อ่าน 16 นาที
เมทัลโลโปรตีน
เมทัลโลโปรตีนเป็นคำทั่วไปสำหรับโปรตีนที่มีไอออนโลหะเป็นโคแฟคเตอร์ โปรตีน ส่วนใหญ่จัดอยู่ในประเภทนี้ ตัวอย่างเช่น โปรตีนของมนุษย์อย่างน้อย 1,000 ชนิด (จากทั้งหมดประมาณ 20,000 ชนิด).
เมทัลโลโปรตีน
เมทัลโลโปรตีนเป็นคำทั่วไปสำหรับโปรตีนที่มีไอออนโลหะเป็นโคแฟคเตอร์ [ 1 ] [ 2 ] โปรตีน ส่วนใหญ่จัดอยู่ในประเภทนี้ ตัวอย่างเช่น โปรตีนของมนุษย์อย่างน้อย 1,000 ชนิด (จากทั้งหมดประมาณ 20,000 ชนิด) มี โดเมนโปรตีนที่จับกับสังกะสี[ 3 ]แม้ว่าอาจจะมีเมทัลโลโปรตีนสังกะสีของมนุษย์มากถึง 3,000 ชนิดก็ตาม[ 4 ]
ความอุดมสมบูรณ์
มีการประมาณว่าโปรตีน ประมาณครึ่งหนึ่ง มีโลหะเป็นองค์ประกอบ [ 5 ] จากการประมาณอีกครั้งหนึ่ง คาดว่าโปรตีนประมาณหนึ่งในสี่ถึงหนึ่งในสามต้องใช้โลหะเพื่อทำหน้าที่[ 6 ]ดังนั้น เมทัลโลโปรตีนจึงมีหน้าที่หลากหลายในเซลล์เช่น การเก็บรักษา การขนส่ง การเร่งปฏิกิริยา ของเอนไซม์และการส่งสัญญาณหรือโรคติดเชื้อ[ 7 ]
โลหะส่วนใหญ่ในร่างกายมนุษย์ จะจับตัวอยู่กับโปรตีน ตัวอย่างเช่น ความเข้มข้นของธาตุเหล็กในร่างกายมนุษย์ที่ค่อนข้างสูงนั้น ส่วนใหญ่เกิดจากธาตุเหล็กในฮีโมโกลบิน
| ตับ | ไต | ปอด | หัวใจ | สมอง | กล้ามเนื้อ | |
|---|---|---|---|---|---|---|
| แมงกานีส ( Mn ) | 138 | 79 | 29 | 27 | 22 | <4-40 |
| Fe ( เหล็ก ) | 16,769 | 7,168 | 24,967 | 5,530 | 4,100 | 3,500 |
| โคบอลต์ (Co ) | <2-13 | <2 | <2-8 | --- | <2 | 150 (?) |
| นิกเกล (Ni ) | <5 | <5-12 | <5 | <5 | <5 | <15 |
| Cu ( ทองแดง ) | 882 | 379 | 220 | 350 | 401 | 85-305 |
| Zn ( สังกะสี ) | 5,543 | 5,018 | 1,470 | 2,772 | 915 | 4,688 |
หลักการเคมีเชิงโคออร์ดิเนชัน
ในเมทัลโลโปรตีน ไอออนของโลหะมักจะถูกประสานโดย ศูนย์กลาง ไนโตรเจนออกซิเจนหรือกำมะถันที่อยู่ใน หมู่ กรดอะมิโนของโปรตีน หมู่ผู้ให้เหล่านี้มักจะมาจากโซ่ข้างบนหมู่กรดอะมิโน โดยเฉพาะอย่างยิ่ง หมู่แทนที่อิมิดาโซลในหมู่ฮิสติ ดีน หมู่แทนที่ ไทโอเลตใน หมู่ ซิสเทอีนและ หมู่ คาร์บอกซิเลตที่มาจากแอสปาร์เทต มีความสำคัญอย่างยิ่ง เนื่องจากความหลากหลายของเมทัลโลโปรตีโอม พบว่าหมู่กรดอะมิโนเกือบทั้งหมดสามารถจับกับศูนย์กลางโลหะได้ โครงสร้างหลักของเปปไทด์ยังให้หมู่ผู้ให้ด้วย ซึ่งรวมถึงอะไมด์ ที่ถูกกำจัดโปรตอน และ ศูนย์กลางออกซิเจน คาร์บอนิลของ อะ ไมด์ การจับของตะกั่ว(II) ในโปรตีนธรรมชาติและโปรตีนสังเคราะห์ได้รับการทบทวนแล้ว[ 9 ]
นอกเหนือจากหมู่ผู้ให้ที่ได้จากกรดอะมิโนแล้วโคแฟคเตอร์ อินทรีย์หลายชนิด ยังทำหน้าที่เป็นลิแกนด์ ด้วย ลิแกนด์ ที่มีชื่อเสียงที่สุดอาจจะเป็น ลิแกนด์ มาโครไซ คลิก N4 แบบสี่ตำแหน่ง ที่รวมอยู่ใน โปรตีน ฮีมลิแกนด์อนินทรีย์ เช่น ซัลไฟด์และออกไซด์ก็พบได้ทั่วไปเช่นกัน
โปรตีนโลหะสำหรับการจัดเก็บและขนส่ง
นี่คือผลิตภัณฑ์ขั้นที่สองของการไฮโดรไลซิสโปรตีน ซึ่งได้จากการบำบัดด้วยกรดและด่างที่มีความเข้มข้นสูงขึ้นเล็กน้อย
ตัวนำออกซิเจน
ฮีโมโกลบินซึ่งเป็นตัวนำออกซิเจนหลักในมนุษย์ มีหน่วยย่อยสี่หน่วย โดยที่ ไอออน เหล็ก (II) จะถูกประสานโดยลิแกนด์มาโครไซคลิกแบบระนาบโปรโตพอร์ไฟริน IX (PIX) และ อะตอม ไนโตรเจนของอิมิดาโซลของ สารตกค้าง ฮิสติ ดี น ตำแหน่งการประสานที่หกประกอบด้วย โมเลกุล น้ำหรือ โมเลกุล ไดออกซิเจนในทางตรงกันข้าม โปรตีนไมโอโกลบินซึ่งพบในเซลล์กล้ามเนื้อมีเพียงหน่วยย่อยเดียวเท่านั้น ตำแหน่งที่ใช้งานอยู่ตั้งอยู่ใน ช่อง ไฮโดรโฟบิกซึ่งมีความสำคัญเนื่องจากหากไม่มีช่องนี้ เหล็ก (II) จะถูกออกซิไดซ์เป็นเหล็ก (III) อย่าง ถาวร ค่าคงที่สมดุลสำหรับการก่อตัวของ HbO2 เป็นเช่นนั้น ออกซิเจนจะถูกดูดซับหรือปล่อยออกมาขึ้นอยู่กับความดันย่อยของออกซิเจนในปอดหรือในกล้ามเนื้อ ในฮีโมโกลบิน หน่วยย่อยทั้งสี่แสดงผลการทำงานร่วมกันที่ช่วยให้การถ่ายโอนออกซิเจนจากฮีโมโกลบินไปยังไมโอโกลบินเป็นไปได้ง่าย[ 10 ]
ในทั้งฮีโมโกลบินและไมโอโกลบินบางครั้งมีการกล่าวอย่างไม่ถูกต้องว่าชนิดที่มีออกซิเจนประกอบด้วยเหล็ก(III) ปัจจุบันเป็นที่ทราบกันแล้วว่า ลักษณะ ไดอะแมกเนติกของชนิดเหล่านี้เป็นเพราะอะตอมเหล็ก(II) อยู่ในสถานะสปินต่ำ ใน ออกซีฮีโมโกลบินอะตอมเหล็กตั้งอยู่ในระนาบของวงแหวนพอร์ไฟริน แต่ใน ดีออก ซีฮีโมโกลบิน ที่ เป็นพาราแมก เนติก อะตอมเหล็กอยู่เหนือระนาบของวงแหวน[ 10 ]การเปลี่ยนแปลงสถานะสปินนี้เป็นผลร่วมกันเนื่องจากการแยกสนามผลึก ที่สูงขึ้น และรัศมีไอออน ที่เล็กกว่า ของ Fe 2+ในส่วนประกอบออกซีฮีโมโกลบิน
เฮเมอริทรินเป็นตัวนำออกซิเจนที่มีธาตุเหล็กอีกชนิดหนึ่ง ตำแหน่งการจับออกซิเจนเป็นศูนย์กลางเหล็กแบบไบนิวเคลียร์ อะตอมของเหล็กจะประสานกับโปรตีนผ่านทาง โซ่ข้าง คาร์บอกซิเลตของกรดกลูตามิกและกรดแอสปาร์ติก และสารตกค้าง ฮิสติดีนห้าตัว การดูดซับ O2 โดยเฮเมอริทรินจะเกิดขึ้นพร้อมกับการออกซิเดชันสองอิเล็กตรอนของศูนย์กลางไบนิวเคลียร์ที่ลดลงเพื่อสร้างเปอร์ออกไซด์ ที่จับอยู่ (OOH− )กลไกการดูดซับและการปล่อยออกซิเจนได้รับการศึกษาอย่างละเอียด[ 11 ] [ 12 ]
เฮโมไซยานินทำหน้าที่ขนส่งออกซิเจนในเลือดของหอย ส่วนใหญ่ และสัตว์ขาปล้อง บางชนิด เช่นปูม้า รองจากฮีโมโกลบินแล้ว เฮโมไซยานินมีความสำคัญทางชีววิทยาในการขนส่งออกซิเจนมากเป็นอันดับสอง เมื่อเกิดปฏิกิริยาออกซิเดชัน อะตอม ทองแดง (I) สองอะตอมที่ตำแหน่งออกฤทธิ์จะถูกออกซิไดซ์เป็นทองแดง (II) และโมเลกุลออกซิเจนจะถูกรีดิวซ์เป็นเปอร์ออกไซด์O₂2− 2[ 13 ] [ 14 ]
คลอโรครูออริน (และ อีริโทร โรครูออรินซึ่ง เป็นตัวนำพาขนาดใหญ่กว่า ) เป็นโปรตีนฮีมที่จับกับออกซิเจน พบในพลาสมาในเลือดของหนอนปล้อง หลายชนิด โดยเฉพาะอย่าง ยิ่ง หนอนปล้องทะเลบางชนิด
ไซโตโครม
ปฏิกิริยา ออกซิเดชันและรีดักชันไม่เป็นที่นิยมในเคมีอินทรีย์เนื่องจากมีโมเลกุลอินทรีย์เพียงไม่กี่ชนิดที่สามารถทำหน้าที่เป็น สาร ออกซิไดซ์หรือสารรีดิวซ์ได้ในทางกลับกันเหล็ก (II) สามารถถูกออกซิไดซ์เป็นเหล็ก (III) ได้ง่าย ฟังก์ชันนี้ถูกนำมาใช้ใน ไซโตโครมซึ่งทำหน้าที่เป็น ตัวนำ อิเล็กตรอน การมีอยู่ของไอออนโลหะทำให้เอนไซม์โลหะ สามารถ ทำหน้าที่ต่างๆ เช่นปฏิกิริยารีดอกซ์ซึ่งไม่สามารถทำได้ง่ายโดยกลุ่มฟังก์ชัน ที่มีจำกัด ที่พบในกรดอะมิโน [ 15 ]อะตอมเหล็กในไซโตโครมส่วนใหญ่จะอยู่ใน กลุ่ม ฮีมความแตกต่างระหว่างไซโตโครมเหล่านั้นอยู่ที่โซ่ข้างที่แตกต่างกัน ตัวอย่างเช่น ไซโตโครม a มีกลุ่ม โปรสเตติก ฮีม a และไซโตโครม b มี กลุ่มโปรสเตติก ฮีม b ความแตกต่างเหล่านี้ส่งผลให้ ศักยภาพรีดอก ซ์ Fe 2+ /Fe 3+ แตกต่างกัน ทำให้ไซโตโครมต่างๆ มีส่วนร่วมในห่วงโซ่การขนส่งอิเล็กตรอนของไมโทคอน เดรีย [ 16 ]
เอนไซม์ ไซโตโครม P450ทำหน้าที่แทรกอะตอมออกซิเจนเข้าไปในพันธะ C−H ซึ่งเป็นปฏิกิริยาออกซิเดชัน[ 17 ] [ 18 ]
รูเบรดอกซิน

รูเบรดอกซินเป็นตัวนำอิเล็กตรอนที่พบในแบคทีเรียและอาร์เคียที่เผาผลาญ กำมะถัน บริเวณออกฤทธิ์ประกอบด้วยไอออนเหล็กที่ถูกประสานกับอะตอมกำมะถันของ หมู่ซิสเท อีน สี่หมู่ซึ่งก่อตัวเป็น รูปทรงสี่เหลี่ยมด้านเท่าเกือบสมบูรณ์รูเบรดอกซินทำการถ่ายโอนอิเล็กตรอนหนึ่งตัวสถานะออกซิเดชันของอะตอมเหล็กเปลี่ยนระหว่างสถานะ +2 และ +3 ในทั้งสองสถานะออกซิเดชัน โลหะจะมีสปินสูงซึ่งช่วยลดการเปลี่ยนแปลงโครงสร้างให้น้อยที่สุด
พลาสโตไซยานิน

พลาสโตไซยานินเป็นหนึ่งในตระกูลโปรตีนทองแดง สีน้ำเงิน ที่เกี่ยวข้องกับปฏิกิริยาการถ่ายโอนอิเล็กตรอน ตำแหน่งการจับ ทองแดงถูกอธิบายว่าเป็นพีระมิดสามเหลี่ยมบิดเบี้ยว[ 19 ]ระนาบสามเหลี่ยมของฐานพีระมิดประกอบด้วยอะตอมไนโตรเจนสองอะตอม (N 1และ N 2 ) จากฮิสติดีนที่แยกจากกันและซัลเฟอร์ (S 1 ) จากซิสเทอีน ซัลเฟอร์ (S 2 ) จากเมไทโอนีนแกนกลางก่อตัวเป็นยอด การบิดเบี้ยวเกิดขึ้นในความยาวพันธะระหว่างลิแกนด์ทองแดงและซัลเฟอร์ การสัมผัส Cu−S 1สั้นกว่า (207 pm ) กว่า Cu−S 2 (282 pm) พันธะ Cu−S 2 ที่ยาวขึ้น ทำให้รูปแบบ Cu(II) ไม่เสถียรและเพิ่มศักยภาพรีดอกซ์ของโปรตีน สีน้ำเงิน (การดูดกลืนสูงสุดที่ 597 นาโนเมตร ) เกิดจากพันธะ Cu−S 1 ซึ่ง มีการถ่ายโอนประจุจากS(pπ) ไปยัง Cu(d x 2 − y 2 ) [ 20 ]
ในพลาสโตไซยานินรูปแบบรีดิวซ์His -87 จะถูกโปรตอนไนซ์โดยมีค่า pKaเท่ากับ4.4การโปรตอนไนซ์จะป้องกันไม่ให้มันทำหน้าที่เป็นลิแกนด์และรูปทรงเรขาคณิตของตำแหน่งทองแดงจะกลายเป็นระนาบสามเหลี่ยม
การกักเก็บและการถ่ายโอนไอออนโลหะ
เหล็ก
ธาตุเหล็กถูกเก็บไว้ในรูปของเหล็ก(III) ในเฟอร์ริตินลักษณะที่แน่นอนของตำแหน่งการจับยังไม่ได้รับการระบุ ธาตุเหล็กดูเหมือนจะอยู่ในรูปของ ผลิตภัณฑ์ ไฮโดรไลซิสเช่น FeO(OH) ธาตุเหล็กถูกขนส่งโดยทรานสเฟอร์รินซึ่งตำแหน่งการจับประกอบด้วยไทโรซีน 2 ตัว กรดแอสปาร์ติก 1 ตัวและฮิสติดีน 1 ตัว[ 21 ]ร่างกายมนุษย์ไม่มีกลไกควบคุมการขับถ่ายธาตุเหล็ก[ 22 ]ซึ่งอาจนำไปสู่ ปัญหา ธาตุเหล็กเกินในผู้ป่วยที่ได้รับการรักษาด้วยการถ่ายเลือดเช่น ในผู้ป่วยโรคธาลัสซีเมียเบต้า ธาตุเหล็กถูกขับออกทางปัสสาวะ[ 23 ]และยังมีความเข้มข้นในน้ำดี[ 24 ]ซึ่งถูกขับออกทางอุจจาระ[ 25 ]
ทองแดง
เซรูโลพลาสมินเป็นโปรตีนหลัก ที่ทำหน้าที่ขนส่ง ทองแดงในเลือด เซรูโลพลาสมินมีฤทธิ์เป็นเอนไซม์ออกซิเดส ซึ่งเกี่ยวข้องกับการออกซิเดชันของ Fe(II) ไปเป็น Fe(III) จึงช่วยในการขนส่ง Fe(III) ในพลาสมาในเลือดโดยทำงานร่วมกับทรานสเฟอร์ริน ซึ่งสามารถขนส่งเหล็กได้เฉพาะในรูป Fe(III) เท่านั้น
แคลเซียม
ออสทีโอพอนตินมีส่วนเกี่ยวข้องกับการสร้างแร่ธาตุในเมทริกซ์นอกเซลล์ของกระดูกและฟัน
เอนไซม์โลหะ
เอนไซม์ที่มีโลหะเป็นองค์ประกอบทั้งหมดมีลักษณะร่วมกันอย่างหนึ่ง คือ ไอออนของโลหะจะจับกับโปรตีนด้วยตำแหน่งการประสานงานที่ไม่เสถียร เพียงตำแหน่งเดียว เช่นเดียวกับ เอนไซม์ ทั้งหมด รูปร่างของตำแหน่งออกฤทธิ์มีความสำคัญอย่างยิ่ง ไอออนของโลหะมักจะอยู่ในช่องที่มีรูปร่างเหมาะสมกับสารตั้งต้น ไอออนของโลหะทำหน้าที่เร่งปฏิกิริยาที่ยากต่อการเกิดขึ้นใน เคมีอินทรีย์
คาร์บอนิกแอนไฮเดรส

ในสารละลายในน้ำคาร์บอนไดออกไซด์จะก่อตัวเป็นกรดคาร์บอนิก
- CO 2 + H 2 O ⇌ H 2 CO 3
ปฏิกิริยานี้จะเกิดขึ้นช้ามากหากไม่มีตัวเร่งปฏิกิริยา แต่จะเกิดขึ้นเร็วมากเมื่อมีไอออน ไฮดรอกไซ ด์อยู่ด้วย
- CO 2 + OH − ⇌ HCO− 3
ปฏิกิริยาที่คล้ายคลึงกันนี้เกิดขึ้นเกือบจะในทันทีด้วยคาร์บอนิกแอนไฮเดรสโครงสร้างของตำแหน่งออกฤทธิ์ในคาร์บอนิกแอนไฮเดรสเป็นที่รู้จักกันดีจากโครงสร้างผลึกจำนวนมาก ประกอบด้วย ไอออน สังกะสี ที่ประสานงานกับอะตอมไนโตรเจนของ อิมิดาโซลสามอะตอมจาก หน่วย ฮิสติดีน สาม หน่วย ตำแหน่งการประสานงานที่สี่ถูกครอบครองโดยโมเลกุลน้ำ ทรงกลมการประสานงานของไอออนสังกะสีมีลักษณะเป็นทรงสี่หน้า โดยประมาณ ไอออนสังกะสีที่มีประจุบวกทำให้โมเลกุลน้ำที่ประสานงานเกิดการโพลาไรซ์ และ การโจมตี แบบนิวคลีโอฟิลิกโดยส่วนไฮดรอกไซด์ที่มีประจุลบต่อคาร์บอนไดออกไซด์จะเกิดขึ้นอย่างรวดเร็ว วัฏจักรเร่งปฏิกิริยาสร้างไอออนไบคาร์บอเนตและไอออนไฮโดรเจน[ 2 ]เป็นสมดุล :
- H₂CO₃ ⇌ HCO− 3+ H +
สนับสนุนการแตกตัวของกรดคาร์บอนิกที่ค่าpH ทางชีวภาพ [ 26 ]
เอนไซม์ที่ต้องอาศัยวิตามินบี12
วิตามินบี12ที่มีโคบอลต์ (หรือที่รู้จักกันในชื่อโคบาลามิน) ทำหน้าที่เร่งปฏิกิริยาการถ่ายโอนหมู่เมทิล (−CH 3 ) ระหว่างโมเลกุลสองโมเลกุล ซึ่งเกี่ยวข้องกับการแตกพันธะ C−Cซึ่งเป็นกระบวนการที่ต้องใช้พลังงานสูงในปฏิกิริยาอินทรีย์ ไอออนโลหะจะลดพลังงานกระตุ้นสำหรับกระบวนการโดยการสร้างพันธะ Co−CH 3 ชั่วคราว [ 27 ]โครงสร้างของโคเอนไซม์ได้รับการกำหนดอย่างมีชื่อเสียงโดยโดโรธี ฮอดจ์กินและเพื่อนร่วมงาน ซึ่งทำให้เธอได้รับ รางวัลโนเบล สาขาเคมี[ 28 ]ประกอบด้วยไอออนโคบอลต์(II) ที่ประสานกับอะตอมไนโตรเจนสี่อะตอมของ วงแหวน คอร์รินและอะตอมไนโตรเจนที่ห้าจาก กลุ่ม อิมิดาโซล ในสถานะพักจะมี พันธะซิกมา Co−C กับอะตอมคาร์บอน 5′ ของอะดีโนซีน[ 29 ]นี่คือ สารประกอบ ออร์กาโนเมทัลลิก ที่เกิดขึ้นตามธรรมชาติ ซึ่งอธิบายถึงหน้าที่ของมันใน ปฏิกิริยา ทรานส์เมทิลเลชัน เช่น ปฏิกิริยาที่ดำเนินการโดยเมไทโอนีนซินเทส
ไนโตรเจเนส (การตรึงไนโตรเจน)
การตรึงไนโตรเจนในชั้นบรรยากาศเป็นกระบวนการที่ต้องใช้พลังงานสูง เนื่องจากเกี่ยวข้องกับการทำลายพันธะสาม ที่เสถียรมาก ระหว่างอะตอมของไนโตรเจน เอนไซม์ไนโตรเจเนสเป็นตัวเร่งปฏิกิริยาในกระบวนการนี้ เอนไซม์ชนิดหนึ่งดังกล่าวพบได้ในแบคทีเรียไรโซเบียม การทำงานของเอนไซม์นี้ประกอบด้วยสามส่วน ได้แก่ อะตอมของ โมลิบเดนัมที่ตำแหน่งออกฤทธิ์กลุ่มเหล็ก-กำมะถันที่เกี่ยวข้องกับการขนส่งอิเล็กตรอนที่จำเป็นในการรีดิวซ์ไนโตรเจน และแหล่งพลังงานที่อุดมสมบูรณ์ในรูปของแมกนีเซียมATPซึ่งได้มาจากความสัมพันธ์แบบพึ่งพาอาศัยกันระหว่างแบคทีเรียและพืชเจ้าบ้าน ซึ่งมักจะเป็นพืชตระกูลถั่วปฏิกิริยานี้สามารถเขียนในเชิงสัญลักษณ์ได้ดังนี้
โดยที่ Pi หมายถึงฟอสเฟต อนินทรีย์ โครงสร้างที่แน่นอนของตำแหน่งออกฤทธิ์นั้นยากที่จะระบุได้ ดูเหมือนว่าจะมีคลัสเตอร์ MoFe 7 S 8ที่สามารถจับกับโมเลกุลไดไนโตรเจนและคาดว่าจะทำให้กระบวนการรีดักชันเริ่มต้นขึ้นได้[ 30 ]นอกจากนี้ยังพบว่าแบคทีเรียและอาร์เคียบางชนิดมีไนโตรเจเนสวาเนเดียมซึ่งมีคลัสเตอร์ VFe 3 S 4และช่วยให้เกิดเส้นทางทางเลือกในการตรึงไนโตรเจนในสภาวะที่ขาดโมลิบเดนัม[ 31 ]อิเล็กตรอนถูกขนส่งโดยคลัสเตอร์ "P" ที่เกี่ยวข้อง ซึ่งประกอบด้วย คลัสเตอร์ Fe 4 S 4 ทรง ลูกบาศก์ สองอัน ที่เชื่อมต่อกันด้วยสะพานซัลเฟอร์[ 32 ]
ซูเปอร์ออกไซด์ดิสมิวเทส

ไอออนซูเปอร์ออกไซด์O₂⁻− 2เกิดขึ้นในระบบชีวภาพโดยการลดออกซิเจน โมเลกุล มีอิเล็กตรอน ที่ไม่จับคู่ ดังนั้นจึงมีพฤติกรรมเหมือนอนุมูลอิสระ เป็นสารออกซิไดซ์ ที่ ทรงพลังคุณสมบัติเหล่านี้ทำให้ไอออนซูเปอร์ออกไซด์เป็นพิษ มาก และถูกนำไปใช้ประโยชน์โดยฟาโกไซต์เพื่อฆ่าจุลินทรีย์ ที่รุกราน มิฉะนั้น ไอออนซูเปอร์ออกไซด์จะต้องถูกทำลายก่อนที่จะก่อให้เกิดความเสียหายที่ไม่พึงประสงค์ในเซลล์ เอนไซม์ ซูเปอร์ออกไซด์ดิสมิวเทสทำหน้าที่นี้ได้อย่างมีประสิทธิภาพมาก[ 33 ]
สถานะออกซิเดชันอย่างเป็นทางการของอะตอมออกซิเจนคือ − 1 ⁄ 2ในสารละลายที่มีค่าpH เป็นกลาง ไอออนซูเปอร์ออกไซด์จะเกิดปฏิกิริยาไม่สมดุลกลายเป็นออกซิเจนโมเลกุลและไฮโดรเจนเปอร์ออกไซด์
- 2 โอ− 2+ 2 H + → O 2 + H 2 O 2
ในทางชีววิทยา ปฏิกิริยาประเภทนี้เรียกว่า ปฏิกิริยา ดิสมิวเทชันซึ่งเกี่ยวข้องกับการออกซิเดชันและการรีดักชันของไอออนซูเปอร์ออกไซด์กลุ่มเอนไซม์ซูเปอร์ออกไซด์ดิ สมิวเทส (SOD) จะเพิ่ม อัตราการเกิดปฏิกิริยาให้ใกล้เคียงกับอัตราที่จำกัดโดยการแพร่[ 34 ]กุญแจสำคัญในการทำงานของเอนไซม์เหล่านี้คือไอออนโลหะที่มีสถานะออกซิเดชันที่แปรผันได้ ซึ่งสามารถทำหน้าที่เป็นตัวออกซิไดซ์หรือตัวรีดิวซ์ก็ได้
- ออกซิเดชัน: M ( n +1)+ + O− 2→ M n + + O 2
- การลดลง: M n + + O− 2+ 2 H + → M ( n +1)+ + H 2 O 2 .
ใน SOD ของมนุษย์ โลหะที่ออกฤทธิ์คือทองแดงในรูป Cu(II) หรือ Cu(I) ซึ่งประสานงานแบบเตตระเฮดรัลโดยหมู่ฮิสติดีนสี่หมู่เอนไซม์นี้ยังมี ไอออน สังกะสีเพื่อความเสถียรและถูกกระตุ้นโดยทองแดงชาเปอโรนสำหรับซูเปอร์ออกไซด์ดิสมิวเทส ( CCS ) ไอ โซเอนไซม์ อื่นๆ อาจมีเหล็กแมงกานีส หรือนิกเกลกิจกรรมของ Ni-SOD เกี่ยวข้องกับนิกเกล(III) ซึ่งเป็นสถานะออกซิเดชันที่ผิดปกติสำหรับธาตุนี้ รูปทรงเรขาคณิตของนิกเกลในบริเวณออกฤทธิ์จะหมุนเวียนจากNi(II) แบบ ระนาบสี่เหลี่ยม ที่มีลิแกนด์ไทโอเลต (Cys 2และ Cys 6 ) และไนโตรเจนของโครงสร้างหลัก (His 1และ Cys 2 ) ไปเป็นNi(III) แบบพีระมิดสี่เหลี่ยม ที่มี ลิแกนด์โซ่ข้าง His 1 ตามแกนเพิ่มเข้ามา [ 35 ]
โปรตีนที่มีคลอโรฟิลล์

คลอโรฟิลล์มีบทบาทสำคัญในการสังเคราะห์แสงมันประกอบด้วยแมกนีเซียมที่ล้อมรอบด้วย วงแหวน คลอรีนอย่างไรก็ตาม ไอออนแมกนีเซียมไม่ได้มีส่วนเกี่ยวข้องโดยตรงกับกระบวนการสังเคราะห์แสง และสามารถถูกแทนที่ด้วยไอออนสองวาเลนซ์อื่นๆ ได้โดยไม่สูญเสียประสิทธิภาพมากนัก แต่โฟตอนจะถูกดูดซับโดยวงแหวนคลอรีน ซึ่งโครงสร้างอิเล็กตรอนของมันเหมาะสมอย่างยิ่งสำหรับวัตถุประสงค์นี้
ในขั้นต้น การดูดกลืนโฟตอนทำให้เกิดการกระตุ้นอิเล็กตรอน ขึ้นสู่ สถานะซิงเกล็ตของแถบ Q สถานะกระตุ้น นี้ จะเกิดการเปลี่ยนสถานะจากสถานะซิงเกล็ตไปสู่สถานะทริปเล็ตซึ่งมีอิเล็กตรอนสองตัวที่มีสปิน ขนานกัน สารชนิดนี้เป็นอนุมูลอิสระที่มีปฏิกิริยาไวมาก และสามารถถ่ายโอนอิเล็กตรอนไปยังตัวรับที่อยู่ติดกับคลอโรฟิลล์ในคลอโรพลาสต์ได้ในกระบวนการนี้ คลอโรฟิลล์จะถูกออกซิไดซ์ ต่อมาในวัฏจักรการสังเคราะห์แสง คลอโรฟิลล์จะถูกรีดิวซ์กลับมาอีกครั้ง การรีดิวซ์นี้จะดึงอิเล็กตรอนจากน้ำ ทำให้เกิดออกซิเจนโมเลกุลเป็นผลิตภัณฑ์ออกซิเดชันสุดท้าย
ไฮโดรจีเนส
ไฮโดรจีเนสถูกจำแนกย่อยออกเป็น 3 ประเภทที่แตกต่างกันตามปริมาณโลหะในบริเวณออกฤทธิ์ ได้แก่ ไฮโดรจีเนสเหล็ก-เหล็ก ไฮโดรจีเนสนิกเกิล-เหล็ก และไฮโดรจีเนสเหล็ก[ 36 ] ไฮโดรจีเนสทั้งหมดเร่งปฏิกิริยา การดูดซับ H 2 แบบย้อนกลับได้ แต่ไฮโดรจีเนส [FeFe] และ [NiFe] เป็นตัวเร่งปฏิกิริยาแบบรีดอกซ์ ที่แท้จริง ซึ่งขับเคลื่อนการออกซิเดชันของ H 2 และ การรีดักชันของ H +
- H 2 ⇌ 2 H + + 2 e −
ไฮโดรจีเนส [Fe] ทำหน้าที่เร่งปฏิกิริยา การแตกตัว แบบเฮเทอโรไลติก ที่ผันกลับได้ ของH 2
- H 2 ⇌ H + + H −

ไรโบไซม์และดีออกซีไรโบไซม์
นับตั้งแต่การค้นพบไรโบไซม์โดยThomas CechและSidney Altmanในช่วงต้นทศวรรษ 1980 ไรโบไซม์ได้รับการพิสูจน์แล้วว่าเป็นเมทัลโลเอนไซม์ประเภทหนึ่งที่แตกต่างออกไป[ 37 ]ไรโบไซม์จำนวนมากต้องการไอออนโลหะในบริเวณที่ออกฤทธิ์เพื่อการเร่งปฏิกิริยาทางเคมี ดังนั้นจึงเรียกว่าเมทัลโลเอนไซม์ นอกจากนี้ ไอออนโลหะยังจำเป็นต่อการรักษาเสถียรภาพโครงสร้างของไรโบไซม์อินทรอนกลุ่ม Iเป็นไรโบไซม์ที่ได้รับการศึกษามากที่สุดซึ่งมีโลหะสามชนิดมีส่วนร่วมในการเร่งปฏิกิริยา[ 38 ]ไรโบไซม์อื่นๆ ที่รู้จัก ได้แก่อินทรอนกลุ่ม II , RNase Pและไรโบไซม์ไวรัสขนาดเล็กหลายชนิด (เช่นแฮมเมอร์เฮด , แฮร์พิน , HDVและVS ) และหน่วยย่อยขนาดใหญ่ของไรโบโซม มีการอธิบายไรโบไซม์หลายประเภท[ 39 ]
ดีออกซีไรโบไซม์ หรือที่เรียกว่า ดีเอ็นเอไซม์ หรือดีเอ็นเอเร่งปฏิกิริยา เป็นตัวเร่งปฏิกิริยาที่สร้างขึ้นจากดีเอ็นเอ ซึ่งผลิตขึ้นครั้งแรกในปี 1994 [ 40 ]ดีเอ็นเอไซม์เกือบทั้งหมดต้องการไอออนโลหะ แม้ว่าไรโบไซม์ส่วนใหญ่จะเร่งปฏิกิริยาการแตกตัวของสารตั้งต้นอาร์เอ็นเอ แต่ดีเอ็นเอไซม์สามารถเร่งปฏิกิริยาได้หลากหลาย รวมถึงการแตกตัวของอาร์เอ็นเอ/ดีเอ็นเอ การเชื่อมต่ออาร์เอ็นเอ/ดีเอ็นเอ การฟอสโฟรีเลชันและการดีฟอสโฟรีเลชันของกรดอะมิโน และการสร้างพันธะคาร์บอน-คาร์บอน[ 41 ]อย่างไรก็ตาม ดีเอ็นเอไซม์ที่เร่งปฏิกิริยาการแตกตัวของอาร์เอ็นเอเป็นชนิดที่ได้รับการศึกษาอย่างกว้างขวางที่สุด ดีเอ็นเอไซม์ 10-23 ซึ่งค้นพบในปี 1997 เป็นหนึ่งในดีเอ็นเอเร่งปฏิกิริยาที่ได้รับการศึกษามากที่สุดและมีการประยุกต์ใช้ทางคลินิกในฐานะตัวแทนในการรักษา[ 42 ]มีการรายงาน DNAzyme ที่เฉพาะเจาะจงกับโลหะหลายชนิด ได้แก่ GR-5 DNAzyme ( เฉพาะ ตะกั่ว ) [ 43 ] CA1-3 DNAzyme ( เฉพาะ ทองแดง ) 39E DNAzyme ( เฉพาะ ยูเรนิล ) [ 44 ]และ NaA43 DNAzyme ( เฉพาะ โซเดียม ) [ 45 ]
โปรตีนโลหะที่ทำหน้าที่ส่งสัญญาณ
แคลโมดูลิน

แคลโมดูลินเป็นตัวอย่างของโปรตีนที่ทำหน้าที่ส่งสัญญาณ โปรตีนชนิดนี้มีขนาดเล็กและประกอบด้วย โมทิฟ EF-hand สี่ โมทิฟ แต่ละโมทิฟสามารถจับกับไอออน Ca 2+ ได้
ใน โดเมนโปรตีนแบบ EF-hand loop ไอออนแคลเซียมจะถูกประสานในโครงสร้างแบบพีระมิดคู่ห้าเหลี่ยมกรดกลูตามิกและ กรด แอสปาร์ติก จำนวน 6 โมเลกุลที่เกี่ยวข้องกับการจับนั้นอยู่ในตำแหน่งที่ 1, 3, 5, 7 และ 9 ของสายโพลีเปปไทด์ ที่ตำแหน่งที่ 12 มีลิแกนด์กลูตาเมตหรือแอสปาร์เทตซึ่งทำหน้าที่เป็น ลิแกนด์ แบบสองตำแหน่งโดยให้สองอะตอมออกซิเจน โมเลกุลที่เก้าในลูปจำเป็นต้องเป็นไกลซีนเนื่องจากข้อกำหนดด้านโครงสร้างของกระดูกสันหลังบริเวณการประสานของไอออนแคลเซียมประกอบด้วยอะตอมออกซิเจนของคาร์บอกซิเลตเท่านั้นและไม่มีอะตอมไนโตรเจน ซึ่งสอดคล้องกับ ลักษณะ ที่แข็งของไอออนแคลเซียม
โปรตีนนี้มีโดเมนสมมาตรโดยประมาณสองโดเมน คั่นด้วยบริเวณ "บานพับ" ที่ยืดหยุ่น การจับกับแคลเซียมทำให้เกิดการเปลี่ยนแปลงโครงสร้างของโปรตีน แคลโมดูลินมีส่วนร่วมใน ระบบ ส่งสัญญาณภายในเซลล์โดยทำหน้าที่เป็นตัวส่งสัญญาณรองที่แพร่กระจายได้ต่อสิ่งเร้าเริ่มต้น[ 46 ] [ 47 ]
โทรโปนิน
ในทั้งกล้ามเนื้อหัวใจและ กล้ามเนื้อโครง ร่าง การสร้างแรงของกล้ามเนื้อถูกควบคุมเป็นหลักโดยการเปลี่ยนแปลงความเข้มข้นของแคลเซียม ภายในเซลล์ โดยทั่วไป เมื่อแคลเซียมสูงขึ้น กล้ามเนื้อจะหดตัว และเมื่อแคลเซียมลดลง กล้ามเนื้อจะคลายตัวโทรโปนินร่วมกับแอคตินและโทรโปไมโอซินเป็นโปรตีนเชิงซ้อนที่แคลเซียมจับเพื่อกระตุ้นการสร้างแรงของกล้ามเนื้อ
ปัจจัยการถอดรหัส
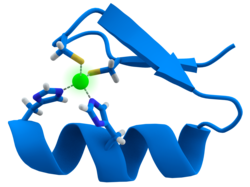
ปัจจัยการถอดรหัสจำนวนมากมีโครงสร้างที่เรียกว่านิ้วสังกะสีซึ่งเป็นโมดูลโครงสร้างที่บริเวณของโปรตีนพับรอบไอออนสังกะสี สังกะสีไม่ได้สัมผัสโดยตรงกับDNAที่โปรตีนเหล่านี้จับ แต่โคแฟคเตอร์นี้มีความสำคัญต่อความเสถียรของสายโปรตีนที่พับแน่น[ 48 ]ในโปรตีนเหล่านี้ ไอออนสังกะสีมักจะถูกประสานงานโดยคู่ของหมู่ข้างเคียงซิสเทอีนและฮิสติดีน
เอนไซม์โลหะอื่นๆ
คาร์บอนมอนอกไซด์ดีไฮโดรจีเนสมีสองประเภท: ประเภทหนึ่งประกอบด้วยเหล็กและโมลิบดีนัม อีกประเภทหนึ่งประกอบด้วยเหล็กและนิกเกล ความคล้ายคลึงและความแตกต่างในกลยุทธ์การเร่งปฏิกิริยาได้รับการทบทวนแล้ว[ 49 ]
Pb 2+ (ตะกั่ว) สามารถแทนที่ Ca 2+ (แคลเซียม) ได้ เช่น แทนที่ด้วยแคลโมดูลินหรือ Zn 2+ (สังกะสี) แทนที่ด้วยเมทัลโลคาร์บอกซีเปปติเดส[ 50 ]
เอนไซม์ที่มีโลหะเป็นองค์ประกอบอื่นๆ บางชนิดแสดงอยู่ในตารางต่อไปนี้ โดยจำแนกตามโลหะที่เกี่ยวข้อง
ดูเพิ่มเติม
ลิงก์ภายนอก
- เมทัลโลโปรตีน ใน หัวข้อทางการ แพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
- สัมมนาของแคทเธอรีน เดรนแนน: ภาพรวมของเมทัลโลโปรตีน
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เมทัลโลโปรตีน
เมทัลโลโปรตีนเป็นคำทั่วไปสำหรับโปรตีนที่มีไอออนโลหะเป็นโคแฟคเตอร์ โปรตีน ส่วนใหญ่จัดอยู่ในประเภทนี้ ตัวอย่างเช่น โปรตีนของมนุษย์อย่างน้อย 1,000 ชนิด (จากทั้งหมดประมาณ 20,000 ชนิด).
ความอุดมสมบูรณ์
มีการประมาณว่า โปรตีน ประมาณครึ่งหนึ่ง มี โลหะเป็นองค์ประกอบ [ 5 ] จาก การประมาณอีกครั้งหนึ่ง คาดว่าโปรตีนประมาณหนึ่งในสี่ถึงหนึ่งในสามต้องใช้โลหะเพื่อทำหน้าที่ [ 6 ] ดังนั้น เมทัลโลโปรตีนจึงมีหน้าที่หลากหลายใน เซลล์ เช่น การเก็บรักษา การขนส่ง...
หลักการเคมีเชิงโคออร์ดิเนชัน
ในเมทัลโลโปรตีน ไอออนของโลหะมักจะถูกประสานโดย ศูนย์กลาง ไนโตรเจน ออกซิเจนหรือ กำมะถัน ที่อยู่ใน หมู่ กรด อะมิโน ของโปรตีน หมู่ผู้ให้เหล่านี้มักจะมาจากโซ่ข้างบนหมู่กรดอะมิโน โดยเฉพาะอย่างยิ่ง หมู่แทนที่ อิมิดาโซล ในหมู่ ฮิสติ ดีน หมู่แทนที่ ไทโอเลต ใน หมู่...
โปรตีนโลหะสำหรับการจัดเก็บและขนส่ง
นี่คือผลิตภัณฑ์ขั้นที่สองของการไฮโดรไลซิสโปรตีน ซึ่งได้จากการบำบัดด้วยกรดและด่างที่มีความเข้มข้นสูงขึ้นเล็กน้อย