อ่าน 13 นาที
โปรตีนที่จับกับอาร์เอ็นเอ
โปรตีนที่จับกับ RNA (มักย่อว่า RBP ) เป็น โปรตีน ที่จับกับ RNA สายคู่หรือสายเดี่ยว [ 1 ] ใน เซลล์ และมีส่วนร่วมในการสร้าง คอมเพล็กซ์ ไรโบนิวคลีโอโปรตีน RBP ประกอบด้วย...
โปรตีนที่จับกับอาร์เอ็นเอ
โปรตีนที่จับกับ RNA (มักย่อว่าRBP ) เป็นโปรตีน ที่จับกับ RNAสายคู่หรือสายเดี่ยว[ 1 ]ในเซลล์และมีส่วนร่วมในการสร้าง คอมเพล็กซ์ ไรโบนิวคลีโอโปรตีน RBP ประกอบด้วยโครงสร้างโมทีฟ ต่างๆ เช่นโมทีฟการจดจำ RNA (RRM) โดเมนการจับ dsRNAนิ้วสังกะสีและอื่นๆ[ 2 ] [ 3 ] พวกมันเป็น โปรตีน ในไซโตพลาสซึมและนิวเคลียสอย่างไรก็ตาม เนื่องจาก RNA ที่โตเต็มที่ส่วนใหญ่จะถูกส่งออกจากนิวเคลียสค่อนข้างเร็ว RBP ส่วนใหญ่ในนิวเคลียสจึงมีอยู่ในรูปของคอมเพล็กซ์ของโปรตีนและpre-mRNAที่เรียกว่าอนุภาคไรโบนิวคลีโอโปรตีนที่ไม่เป็นเนื้อเดียวกัน (hnRNP) RBP มีบทบาทสำคัญในกระบวนการต่างๆ ของเซลล์ เช่น การทำงานของเซลล์ การขนส่ง และการกำหนดตำแหน่ง โดยเฉพาะอย่างยิ่งพวกมันมีบทบาทสำคัญในการควบคุม RNA หลังการถอดรหัส เช่นการตัดต่อการเติมโพลีอะดีนีน การทำให้mRNA เสถียร การกำหนดตำแหน่งของ mRNA และการแปล เซลล์ ยูคาริโอติกแสดงออกถึง RBP ที่หลากหลายซึ่งมีกิจกรรมการจับ RNA ที่เป็นเอกลักษณ์และการโต้ตอบระหว่างโปรตีนกับโปรตีนตามฐานข้อมูล RBP ของยูคาริโอติก (EuRBPDB) มี 2961 ยีนที่เข้ารหัส RBP ในมนุษย์ในระหว่างวิวัฒนาการความหลากหลายของ RBP เพิ่มขึ้นอย่างมากพร้อมกับการเพิ่มขึ้นของจำนวนอินทรอนความหลากหลายนี้ทำให้เซลล์ยูคาริโอติกสามารถใช้เอ็กซอน RNA ในการจัดเรียงต่างๆ ทำให้เกิด RNP (ไรโบนิวคลีโอโปรตีน) ที่เป็นเอกลักษณ์สำหรับ RNA แต่ละชนิด แม้ว่า RBP จะมีบทบาทสำคัญในการควบคุมการแสดงออกของยีนหลังการถอดรหัส แต่มี RBP เพียงไม่กี่ชนิดเท่านั้นที่ได้รับการศึกษาอย่างเป็นระบบ ปัจจุบันเป็นที่ชัดเจนแล้วว่าการโต้ตอบระหว่าง RNA กับ RBP มีบทบาทสำคัญในกระบวนการทางชีววิทยาหลายอย่างในสิ่งมีชีวิต[ 4 ] [ 5 ] [ 6 ]
โครงสร้าง
RBP จำนวนมากมีโครงสร้างแบบโมดูลาร์และประกอบด้วยการทำซ้ำหลายครั้งของโดเมนพื้นฐานเฉพาะเพียงไม่กี่โดเมนซึ่งมักจะมีลำดับที่จำกัด RBP ที่แตกต่างกันจะมีลำดับเหล่านี้เรียงกันในรูปแบบต่างๆ การจดจำ RNA เฉพาะของโปรตีนเฉพาะนั้นวิวัฒนาการมาจากการจัดเรียงใหม่ของโดเมนพื้นฐานเพียงไม่กี่โดเมนเหล่านี้ โดเมนพื้นฐานแต่ละโดเมนจะจดจำ RNA แต่โปรตีนเหล่านี้จำนวนมากต้องการสำเนาหลายชุดของโดเมนทั่วไปโดเมนใดโดเมนหนึ่งเพื่อการทำงาน[ 2 ]
ความหลากหลาย
เมื่อRNA ในนิวเคลียส เกิดขึ้นจากRNA polymerase RNA ที่ถอดรหัสแล้วจะถูกปกคลุมด้วยโปรตีนที่จับกับ RNA ทันที ซึ่งควบคุมทุกแง่มุมของการเผาผลาญและการทำงานของ RNA รวมถึงการสร้าง RNA การเจริญเติบโต การขนส่ง การกำหนดตำแหน่งในเซลล์ และความเสถียร RBP ทั้งหมดจับกับ RNA แต่พวกมันจับด้วยความจำเพาะและความสัมพันธ์ของลำดับ RNA ที่แตกต่างกัน ซึ่งทำให้ RBP มีความหลากหลายเช่นเดียวกับเป้าหมายและหน้าที่ของพวกมัน[ 5 ]เป้าหมายเหล่านี้รวมถึงmRNA ซึ่งเป็นรหัสสำหรับโปรตีน รวมถึง RNA ที่ไม่เข้ารหัสที่มีฟังก์ชันการทำงานจำนวนหนึ่งNcRNA เกือบทั้งหมดทำงานเป็น สารประกอบ ไรโบโปรตีนและไม่ใช่ RNA เปล่าๆ RNA ที่ไม่เข้ารหัสเหล่านี้รวมถึงmicroRNA , small interfering RNA (siRNA) รวมถึง spliceosomal small nuclear RNA (snRNA) [ 7 ]
การทำงาน
กระบวนการและการดัดแปลง RNA
การตัดต่อทางเลือก
การสลับการต่อเชื่อม (Alternative splicing ) เป็นกลไกที่ทำให้เกิด mRNA (messenger RNA) ในรูปแบบต่างๆ จากยีน เดียวกัน เป็นกลไกการควบคุมที่การเปลี่ยนแปลงในการรวมเอ็กซอนเข้าไปใน mRNA นำไปสู่การผลิตโปรตีนที่เกี่ยวข้องมากกว่าหนึ่งชนิด จึงเป็นการขยายผลลัพธ์ทางจีโนมที่เป็นไปได้ RBP ทำหน้าที่อย่างกว้างขวางในการควบคุมกระบวนการนี้ โปรตีนที่จับกับ RNA บางชนิด เช่น โปรตีนที่จับกับ RNA เฉพาะในเซลล์ประสาท (neuronal specific RNA-binding proteins) โดยเฉพาะNOVA1ควบคุมการสลับการต่อเชื่อมของ hnRNA บางส่วนโดยการจดจำและจับกับลำดับเฉพาะใน RNA (YCAY โดยที่ Y หมายถึงไพริมิดีน, U หรือ C) [ 5 ]จากนั้นโปรตีนเหล่านี้จะดึงดูดโปรตีนสไปลโซมไปยังตำแหน่งเป้าหมายนี้โปรตีน SRก็เป็นที่รู้จักกันดีในบทบาทของการสลับการต่อเชื่อมผ่านการดึงดูดsnRNPที่สร้างสไปลโซมโดยเฉพาะ U1 snRNP และ U2AF snRNP อย่างไรก็ตาม RBP ก็เป็นส่วนหนึ่งของสไปลโซมเองด้วย สไปลโซมเป็นคอมเพล็กซ์ของ snRNA และหน่วยย่อยโปรตีน และทำหน้าที่เป็นตัวแทนเชิงกลที่กำจัดอินทรอนและเชื่อมต่อเอ็กซอนที่อยู่ข้างเคียง[ 7 ]นอกเหนือจากคอมเพล็กซ์สไปลโซมหลักแล้ว RBP ยังจับกับตำแหน่งของ องค์ประกอบ RNA ที่ออกฤทธิ์แบบ Cisซึ่งมีอิทธิพลต่อการรวมหรือการยกเว้นเอ็กซอนในระหว่างการสไปลซิง ตำแหน่งเหล่านี้เรียกว่าตัวเสริมการสไปลซิงเอ็กซอน (ESEs) ตัวยับยั้งการสไปลซิงเอ็กซอน (ESSs) ตัวเสริมการสไปลซิงอินทรอน (ISEs) และตัวยับยั้งการสไปลซิงอินทรอน (ISSs) และขึ้นอยู่กับตำแหน่งการจับ RBP จะทำหน้าที่เป็นตัวยับยั้งหรือตัวเสริมการสไปลซิง[ 8 ]
การแก้ไขอาร์เอ็นเอ

รูปแบบการแก้ไข RNA ที่ได้รับการศึกษาอย่างกว้างขวางที่สุดเกี่ยวข้องกับ โปรตีน ADARโปรตีนนี้ทำงานผ่านการดัดแปลงหลังการถอดรหัสของ mRNA โดยการเปลี่ยน เนื้อหา นิวคลีโอไทด์ของ RNA ซึ่งทำได้โดยการเปลี่ยนอะดีโนซีนเป็นอิโนซีนในปฏิกิริยาเอนไซม์ที่เร่งปฏิกิริยาโดย ADAR กระบวนการนี้เปลี่ยนลำดับ RNA จากที่เข้ารหัสโดยจีโนม อย่างมีประสิทธิภาพ และขยายความหลากหลายของผลิตภัณฑ์ยีน การแก้ไข RNA ส่วนใหญ่เกิดขึ้นในบริเวณที่ไม่เข้ารหัสของ RNA อย่างไรก็ตาม พบว่า RNA ที่เข้ารหัสโปรตีนบางส่วนสามารถแก้ไขได้ ส่งผลให้ลำดับกรดอะมิโนของโปรตีนแตกต่างกัน ตัวอย่างเช่น mRNA ของตัวรับกลูตาเมตซึ่งกลูตามีนถูกเปลี่ยนเป็นอาร์จินีน ทำให้เกิดการเปลี่ยนแปลงในการทำงานของโปรตีน[ 5 ]
โพลีอะดีนิเลชัน
การเติมโพลีอะ ดีนิเลชันคือการเพิ่ม "หาง" ของสารตกค้างอะดีนิเลตให้กับทรานสคริปต์ RNA ประมาณ 20 เบสลงไปทางด้านล่างของลำดับ AAUAAA ภายในบริเวณที่ไม่ถูกแปลรหัสสามไพร ม์ การเติมโพลีอะดีนิเลชันของ mRNA มีผลอย่างมากต่อการขนส่งนิวเคลียร์ประสิทธิภาพการแปล และความเสถียร ทั้งหมดนี้รวมถึงกระบวนการเติมโพลีอะดีนิเลชันขึ้นอยู่กับการจับของ RBP เฉพาะ mRNA ของยูคาริโอตทั้งหมด ยกเว้นบางส่วน จะถูกประมวลผลเพื่อให้ได้รับหางโพลี (A) 3' ประมาณ 200 นิวคลีโอไทด์ หนึ่งในคอมเพล็กซ์โปรตีนที่จำเป็นในกระบวนการนี้คือCPSF CPSF จับกับลำดับหาง 3' (AAUAAA) และร่วมกับโปรตีนอีกตัวหนึ่งที่เรียกว่าโปรตีนจับโพลี (A)จะดึงดูดและกระตุ้นการทำงานของโพลี (A) โพลีเมอเรส โพลี (A) โพลีเมอเรสไม่สามารถทำงานได้ด้วยตัวเองและต้องการการจับของโปรตีนอื่นๆ เหล่านี้เพื่อให้ทำงานได้อย่างถูกต้อง[ 5 ]
ส่งออก
หลังจากการประมวลผลเสร็จสมบูรณ์ mRNA จำเป็นต้องถูกขนส่งจากนิวเคลียสของเซลล์ไปยังไซโตพลาสซึมกระบวนการนี้มีสามขั้นตอน ได้แก่ การสร้างคอมเพล็กซ์ตัวขนส่งในนิวเคลียส ตามด้วยการเคลื่อนย้ายคอมเพล็กซ์ผ่านคอมเพล็กซ์รูพรุนนิวเคลียร์และสุดท้ายคือการปล่อยสินค้าเข้าสู่ไซโตพลาสซึม จากนั้นตัวขนส่งจะถูกนำกลับมาใช้ใหม่ เชื่อกันว่าเฮเทอโรไดเมอร์ TAP/NXF1:p15 เป็นตัวสำคัญในการส่งออก mRNA การแสดงออกมากเกินไปของ TAP ใน กบ Xenopus laevisจะเพิ่มการส่งออกทรานสคริปต์ที่ปกติแล้วส่งออกได้ไม่ดี อย่างไรก็ตาม TAP ต้องการโปรตีนอะแดปเตอร์เนื่องจากไม่สามารถโต้ตอบกับ mRNA ได้โดยตรง โปรตีน Aly/REF จะโต้ตอบและจับกับ mRNA เพื่อดึงดูด TAP [ 5 ]
การระบุตำแหน่งของ mRNA
การกำหนดตำแหน่งของ mRNA มีความสำคัญต่อการควบคุมการแสดงออกของยีนโดยอนุญาตให้มีการผลิตโปรตีนที่ถูกควบคุมตามตำแหน่ง ผ่านการกำหนดตำแหน่งของ mRNA โปรตีนจะถูกแปลไปยังตำแหน่งเป้าหมายที่ตั้งใจไว้ของเซลล์ สิ่งนี้มีความสำคัญอย่างยิ่งในช่วงการพัฒนาในระยะแรกเมื่อการแบ่งเซลล์อย่างรวดเร็วทำให้เซลล์ต่างๆ มี mRNA หลายชุด ซึ่งอาจนำไปสู่ชะตากรรมของเซลล์ที่แตกต่างกันอย่างมาก โปรตีนที่จับกับ RNA (RBPs) มีบทบาทสำคัญในการกำหนดตำแหน่งของ mRNA นี้ ซึ่งทำให้มั่นใจได้ว่าโปรตีนจะถูกแปลเฉพาะในบริเวณที่ตั้งใจไว้เท่านั้น หนึ่งในโปรตีนเหล่านี้คือZBP1 ZBP1 จับกับ mRNA ของเบต้า-แอคตินที่ตำแหน่งของการถอดรหัสและเคลื่อนที่ไปพร้อมกับ mRNA เข้าสู่ไซโตพลาสซึม จากนั้นมันจะกำหนดตำแหน่งของ mRNA นี้ไปยัง บริเวณ ลามิลลาของเซลล์ประเภทอสมมาตรหลายชนิด ซึ่งสามารถแปลได้[ 5 ]ในปี 2008 มีการเสนอว่าFMRPมีส่วนเกี่ยวข้องกับการกำหนดตำแหน่งของ mRNA เดนไดรต์หลายชนิดในเดนไดรต์ของเซลล์ประสาทฮิปโปแคมปัสที่เพาะเลี้ยงโดยการกระตุ้น[ 9 ]การศึกษาล่าสุดเกี่ยวกับ RNA ที่จับกับ FMRP ที่มีอยู่ในเดนไดรต์ที่แยกส่วนของเซลล์ประสาทฮิปโปแคมปัส CA1 เผยให้เห็นว่าไม่มีการเปลี่ยนแปลงตำแหน่งในสมองของหนูป่าเทียบกับหนูที่ไม่มี FMRP [ 10 ]
การแปล
การควบคุมการแปลเป็นกลไกที่รวดเร็วในการควบคุมการแสดงออกของยีน แทนที่จะควบคุมการแสดงออกของยีนในระดับการถอดรหัส mRNA จะถูกถอดรหัสแล้ว แต่การดึงดูดไรโบโซมจะถูกควบคุม ซึ่งช่วยให้เกิดการสร้างโปรตีนอย่างรวดเร็วเมื่อสัญญาณกระตุ้นการแปล นอกจากบทบาทของ ZBP1 ในการกำหนดตำแหน่งของ mRNA ของ B-actin แล้ว ZBP1 ยังมีส่วนเกี่ยวข้องกับการยับยั้งการแปลของ mRNA ของเบต้า-แอคตินโดยการปิดกั้นการเริ่มต้นการแปล ZBP1 จะต้องถูกกำจัดออกจาก mRNA เพื่อให้ไรโบโซมสามารถจับได้อย่างถูกต้องและเริ่มการแปลได้[ 5 ]
ปฏิสัมพันธ์ระหว่างโปรตีนและอาร์เอ็นเอ

โปรตีนที่จับกับ RNA แสดงการจดจำเป้าหมาย RNA ที่เฉพาะเจาะจงสูงโดยการจดจำลำดับ โครงสร้าง ลวดลาย และการดัดแปลง RNA [ 11 ]การจับที่เฉพาะเจาะจงของโปรตีนที่จับกับ RNA ช่วยให้พวกมันสามารถแยกแยะเป้าหมายและควบคุมการทำงานของเซลล์ต่างๆ ผ่านการควบคุมการสร้าง การเจริญเติบโต และอายุขัยของ RNA ที่ถอดรหัส ปฏิสัมพันธ์นี้เริ่มต้นในระหว่างการถอดรหัส เนื่องจาก RBP บางตัวยังคงจับกับ RNA จนกว่าจะถูกย่อยสลาย ในขณะที่บางตัวจับกับ RNA เพียงชั่วคราวเพื่อควบคุม การตัดต่อ การประมวลผล การขนส่ง และการกำหนดตำแหน่งของRNA [ 12 ] วิธี การตกตะกอนภูมิคุ้มกันแบบเชื่อมโยงข้าม (CLIP) ใช้เพื่อระบุตำแหน่งการจับ RNA โดยตรงของโปรตีนที่จับกับ RNA ในเนื้อเยื่อและสิ่งมีชีวิตต่างๆ อย่างเข้มงวด ในส่วนนี้ จะกล่าวถึงโดเมนที่จับกับ RNA ที่ได้รับการศึกษาอย่างกว้างขวางที่สุด 3 ประเภท (ลวดลายการจดจำ RNA ลวดลายการจับกับ RNA สองสาย ลวดลายนิ้วสังกะสี)
ลวดลายการจดจำอาร์เอ็นเอ (RRM)
โมทีฟการจดจำอาร์เอ็นเอ (RNA recognition motif หรือ RRM ) ซึ่งเป็นโมทีฟที่จับกับอาร์เอ็นเอได้บ่อยที่สุด เป็นโดเมนโปรตีนขนาดเล็กที่มีกรดอะมิโน 75-85 ตัว โดยจะสร้างแผ่นเบต้า (β-sheet) สี่สาย ประกบกับเกลียวอัลฟา (α-helices) สองเกลียว โมทีฟการจดจำนี้มีบทบาทในหน้าที่ต่างๆ ของเซลล์มากมาย โดยเฉพาะอย่างยิ่งในกระบวนการประมวลผล mRNA/rRNA การตัดต่อ การควบคุมการแปล การส่งออกอาร์เอ็นเอ และความเสถียรของอาร์เอ็นเอ มีการระบุโครงสร้างของ RRM จำนวน 10 โครงสร้าง ผ่านการวิเคราะห์ด้วยสเปก โทรสโกปีNMRและ การ ตกผลึกด้วยรังสีเอกซ์ โครงสร้างเหล่านี้แสดงให้เห็นถึงความซับซ้อนของการจดจำโปรตีน-อาร์เอ็นเอของ RRM เนื่องจากเกี่ยวข้องกับการโต้ตอบระหว่างอาร์เอ็นเอ-อาร์เอ็นเอ และโปรตีน-โปรตีน นอกเหนือจากการโต้ตอบระหว่างโปรตีน-อาร์เอ็นเอ แม้จะมีความซับซ้อน แต่โครงสร้างทั้งสิบก็มีลักษณะร่วมกันบางประการ พบว่าแผ่นเบต้าสี่สายบนพื้นผิวโปรตีนหลักของ RRM ทุกตัวมีปฏิสัมพันธ์กับอาร์เอ็นเอ ซึ่งมักจะสัมผัสกับนิวคลีโอไทด์สองหรือสามตัวในลักษณะเฉพาะ นอกจากนี้ ความสามารถในการจับ RNA ที่แข็งแกร่งและความจำเพาะต่อการเปลี่ยนแปลงเกิดขึ้นได้จากการโต้ตอบระหว่างตัวเชื่อมระหว่างโดเมนและ RNA และระหว่าง RRM ด้วยกันเอง ความยืดหยุ่นของ RRM นี้อธิบายได้ว่าทำไม RRM จึงเป็นโดเมนที่มีมากที่สุดและทำไมจึงมีบทบาทสำคัญในหน้าที่ทางชีววิทยาต่างๆ[ 12 ]
ลวดลายการจับ RNA สองสาย
| ลวดลายการจับ RNA สองสาย | |||||||
|---|---|---|---|---|---|---|---|
 | |||||||
| ตัวระบุ | |||||||
| เครื่องหมาย | ดรัม | ||||||
| พีแฟม | พีเอฟ14709 | ||||||
| ตระกูลพีแฟม | ซีแอล0196 | ||||||
| อินเตอร์โปร | IPR014720 | ||||||
| แคท | 1di2 | ||||||
| สโคป2 | 1di2 / SCOPe / SUPFAM | ||||||
| |||||||
| ใช้กลุ่ม Pfam สำหรับซูเปอร์แฟมิลีที่คล้ายคลึงกัน | |||||||
โดเมนจับ RNA สองสาย (dsRM, dsRBD) ซึ่งประกอบด้วยกรดอะมิโน 70-75 ตัว มีบทบาทสำคัญในการประมวลผล RNAการกำหนดตำแหน่งRNA การแทรกแซง RNAการแก้ไข RNAและการยับยั้งการแปลโปรตีน โครงสร้างทั้งสามของโดเมนที่ได้รับการศึกษาจนถึงปี 2005 มีลักษณะร่วมกันที่อธิบายว่าทำไม dsRM จึงจับกับ dsRNA แทนที่จะเป็น dsDNA พบว่า dsRM มีปฏิสัมพันธ์ตามแนวเกลียวคู่ของ RNA ผ่านทั้งเกลียวอัลฟาและลูป β1-β2 ยิ่งไปกว่านั้น โครงสร้าง dsRBM ทั้งสามแบบยังสัมผัสกับโครงสร้างน้ำตาลฟอสเฟตของร่องหลักและร่องรองหนึ่งร่อง ซึ่งเชื่อมโยงโดยลูป β1-β2 ร่วมกับ บริเวณ ปลาย Nของเกลียวอัลฟา 2 ปฏิสัมพันธ์นี้เป็นการปรับตัวเฉพาะสำหรับรูปร่างของเกลียวคู่ RNA เนื่องจากเกี่ยวข้องกับ 2'-ไฮดรอกซิลและออกซิเจนของฟอสเฟต แม้ว่า dsRBM จะมีลักษณะโครงสร้างทั่วไปเหมือนกัน แต่ก็มีโครงสร้างทางเคมีที่แตกต่างกัน ซึ่งช่วยให้มีความเฉพาะเจาะจงสำหรับโครงสร้าง RNA ที่หลากหลาย รวมถึงก้านห่วง ห่วงภายใน ส่วนโป่ง หรือเกลียวที่มีการจับคู่ผิดพลาด[ 12 ]
นิ้วสังกะสี
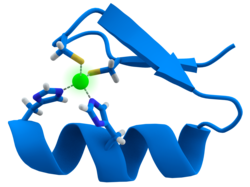
โดเมน ซิงค์ฟิงเกอร์ชนิด CCHH เป็น โดเมนจับกับ DNA ที่พบได้บ่อยที่สุดในจีโนม ของสิ่งมีชีวิตยูคาริโอต เพื่อให้สามารถจดจำลำดับ DNA ได้อย่างจำเพาะเจาะจงสูง จึงมีการใช้ซิงค์ฟิงเกอร์หลายตัวในลักษณะโมดูลาร์ ซิงค์ฟิงเกอร์มีโครงสร้างโปรตีนแบบ ββα ซึ่งβ-hairpinและ α-helix เชื่อมต่อกันด้วยอะตอมสังกะสี (Zn)2+นอกจากนี้ ปฏิสัมพันธ์ระหว่างหมู่ข้างเคียงของโปรตีนในเกลียวอัลฟา (α-helix) กับเบสของดีเอ็นเอในร่องหลัก (major groove) ช่วยให้เกิดการจดจำลำดับดีเอ็นเอจำเพาะ แม้ว่าจะสามารถจดจำดีเอ็นเอได้อย่างกว้างขวาง แต่เมื่อไม่นานมานี้มีการค้นพบว่าซิงค์ฟิงเกอร์ยังมีความสามารถในการจดจำอาร์เอ็นเอได้อีกด้วย นอกเหนือจากซิงค์ฟิงเกอร์ชนิด CCHH แล้ว ซิงค์ฟิงเกอร์ชนิด CCCH ก็เพิ่งถูกค้นพบว่าใช้การจดจำลำดับจำเพาะของอาร์เอ็นเอสายเดี่ยวผ่านปฏิสัมพันธ์ระหว่างพันธะไฮโดรเจน ระหว่างโมเลกุล และขอบวัตสัน-คริก (Watson-Crick edges) ของเบสอาร์เอ็นเอ ซิงค์ฟิงเกอร์ชนิด CCHH ใช้สองวิธีในการจับกับอาร์เอ็นเอ วิธีแรก ซิงค์ฟิงเกอร์จะออกแรงปฏิสัมพันธ์แบบไม่จำเพาะกับโครงสร้างหลักของเกลียวคู่ในขณะที่วิธีที่สองช่วยให้ซิงค์ฟิงเกอร์สามารถจดจำเบสแต่ละตัวที่ยื่นออกมาได้อย่างจำเพาะเจาะจง แตกต่างจากชนิด CCHH ซิงค์ฟิงเกอร์ชนิด CCCH แสดงโหมดการจับกับอาร์เอ็นเออีกแบบหนึ่ง ซึ่งอาร์เอ็นเอสายเดี่ยวจะถูกระบุในลักษณะจำเพาะต่อลำดับ โดยรวมแล้ว นิ้วสังกะสีสามารถจดจำ DNA ได้โดยตรงผ่านการจับกับลำดับ dsDNA และ RNA ผ่านการจับกับลำดับ ssRNA [ 12 ]
บทบาทในการพัฒนาตัวอ่อน

การควบคุมการถอดรหัสและ การควบคุมหลังการถอดรหัส ของ RNA โดยโปรตีนที่จับกับ RNA มีบทบาทในการควบคุมรูปแบบการแสดงออกของยีนในระหว่างการพัฒนา[ 13 ]การวิจัยอย่างกว้างขวางเกี่ยวกับหนอนตัวกลมC. elegansได้ระบุว่าโปรตีนที่จับกับ RNA เป็นปัจจัยสำคัญในระหว่างการพัฒนาของ เซลล์ สืบพันธุ์ และตัวอ่อนในระยะเริ่มต้น หน้าที่เฉพาะของพวกมันเกี่ยวข้องกับการพัฒนาของเนื้อเยื่อ ร่างกาย ( เซลล์ประสาทเนื้อเยื่อใต้ผิวหนังกล้ามเนื้อและเซลล์ขับถ่าย) รวมถึงการให้สัญญาณกำหนดเวลาสำหรับเหตุการณ์การพัฒนา อย่างไรก็ตาม การค้นพบกลไกเบื้องหลังการทำงานของ RBP ในการพัฒนานั้นเป็นเรื่องที่ท้าทายอย่างยิ่ง เนื่องจากความยากลำบากในการระบุเป้าหมาย RNA ของพวกมัน เนื่องจาก RBP ส่วนใหญ่มักมีเป้าหมาย RNA หลายตัว[ 14 ]อย่างไรก็ตาม เป็นที่ปฏิเสธไม่ได้ว่า RBP มีบทบาทสำคัญในการควบคุมเส้นทางการพัฒนาอย่างเป็นระบบ
การพัฒนาของเซลล์สืบพันธุ์
ในDrosophila melanogasterยีนที่เข้ารหัสโปรตีนจับ RNA ได้แก่ Elav, Sxl และ tra-2 ซึ่งมีความสำคัญต่อการกำหนดเพศ ในระยะเริ่มต้น และการรักษาสถานะทางเพศของร่างกาย[ 15 ]ยีนเหล่านี้มีผลต่อระดับหลังการถอดรหัสโดยการควบคุมการตัดต่อเฉพาะเพศในDrosophila Sxl ทำหน้าที่ควบคุมยีนtra ที่ทำให้เกิดเพศหญิงในเชิงบวก เพื่อสร้าง mRNA ของ tra ที่ใช้งานได้ในเพศเมีย ในC. elegansโปรตีนจับ RNA ได้แก่ FOG-1, MOG-1/-4/-5 และ RNP-4 ควบคุมการกำหนดเพศของเซลล์สืบพันธุ์และเซลล์ร่างกาย นอกจากนี้ RBP หลายชนิด เช่น GLD-1, GLD-3, DAZ-1, PGL-1 และ OMA-1/-2 ทำหน้าที่ควบคุมในระหว่างการดำเนินไปของระยะโปรเฟสของไมโอซิสการสร้างเซลล์สืบพันธุ์และการเจริญเติบโตของไข่[ 14 ]
การพัฒนาทางร่างกาย
นอกเหนือจากหน้าที่ของ RBP ในการพัฒนาเซลล์สืบพันธุ์แล้ว การควบคุมหลังการถอดรหัสยังมีบทบาทสำคัญในการพัฒนาเซลล์ร่างกายด้วย แตกต่างจาก RBP ที่เกี่ยวข้องกับการพัฒนาเซลล์สืบพันธุ์และตัวอ่อนระยะแรก RBP ที่ทำหน้าที่ในการพัฒนาเซลล์ร่างกายจะควบคุมการตัดต่อทางเลือกเฉพาะเนื้อเยื่อของเป้าหมาย mRNA ตัวอย่างเช่น MEC-8 และ UNC-75 ที่มีโดเมน RRM จะอยู่บริเวณใต้ผิวหนังและระบบประสาทตามลำดับ[ 14 ]นอกจากนี้ RBP ที่มี RRM อีกตัวหนึ่งคือ EXC-7 ยังพบว่าอยู่ในเซลล์ท่อขับถ่ายของตัวอ่อนและทั่วทั้งระบบประสาทในระหว่างการพัฒนาเซลล์ร่างกาย
การพัฒนาของเซลล์ประสาท
ZBP1แสดงให้เห็นว่าควบคุม การสร้าง เดนไดรต์ในเซลล์ประสาทฮิปโปแคมปัส[ 16 ]โปรตีนที่จับกับ RNA อื่นๆ ที่เกี่ยวข้องกับการสร้างเดนไดรต์ ได้แก่Pumilioและ Nanos [ 17 ] FMRP , CPEBและStaufen 1 [ 18 ]
บทบาทในโรคมะเร็ง
RBP กำลังมีบทบาทสำคัญในการพัฒนาเนื้องอก[ 19 ] RBP หลายร้อยตัวมีการควบคุมที่ผิดปกติอย่างเห็นได้ชัดในมะเร็งของมนุษย์และแสดงให้เห็นการลดระดับที่เด่นชัดในเนื้องอกที่เกี่ยวข้องกับเนื้อเยื่อปกติ[ 19 ] RBP หลายตัวมีการ แสดงออกที่แตกต่างกันในมะเร็งประเภทต่างๆ เช่น KHDRBS1 (Sam68) [ 20 ] [ 21 ] ELAVL1 (HuR) [ 22 ] [ 23 ] FXR1 [ 24 ]และUHMK1 [ 25 ]สำหรับ RBP บางตัว การเปลี่ยนแปลงในการแสดงออกเกี่ยวข้องกับการเปลี่ยนแปลงจำนวนสำเนา (CNV) เช่น การเพิ่ม CNV ของ BYSL ในเซลล์มะเร็งลำไส้ใหญ่[ 19 ]และ ESRP1, CELF3 ในมะเร็งเต้านม, RBM24 ในมะเร็งตับ, IGF2BP2, IGF2BP3 ในมะเร็งปอด หรือการสูญเสีย CNV ของ KHDRBS2 ในมะเร็งปอด[ 26 ]การเปลี่ยนแปลงการแสดงออกบางอย่างเกิดจากการกลายพันธุ์ที่ส่งผลต่อโปรตีนบน RBP เหล่านี้ เช่น NSUN6, ZC3H13, ELAC1, RBMS3และ ZGPAT, SF3B1, SRSF2, RBM10, U2AF1, SF3B1, PPRC1, RBMXL1, HNRNPCL1 เป็นต้น[ 19 ] [ 26 ] [ 27 ] [ 28 ] [ 29 ]การศึกษาหลายชิ้นเชื่อมโยงการเปลี่ยนแปลงการแสดงออกของ RBP นี้กับการตัดต่อทางเลือกที่ผิดปกติในมะเร็ง[ 26 ] [ 30 ] [ 31 ]
งานวิจัยปัจจุบัน

เนื่องจากโปรตีนที่จับกับ RNA มีบทบาทสำคัญในการควบคุมการทำงานของเซลล์หลายอย่าง จึงเป็นหัวข้อที่นักวิจัยหลายคนให้ความสนใจเป็นอย่างมาก ด้วยความสำคัญในด้านชีววิทยา จึงมีการค้นพบมากมายเกี่ยวกับศักยภาพของโปรตีนที่จับกับ RNA ในช่วงไม่นานมานี้[ 12 ]การพัฒนาล่าสุดในการระบุโปรตีนที่จับกับ RNA ด้วยวิธีการทดลองได้ขยายจำนวนโปรตีนที่จับกับ RNA อย่างมีนัยสำคัญ[ 32 ] [ 33 ] [ 34 ]
โปรตีนที่จับกับ RNA ชื่อ Sam68 ควบคุมการแบ่งส่วนเชิงพื้นที่และเวลาของการเผาผลาญ RNA เพื่อให้ การทำงานของ ไซแนปส์ในเดนไดรต์ เป็นไปอย่างเหมาะสม การสูญเสีย Sam68 ส่งผลให้เกิดการควบคุมหลังการถอดรหัสที่ผิดปกติ และในที่สุดนำไปสู่ความผิดปกติทางระบบประสาทเช่นกลุ่มอาการสั่น/เดินเซที่เกี่ยวข้องกับกลุ่มอาการฟราจิลเอ็กซ์พบว่า Sam68 มีปฏิสัมพันธ์กับ mRNA ที่เข้ารหัสβ-actinซึ่งควบคุมการสร้างไซแนปส์ของหนามเดนไดรต์ด้วยส่วนประกอบของโครงร่างเซลล์ดังนั้น Sam68 จึงมีบทบาทสำคัญในการควบคุมจำนวนไซแนปส์ผ่านการควบคุมการเผาผลาญ mRNA ของ β-actin หลังไซแนปส์[ 35 ]
โปรตีนจับ RNA ตระกูล CELF เฉพาะเซลล์ประสาท UNC-75 จับกับ mRNA สาย UUGUUGUGUUGU โดยเฉพาะผ่านโมทีฟการจดจำ RNA สามโมทีฟสำหรับการเลือกเอ็กซอน 7a ใน เซลล์ประสาท ของ C. elegansเนื่องจากเอ็กซอน 7a ถูกข้ามไปเนื่องจากไซต์การเชื่อมต่อที่อ่อนแอในเซลล์ที่ไม่ใช่เซลล์ประสาท จึงพบว่า UNC-75 กระตุ้นการเชื่อมต่อระหว่างเอ็กซอน 7a และเอ็กซอน 8 เฉพาะในเซลล์ประสาทเท่านั้น[ 36 ]
โปรตีนCIRBP ที่จับกับ RNA ที่เหนี่ยวนำ โดย ความเย็น มีบทบาทในการควบคุมการตอบสนองของเซลล์เมื่อเผชิญกับความเครียดของเซลล์หลายชนิด รวมถึงแสงอัลตราไวโอเลต ความยาวคลื่นสั้น ภาวะ ขาดออกซิเจนและภาวะอุณหภูมิต่ำการวิจัยนี้ให้ผลลัพธ์ที่อาจบ่งชี้ถึงความสัมพันธ์ระหว่างสภาวะของโรคกับการอักเสบ[ 37 ]
พบว่าโปรตีนจับ RNA ตระกูลเซอรีน-อาร์จินีน Slr1 มีบทบาทในการควบคุมการเจริญเติบโตแบบมีขั้วในCandida albicansการกลายพันธุ์ของ Slr1 ในหนูส่งผลให้การสร้างเส้นใยลดลงและลดความเสียหายต่อเซลล์เยื่อบุผิวและ เซลล์บุผนังหลอดเลือด ซึ่งนำไปสู่การมีอัตราการรอดชีวิตที่ยาวนานขึ้นเมื่อเทียบกับสายพันธุ์ Slr1 ชนิดปกติ ดังนั้น การวิจัยนี้จึงแสดงให้เห็นว่าโปรตีนคล้าย SR อย่าง Slr1 มีบทบาทในการกระตุ้นการสร้างเส้นใยและความรุนแรงในC. albicans [ 38 ]
ดูเพิ่มเติม
ลิงก์ภายนอก
- แพลตฟอร์ม starBase : แพลตฟอร์มสำหรับถอดรหัสตำแหน่งการจับของโปรตีนที่จับกับ RNA (RBPs) จากชุดข้อมูล CLIP-Seqขนาดใหญ่(HITS-CLIP, PAR-CLIP, iCLIP, CLASH)
- ฐานข้อมูล RBPDB : ฐานข้อมูลโปรตีนที่จับกับ RNA
- oRNAment : ฐานข้อมูลของตำแหน่งการจับของ RBP ที่คาดการณ์ได้ใน RNA ทั้งส่วนที่เข้ารหัสและไม่เข้ารหัสในสิ่งมีชีวิตหลากหลายชนิด
- ฐานข้อมูล ATTRACt : ฐานข้อมูลของโปรตีนที่จับกับ RNA และลวดลายที่เกี่ยวข้อง
- SplicedAid-F : ฐานข้อมูลโปรตีนที่จับกับ RNA ของมนุษย์ซึ่งได้จากการขูดด้วยมือ
- RsiteDB : ฐานข้อมูลตำแหน่งการจับของ RNA
- SPOT-Seq-RNA : การทำนายโปรตีนที่จับกับ RNA และโครงสร้างเชิงซ้อนของพวกมันโดยใช้แม่แบบ
- SPOT-Struct-RNA ถูกเก็บถาวรเมื่อวันที่ 24 พฤษภาคม 2019 ในWayback Machine : การทำนายโปรตีนที่จับกับ RNA จากโครงสร้างสามมิติ
- โครงการ ENCODE : ชุดข้อมูลจีโนมิกส์ (เช่น RNA Bind-n-seq, eCLIP, RBP targeted shRNA RNA-seq) สำหรับ RBP
- ฐานข้อมูลภาพ RBP ที่เก็บถาวรเมื่อวันที่ 21 ตุลาคม 2018 ในWayback Machine : ภาพแสดงตำแหน่งของ RBP ภายในเซลล์
- ซอฟต์แวร์ RBPSpot : ซอฟต์แวร์ที่ใช้การเรียนรู้เชิงลึก (Deep-Learning) เพื่อตรวจจับปฏิสัมพันธ์ระหว่าง RBP และ RNA อย่างแม่นยำสูง นอกจากนี้ยังมีโมดูลสำหรับสร้างแบบจำลองปฏิสัมพันธ์ระหว่าง RBP และ RNA ใหม่ๆ อีกด้วย
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ โปรตีนที่จับกับอาร์เอ็นเอ
โปรตีนที่จับกับ RNA (มักย่อว่า RBP ) เป็น โปรตีน ที่จับกับ RNA สายคู่หรือสายเดี่ยว [ 1 ] ใน เซลล์ และมีส่วนร่วมในการสร้าง คอมเพล็กซ์ ไรโบนิวคลีโอโปรตีน RBP ประกอบด้วย...
โครงสร้าง
RBP จำนวนมากมีโครงสร้างแบบโมดูลาร์และประกอบด้วยการทำซ้ำหลายครั้งของโดเมนพื้นฐานเฉพาะเพียงไม่กี่โดเมนซึ่งมักจะมีลำดับที่จำกัด RBP ที่แตกต่างกันจะมีลำดับเหล่านี้เรียงกันในรูปแบบต่างๆ การจดจำ RNA...
ความหลากหลาย
เมื่อ RNA ในนิวเคลียส เกิดขึ้นจาก RNA polymerase RNA ที่ถอดรหัสแล้วจะถูกปกคลุมด้วยโปรตีนที่จับกับ RNA ทันที ซึ่งควบคุมทุกแง่มุมของการเผาผลาญและการทำงานของ RNA รวมถึงการสร้าง RNA การเจริญเติบโต การขนส่ง การกำหนดตำแหน่งในเซลล์ และความเสถียร RBP ทั้งหมดจับกับ...
กระบวนการและการดัดแปลง RNA
การสลับการต่อเชื่อม (Alternative splicing ) เป็นกลไกที่ทำให้เกิด mRNA (messenger RNA) ในรูปแบบต่างๆ จาก ยีน เดียวกัน เป็นกลไกการควบคุมที่การเปลี่ยนแปลงในการรวมเอ็ กซอน เข้าไปใน mRNA นำไปสู่การผลิตโปรตีนที่เกี่ยวข้องมากกว่าหนึ่งชนิด...