลานในอนาคตด้วย เนื่องจากยีนที่เปลี่ยนแปลงไปจะอยู่ในอสุจิหรือไข่ของพวกเขา[ 87 ] [ 88 ]ในปี 2001 นักวิจัยชาวออสเตรเลีย โรนัลด์ แจ็กสัน และเอียน แรมชอว์ ถูกวิพากษ์วิจารณ์จากการตีพิมพ์บทความในวารสาร Journal of Virologyที่สำรวจความเป็นไปได้ในการควบคุมหนู ซึ่งเป็นศัตรูพืชที่สำคัญในออสเตรเลีย โดยการติดเชื้อ ไวรัส ไข้ทรพิษหนู ที่เปลี่ยนแปลงไป ซึ่งจะทำให้เป็นหมัน เนื่องจากข้อมูลที่ละเอียดอ่อนดังกล่าวอาจนำไปสู่การผลิตอาวุธชีวภาพโดยผู้ก่อการร้ายทางชีวภาพที่อาจใช้ความรู้ดังกล่าวในการสร้างสายพันธุ์ไวรัสไข้ทรพิษอื่นๆ ที่ดื้อต่อวัคซีน เช่นไข้ทรพิษซึ่งอาจส่งผลกระทบต่อมนุษย์ได้[ 88 ] [ 96 ]นอกจากนี้ ยังมีความกังวลเพิ่มเติมเกี่ยวกับความเสี่ยงทางนิเวศวิทยาของการปล่อยยีนไดรฟ์เข้าสู่ประชากรป่า[ 88 ] [ 97 ] [ 98 ]
รางวัลโนเบล
ในปี พ.ศ. 2550 รางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ได้มอบให้แก่Mario Capecchi , Martin EvansและOliver Smithies "สำหรับการค้นพบหลักการในการแนะนำการดัดแปลงยีนเฉพาะในหนูโดยใช้เซลล์ต้นกำเนิดตัวอ่อน" [ 21 ]
ในปี 2020 รางวัลโนเบลสาขาเคมีได้มอบให้แก่Emmanuelle CharpentierและJennifer Doudnaสำหรับ "การพัฒนาวิธีการแก้ไขจีโนม" [ 99 ]
ดูเพิ่มเติม
- คริสเปอร์/ซีเอฟ1
- หนูขนปุย
- การแก้ไขอาร์เอ็นเอ
- การแก้ไขเอพิเจโนม
- การตัดต่อหลัก
- ทรานสโพซอนในฐานะเครื่องมือทางพันธุกรรม
- เทคโนโลยีการคัดเลือกเชื้อพันธุ์
- NgAgoเป็นเอนโดนิวคลีเอส Argonaute ที่นำทางด้วย ssDNA
อ่านเพิ่มเติม
- Saurabh S (มีนาคม 2021). "การแก้ไขจีโนม: การปฏิวัติการปรับปรุงพันธุ์พืช". Plant Molecular Biology Reporte . 39 (4): 752– 772. doi : 10.1007/s11105-021-01286-7 . S2CID 233713026 .
- Iancu, Daniela (2023). "บทที่ 1 - การแก้ไขจีโนม: จากสุขภาพของมนุษย์สู่ "เด็กที่สมบูรณ์แบบ"ใน Hostiuc, Sorin (บรรณาธิการ). จริยธรรมทางคลินิก ณ จุดตัดของเทคโนโลยีทางพันธุกรรมและการสืบพันธุ์ (ฉบับที่ 2). สำนักพิมพ์ Academic Press. หน้า 1–32 . doi : 10.1016/B978-0-443-19045-2.00003-9 . ISBN 978-0-443-19045-2.
- "ฉบับพิเศษว่าด้วยการแก้ไขพันธุกรรมในเซลล์สืบพันธุ์ของมนุษย์ " จริยธรรมชีวภาพ34 2020
- "ยีนมนุษย์ที่ปรับแต่งได้: คำมั่นสัญญาและอันตรายใหม่" Scientific American สืบค้นเมื่อ21 กุมภาพันธ์ 2019
- Connor S (25 เมษายน 2557). "ความแตกแยกทางวิทยาศาสตร์ - ความก้าวหน้าด้านจีโนมมนุษย์แบ่งแยกอดีตเพื่อนร่วมงาน" . The Independent . สืบค้นเมื่อ11 กุมภาพันธ์ 2559 .
- "การแก้ไขจีโนมคืออะไร?" . yourgenome.org . สืบค้นเมื่อ25 มีนาคม 2025 .
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การแก้ไขจีโนม
การแก้ไขจีโนมหรือวิศวกรรมจีโนมหรือการแก้ไขยีน เป็น วิศวกรรมพันธุกรรมประเภทหนึ่งที่แทรก ลบ แก้ไข หรือแทนที่DNA ใน จีโนมของสิ่งมีชีวิต แตกต่างจากเทคนิควิศวกรรมพันธุกรรม ในยุคแรกๆ
ประวัติศาสตร์
การแก้ไขจีโนมได้รับการบุกเบิกในช่วงทศวรรษ 1990 [ 6 ] ก่อนการเกิดขึ้นของแพลตฟอร์มการแก้ไขยีนแบบนิวคลีเอสที่ใช้กันทั่วไปในปัจจุบัน แต่การใช้งานถูกจำกัดด้วยประสิทธิภาพการแก้ไขที่ต่ำ การแก้ไขจีโนมด้วยนิวคลีเอสที่ได้รับการดัดแปลงทางวิศวกรรม กล่าวคือ...
พื้นหลัง
วิศวกรรมพันธุกรรม ซึ่งเป็นวิธีการนำองค์ประกอบทางพันธุกรรมใหม่เข้าสู่สิ่งมีชีวิต มีมาตั้งแต่ทศวรรษ 1970 ข้อเสียอย่างหนึ่งของเทคโนโลยีนี้คือลักษณะสุ่มของ การแทรก DNA เข้าไปใน จีโนม ของโฮสต์ซึ่งอาจทำให้ยีนอื่นๆ ภายในสิ่งมีชีวิตเสียหายหรือเปลี่ยนแปลงได้...
การกำหนดเป้าหมายยีน
วิธีการในยุคแรกๆ ที่ใช้ในการกำหนดเป้าหมายยีนไปยังตำแหน่งเฉพาะภายในจีโนมของสิ่งมีชีวิต (เรียกว่า การกำหนดเป้าหมายยีน ) อาศัย การรวมตัวกันแบบโฮโมโลจัส (HR) [ 19 ] โดยการสร้างโครงสร้าง DNA ที่มีแม่แบบที่ตรงกับลำดับจีโนมเป้าหมาย ทำให้กระบวนการ HR...