อ่าน 52 นาที
ดีเอ็นเอ
กรดดีออกซีไรโบนิวคลีอิก ( การออกเสียง ⓘ ; [ 1 ] DNA ) เป็น พอลิเมอร์ ที่ประกอบด้วย โพลีนิวคลีโอไทด์ สายที่พันกันเป็น เกลียวคู่ ลิเมอร์นี้บรรจุ ทางพันธุกรรม สำหรับการพัฒนา การทำงาน...
ดีเอ็นเอ


| ส่วนหนึ่งของชุดบทความเกี่ยวกับ |
| พันธุศาสตร์ |
|---|
 |
|
กรดดีออกซีไรโบนิวคลีอิก (ⓘ ; [ 1 ] DNA) เป็นพอลิเมอร์ที่ประกอบด้วยโพลีนิวคลีโอไทด์สายที่พันกันเป็นเกลียวคู่ลิเมอร์นี้บรรจุทางพันธุกรรมสำหรับการพัฒนา การทำงาน การเจริญเติบโต และการสืบพันธุ์ของสิ่งมีชีวิตและไวรัสDNA และกรดไรโบนิวคลีอิก(RNA) เป็นกรดนิวคลีอิกร่วมกับโปรตีนไขมันและคาร์โบไฮเดรตเชิงซ้อน (พอลิแซ็กคาไรด์) กรดนิวคลีอิกเป็นหนึ่งในสี่ประเภทหลักของโมเลกุลที่จำเป็นต่อสิ่งมีชีวิตแบบ
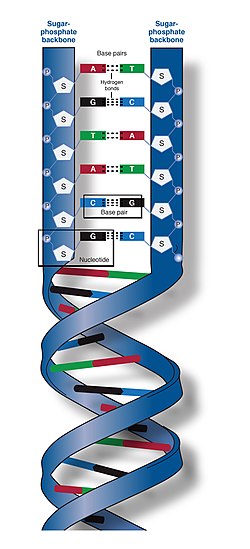
สาย DNA สองสายนี้เรียกว่าโพลีนิวคลีโอไทด์ เนื่องจากประกอบด้วยหน่วยโมโนเมอร์ที่ เรียบง่ายกว่าที่เรียกว่านิ ว คลีโอ ไทด์[ 2 ] [ 3 ] นิวคลีโอ ไทด์แต่ละตัวประกอบด้วยนิว คลีโอเบส ที่มีไนโตรเจน 4 ชนิด ( ไซโตซีน [C], กัวนีน [G], อะดีนีน [A] หรือไทมีน [T]) น้ำตาลที่เรียกว่าดีออกซีไรโบสและหมู่ฟอสเฟต นิว คลีโอ ไทด์จะเชื่อมต่อกันเป็นโซ่ด้วยพันธะโควา เลนต์ (เรียกว่าพันธะฟอสโฟไดเอสเทอร์ ) ระหว่างน้ำตาลของนิวคลีโอไทด์หนึ่งกับฟอสเฟตของนิวคลีโอไทด์ถัดไป ทำให้เกิดโครงสร้างหลักแบบสลับน้ำตาล-ฟอสเฟต เบส ที่มี ไนโตรเจนของสายโพลีนิวคลีโอไทด์สองสายที่แยกจากกันจะเชื่อมต่อกันตาม กฎ การจับคู่เบส (A กับ T และ C กับ G) ด้วยพันธะไฮโดรเจนเพื่อสร้าง DNA สองสาย เบสไนโตรเจนที่เป็นคู่สมแบ่งออกเป็นสองกลุ่ม คือ ไพริมิดีนที่มีวงแหวนเดี่ยว และ พิวรีนที่มีวงแหวนคู่ในดีเอ็นเอ ไพริมิดีนได้แก่ ไทมีนและไซโตซีน ส่วนพิวรีนได้แก่ อะดีนีนและกัวนีน
สาย DNA สองสายเก็บข้อมูลทางชีวภาพ เดียวกัน ข้อมูลนี้จะถูกจำลองเมื่อสายทั้งสองแยกออกจากกัน สาย DNA ทั้งสองสายวิ่งไปในทิศทางตรงกันข้ามกัน จึงเรียกว่า แอนติ พาราเล ล แต่ละโมเลกุลของน้ำตาลจะมีนิวคลีโอเบส (หรือ เบส ) สี่ชนิดติดอยู่ลำดับของนิวคลีโอเบสทั้งสี่ชนิดนี้ตามแนวกระดูกสันหลังเป็นตัวเข้ารหัสข้อมูลทางพันธุกรรม สาย RNA ถูกสร้างขึ้นโดยใช้สาย DNA เป็นแม่แบบในกระบวนการที่เรียกว่าการถอดรหัสโดยที่เบสของ DNA จะถูกแลกเปลี่ยนกับเบสที่สอดคล้องกัน ยกเว้นในกรณีของไทมีน (T) ซึ่ง RNA จะแทนที่ด้วยยูราซิล (U) [ 4 ]ภายใต้รหัสพันธุกรรมสาย RNA เหล่านี้จะระบุลำดับของกรดอะมิโนภายในโปรตีนในกระบวนการที่เรียกว่าการแปล
ภายในเซลล์ยูคาริโอต DNA จะถูกจัดเรียงเป็นโครงสร้างยาวที่เรียกว่าโครโมโซมก่อนการแบ่งเซลล์ตาม ปกติ โครโมโซมเหล่านี้จะถูกจำลองขึ้นในกระบวนการจำลอง DNA ทำให้แต่ละเซลล์ลูกมีโครโมโซมครบชุด สิ่ง มีชีวิตยูคา ริโอต (สัตว์ พืช เชื้อรา และโปรติสต์)เก็บDNAส่วนใหญ่ไว้ภายในนิวเคลียสของเซลล์ในรูปของDNA นิวเคลียร์ และบางส่วนอยู่ในไมโทคอนเดรียในรูป ของ DNA ไมโทคอนเดรียหรือในคลอโรพลาสต์ ใน รูปของ DNA คลอโรพลาสต์ [ 5 ] ในทางตรงกันข้ามโปรคาริโอต ( แบคทีเรียและอาร์เคีย ) เก็บ DNA ไว้เฉพาะในไซโทพลาซึมใน รูป ของโครโมโซมวงกลมภายในโครโมโซมของยูคาริโอต โปรตีน โครมาตินเช่นฮิสโตนจะทำให้ DNA อัดแน่นและจัดระเบียบ โครงสร้างที่อัดแน่นเหล่านี้จะนำทางปฏิสัมพันธ์ระหว่าง DNA และโปรตีนอื่นๆ ช่วยควบคุมว่าส่วนใดของ DNA จะถูกถอดรหัส
คุณสมบัติ
ดีเอ็นเอเป็นพอลิเมอร์ สายยาว ที่สร้างจากหน่วยซ้ำๆ ที่เรียกว่านิวคลีโอไทด์ [ 6 ] [ 7 ] โดย ปกติดีเอ็นเอไม่ได้อยู่ในรูปสายเดี่ยว แต่เป็นสายคู่ที่ยึดติดกันแน่น[ 8 ] [ 9 ]สายยาวสองสายนี้จะพันกันเป็นเกลียวคู่ นิวคลี โอไทด์ประกอบด้วยส่วนของโครงสร้างหลักของโมเลกุล (ซึ่งยึดสายโซ่ไว้ด้วยกัน) และนิวคลีโอเบส (ซึ่งมีปฏิสัมพันธ์กับสายดีเอ็นเออีกสายหนึ่งในเกลียว) นิวคลีโอเบสที่เชื่อมต่อกับน้ำตาลเรียกว่านิวคลีโอไซด์และเบสที่เชื่อมต่อกับน้ำตาลและหมู่ฟอสเฟตหนึ่งหมู่หรือมากกว่าเรียกว่านิวคลีโอไทด์ พอลิเมอร์ชีวภาพที่ประกอบด้วยนิวคลีโอไทด์ที่เชื่อมต่อกันหลายตัว (เช่นในดีเอ็นเอ) เรียกว่าพอลินิวคลีโอไทด์[ 10 ]
โครงสร้างของ DNA นั้นมีพลวัตตลอดความยาว สามารถม้วนตัวเป็นวงแน่นและรูปร่างอื่นๆ ได้[ 11 ]ในทุกสปีชีส์ DNA ประกอบด้วยสายเกลียวสองสายที่ยึดติดกันด้วยพันธะไฮโดรเจนสายทั้งสองจะม้วนตัวรอบแกนเดียวกัน (แต่ไม่ได้อยู่ตรงข้ามกันโดยตรง) และมีระยะห่าง เท่ากัน ที่ 34 อังสตรอม (3.4 นาโนเมตร ) หากไม่คำนึงถึงเอกลักษณ์ของเบสกลุ่มสายคือ D 1 (q) โดยที่ q หมายถึงจำนวนรอบต่อคู่เบส ซึ่งประมาณหนึ่งในสิบของรอบเต็ม ในรูปแบบปกติ (B-DNA) การบิดจะเป็นแบบมือขวา เหมือนสกรูปกติที่หมุนตามเข็มนาฬิกาไปเรื่อยๆ คู่ของสายมีรัศมี 10 อังสตรอม (1.0 นาโนเมตร) [ 8 ]จากการศึกษาอีกชิ้นหนึ่ง เมื่อวัดในสารละลายที่แตกต่างกัน สาย DNA มีความกว้าง 22–26 Å (2.2–2.6 nm) และหน่วยนิวคลีโอไทด์หนึ่งหน่วยมีความยาว 3.3 Å (0.33 nm) [ 12 ]ความหนาแน่นลอยตัวของ DNA ส่วนใหญ่คือ 1.7 กรัม/ ซม³ [ 13 ]
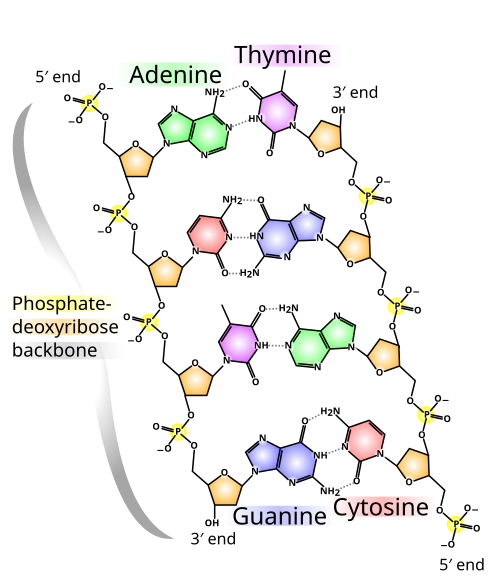
โครงสร้างหลักของสาย DNA สร้างขึ้นจากกลุ่มฟอสเฟตและน้ำตาล ที่สลับกัน [ 14 ]น้ำตาลใน DNA คือ2-ดีออกซีไรโบสซึ่งเป็น น้ำตาล เพนโทส (น้ำตาลห้าคาร์บอน ) น้ำตาลเหล่านี้เชื่อมต่อกันด้วยกลุ่มฟอสเฟตที่สร้างพันธะฟอสโฟไดเอสเทอร์ ระหว่าง อะตอมคาร์บอนที่สามและห้าของวงแหวนน้ำตาลที่อยู่ติดกัน อะตอมเหล่านี้เรียกว่า คาร์บอน ปลาย 3′ (ปลายสามไพรม์) และ คาร์บอน ปลาย 5′ (ปลายห้าไพรม์) โดยใช้สัญลักษณ์ไพรม์เพื่อแยกความแตกต่างระหว่างอะตอมคาร์บอนเหล่านี้กับอะตอมคาร์บอนของเบสที่ดีออกซีไรโบสสร้างพันธะไกลโคไซด์[ 9 ]
ดังนั้น โดยปกติแล้วสายดีเอ็นเอจะมีปลายด้านหนึ่งที่มีหมู่ฟอสเฟตติดอยู่กับคาร์บอน 5′ ของไรโบส (ฟอสฟอริล 5′) และปลายอีกด้านหนึ่งที่มีหมู่ไฮดรอกซิลอิสระติดอยู่กับคาร์บอน 3′ ของไรโบส (ไฮดรอกซิล 3′) การเรียงตัวของคาร์บอน 3′ และ 5′ ตามโครงสร้างน้ำตาล-ฟอสเฟตทำให้สายดีเอ็นเอแต่ละสายมีทิศทาง (บางครั้งเรียกว่าขั้ว) ใน เกลียวคู่ของกรดนิวคลีอิกทิศทางของนิวคลีโอไทด์ในสายหนึ่งจะตรงข้ามกับทิศทางในอีกสายหนึ่ง: สายทั้งสองเป็นแบบแอนติพาราเลลปลายที่ไม่สมมาตรของสายดีเอ็นเอจะมีทิศทางเป็นปลาย 5′ และปลาย 3′ โดยปลาย 5′ มีหมู่ฟอสเฟตที่ปลายสุด และปลาย 3′ มีหมู่ไฮดรอกซิลที่ปลายสุด ความแตกต่างที่สำคัญอย่างหนึ่งระหว่าง DNA และRNAคือน้ำตาล โดย 2-ดีออกซีไรโบสใน DNA จะถูกแทนที่ด้วยน้ำตาลเพนโทสที่เกี่ยวข้องคือไรโบ ส ใน RNA [ 9 ]

{kind=link}
โครงสร้างเกลียวคู่ของ DNA มีความเสถียรโดยแรงสองแรงหลัก ได้แก่พันธะไฮโดรเจนระหว่างนิวคลีโอไทด์และ ปฏิกิริยาการ เรียงซ้อนของเบสระหว่างนิวคลีโอเบสอะโรมา ติก [ 16 ]เบสทั้งสี่ที่พบใน DNA คืออะดีนีน ( A ), ไซโตซีน ( C ), กัวนีน ( G ) และไทมีน ( T ) เบสทั้งสี่นี้จะเชื่อมต่อกับน้ำตาลฟอสเฟตเพื่อสร้างนิวคลีโอไทด์ที่สมบูรณ์ ดังแสดงในอะดีโนซีนโมโนฟอสเฟตอะดีนีนจับคู่กับไทมีนและกัวนีนจับคู่กับไซโตซีน ก่อให้เกิดคู่เบสATและGC [ 17 ] [ 18 ]
การจำแนกประเภทนิวคลีโอเบส
นิวคลีโอเบสแบ่งออกเป็นสองประเภท ได้แก่พิวรีน A และ G ซึ่ง เป็น สารประกอบเฮเทอโรไซคลิกห้าและหกสมาชิกที่หลอมรวมกันและไพริมิดีน ซึ่ง เป็นวงแหวนหกสมาชิกCและT [ 9 ]นิวคลีโอเบสไพริมิดีนตัวที่ห้า คือยูราซิล ( U ) มักจะเข้ามาแทนที่ไทมีนใน RNA และแตกต่างจากไทมีนตรงที่ไม่มีหมู่เมทิลบนวงแหวน นอกจาก RNA และ DNA แล้ว ยัง มีการสร้าง อะนาล็อกของกรดนิวคลีอิก เทียมจำนวนมาก เพื่อศึกษาคุณสมบัติของกรดนิวคลีอิก หรือเพื่อใช้ในด้านเทคโนโลยีชีวภาพ[ 19 ]
ฐานที่ไม่เป็นไปตามแบบแผน
เบสที่ถูกดัดแปลงเกิดขึ้นใน DNA เบสแรกที่ถูกค้นพบคือ5-เมทิลไซโตซีนซึ่งพบในจีโนมของMycobacterium tuberculosisในปี พ.ศ. 2468 [ 20 ]เหตุผลของการมีเบสที่ไม่เป็นไปตามแบบแผนเหล่านี้ในไวรัสแบคทีเรีย ( แบคทีริโอเฟจ ) ก็เพื่อหลีกเลี่ยงเอนไซม์จำกัดที่มีอยู่ในแบคทีเรีย ระบบเอนไซม์นี้ทำหน้าที่อย่างน้อยบางส่วนเป็นระบบภูมิคุ้มกันระดับโมเลกุลที่ปกป้องแบคทีเรียจากการติดเชื้อไวรัส[ 21 ]การดัดแปลงเบสไซโตซีนและอะดีนีน ซึ่งเป็นเบส DNA ที่พบได้บ่อยและถูกดัดแปลง มีบทบาทสำคัญใน การควบคุมการแสดงออกของยีน แบบเอพิเจเนติกในพืชและสัตว์[ 22 ]
เป็นที่ทราบกันว่ามีเบสที่ไม่เป็นไปตามแบบแผนจำนวนหนึ่งเกิดขึ้นใน DNA [ 23 ]ส่วนใหญ่เป็นการดัดแปลงเบสที่เป็นไปตามแบบแผนบวกกับยูราซิล
- อะดีนีนที่ดัดแปลงแล้ว
- เอ็น6-คาร์บาโมอิล-เมทิลอะดีนีน
- เอ็น6-เมทิลอะดีนีน
- กัวนีนที่ดัดแปลงแล้ว
- 7-ดีอะซากัวนีน
- 7-เมทิลกัวนีน
- ไซโตซีนที่ถูกดัดแปลง
- เอ็น4-เมทิลไซโตซีน
- 5-คาร์บอกซิลไซโตซีน
- 5-ฟอร์มิลไซโตซีน
- 5-ไกลโคซิลไฮดรอกซีเมทิลไซโตซีน
- 5-ไฮดรอกซีไซโตซีน
- 5-เมทิลไซโตซีน
- ไทมิดีนดัดแปลง
- α-กลูตาไมไทมิดีน
- α-พิวเทรซินิลไทมีน
- ยูราซิลและการดัดแปลง
- ฐานเจ
- ยูราซิล
- 5-ไดไฮดรอกซีเพนทอราซิล
- 5-ไฮดรอกซีเมทิลดีออกซียูราซิล
- คนอื่น
- ดีออกซีอาร์คีโอซีน
- 2,6-ไดอะมิโนพิวรีน (2-อะมิโนอะดีนีน)
ร่อง

สายเกลียวคู่ก่อตัวเป็นโครงสร้างหลักของ DNA อาจพบเกลียวคู่อีกอันหนึ่งตามช่องว่างหรือร่องระหว่างสาย ช่องว่างเหล่านี้อยู่ติดกับคู่เบสและอาจเป็นตำแหน่งการจับเนื่องจากสายไม่ได้อยู่สมมาตรกัน ร่องจึงมีขนาดไม่เท่ากัน ร่องหลักมีความกว้าง 22 อังสตรอม (2.2 นาโนเมตร) ในขณะที่ร่องรองมีความกว้าง 12 อังสตรอม (1.2 นาโนเมตร) [ 24 ]เนื่องจากความกว้างของร่องหลักที่มากกว่า ขอบของเบสจึงเข้าถึงได้ง่ายกว่าในร่องหลักมากกว่าในร่องรอง ส่งผลให้โปรตีน เช่นปัจจัยการถอดรหัสที่สามารถจับกับลำดับเฉพาะใน DNA สองสาย มักจะสัมผัสกับด้านข้างของเบสที่เปิดเผยในร่องหลัก[ 25 ]สถานการณ์นี้แตกต่างกันไปตามโครงสร้าง DNA ที่ผิดปกติภายในเซลล์(ดูด้านล่าง) แต่ร่องหลักและร่องรองจะถูกตั้งชื่อเพื่อสะท้อนถึงความแตกต่างในความกว้างที่จะเห็นได้หาก DNA ถูกบิดกลับไปเป็นรูป ทรง Bปกติ
การจับคู่เบส
 |
 |
ในเกลียวคู่ของ DNA นิวคลีโอเบสแต่ละชนิดบนสายหนึ่งจะจับคู่กับนิวคลีโอเบสเพียงชนิดเดียวบนสายอื่น เรียกว่าการจับคู่เบสแบบเสริมกัน พิวรีนสร้างพันธะไฮโดรเจนกับไพริมิดีน อะดีนีนจับคู่กับไทมีนเพียงสองพันธะไฮโดรเจน และไซโตซีนจับคู่กับกัวนีนเพียงสามพันธะไฮโดรเจน การจัดเรียงของนิวคลีโอไทด์สองตัวที่จับคู่กันข้ามเกลียวคู่ (จากวงแหวนคาร์บอนหกตัวไปยังวงแหวนคาร์บอนหกตัว) เรียกว่าคู่เบสแบบวัตสัน-คริก DNA ที่มีปริมาณ GC สูง จะมีเสถียรภาพมากกว่า DNA ที่มีปริมาณGC ต่ำ คู่เบสแบบฮูกสตีน (พันธะไฮโดรเจนระหว่างวงแหวนคาร์บอนหกตัวกับวงแหวนคาร์บอนห้าตัว) เป็นรูปแบบที่หายากของการจับคู่เบส[ 26 ]เนื่องจากพันธะไฮโดรเจนไม่ใช่พันธะโควาเลนต์จึงสามารถแตกและเชื่อมต่อใหม่ได้ค่อนข้างง่าย สาย DNA สองสายในเกลียวคู่สามารถแยกออกจากกันได้เหมือนซิป ไม่ว่าจะด้วยแรงทางกลหรืออุณหภูมิสูง[ 27 ]ผลจากการจับคู่เบสนี้ ข้อมูลทั้งหมดในลำดับเกลียวคู่ของ DNA จะถูกทำซ้ำบนแต่ละสาย ซึ่งมีความสำคัญต่อการจำลองแบบ DNA ปฏิสัมพันธ์ที่ย้อนกลับได้และเฉพาะเจาะจงระหว่างคู่เบสที่เสริมกันนี้มีความสำคัญต่อการทำงานทั้งหมดของ DNA ในสิ่งมีชีวิต[ 7 ]
ssDNA เทียบกับ dsDNA
โมเลกุล DNA ส่วนใหญ่ประกอบด้วยสายพอลิเมอร์สองสายที่เชื่อมต่อกันในลักษณะเกลียวด้วยพันธะที่ไม่ใช่โคเวเลนต์ โครงสร้างแบบสองสาย (dsDNA) นี้คงอยู่ได้ส่วนใหญ่ด้วยปฏิกิริยาการเรียงซ้อนของเบสภายในสาย ซึ่งแข็งแรงที่สุดสำหรับ เบส G และ Cสายทั้งสองสามารถแยกออกจากกันได้—กระบวนการนี้เรียกว่าการหลอมละลาย—เพื่อสร้างโมเลกุล DNA สายเดี่ยว (ssDNA) สองโมเลกุล การหลอมละลายเกิดขึ้นที่อุณหภูมิสูง เกลือต่ำ และค่า pH สูง (ค่า pH ต่ำก็ทำให้ DNA หลอมละลายได้เช่นกัน แต่เนื่องจาก DNA ไม่เสถียรเนื่องจากการกำจัดเบสพิวรีนด้วยกรด จึงไม่ค่อยได้ใช้ค่า pH ต่ำ)
ความเสถียรของโครงสร้าง dsDNA ไม่ได้ขึ้นอยู่กับ ปริมาณ GC ( เปอร์เซ็นต์ของคู่เบส G,C ) เพียงอย่างเดียว แต่ยังขึ้นอยู่กับลำดับ (เนื่องจากการเรียงตัวเป็นแบบจำเพาะตามลำดับ) และความยาวด้วย (โมเลกุลที่ยาวกว่าจะมีความเสถียรมากกว่า) สามารถวัดความเสถียรได้หลายวิธี วิธีที่นิยมใช้คืออุณหภูมิหลอมเหลว (หรือเรียกว่าค่า Tm ) ซึ่งเป็นอุณหภูมิที่โมเลกุลแบบสองสาย 50% เปลี่ยนเป็นโมเลกุลแบบสายเดี่ยว อุณหภูมิหลอมเหลวขึ้นอยู่กับความแรงของไอออนและความเข้มข้นของ DNA ดังนั้นทั้งเปอร์เซ็นต์ของ คู่เบส GCและความยาวโดยรวมของเกลียวคู่ DNA จึงเป็นตัวกำหนดความแข็งแรงของการเชื่อมโยงระหว่างสองสายของ DNA เกลียว DNA ยาวที่มี ปริมาณ GC สูง จะมีสายที่โต้ตอบกันอย่างแข็งแรงกว่า ในขณะที่เกลียวสั้นที่มี ปริมาณ AT สูง จะมีสายที่โต้ตอบกันอย่างอ่อนกว่า[ 28 ]ในทางชีววิทยา ส่วนของเกลียวคู่ DNA ที่จำเป็นต้องแยกออกจากกันได้ง่าย เช่น กล่อง TATAAT Pribnowในโปรโมเตอร์ บางตัว มักจะมี ปริมาณ AT สูง ทำให้สาย DNA แยกออกจากกันได้ง่ายขึ้น[ 29 ]
ในห้องปฏิบัติการ ความแข็งแรงของปฏิสัมพันธ์นี้สามารถวัดได้โดยการหาอุณหภูมิหลอมเหลวT mที่จำเป็นในการทำลายพันธะไฮโดรเจนครึ่งหนึ่ง เมื่อคู่เบสทั้งหมดในเกลียวคู่ DNA หลอมเหลว สายจะแยกออกจากกันและมีอยู่ในสารละลายเป็นโมเลกุลอิสระสองโมเลกุล โมเลกุล DNA สายเดี่ยวเหล่านี้ไม่มีรูปร่างทั่วไปเพียงแบบเดียว แต่บางโครงสร้างมีความเสถียรมากกว่าโครงสร้างอื่น[ 30 ]
จำนวน

ในมนุษย์จีโนมนิวเคลียร์แบบดิพลอยด์ของ เพศหญิง ต่อเซลล์ทั้งหมดมีความยาว 6.37 กิกะเบสแพร์ (Gbp) ยาว 208.23 เซนติเมตร และหนัก 6.51 พิโคกรัม (pg) [ 31 ]ค่าของเพศชายคือ 6.27 Gbp, 205.00 เซนติเมตร, 6.41 pg [ 31 ]โพลีเมอร์ DNA แต่ละตัวสามารถบรรจุนิวคลีโอไทด์ได้หลายร้อยล้านตัว เช่นในโครโมโซม 1โครโมโซม 1 เป็นโครโมโซม ที่ใหญ่ที่สุดของมนุษย์ มีเบสแพร์ประมาณ 220 ล้านคู่และจะเป็นถ้ายืดตรงจะยาว85 มม . [ 32 ]
ในยูคาริโอตนอกเหนือจากDNA ใน นิวเคลียส แล้ว ยังมีDNA ในไมโทคอนเดรีย (mtDNA) ซึ่งเข้ารหัสโปรตีนบางชนิดที่ใช้โดยไมโทคอนเดรีย โดยปกติ mtDNA จะมีขนาดค่อนข้างเล็กเมื่อเทียบกับ DNA ในนิวเคลียส ตัวอย่างเช่นDNA ในไมโทคอนเดรียของมนุษย์ก่อตัวเป็นโมเลกุลวงกลมปิด ซึ่งแต่ละโมเลกุลประกอบด้วยคู่เบส DNA 16,569 คู่ [ 33 ] [ 34 ] [ 35 ]โดยแต่ละโมเลกุลดังกล่าวโดยปกติจะมีชุดยีนไมโทคอนเดรียครบถ้วน ไมโทคอนเดรียของมนุษย์แต่ละอันมีโมเลกุล mtDNA ดังกล่าวโดยเฉลี่ยประมาณ 5 โมเลกุล[ 35 ]เซลล์มนุษย์แต่ละ เซลล์ มีไมโทคอนเดรียประมาณ 100 ไมโทคอนเดรีย ทำให้มีโมเลกุล mtDNA รวมประมาณ 500 โมเลกุลต่อเซลล์มนุษย์[ 35 ]อย่างไรก็ตาม จำนวนไมโทคอนเดรียต่อเซลล์ก็แตกต่างกันไปตามชนิดของเซลล์ และเซลล์ไข่สามารถมีไมโทคอนเดรียได้ถึง 100,000 ไมโทคอนเดรีย ซึ่งเทียบเท่ากับสำเนาจีโนมไมโทคอนเดรียมากถึง 1,500,000 ชุด (คิดเป็น 90% ของ DNA ในเซลล์) [ 36 ]
ความหมายและความไม่มีความหมาย
ลำดับDNAเรียกว่าลำดับ "เซนส์" หากเป็นลำดับเดียวกับสำเนาของอาร์เอ็นเอส่งสารที่ถูกแปลเป็นโปรตีน[ 37 ]ลำดับบนสายตรงข้ามเรียกว่าลำดับ "แอนติเซนส์" ทั้งลำดับเซนส์และแอนติเซนส์สามารถมีอยู่ได้ในส่วนต่างๆ ของสาย DNA เดียวกัน (กล่าวคือ ทั้งสองสายสามารถมีทั้งลำดับเซนส์และแอนติเซนส์ได้) ในทั้งโปรคาริโอตและยูคาริโอต มีการสร้างลำดับอาร์เอ็นเอแอนติเซนส์ แต่หน้าที่ของอาร์เอ็นเอเหล่านี้ยังไม่ชัดเจนนัก[ 38 ]ข้อเสนอหนึ่งคือ อาร์เอ็นเอแอนติเซนส์มีส่วนเกี่ยวข้องในการควบคุมการแสดงออกของยีนผ่านการจับคู่เบสของอาร์เอ็นเอ-อาร์เอ็นเอ[ 39 ]
ลำดับดีเอ็นเอจำนวนเล็กน้อยในโปรคาริโอตและยูคาริโอต และจำนวนมากในพลาสมิดและไวรัสทำให้ความแตกต่างระหว่างสายดีเอ็นเอแบบมีทิศทางและแบบไม่มีทิศทางไม่ชัดเจนเนื่องจากมียีนที่ทับซ้อนกัน [ 40 ]ในกรณีเหล่านี้ ลำดับดีเอ็นเอบางส่วนทำหน้าที่สองอย่าง คือเข้ารหัสโปรตีนหนึ่งตัวเมื่ออ่านตามสายหนึ่ง และเข้ารหัสโปรตีนตัวที่สองเมื่ออ่านในทิศทางตรงกันข้ามตามสายอื่น ในแบคทีเรียการทับซ้อนนี้อาจเกี่ยวข้องกับการควบคุมการถอดรหัสยีน[ 41 ]ในขณะที่ในไวรัส ยีนที่ทับซ้อนกันจะเพิ่มปริมาณข้อมูลที่สามารถเข้ารหัสได้ภายในจีโนมไวรัสขนาดเล็ก[ 42 ]
การขดตัวซ้อน
ดีเอ็นเอสามารถบิดตัวได้เหมือนเชือกในกระบวนการที่เรียกว่าการขดตัวของดีเอ็นเอ (DNA supercoiling ) เมื่อดีเอ็นเออยู่ในสภาวะ "ผ่อนคลาย" สายดีเอ็นเอจะพันรอบแกนของเกลียวคู่ทุกๆ 10.4 คู่เบส แต่ถ้าดีเอ็นเอบิดตัว สายดีเอ็นเอจะพันกันแน่นขึ้นหรือหลวมลง[ 43 ]ถ้าดีเอ็นเอบิดตัวไปในทิศทางของเกลียว จะเรียกว่าการขดตัวแบบบวก (positive supercoiling) และเบสจะยึดติดกันแน่นขึ้น ถ้าบิดตัวไปในทิศทางตรงกันข้าม จะเรียกว่าการขดตัวแบบลบ (negative supercoiling) และเบสจะแยกออกจากกันได้ง่ายขึ้น ในธรรมชาติ ดีเอ็นเอส่วนใหญ่มีการขดตัวแบบลบเล็กน้อยที่เกิดจากเอนไซม์ที่เรียกว่าโทโปไอโซเมอเรส (topoisomerases ) [ 44 ]เอนไซม์เหล่านี้ยังจำเป็นต่อการบรรเทาความเครียดจากการบิดตัวที่เกิดขึ้นในสายดีเอ็นเอระหว่างกระบวนการต่างๆ เช่นการถอดรหัสและการจำลองดีเอ็นเอ[ 45 ]
โครงสร้าง DNA ทางเลือก

DNA มีโครงสร้างที่ เป็นไปได้หลายแบบ ซึ่งรวมถึงรูปแบบA-DNA , B-DNAและZ-DNA แม้ว่าจะมีเพียง B-DNA และ Z-DNA เท่านั้นที่ได้รับการสังเกตโดยตรงในสิ่งมีชีวิตที่มีการทำงาน [ 14 ]โครงสร้างที่ DNA ใช้ขึ้นอยู่กับระดับไฮเดรชั่น ลำดับ DNA ปริมาณและทิศทางของการขดตัว การดัดแปลงทางเคมีของเบส ชนิดและความเข้มข้นของไอออน โลหะ และการมีอยู่ของโพลีเอมีนในสารละลาย[ 46 ]
รายงานฉบับแรกที่ตีพิมพ์เกี่ยวกับ รูปแบบ การเลี้ยวเบนรังสีเอกซ์ ของ A-DNA— และ B-DNA ด้วย—ใช้การวิเคราะห์ตามฟังก์ชันแพตเตอร์สันซึ่งให้ข้อมูลโครงสร้างเพียงเล็กน้อยสำหรับเส้นใย DNA ที่มีทิศทาง[ 47 ] [ 48 ]การวิเคราะห์ทางเลือกได้รับการเสนอโดย Wilkins et al.ในปี 1953 สำหรับรูปแบบการเลี้ยวเบนและการกระเจิงรังสีเอกซ์ของ B-DNA ในร่างกาย ของเส้นใย DNA ที่มีน้ำมากในแง่ของกำลังสองของ ฟังก์ชันเบสเซล [ 49 ] ในวารสารเดียวกันJames WatsonและFrancis Crickได้นำเสนอ การวิเคราะห์ แบบจำลองโมเลกุลของรูปแบบการเลี้ยวเบนรังสีเอกซ์ของ DNA เพื่อแนะนำว่าโครงสร้างเป็นเกลียวคู่[ 8 ]
แม้ว่ารูปแบบ B-DNAจะพบได้บ่อยที่สุดภายใต้สภาวะที่พบในเซลล์[ 50 ]แต่ก็ไม่ใช่โครงสร้างที่กำหนดไว้อย่างชัดเจน แต่เป็นกลุ่มของโครงสร้าง DNA ที่เกี่ยวข้อง[ 51 ]ซึ่งเกิดขึ้นที่ระดับความชุ่มชื้นสูงที่มีอยู่ในเซลล์ รูปแบบการเลี้ยวเบนและการกระเจิงของรังสีเอกซ์ที่สอดคล้องกันนั้นมีลักษณะเฉพาะของพาราคริสตัลโมเลกุลที่มีระดับ ความไม่เป็นระเบียบสูง [ 52 ] [ 53 ]
เมื่อเปรียบเทียบกับ B-DNA แล้ว A-DNA จะเป็น เกลียว ขวา ที่กว้างกว่า มีร่องเล็กที่ตื้นและกว้าง และร่องใหญ่ที่แคบและลึกกว่า รูปแบบ A เกิดขึ้นภายใต้สภาวะที่ไม่ใช่สรีรวิทยาในตัวอย่าง DNA ที่ขาดน้ำบางส่วน ในขณะที่ในเซลล์อาจผลิตขึ้นในรูปแบบการจับคู่แบบไฮบริดของสาย DNA และ RNA และในเอนไซม์-DNA คอมเพล็กซ์[ 54 ] [ 55 ]ส่วนของ DNA ที่เบสได้รับการดัดแปลงทางเคมีโดยการเมทิลเลชันอาจมีการเปลี่ยนแปลงโครงสร้างที่ใหญ่ขึ้นและมีรูปแบบ Zโดยที่สายจะหมุนรอบแกนเกลียวในเกลียวซ้าย ซึ่งตรงข้ามกับรูปแบบ B ที่พบได้ทั่วไป[ 56 ]โครงสร้างที่ผิดปกติเหล่านี้สามารถจดจำได้โดยโปรตีนที่จับกับ Z-DNA โดยเฉพาะ และอาจเกี่ยวข้องกับการควบคุมการถอดรหัส[ 57 ]
เคมีดีเอ็นเอทางเลือก
เป็นเวลาหลายปีที่นักชีววิทยานอก โลก ได้เสนอการมีอยู่ของชีวภาคเงา ซึ่งเป็น ชีวภาคจุลินทรีย์ที่คาดการณ์ไว้ของโลกที่ใช้กระบวนการทางชีวเคมีและโมเลกุลที่แตกต่างอย่างสิ้นเชิงจากสิ่งมีชีวิตที่รู้จักในปัจจุบัน หนึ่งในข้อเสนอคือการมีอยู่ของสิ่งมีชีวิตที่ใช้สารหนูแทนฟอสฟอรัสในดีเอ็นเอรายงานในปี 2010 เกี่ยวกับความเป็นไปได้ในแบคทีเรียGFAJ-1ได้รับการประกาศ[ 58 ] [ 59 ]แม้ว่าการวิจัยจะถูกโต้แย้ง[ 59 ] [ 60 ]และหลักฐานชี้ให้เห็นว่าแบคทีเรียป้องกันการรวมตัวของสารหนูเข้าสู่โครงสร้างดีเอ็นเอและโมเลกุลชีวภาพอื่น ๆ อย่างแข็งขัน[ 61 ]
โครงสร้างควอดรูเพล็กซ์
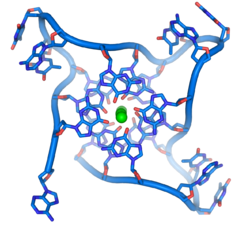
ที่ปลายของโครโมโซมเชิงเส้นจะมีบริเวณ DNA พิเศษที่เรียกว่าเทโลเมียร์หน้าที่หลักของบริเวณเหล่านี้คือการอนุญาตให้เซลล์จำลองปลายโครโมโซมโดยใช้เอนไซม์เทโลเมอเรสเนื่องจากเอนไซม์ที่ปกติใช้ในการจำลอง DNA ไม่สามารถคัดลอกปลาย 3′ สุดขั้วของโครโมโซมได้[ 63 ]ปลายโครโมโซมพิเศษเหล่านี้ยังช่วยปกป้องปลาย DNA และหยุด ระบบ ซ่อมแซม DNAในเซลล์ไม่ให้มองว่าเป็นความเสียหายที่ต้องแก้ไข[ 64 ]ในเซลล์มนุษย์เทโลเมียร์มักจะเป็นความยาวของ DNA สายเดี่ยวที่มีการทำซ้ำหลายพันครั้งของลำดับ TTAGGG อย่างง่าย[ 65 ]
ลำดับที่มีกัวนีนเป็นองค์ประกอบหลักเหล่านี้อาจทำให้ปลายโครโมโซมมีเสถียรภาพโดยการสร้างโครงสร้างของชุดหน่วยสี่เบสที่เรียงซ้อนกัน แทนที่จะเป็นคู่เบสแบบปกติที่พบในโมเลกุล DNA อื่นๆ โดยที่เบสกัวนีนสี่ตัวที่เรียกว่ากัวนีนเตตระดจะก่อตัวเป็นแผ่นเรียบ หน่วยสี่เบสแบบแบนเหล่านี้จะเรียงซ้อนกันเพื่อสร้างโครงสร้างG-quadruplex ที่มีเสถียรภาพ [ 66 ]โครงสร้างเหล่านี้มีเสถียรภาพโดยพันธะไฮโดรเจนระหว่างขอบของเบสและการคีเลตของไอออนโลหะที่อยู่ตรงกลางของแต่ละหน่วยสี่เบส[ 67 ]นอกจากนี้ยังสามารถสร้างโครงสร้างอื่นๆ ได้ โดยชุดเบสสี่ตัวตรงกลางอาจมาจากสายเดี่ยวที่พับรอบเบส หรือสายขนานหลายสายที่แตกต่างกัน โดยแต่ละสายจะให้เบสหนึ่งตัวแก่โครงสร้างตรงกลาง
นอกจากโครงสร้างที่ซ้อนกันเหล่านี้แล้ว เทโลเมียร์ยังสร้างโครงสร้างห่วงขนาดใหญ่ที่เรียกว่าห่วงเทโลเมียร์ หรือ T-loop อีกด้วย โดยที่ DNA สายเดี่ยวจะม้วนตัวเป็นวงกลมยาวโดยมีโปรตีนที่จับกับเทโลเมียร์เป็นตัวช่วยในการทรงตัว[ 68 ]ที่ปลายสุดของ T-loop นั้น DNA สายเดี่ยวของเทโลเมียร์จะถูกยึดไว้กับบริเวณของ DNA สายคู่โดยสายเทโลเมียร์ ทำให้โครงสร้างเกลียวคู่ของ DNA แตกออก และเกิดการจับคู่เบสกับหนึ่งในสองสาย โครงสร้างสามสายนี้เรียกว่าห่วงการแทนที่ หรือD-loop [ 66 ]
ดีเอ็นเอแบบแตกแขนง
 |  |
| สาขาเดียว | หลายสาขา |
ใน DNA การแตกแขนงเกิดขึ้นเมื่อมีบริเวณที่ไม่เข้าคู่กันอยู่ที่ปลายของสายคู่ DNA ที่ปกติแล้วเข้าคู่กัน อย่างไรก็ตาม DNA แบบแตกแขนงสามารถเกิดขึ้นได้หากมีการนำสาย DNA ที่สามเข้ามาและมีบริเวณที่อยู่ติดกันซึ่งสามารถไฮบริดกับบริเวณที่แตกแขนงของสายคู่ที่มีอยู่ก่อนแล้วได้ แม้ว่าตัวอย่างที่ง่ายที่สุดของ DNA แบบแตกแขนงจะเกี่ยวข้องกับสาย DNA เพียงสามสาย แต่คอมเพล็กซ์ที่เกี่ยวข้องกับสายเพิ่มเติมและแขนงหลายแขนงก็เป็นไปได้เช่นกัน[ 69 ] DNA แบบแตกแขนงสามารถใช้ในนาโนเทคโนโลยีเพื่อสร้างรูปทรงเรขาคณิต ดูส่วนการใช้งานในเทคโนโลยีด้านล่าง
ฐานเทียม
นิวคลีโอเบสเทียมหลายชนิดได้รับการสังเคราะห์และรวมเข้ากับดีเอ็นเออะนาล็อกแปดเบสที่เรียกว่าHachimoji DNA ได้สำเร็จ เบสเทียมเหล่านี้มีชื่อว่า S, B, P และ Z ซึ่งสามารถจับคู่กันได้ในแบบที่คาดการณ์ได้ (S–B และ P–Z) รักษาโครงสร้างเกลียวคู่ของดีเอ็นเอ และถอดรหัสเป็นอาร์เอ็นเอ การมีอยู่ของพวกมันอาจถือเป็นข้อบ่งชี้ว่าไม่มีอะไรพิเศษเกี่ยวกับนิวคลีโอเบสธรรมชาติสี่ชนิดที่วิวัฒนาการบนโลก[ 70 ] [ 71 ]ในทางกลับกัน ดีเอ็นเอมีความสัมพันธ์อย่างใกล้ชิดกับอาร์เอ็นเอซึ่งไม่เพียงแต่ทำหน้าที่เป็นตัวถอดรหัสของดีเอ็นเอเท่านั้น แต่ยังทำหน้าที่เป็นเครื่องจักรโมเลกุลที่ทำงานหลายอย่างในเซลล์ เพื่อจุดประสงค์นี้ ดีเอ็นเอต้องพับตัวเป็นโครงสร้าง มีการแสดงให้เห็นว่าเพื่อให้สามารถสร้างโครงสร้างที่เป็นไปได้ทั้งหมด จำเป็นต้องมีเบสอย่างน้อยสี่เบสสำหรับอาร์เอ็นเอที่สอดคล้องกัน[ 72 ]ในขณะที่จำนวนที่มากกว่านั้นก็เป็นไปได้เช่นกัน แต่จะขัดกับหลักการตามธรรมชาติของการใช้ความพยายามน้อยที่สุด
ความเป็นกรด
หมู่ฟอสเฟตของ DNA ทำให้ มีคุณสมบัติ เป็นกรด คล้าย กับกรดฟอสฟอริกและสามารถถือได้ว่าเป็นกรดแก่เมื่อค่า pH ปกติของเซลล์เป็นปกติ DNA จะแตกตัวเป็นไอออนอย่างสมบูรณ์ ปล่อยโปรตอน ออกมา ซึ่งจะทิ้งประจุลบไว้บนหมู่ฟอสเฟต ประจุลบเหล่านี้ช่วยปกป้อง DNA จากการแตกตัวด้วยไฮโดรไลซิสโดยการขับไล่นิวคลีโอไฟล์ที่อาจไฮโดรไลซิส DNA ได้[ 73 ]
ลักษณะที่มองเห็นได้ด้วยตาเปล่า

DNA บริสุทธิ์ที่สกัดจากเซลล์จะก่อตัวเป็นก้อนสีขาวเป็นเส้นๆ[ 74 ]
การดัดแปลงทางเคมีและการบรรจุ DNA ที่เปลี่ยนแปลงไป
การดัดแปลงเบสและการบรรจุดีเอ็นเอ
 | 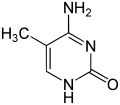 |  |
| ไซโตซีน | 5-เมทิลไซโตซีน | ไทมีน |
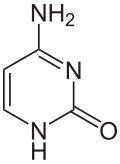
การแสดงออกของยีนได้รับอิทธิพลจากวิธีการบรรจุ DNA ในโครโมโซม ในโครงสร้างที่เรียกว่าโครมาตินการดัดแปลงเบสอาจเกี่ยวข้องกับการบรรจุ โดยบริเวณที่มีการแสดงออกของยีนต่ำหรือไม่มีเลยมักจะมีระดับเมทิลเลชันของ เบส ไซโตซีน สูง การบรรจุ DNA และอิทธิพลต่อการแสดงออกของยีนยังสามารถเกิดขึ้นได้จากการดัดแปลงโควาเลนต์ของ แกนโปรตีน ฮิสโตนซึ่ง DNA ถูกพันรอบในโครงสร้างโครมาติน หรือโดยการปรับโครงสร้างใหม่ที่ดำเนินการโดยคอมเพล็กซ์การปรับโครงสร้างโครมาติน (ดูการปรับโครงสร้างโครมาติน ) นอกจากนี้ยังมีการสื่อสารระหว่างเมทิลเลชันของ DNA และการดัดแปลงฮิสโตน ดังนั้นจึงสามารถส่งผลต่อโครมาตินและการแสดงออกของยีนได้อย่างประสานงานกัน[ 75 ]
ตัวอย่างเช่น การเมทิลเลชันของไซโตซีนทำให้เกิด5-เมทิลไซโตซีนซึ่งมีความสำคัญต่อ การปิดใช้งาน โครโมโซมX [ 76 ]ระดับเฉลี่ยของการเมทิลเลชันจะแตกต่างกันไปในแต่ละสิ่งมีชีวิต—หนอนCaenorhabditis elegansขาดการเมทิลเลชันของไซโตซีน ในขณะที่สัตว์มีกระดูกสันหลังมีระดับที่สูงกว่า โดยมี DNA มากถึง 1% ที่มี 5-เมทิลไซโตซีน[ 77 ]แม้ว่า 5-เมทิลไซโตซีนจะมีความสำคัญ แต่ก็สามารถ เกิดการดีอะมิเน ชันเพื่อเหลือเบสไทมีนได้ ดังนั้นไซโตซีนที่ถูกเมทิลเลชันจึงมีแนวโน้มที่จะเกิดการกลายพันธุ์ได้ง่ายเป็นพิเศษ[ 78 ] การดัดแปลงเบสอื่นๆ ได้แก่ การเมทิลเลชันของอะดีนีนในแบคทีเรีย การมีอยู่ของ 5-ไฮดรอกซีเมทิลไซโตซีนในสมอง [ 79 ] และการไกลโคซิเลชันของยูราซิลเพื่อสร้าง "เบส J" ในคิเนโตพลาสติด[ 80 ] [ 81 ]
ความเสียหาย

ดีเอ็นเอสามารถถูกทำลายได้ด้วยสารก่อกลาย พันธุ์หลายชนิด ซึ่งจะเปลี่ยนลำดับของดีเอ็นเอสารก่อกลายพันธุ์ได้แก่ สารออกซิไดซ์ สารอัลคิเลตและรังสีแม่เหล็กไฟฟ้าพลังงานสูงเช่นรังสีอัลตราไวโอเลต และ รังสีเอ็กซ์ประเภทของความเสียหายของดีเอ็นเอที่เกิดขึ้นจะขึ้นอยู่กับประเภทของสารก่อกลายพันธุ์ ตัวอย่างเช่น รังสี UV สามารถทำลายดีเอ็นเอได้โดยการสร้างไทมีนไดเมอร์ซึ่งเป็นการเชื่อมโยงระหว่างเบสไพริมิดีน[ 83 ]ในทางกลับกัน สารออกซิไดซ์ เช่นอนุมูลอิสระหรือไฮโดรเจนเปอร์ออกไซด์ก่อให้เกิดความเสียหายหลายรูปแบบ รวมถึงการดัดแปลงเบส โดยเฉพาะอย่างยิ่งกัวโนซีน และการแตกของสายคู่[ 84 ]เซลล์มนุษย์ทั่วไปมีเบสประมาณ 150,000 เบสที่ได้รับความเสียหายจากออกซิเดชัน[ 85 ]ในบรรดารอยโรคจากออกซิเดชันเหล่านี้ การแตกของสายคู่เป็นอันตรายที่สุด เนื่องจากซ่อมแซมได้ยากและสามารถก่อให้เกิดการกลายพันธุ์แบบจุดการ แทรก การลบออกจากลำดับดีเอ็นเอ และการ ย้ายตำแหน่ง ของโครโมโซม[ 86 ]การกลายพันธุ์เหล่านี้สามารถก่อให้เกิดมะเร็งได้ เนื่องจากข้อจำกัดโดยธรรมชาติของกลไกการซ่อมแซม DNA หากมนุษย์มีอายุยืนยาวพอ พวกเขาทุกคนก็จะเป็นมะเร็งในที่สุด[ 87 ] [ 88 ]ความเสียหายของ DNA ที่เกิดขึ้นตามธรรมชาติอันเนื่องมาจากกระบวนการของเซลล์ปกติที่สร้างอนุมูลออกซิเจน กิจกรรมไฮโดรไลซิสของน้ำในเซลล์ ฯลฯ ก็เกิดขึ้นบ่อยครั้งเช่นกัน แม้ว่าความเสียหายส่วนใหญ่เหล่านี้จะได้รับการซ่อมแซม แต่ในเซลล์ใดๆ ก็อาจยังมีความเสียหายของ DNA หลงเหลืออยู่บ้าง แม้จะมีการทำงานของกระบวนการซ่อมแซมก็ตาม ความเสียหายของ DNA ที่เหลืออยู่เหล่านี้จะสะสมมากขึ้นตามอายุในเนื้อเยื่อหลังการแบ่งตัวของสัตว์เลี้ยงลูกด้วยนม การสะสมนี้ดูเหมือนจะเป็นสาเหตุสำคัญที่อยู่เบื้องหลังความชรา[ 89 ] [ 90 ] [ 91 ]
สารก่อกลายพันธุ์หลายชนิดสามารถแทรกตัวเข้าไปในช่องว่างระหว่างคู่เบสที่อยู่ติดกันได้ ซึ่งเรียกว่าการแทรกตัว (intercalation ) สารแทรกตัวส่วนใหญ่เป็นโมเลกุลอะโรมาติกและแบนราบ ตัวอย่างเช่นเอทิเดียมโบรไมด์อะคริดีนดาวโนไมซินและด็อกโซรูบิซินเพื่อให้สารแทรกตัวสามารถแทรกตัวเข้าไประหว่างคู่เบสได้ เบสจะต้องแยกออกจากกัน ทำให้สายดีเอ็นเอเสียรูปทรงโดยการคลายเกลียวของเกลียวคู่ ซึ่งจะยับยั้งทั้งการถอดรหัสและการจำลองดีเอ็นเอ ทำให้เกิดความเป็นพิษและการกลายพันธุ์[ 92 ]ดังนั้น สารแทรกตัวในดีเอ็นเอจึงอาจเป็นสารก่อมะเร็งและในกรณีของทาลิโดไมด์ อาจเป็นสารก่อความ พิการ แต่กำเนิด [ 93 ]ส่วนสารอื่นๆ เช่นเบนโซ[ a ]ไพรีนไดออลอีพอกไซด์และอะฟลาทอกซินจะสร้างสารประกอบดีเอ็นเอที่ทำให้เกิดข้อผิดพลาดในการจำลอง[ 94 ]อย่างไรก็ตาม เนื่องจากความสามารถในการยับยั้งการถอดรหัสและการจำลองดีเอ็นเอ สารพิษที่คล้ายคลึงกันอื่นๆ จึงถูกนำมาใช้ในเคมีบำบัดเพื่อยับยั้งเซลล์มะเร็ง ที่เติบโตอย่างรวดเร็ว [ 95 ]
หน้าที่ทางชีวภาพ

โดยทั่วไป DNA จะอยู่ในรูปโครโมโซม เชิงเส้น ในยูคาริโอตและโครโมโซมวงกลมในโปรคาริโอตชุดของโครโมโซมในเซลล์ประกอบขึ้น เป็น จีโนม ของเซลล์ นั้นจีโนมของมนุษย์มี DNA ประมาณ 3 พันล้านคู่เบสเรียงตัวเป็น 46 โครโมโซม[ 96 ]ข้อมูลที่บรรจุอยู่ใน DNA จะถูกเก็บไว้ในลำดับของชิ้นส่วน DNA ที่เรียกว่ายีนการส่งต่อ ข้อมูลทางพันธุกรรมใน ยีนเกิดขึ้นผ่านการจับคู่เบสที่เสริมกัน ตัวอย่างเช่น ในการถอดรหัส เมื่อเซลล์ใช้ข้อมูลในยีน ลำดับ DNA จะถูกคัดลอกเป็นลำดับ RNA ที่เสริมกันผ่านแรงดึงดูดระหว่าง DNA และนิวคลีโอไทด์ RNA ที่ถูกต้อง โดยปกติแล้ว สำเนา RNA นี้จะถูกนำไปใช้สร้างลำดับโปรตีน ที่ตรงกัน ในกระบวนการที่เรียกว่าการแปลซึ่งขึ้นอยู่กับปฏิสัมพันธ์เดียวกันระหว่างนิวคลีโอไทด์ RNA ในอีกวิธีหนึ่ง เซลล์อาจคัดลอกข้อมูลทางพันธุกรรมในกระบวนการที่เรียกว่าการจำลองแบบ DNAรายละเอียดของฟังก์ชันเหล่านี้จะกล่าวถึงในบทความอื่น ๆ ในที่นี้จะเน้นไปที่ปฏิสัมพันธ์ระหว่าง DNA กับโมเลกุลอื่นๆ ที่เป็นตัวกลางในการทำงานของจีโนม
จีโนม
ดีเอ็นเอจีโนมิกถูกบรรจุอย่างแน่นหนาและเป็นระเบียบในกระบวนการที่เรียกว่าการควบแน่นของดีเอ็นเอเพื่อให้พอดีกับปริมาตรเล็กๆ ที่มีอยู่ในเซลล์ ในยูคาริโอต ดีเอ็นเอจะอยู่ในนิวเคลียสของเซลล์โดยมีปริมาณเล็กน้อยในไมโทคอนเดรียและคลอโรพลาสต์ในโปรคาริโอต ดีเอ็นเอจะถูกเก็บไว้ภายในโครงสร้างที่มีรูปร่างไม่สม่ำเสมอในไซโตพลาสซึมที่เรียกว่านิวคลีออยด์[ 97 ]
ข้อมูลทางพันธุกรรมที่มีหน้าที่สำคัญในจีโนมนั้นตั้งอยู่ในยีน ลำดับควบคุม จุดเริ่มต้นของการจำลองแบบ เซนโทรเมียร์ เทโลเมียร์ และส่วนต่างๆ ที่จำเป็นสำหรับโครงสร้างสามมิติของโครมาติน ในยูคาริโอตที่ซับซ้อนหลายชนิด มีเพียงส่วนน้อยของลำดับทั้งหมดเท่านั้นที่อุทิศให้กับองค์ประกอบที่มีหน้าที่สำคัญต่างๆ ตัวอย่างเช่น ในมนุษย์ จีโนมน้อยกว่า 10% มีบทบาทหน้าที่ที่กำหนดไว้ และส่วนที่เหลือ (90%) อาจเป็นดีเอ็นเอไร้ประโยชน์ (ดูเพิ่มเติมที่ [ดีเอ็นเอที่ไม่เข้ารหัส])
การถอดเสียงและการแปล
ยีนคือลำดับของดีเอ็นเอที่บรรจุข้อมูลทางพันธุกรรมและสามารถส่งผลต่อลักษณะภายนอกของสิ่งมีชีวิตได้ ภายในยีน ลำดับของเบสตามสายดีเอ็นเอจะกำหนด ลำดับของ อาร์เอ็นเอผู้ส่งสารซึ่งจะกำหนดลำดับของโปรตีนหนึ่งตัวหรือมากกว่านั้น ความสัมพันธ์ระหว่างลำดับนิวคลีโอไทด์ของยีนและ ลำดับ กรดอะมิโนของโปรตีนถูกกำหนดโดยกฎการแปลซึ่งเรียกรวมกันว่ารหัสพันธุกรรม รหัสพันธุกรรมประกอบด้วย "คำ" สามตัวอักษรที่เรียกว่าโคดอนซึ่งเกิดจากลำดับของนิวคลีโอไทด์สามตัว (เช่น ACT, CAG, TTT)
ในกระบวนการถอดรหัสพันธุกรรม โคดอนของยีนจะถูกคัดลอกไปยังอาร์เอ็นเอส่งสาร(mRNA) โดยเอนไซม์ อาร์เอ็นเอพอลิเมอเรส จากนั้นสำเนาอาร์เอ็นเอจะถูกถอดรหัสโดยไรโบโซมซึ่งจะอ่านลำดับอาร์เอ็นเอโดยการจับคู่เบสระหว่างอาร์เอ็นเอส่งสารกับ อาร์เอ็นเอ ถ่ายโอน (tRNA) ซึ่งทำหน้าที่ขนส่งกรดอะมิโน เนื่องจากมีเบส 4 ชนิดในแต่ละชุดตัวอักษร 3 ตัว จึงมีโคดอนที่เป็นไปได้ 64 แบบ (4³ ชุด ) โคดอนเหล่านี้เข้ารหัสกรดอะมิโนมาตรฐาน 20 ชนิด ทำให้กรดอะมิโนส่วนใหญ่มีโคดอนที่เป็นไปได้มากกว่าหนึ่งแบบ นอกจากนี้ยังมีโคดอน "หยุด" หรือ "ไร้ความหมาย" สามแบบที่บ่งบอกถึงจุดสิ้นสุดของบริเวณการเข้ารหัส ได้แก่ โคดอน TAG, TAA และ TGA (UAG, UAA และ UGA บน mRNA)
การจำลองแบบ

การแบ่งเซลล์เป็นสิ่งจำเป็นสำหรับการเจริญเติบโตของสิ่งมีชีวิต แต่เมื่อเซลล์แบ่งตัว เซลล์จะต้องจำลองดีเอ็นเอในจีโนมเพื่อให้เซลล์ลูกทั้งสองมีข้อมูลทางพันธุกรรมเหมือนกับเซลล์แม่ โครงสร้างแบบสองสายของดีเอ็นเอเป็นกลไกที่ง่ายสำหรับการจำลองดีเอ็นเอโดยที่สายทั้งสองจะแยกออกจากกัน จากนั้นลำดับดีเอ็นเอที่เสริมกัน ของแต่ละสายจะถูกสร้างขึ้นใหม่โดย เอนไซม์ที่เรียกว่าดีเอ็นเอพอลิเมอเรสเอนไซม์นี้สร้างสายที่เสริมกันโดยการหาเบสที่ถูกต้องผ่านการจับคู่เบสที่เสริมกันและเชื่อมต่อเข้ากับสายเดิม เนื่องจากดีเอ็นเอพอลิเมอเรสสามารถขยายสายดีเอ็นเอได้เฉพาะในทิศทาง 5′ ถึง 3′ เท่านั้น จึงใช้กลไกที่แตกต่างกันในการคัดลอกสายแอนติพาราเลลของเกลียวคู่[ 98 ]ด้วยวิธีนี้ เบสบนสายเดิมจะกำหนดว่าเบสใดจะปรากฏบนสายใหม่ และเซลล์ก็จะได้สำเนาดีเอ็นเอที่สมบูรณ์แบบ
กรดนิวคลีอิกนอกเซลล์
ดีเอ็นเอภายนอกเซลล์ (eDNA) ที่ไม่มีเปลือกหุ้ม ซึ่งส่วนใหญ่ถูกปล่อยออกมาจากการตายของเซลล์นั้น พบได้ทั่วไปในสิ่งแวดล้อม ความเข้มข้นในดินอาจสูงถึง 2 μg/L และความเข้มข้นในสภาพแวดล้อมทางน้ำตามธรรมชาติอาจสูงถึง 88 μg/L [ 99 ]มีการเสนอหน้าที่ที่เป็นไปได้ต่างๆ ของ eDNA ได้แก่ อาจเกี่ยวข้องกับการถ่ายทอดยีนในแนวนอน [ 100 ] อาจให้สารอาหาร[ 101 ]และอาจทำหน้าที่เป็นบัฟเฟอร์เพื่อดึงดูดหรือปรับสมดุลไอออนหรือยาปฏิชีวนะ[ 102 ]ดีเอ็นเอภายนอกเซลล์ทำหน้าที่เป็นส่วนประกอบของเมทริกซ์ภายนอกเซลล์ที่มีฟังก์ชันการทำงานในไบโอฟิล์มของแบคทีเรียหลายชนิด อาจทำหน้าที่เป็นปัจจัยการรับรู้เพื่อควบคุมการยึดเกาะและการกระจายตัวของเซลล์ชนิดเฉพาะในไบโอฟิล์ม[ 103 ]อาจมีส่วนช่วยในการสร้างไบโอฟิล์ม[ 104 ]และอาจมีส่วนช่วยในความแข็งแรงทางกายภาพและความต้านทานต่อความเครียดทางชีวภาพของไบโอฟิล์ม[ 105 ]
ดีเอ็นเอของทารกในครรภ์ที่ปราศจากเซลล์จะพบได้ในเลือดของมารดา และสามารถจัดลำดับเพื่อกำหนดข้อมูลจำนวนมากเกี่ยวกับทารกในครรภ์ที่กำลังพัฒนา[ 106 ]
ภายใต้ชื่อDNA สิ่งแวดล้อม eDNA ได้มีการนำมาใช้เพิ่มมากขึ้นในวิทยาศาสตร์ธรรมชาติในฐานะเครื่องมือสำรวจทางนิเวศวิทยาการติดตามการเคลื่อนไหวและการปรากฏตัวของสายพันธุ์ในน้ำ อากาศ หรือบนบก และการประเมินความหลากหลายทางชีวภาพของพื้นที่[ 107 ] [ 108 ]
ปฏิสัมพันธ์กับโปรตีน
หน้าที่ทั้งหมดของ DNA ขึ้นอยู่กับการโต้ตอบกับโปรตีนการโต้ตอบระหว่างโปรตีน กับ DNA อาจเกิดขึ้นแบบไม่จำเพาะเจาะจง หรือโปรตีนอาจจับกับลำดับ DNA เพียงลำดับเดียวอย่างจำเพาะเจาะจง นอกจากนี้ เอนไซม์ยังสามารถจับกับ DNA ได้ และในบรรดาเอนไซม์เหล่านี้ เอนไซม์พอลิเมอเรสที่คัดลอกลำดับเบสของ DNA ในกระบวนการถอดรหัสและการจำลองแบบ DNA มีความสำคัญเป็นพิเศษ
โปรตีนที่จับกับดีเอ็นเอ

โปรตีนโครงสร้างที่จับกับ DNA เป็นตัวอย่างที่เข้าใจกันดีของการโต้ตอบระหว่าง DNA กับโปรตีนที่ไม่จำเพาะเจาะจง ภายในโครโมโซม DNA จะถูกยึดไว้ในคอมเพล็กซ์กับโปรตีนโครงสร้าง โปรตีนเหล่านี้จัดระเบียบ DNA ให้เป็นโครงสร้างที่กะทัดรัดเรียกว่าโครมาตินในยูคาริโอต โครงสร้างนี้เกี่ยวข้องกับการจับกันของ DNA กับคอมเพล็กซ์ของโปรตีนพื้นฐานขนาดเล็กที่เรียกว่าฮิสโตนในขณะที่ในโปรคาริโอตมีโปรตีนหลายประเภทเข้ามาเกี่ยวข้อง[ 109 ] [ 110 ]ฮิสโตนก่อตัวเป็นคอมเพล็กซ์รูปทรงแผ่นดิสก์ที่เรียกว่านิวคลีโอโซมซึ่งประกอบด้วย DNA สองสายคู่ที่พันรอบพื้นผิวอย่างสมบูรณ์ การโต้ตอบที่ไม่จำเพาะเจาะจงเหล่านี้เกิดขึ้นผ่านสารตกค้างพื้นฐานในฮิสโตน ทำให้เกิดพันธะไอออนิกกับโครงสร้างน้ำตาลฟอสเฟตที่เป็นกรดของ DNA และจึงไม่ขึ้นอยู่กับลำดับเบสเป็นส่วนใหญ่[ 111 ]การดัดแปลงทางเคมีของสารตกค้างกรดอะมิโนพื้นฐานเหล่านี้ ได้แก่เมทิลเลชัน ฟอสโฟริเลชันและอะเซทิเลชัน[ 112 ]การเปลี่ยนแปลงทางเคมีเหล่านี้เปลี่ยนแปลงความแข็งแรงของปฏิสัมพันธ์ระหว่าง DNA และฮิสโตน ทำให้ DNA เข้าถึงปัจจัยการถอดรหัส ได้มากขึ้นหรือน้อยลง และเปลี่ยนแปลงอัตราการถอดรหัส[ 113 ]โปรตีนที่จับกับ DNA แบบไม่จำเพาะอื่นๆ ในโครมาติน ได้แก่ โปรตีนกลุ่มที่มีการเคลื่อนที่สูง ซึ่งจับกับ DNA ที่โค้งงอหรือบิดเบี้ยว[ 114 ]โปรตีนเหล่านี้มีความสำคัญในการโค้งงออาร์เรย์ของนิวคลีโอโซมและจัดเรียงให้เป็นโครงสร้างขนาดใหญ่ที่ประกอบเป็นโครโมโซม[ 115 ]
โปรตีนที่จับกับ DNA กลุ่มหนึ่งที่แตกต่างออกไปคือโปรตีนที่จับกับ DNA สายเดี่ยวโดยเฉพาะ ในมนุษย์โปรตีนการจำลองแบบ Aเป็นสมาชิกที่เข้าใจได้ดีที่สุดในตระกูลนี้และถูกใช้ในกระบวนการที่เกลียวคู่ถูกแยกออกจากกัน รวมถึงการจำลองแบบ DNA การรวมตัวใหม่ และการซ่อมแซม DNA [ 116 ]โปรตีนที่จับเหล่านี้ดูเหมือนจะทำให้ DNA สายเดี่ยวมีเสถียรภาพและป้องกันไม่ให้เกิดก้านห่วงหรือถูกย่อยสลายโดยนิวคลีเอส
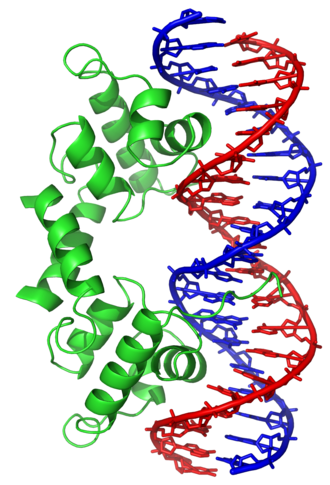
ในทางตรงกันข้าม โปรตีนอื่นๆ ได้วิวัฒนาการเพื่อจับกับลำดับ DNA ที่เฉพาะเจาะจง โปรตีนที่ได้รับการศึกษาอย่างเข้มข้นที่สุดคือปัจจัยการถอดรหัส ต่างๆ ซึ่งเป็นโปรตีนที่ควบคุมการถอดรหัส ปัจจัยการถอดรหัสแต่ละตัวจะจับกับลำดับ DNA ชุดใดชุดหนึ่งโดยเฉพาะ และกระตุ้นหรือยับยั้งการถอดรหัสของยีนที่มีลำดับเหล่านี้อยู่ใกล้กับโปรโมเตอร์ ปัจจัยการถอดรหัสทำเช่นนี้ได้สองวิธี วิธีแรก พวกมันสามารถจับกับ RNA polymerase ที่รับผิดชอบในการถอดรหัส ไม่ว่าจะโดยตรงหรือผ่านโปรตีนตัวกลางอื่นๆ ซึ่งจะทำให้ polymerase อยู่ที่โปรโมเตอร์และอนุญาตให้เริ่มการถอดรหัสได้[ 118 ]หรืออีกทางหนึ่ง ปัจจัยการถอดรหัสสามารถจับกับเอนไซม์ที่ปรับเปลี่ยนฮิสโตนที่โปรโมเตอร์ ซึ่งจะเปลี่ยนการเข้าถึงแม่แบบ DNA ให้กับ polymerase [ 119 ]
เนื่องจากเป้าหมาย DNA เหล่านี้สามารถเกิดขึ้นได้ทั่วทั้งจีโนมของสิ่งมีชีวิต การเปลี่ยนแปลงในกิจกรรมของปัจจัยการถอดรหัสประเภทหนึ่งสามารถส่งผลกระทบต่อยีนหลายพันยีนได้[ 120 ]ด้วยเหตุนี้ โปรตีนเหล่านี้จึงมักเป็นเป้าหมายของ กระบวนการ ส่งสัญญาณที่ควบคุมการตอบสนองต่อการเปลี่ยนแปลงของสิ่งแวดล้อมหรือการแบ่งแยกและการพัฒนาของเซลล์ ความจำเพาะของการโต้ตอบของปัจจัยการถอดรหัสเหล่านี้กับ DNA มาจากการที่โปรตีนสร้างการสัมผัสหลายครั้งกับขอบของเบส DNA ทำให้พวกมันสามารถ "อ่าน" ลำดับ DNA ได้ การโต้ตอบกับเบสส่วนใหญ่เกิดขึ้นในร่องหลัก ซึ่งเป็นบริเวณที่เบสสามารถเข้าถึงได้มากที่สุด[ 25 ]
เอนไซม์ดัดแปลงดีเอ็นเอ
นิวคลีเอสและไลเกส

นิวคลีเอสเป็นเอนไซม์ที่ตัดสาย DNA โดยการเร่งปฏิกิริยา ไฮโดรไลซิส ของพันธะฟอสโฟไดเอสเทอร์ นิวคลีเอสที่ไฮโดรไลซิสนิวคลีโอไทด์จากปลายสาย DNA เรียกว่าเอ็กโซนิว คลีเอส ในขณะที่เอนโดนิ วคลี เอสจะตัดภายในสาย นิวคลีเอสที่ใช้บ่อยที่สุดในชีววิทยาโมเลกุลคือ เอนโดนิวคลีเอ สแบบจำกัด ซึ่งตัด DNA ที่ลำดับเฉพาะ ตัวอย่างเช่น เอนไซม์ EcoRV ที่แสดงทางด้านซ้ายจะจดจำลำดับ 6 เบส 5′-GATATC-3′ และทำการตัดที่เส้นแนวนอน ในธรรมชาติ เอนไซม์เหล่านี้ปกป้องแบคทีเรียจาก การติดเชื้อไวรัส แบคทีริโอเฟจโดยการย่อย DNA ของไวรัสแบคทีริโอเฟจเมื่อมันเข้าสู่เซลล์แบคทีเรีย ซึ่งทำหน้าที่เป็นส่วนหนึ่งของระบบการดัดแปลงแบบจำกัด[ 122 ]ในเทคโนโลยี นิวคลีเอสที่จำเพาะต่อลำดับเหล่านี้ถูกใช้ในการโคลนนิ่งโมเลกุลและ การ ตรวจ สอบลายนิ้ว มือ DNA
เอนไซม์ที่เรียกว่าDNA ligaseสามารถเชื่อมต่อสาย DNA ที่ถูกตัดหรือขาดได้[ 123 ] Ligase มีความสำคัญอย่างยิ่งใน การจำลองแบบ DNA สายล้าหลังเนื่องจากพวกมันเชื่อมต่อส่วนสั้นๆ ของ DNA ที่ผลิตขึ้นที่จุดแยกการจำลองแบบเข้ากับสำเนาที่สมบูรณ์ของแม่แบบ DNA นอกจากนี้ยังใช้ในการซ่อมแซม DNAและการรวมตัวทางพันธุกรรมอีก ด้วย [ 123 ]
โทโปไอโซเมอเรสและเฮลิเคส
โทโปไอโซเมอเรสเป็นเอนไซม์ที่มีทั้งกิจกรรมนิวคลีเอสและไลเกส โปรตีนเหล่านี้เปลี่ยนแปลงปริมาณการขดตัวของ DNA เอนไซม์บางชนิดทำงานโดยการตัดเกลียว DNA และอนุญาตให้ส่วนหนึ่งหมุนได้ ซึ่งจะช่วยลดระดับการขดตัว จากนั้นเอนไซม์จะเชื่อมรอยแตกของ DNA [ 44 ]เอนไซม์ประเภทอื่น ๆ สามารถตัดเกลียว DNA หนึ่งเกลียวแล้วส่งสาย DNA ที่สองผ่านรอยแตกนี้ก่อนที่จะเชื่อมเกลียวเข้าด้วยกันอีกครั้ง[ 124 ]โทโปไอโซเมอเรสจำเป็นสำหรับกระบวนการหลายอย่างที่เกี่ยวข้องกับ DNA เช่น การจำลองแบบ DNA และการถอดรหัส[ 45 ]
เฮลิเคสเป็นโปรตีนประเภทมอเตอร์โมเลกุลพวกมันใช้พลังงานเคมีในนิวคลีโอไซด์ไตรฟอสเฟตซึ่งส่วนใหญ่เป็นอะดีโนซีนไตรฟอสเฟต (ATP) เพื่อทำลายพันธะไฮโดรเจนระหว่างเบสและคลายเกลียวคู่ของ DNA ให้เป็นสายเดี่ยว[ 125 ]เอนไซม์เหล่านี้จำเป็นสำหรับกระบวนการส่วนใหญ่ที่เอนไซม์ต้องเข้าถึงเบสของ DNA
โพลีเมอเรส
เอนไซม์ พอลิเมอเรสเป็นเอนไซม์ที่สังเคราะห์สายพอลินิวคลีโอไทด์จากนิวคลีโอไซด์ ไตรฟอสเฟต ลำดับของผลิตภัณฑ์ถูกสร้างขึ้นโดยอิงจากสายพอลินิวคลีโอไทด์ที่มีอยู่ ซึ่งเรียก ว่า แม่แบบเอนไซม์เหล่านี้ทำงานโดยการเพิ่มนิวคลีโอไท ด์ซ้ำๆ เข้าไปที่ หมู่ไฮดรอกซิล 3′ ที่ปลายของสายพอลินิวคลีโอไทด์ที่กำลังเติบโต ดังนั้น พอลิเมอเรสทั้งหมดจึงทำงานในทิศทาง 5′ ถึง 3′ [ 126 ]ในบริเวณที่ทำงานของเอนไซม์เหล่านี้ นิวคลีโอไซด์ไตรฟอสเฟตที่เข้ามาจะจับคู่กับแม่แบบ ซึ่งทำให้พอลิเมอเรสสามารถสังเคราะห์สายเสริมของแม่แบบได้อย่างแม่นยำ พอลิเมอเรสถูกจัดประเภทตามชนิดของแม่แบบที่ใช้
ในการจำลองดีเอ็นเอ ดีเอ็นเอพอลิเมอเรส ที่ขึ้นอยู่กับดีเอ็นเอ จะสร้างสำเนาของสายพอลินิวคลีโอไทด์ของดีเอ็นเอ เพื่อรักษาข้อมูลทางชีวภาพ จำเป็นอย่างยิ่งที่ลำดับของเบสในแต่ละสำเนาจะต้องเสริมกันอย่างแม่นยำกับลำดับของเบสในสายแม่แบบ ดีเอ็นเอพอลิเมอเรสหลายตัวมี กิจกรรม การตรวจสอบความถูกต้อง ในที่นี้ พอลิเมอเรสจะตรวจจับข้อผิดพลาดที่เกิดขึ้นเป็นครั้งคราวในปฏิกิริยาการสังเคราะห์โดยการขาดการจับคู่เบสระหว่างนิวคลีโอไทด์ที่ไม่ตรงกัน หากตรวจพบความไม่ตรงกัน กิจกรรม เอ็กโซนิวคลีเอส 3′ ถึง 5′ จะถูกกระตุ้นและเบสที่ไม่ถูกต้องจะถูกกำจัดออกไป[ 127 ]ในสิ่งมีชีวิตส่วนใหญ่ ดีเอ็นเอพอลิเมอเรสทำงานในคอมเพล็กซ์ขนาดใหญ่ที่เรียกว่ารีพลิโซมซึ่งประกอบด้วยหน่วยย่อยเสริมหลายหน่วย เช่นดีเอ็นเอแคลมป์หรือเฮลิเคส[ 128 ]
เอนไซม์ DNA polymerase ที่ขึ้นอยู่กับ RNA เป็นเอนไซม์ polymerase ชนิดพิเศษที่คัดลอกลำดับของสาย RNA ไปเป็น DNA ซึ่งรวมถึงreverse transcriptaseซึ่งเป็น เอนไซม์ ของไวรัสที่เกี่ยวข้องกับการติดเชื้อของเซลล์โดยretrovirusและtelomeraseซึ่งจำเป็นสำหรับการจำลองแบบของ telomeres [ 63 ] [ 129 ]ตัวอย่างเช่น HIV reverse transcriptase เป็นเอนไซม์สำหรับการจำลองแบบของไวรัสเอดส์[ 129 ] Telomerase เป็นเอนไซม์ polymerase ที่ผิดปกติเนื่องจากมีแม่แบบ RNA ของตัวเองเป็นส่วนหนึ่งของโครงสร้าง มันสังเคราะห์telomeresที่ปลายโครโมโซม Telomeres ป้องกันการรวมตัวของปลายโครโมโซมที่อยู่ติดกันและปกป้องปลายโครโมโซมจากความเสียหาย[ 64 ]
การถอดรหัสจะดำเนินการโดยเอนไซม์RNA polymerase ที่ขึ้นอยู่กับ DNA ซึ่งคัดลอกลำดับของสาย DNA ไปเป็น RNA ในการเริ่มต้นการถอดรหัสยีน เอนไซม์ RNA polymerase จะจับกับลำดับของ DNA ที่เรียกว่า promoter และแยกสาย DNA ออกจากกัน จากนั้นจะคัดลอกลำดับยีนไปเป็น สารถอดรหัส messenger RNAจนกระทั่งถึงบริเวณของ DNA ที่เรียกว่าterminatorซึ่งจะหยุดและแยกตัวออกจาก DNA เช่นเดียวกับ DNA polymerase ที่ขึ้นอยู่กับ DNA ของมนุษย์RNA polymerase IIซึ่งเป็นเอนไซม์ที่ถอดรหัสยีนส่วนใหญ่ในจีโนมของมนุษย์ ทำงานเป็นส่วนหนึ่งของโปรตีนเชิงซ้อน ขนาดใหญ่ ที่มีหน่วยย่อยควบคุมและหน่วยย่อยเสริมหลายหน่วย[ 130 ]
การรวมตัวทางพันธุกรรม
 |
 |

โดยปกติแล้วเกลียว DNA จะไม่โต้ตอบกับส่วนอื่นๆ ของ DNA และในเซลล์ของมนุษย์ โครโมโซมต่างๆ ยังครอบครองพื้นที่แยกกันในนิวเคลียสที่เรียกว่า " อาณาเขตโครโมโซม " [ 132 ]การแยกทางกายภาพของโครโมโซมต่างๆ นี้มีความสำคัญต่อความสามารถของ DNA ในการทำหน้าที่เป็นแหล่งเก็บข้อมูลที่เสถียร เนื่องจากหนึ่งในไม่กี่ครั้งที่โครโมโซมโต้ตอบกันคือการไขว้กันของโครโมโซมซึ่งเกิดขึ้นระหว่างการสืบพันธุ์แบบอาศัยเพศเมื่อ เกิด การรวมตัวทางพันธุกรรม การไขว้กันของโครโมโซมคือเมื่อเกลียว DNA สองเกลียวแตก สลับส่วนหนึ่งแล้วเชื่อมต่อกันใหม่
การรวมตัวใหม่ช่วยให้โครโมโซมสามารถแลกเปลี่ยนข้อมูลทางพันธุกรรมและสร้างการรวมกันของยีนใหม่ ซึ่งจะเพิ่มประสิทธิภาพของการคัดเลือกโดยธรรมชาติและอาจมีความสำคัญต่อวิวัฒนาการอย่างรวดเร็วของโปรตีนใหม่[ 133 ]การรวมตัวทางพันธุกรรมยังสามารถเกี่ยวข้องกับการซ่อมแซม DNA โดยเฉพาะอย่างยิ่งในการตอบสนองของเซลล์ต่อการแตกของสายคู่[ 134 ]
รูปแบบที่พบได้บ่อยที่สุดของการไขว้กันของโครโมโซมคือการรวมตัวกันแบบโฮโมโลกัสซึ่งโครโมโซมทั้งสองที่เกี่ยวข้องมีลำดับที่คล้ายคลึงกันมากการรวมตัวกันแบบไม่โฮโมโลกัสอาจเป็นอันตรายต่อเซลล์ได้ เนื่องจากสามารถทำให้เกิดการย้ายตำแหน่งของโครโมโซม และความผิดปกติทางพันธุกรรม ได้ปฏิกิริยาการรวมตัวกันถูกเร่งปฏิกิริยาโดยเอนไซม์ที่เรียกว่ารีคอมบิเนสเช่นRAD51 [ 135 ]ขั้นตอนแรกในการรวมตัวกันคือการแตกของสายคู่ที่เกิดจากเอนโดนิวคลีเอสหรือความเสียหายต่อ DNA [ 136 ]จากนั้นขั้นตอนต่างๆ ที่เร่งปฏิกิริยาบางส่วนโดยรีคอมบิเนสจะนำไปสู่การเชื่อมต่อของเกลียวทั้งสองโดยอย่างน้อยหนึ่งฮอลลิเดย์จังก์ชันซึ่งส่วนของสายเดี่ยวในแต่ละเกลียวจะเชื่อมต่อกับสายเสริมในเกลียวอื่น จุดเชื่อมต่อฮอลลิเดย์เป็นโครงสร้างจุดเชื่อมต่อแบบทรงสี่หน้าซึ่งสามารถเคลื่อนที่ไปตามคู่โครโมโซม โดยสลับสายหนึ่งกับอีกสายหนึ่ง ปฏิกิริยาการรวมตัวใหม่จะหยุดลงโดยการตัดจุดเชื่อมต่อและการเชื่อมต่อใหม่ของ DNA ที่ปล่อยออกมา[ 137 ]เฉพาะสายที่มีขั้วเหมือนกันเท่านั้นที่จะแลกเปลี่ยน DNA ในระหว่างการรวมตัวใหม่ มีการตัดสองประเภท ได้แก่ การตัดแบบตะวันออก-ตะวันตก และการตัดแบบเหนือ-ใต้ การตัดแบบเหนือ-ใต้จะตัดสาย DNA ทั้งสองสาย ในขณะที่การตัดแบบตะวันออก-ตะวันตกจะมีสาย DNA หนึ่งสายที่ยังคงสภาพสมบูรณ์ การก่อตัวของจุดเชื่อมต่อฮอลลิเดย์ในระหว่างการรวมตัวใหม่ทำให้เกิดความหลากหลายทางพันธุกรรม การแลกเปลี่ยนยีนบนโครโมโซม และการแสดงออกของจีโนมไวรัสชนิดป่าได้
วิวัฒนาการ
ดีเอ็นเอมีข้อมูลทางพันธุกรรมที่ทำให้สิ่งมีชีวิตทุกรูปแบบสามารถทำงาน เติบโต และสืบพันธุ์ได้ อย่างไรก็ตาม ยังไม่ชัดเจนว่าในประวัติศาสตร์ 4 พันล้านปีของสิ่งมีชีวิตดีเอ็นเอทำหน้าที่นี้มานานแค่ไหน เนื่องจากมีการเสนอว่าสิ่งมีชีวิตในยุคแรกเริ่มอาจใช้อาร์เอ็นเอเป็นสารพันธุกรรม[ 138 ] [ 139 ]อาร์เอ็นเออาจทำหน้าที่เป็นส่วนสำคัญของกระบวนการเผาผลาญของเซลล์ ในยุคแรก เนื่องจากสามารถส่งต่อข้อมูลทางพันธุกรรมและดำเนินการเร่งปฏิกิริยาในฐานะส่วนหนึ่งของไรโบไซม์ได้[ 140 ]โลกอาร์เอ็นเอโบราณนี้ซึ่งกรดนิวคลีอิกจะถูกใช้ทั้งในการเร่งปฏิกิริยาและพันธุกรรม อาจมีอิทธิพลต่อวิวัฒนาการของรหัสพันธุกรรมในปัจจุบันซึ่งมีพื้นฐานมาจากเบสนิวคลีโอไทด์สี่ชนิด สิ่งนี้จะเกิดขึ้นเนื่องจากจำนวนเบสที่แตกต่างกันในสิ่งมีชีวิตดังกล่าวเป็นการแลกเปลี่ยนระหว่างจำนวนเบสน้อยที่เพิ่มความแม่นยำในการจำลองแบบและจำนวนเบสมากที่เพิ่มประสิทธิภาพการเร่งปฏิกิริยาของไรโบไซม์[ 141 ]อย่างไรก็ตาม ไม่มีหลักฐานโดยตรงเกี่ยวกับระบบพันธุกรรมโบราณ เนื่องจากการกู้คืน DNA จากฟอสซิลส่วนใหญ่เป็นไปไม่ได้ เพราะ DNA อยู่รอดในสิ่งแวดล้อมได้น้อยกว่าหนึ่งล้านปี และจะค่อยๆ สลายตัวเป็นชิ้นส่วนสั้นๆ ในสารละลาย[ 142 ]มีการกล่าวอ้างถึง DNA ที่เก่ากว่านั้น โดยเฉพาะอย่างยิ่งรายงานการแยกแบคทีเรียที่มีชีวิตจากผลึกเกลือที่มีอายุ 250 ล้านปี[ 143 ]แต่การกล่าวอ้างเหล่านี้เป็นที่ถกเถียงกัน[ 144 ] [ 145 ]
หน่วยพื้นฐานของ DNA ( อะดีนีนกัวนีนและโมเลกุลอินทรีย์ ที่เกี่ยวข้อง ) อาจเกิดขึ้นนอกโลกในอวกาศ[ 146 ] [ 147 ] [ 148 ] สารประกอบอินทรีย์ DNA และRNA ที่ซับซ้อน ของสิ่งมีชีวิตรวมถึงยูราซิลไซโตซีนและไทมีนก็เกิดขึ้นในห้องทดลองภายใต้สภาวะที่เลียนแบบสภาวะที่พบในอวกาศ โดยใช้สารเคมีเริ่มต้น เช่นไพริมิดีนที่พบในอุกกาบาตไพริมิดีน เช่นเดียวกับ โพลีไซคลิกอะโรมาติกไฮโดรคาร์บอน (PAHs) ซึ่งเป็นสารเคมีที่มีคาร์บอนมากที่สุดในจักรวาลอาจเกิดขึ้นในดาวยักษ์แดงหรือในฝุ่นและก๊าซ ในอวกาศระหว่างดาว [ 149 ]
ดีเอ็นเอโบราณได้รับการกู้คืนจากสิ่งมีชีวิตโบราณในช่วงเวลาที่สามารถสังเกตวิวัฒนาการของจีโนมได้โดยตรง รวมถึงจากสิ่งมีชีวิตที่สูญพันธุ์ไปแล้วที่มีอายุหลายล้านปี เช่นแมมมอธขนปุย[ 150 ] [ 151 ]
การใช้งานในด้านเทคโนโลยี
วิศวกรรมพันธุกรรม
มีการพัฒนาวิธีการต่างๆ เพื่อทำให้ DNA บริสุทธิ์จากสิ่งมีชีวิต เช่นการสกัดด้วยฟีนอล-คลอโรฟอร์มและเพื่อจัดการกับ DNA ในห้องปฏิบัติการ เช่นการย่อยด้วยเอนไซม์จำกัดและปฏิกิริยาลูกโซ่พอลิเมอเรส ชีววิทยาและชีวเคมีสมัยใหม่ใช้เทคนิคเหล่านี้อย่างเข้มข้นในเทคโนโลยี DNA รีคอมบิแนนท์DNA รีคอมบิแนนท์คือลำดับ DNA ที่มนุษย์สร้างขึ้นซึ่งประกอบขึ้นจากลำดับ DNA อื่นๆ สามารถถ่ายโอนเข้าไปในสิ่งมีชีวิตในรูปของพลาสมิดหรือในรูปแบบที่เหมาะสมโดยใช้เวกเตอร์ไวรัส [ 152 ]สิ่ง มีชีวิต ดัดแปลงพันธุกรรมที่ผลิตขึ้นสามารถนำไปใช้ในการผลิตผลิตภัณฑ์ต่างๆ เช่นโปรตีน รีคอมบิแนนท์ ซึ่งใช้ใน การ วิจัยทางการแพทย์ [ 153 ] หรือปลูกในทางการเกษตร[ 154 ] [ 155 ]
การวิเคราะห์ดีเอ็นเอ
นักวิทยาศาสตร์นิติเวชสามารถใช้ DNA ในเลือดน้ำอสุจิ ผิวหนัง น้ำลาย หรือเส้นผมที่พบในที่เกิดเหตุเพื่อระบุ DNA ที่ตรงกันของบุคคล เช่น ผู้กระทำความผิด[ 156 ]กระบวนการนี้เรียกว่าการสร้างโปรไฟล์ DNAหรือที่เรียกว่าการตรวจสอบลายนิ้วมือ DNAในการสร้างโปรไฟล์ DNA จะมีการเปรียบเทียบความยาวของส่วนที่แปรผันของ DNA ที่ซ้ำกัน เช่นshort tandem repeatsและminisatellitesระหว่างบุคคลต่างๆ วิธีนี้มักเป็นเทคนิคที่เชื่อถือได้มากในการระบุ DNA ที่ตรงกัน[ 157 ]อย่างไรก็ตาม การระบุตัวตนอาจซับซ้อนหากที่เกิดเหตุปนเปื้อนด้วย DNA จากหลายคน [ 158 ]การสร้างโปรไฟล์ DNA ได้รับการพัฒนาขึ้นในปี 1984 โดยนักพันธุศาสตร์ชาวอังกฤษ เซอร์อเล็ก เจฟฟรีย์ส [ 159 ] และถูกนำมาใช้ในนิติเวชศาสตร์เป็นครั้งแรกเพื่อตัดสินลงโทษโคลิน พิทช์ฟอร์ก ในคดีฆาตกรรมเอ็นเดอร์บี ในปี 1988 [ 160 ]
การพัฒนาด้านนิติวิทยาศาสตร์และความสามารถในการตรวจหาดีเอ็นเอจากตัวอย่างเลือด ผิวหนัง น้ำลาย หรือเส้นผมในปริมาณน้อยมาก ทำให้มีการตรวจสอบคดีหลายคดีใหม่ หลักฐานที่ก่อนหน้านี้เป็นไปไม่ได้ทางวิทยาศาสตร์สามารถค้นพบได้ในขณะที่ทำการตรวจสอบครั้งแรก เมื่อรวมกับการยกเลิก กฎหมาย ห้ามดำเนินคดีซ้ำซ้อนในบางพื้นที่ ทำให้สามารถเปิดคดีใหม่ได้ในกรณีที่การพิจารณาคดีครั้งก่อนไม่สามารถหาหลักฐานเพียงพอที่จะโน้มน้าวคณะลูกขุนได้ ผู้ที่ถูกตั้งข้อหาในคดีอาญาร้ายแรงอาจต้องให้ตัวอย่างดีเอ็นเอเพื่อการตรวจสอบ ข้อแก้ตัวที่ชัดเจนที่สุดสำหรับการตรวจสอบดีเอ็นเอทางนิติวิทยาศาสตร์คือการอ้างว่าเกิดการปนเปื้อนของหลักฐาน สิ่งนี้ส่งผลให้มีขั้นตอนการจัดการที่เข้มงวดและละเอียดถี่ถ้วนมากขึ้นสำหรับคดีอาญาร้ายแรงใหม่ๆ
การตรวจวิเคราะห์ดีเอ็นเอยังถูกนำมาใช้อย่างประสบความสำเร็จในการระบุตัวตนเหยื่อจากเหตุการณ์ที่มีผู้เสียชีวิตจำนวนมาก[ 161 ]ศพหรือชิ้นส่วนร่างกายในอุบัติเหตุร้ายแรง และเหยื่อแต่ละรายในหลุมฝังศพหมู่ในสงคราม โดยการจับคู่กับสมาชิกในครอบครัว
การตรวจวิเคราะห์ดีเอ็นเอยังใช้ในการตรวจพิสูจน์ความเป็นพ่อโดยใช้ดีเอ็นเอเพื่อตรวจสอบว่าบุคคลใดเป็นพ่อแม่หรือปู่ย่าตายายทางชีววิทยาของเด็กหรือไม่ โดยความน่าจะเป็นของความเป็นพ่อแม่โดยทั่วไปอยู่ที่ 99.99% เมื่อผู้ที่ถูกกล่าวหาว่าเป็นพ่อแม่มีความสัมพันธ์ทางชีววิทยากับเด็ก โดยปกติแล้วการจัดลำดับดีเอ็นเอจะดำเนินการหลังคลอด แต่มีวิธีการใหม่ๆ ในการตรวจพิสูจน์ความเป็นพ่อในขณะที่แม่ยังตั้งครรภ์อยู่[ 162 ]
เอนไซม์ดีเอ็นเอหรือดีเอ็นเอเร่งปฏิกิริยา
ดีออกซี ไรโบไซม์ หรือที่เรียกว่า ดีเอ็นเอไซม์ หรือดีเอ็นเอเร่งปฏิกิริยา ถูกค้นพบครั้งแรกในปี 1994 [ 163 ] ส่วนใหญ่เป็นลำดับดีเอ็นเอสายเดี่ยวที่แยกได้จากกลุ่มลำดับดีเอ็นเอแบบสุ่มจำนวนมากผ่านวิธีการผสมผสานที่เรียกว่า การคัดเลือก ในหลอดทดลองหรือวิวัฒนาการอย่างเป็นระบบของลิแกนด์โดยการเพิ่มความเข้มข้นแบบทวีคูณ (SELEX) ดีเอ็นเอไซม์เร่งปฏิกิริยาเคมีได้หลากหลายชนิด รวมถึงการตัด RNA-DNA การเชื่อมต่อ RNA-DNA การฟอสโฟรีเลชัน-ดีฟอสโฟรีเลชันของกรดอะมิโน การสร้างพันธะคาร์บอน-คาร์บอน เป็นต้น ดีเอ็นเอไซม์สามารถเพิ่มอัตราการเร่งปฏิกิริยาของปฏิกิริยาเคมีได้มากถึง 100,000,000,000 เท่าเมื่อเทียบกับปฏิกิริยาที่ไม่มีตัวเร่งปฏิกิริยา[ 164 ]ดีเอ็นเอไซม์ประเภทที่ได้รับการศึกษาอย่างกว้างขวางที่สุดคือประเภทที่ตัด RNA ซึ่งถูกนำมาใช้ในการตรวจจับไอออนโลหะต่างๆ และออกแบบตัวแทนบำบัดโรค มีการรายงาน DNAzyme ที่เฉพาะเจาะจงกับโลหะหลายชนิด ได้แก่ GR-5 DNAzyme (เฉพาะตะกั่ว) [ 163 ] CA1-3 DNAzymes (เฉพาะทองแดง) [ 165 ] 39E DNAzyme (เฉพาะยูเรนิล) และ NaA43 DNAzyme (เฉพาะโซเดียม) [ 166 ] NaA43 DNAzyme ซึ่งมีรายงานว่ามีความเลือกจำเพาะต่อโซเดียมมากกว่าไอออนโลหะอื่นๆ มากกว่า 10,000 เท่า ถูกนำมาใช้สร้างเซนเซอร์โซเดียมแบบเรียลไทม์ในเซลล์
ชีวสารสนเทศ
ชีวสารสนเทศศาสตร์เกี่ยวข้องกับการพัฒนาเทคนิคในการจัดเก็บขุดค้นข้อมูลค้นหา และจัดการข้อมูลทางชีววิทยา รวมถึงข้อมูลลำดับกรดนิวคลีอิก DNA ซึ่งนำไปสู่ความก้าวหน้าในการประยุกต์ใช้อย่างกว้างขวางใน วิทยาศาสตร์คอมพิวเตอร์โดยเฉพาะอย่างยิ่งอัลกอริทึมการค้นหาสตริงการเรียนรู้ของเครื่องและทฤษฎีฐานข้อมูล [ 167 ] อัลกอริทึมการค้นหาหรือการจับคู่สตริง ซึ่งค้นหาการเกิดขึ้นของลำดับตัวอักษรภายในลำดับตัวอักษรที่ใหญ่กว่า ได้รับการพัฒนาขึ้นเพื่อค้นหาลำดับนิวคลีโอไทด์ที่เฉพาะเจาะจง[ 168 ]ลำดับ DNA อาจถูกจัดเรียงกับลำดับ DNA อื่นๆ เพื่อระบุลำดับที่เหมือนกันและระบุตำแหน่งการกลายพันธุ์ เฉพาะ ที่ทำให้แตกต่างกัน เทคนิคเหล่านี้ โดยเฉพาะอย่างยิ่งการจัดเรียงลำดับหลายลำดับถูกนำมาใช้ในการศึกษา ความสัมพันธ์ ทางวิวัฒนาการและหน้าที่ของโปรตีน[ 169 ]ชุดข้อมูลที่แสดงถึงลำดับ DNA ของจีโนมทั้งหมด เช่น ชุดข้อมูลที่ผลิตโดยโครงการจีโนมมนุษย์เป็นเรื่องยากที่จะใช้หากไม่มีคำอธิบายประกอบที่ระบุตำแหน่งของยีนและองค์ประกอบควบคุมบนแต่ละโครโมโซม ภูมิภาคของลำดับดีเอ็นเอที่มีรูปแบบลักษณะเฉพาะที่เกี่ยวข้องกับยีนที่เข้ารหัสโปรตีนหรืออาร์เอ็นเอสามารถระบุได้ด้วย อัลกอริทึม การค้นหายีนซึ่งช่วยให้นักวิจัยสามารถทำนายการมีอยู่ของผลิตภัณฑ์ยีน เฉพาะ และหน้าที่ที่เป็นไปได้ในสิ่งมีชีวิตได้ แม้กระทั่งก่อนที่จะมีการแยกออกมาทดลอง[ 170 ]นอกจากนี้ยังสามารถเปรียบเทียบจีโนมทั้งหมดได้ ซึ่งสามารถให้ความกระจ่างเกี่ยวกับประวัติวิวัฒนาการของสิ่งมีชีวิตเฉพาะและอนุญาตให้ตรวจสอบเหตุการณ์วิวัฒนาการที่ซับซ้อนได้
นาโนเทคโนโลยีดีเอ็นเอ

นาโนเทคโนโลยีดีเอ็นเอใช้ คุณสมบัติ การจดจำโมเลกุล ที่เป็นเอกลักษณ์ ของดีเอ็นเอและกรดนิวคลีอิกอื่นๆ เพื่อสร้างคอมเพล็กซ์ดีเอ็นเอแบบแตกแขนงที่ประกอบตัวเองได้ซึ่งมีคุณสมบัติที่เป็นประโยชน์[ 172 ]ดังนั้นดีเอ็นเอจึงถูกใช้เป็นวัสดุโครงสร้างแทนที่จะเป็นตัวนำข้อมูลทางชีวภาพ ซึ่งนำไปสู่การสร้างโครงตาข่ายแบบคาบสองมิติ (ทั้งแบบใช้กระเบื้องและแบบใช้ดีเอ็นเอโอริกามิ ) และโครงสร้างสามมิติในรูปทรงของทรงหลายเหลี่ยม [ 173 ] อุปกรณ์นาโนเชิงกลและการประกอบตัวเองแบบอัลกอริทึมก็ได้รับการสาธิตแล้วเช่นกัน[ 174 ]และโครงสร้างดีเอ็นเอเหล่านี้ถูกใช้เป็นแม่แบบในการจัดเรียงโมเลกุลอื่นๆ เช่นอนุภาคนาโนทองคำและโปรตีนสเตรปตาไวดีน[ 175 ]ดีเอ็นเอและกรดนิวคลีอิกอื่นๆ เป็นพื้นฐานของแอพ ทาเมอร์ ซึ่งเป็นลิแกนด์โอลิโกนิวคลีโอไทด์สังเคราะห์สำหรับโมเลกุลเป้าหมายเฉพาะที่ใช้ในงานด้านเทคโนโลยีชีวภาพและการแพทย์หลายประเภท[ 176 ]
ประวัติศาสตร์และมานุษยวิทยา
เนื่องจาก DNA สะสมการกลายพันธุ์เมื่อเวลาผ่านไป ซึ่งจะถูกถ่ายทอดทางพันธุกรรม จึงมีข้อมูลทางประวัติศาสตร์ และด้วยการเปรียบเทียบลำดับ DNA นักพันธุศาสตร์สามารถอนุมานประวัติวิวัฒนาการของสิ่งมีชีวิต หรือฟิโลเจเนติกส์ได้[ 177 ]สาขาฟิโลเจเนติกส์นี้เป็นเครื่องมือที่มีประสิทธิภาพในชีววิทยาเชิงวิวัฒนาการหากเปรียบเทียบลำดับ DNA ภายในสปีชีส์นักพันธุศาสตร์ประชากรสามารถเรียนรู้ประวัติของประชากรเฉพาะกลุ่มได้ ซึ่งสามารถนำไปใช้ในการศึกษาต่างๆ ตั้งแต่พันธุศาสตร์เชิงนิเวศวิทยาไปจนถึง มานุษยวิทยา
การจัดเก็บข้อมูล
DNA ในฐานะอุปกรณ์จัดเก็บข้อมูลมีศักยภาพมหาศาล เนื่องจากมีความหนาแน่นในการจัดเก็บข้อมูลสูงกว่าอุปกรณ์อิเล็กทรอนิกส์มาก อย่างไรก็ตาม ต้นทุนสูง เวลาในการอ่านและเขียนที่ช้า ( ความหน่วงของหน่วยความจำ ) และความน่าเชื่อถือ ที่ไม่เพียงพอ ทำให้การใช้งานจริงเป็นไปได้ยาก[ 178 ] [ 179 ]
ประวัติศาสตร์

ดีเอ็นเอถูกแยกออกมาเป็นครั้งแรกโดยแพทย์ชาวสวิสชื่อฟรีดริช มีเชอร์ซึ่งในปี 1869 ได้ค้นพบสารขนาดเล็กในหนองของผ้าพันแผลผ่าตัดที่ถูกทิ้งแล้ว เนื่องจากสารนี้อยู่ในนิวเคลียสของเซลล์ เขาจึงเรียกมันว่า "นิวคลีอิน" [ 180 ] [ 181 ]ในปี 1878 อัลเบรชต์ คอสเซล ได้แยกส่วนประกอบที่ไม่ใช่โปรตีนของ "นิวคลีอิน" ซึ่งก็คือกรดนิวคลีอิก และต่อมาได้แยก นิวคลีโอเบสหลักทั้งห้าของมันออกมา[ 182 ] [ 183 ]
ในปี พ.ศ. 2452 โฟบัส เลเวน ได้ระบุหน่วยนิวคลีโอไทด์เบส น้ำตาล และฟอสเฟตของ RNA (ซึ่งในขณะนั้นเรียกว่า "กรดนิวคลีอิกของยีสต์") [ 184 ] [ 185 ] [ 186 ]ในปี พ.ศ. 2462 เลเวนได้ระบุน้ำตาลดีออกซีไรโบสใน "กรดนิวคลีอิกของต่อมไทมัส" (DNA) [ 187 ]เลเวนเสนอว่า DNA ประกอบด้วยสายของหน่วยนิวคลีโอไทด์สี่หน่วยที่เชื่อมต่อกันผ่านกลุ่มฟอสเฟต (" สมมติฐานเตตระนิวคลีโอไทด์ ") เลเวนคิดว่าสายโซ่นั้นสั้นและเบสจะซ้ำกันในลำดับที่แน่นอน ในปี พ.ศ. 2460 นิโคไล โคลต์ซอฟ เสนอว่าลักษณะทางพันธุกรรมจะถูกถ่ายทอดผ่าน "โมเลกุลทางพันธุกรรมขนาดใหญ่" ที่ประกอบด้วย "สายคู่ที่สะท้อนกันซึ่งจะจำลองแบบกึ่งอนุรักษ์โดยใช้แต่ละสายเป็นแม่แบบ" [ 188 ] [ 189 ]ในปี พ.ศ. 2461 เฟรเดอริค กริฟฟิธค้นพบในการทดลอง ของเขาว่า ลักษณะ ของเชื้อ นิวโมค็อกคัสชนิด "เรียบ" สามารถถ่ายทอดไปยังเชื้อชนิด "หยาบ" ได้โดยการผสมเชื้อชนิด "เรียบ" ที่ตายแล้วกับเชื้อชนิด "หยาบ" ที่ยังมีชีวิตอยู่[ 190 ] [ 191 ]ระบบนี้ให้ข้อเสนอแนะที่ชัดเจนเป็นครั้งแรกว่า DNA เป็นตัวนำข้อมูลทางพันธุกรรม
ในปี พ.ศ. 2476 ขณะศึกษาไข่เม่นทะเล ที่ยังบริสุทธิ์ Jean Brachetเสนอว่า DNA พบอยู่ในนิวเคลียสของเซลล์และRNAพบเฉพาะในไซโตพลาซึมเท่านั้น ในขณะนั้น "กรดนิวคลีอิกของยีสต์" (RNA) เชื่อกันว่าพบเฉพาะในพืช ในขณะที่ "กรดนิวคลีอิกของต่อมไทมัส" (DNA) พบเฉพาะในสัตว์เท่านั้น โดยเชื่อกันว่า DNA เป็นเทตราเมอร์ มีหน้าที่ในการปรับสมดุลค่า pH ของเซลล์[ 192 ] [ 193 ]ในปี พ.ศ. 2480 William Astburyได้สร้างรูปแบบการเลี้ยวเบนของรังสีเอกซ์ครั้งแรกที่แสดงให้เห็นว่า DNA มีโครงสร้างที่สม่ำเสมอ[ 194 ]
ในปี พ.ศ. 2486 Oswald Averyพร้อมด้วยเพื่อนร่วมงานColin MacLeodและMaclyn McCartyได้ระบุว่า DNA เป็นหลักการเปลี่ยนแปลงซึ่งสนับสนุนข้อเสนอแนะของ Griffith ( การทดลอง Avery–MacLeod–McCarty ) [ 195 ] Erwin Chargaffได้พัฒนาและเผยแพร่ข้อสังเกตที่ปัจจุบันรู้จักกันในชื่อกฎของ Chargaffโดยระบุว่าใน DNA จากสิ่งมีชีวิตชนิดใดก็ตาม ปริมาณของกัวนีนควรเท่ากับไซโตซีนและปริมาณของอะดีนีนควรเท่ากับไทมีน[ 196 ] [ 197 ]

ในปี พ.ศ. 2494 อเล็กซ์ ทอดด์และผู้ร่วมงานที่มหาวิทยาลัยเคมบริดจ์ได้กำหนดโครงสร้างหลักของดีเอ็นเอโดยวิธีทางชีวเคมีผ่านการเชื่อมต่ออะตอมคาร์บอนที่ 3 และ 5 ของน้ำตาลกับฟอสเฟตอย่างต่อเนื่อง ซึ่งจะช่วยยืนยันผลงานโครงสร้างด้วยรังสีเอกซ์ของวัตสันและครีกในภายหลัง[ 198 ] ต่อมาทอดด์ได้รับ รางวัลโนเบลสาขาเคมีในปี พ.ศ. 2490 จากการค้นพบนี้และการค้นพบอื่นๆ ที่เกี่ยวข้องกับดีเอ็นเอ[ 199 ]
ปลายปี พ.ศ. 2494 ฟรานซิส คริกเริ่มทำงานร่วมกับเจมส์ วัตสันที่ห้องปฏิบัติการคาเวนดิชภายในมหาวิทยาลัยเคมบริดจ์ บทบาทของ DNA ในการถ่ายทอดทางพันธุกรรมได้รับการยืนยันในปี พ.ศ. 2495 เมื่ออัลเฟรด เฮอร์ชีย์และมาร์ธา เชสในการทดลองเฮอร์ชีย์-เชสแสดงให้เห็นว่า DNA เป็นสารพันธุกรรมของแบคทีเรียเอนเทอโรแบคทีเรียฟาจ T2 [ 200 ]

ในเดือนพฤษภาคม พ.ศ. 2495 เรย์มอนด์ กอสลิงนักศึกษาปริญญาโทที่ทำงานภายใต้การดูแลของโรซาลินด์ แฟ รงคลิน ได้ถ่าย ภาพ การเลี้ยวเบนของรังสี เอกซ์ ซึ่งติดป้ายกำกับว่า " ภาพถ่ายหมายเลข 51 " [ 201 ]ที่ระดับความชุ่มชื้นสูงของดีเอ็นเอ ภาพถ่ายนี้ถูกมอบให้แก่วัตสันและครีกโดยมอริซ วิลกินส์ และมีความสำคัญอย่างยิ่งต่อการที่พวกเขาได้รับโครงสร้างที่ถูกต้องของดีเอ็นเอ แฟรงคลินบอกครีกและวัตสันว่ากระดูกสันหลังต้องอยู่ด้านนอก ก่อนหน้านั้น ไลนัส พอลลิง และวัตสันและครีก มีแบบจำลองที่ผิดพลาดโดยมีโซ่อยู่ด้านในและเบสชี้ออกไปด้านนอก การระบุกลุ่มพื้นที่สำหรับผลึกดีเอ็นเอของแฟรงคลินพิสูจน์ว่าเธอถูกต้อง[ 202 ]ในเดือนกุมภาพันธ์ พ.ศ. 2496 ไลนัส พอลลิงและโรเบิร์ต คอรีย์เสนอแบบจำลองสำหรับกรดนิวคลีอิกที่มีโซ่พันกันสามโซ่ โดยมีฟอสเฟตอยู่ใกล้แกน และเบสอยู่ด้านนอก[ 203 ]วัตสันและคริกสร้างแบบจำลองของพวกเขาเสร็จสมบูรณ์ ซึ่งปัจจุบันได้รับการยอมรับว่าเป็นแบบจำลองที่ถูกต้องแรกของเกลียวคู่ของดีเอ็นเอเมื่อวันที่ 28 กุมภาพันธ์ พ.ศ. 2496 คริกขัดจังหวะช่วงเวลาอาหารกลางวันของลูกค้าที่ผับThe Eagle ในเคมบริดจ์ ประเทศอังกฤษ เพื่อประกาศว่าเขาและวัตสันได้ "ค้นพบความลับของชีวิต" [ 204 ]

วารสารNature ฉบับวันที่ 25 เมษายน พ.ศ. 2496 ได้ตีพิมพ์บทความชุดห้าบทความเกี่ยวกับโครงสร้างเกลียวคู่ของ DNA ตามแนวคิดของ Watson และ Crick พร้อมหลักฐานสนับสนุน[ 205 ]โครงสร้างดังกล่าวได้รับการรายงานในจดหมายชื่อ " โครงสร้างโมเลกุลของกรดนิวคลีอิก โครงสร้างของกรดดีออกซีไรโบสนิวคลีอิก"ซึ่งพวกเขากล่าวว่า "เราสังเกตเห็นว่าการจับคู่เฉพาะที่เราได้ตั้งสมมติฐานไว้นั้นชี้ให้เห็นถึงกลไกการคัดลอกที่เป็นไปได้สำหรับสารพันธุกรรม" [ 8 ]จดหมายฉบับนี้ตามมาด้วยจดหมายจาก Franklin และ Gosling ซึ่งเป็นการตีพิมพ์ข้อมูลการเลี้ยวเบนของรังสีเอกซ์และวิธีการวิเคราะห์ดั้งเดิมของพวกเขาเป็นครั้งแรก[ 48 ] [ 206 ]จากนั้นตามมาด้วยจดหมายจาก Wilkins และเพื่อนร่วมงานอีกสองคน ซึ่งมีการวิเคราะห์ รูปแบบรังสีเอกซ์ของ B-DNA ในร่างกายและสนับสนุนการมีอยู่ของโครงสร้าง Watson และ Crick ในร่างกาย[ 49 ]
ในเดือนเมษายน พ.ศ. 2566 นักวิทยาศาสตร์สรุปโดยอาศัยหลักฐานใหม่ว่า โรซาลินด์ แฟรงคลิน เป็นผู้มีส่วนร่วมและเป็น "ผู้เล่นที่เท่าเทียมกัน" ในกระบวนการค้นพบดีเอ็นเอ ไม่ใช่เป็นอย่างอื่นอย่างที่อาจมีการนำเสนอในภายหลังจากการค้นพบ[ 207 ] [ 208 ] [ 209 ]ในปี พ.ศ. 2505 หลังจากที่แฟรงคลินเสียชีวิต วัตสัน คริก และวิลกินส์ ได้รับรางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ร่วม กัน [ 210 ]รางวัลโนเบลจะมอบให้แก่ผู้ที่ยังมีชีวิตอยู่เท่านั้น การถกเถียงยังคงดำเนินต่อไปว่าใครควรได้รับเครดิตสำหรับการค้นพบ[ 211 ]
ในการนำเสนอที่มีอิทธิพลในปี 1957 คริกได้วางรากฐานหลักการพื้นฐานของชีววิทยาโมเลกุลซึ่งทำนายความสัมพันธ์ระหว่าง DNA, RNA และโปรตีน และได้อธิบาย "สมมติฐานตัวปรับ" [ 212 ]การยืนยันขั้นสุดท้ายของกลไกการจำลองแบบที่บ่งบอกโดยโครงสร้างเกลียวคู่เกิดขึ้นในปี 1958 ผ่านการทดลองของเมเซลสัน-สตาห์ล [ 213 ] งานเพิ่มเติมโดยคริกและผู้ร่วมงานแสดงให้เห็นว่ารหัสพันธุกรรมนั้นขึ้นอยู่กับเบสสามตัวที่ไม่ทับซ้อนกัน เรียกว่าโคดอนทำให้ฮาร์ โกบินด์ โครานาโรเบิร์ต ดับเบิลยู ฮอลลีย์และมาร์แชล วอร์เรน นิเรนเบิร์กสามารถถอดรหัสพันธุกรรมได้[ 214 ]การค้นพบเหล่านี้แสดงถึงการกำเนิดของชีววิทยาโมเลกุล[ 215 ]
ในปี 1986 การวิเคราะห์ดีเอ็นเอถูกนำมาใช้ในการสืบสวนคดีอาญาเป็นครั้งแรก เมื่อตำรวจในสหราชอาณาจักรขอให้Alec Jeffreysจากมหาวิทยาลัยเลสเตอร์พิสูจน์หรือหักล้างความเกี่ยวข้องในคดีเฉพาะของผู้ต้องสงสัยที่อ้างว่าบริสุทธิ์ แม้ว่าผู้ต้องสงสัยจะสารภาพว่าได้ก่อเหตุข่มขืนและฆาตกรรมเมื่อไม่นานมานี้ แต่เขากลับปฏิเสธความเกี่ยวข้องในอาชญากรรมที่คล้ายคลึงกันซึ่งเกิดขึ้นเมื่อสามปีก่อน อย่างไรก็ตาม รายละเอียดของทั้งสองคดีนั้นคล้ายคลึงกันมากจนตำรวจสรุปว่าอาชญากรรมทั้งสองถูกกระทำโดยบุคคลเดียวกัน แต่ข้อกล่าวหาทั้งหมดต่อผู้ต้องสงสัยถูกยกเลิกเมื่อการทดสอบดีเอ็นเอของ Jeffreys พิสูจน์ว่าผู้ต้องสงสัยบริสุทธิ์ ทั้งในคดีฆาตกรรมก่อนหน้านี้และคดีที่เขาสารภาพ นอกจากนี้ การวิเคราะห์ดีเอ็นเอในลักษณะเดียวกันยังนำไปสู่การระบุตัวตนของผู้ต้องสงสัยอีกคนหนึ่ง ( Colin Pitchfork ) ซึ่งในปี 1988 ถูกตัดสินว่ามีความผิดในคดีข่มขืนและฆาตกรรมทั้งสองคดี[ 216 ] [ 217 ]
ดูเพิ่มเติม
- ออโตโซม – โครโมโซมใดๆ ก็ตามที่ไม่ใช่โครโมโซมเพศ
- ผลึกศาสตร์ – การศึกษาทางวิทยาศาสตร์เกี่ยวกับโครงสร้างของผลึก
- วันดีเอ็นเอ – วันหยุดที่เฉลิมฉลองในวันที่ 25 เมษายน
- ไมโครอาร์เรย์ดีเอ็นเอ – ชุดจุดดีเอ็นเอขนาดเล็กที่ยึดติดอยู่บนพื้นผิวแข็ง
- การจัดลำดับดีเอ็นเอ – กระบวนการกำหนดลำดับกรดนิวคลีอิก
- ดีเอ็นเอจากสิ่งแวดล้อม – ดีเอ็นเอที่เก็บตัวอย่างจากสิ่งแวดล้อม ไม่ใช่จากสิ่งมีชีวิตแต่ละตัวโดยตรง
- ความผิดปกติทางพันธุกรรม – ปัญหาสุขภาพที่เกิดจากความผิดปกติของจีโนม
- พันธุศาสตร์เชิงลำดับวงศ์ตระกูล – การตรวจดีเอ็นเอเพื่อหาความสัมพันธ์ทางสายเลือด
- แฮพลโลไทป์ – กลุ่มยีนที่ได้รับมาจากพ่อหรือแม่ฝ่ายใดฝ่ายหนึ่ง
- ไมโอซิส – การแบ่งเซลล์เพื่อสร้างเซลล์สืบพันธุ์แบบแฮพลอยด์
- สัญลักษณ์กรดนิวคลีอิก – สัญลักษณ์สากลของนิวคลีโอไทด์ในดีเอ็นเอ
- ลำดับกรดนิวคลีอิก – ลำดับของนิวคลีโอไทด์ในกรดนิวคลีอิก
- รูปแบบของดีเอ็นเอในสารานุกรมโปรตีโอพีเดีย
- ดีเอ็นเอไรโบโซม – ยีนที่เข้ารหัสอาร์เอ็นเอไรโบโซม
- เทคนิคการวิเคราะห์ดีเอ็นเอแบบเซาเทิร์นบลอต
- เทคนิคการกระเจิงรังสีเอกซ์ – กลุ่มเทคนิคการวิเคราะห์
- กรดนิวคลีอิกสังเคราะห์ – สารอะนาล็อกของกรดนิวคลีอิกสังเคราะห์
อ่านเพิ่มเติม
- Berry A, Watson J (2003). DNA: ความลับของชีวิต . นิวยอร์ก: Alfred A. Knopf. ISBN 0-375-41546-7.
{{cite book}}: CS1 maint: การตั้งค่าที่ถูกแทนที่ ( ลิงก์ ) - Calladine CR, Drew HR, Luisi BF, Travers AA (2003). ทำความเข้าใจดีเอ็นเอ: โมเลกุลและวิธีการทำงาน . อัมสเตอร์ดัม: สำนักพิมพ์ Elsevier Academic Press. ISBN 0-12-155089-3.
- Carina D, Clayton J (2003). 50 ปีแห่งดีเอ็นเอ . เบซิงสโตก: Palgrave Macmillan. ISBN 1-4039-1479-6.
- Judson HF (1979). วันที่แปดแห่งการสร้างสรรค์: ผู้สร้างการปฏิวัติในชีววิทยา (ฉบับที่ 2). สำนักพิมพ์ Cold Spring Harbor Laboratory Press. ISBN 0-671-22540-5.
- Olby RC (1994). เส้นทางสู่เกลียวคู่: การค้นพบดีเอ็นเอ . นิวยอร์ก: Dover Publications. ISBN 0-486-68117-3.หนังสือเล่มนี้ตีพิมพ์ครั้งแรกในเดือนตุลาคม ปี 1974 โดยสำนักพิมพ์ MacMillan พร้อมคำนำโดย Francis Crick นับเป็นตำราเกี่ยวกับดีเอ็นเอที่ได้รับการยอมรับอย่างกว้างขวาง และได้รับการแก้ไขเพิ่มเติมในปี 1994 โดยมีภาคผนวกเก้าหน้า
- Olby R (มกราคม 2546). "การเปิดตัวอย่างเงียบๆ ของเกลียวคู่" Nature . 421 ( 6921): 402–05 . Bibcode : 2003Natur.421..402O . doi : 10.1038/nature01397 . PMID 12540907 .
- Olby RC (2009). ฟรานซิส คริก: ชีวประวัติ . เพลนวิว, นิวยอร์ก: สำนักพิมพ์โคลด์สปริงฮาร์เบอร์แล็บ. ISBN 978-0-87969-798-3.
- Micklas D (2003). วิทยาศาสตร์ดีเอ็นเอ: หลักสูตรเบื้องต้น . สำนักพิมพ์ Cold Spring Harbor. ISBN 978-0-87969-636-8.
- Ridley M (2006). Francis Crick: ผู้ค้นพบรหัสพันธุกรรม . Ashland, OH: Eminent Lives, Atlas Books. ISBN 0-06-082333-X.
- Rosenfeld I (2010). DNA: คู่มือภาพประกอบเกี่ยวกับโมเลกุลที่เขย่าโลก . สำนักพิมพ์มหาวิทยาลัยโคลัมเบีย. ISBN 978-0-231-14271-7.
- Schultz M, Cannon Z (2009). The Stuff of Life: A Graphic Guide to Genetics and DNA . Hill and Wang. ISBN 978-0-8090-8947-5.
- Stent GS , Watson J (1980). The Double Helix: A Personal Account of the Discovery of the Structure of DNA . นิวยอร์ก: Norton. ISBN 0-393-95075-1.
- Watson J (2004). DNA: ความลับของชีวิต . Random House. ISBN 978-0-09-945184-6.
- วิลกินส์ เอ็ม (2003). ชายคนที่สามแห่งเกลียวคู่: อัตชีวประวัติของมอริซ วิลกินส์ เคมบริดจ์ ประเทศอังกฤษ: สำนักพิมพ์มหาวิทยาลัยISBN 0-19-860665-6.
ลิงก์ภายนอก
- การทำนายตำแหน่งการจับกับ DNA บนโปรตีน
- เกมดีเอ็นเอเกลียวคู่จากเว็บไซต์อย่างเป็นทางการของรางวัลโนเบล
- ดีเอ็นเอภายใต้กล้องจุลทรรศน์อิเล็กตรอน
- ศูนย์การเรียนรู้ดีเอ็นเอโดลัน
- โครงสร้างเกลียวคู่: 50 ปีแห่งดีเอ็นเอ , เนเจอร์
- โปรตีโอพีเดียดีเอ็นเอ
- รูปแบบของดีเอ็นเอในสารานุกรมโปรตีโอพีเดีย
- เครื่องมือสำรวจเธรด ENCODEหน้าหลัก ENCODE ที่Nature
- ศูนย์การศึกษาเทคโนโลยีชีวภาพแห่งชาติ1953–2003 (Double Helix 1953–2003)
- สื่อการสอนพันธุศาสตร์สำหรับครู – คู่มือการเรียนรู้ เรื่อง ดีเอ็นเอตั้งแต่เริ่มต้น
- โมเลกุลประจำเดือนของPDB : DNA
- "พบเบาะแสสำคัญเกี่ยวกับเคมีของการถ่ายทอดทางพันธุกรรม"หนังสือพิมพ์เดอะนิวยอร์กไทมส์มิถุนายน 1953 นี่คือรายงานข่าวในหนังสือพิมพ์อเมริกันฉบับแรกเกี่ยวกับการค้นพบโครงสร้างดีเอ็นเอ
- ดีเอ็นเอตั้งแต่เริ่มต้นอีกหนึ่งเว็บไซต์จากศูนย์การเรียนรู้ดีเอ็นเอ เกี่ยวกับดีเอ็นเอ ยีน และพันธุกรรม ตั้งแต่เมนเดลจนถึงโครงการจีโนมมนุษย์
- ทะเบียนเอกสารส่วนตัวของฟรานซิส คริก ปี 1938 – 2007ณ ห้องสมุดเอกสารพิเศษแมนเดวิลล์มหาวิทยาลัยแคลิฟอร์เนีย ซานดิเอโก
- จดหมายที่เขียนด้วยลายมือของคริกจำนวนเจ็ดหน้า ส่งถึงไมเคิล ลูกชายวัย 12 ปีของเขาในปี 1953 ซึ่งอธิบายโครงสร้างของดีเอ็นเอดูได้จากบทความ "เหรียญรางวัลของคริกถูกนำออกประมูล" ในวารสาร Nature ฉบับวันที่ 5 เมษายน 2013
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ดีเอ็นเอ
กรดดีออกซีไรโบนิวคลีอิก ( การออกเสียง ⓘ ; [ 1 ] DNA ) เป็น พอลิเมอร์ ที่ประกอบด้วย โพลีนิวคลีโอไทด์ สายที่พันกันเป็น เกลียวคู่ ลิเมอร์นี้บรรจุ ทางพันธุกรรม สำหรับการพัฒนา การทำงาน...
คุณสมบัติ
ดีเอ็นเอเป็น พอลิเมอร์ สายยาว ที่สร้างจากหน่วยซ้ำๆ ที่เรียกว่า นิวคลีโอไทด์ [ 6 ] [ 7 ] โดย ปกติดีเอ็นเอไม่ได้อยู่ในรูปสายเดี่ยว แต่เป็นสายคู่ที่ยึดติดกันแน่น [ 8 ] [ 9 ] สายยาวสองสายนี้จะพันกันเป็นเกลียวคู่ นิวคลี โอ ไทด์ประกอบด้วยส่วนของโครงสร้างหลัก ของ...
การจำแนกประเภทนิวคลีโอเบส
นิวคลีโอเบสแบ่งออกเป็นสองประเภท ได้แก่ พิวรีน A และ G ซึ่ง เป็น สารประกอบเฮเทอโรไซคลิก ห้าและหกสมาชิกที่หลอมรวมกันและ ไพริมิดีน ซึ่ง เป็น วงแหวนหกสมาชิก C และ T [ 9 ] นิวคลีโอเบสไพริมิดีนตัวที่ห้า คือ ยูราซิล ( U ) มักจะเข้ามาแทนที่ไทมีนใน RNA...
ฐานที่ไม่เป็นไปตามแบบแผน
เบสที่ถูกดัดแปลงเกิดขึ้นใน DNA เบสแรกที่ถูกค้นพบคือ 5-เมทิลไซโตซีน ซึ่งพบใน จีโนม ของ Mycobacterium tuberculosis ในปี พ.ศ.