อ่าน 20 นาที
คอมเพล็กซ์ความเข้ากันได้ทางพันธุกรรมหลัก
คอมเพล็กซ์ ฮิสโตคอมแพติบิลิตีหลัก ( MHC ) เป็น บริเวณ ขนาดใหญ่ บนดีเอ็นเอของสัตว์มีกระดูกสันหลัง ซึ่งประกอบด้วยชุด ยีน ที่เชื่อมโยงกันอย่างใกล้ชิดและ มีลักษณะเป็นโพลีมอร์ฟิก...
คอมเพล็กซ์ความเข้ากันได้ทางพันธุกรรมหลัก
| โมเลกุลคอมเพล็กซ์ความเข้ากันได้ทางพันธุกรรมหลัก | |||||||
|---|---|---|---|---|---|---|---|
 โปรตีนคอมเพล็กซ์ฮิสโตคอมแพติบิลิตีหลัก (คลาส I) แสดงด้วยสีส้มและสีชมพู โดยมีเปปไทด์ที่แสดงด้วยสีแดง เยื่อหุ้มเซลล์แสดงด้วยสีเทา โดเมนทรานส์เมมเบรนและไซโทพลาสมิกแสดงในรูปแบบการ์ตูน ( PDB : 1hsa ) | |||||||
| ตัวระบุ | |||||||
| เครื่องหมาย | เอชแอลเอ | ||||||
| อินเตอร์โปร | IPR001039 | ||||||
| เมมเบรน | 63 | ||||||
| |||||||
คอมเพล็กซ์ฮิสโตคอมแพติบิลิตีหลัก ( MHC ) เป็นบริเวณ ขนาดใหญ่ บนดีเอ็นเอของสัตว์มีกระดูกสันหลัง ซึ่งประกอบด้วยชุดยีน ที่เชื่อมโยงกันอย่างใกล้ชิดและ มีลักษณะเป็นโพลีมอร์ฟิก โดยยีนเหล่านี้จะสร้าง โปรตีนบนพื้นผิว เซลล์ ซึ่ง จำเป็นต่อระบบภูมิคุ้มกันแบบปรับตัว โปรตีน บนพื้นผิวเซลล์เหล่านี้เรียกว่าโมเลกุล MHC
ชื่อของมันมาจากการค้นพบในระหว่างการศึกษาความเข้ากันได้ของเนื้อเยื่อที่ปลูกถ่าย[ 1 ]การศึกษาในภายหลังเปิดเผยว่าการปฏิเสธเนื้อเยื่อเนื่องจากความไม่เข้ากันเป็นเพียงแง่มุมหนึ่งของหน้าที่ทั้งหมดของโมเลกุล MHC ซึ่งก็คือการจับกับแอนติเจนที่ได้มาจากโปรตีนของร่างกายเองหรือจากเชื้อโรค และนำการนำเสนอแอนติเจนไปยังพื้นผิวเซลล์เพื่อให้เซลล์ T ที่เหมาะสมจดจำ ได้[ 2 ]โมเลกุล MHC เป็นตัวกลางในการโต้ตอบของเม็ดเลือด ขาว หรือที่เรียกว่าเซลล์เม็ดเลือดขาว (WBCs) กับเม็ดเลือดขาวอื่นๆ หรือกับเซลล์ของร่างกาย MHC กำหนดความเข้ากันได้ของผู้บริจาคสำหรับการปลูกถ่ายอวัยวะรวมถึงความไวต่อโรคภูมิต้านตนเองด้วย
ในเซลล์ โมเลกุล โปรตีน ของ ฟีโนไทป์ของโฮสต์เองหรือของเอนทิตีทางชีวภาพอื่นๆ จะถูกสังเคราะห์และสลายตัวอย่างต่อเนื่อง โมเลกุล MHC แต่ละตัวบนพื้นผิวเซลล์จะแสดงเปปไทด์ขนาดเล็ก (เศษส่วนโมเลกุลของโปรตีน) ที่เรียกว่าเอพิโทป [ 3 ] แอนติเจนของตัวเองที่นำเสนอจะป้องกันระบบภูมิคุ้มกันของสิ่งมีชีวิตจากการโจมตีเซลล์ของตัวเอง การนำเสนอโปรตีนที่ได้จากเชื้อโรคส่งผลให้ระบบภูมิคุ้มกันกำจัดเซลล์ที่ติดเชื้อ
ความหลากหลายของ การนำเสนอแอนติเจนของตนเองของแต่ละบุคคลซึ่งถูกควบคุมโดยแอนติเจน MHC เกิดขึ้นได้อย่างน้อยสามวิธี: (1) คลัง MHC ของสิ่งมีชีวิตเป็นแบบโพลีจีนิก (ผ่านยีนหลายตัวที่โต้ตอบกัน); (2) การแสดงออกของ MHC เป็นแบบโคโดมิแนนท์ (จากทั้งสองชุดของอัลลีล ที่สืบทอดมา); (3) ยีน MHC มีหลายรูปแบบ (แตกต่างกันไปในแต่ละสิ่งมีชีวิตภายในสปีชีส์ ) [ 4 ] มีการสังเกตการคัดเลือกทางเพศ ในหนูตัวผู้ ที่เลือกผสมพันธุ์กับหนูตัวเมียที่มี MHC ต่างกัน[ 5 ]นอกจากนี้ อย่างน้อยสำหรับการนำเสนอ MHC I ยังมีหลักฐานของการตัดต่อ เปปไทด์แอนติเจน ซึ่งสามารถรวมเปปไทด์จากโปรตีนที่แตกต่างกัน ทำให้ความหลากหลายของแอนติเจนเพิ่มขึ้นอย่างมาก[ 6 ]
การค้นพบ
คำอธิบายแรกของ MHC เกิดขึ้นโดยปีเตอร์ โกเรอร์นักภูมิคุ้มกันวิทยา ชาวอังกฤษ ในปี 1936 [ 7 ] ยีน MHC ถูกระบุครั้งแรกในหนูสายพันธุ์ผสมพันธุ์คลาเรนซ์ ลิตเติลปลูกถ่ายเนื้องอกข้ามสายพันธุ์ต่างๆ และพบว่ามีการปฏิเสธเนื้องอกที่ปลูกถ่ายตามสายพันธุ์ของผู้รับและผู้ให้[ 8 ]จอร์จ สเนลล์ผสมพันธุ์หนูสองสายพันธุ์อย่างเลือกสรร ได้สายพันธุ์ใหม่ที่เกือบจะเหมือนกับสายพันธุ์ต้นกำเนิด แต่แตกต่างกันอย่างสำคัญในความเข้ากันได้ของเนื้อเยื่อนั่นคือ ความเข้ากันได้ของเนื้อเยื่อเมื่อปลูกถ่าย และด้วยเหตุนี้จึงระบุตำแหน่ง ของ MHC ได้[ 9 ] ต่อมาฌอง ดอสเซต์ได้แสดงให้เห็นถึงการมีอยู่ของยีน MHC ในมนุษย์และอธิบายแอนติเจนเม็ดเลือดขาวของมนุษย์ตัวแรก ซึ่งเป็นโปรตีนที่เราเรียกว่า HLA-A2 ในปัจจุบัน หลายปีต่อมา บารูจ เบนาเซราฟแสดงให้เห็นว่ายีน MHC ที่มีลักษณะหลายรูปแบบไม่เพียงแต่กำหนดองค์ประกอบของแอนติเจนเฉพาะบุคคลเท่านั้น แต่ยังควบคุมปฏิสัมพันธ์ระหว่างเซลล์ต่างๆ ของระบบภูมิคุ้มกันอีกด้วย นักวิทยาศาสตร์ทั้งสามคนนี้ได้รับรางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ประจำปี 1980 [ 10 ]จากการค้นพบเกี่ยวกับ "โครงสร้างที่กำหนดโดยพันธุกรรมบนพื้นผิวเซลล์ที่ควบคุมปฏิกิริยาภูมิคุ้มกัน"
ลำดับและคำอธิบายประกอบ MHC ที่สมบูรณ์ครั้งแรกได้รับการตีพิมพ์สำหรับมนุษย์ในปี 1999 โดยกลุ่มศูนย์การจัดลำดับจากสหราชอาณาจักร สหรัฐอเมริกา และญี่ปุ่นในวารสารNature [ 11 ]มันเป็น "MHC เสมือน" เนื่องจากเป็นโมเสกจากบุคคลต่างๆ ตำแหน่ง MHC ที่สั้นกว่ามากจากไก่ได้รับการตีพิมพ์ในวารสาร Nature ฉบับเดียวกัน[ 12 ]มีการ จัดลำดับสายพันธุ์อื่นๆ อีกหลายชนิด และมีการศึกษาการวิวัฒนาการของ MHC เช่น ในโอพอสซัม หางสั้นสีเทา ( Monodelphis domestica ) ซึ่งเป็นสัตว์มี ถุงหน้าท้อง MHC มีความยาว 3.95 Mb ให้ผลลัพธ์เป็น 114 ยีน โดย 87 ยีนมีร่วมกับมนุษย์[ 13 ] ความแปรผัน ของจีโนไทป์ MHC ของสัตว์ มีถุงหน้าท้อง อยู่ระหว่างสัตว์เลี้ยงลูกด้วยนมยูเทอเรียนและนกซึ่งถือเป็นการเข้ารหัส MHC ขั้นต่ำ แต่มีโครงสร้างที่ใกล้เคียงกับสัตว์ที่ไม่ใช่สัตว์เลี้ยงลูก ด้วยนม มากกว่า ฐานข้อมูล IPD-MHC [ 14 ]ถูกสร้างขึ้นเพื่อเป็นแหล่งเก็บข้อมูลส่วนกลางสำหรับลำดับของ Major Histocompatibility Complex (MHC) จากหลายสายพันธุ์ ณ วันที่เผยแพร่ 19 ธันวาคม 2019 ฐานข้อมูลนี้มีข้อมูลเกี่ยวกับ 77 สายพันธุ์
ยีน
ตำแหน่ง MHC มีอยู่ในสัตว์มีกระดูกสันหลังที่มีขากรรไกร ทั้งหมด สันนิษฐานว่าเกิดขึ้นเมื่อประมาณ 450 ล้านปีก่อน[ 15 ]แม้ว่าจำนวนยีนที่รวมอยู่ใน MHC ของสปีชีส์ต่างๆ จะแตกต่างกัน แต่โครงสร้างโดยรวมของตำแหน่งนั้นค่อนข้างคล้ายกัน MHC ทั่วไปประกอบด้วยยีนและยีนเทียมประมาณหนึ่งร้อยยีน ซึ่งไม่ได้เกี่ยวข้องกับภูมิคุ้มกันทั้งหมด ในมนุษย์บริเวณ MHC อยู่บนโครโมโซม 6ระหว่างเครื่องหมายทาง พันธุกรรม MOGและCOL11A2 (จาก 6p22.1 ถึง 6p21.3 ประมาณ 29Mb ถึง 33Mb บนชุดประกอบ hg38) และประกอบด้วยยีน 224 ยีน ครอบคลุม 3.6 เมกะเบสแพร์ (3,600,000 เบส) [ 11 ]ประมาณครึ่งหนึ่งมีหน้าที่ทางภูมิคุ้มกันที่รู้จัก MHC ของมนุษย์ยังเรียกว่าคอมเพล็กซ์ HLA ( แอนติเจนเม็ดเลือดขาวของมนุษย์ ) (มักเรียกสั้นๆ ว่า HLA) ในทำนองเดียวกัน มี SLA (แอนติเจนเม็ดเลือดขาวของสุกร), BoLA (แอนติเจนเม็ดเลือดขาวของวัว), DLA สำหรับสุนัข เป็นต้น อย่างไรก็ตาม ในอดีต MHC ใน หนูเรียกว่าระบบความเข้ากันได้ทางเนื้อเยื่อ 2 หรือ H-2 ในขณะที่ในหนูแรตเรียกว่าคอมเพล็กซ์ RT1 และในไก่เรียกว่าตำแหน่ง B
ตระกูลยีน MHC แบ่งออกเป็นสามกลุ่มย่อย ได้แก่MHC คลาส I , MHC คลาส IIและMHC คลาส IIIในบรรดายีนทั้งหมดที่มีอยู่ใน MHC นั้น มียีนสองประเภทที่เข้ารหัสโปรตีน โมเลกุล MHC คลาส Iและ โมเลกุล MHC คลาส IIซึ่งเกี่ยวข้องโดยตรงกับการนำเสนอแอนติเจนยีนเหล่านี้มีความหลากหลายสูง โดยมีอัลลีลของ HLA คลาส I จำนวน 19031 อัลลีล และของ HLA คลาส II จำนวน 7183 อัลลีล ถูกบันทึกไว้สำหรับมนุษย์ในฐานข้อมูล IMGT [ 16 ]
| ระดับ | การเข้ารหัส | การแสดงออก |
|---|---|---|
| ฉัน | (1) โปรตีนที่จับกับเปปไทด์ ซึ่งเลือกลำดับกรดอะมิโนสั้นๆ สำหรับการนำเสนอแอนติเจนรวมทั้ง (2) โมเลกุลที่ช่วยในการประมวลผลแอนติเจน (เช่นTAPและtapasin ) | สายโซ่หนึ่งเรียกว่า α ซึ่งมีตัวรับคือ ตัวรับ CD8ซึ่งพบได้ในเซลล์ T ชนิดทำลายเซลล์เป้าหมาย และตัวรับยับยั้งที่พบในเซลล์ NK |
| 2. | (1) โปรตีนที่จับกับเปปไทด์ และ (2) โปรตีนที่ช่วยในการบรรจุแอนติเจนลงบนโปรตีนที่จับกับเปปไทด์ของ MHC คลาส II (เช่นMHC II DM , MHC II DQ , MHC II DRและMHC II DP ) | ประกอบด้วยสองสายโซ่ เรียกว่า α และ β ซึ่งมีตัว รับ CD4ที่พบในเซลล์ทีช่วย (helper T cells) เป็น ลิแกนด์ |
| 3. | โปรตีนภูมิคุ้มกันอื่นๆ นอกเหนือจากการประมวลผลและการนำเสนอแอนติเจน เช่น ส่วนประกอบของระบบคอมพลีเมนต์ (เช่นC2 , C4 , แฟคเตอร์ B ) ไซโตไคน์ในการส่งสัญญาณภูมิคุ้มกัน (เช่นTNF-α ) และโปรตีนช็อกความร้อนที่ช่วยปกป้องเซลล์จากความเครียด | หลากหลาย |
โปรตีน

เอ็มเอชซี คลาส ไอ
โมเลกุล MHC คลาส Iพบได้ใน เซลล์ ที่มีนิวเคลียส ทุกเซลล์ รวมถึงเกล็ดเลือด ด้วย กล่าวคือ พบได้ในเซลล์ทุกเซลล์ยกเว้นเม็ดเลือดแดง MHC คลาส I ทำหน้าที่นำเสนออีพิโทปให้กับเซลล์ที ชนิดฆ่า (killer T cells) หรือที่เรียกว่า เซลล์ ทีชนิดทำลายเซลล์ ( cytotoxic T lymphocytesหรือ CTLs) CTL จะแสดงตัวรับ CD8 นอกเหนือจาก ตัวรับที เซลล์ (TCRs) เมื่อตัวรับ CD8 ของ CTL จับกับโมเลกุล MHC คลาส I หาก TCR ของ CTL เข้ากับอีพิโทปภายในโมเลกุล MHC คลาส I CTL จะกระตุ้นให้เซลล์นั้นตายตามโปรแกรมโดยกระบวนการอะพอพโทซิสดังนั้น MHC คลาส I จึงช่วยในการสร้างภูมิคุ้มกันระดับเซลล์ซึ่งเป็นวิธีการหลักในการจัดการกับเชื้อโรคภายในเซลล์เช่นไวรัสและแบคทีเรีย บางชนิด รวมถึงแบคทีเรียรูปแบบ LแบคทีเรียสกุลMycoplasmaและแบคทีเรียสกุลRickettsiaในมนุษย์ MHC คลาส I ประกอบด้วยโมเลกุล HLA-A , HLA-BและHLA-C
โครงสร้างผลึกแรกของโมเลกุล MHC คลาส I คือ HLA-A2 ของมนุษย์ ได้รับการตีพิมพ์ในปี 1989 [ 17 ]โครงสร้างดังกล่าวเผยให้เห็นว่าโมเลกุล MHC-I เป็นเฮเทอโรไดเมอร์ พวกมันมีซับยูนิตหนัก α ที่มีรูปแบบหลากหลาย ซึ่งยีนของมันอยู่ภายในโลคัส MHC และ ซับยูนิต ไมโครโกลบูลินβ2ขนาด เล็กที่ไม่เปลี่ยนแปลง ซึ่งยีนของมันมักจะอยู่ภายนอกโลคัส โซ่หนักที่มีรูปแบบหลากหลายของโมเลกุล MHC-I ประกอบด้วยบริเวณนอกเซลล์ที่ปลาย N ซึ่งประกอบด้วยสามโดเมน α1, α2 และ α3 เกลียวทรานส์เมมเบรนเพื่อยึดโมเลกุล MHC-I ไว้บนพื้นผิวเซลล์ และหางไซโตพลาสมิกสั้นๆ สองโดเมน α1 และ α2 สร้างร่องจับเปปไทด์ลึกระหว่างเกลียว α ยาวสองอัน และพื้นของร่องที่เกิดจากสาย β แปดสาย โดเมนคล้ายอิมมูโนโกลบูลิน α3 เกี่ยวข้องกับการโต้ตอบกับตัวรับร่วมCD8 β 2ไมโครโกลบูลินช่วยให้คอมเพล็กซ์มีความเสถียรและมีส่วนร่วมในการจดจำคอมเพล็กซ์เปปไทด์-MHC คลาส I โดยตัวรับร่วมCD8 [ 18 ] เปปไทด์จะจับกับ MHC-I โดยไม่มีพันธะโควาเลนต์ โดยจะถูกยึดไว้ด้วยช่องหลายช่องบนพื้นของร่องจับเปปไทด์ โซ่ข้างของกรดอะมิโนที่มีความหลากหลายมากที่สุดในอัลลีลของมนุษย์จะเติมเต็มส่วนกลางและส่วนที่กว้างที่สุดของร่องจับ ในขณะที่โซ่ข้างที่อนุรักษ์ไว้จะรวมกลุ่มกันที่ปลายที่แคบกว่าของร่อง

โมเลกุล MHC แบบคลาสสิกจะนำเสนออีพิโทปให้กับ TCR ของเซลล์ T CD8+ โมเลกุลที่ไม่ใช่แบบคลาสสิก (MHC คลาส IB) แสดงให้เห็นถึงความหลากหลายทางพันธุกรรม รูปแบบการแสดงออก และแอนติเจนที่นำเสนออย่างจำกัด กลุ่มนี้แบ่งย่อยออกเป็นกลุ่มที่เข้ารหัสภายในตำแหน่ง MHC (เช่น HLA-E, -F, -G) และกลุ่มที่ไม่ได้เข้ารหัส (เช่นลิแกนด์ที่เกิดจากความเครียดเช่น ULBPs, Rae1 และ H60) แอนติเจน/ลิแกนด์สำหรับโมเลกุลเหล่านี้จำนวนมากยังคงไม่เป็นที่รู้จัก แต่พวกมันสามารถโต้ตอบกับเซลล์ T CD8+ เซลล์ NKT และเซลล์ NK ได้ สายพันธุ์ MHC คลาส I ที่ไม่ใช่แบบคลาสสิกที่เก่าแก่ที่สุดในมนุษย์นั้นสันนิษฐานได้ว่าเป็นสายพันธุ์ที่รวมถึงโมเลกุล CD1 และ PROCR (หรือที่รู้จักกันในชื่อEPCR ) สายพันธุ์นี้อาจเกิดขึ้นก่อนกำเนิดของสัตว์มีกระดูกสันหลังสี่ขา[ 19 ]อย่างไรก็ตาม สายพันธุ์ MHC class I ที่ไม่ใช่แบบคลาสสิกเพียงสายพันธุ์เดียวที่มีหลักฐานว่าก่อตั้งขึ้นก่อนการแยกวิวัฒนาการของ Actinopterygii (ปลาครีบแข็ง) และ Sarcopterygii (ปลาครีบเนื้อและสัตว์สี่ขา) คือสายพันธุ์ Z ซึ่งสมาชิกพบร่วมกันในแต่ละสปีชีส์กับ MHC class I แบบคลาสสิกในปลาปอดและปลาครีบแข็งทั้งหมด[ 20 ]เหตุใดสายพันธุ์ Z จึงได้รับการอนุรักษ์ไว้อย่างดีในปลาครีบแข็งแต่สูญหายไปในสัตว์สี่ขาจึงยังไม่เป็นที่เข้าใจ
เอ็มเอชซี คลาส II
โมเลกุล MHC class IIสามารถแสดงออกได้แบบมีเงื่อนไขในเซลล์ทุกชนิด แต่โดยปกติแล้วจะพบเฉพาะในเซลล์นำเสนอแอนติเจน (APC) เฉพาะทาง ได้แก่แมโครฟาจเซลล์บีและโดยเฉพาะอย่างยิ่งเซลล์เดนดริติก (DC) เซลล์ APC จะรับ โปรตีน แอนติเจน เข้าไป ทำกระบวนการแอนติเจนและส่งคืนโมเลกุลส่วนหนึ่ง ซึ่งเรียกว่าเอพิโทปแล้วนำไปแสดงบนพื้นผิวของเซลล์ APC โดยเชื่อมต่อกับโมเลกุล MHC class II ( การนำเสนอแอนติเจน ) บนพื้นผิวของเซลล์ เอพิโทปสามารถถูกจดจำได้โดยโครงสร้างทางภูมิคุ้มกัน เช่นตัวรับทีเซลล์ (TCR) บริเวณโมเลกุลที่จับกับเอพิโทปเรียกว่าพาราโทป
บนพื้นผิวของเซลล์ทีเฮลเปอร์จะมีตัวรับ CD4 เช่นเดียวกับ TCR เมื่อโมเลกุล CD4 ของเซลล์ทีเฮลเปอร์ที่ยังไม่ได้รับการกระตุ้น (naive T cell) จับกับโมเลกุล MHC class II ของเซลล์นำเสนอแอนติเจน (APC) TCR ของเซลล์นั้นจะสามารถพบและจับกับเอพิโทปที่เชื่อมต่ออยู่ภายใน MHC class II ได้ เหตุการณ์นี้จะกระตุ้นเซลล์ทีที่ยังไม่ได้รับ การกระตุ้น นั้น ตามสภาพแวดล้อมเฉพาะที่ กล่าวคือ ความสมดุลของไซโตไคน์ที่หลั่งออกมาจาก APC ในสภาพแวดล้อมจุลภาคเซลล์ทีเฮลเปอร์ ที่ยังไม่ได้รับการกระตุ้น (Th 0 ) จะแบ่งตัวเป็นเซลล์ทีความจำ (memory Th cell) หรือเซลล์ทีออกฤทธิ์ (effector Th cell) ที่มีฟีโนไทป์เป็นประเภทที่ 1 (Th 1 ), ประเภทที่ 2 (Th 2 ), ประเภทที่ 17 (Th 17 ) หรือเซลล์ควบคุม/ยับยั้ง (T reg ) ซึ่งเป็นการแบ่งตัวขั้นสุดท้ายของเซลล์ทีที่ได้รับการระบุไว้แล้ว
ดังนั้น MHC class II จึงเป็นตัวกลางในการสร้างภูมิคุ้มกันต่อแอนติเจน หรือหาก APC กระตุ้นเซลล์ Th 0 ให้เปลี่ยนไปเป็น เซลล์ T reg เป็นหลัก ก็จะทำให้เกิด ความทนทานต่อภูมิคุ้มกันต่อแอนติเจน นั้น การเปลี่ยนแปลงขั้วในระหว่างการสัมผัสกับแอนติเจนครั้งแรกเป็นกุญแจสำคัญในการกำหนด โรคเรื้อรังหลายชนิดเช่นโรคอักเสบในลำไส้และโรคหอบหืดโดยการเบี่ยงเบนการตอบสนองทางภูมิคุ้มกันที่เซลล์ Th หน่วยความจำประสานงานเมื่อการเรียกคืนหน่วยความจำของพวกมันถูกกระตุ้นเมื่อสัมผัสกับแอนติเจนที่คล้ายกันเป็นครั้งที่สอง เซลล์ B แสดง MHC class II เพื่อนำเสนอแอนติเจนแก่เซลล์ Th 0แต่เมื่อตัวรับเซลล์ B ของพวกมัน จับกับอีพิโทปที่ตรงกัน ซึ่งเป็นการโต้ตอบที่ไม่ได้เกิดขึ้นผ่าน MHC เซลล์ B ที่ถูกกระตุ้น เหล่านี้ จะหลั่งอิมมูโนโกลบูลินที่ละลายได้ ซึ่งเป็นโมเลกุล แอนติบอดีที่เป็นตัวกลางในการสร้างภูมิคุ้มกันแบบฮิว โมรั ล
โมเลกุล MHC คลาส II ก็เป็นเฮเทอโรไดเมอร์เช่นกัน ยีนสำหรับซับยูนิต α และ β ต่างก็เป็นโพลีมอร์ฟิกและตั้งอยู่ในบริเวณย่อย MHC คลาส II ร่องการจับเปปไทด์ของโมเลกุล MHC-II เกิดจากโดเมน N-เทอร์มินัลของซับยูนิตทั้งสองของเฮเทอโรไดเมอร์ α 1และ β 1ซึ่งแตกต่างจากโมเลกุล MHC-I ที่เกี่ยวข้องกับสองโดเมนของสายโซ่เดียวกัน นอกจากนี้ ซับยูนิตทั้งสองของ MHC-II ยังมีเฮลิกซ์ทรานส์เมมเบรนและโดเมนอิมมูโนโกลบูลิน α 2หรือ β 2ที่สามารถจดจำได้โดยโครีเซปเตอร์CD4 [ 21 ]ด้วยวิธีนี้ โมเลกุล MHC จะนำทางชนิดของลิมโฟไซต์ที่อาจจับกับแอนติเจนที่กำหนดด้วยความสัมพันธ์สูง เนื่องจากลิมโฟไซต์ที่แตกต่างกันจะแสดงโครีเซปเตอร์ T-Cell (TCR) ที่แตกต่างกัน
โมเลกุล MHC คลาส II ในมนุษย์มีไอโซไทป์ ห้าถึงหกแบบ โมเลกุลแบบคลาสสิกนำเสนอเปปไทด์ให้กับลิมโฟไซต์ CD4+ โมเลกุลแบบไม่คลาสสิกหรือที่รู้จักกันในชื่อโมเลกุลเสริม มีหน้าที่ภายในเซลล์ พวกมันไม่ได้ถูกเปิดเผยบนเยื่อหุ้มเซลล์ แต่พบในเยื่อหุ้มภายใน ซึ่งพวกมันช่วยในการบรรจุเปปไทด์แอนติเจนลงบนโมเลกุล MHC คลาส II แบบคลาสสิก โมเลกุล MHC คลาส II แบบไม่คลาสสิกที่สำคัญอย่าง DM พบได้เฉพาะในระดับวิวัฒนาการของปลาปอด[ 22 ]แม้ว่าในปลาที่ดั้งเดิมกว่านั้นก็พบทั้ง MHC คลาส II แบบคลาสสิกและไม่คลาสสิกเช่นกัน[ 23 ] [ 24 ]
| ลำดับที่ | คุณสมบัติ[ 25 ] | เอ็มเอชซี คลาส I | เอ็มเอชซีคลาส II |
|---|---|---|---|
| 1 | ประกอบเป็นสายโซ่โพลีเปปไทด์ | สายโซ่ α (45 กิโลดาลตันในมนุษย์) สายโซ่ β 2 (12 กิโลดาลตันในมนุษย์) | สายโซ่ α (30–34 กิโลดาลตันในมนุษย์) สายโซ่เบตา (26–29 กิโลดาลตันในมนุษย์) |
| 2 | โดเมนจับแอนติเจน | โดเมน α 1และ α 2 | โดเมน α 1และ β 1 |
| 3 | จับกับโปรตีนแอนติเจนของ | กรดอะมิโนตกค้าง 8–10 ตัว | กรดอะมิโนตกค้าง 13–18 ตัว |
| 4 | รอยแยกการโค้งงอของเปปไทด์ | พื้นประกอบด้วยแผ่นเบต้า และด้านข้างประกอบด้วยแผ่นอัลฟา เกลียวที่ถูกปิดกั้นที่ปลายทั้งสองข้าง | พื้นประกอบด้วยแผ่นเบต้า และด้านข้างประกอบด้วยแผ่นอัลฟา เกลียวที่เปิดออกทั้งสองด้าน |
| 5 | ลวดลายเปปไทด์แอนติเจนิก เกี่ยวข้องกับการผูกมัด | สารตกค้างของจุดยึดที่ตำแหน่งอะมิโนและ ปลายคาร์บอน | เศษสมอเรือกระจายตัวอยู่เกือบสม่ำเสมอ ตามแนวเปปไทด์ |
| 6 | นำเสนอเปปไทด์แอนติเจนิกให้กับ | เซลล์ T CD8+ | เซลล์ T CD4+ |
เอ็มเอชซี คลาส III
แตกต่างจากโมเลกุลคลาส I และ II โมเลกุลคลาส III มีบทบาททางสรีรวิทยาและถูกเข้ารหัสอยู่ระหว่างคลาส I และ II บนแขนสั้นของโครโมโซมคู่ที่ 6 ของมนุษย์ โมเลกุลคลาส III ประกอบด้วยโปรตีนที่หลั่งออกมาหลายชนิดที่มีหน้าที่ทางภูมิคุ้มกัน ได้แก่ ส่วนประกอบของระบบคอมพลีเมนต์ (เช่นC2 , C4และB factor ), ไซโตไคน์ (เช่นTNF-α , LTAและLTB ) และโปรตีนช็อกความร้อน
การทำงาน
MHC คือแอนติเจนในเนื้อเยื่อที่ช่วยให้ระบบภูมิคุ้มกัน (โดยเฉพาะเซลล์ T) สามารถจับ รับรู้ และทนต่อตัวเองได้ (การรับรู้ตัวเอง) นอกจากนี้ MHC ยังทำหน้าที่เป็นตัวนำพาเปปไทด์ภายในเซลล์ที่จับกับ MHC และนำเสนอต่อตัวรับเซลล์ T (TCR) ในฐานะแอนติเจนแปลกปลอมที่อาจเกิดขึ้นได้ MHC มีปฏิสัมพันธ์กับ TCR และตัวรับร่วมของมันเพื่อปรับเงื่อนไขการจับให้เหมาะสมที่สุดสำหรับปฏิกิริยาระหว่าง TCR กับแอนติเจน ทั้งในแง่ของความสัมพันธ์และความจำเพาะในการจับแอนติเจน และประสิทธิภาพในการส่งสัญญาณ
โดยพื้นฐานแล้ว คอมplex MHC-peptide เป็นคอมplex ของ auto-antigen/allo-antigen เมื่อเกิดการจับกัน เซลล์ T ควรจะทนต่อ auto-antigen ได้ แต่จะทำงานเมื่อสัมผัสกับ allo-antigen สภาวะของโรคเกิดขึ้นเมื่อหลักการนี้ถูกรบกวน
การนำเสนอแอนติเจน : โมเลกุล MHC จับกับตัวรับเซลล์ Tและตัวรับร่วมCD4 / CD8 บน ลิมโฟไซต์ Tและ เอพิโท ป แอนติเจน ที่อยู่ในร่องจับเปปไทด์ของโมเลกุล MHC จะทำปฏิกิริยากับโดเมน Ig-Like ที่แปรผันได้ของ TCR เพื่อกระตุ้นการทำงานของเซลล์ T [ 26 ]
ปฏิกิริยาภูมิคุ้มกันตนเอง : การมีโมเลกุล MHC บางชนิดสามารถเพิ่มความเสี่ยงต่อโรคภูมิคุ้มกันตนเองได้มากกว่าชนิดอื่นๆHLA-B27 เป็นตัวอย่างหนึ่ง ยังไม่เป็นที่แน่ชัดว่าการมีเนื้อเยื่อชนิด HLA-B27 เพิ่มความเสี่ยงต่อ โรคกระดูกสันหลังอักเสบ และโรคอักเสบอื่นๆ ที่เกี่ยวข้อง ได้อย่างไรแต่มีการตั้งสมมติฐานเกี่ยวกับกลไกที่เกี่ยวข้องกับการนำเสนอแอนติเจนที่ผิดปกติหรือการกระตุ้นเซลล์ T
การจดจำเนื้อเยื่อต่างชนิด: โมเลกุล MHC ที่จับคู่กับอีพิโทปของเปปไทด์นั้นโดยพื้นฐานแล้วเป็นลิแกนด์สำหรับ TCR เซลล์ T จะถูกกระตุ้นโดยการจับกับร่องจับเปปไทด์ของโมเลกุล MHC ใดๆ ก็ตามที่พวกมันไม่ได้รับการฝึกฝนให้จดจำในระหว่างการคัดเลือกเชิงบวกในต่อมไท มั ส
การประมวลผลและการนำเสนอแอนติเจน
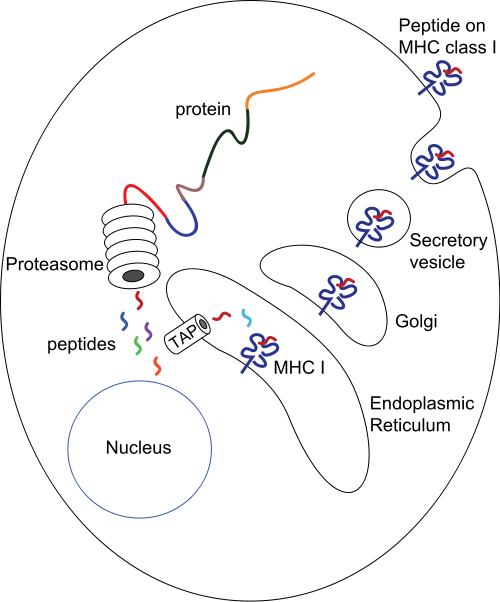
เปปไทด์จะถูกประมวลผลและนำเสนอโดยสองกระบวนการหลัก:
- ในMHC class IIเซลล์ฟาโกไซต์เช่นแมโครฟาจและเซลล์เดนไดรต์ ที่ยังไม่เจริญเต็มที่ จะดูดซับเอนทิตีโดย กระบวนการ ฟาโกไซโท ซิส เข้าไปในฟาโกโซม —แม้ว่าเซลล์ B จะแสดงกระบวนการ เอนโดไซ โทซิส ทั่วไปเข้าไปในเอนโดโซม —ซึ่งจะรวมเข้ากับไลโซโซมโดยเอนไซม์ที่เป็นกรดของไลโซโซมจะตัดโปรตีนที่ถูกดูดซับออกเป็นเปปไทด์หลายชนิด ผ่านพลวัตทางกายภาพและ เคมี ในการโต้ตอบระดับโมเลกุลกับ MHC class II ชนิดต่างๆ ที่มีอยู่ในโฮสต์ ซึ่งถูกเข้ารหัสไว้ในจีโนมของโฮสต์ เปปไทด์เฉพาะจะแสดงความเด่นทางภูมิคุ้มกันและโหลดลงบนโมเลกุล MHC class II โมเลกุลเหล่านี้จะถูกลำเลียงไปยังและแสดงออกภายนอกบนพื้นผิวเซลล์[ 27 ]
- ในMHC คลาส Iเซลล์ที่มีนิวเคลียสโดยทั่วไปจะแสดงเปปไทด์ในไซโตพลาสซึม ซึ่งส่วนใหญ่เป็นเปปไทด์ของเซลล์เองที่ได้มาจากกระบวนการหมุนเวียนของโปรตีนและผลิตภัณฑ์ไรโบโซมที่บกพร่อง ในระหว่างการติดเชื้อไวรัส การติดเชื้อจุลินทรีย์ภายในเซลล์ หรือการเปลี่ยนแปลงไปเป็นเซลล์มะเร็ง โปรตีนที่ถูกย่อยสลายในโปรตีโอโซม เหล่านี้ จะถูกบรรจุลงบนโมเลกุล MHC คลาส I และแสดงอยู่บนพื้นผิวเซลล์ เซลล์ T ลิมโฟไซต์สามารถตรวจจับเปปไทด์ที่แสดงอยู่ที่ 0.1–1% ของโมเลกุล MHC ได้

| ลักษณะเฉพาะ | เส้นทาง MHC-I | เส้นทาง MHC-II |
|---|---|---|
| องค์ประกอบของสารเชิงซ้อนเปปไทด์-MHC ที่เสถียร | ไมโครโกลบูลิน 2สายโซ่โพลีมอร์ฟิก α และ β โดยมีเปปไทด์จับกับสายโซ่ α | สายโซ่โพลีมอร์ฟิก α และ β โดยเปปไทด์จะจับกับทั้งสองสาย |
| ประเภทของเซลล์นำเสนอแอนติเจน (APC) | เซลล์ที่มีนิวเคลียสทั้งหมด | เซลล์เดนไดรติก , เซลล์ฟาโกไซต์แบบโมโนนิวเคลียร์, ลิมโฟไซต์บี , เซลล์บุผนังหลอดเลือดบางส่วน, เนื้อเยื่อบุผิวของต่อมไทมัส |
| ลิมโฟไซต์ T ที่สามารถตอบสนองได้ | เซลล์ทีลิมโฟไซต์ชนิดทำลายเซลล์เป้าหมาย (CD8+) | เซลล์ทีลิมโฟไซต์ผู้ช่วย (CD4+) |
| ที่มาของโปรตีนแอนติเจน | โปรตีน ในไซโตพลาสซึม (ส่วนใหญ่สังเคราะห์โดยเซลล์ อาจเข้ามาจากสิ่งแวดล้อมภายนอกเซลล์ผ่านทางฟาโกโซม ได้เช่นกัน ) | โปรตีนที่พบในเอนโดโซมหรือไลโซโซม (ส่วนใหญ่ถูกนำเข้ามาจากสิ่งแวดล้อมภายนอกเซลล์) |
| เอนไซม์ที่รับผิดชอบในการสร้างเปปไทด์ | โปรตีเอโซมในไซโตพลาสซึม | เอนไซม์โปรตีเอสจากเอนโดโซมและไลโซโซม (เช่นแคเทปซิน ) |
| ตำแหน่งการบรรจุเปปไทด์บนโมเลกุล MHC | เอนโดพลาสมิกเรติคูลัม | ช่องเวสิเคิลเฉพาะ |
| โมเลกุลที่มีส่วนเกี่ยวข้องในการขนส่งเปปไทด์และบรรจุเปปไทด์เหล่านั้นลงบนโมเลกุล MHC | TAP (transporter associated with antigen processing) | DM, โซ่ไม่เปลี่ยนแปลง |
ข้อจำกัดในการจดจำของทีลิมโฟไซต์
ในระหว่างการพัฒนาในต่อมไทมัส เซลล์ลิมโฟไซต์ T จะถูกคัดเลือกให้จดจำโมเลกุล MHC ของร่างกายเอง แต่ไม่จดจำแอนติเจนอื่นๆ ของร่างกาย หลังจากกระบวนการคัดเลือก เซลล์ลิมโฟไซต์ T แต่ละเซลล์จะแสดงความจำเพาะสองด้าน กล่าวคือ ตัวรับ T (TCR) จดจำ MHC ของร่างกายเอง แต่จดจำเฉพาะแอนติเจนที่ไม่ใช่ของร่างกายเท่านั้น
การจำกัดการทำงานของ MHC เกิดขึ้นระหว่างการพัฒนาของลิมโฟไซต์ในต่อมไทมัสผ่านกระบวนการที่เรียกว่าการคัดเลือกเชิงบวก (positive selection ) เซลล์ T ที่ไม่ได้รับสัญญาณการอยู่รอดเชิงบวก — ซึ่งส่วนใหญ่เกิดจากเซลล์เยื่อบุผิวของต่อมไทมัสที่นำเสนอเปปไทด์ของตนเองที่จับกับโมเลกุล MHC — ไปยัง TCR ของพวกมัน จะเกิดภาวะอะพอพโทซิส (apoptosis) การคัดเลือกเชิงบวกทำให้มั่นใจได้ว่าเซลล์ T ที่เจริญเต็มที่แล้วสามารถจดจำโมเลกุล MHC ได้อย่างมีประสิทธิภาพในบริเวณรอบนอก (เช่น ในส่วนอื่นๆ ของร่างกาย)
ตัวรับทีเซลล์ (TCR) ของทีลิมโฟไซต์จะจดจำเฉพาะอีพิโทปแบบเรียงลำดับหรือที่เรียกว่าอีพิโทปเชิงเส้น ซึ่งเป็นเปปไทด์เท่านั้น และเฉพาะเมื่อจับคู่กับโมเลกุล MHC เท่านั้น ( อย่างไรก็ตามโมเลกุลแอนติบอดีที่หลั่งออกมาจากบีเซลล์ที่ถูกกระตุ้น จะจดจำ อีพิ โทปที่หลากหลาย ทั้ง เปป ไทด์ ไขมันคาร์โบไฮเดรตและกรดนิวคลีอิกและจดจำอีพิโทปแบบมี โครงสร้าง สามมิติ )
ในการเลือกคู่ครองทางเพศ
โมเลกุล MHC ช่วยให้ระบบภูมิคุ้มกันเฝ้าระวังประชากรของโมเลกุลโปรตีนในเซลล์โฮสต์ และความหลากหลายของ MHC ที่มากขึ้นช่วยให้การนำเสนอแอนติเจน มีความหลากหลายมากขึ้น ในปี 1976 Yamazaki และคณะได้แสดงให้เห็นถึงการเลือกคู่ครองทางเพศ ของหนูตัวผู้สำหรับหนูตัวเมียที่มี MHC ที่แตกต่างกัน ผลลัพธ์ที่คล้ายกันนี้ได้รับจากปลา[ 29 ] ข้อมูลบางส่วนพบว่าอัตราการแท้งบุตรในช่วงต้น ของการตั้งครรภ์ ในคู่รักมนุษย์ที่มีพันธุกรรม MHC ที่แตกต่างกันนั้น ต่ำกว่า [ 30 ]
MHC อาจเกี่ยวข้องกับการเลือกคู่ครองในประชากรมนุษย์บางกลุ่ม ซึ่งเป็นทฤษฎีที่ได้รับการสนับสนุนจากการศึกษาของ Ober และคณะในปี 1997 [ 31 ]รวมถึง Chaix และคณะในปี 2008 [ 32 ]อย่างไรก็ตาม ผลการค้นพบหลังๆ นี้เป็นที่ถกเถียงกัน[ 33 ]หากมีอยู่จริง ปรากฏการณ์นี้อาจถูกควบคุมโดยการดมกลิ่นเนื่องจากฟีโนไทป์ MHC ดูเหมือนจะมีส่วนเกี่ยวข้องอย่างมากกับความแรงและความน่าพึงพอใจของกลิ่นที่รับรู้ของสารประกอบจากเหงื่อเอสเทอร์ ของ กรดไขมันเช่นเมทิลอันเดคาโนเอตเมทิลเดคาโน เอ ตเมทิลโนนาโน เอ ตเมทิลออกตาโน เอต และเมทิลเฮกซาโนเอต แสดงให้เห็นถึงความเชื่อมโยงที่แข็งแกร่งกับ MHC [ 34 ]
ในปี 1995 Claus Wedekindพบว่าในกลุ่มนักศึกษาหญิงที่ได้ดมกลิ่นเสื้อยืดที่นักศึกษาชายสวมใส่เป็นเวลาสองคืน (โดยไม่ใช้ผลิตภัณฑ์ระงับกลิ่นกาย น้ำหอม หรือสบู่ที่มีกลิ่นหอม) ผู้หญิงส่วนใหญ่เลือกเสื้อที่ผู้ชายที่มี MHC แตกต่างกันสวมใส่ ซึ่งความชอบนี้จะกลับกันหากผู้หญิงเหล่านั้นใช้ยาคุมกำเนิด[ 35 ]ในปี 2005 ในกลุ่มตัวอย่าง 58 คน ผู้หญิงมีความลังเลมากขึ้นเมื่อได้รับ MHC ที่คล้ายกับของตนเอง[ 36 ]แม้ว่าผู้หญิงที่ใช้ยาคุมกำเนิดจะไม่แสดงความชอบเป็นพิเศษ[ 37 ]ไม่มีงานวิจัยใดแสดงให้เห็นถึงขอบเขตที่ความชอบกลิ่นกำหนดการเลือกคู่ครอง (หรือในทางกลับกัน)
ความหลากหลายทางวิวัฒนาการ
สัตว์เลี้ยงลูกด้วยนมส่วนใหญ่มี MHC รูปแบบต่างๆ คล้ายกับของมนุษย์ ซึ่งมีความหลากหลายของอัลลีล อย่างมาก โดยเฉพาะอย่างยิ่งในยีนคลาสสิกทั้งเก้ายีน ซึ่งดูเหมือนว่าส่วนใหญ่เกิดจากการทำซ้ำของยีน แม้ว่า บริเวณ MHC ของมนุษย์จะมีpseudogenes จำนวนมาก ก็ตาม[ 38 ]ตำแหน่งที่มีความหลากหลายมากที่สุด ได้แก่ HLA-A, HLA-B และ HLA-C มีอัลลีลที่รู้จักประมาณ 6000, 7200 และ 5800 ตามลำดับ[ 39 ]อัลลีล HLA หลายตัวมีอายุเก่าแก่ บางครั้งมีความคล้ายคลึงกับอัลลีล MHC ของชิมแปนซีมากกว่าอัลลีลของมนุษย์บางตัวในยีนเดียวกัน[ 40 ]
ความหลากหลายของอัลลีล MHC เป็นความท้าทาย สำหรับ นักชีววิทยาวิวัฒนาการในการหาคำอธิบาย ส่วนใหญ่เสนอว่าเกิด จาก การคัดเลือกแบบสมดุล (ดูpolymorphism (biology) ) ซึ่งเป็น กระบวนการ คัดเลือกตามธรรมชาติที่ไม่มีอัลลีลใดเหมาะสมที่สุดอย่างแท้จริง เช่นการคัดเลือกที่ขึ้นอยู่กับความถี่[ 41 ]และความได้เปรียบของเฮเทอโรไซโกต์วิวัฒนาการร่วมของเชื้อโรค ซึ่งเป็นประเภทหนึ่งของการคัดเลือกแบบสมดุล ระบุว่าอัลลีลทั่วไปอยู่ภายใต้แรงกดดันจากเชื้อโรคมากที่สุด ผลักดันให้เกิดการคัดเลือกเชิงบวกของอัลลีลที่ไม่พบบ่อย—เป้าหมายที่เคลื่อนที่ได้สำหรับเชื้อโรค เมื่อแรงกดดันจากเชื้อโรคต่ออัลลีลที่เคยพบบ่อยลดลง ความถี่ของอัลลีลเหล่านั้นในประชากรจะคงที่ และยังคงหมุนเวียนอยู่ในประชากรขนาดใหญ่[ 42 ]การเปลี่ยนแปลงทางพันธุกรรมก็เป็นแรงผลักดันหลักในบางชนิดเช่นกัน[ 43 ] [ 44 ]เป็นไปได้ว่าผลรวมของปัจจัยเหล่านี้บางส่วนหรือทั้งหมดทำให้เกิดความหลากหลายทางพันธุกรรม[ 45 ]
นอกเหนือจากความหลากหลายทางพันธุกรรมของอัลลีลแล้ว ระดับความแตกต่างทางวิวัฒนาการระหว่างอัลลีล MHC ภายในแต่ละบุคคลยังมีอิทธิพลต่อการทำงานของระบบภูมิคุ้มกันอีกด้วย คู่อัลลีลที่แตกต่างกันมากขึ้นจะช่วยให้สามารถนำเสนอชุดเปปไทด์ที่กว้างขึ้นและซ้ำซ้อนน้อยลง ซึ่งอาจช่วยเพิ่มการจดจำภูมิคุ้มกันได้ แนวคิดนี้ บางครั้งเรียกว่าความแตกต่างทางวิวัฒนาการของ MHC หรือ HLA ขยายแบบจำลองคลาสสิกของความหลากหลายของ MHC และข้อได้เปรียบของเฮเทอโรไซโกต์โดยพิจารณาความแปรผันของอัลลีลเป็นแบบต่อเนื่องแทนที่จะเป็นสถานะไบนารี[ 46 ] [ 47 ] [ 48 ]
ความหลากหลายของ MHC ยังได้รับการเสนอแนะว่าเป็นตัวบ่งชี้ที่เป็นไปได้สำหรับการอนุรักษ์ เนื่องจากประชากรขนาดใหญ่และมีเสถียรภาพมักแสดงความหลากหลายของ MHC มากกว่าประชากรขนาดเล็กและแยกตัว[ 49 ] [ 50 ]ประชากรขนาดเล็กที่กระจัดกระจายซึ่งประสบกับภาวะคอขวดของประชากรมักมีความหลากหลายของ MHC ต่ำกว่า ตัวอย่างเช่น พบความหลากหลายของ MHC ที่ค่อนข้างต่ำในเสือชีตาห์ ( Acinonyx jubatus ) [ 51 ]บีเวอร์ยูเรเซีย ( Castor fiber ) [ 52 ]และแพนด้ายักษ์ ( Ailuropoda melanoleuca ) [ 53 ]ในปี 2550 ความหลากหลายของ MHC ที่ต่ำถูกระบุว่ามีบทบาทในความอ่อนแอต่อโรคในปีศาจแทสเมเนียน ( Sarcophilus harrisii ) ซึ่งเป็นสัตว์พื้นเมืองของเกาะแทสเมเนีย ที่แยกตัวออกไป โดยแอนติเจนของเนื้องอกที่ถ่ายทอดได้ ซึ่งเกี่ยวข้องกับโรคเนื้องอกที่ใบหน้าของปีศาจ ดูเหมือนจะถูกรับรู้ ว่าเป็นแอนติเจนของตัวเอง[ 54 ]เพื่อชดเชยการผสมพันธุ์ในสายเลือดเดียวกันได้มีการเสนอแนะให้พยายามรักษาความหลากหลายทางพันธุกรรมในประชากรของสัตว์ใกล้สูญพันธุ์และสัตว์ที่ถูกกักขัง
ในปลาที่มีครีบเป็นเส้น เช่น ปลาเทราต์สายรุ้ง ความหลากหลายทางพันธุกรรมของอัลลีลใน MHC คลาส II คล้ายคลึงกับที่พบในสัตว์เลี้ยงลูกด้วยนม และส่วนใหญ่จะอยู่ในร่องการจับเปปไทด์[ 55 ]อย่างไรก็ตาม ใน MHC คลาส I ของปลาเทเลออสหลายชนิด ความหลากหลายทางพันธุกรรมของอัลลีลนั้นรุนแรงกว่าในสัตว์เลี้ยงลูกด้วยนมมาก ในแง่ที่ว่าระดับความเหมือนของลำดับระหว่างอัลลีลอาจต่ำมาก และความแปรผันขยายออกไปไกลเกินกว่าร่องการจับเปปไทด์[ 55 ] [ 56 ] [ 20 ]มีการคาดการณ์ว่าความแปรผันของอัลลีล MHC คลาส I ประเภทนี้มีส่วนทำให้เกิดการปฏิเสธการปลูกถ่ายอวัยวะ ซึ่งอาจมีความสำคัญเป็นพิเศษในปลาเพื่อหลีกเลี่ยงการปลูกถ่ายเซลล์มะเร็งผ่านผิวหนังเยื่อบุ[ 57 ]
ตำแหน่ง MHC (6p21.3) มีตำแหน่งพาราโลจัสอีก 3 ตำแหน่งในจีโนมมนุษย์ ได้แก่ 19pl3.1, 9q33–q34 และ 1q21–q25 เชื่อกันว่าตำแหน่งเหล่านี้เกิดขึ้นจากการจำลองแบบสองรอบในสัตว์มีกระดูกสันหลังของตำแหน่ง ProtoMHC เดียว และการจัดระเบียบโดเมนใหม่ของยีน MHC เป็นผลมาจากการจำลองแบบซิสและการสับเปลี่ยนเอ็กซอนในภายหลังในกระบวนการที่เรียกว่า "MHC Big Bang" [ 58 ]ยีนในตำแหน่งนี้เชื่อมโยงกับภูมิคุ้มกันภายใน เซลล์ใน Metazoan พื้นฐานTrichoplax adhaerensอย่าง เห็นได้ชัด [ 59 ]
ในการปฏิเสธการปลูกถ่าย
ในขั้นตอนการปลูกถ่าย เช่น อวัยวะหรือสเต็มเซลล์โมเลกุล MHC ทำหน้าที่เป็นแอนติเจนและสามารถกระตุ้นการตอบสนองทางภูมิคุ้มกันในผู้รับ ทำให้เกิดการปฏิเสธการปลูกถ่าย โมเลกุล MHC ได้รับการระบุและตั้งชื่อตามบทบาทของมันในการ ปฏิเสธ การปลูกถ่ายระหว่างหนูสายพันธุ์ต่างๆ แม้ว่าจะใช้เวลากว่า 20 ปีในการชี้แจงบทบาทของ MHC ในการนำเสนอแอนติเจนเปปไทด์ให้กับลิมโฟไซต์ T ที่เป็นพิษต่อเซลล์ (CTLs) [ 60 ]
เซลล์ของมนุษย์แต่ละเซลล์แสดงออกถึงอัลลีล MHC คลาส I จำนวน 6 อัลลีล (อัลลีล HLA-A, -B และ -C จากพ่อและแม่คนละ 1 อัลลีล) และอัลลีล MHC คลาส II จำนวน 6-8 อัลลีล (อัลลีล HLA-DP และ -DQ และอัลลีล HLA-DR หนึ่งหรือสองอัลลีลจากพ่อและแม่คนละ 1 อัลลีล และการผสมผสานของอัลลีลเหล่านี้) ความหลากหลายของ MHC ในประชากรมนุษย์นั้นสูงมาก อย่างน้อย 350 อัลลีลสำหรับยีน HLA-A, 620 อัลลีลสำหรับ HLA-B, 400 อัลลีลสำหรับ DR และ 90 อัลลีลสำหรับ DQ บุคคลสองคนใดๆ ที่ไม่ใช่ฝาแฝดเหมือนกัน สามแฝด หรือการเกิดหลายคนมากกว่านั้น จะแสดงโมเลกุล MHC ที่แตกต่างกัน โมเลกุล MHC ทั้งหมดสามารถเป็นตัวกลางในการปฏิเสธการปลูกถ่าย แต่ HLA-C และ HLA-DP ซึ่งแสดงความหลากหลายทางพันธุกรรมต่ำ ดูเหมือนจะมีความสำคัญน้อยที่สุด
เมื่อเจริญเติบโตในต่อมไทมัส ลิมโฟไซต์ T จะถูกคัดเลือกจากความไม่สามารถของ TCR ในการจดจำแอนติเจนของตนเอง แต่ลิมโฟไซต์ T สามารถทำปฏิกิริยากับร่องจับเปปไทด์ ของ MHC ผู้ให้ ซึ่งเป็นบริเวณแปรผันของ MHC ที่จับอีพิโทปของแอนติเจนที่นำเสนอเพื่อให้ TCR หรือ พาราโทปที่เข้ากันจดจำได้ลิมโฟไซต์ T ของผู้รับจะถือว่าร่องจับเปปไทด์ที่ไม่เข้ากันนั้นเป็นแอนติเจนที่ไม่ใช่ของตนเอง
การปฏิเสธการปลูกถ่ายอวัยวะมีหลายประเภทที่ทราบกันว่ามีกลไกเกี่ยวข้องกับ MHC (HLA):
- การปฏิเสธแบบเฉียบพลันมากเกิดขึ้นเมื่อก่อนการปลูกถ่าย ผู้รับการปลูกถ่ายได้สร้างแอนติบอดีต่อ HLA ไว้แล้ว อาจเกิดจากการถ่ายเลือดมาก่อน (เนื้อเยื่อของผู้บริจาคที่มีลิมโฟไซต์ที่แสดงโมเลกุล HLA) จากแอนติบอดีต่อ HLA ที่สร้างขึ้นระหว่างตั้งครรภ์ (ซึ่งมุ่งเป้าไปที่ HLA ของพ่อที่แสดงโดยทารกในครรภ์) หรือจากการปลูกถ่ายครั้งก่อน
- การปฏิเสธเซลล์แบบเฉียบพลันเกิดขึ้นเมื่อลิมโฟไซต์ T ของผู้รับถูกกระตุ้นโดยเนื้อเยื่อของผู้บริจาค ทำให้เกิดความเสียหายผ่านกลไกต่างๆ เช่น การทำลายโดยตรงจากเซลล์ CD8
- การปฏิเสธเนื้อเยื่อแบบเฉียบพลันจากปฏิกิริยาของสารน้ำ และความผิดปกติเรื้อรังเกิดขึ้นเมื่อผู้รับการปลูกถ่ายสร้างแอนติบอดีต่อ HLA โดยมุ่งเป้าไปที่โมเลกุล HLA ที่มีอยู่บนเซลล์เยื่อบุผนังหลอดเลือดของเนื้อเยื่อที่ปลูกถ่าย
ในทุกสถานการณ์ข้างต้น ภูมิคุ้มกันจะมุ่งเป้าไปที่อวัยวะที่ปลูกถ่าย ทำให้เกิดความเสียหาย การทดสอบปฏิกิริยาข้ามสายพันธุ์ระหว่างเซลล์ผู้บริจาคและซีรั่มของผู้รับมีจุดมุ่งหมายเพื่อตรวจหาแอนติบอดีต่อ HLA ที่สร้างขึ้นแล้วในผู้รับ ซึ่งจะจดจำโมเลกุล HLA ของผู้บริจาค เพื่อป้องกันการปฏิเสธแบบเฉียบพลัน ในสถานการณ์ปกติ จะมีการประเมินความเข้ากันได้ระหว่างโมเลกุล HLA-A, -B และ -DR ยิ่งจำนวนความไม่เข้ากันสูงเท่าไร อัตราการรอดชีวิตในห้าปีก็จะยิ่งต่ำลงเท่านั้น ฐานข้อมูลผู้บริจาคทั่วโลกช่วยเพิ่มประสิทธิภาพในการค้นหาผู้บริจาคที่เข้ากันได้
การมีส่วนร่วมในการปฏิเสธการปลูกถ่ายแบบอัลโลจีนิกดูเหมือนจะเป็นคุณลักษณะดั้งเดิมของโมเลกุล MHC เนื่องจากมีการสังเกต ความสัมพันธ์ระหว่างการปฏิเสธการปลูกถ่ายและการจับคู่ (ไม่ตรงกัน) ของ MHC คลาส I [ 61 ] [ 62 ]และ MHC คลาส II [ 63 ] ในปลาด้วยเช่นกัน
ชีววิทยาของ HLA

โมเลกุล MHC คลาส I และ II ของมนุษย์เรียกอีกอย่างว่าแอนติเจนเม็ดเลือดขาวของมนุษย์ (HLA) เพื่อความชัดเจนในการใช้งาน เอกสารทางการแพทย์บางฉบับใช้ HLA เพื่อหมายถึงโมเลกุลโปรตีน HLA โดยเฉพาะ และสงวนคำว่า MHC ไว้สำหรับบริเวณของจีโนมที่เข้ารหัสโมเลกุลนี้ แต่การใช้คำเช่นนี้ไม่ได้เป็นไปอย่างสม่ำเสมอ
ยีน HLA ที่ได้รับการศึกษามากที่สุดคือยีน MHC คลาสสิกทั้งเก้ายีน ได้แก่HLA-A , HLA-B , HLA-C , HLA- DPA1 , HLA-DPB1 , HLA-DQA1 , HLA-DQB1, HLA-DRAและ HLA -DRB1ในมนุษย์ กลุ่มยีน MHC แบ่งออกเป็นสามกลุ่ม ได้แก่ กลุ่มที่ I, II และ III ยีน A, B และ C อยู่ในกลุ่ม MHC กลุ่มที่ I ในขณะที่ยีน D ทั้งหกยีนอยู่ในกลุ่มที่ II
อัลลีล MHC แสดงออกในลักษณะโคโดมิแนนท์[ 64 ]ซึ่งหมายความว่าอัลลีล (ตัวแปร) ที่ได้รับสืบทอดมาจากทั้งพ่อและแม่จะแสดงออกอย่างเท่าเทียมกัน:
- แต่ละคนมีอัลลีล 2 ตัวของยีนคลาส I ทั้ง 3 ยีน ( HLA-A, HLA-BและHLA-C ) ดังนั้นจึงสามารถแสดงออก MHC-I ได้ 6 ชนิดที่แตกต่างกัน (ดูภาพประกอบ)
- ในตำแหน่งยีนคลาส II แต่ละคนจะได้รับยีน HLA-DP คู่หนึ่ง (DPA1 และ DPB1 ซึ่งเข้ารหัสสายโซ่ α และ β) ยีนHLA-DQ คู่หนึ่ง ( DQA1และDQB1สำหรับสายโซ่ α และ β) ยีนHLA-DRα หนึ่งตัว ( DRA1 ) และยีน HLA-DRβหนึ่งตัวหรือมากกว่า( DRB1และDRB3, -4หรือ-5 ) นั่นหมายความว่าบุคคลที่เป็นเฮเทโรไซกัสสามารถได้รับอัลลีลคลาส II ที่ทำงานได้หกหรือแปดตัว โดยสามตัวหรือมากกว่าจากพ่อและแม่แต่ละคน บทบาทของDQA2หรือDQB2ยังไม่ได้รับการยืนยัน ส่วนDRB2, DRB6, DRB7, DRB8และDRB9เป็นยีนเทียม
ชุดของอัลลีลที่มีอยู่ในแต่ละโครโมโซมเรียกว่าแฮพลอไทป์ MHC ในมนุษย์ อัลลีล HLA แต่ละตัวจะถูกตั้งชื่อด้วยตัวเลข ตัวอย่างเช่น สำหรับบุคคลหนึ่ง แฮพลอไทป์ของเขาอาจเป็น HLA-A2, HLA-B5, HLA-DR3 เป็นต้น... บุคคลที่เป็นเฮเทโรไซกัสแต่ละคนจะมีแฮพลอไทป์ MHC สองแบบ แบบหนึ่งจากโครโมโซมของพ่อ และอีกแบบหนึ่งจากโครโมโซมของแม่
ยีน MHC มีความหลากหลายทางพันธุกรรมสูงมาก มีอัลลีลที่แตกต่างกันมากมายในแต่ละบุคคลภายในประชากรเดียวกัน ความหลากหลายทางพันธุกรรมนี้สูงมากจนในประชากรผสม (ที่ไม่แต่งงานในกลุ่มเดียวกัน ) ไม่มีบุคคลใดสองคนที่มีโมเลกุล MHC เหมือนกันทุกประการ ยกเว้นฝาแฝดเหมือนกัน
บริเวณที่มีความหลากหลายทางพันธุกรรมในแต่ละอัลลีลนั้นตั้งอยู่ในบริเวณที่เปปไทด์สามารถสัมผัสได้ ในบรรดาเปปไทด์ทั้งหมดที่ MHC สามารถแสดงได้นั้น มีเพียงส่วนน้อยเท่านั้นที่จะจับกับอัลลีล HLA ใดๆ ได้อย่างแข็งแรงพอ ดังนั้น การที่แต่ละยีนมีสองอัลลีล โดยแต่ละอัลลีลเข้ารหัสความจำเพาะต่อแอนติเจนที่ไม่ซ้ำกัน จะทำให้สามารถนำเสนอเปปไทด์ได้หลากหลายมากขึ้น
ในทางกลับกัน ภายในประชากร การมีอัลลีลที่แตกต่างกันมากมายทำให้มั่นใจได้ว่าจะมีบุคคลที่มีโมเลกุล MHC เฉพาะที่สามารถบรรจุเปปไทด์ที่ถูกต้องเพื่อจดจำจุลินทรีย์เฉพาะได้เสมอ วิวัฒนาการของความหลากหลายทางพันธุกรรมของ MHC ทำให้มั่นใจได้ว่าประชากรจะไม่พ่ายแพ้ต่อเชื้อโรคชนิดใหม่หรือเชื้อโรคที่กลายพันธุ์ เพราะอย่างน้อยที่สุดก็จะมีบุคคลบางส่วนที่สามารถพัฒนาการตอบสนองทางภูมิคุ้มกันที่เพียงพอเพื่อเอาชนะเชื้อโรคได้ การเปลี่ยนแปลงในโมเลกุล MHC (ซึ่งเป็นสาเหตุของความหลากหลายทางพันธุกรรม) เป็นผลมาจากการสืบทอดโมเลกุล MHC ที่แตกต่างกัน และไม่ได้เกิดจากการ รวมตัวกันใหม่เช่นเดียวกับกรณีของตัวรับ แอนติเจน
เนื่องจากความหลากหลายของอัลลีล ในระดับสูง ที่พบในยีน MHC จึงดึงดูดความสนใจของนักชีววิทยาวิวัฒนาการ หลายคน [ 65 ]รวมถึงผู้เชี่ยวชาญด้านโรค[ 66 ]
ดูเพิ่มเติม
- ภูมิคุ้มกันแบบอาศัยเซลล์
- การเลือกเพศที่ไม่เข้าคู่กัน
- ภูมิคุ้มกันแบบฮิวโมรัล
- เอ็มเอชซีมัลติเมอร์
- ฟีโรโมน
- สเตรปทาเมอร์
- การปฏิเสธการปลูกถ่าย
หมายเหตุและเอกสารอ้างอิง
- ^ Hull P (สิงหาคม 1970). "บันทึกเกี่ยวกับการสังเกตของดร. Snell เกี่ยวกับความหลากหลายทางพันธุกรรมของตำแหน่ง H-2" . Heredity . 25 (3): 461– 5. Bibcode : 1970Hered..25..461H . doi : 10.1038/hdy.1970.47 . PMID 5275401 .
- ^ Janeway Jr CA, Travers P, Walport M และคณะ (2001). "คอมเพล็กซ์ฮิสโตคอมแพติบิลิตีหลักและหน้าที่ของมัน"ภูมิคุ้มกันวิทยา: ระบบภูมิคุ้มกันในสุขภาพและโรค (ฉบับที่ 5). นิวยอร์ก: การ์แลนด์ ไซแอนซ์
- ^ Kimball JW (11 กุมภาพันธ์ 2011). "โมเลกุลฮิสโตคอมแพติบิลิตี" . Kimball's Biology Pages . เก็บถาวรจากต้นฉบับเมื่อ 4 กุมภาพันธ์ 2016.
- ^ Janeway Jr CA, Travers P, Walport M และคณะ (2001). "คอมเพล็กซ์ฮิสโตคอมแพติบิลิตีหลักและหน้าที่ของมัน"ภูมิคุ้มกันวิทยา: ระบบภูมิคุ้มกันในสุขภาพและโรค (ฉบับที่ 5). นิวยอร์ก: การ์แลนด์ ไซแอนซ์
- ^ Yamazaki K, Boyse EA, Miké V, Thaler HT, Mathieson BJ, Abbott J และคณะ (พฤศจิกายน 1976). "การควบคุมความชอบในการผสมพันธุ์ในหนูโดยยีนในคอมเพล็กซ์ฮิสโตคอมแพติบิลิตีหลัก"วารสารการแพทย์ทดลอง 144 ( 5): 1324– 35. doi : 10.1084/jem.144.5.1324 . PMC 2190468 . PMID 1032893 .
- ^ Vigneron N, Stroobant V, Chapiro J, Ooms A, Degiovanni G, Morel S และคณะ (เมษายน 2547). "เปปไทด์แอนติเจนิกที่ผลิตโดยการตัดต่อเปปไทด์ในโปรตีเอโซม" Science . 304 (5670): 587– 90. Bibcode : 2004Sci...304..587V . doi : 10.1126/science.1095522 . PMID 15001714 . S2CID 33796351 .
- ^ Klein J (1986). "เมล็ดพันธุ์แห่งกาลเวลา: ห้าสิบปีที่แล้ว Peter A. Gorer ค้นพบคอมเพล็กซ์ H-2" Immunogenetics . 24 (6): 331– 8. doi : 10.1007/bf00377947 . PMID 3539775 . S2CID 28211127 .
- ^ Little CC 1941, "พันธุศาสตร์ของการปลูกถ่ายเนื้องอก", หน้า 279–309, ใน Biology of the Laboratory Mouse , บรรณาธิการโดย Snell GD, นิวยอร์ก: Dover.
- ^ Snell GD, Higgins GF (พฤษภาคม 1951). "อัลลีลที่ตำแหน่งฮิสโตคอมแพติบิลิตี-2 ในหนูที่กำหนดโดยการปลูกถ่ายเนื้องอก"พันธุศาสตร์ 36 ( 3): 306– 10. doi : 10.1093/genetics/36.3.306 . PMC 1209522 . PMID 14840651 .
- ^ "รางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ ประจำปี 1980" 10 ตุลาคม 1980
สภาโนเบลแห่งสถาบันคาโรลินสกาได้ตัดสินใจในวันนี้ที่จะมอบรางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ประจำปี 1980 ร่วมกันให้แก่ บารุจ เบนาเซราฟ, ฌอง ดอสเซต์ และ จอร์จ สเนลล์
- ^ a bกลุ่มวิจัยการจัดลำดับ MHC (ตุลาคม 1999) "ลำดับสมบูรณ์และแผนที่ยีนของคอมเพล็กซ์ความเข้ากันได้ทางเนื้อเยื่อหลักของมนุษย์ กลุ่มวิจัยการจัดลำดับ MHC" Nature . 401 (6756): 921– 3. Bibcode : 1999Natur.401..921T . doi : 10.1038/44853 . PMID 10553908 . S2CID 186243515 .
- ^ Kaufman J, Milne S, Göbel TW, Walker BA, Jacob JP, Auffray C และคณะ (ตุลาคม 1999). "ตำแหน่ง B ของไก่เป็นคอมเพล็กซ์ฮิสโตคอมแพติบิลิตีหลักที่จำเป็นขั้นต่ำ" Nature . 401 (6756): 923– 5. Bibcode : 1999Natur.401..923K . doi : 10.1038/44856 . PMID 10553909 . S2CID 4387040 .
- ^ Belov K, Deakin JE, Papenfuss AT, Baker ML, Melman SD, Siddle HV และคณะ (มีนาคม 2549) "การสร้างซูเปอร์คอมเพล็กซ์ภูมิคุ้มกันของสัตว์เลี้ยงลูกด้วยนมบรรพบุรุษขึ้นใหม่จากคอมเพล็กซ์ฮิสโตคอมแพติบิลิตีหลักของสัตว์มีถุงหน้าท้อง" PLOS Biology 4 ( 3) e46. doi : 10.1371/journal.pbio.0040046 . PMC 1351924 . PMID 16435885 .
- ^ "ฐานข้อมูล IPD-MHC" . EMBL-EBI .
- ^ Kulski JK, Shiina T, Anzai T, Kohara S, Inoko H (ธันวาคม 2002). "การวิเคราะห์จีโนมเปรียบเทียบของ MHC: วิวัฒนาการของบล็อกการทำซ้ำคลาส I ความหลากหลายและความซับซ้อนจากฉลามสู่มนุษย์" Immunological Reviews . 190 (1): 95– 122. Bibcode : 2002ImRv..190...95K . doi : 10.1034/j.1600-065x.2002.19008.x . PMID 12493009 . S2CID 41765680 .
- ^ "ระบบข้อมูลพันธุศาสตร์ภูมิคุ้มกันระหว่างประเทศ" . เก็บถาวรจากต้นฉบับเมื่อ 2012-07-17 . เรียกดูเมื่อ2020-03-11 .
- ^ Saper MA, Bjorkman PJ, Wiley DC (พฤษภาคม 1991). "โครงสร้างที่ละเอียดขึ้นของแอนติเจนความเข้ากันได้ของเนื้อเยื่อมนุษย์ HLA-A2 ที่ความละเอียด 2.6 อังสตรอม" วารสารชีววิทยาโมเลกุล 219 ( 2): 277– 319. doi : 10.1016/0022-2836(91)90567-p . PMID 2038058 .
- ^ Gao GF, Tormo J, Gerth UC, Wyer JR, McMichael AJ, Stuart DI และคณะ (มิถุนายน 1997). "โครงสร้างผลึกของสารประกอบเชิงซ้อนระหว่าง CD8alpha(alpha) ของมนุษย์และ HLA-A2" Nature . 387 (6633): 630– 4. Bibcode : 1997Natur.387..630G . doi : 10.1038/42523 . PMID 9177355 . S2CID 4267617 .
- ^ Dijkstra JM, Yamaguchi T, Grimholt U (กรกฎาคม 2018). "การอนุรักษ์ลำดับโมทีฟบ่งชี้ว่าสายพันธุ์ MHC class I ที่ไม่ใช่แบบคลาสสิก CD1/PROCR และ UT ถูกสร้างขึ้นก่อนการกำเนิดของสัตว์สี่ขา" Immunogenetics . 70 (7): 459– 476. doi : 10.1007/s00251-017-1050-2 . PMID 29270774 . S2CID 24591879 .
- ^ a b Grimholt U, Tsukamoto K, Azuma T, Leong J, Koop BF, Dijkstra JM (มีนาคม 2015). "การวิเคราะห์ลำดับ MHC class I ของปลาเทเลออสอย่างครอบคลุม" . BMC Evolutionary Biology . 15 (1) 32. Bibcode : 2015BMCEE..15...32G . doi : 10.1186/s12862-015-0309-1 . PMC 4364491 . PMID 25888517 .
- ^ Wang XX, Li Y, Yin Y, Mo M, Wang Q, Gao W และคณะ (กันยายน 2011). "การพัฒนาความสัมพันธ์ของ CD4 ในมนุษย์โดยการแสดงผลบนพื้นผิวของยีสต์และโครงสร้างผลึกของคอมเพล็กซ์ CD4-HLA-DR1" Proceedings of the National Academy of Sciences of the United States of America . 108 (38): 15960– 5. Bibcode : 2011PNAS..10815960W . doi : 10.1073/pnas.1109438108 . PMC 3179091 . PMID 21900604 .
- ^ Dijkstra JM, Yamaguchi T (มีนาคม 2019). "ลักษณะดั้งเดิมของเส้นทางการนำเสนอ MHC คลาส II และแบบจำลองสำหรับต้นกำเนิดที่เป็นไปได้ของโมเลกุล MHC" Immunogenetics . 71 (3): 233– 249. doi : 10.1007/s00251-018-1090-2 . PMID 30377750 . S2CID 53110357 .
- ^ Dijkstra JM, Grimholt U, Leong J, Koop BF, Hashimoto K (พฤศจิกายน 2013). "การวิเคราะห์อย่างครอบคลุมของยีน MHC class II ในจีโนมปลาเทเลออสเผยให้เห็นว่าระบบ DM ที่บรรจุเปปไทด์นั้นไม่จำเป็นในสัตว์มีกระดูกสันหลังส่วนใหญ่" . BMC Evolutionary Biology . 13 (1) 260. Bibcode : 2013BMCEE..13..260D . doi : 10.1186/1471-2148-13-260 . PMC 4219347 . PMID 24279922 .
- ^ Almeida T, Gaigher A, Muñoz-Mérida A, Neves F, Castro LF, Flajnik MF และคณะ (ตุลาคม 2020). "ยีนคลาส II ของปลากระดูกอ่อนเผยให้เห็นสายอัลลีลเก่าที่ไม่เคยมีมาก่อนและยืนยันการเกิดขึ้นของ DM ในช่วงวิวัฒนาการตอนปลาย" . Molecular Immunology . 128 : 125– 138. doi : 10.1016/j.molimm.2020.10.003 . PMC 8010645 . PMID 33126081 .
- ^ Khan FH (2009). องค์ประกอบของภูมิคุ้มกันวิทยา . เดลี: Pearson Education. ISBN 978-81-317-1158-3. OCLC 276274663 .
- ↑คินต์ ทีเจ, โกลด์สบี อาร์เอ, ออสบอร์น บีเอ, คูบี เจ (2550) ภูมิคุ้มกันวิทยาคูบี . มักมิลลัน. ไอเอสบีเอ็น 978-1-4292-0211-4สืบค้นข้อมูลเมื่อวันที่ 28 พฤศจิกายน 2553
- ^ Nesmiyanov P (2020). "การนำเสนอแอนติเจนและคอมเพล็กซ์ความเข้ากันได้ทางเนื้อเยื่อหลัก" . Reference Module in Biomedical Sciences : 90– 98. doi : 10.1016/B978-0-12-818731-9.00029-X . ISBN 978-0-12-801238-3S2CID 234948691 – ผ่าน ทาง Elsevier
- ^ Murphy (2012). "การจดจำแอนติเจนโดยทีเซลล์". อิมมูโนไบโอโลยีของ Janeway (ฉบับที่ 8). Garland Science. หน้า 138–153 .
- ^ Boehm T, Zufall F (กุมภาพันธ์ 2549). "เปปไทด์ MHC และการประเมินทางประสาทสัมผัสของจีโนไทป์". Trends in Neurosciences . 29 (2): 100– 7. doi : 10.1016/j.tins.2005.11.006 . hdl : 11858/00-001M-0000-002B-9268-D . PMID 16337283 . S2CID 15621496 .
- ^ Haig D (พฤศจิกายน 1997). "ปฏิสัมพันธ์ระหว่างมารดาและทารกในครรภ์และความหลากหลายทางพันธุกรรมของ MHC". Journal of Reproductive Immunology . 35 (2): 101– 9. doi : 10.1016/s0165-0378(97)00056-9 . PMID 9421795 .
- ^ Ober C, Weitkamp LR, Cox N, Dytch H, Kostyu D, Elias S (กันยายน 1997). "HLA และการเลือกคู่ครองในมนุษย์" . American Journal of Human Genetics . 61 (3): 497– 504. Bibcode : 1997AmJHG..61..497O . doi : 10.1086/515511 . PMC 1715964 . PMID 9326314 .
- ^ Chaix R, Cao C, Donnelly P (กันยายน 2551). "การเลือกคู่ครองในมนุษย์ขึ้นอยู่กับ MHC หรือไม่?" PLOS Genetics . 4 ( 9) e1000184. doi : 10.1371/journal.pgen.1000184 . PMC 2519788 . PMID 18787687 .
- ^ Derti A, Cenik C, Kraft P, Roth FP (เมษายน 2010). "ไม่มีหลักฐานสำหรับการเลือกคู่ครองที่ขึ้นอยู่กับ MHC ภายในประชากร HapMap" . PLOS Genetics . 6 (4) e1000925. doi : 10.1371/journal.pgen.1000925 . PMC 2861700 . PMID 20442868 .
- ^ Janeš D, Klun I, Vidan-Jeras B, Jeras M, Kreft S (2010). "อิทธิพลของ MHC ต่อการรับรู้กลิ่นของสารเคมี 43 ชนิดและกลิ่นตัว"วารสารชีววิทยาแห่งยุโรปกลาง 5 ( 3): 324– 330. doi : 10.2478/s11535-010-0020-6 .
- ^ Wedekind C, Seebeck T, Bettens F, Paepke AJ (มิถุนายน 1995). "ความชอบคู่ครองที่ขึ้นอยู่กับ MHC ในมนุษย์". Proceedings. Biological Sciences . 260 (1359): 245– 9. Bibcode : 1995RSPSB.260..245W . doi : 10.1098/rspb.1995.0087 . PMID 7630893 . S2CID 34971350 .
- ^ Santos PS, Schinemann JA, Gabardo J, Bicalho MD (เมษายน 2548). "หลักฐานใหม่ที่แสดงว่า MHC มีอิทธิพลต่อการรับรู้กลิ่นในมนุษย์: การศึกษาในนักเรียนชาวบราซิลตอนใต้ 58 คน". Hormones and Behavior . 47 (4): 384– 8. Bibcode : 2005HoBeh..47..384S . doi : 10.1016/j.yhbeh.2004.11.005 . PMID 15777804 . S2CID 8568275 .
- ^ Bryner J (12 สิงหาคม 2551). "ยาคุมกำเนิดทำให้ผู้หญิงเลือกคู่ครองที่ไม่ดี" . Live Science . Future US Inc.
- ^ Sznarkowska A, Mikac S, Pilch M (พฤษภาคม 2020). "การควบคุม MHC Class I: มุมมองต้นกำเนิด"มะเร็ง12 ( 5 ): 1155. doi : 10.3390/cancers12051155 . PMC 7281430 . PMID 32375397 .
- ^ "หมายเลขอัลลีล HLA" . hla.alleles.org . เก็บถาวรจากต้นฉบับเมื่อ 2014-12-16 . เรียกดูเมื่อ2014-12-03 .
- ^ Leffler E, Gao Z, Pfeifer S, Ségurel L, Auton A, Venn O และคณะ (2013). "ตัวอย่างการคัดเลือกสมดุลโบราณหลายครั้งที่พบร่วมกันในมนุษย์และชิมแปนซี" Science . 339 (6127): 1578– 1582. doi : 10.1126/science.1234070 .
- ^ van Oosterhout C (กุมภาพันธ์ 2552). "ทฤษฎีใหม่ของการวิวัฒนาการของ MHC: นอกเหนือจากการคัดเลือกบนยีนภูมิคุ้มกัน" Proceedings . Biological Sciences . 276 (1657): 657– 65. doi : 10.1098/rspb.2008.1299 . PMC 2660941 . PMID 18986972 .
- ^ Manczinger M, Boross G, Kemény L, Müller V, Lenz TL, Papp B และคณะ (มกราคม 2019). "ความหลากหลายของเชื้อโรคขับเคลื่อนวิวัฒนาการของอัลลีล MHC-II ทั่วไปในประชากรมนุษย์" PLOS Biology 17 ( 1) e3000131. doi : 10.1371/journal.pbio.3000131 . PMC 6372212 . PMID 30703088 .
- ^ Zeisset I, Beebee TJ (2014). "การเปลี่ยนแปลงแบบสุ่มมากกว่าการคัดเลือกเป็นตัวกำหนดรูปแบบความหลากหลายของอัลลีล MHC class II ในระดับขอบเขตทางชีวภูมิศาสตร์ในคางคก Bufo calamita" . PLOS ONE . 9 (6) e100176. Bibcode : 2014PLoSO...9j0176Z . doi : 10.1371/journal.pone.0100176 . PMC 4061088 . PMID 24937211 .
- ^ Cortázar-Chinarro M, Lattenkamp EZ, Meyer-Lucht Y, Luquet E, Laurila A, Höglund J (สิงหาคม 2017). "การลอยตัว การคัดเลือก หรือการอพยพ? กระบวนการที่มีผลต่อความแตกต่างทางพันธุกรรมและความแปรผันตามแนวละติจูดในสัตว์สะเทินน้ำสะเทินบก" . BMC Evolutionary Biology . 17 (1) 189. Bibcode : 2017BMCEE..17..189C . doi : 10.1186/s12862-017-1022-z . PMC 5557520 . PMID 28806900 .
- ^ Apanius V, Penn D, Slev PR, Ruff LR, Potts WK (2017). "ลักษณะของการคัดเลือกบนคอมเพล็กซ์ฮิสโตคอมแพติบิลิตีหลัก" บทวิจารณ์เชิงวิพากษ์ในภูมิคุ้มกันวิทยา 37 ( 2– 6 ): 75– 120. doi : 10.1615/CritRevImmunol.v37.i2-6.10 . PMID 29773018 .
- ^ Chowell D (2019). "การแยกตัวทางวิวัฒนาการของจีโนไทป์ HLA คลาส I ส่งผลต่อประสิทธิภาพของการรักษาด้วยภูมิคุ้มกันบำบัดมะเร็ง" Nature Medicine 25 ( 11): 1715– 1720. doi : 10.1038/s41591-019-0639-4 . PMC 7938381 .
- ^ Viard M (2024). "ผลกระทบของความแตกต่างในการทำงานของ HLA คลาส I ต่อการควบคุมเชื้อ HIV" Science . 383 (6680): 319– 325. doi : 10.1126/science.adk0777 .
- ^ "ข้อได้เปรียบของอัลลีลที่แตกต่างกันในยีน MHC ของมนุษย์" อิมมูโนเจเนติกส์ 1990. doi : 10.1007/BF02918202 .
- ^ Sommer S (ตุลาคม 2548). "ความสำคัญของความแปรปรวนของยีนภูมิคุ้มกัน (MHC) ในนิเวศวิทยาเชิงวิวัฒนาการและการอนุรักษ์" . Frontiers in Zoology . 2 (16) 16. doi : 10.1186/1742-9994-2-16 . PMC 1282567 . PMID 16242022 .
- ^ Manlik O, Krützen M, Kopps AM, Mann J, Bejder L, Allen SJ และคณะ (มิถุนายน 2019). "ความหลากหลายของ MHC เป็นตัวบ่งชี้ที่ดีกว่าสำหรับการอนุรักษ์มากกว่าความหลากหลายทางพันธุกรรมที่เป็นกลางหรือไม่? กรณีศึกษาของประชากรโลมาสองกลุ่มที่แตกต่างกัน" . Ecology and Evolution . 9 (12): 6986– 6998. Bibcode : 2019EcoEv...9.6986M . doi : 10.1002/ece3.5265 . PMC 6662329 . PMID 31380027 .
- ^ Castro-Prieto A, Wachter B, Sommer S (เมษายน 2011). "การทบทวนแบบจำลองเสือชีตาห์: ความหลากหลายของ MHC ในประชากรเสือชีตาห์ที่อาศัยอยู่ตามธรรมชาติที่ใหญ่ที่สุดในโลก" . Molecular Biology and Evolution . 28 (4): 1455– 68. doi : 10.1093/molbev/msq330 . PMC 7187558 . PMID 21183613 .
- ^ Babik W, Durka W, Radwan J (ธันวาคม 2005). "ความหลากหลายของลำดับยีน MHC DRB ในบีเวอร์ยูเรเซีย (Castor fiber)" . Molecular Ecology . 14 (14): 4249– 57. Bibcode : 2005MolEc..14.4249B . doi : 10.1111/j.1365-294X.2005.02751.x . PMID 16313590 . S2CID 22260395 .
- ^ Zhu L, Ruan XD, Ge YF, Wan QH, Fang SG (มิถุนายน 2550). "ความหลากหลาย DQA ของคอมเพล็กซ์ฮิสโตคอมแพติบิลิตีหลักคลาส II ต่ำในแพนด้ายักษ์ (Ailuropoda melanoleuca)" . BMC Genetics . 8 (1) 29. Bibcode : 2007BMCGe...8...29Z . doi : 10.1186/1471-2156-8-29 . PMC 1904234 . PMID 17555583 .
- ^ Siddle HV, Kreiss A, Eldridge MD, Noonan E, Clarke CJ, Pyecroft S และคณะ (ตุลาคม 2550). "การแพร่กระจายของเนื้องอกโคลนที่ร้ายแรงโดยการกัดเกิดขึ้นเนื่องจากความหลากหลายของ MHC ลดลงในสัตว์มีถุงหน้าท้องกินเนื้อที่ใกล้สูญพันธุ์" Proceedings of the National Academy of Sciences of the United States of America . 104 (41): 16221– 6. doi : 10.1073/pnas.0704580104 . PMC 1999395 . PMID 17911263 .
- ^ a b Shum BP, Guethlein L, Flodin LR, Adkison MA, Hedrick RP, Nehring RB และคณะ (มีนาคม 2544). "รูปแบบวิวัฒนาการของ MHC คลาส I และ II ในปลาแซลมอนแตกต่างจากแบบแผนของไพรเมต"วารสารภูมิคุ้มกันวิทยา 166 ( 5): 3297– 308. Bibcode : 2001JImm..166.3297S . doi : 10.4049/jimmunol.166.5.3297 . PMID 11207285 . S2CID 5725603 .
- ^ Aoyagi K, Dijkstra JM, Xia C, Denda I, Ototake M, Hashimoto K และคณะ (มกราคม 2545) "ยีน MHC คลาส I แบบคลาสสิกที่ประกอบด้วยสายพันธุ์ลำดับที่แตกต่างกันอย่างมากมีโลคัสเดียวร่วมกันในปลาเทราต์สายรุ้ง (Oncorhynchus mykiss)"วารสารภูมิคุ้มกันวิทยา 168 ( 1): 260–73 . Bibcode : 2002JImm..168..260A . doi : 10.4049/jimmunol.168.1.260 . PMID 11751970. S2CID 36838421 .
- ^ Yamaguchi T, Dijkstra JM (เมษายน 2019). "ยีน Major Histocompatibility Complex (MHC) และความต้านทานโรคในปลา" . Cells . 8 (4): 378. doi : 10.3390/cells8040378 . PMC 6523485 . PMID 31027287 .
- ^ Abi Rached L, McDermott MF, Pontarotti P (กุมภาพันธ์ 1999). "การระเบิดครั้งใหญ่ของ MHC". Immunological Reviews . 167 (1): 33– 44. doi : 10.1111/j.1600-065X.1999.tb01380.x . PMID 10319249 . S2CID 29886370 .
- ↑ซูร์วาลี เจ, จูโน แอล, ธีโปต ดี, กรูซี เอส, ปอนตารอตติ พี, ดู ปาสคิเยร์ แอล, และคณะ (กันยายน 2557). "โปรโต-MHC ของพลาโคซัว ซึ่งเป็นบริเวณที่เชี่ยวชาญด้านความเครียดของเซลล์และวิถีการแพร่กระจาย/โปรตีโอโซม " วารสารวิทยาภูมิคุ้มกัน . 193 (6): 2891– 901. ดอย : 10.4049/jimmunol.1401177 . PMID25114105 .
- ^ Abbas AB, Lichtman AH (2009). "บทที่ 10 การตอบสนองทางภูมิคุ้มกันต่อเนื้องอกและการปลูกถ่าย" ภูมิคุ้มกันวิทยาพื้นฐาน หน้าที่และความผิดปกติของระบบภูมิคุ้มกัน (ฉบับที่ 3) Saunders (Elsevier) ISBN 978-1-4160-4688-2.
- ^ Sarder MR, Fischer U, Dijkstra JM, Kiryu I, Yoshiura Y, Azuma T และคณะ (สิงหาคม 2546) "กลุ่มการเชื่อมโยง MHC คลาส I เป็นตัวกำหนดหลักในการปฏิเสธเม็ดเลือดแดงต่างสายพันธุ์ในร่างกายของปลาเทราต์สายรุ้ง (Oncorhynchus mykiss)" Immunogenetics 55 ( 5): 315– 24. doi : 10.1007/s00251-003-0587-4 . PMID 12879308 . S2CID 21437633 .
- ^ Quiniou SM, Wilson M, Bengtén E, Waldbieser GC, Clem LW, Miller NW (2005). "การวิเคราะห์ MHC RFLP ในครอบครัวพี่น้องร่วมสายเลือดของปลาดุกช่อง: การระบุบทบาทของโมเลกุล MHC ในการตอบสนองแบบไซโทท็อกซิกอัลโลจีนิกโดยธรรมชาติ" ภูมิคุ้มกัน วิทยาเชิงพัฒนาการและเปรียบเทียบ29 (5): 457– 67. doi : 10.1016/j.dci.2004.08.008 . PMID 15707666 .
- ^ Cardwell TN, Sheffer RJ, Hedrick PW (สิงหาคม 2544). "ความแปรผันของ MHC และการปลูกถ่ายเนื้อเยื่อในปลา". วารสารพันธุกรรม 92 ( 4): 305– 8. doi : 10.1093/jhered/92.4.305 . PMID 11535641 .
- ^ Abbas AB, Lichtman AH (2009). "บทที่ 3 การจับและการนำเสนอแอนติเจนต่อลิมโฟไซต์" ภูมิคุ้มกันวิทยาพื้นฐาน หน้าที่และความผิดปกติของระบบภูมิคุ้มกัน (ฉบับที่ 3) Saunders (Elsevier) ISBN 978-1-4160-4688-2.
- ^ Spurgin LG, Richardson DS (เมษายน 2010). "เชื้อโรคขับเคลื่อนความหลากหลายทางพันธุกรรมอย่างไร: MHC กลไก และความเข้าใจผิด" Proceedings. Biological Sciences . 277 ( 1684): 979– 88. doi : 10.1098/rspb.2009.2084 . PMC 2842774 . PMID 20071384 .
- ^ Kinget L, Naulaerts S, Govaerts J, Vanmeerbeek I, Sprooten J, Laureano RS และคณะ (มิถุนายน 2024). "ลายเซ็น HLA ที่ฝังสถาปัตยกรรมเชิงพื้นที่เพื่อทำนายการตอบสนองทางคลินิกต่อภูมิคุ้มกันบำบัดในมะเร็งเซลล์ไต" Nature Medicine . 30 (6): 1667– 1679. doi : 10.1038/s41591-024-02978-9 . PMID 38773341 .
{{cite journal}}: CS1 maint: การตั้งค่าที่ถูกแทนที่ ( ลิงก์ )
บรรณานุกรม
- เดวิส ดีเอ็น (2014). ยีนแห่งความเข้ากันได้ . ลอนดอน: สำนักพิมพ์เพนกวิน . ISBN 978-0-241-95675-5.
ลิงก์ภายนอก
- Major+Histocompatibility+Complexที่ US National Library of Medicine Medical Subject Headings (MeSH)
- Molecular Individuality Archived 2013-01-29 at the Wayback Machine —German online book (2012)
- เซิร์ฟเวอร์ NetMHC 3.0 —ทำนายการจับกันของเปปไทด์กับอัลลีล MHC (HLA) ที่แตกต่างกันหลายชนิด
- กลุ่มทีเซลล์ — มหาวิทยาลัยคาร์ดิฟฟ์
- เรื่องราวของ 2YF6: ไก่ MHC
- ฐานข้อมูลโปรตีน RCSB: โมเลกุลประจำเดือน—คอมเพล็กซ์ความเข้ากันได้ทางพันธุกรรมหลัก
- dbMHC Home ฐานข้อมูล Major Histocompatibility Complex ของ NCBI
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ คอมเพล็กซ์ความเข้ากันได้ทางพันธุกรรมหลัก
คอมเพล็กซ์ ฮิสโตคอมแพติบิลิตีหลัก ( MHC ) เป็น บริเวณ ขนาดใหญ่ บนดีเอ็นเอของสัตว์มีกระดูกสันหลัง ซึ่งประกอบด้วยชุด ยีน ที่เชื่อมโยงกันอย่างใกล้ชิดและ มีลักษณะเป็นโพลีมอร์ฟิก...
การค้นพบ
คำอธิบายแรกของ MHC เกิดขึ้นโดย ปีเตอร์ โกเรอร์ นักภูมิคุ้มกันวิทยา ชาวอังกฤษ ในปี 1936 [ 7 ] ยีน MHC ถูกระบุครั้งแรกในหนูสายพันธุ์ผสมพันธุ์ คลาเรนซ์ ลิตเติล ปลูกถ่ายเนื้องอกข้ามสายพันธุ์ต่างๆ...
ยีน
ตำแหน่ง MHC มีอยู่ใน สัตว์มีกระดูกสันหลังที่มีขากรรไกร ทั้งหมด สันนิษฐานว่าเกิดขึ้นเมื่อประมาณ 450 ล้านปีก่อน [ 15 ] แม้ว่าจำนวนยีนที่รวมอยู่ใน MHC ของสปีชีส์ต่างๆ จะแตกต่างกัน แต่โครงสร้างโดยรวมของตำแหน่งนั้นค่อนข้างคล้ายกัน MHC...
โปรตีน
ตัวรับทีเซลล์ ที่จับคู่กับ MHC-I และ MHC-II