อ่าน 18 นาที
ซินาปซิดา
Synapsida [ a ] เป็นกลุ่ม สัตว์มีกระดูกสันหลัง สี่ขา ที่มีความหลากหลาย ซึ่งรวมถึง สัตว์เลี้ยงลูกด้วยนม ทั้งหมด และญาติที่สูญพันธุ์ไปแล้ว เป็นหนึ่งในสอง กลุ่ม หลัก ของกลุ่ม Amniota...
ซินาปซิดา
| ซินาปซิดา ช่วงเวลา: | |
|---|---|
 | |
| ตัวอย่างของสัตว์ในกลุ่มซินาปซิด (จากซ้ายไปขวา บนลงล่าง): Cotylorhynchus , Dimetrodon , Inostrancevia , Moschops , Castorocauda , Adelobasileus , Tachyglossus aculeatusและPanthera tigris | |
| การจำแนกทางวิทยาศาสตร์ | |
| อาณาจักร: | แอนิมอลเลีย |
| ไฟลัม: | คอร์ดาต้า |
| กลุ่มสายพันธุ์ : | เตตระโพดา |
| กลุ่มสายพันธุ์ : | เรปทิลิโอมอร์ฟา |
| กลุ่มสายพันธุ์ : | แอมนิโอตา |
| กลุ่มสายพันธุ์ : | ซินาปซิดาออสบอร์น , 1903 |
| กลุ่มย่อย | |
| |
| คำพ้องความหมาย | |
Theropsida ( Seeley , 1895) [ 5 ] " Pelycosauria " (รวมถึง therapsids ตามการจัดกลุ่ม) | |
Synapsida [ a ]เป็นกลุ่มสัตว์มีกระดูกสันหลังสี่ขา ที่มีความหลากหลาย ซึ่งรวมถึงสัตว์เลี้ยงลูกด้วยนม ทั้งหมด และญาติที่สูญพันธุ์ไปแล้ว เป็นหนึ่งในสองกลุ่ม หลัก ของกลุ่มAmniota อีกกลุ่มหนึ่งคือกลุ่ม Sauropsidaที่มีความหลากหลายมากกว่า(ซึ่งรวมถึงสัตว์เลื้อยคลานและนก ที่ยังมีชีวิตอยู่ทั้งหมด ) Synapsida มี ช่องเปิดขมับล่างเพียงช่องเดียว ซึ่ง เป็นช่องเปิดที่อยู่ต่ำในส่วนบนของกะโหลกศีรษะ ด้านหลัง เบ้าตาแต่ละข้างช่องเปิดเหล่านี้สร้างส่วนโค้งของกระดูกที่ช่วยให้กล้ามเนื้อขากรรไกรขนาดใหญ่ยึดติดได้และทำให้การกัดมีประสิทธิภาพมากขึ้น[ 6 ]แม้ว่าก่อนหน้านี้จะมีการโต้แย้งว่าช่องเปิดขมับล่างเพียงช่องเดียวเป็นลักษณะเฉพาะของ Synapsida แต่ก็พบได้ใน Sauropsida ดั้งเดิมหลายชนิด เมื่อเร็วๆ นี้มีการโต้แย้งว่าบรรพบุรุษร่วมสุดท้ายของ Synapsida และ Sauropsida มีช่องเปิดขมับล่างเพียงช่องเดียวร่วมกัน[ 7 ]
สัตว์ มีกระดูกสันหลังกลุ่มแอมนิโอต พื้นฐาน (เรปทิลิโอเมอร์ฟ ) ซึ่งเป็นต้นกำเนิดของซินาปซิดนั้น ในอดีตถูกเรียกว่า "สัตว์เลื้อยคลาน" เฉยๆ ดังนั้น ซินาปซิดที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนมจึงถูกอธิบายว่าเป็น สัตว์ เลื้อยคลานที่คล้ายสัตว์เลี้ยงลูกด้วยนมในระบบอนุกรมวิธานแบบดั้งเดิม และซินาปซิดที่ไม่ใช่เทอราปซิดก็ถูกเรียกว่าเพลิโคซอร์หรือซินาปซิดระดับเพลิ โคซอร์ด้วย คำศัพท์ เหล่านี้ ซึ่งจัดอยู่ในกลุ่ม พาราไฟเลติก นั้น ไม่เป็นที่นิยมแล้ว และใช้กันอย่างไม่เป็นทางการ (หรืออาจจะไม่ใช้เลย) ในวรรณกรรมสมัยใหม่ เนื่องจากปัจจุบันเป็นที่ทราบกันดีว่า สัตว์เลื้อยคลาน ที่ยังมีชีวิตอยู่ ทั้งหมด มีความสัมพันธ์ใกล้ชิดกันเองและกับนกมากกว่าซินาปซิด ดังนั้นคำว่า "สัตว์เลื้อยคลาน" จึงถูกนิยามใหม่ให้หมายถึงเฉพาะสมาชิกของซอรอปซิดาหรือแม้แต่กลุ่มย่อยของมัน ใน แง่ของค ลัดิสติก ซินาปซิดนั้นแท้จริงแล้วเป็นกลุ่มพี่น้องโมโนไฟเลติก ของซอรอปซิดมากกว่าที่จะเป็นส่วนหนึ่งของสายพันธุ์ซอรอปซิด[ 8 ] [ 9 ] [ 10 ] [ 11 ]ทางเลือกสมัยใหม่ที่บางครั้งใช้ ได้แก่สัตว์เลี้ยงลูกด้วยนมดั้งเดิมสัตว์ เลี้ยง ลูกด้วยนมดั้งเดิม สัตว์เลี้ยงลูกด้วยนมกึ่งสัตว์เลี้ยงลูกด้วยนม หรือสัตว์เลี้ยงลูกด้วยนมทุก ชนิด [ 12 ] [ 13 ] [ 14 ]สายพันธุ์ส่วนใหญ่ของซินาปซิดระดับเพลิโคซอร์ถูกแทนที่ด้วยเทอราปซิดที่ก้าวหน้ากว่า ซึ่งวิวัฒนาการมาจาก เพ ลิโคซอร์สเฟนาโคดอนเทียน ในช่วงปลายยุคเพอร์เมียนตอนต้น การเปลี่ยนแปลงนี้เรียกว่า ช่องว่าง/การสูญ พันธุ์ ของโอลสัน
ซินาปซิดส์ยุคแรกสุดปรากฏขึ้นในช่วงปลายยุคคาร์บอนิเฟอรัสประมาณ 315 ถึง 307 ล้านปีก่อน[ 15 ]ซินาปซิดส์เป็นสัตว์มีกระดูกสันหลังบนบกที่ใหญ่ที่สุดในยุคเพอร์เมียน (299 ถึง 251 ล้านปีก่อน) รองลงมาก็คือพาราเรปไทล์พาเรอาซอเรียนขนาดใหญ่บางชนิด เช่น สคูโตซอรัส พวกมันเป็นนักล่าบนบกที่โดดเด่นในยุคพาลีโอ โซอิกตอนปลายและยุคมีโซโซอิกตอนต้นโดยมียูเพลิโคซอร์เช่นไดเมโทรดอนไททาโนโฟนัสและอิโนสทรานเซเวียเป็นนักล่าสูงสุดในยุคเพอร์เมียน และเทริโอดอนต์เช่นมอสโคไรนัสในช่วงต้นยุคไทรแอสสิก ประชากรและความหลากหลายของซินาปซิดลดลงอย่างมากจากเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในยุคคาปิทาเนียนและเหตุการณ์การสูญพันธุ์ในยุคเพอร์เมียน-ไทรแอสสิกและมีเพียงสองกลุ่มของเทอราปซิด ได้แก่ไดซิโนดอนต์และยูเทอริโอดอนต์ (ประกอบด้วยเทอโรเซฟาเลียนและไซโนดอนต์ ) ที่ทราบว่ารอดชีวิตมาจนถึงยุคไทรแอสสิก เทอราปซิดเหล่านี้ฟื้นตัวขึ้นมาในฐานะกลุ่มสิ่งมีชีวิตที่สูญพันธุ์ในช่วงต้นยุคมีโซโซอิก โดยไดซิโน ดอนต์ลิส โทรซอ รัส มีสัดส่วนมากถึง 95% ของสิ่งมีชีวิตบนบกทั้งหมดในบางช่วงเวลา[ 16 ] [ 17 ]แต่ก็ลดลงอีกครั้งหลังจากเหตุการณ์รอยต่อสมิธเทียน-สปาเธียนโดยนิเวศวิทยา ที่โดดเด่นของพวกมัน ส่วนใหญ่ถูกแทนที่โดยการเพิ่มขึ้นของ ซอรอปซิด อาร์โคซอเรียน โดยเริ่มจากซูโดซูเชียนจากนั้นก็เป็นเทโรซอร์และไดโนเสาร์กลุ่มไซโนดอนต์Probainognathiaซึ่งรวมถึงกลุ่มMammaliaformesเป็นไซแนปซิดเพียงกลุ่มเดียวที่รอดชีวิตมาได้หลังยุคไทรแอสสิก[ 18 ]ในช่วงยุคจูราสสิกไซโนดอนต์ส่วนใหญ่หากินในเวลากลางคืนเพื่อหลีกเลี่ยงการแข่งขันกับไดโนเสาร์ หลังจากเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในยุคครีเทเชียส-พาลีโอจีนได้กวาดล้างไดโนเสาร์และเทโรซอร์ที่ไม่ใช่นกทั้งหมด ไซแนปซิด (ในฐานะสัตว์เลี้ยงลูกด้วยนม) ก็กลับมาครองอำนาจอีกครั้งในช่วงยุคซีโนโซอิกโดยปัจจุบันมีประมาณ 6,800 ชนิดที่ยังมีชีวิตอยู่[ 19 ]
การจำแนกประเภทแบบลินเนียนและแบบคลาดิสติก
ในช่วงต้นศตวรรษที่ 20 เชื่อกันว่าไซแนปซิดเป็นหนึ่งในสี่กลุ่มย่อยหลักของสัตว์เลื้อยคลานอย่างไรก็ตาม ความคิดนี้ถูกหักล้างเมื่อตรวจสอบซากโครงกระดูกอย่างละเอียด เนื่องจากไซแนปซิดแตกต่างจากสัตว์เลื้อยคลานตรงที่มีช่องเปิดบริเวณขมับที่โดดเด่น ช่องเปิดเหล่านี้ใน กระดูก กะโหลกศีรษะช่วยให้กล้ามเนื้อขากรรไกรขนาดใหญ่สามารถยึดเกาะได้ จึงทำให้การกัดมีประสิทธิภาพมากขึ้น
ต่อมา Synapsids ถูกพิจารณาว่าเป็นสายพันธุ์สัตว์เลื้อยคลานในยุคหลังที่พัฒนาเป็นสัตว์เลี้ยงลูกด้วยนมโดยค่อยๆวิวัฒนาการลักษณะคล้ายสัตว์เลี้ยงลูกด้วยนมมากขึ้นเรื่อยๆ จึงได้ชื่อว่า "สัตว์เลื้อยคลานคล้ายสัตว์เลี้ยงลูกด้วยนม" (หรือที่รู้จักกันในชื่อpelycosaurs ) ซึ่งกลายเป็นคำเรียกแบบดั้งเดิมสำหรับ synapsids ในยุค Paleozoic (ยุคแรก) ทั้งหมด อย่างไรก็ตาม การศึกษาล่าสุดได้หักล้างแนวคิดนี้เช่นกัน และปัจจุบันสัตว์เลื้อยคลานถูกจัดอยู่ในSauropsida (sauropsids) ซึ่งเป็นกลุ่มพี่น้องกับ synapsids ทำให้ synapsids เป็นกลุ่มอนุกรมวิธาน ของตัวเอง [ 8 ] [ 10 ]
ด้วยเหตุนี้ คำศัพท์ พาราไฟเลติกอย่าง "สัตว์เลื้อยคลานคล้ายสัตว์เลี้ยงลูกด้วยนม" และ "เพลิโคซอร์" จึงถูกมองว่าล้าสมัยและไม่เป็นที่นิยมในเอกสารทางเทคนิค และมีการใช้คำ ว่า สัตว์เลี้ยง ลูกด้วยนมต้นกำเนิด (หรือบางครั้ง เรียกว่า โปรโตแมมมอลหรือพาราแมมมอล ) แทนในทางวิวัฒนาการปัจจุบันเป็นที่เข้าใจกันว่าไซแนปซิดประกอบเป็นสาขาอิสระของ ต้นไม้ แห่งชีวิต[ 20 ]ความเป็นโมโนฟิเลติกของไซแนปซิดาไม่เป็นที่สงสัย และวลีต่างๆ เช่น "ไซแนปซิดาประกอบด้วยสัตว์เลี้ยงลูกด้วยนม" และ "ไซแนปซิดให้กำเนิดสัตว์เลี้ยงลูกด้วยนม" ต่างก็แสดงถึงสมมติฐานทางวิวัฒนาการเดียวกัน คำศัพท์นี้สะท้อนถึง แนวทาง คลาดิสติก สมัยใหม่ เกี่ยวกับความสัมพันธ์ของสัตว์ ซึ่งกลุ่มที่ถูกต้องเพียงกลุ่มเดียวคือกลุ่มที่รวมลูกหลานทั้งหมดของบรรพบุรุษร่วมกัน: กลุ่มเหล่านี้เรียกว่า กลุ่ม โมโนฟิเลติกหรือคลาเดส
นอกจากนี้Reptilia (สัตว์เลื้อยคลาน) ได้รับการแก้ไขให้เป็นกลุ่มโมโนฟิเลติกและถือว่าแตกต่างอย่างสิ้นเชิงจาก Synapsida โดยอยู่ในSauropsidaซึ่งเป็นกลุ่มพี่น้องของ Synapsida ภายใน Amniota [ 21 ]
ไซแนปซิดดั้งเดิมและไซแนปซิดขั้นสูง
ตามธรรมเนียมแล้ว ซินาปซิดจะถูกแบ่งออกเป็น กลุ่ม เทราปซิดซึ่งเป็นกลุ่มซินาปซิดขั้นสูงและเป็นสาขาที่สัตว์เลี้ยงลูกด้วยนมวิวัฒนาการขึ้นมา และสัตว์เลี้ยงลูกด้วยนมดั้งเดิม (เดิมเรียกว่าเพลิโคซอร์ ) ซึ่งประกอบด้วยซินาปซิดอีก 6 วงศ์ที่ดั้งเดิมกว่า[ 22 ]สัตว์เลี้ยงลูกด้วยนมดั้งเดิมทั้งหมดมีลักษณะคล้ายกิ้งก่า มีการเดินแบบกางขาและอาจมีเกล็ด แข็ง ในขณะที่เทราปซิดมักมีท่าทางที่ยืนตรงกว่าและอาจมีขน อย่างน้อยในบางรูปแบบ ในการจำแนกทางอนุกรมวิธานแบบดั้งเดิม ซินาปซิดาประกอบด้วยสองระดับ ที่แตกต่างกัน : สัตว์เลี้ยงลูกด้วยนมดั้งเดิมที่มีลำตัวเตี้ยได้ให้กำเนิดเทราปซิดที่ยืนตรงกว่า ซึ่งในทางกลับกันได้ให้กำเนิดสัตว์เลี้ยงลูกด้วยนม ในการจำแนกประเภทสัตว์มีกระดูกสันหลังแบบดั้งเดิม สัตว์เลี้ยงลูกด้วยนมดั้งเดิมและเทราปซิดต่างก็ถือว่าเป็นอันดับของชั้นย่อยซินาปซิดา[ 23 ] [ 8 ]
การใช้คำว่า "synapsid" และ "therapsid" ในเชิงปฏิบัติและเชิงวิวัฒนาการ
ในระบบการตั้งชื่อทางวิวัฒนาการคำเหล่านี้ถูกใช้แตกต่างออกไปเล็กน้อย เนื่องจากมีการรวมกลุ่มย่อยที่แตกแขนงออกมาด้วย บทความส่วนใหญ่ที่ตีพิมพ์ในช่วงศตวรรษที่ 21 ถือว่า "Pelycosaur" เป็นกลุ่มที่ไม่เป็นทางการของสมาชิกดั้งเดิม ในขณะที่ Therapsida ยังคงถูกใช้เป็นกลุ่มที่รวมทั้งวงศ์ Therapsid ดั้งเดิมและสัตว์เลี้ยงลูกด้วยนม
แม้ว่า Synapsida และ Therapsida จะรวมถึงสัตว์เลี้ยงลูกด้วยนมในปัจจุบัน แต่ในทางปฏิบัติแล้ว คำทั้งสองนี้มักใช้เฉพาะเมื่อกล่าวถึง สมาชิก ดั้งเดิมที่อยู่นอกเหนือกลุ่มMammaliaformesเท่านั้น
ลักษณะเฉพาะ
ช่องเปิดกะโหลกศีรษะ

สัตว์ในไฟลั ม Synapsids วิวัฒนาการช่อง เปิด ขมับ (temporal fenestra) อยู่ด้านหลัง เบ้าตาแต่ละข้างบนพื้นผิวด้านข้างของกะโหลกศีรษะ ซึ่งอาจเป็นจุดยึดใหม่สำหรับกล้ามเนื้อขากรรไกร เดิมที ช่องเปิดในกะโหลกศีรษะทำให้ส่วนในของกะโหลกถูกปกคลุมด้วยกล้ามเนื้อขากรรไกรเท่านั้น แต่ในสัตว์ในไฟลัม Terapsids ชั้นสูงและสัตว์เลี้ยงลูกด้วยนม กระดูกสฟีนอยด์ได้ขยายตัวเพื่อปิดช่องเปิด ทำให้ขอบล่างของช่องเปิดกลายเป็นส่วนโค้งที่ยื่นออกมาจากขอบล่างของกะโหลกสมอง
แม้ว่าในอดีตจะมีการโต้แย้งว่าลักษณะนี้เป็นลักษณะเฉพาะของซินาปซิด แต่การวิจัยล่าสุดแสดงให้เห็นว่าซอรอปซิดดั้งเดิมหลายชนิดก็มีช่องขมับล่างเพียงช่องเดียวเช่นกัน และยังมีการเสนอแนะว่าซอรอปซิดหลายชนิดที่คิดว่ามี กะโหลกแบบ อนาปซิดซึ่งไม่มีช่องขมับเลยนั้นไม่ใช่สัตว์เลื้อยคลาน แต่เป็นสเต็ม-แอมนิโอต ซึ่งหมายความว่าลักษณะนี้อาจมีอยู่ในบรรพบุรุษร่วมสุดท้ายของซินาปซิดและซอรอปซิด และไม่ใช่ลักษณะเฉพาะของซินาปซิดา[ 7 ]
บรรพบุรุษของซินาปซิดมีช่องเปิดบนหลังคากะโหลก (ช่องไพเนียล) สำหรับตาข้างขมับซึ่งเป็นอวัยวะที่ไวต่อแสงที่ยังคงพบในสัตว์ครึ่งบกครึ่งน้ำและสัตว์เลื้อยคลานบางชนิด ใช้สำหรับการปรับเทียบการควบคุมอุณหภูมิและจังหวะชีวิตประจำวันรวมถึงหน้าที่อื่นๆ การทำงานของตาหายไปและช่องไพเนียลปิดลงใน สายพันธุ์ ไซโนดอนต์ซึ่งเป็นบรรพบุรุษของสัตว์เลี้ยงลูกด้วยนม ( Probainognathia ) [ 24 ]
ฟัน




ไซแนปซิดหลายชนิดรวมถึงสัตว์เลี้ยงลูกด้วยนมที่ยังมีชีวิตอยู่มีลักษณะเด่นคือฟันเฮเทอโรดอนต์ ที่มีขนาดและ/หรือรูปร่างแตกต่างกันไปตามแถวฟัน ลักษณะนี้เริ่มปรากฏให้เห็นในไซแนปซิด "เพลิโคซอร์" ยุคแรกๆ หลายชนิด แต่กลับเด่นชัดขึ้นใน ไซแนปซิดสเฟนาโคดอน เทียนขั้นสูง โดยเฉพาะในเทอแรปซิด [ 25 ]ซึ่งฟันจะแยกออกเป็นฟันตัด ฟันเขี้ยว และฟันหลังเขี้ยว[ 26 ]ไดไซโนดอนต์เป็นไซแนปซิดที่มีลักษณะเฉพาะคือสูญเสียฟันส่วนใหญ่ (หรือทั้งหมดในกรณีของไดไซโนดอนต์บางชนิด) ไปเป็นจะงอยปากคล้ายเต่าที่ทำจากเคราติน[ 27 ]ปุ่มฟันปรากฏขึ้นครั้งแรกในบรรพบุรุษของสัตว์เลี้ยงลูกด้วยนมในกลุ่มไซโนดอนต์[ 25 ]เชื่อกันว่าไซแนปซิดที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนมส่วนใหญ่มีการเปลี่ยนฟันอย่างต่อเนื่องตลอดชีวิต ( โพลีฟิโอดอนตี ) [ 28 ]โดยไซโนดอนต์ขั้นสูงมีวิวัฒนาการของวงจรการเปลี่ยนฟันที่จำกัด ซึ่งสิ้นสุดลงด้วยการเกิดขึ้นของฟันสองชุด ( ไดฟิโอดอนตี ) ในMammaliformes (หรืออาจจะเป็นกลุ่มที่กว้างกว่าเล็กน้อยMammaliamorpha [ 29 ] ) [ 30 ]
ขากรรไกรและหู

เช่นเดียวกับสัตว์เลื้อยคลาน ขากรรไกรล่างของไซแนปซิดที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนมส่วนใหญ่ประกอบด้วยกระดูกหลักหลายชิ้น ได้แก่กระดูกเดนทารี (กระดูกที่รองรับฟัน) รวมถึงกระดูกอาร์ติคูลาร์แอ งกูลาร์ ซูแองกูลาร์และสปลีเนียลในไซโนดอนต์ กระดูกขากรรไกรเหล่านี้ ยกเว้นเดนทารี จะมีขนาดเล็ลงและอาจหายไป หรือในกรณีของแองกูลาร์และอาร์ติคูลาร์ จะค่อยๆ เคลื่อนตัวไปสร้างเป็นกระดูกหู ในขณะที่สัตว์เลี้ยงลูกด้วยนมในปัจจุบันมีกระดูกมัลเลียสอินคั ส ทิมพานิกและสเตปส์ไซ แนปซิด ดั้งเดิม (เช่นเดียวกับสัตว์มีกระดูกสันหลังสี่ขาอื่นๆ ทั้งหมด) มีเพียงกระดูกสเตปส์เท่านั้น กระดูกมัลเลียสมีต้นกำเนิดมาจากกระดูกอาร์ติคูลาร์ และกระดูกทิมพานิก (หรือที่รู้จักกันอย่างแพร่หลายในชื่อเอ็กโททิมพานิก ) มีต้นกำเนิดมาจากกระดูกแองกูลาร์ ในขณะที่กระดูกอินคัสมีต้นกำเนิดมาจากกระดูกควอดเรต (กระดูกกะโหลก) [ 31 ] [ 32 ] [ 33 ] [ 34 ]
โครงสร้างขากรรไกรของสัตว์เลี้ยงลูกด้วยนมยังแตกต่างกันด้วยข้อต่อขา กรรไกรเดนทารี-สควาโมซั ล ในข้อต่อขากรรไกรแบบนี้ เดนทารีจะเชื่อมต่อกับส่วนเว้าใน สควาโมซั ลที่เรียกว่าเบ้ากลีนอยด์ ในทางตรงกันข้าม สัตว์มีกระดูกสันหลังที่มีขากรรไกรอื่นๆ ทั้งหมด รวมถึงสัตว์เลื้อยคลานและไซแนปซิดที่ไม่ใช่ไซโนดอนต์ จะมีข้อต่อขากรรไกรที่กระดูกชิ้นเล็กชิ้นหนึ่งของขากรรไกรล่าง คือกระดูกอาร์ติคูลาร์ จะเชื่อมต่อกับกระดูกของกะโหลกศีรษะที่เรียกว่ากระดูกควอดเรตเพื่อสร้างข้อต่อขากรรไกรอาร์ติคูลาร์-ควอดเรต ไซโนดอนต์ที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนมแสดงข้อต่อที่อยู่ระหว่างสองสภาวะนี้[ 31 ]
เพดานปาก
บรรพบุรุษของซินาปซิดมีฟันเพดานปากซึ่งอยู่บนกระดูกของเพดานปาก ฟันเหล่านี้หายไปในบรรพบุรุษของสัตว์เลี้ยงลูกด้วยนมที่เป็นไซโนดอน[ 35 ]
เมื่อเวลาผ่านไป ซินาปซิดส์เริ่มมีลักษณะคล้ายสัตว์เลี้ยงลูกด้วยนมมากขึ้นและคล้ายสัตว์เลื้อยคลานน้อยลง พวกมันจึงเริ่มพัฒนาเพดานปากส่วนที่สองซึ่งแยกช่องปากและโพรงจมูก ออกจากกัน ในซินาปซิดส์ยุคแรก เพดานปากส่วนที่สองเริ่มก่อตัวขึ้นที่ด้านข้างของกระดูกขากรรไกร บน โดยที่ช่องปากและรูจมูกยังคงเชื่อมต่อกันอยู่
ในที่สุด เพดานปากทั้งสองข้างก็เริ่มโค้งเข้าหากัน เกิดเป็นรูปตัว U แทนที่จะเป็นรูปตัว C เพดานปากยังเริ่มยื่นออกไปทางลำคอ ยึดปากทั้งหมดไว้ และสร้างกระดูกเพดานปาก ที่สมบูรณ์ กระดูกขากรรไกรบนก็ปิดสนิทเช่นกัน ในฟอสซิลของยูเทอริโอดอนต์ กลุ่มแรกๆ จะเห็นจุดเริ่มต้นของเพดานปากได้อย่างชัดเจน ทรินาโซดอนในยุคหลังมีเพดานปากที่สมบูรณ์และปิดสนิท ทำให้เกิดความก้าวหน้าที่ชัดเจน[ 36 ]
ผิวหนังและขน

นอกจากผิวหนังที่มีต่อมและปกคลุมด้วยขนซึ่งพบในสัตว์เลี้ยงลูกด้วยนมส่วนใหญ่ในปัจจุบันแล้ว ซินาปซิดทั้งในปัจจุบันและที่สูญพันธุ์ไปแล้วยังมีผิวหนังที่ดัดแปลงหลากหลายรูปแบบ รวมถึง ออสทีโอ เดอร์ม (เกราะกระดูกที่ฝังอยู่ในผิวหนัง) สคิวต์ (โครงสร้างป้องกันของชั้นหนังแท้ซึ่งมักมีเปลือกแข็งปกคลุม) ขน หรือขนสัตว์ และ โครงสร้างคล้าย เกล็ด (มักเกิดจากขนที่ดัดแปลง เช่นในตัวนิ่มและสัตว์ฟันแทะ บางชนิด ) ในขณะที่ผิวหนังของสัตว์เลื้อยคลานค่อนข้างบาง แต่ผิวหนังของสัตว์เลี้ยงลูกด้วยนมมีชั้นหนังแท้ ที่หนา [ 37 ]
แม้ว่าก่อนหน้านี้ลักษณะของผิวหนังของซินาปซิดในยุคแรกจะยังไม่ชัดเจน แต่ปัจจุบันได้มีการแสดงให้เห็นอย่างชัดเจนแล้วว่าซินาปซิด "เพลิโคซอร์" ในยุคแรกนั้นปกคลุมด้วยเกล็ดผิวหนังคล้ายกับของสัตว์เลื้อยคลาน (โดยที่ลำตัวของสเฟนาโคดอนทิดอย่างน้อยบางชนิดมีเกล็ดรูปสี่เหลี่ยมผืนผ้าอยู่ใต้ลำตัว ในขณะที่แขนขาของพวกมันปกคลุมด้วยเกล็ดรูปหกเหลี่ยม) มีการเสนอแนะว่าเกล็ดผิวหนังวิวัฒนาการขึ้นในบรรพบุรุษร่วมกันก่อนที่จะเกิดการแยกตัวระหว่างสัตว์เลื้อยคลานและซินาปซิด[ 38 ]
ปัจจุบันยังไม่ทราบแน่ชัดว่าลักษณะของสัตว์เลี้ยงลูกด้วยนม เช่นขนตามร่างกายและต่อมน้ำนมปรากฏขึ้นครั้งแรกเมื่อใด เนื่องจากฟอสซิลให้หลักฐานโดยตรงเกี่ยวกับเนื้อเยื่ออ่อนได้น้อยมาก[ 39 ]ร่องรอยผิวหนังที่ชัดเจนเพียงอย่างเดียวที่รู้จักจากเทอแรปซิดที่ไม่ใช่ไซโนดอนต์ คือ ร่องรอยของไดโนเซฟาเลียน เอสเตมเมโนซูคัสจากยุคเพอร์เมียนตอนกลางของรัสเซีย และไดไซโนดอนต์ ลิสโทรซอ รัส จากยุคไทรแอสสิกตอนต้นของแอฟริกาใต้ ซึ่งทั้งสองชนิดแสดงให้เห็น "ตุ่มรูปไข่ ที่โป่งออกมา โดยมีแนวโน้มที่จะยื่นออกมาบางส่วนซึ่งกันและกัน ทำให้เกิดพื้นผิวที่เป็นตุ่มหนองโดยรวม" [ 40 ]โครงสร้างคล้ายเส้นผมในอุจจาระยุคเพอร์เมียนตอนปลาย จากรัสเซียและอาจรวมถึงแอฟริกาใต้ อาจบ่งชี้ว่าเทอแรปซิดบางชนิดอาจมีขนในช่วงเวลานี้[ 41 ]แม้ว่าลักษณะของโครงสร้างเหล่านี้จะคลุมเครือ[ 42 ]
มีการเสนอว่า หนวดอาจวิวัฒนาการขึ้นครั้งแรกในไซโนดอนต์ที่อยู่ในกลุ่มProzostrodontiaโดยพิจารณาจากกายวิภาคของกะโหลกศีรษะ[ 42 ]ฟอสซิลที่เก่าแก่ที่สุดที่แสดงร่องรอยของขนอย่างชัดเจนคือCastorocaudaจากยุค Callovian (ปลาย ยุคจูราสสิกตอนกลาง) และharamiyidans ร่วมสมัยหลายชนิด ซึ่งทั้งสองชนิด เป็น mammaliaformที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนม[ 43 ] [ 44 ] (อย่างไรก็ตาม โปรดดูด้านล่าง) นอกจากนี้ยังมีการตั้งสมมติฐานว่าสมาชิกดั้งเดิมของไซโนดอนต์อาจมีขนหรือสิ่งปกคลุมคล้ายขนโดยพิจารณาจากการเผาผลาญเลือดอุ่นที่อนุมานได้[ 45 ]ในขณะที่หลักฐานโดยตรงมากขึ้นเกี่ยวกับขนในไซโนดอนต์ยุคแรกได้รับการเสนอในรูปแบบของหลุมเล็กๆ บนจมูกซึ่งอาจเกี่ยวข้องกับหนวดหลุมดังกล่าวก็พบได้ในสัตว์เลื้อยคลานบางชนิดที่ไม่มีหนวดเช่นกัน[ 45 ]มีหลักฐานว่าไซโนดอนต์ที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนมบางชนิดที่มีวิวัฒนาการพื้นฐานกว่าCastorocaudaเช่นMorganucodonมีต่อม Harderianซึ่งเกี่ยวข้องกับการดูแลและบำรุงรักษาขน การที่ไม่มีต่อมเหล่านี้ในไซโนดอนต์ที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนมอาจบ่งชี้ว่าขนไม่ได้เกิดขึ้นจนกระทั่งถึงจุดนั้นในวิวัฒนาการของไซแนปซิด[ 45 ]เป็นไปได้ว่าขนและลักษณะที่เกี่ยวข้องกับสัตว์เลือดอุ่นที่แท้จริงไม่ได้ปรากฏขึ้นจนกระทั่งไซแนปซิดบางชนิดมีขนาดเล็กมากและออกหากินในเวลากลางคืน ซึ่งจำเป็นต้องมีการเผาผลาญที่สูงขึ้น[ 45 ]การกลายพันธุ์ของยีน MSX2 ในหนูแสดงให้เห็นว่าเมื่อยีนไม่ทำงาน หนูจะไม่สามารถพัฒนาขนได้ และช่องไพเนียลซึ่งปิดอยู่แต่เดิมในสัตว์เลี้ยงลูกด้วยนมจะเปิดออก นี่อาจบ่งชี้ว่าการพัฒนาทั้งสองอย่างเกิดขึ้นในเวลาเดียวกันในบรรพบุรุษไซโนดอนต์ของสัตว์เลี้ยงลูกด้วยนม[ 39 ]
ปาตาเกีย
การเคลื่อนที่ในอากาศเริ่มต้นขึ้นครั้งแรกใน ไซโนดอน ต์ ฮารามิ ยิดันที่ ไม่ใช่สัตว์เลี้ยงลูกด้วยนมโดย อาร์โบโร ฮารามิยะเซียนโชว ไมโอ พาทาเจียม และวิเลโวโลดอนมีเยื่อปีกที่ปกคลุมด้วยขนซึ่งได้รับการอนุรักษ์ไว้อย่างดีเยี่ยม ซึ่งทอดยาวไปทั่วแขนขาและหาง นิ้วของพวกมันยาวคล้ายกับของค้างคาวและโคลูโกและน่าจะมีบทบาทคล้ายกันทั้งในการค้ำยันปีกและในการเกาะกิ่งไม้[ 46 ]
ในกลุ่มสัตว์เลี้ยงลูกด้วยนมที่แท้จริง การเคลื่อนที่ในอากาศเกิดขึ้นครั้งแรกในยูทริโคโนดอนต์ในกลุ่มโวลาติโคเทอเรียม ฟอสซิลของ โวลาติ โคเทอเรียม มีแผ่น เนื้อใต้ปีกที่มีขนปกคลุมอย่างสวยงาม มีรอยย่นละเอียด และมีขนาดใหญ่มาก ห่อหุ้มมือและเท้าที่เก็บรักษาไว้ไม่ดี และขยายไปจนถึงโคนหาง[ 47 ]อาร์เจนโตโคโนดอนซึ่งเป็นญาติใกล้ชิด มีกระดูกต้นขาที่คล้ายคลึงกันซึ่งปรับตัวให้เข้ากับแรงกดดันในการบิน แสดงให้เห็นถึงวิถีชีวิตที่คล้ายคลึงกัน[ 48 ]
สัตว์เลี้ยงลูกด้วยนม เทเรียน จะสามารถบินและร่อนได้ด้วยพลังงานก็ต่อเมื่อนักบินยุคแรกเหล่านี้สูญพันธุ์ไปนานแล้ว โดยสัตว์เลี้ยงลูกด้วย นมเมตา เทอเรียน และค้างคาวที่ร่อนได้ที่เก่าแก่ที่สุดเท่าที่รู้จักนั้นวิวัฒนาการขึ้นในยุคพาลีโอซีน[ 49 ]
ระบบสืบพันธุ์และต่อมน้ำนม

ไซแนปซิดที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนมส่วนใหญ่หรือทั้งหมดอาจวางไข่ที่มีลักษณะอ่อนนุ่มคล้ายหนัง เหมือนกับไข่ของโมโนทรีมซึ่งไม่มีเปลือกแข็งหนาเหมือนที่พบในไข่ของสัตว์เลื้อยคลานและนกในปัจจุบันหลายชนิด[ 50 ]นี่อาจอธิบายได้ว่าทำไมจึงไม่มีหลักฐานฟอสซิลของเปลือกไข่ของไซแนปซิดจนถึงปัจจุบัน[ 50 ] [ 51 ]การวางไข่ในไซแนปซิดที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนมดูเหมือนจะได้รับการยืนยันโดยตัวอ่อนที่ขดแน่นของไดไซโนดอนเทอแรปซิดลิสโทรซอรัสที่พบในยุคไทรแอสสิกตอนต้นของแอฟริกาใต้ ซึ่งน่าจะถูกวางในไข่ที่อ่อนนุ่มคล้ายหนัง (ที่ไม่ได้รับการเก็บรักษาไว้) [ 50 ]เนื่องจากไข่เหล่านี้มีความเสี่ยงต่อการ แห้ง การหลั่งจาก ต่อมคล้าย อะโพครีนอาจช่วยรักษาความชุ่มชื้นของไข่ได้[ 52 ]
ตามที่ Oftedal กล่าวไว้ สัตว์กลุ่มซินาปซิดในยุคแรกอาจฝังไข่ลงในดินที่มีความชื้นสูง เพื่อให้ไข่ชุ่มชื้นด้วยการสัมผัสกับผิวหนังที่ชื้น หรืออาจอุ้มไข่ไว้ในถุงที่ชื้นคล้ายกับสัตว์กลุ่มโมโนทรีม ( เม่นหนามอุ้มไข่และลูกอ่อนไว้ในถุงชั่วคราว[ 53 ] [ 54 ] ) แม้ว่าวิธีนี้จะจำกัดการเคลื่อนที่ของพ่อแม่ก็ตาม วิธีหลังนี้อาจเป็นรูปแบบดั้งเดิมของการดูแลไข่ในสัตว์กลุ่มซินาปซิดมากกว่าการฝังไข่ และข้อจำกัดในการเคลื่อนที่ของพ่อแม่จะได้รับการแก้ไขโดยการ "พัก" ไข่ไว้ในรังระหว่างการหาอาหารหรือกิจกรรมอื่นๆ และให้ความชุ่มชื้นเป็นระยะ ทำให้สามารถวางไข่ได้มากกว่าที่จะใส่ไว้ในถุง (หรือหลายถุง) ในคราวเดียว และไข่ขนาดใหญ่ซึ่งจะลำบากในการอุ้มในถุงก็จะดูแลได้ง่ายกว่า พื้นฐานของการคาดเดาของ Oftedalคือข้อเท็จจริงที่ว่ากบหลายชนิดสามารถแบกไข่หรือลูกอ๊อดที่ติดอยู่กับผิวหนัง หรือฝังอยู่ใน "ถุง" บนผิวหนัง และซาลาแมนเดอร์ ส่วนใหญ่ จะม้วนตัวรอบไข่เพื่อรักษาความชุ่มชื้น ซึ่งทั้งสองกลุ่มนี้ยังมีผิวหนังที่มีต่อมอีกด้วย[ 51 ]ขนาดไข่ที่ค่อนข้างใหญ่ของLystrosaurusบ่งชี้ว่าไดไซโนดอนต์และอาจรวมถึงไซแนปซิดที่ไม่ใช่ไซโนดอนต์อื่นๆ มีไข่แดงเพียงพอที่จะพัฒนาได้เพียงพอโดยไม่ต้องใช้น้ำนม ซึ่งบ่งชี้ว่าพวกมันขาดน้ำนม[ 50 ]
ต่อมที่เกี่ยวข้องกับกลไกนี้จะวิวัฒนาการไปเป็นต่อมน้ำนมที่แท้จริงในภายหลัง โดยมีโหมดการหลั่งหลายแบบร่วมกับรูขุมขน การวิเคราะห์เปรียบเทียบต้นกำเนิดวิวัฒนาการของส่วนประกอบในน้ำนมสนับสนุนสถานการณ์ที่สารคัดหลั่งจากต่อมเหล่านี้วิวัฒนาการไปเป็นน้ำนมที่ซับซ้อนและอุดมไปด้วยสารอาหารมานานก่อนที่สัตว์เลี้ยงลูกด้วยนมที่แท้จริงจะเกิดขึ้น (โดยส่วนประกอบบางอย่างอาจมีมาก่อนการแยกสายพันธุ์ระหว่างไซแนปซิดและซอรอปซิด) ไซโนดอนต์เกือบจะสามารถผลิตสิ่งนี้ได้อย่างแน่นอน ซึ่งทำให้มวลของไข่แดงและขนาดของไข่ลดลงอย่างต่อเนื่อง ส่งผลให้ ลูกอ่อนที่ฟักออกมา มีลักษณะช่วยเหลือตัวเอง น้อยลงเรื่อยๆ เนื่องจากน้ำนมกลายเป็นแหล่งโภชนาการหลัก ซึ่งทั้งหมดนี้เห็นได้จากขนาดตัวที่เล็ก การมีกระดูกเอ พิพิวบิก และ การเปลี่ยนฟันที่จำกัดในไซโนดอนต์ขั้นสูง เช่นเดียวกับในสัตว์เลี้ยงลูกด้วยนม[ 52 ] [ 55 ]
วงจรการเผาผลาญและกิจกรรม
มีการถกเถียงกันว่าภาวะเลือดอุ่นเกิดขึ้นเมื่อใดในสายพันธุ์ซินาปซิด บางคนเสนอว่ามันเกิดขึ้นในบรรพบุรุษร่วมของซินาปซิด[ 56 ]ในขณะที่บางคนเสนอว่าภาวะเลือดอุ่นมีอยู่เฉพาะในสมาชิกของเทราปซิดา (อาจมีต้นกำเนิดแยกกันในไดซิโนดอนต์และซิโนดอนต์[ 57 ] ) [ 58 ]และซินาปซิดมีบรรพบุรุษเป็นสัตว์เลือดเย็นเหมือนสัตว์เลื้อยคลาน[ 59 ]
จากขนาดของเบ้าตาและวงแหวนสเคลรา คาด ว่าไซแนปซิดที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนมบางชนิดน่าจะเป็น สัตว์ หากินกลางวัน (ออกหากินในเวลากลางวัน) ในขณะที่บางชนิดน่าจะเป็นสัตว์หากินกลางคืน[ 60 ]
ประวัติวิวัฒนาการ


ตลอดช่วงวิวัฒนาการของซินาปซิด กลุ่มบรรพบุรุษในช่วงเริ่มต้นของการแผ่ขยายแบบปรับตัวมักจะเป็นสัตว์กินเนื้อที่พัฒนาแล้ว การแผ่ขยายแบบปรับตัวของซินาปซิดโดยทั่วไปเกิดขึ้นหลังจากเหตุการณ์การสูญพันธุ์ที่ทำให้ชีวภาคหมดไปและทิ้งช่องว่างไว้ให้กลุ่มสิ่งมีชีวิตที่วิวัฒนาการใหม่เข้ามาเติมเต็ม ในกลุ่มซินาปซิดที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนม กลุ่มสิ่งมีชีวิตที่ก่อให้เกิดสายพันธุ์ที่มีความหลากหลายอย่างรวดเร็วมีทั้งขนาดเล็กและขนาดใหญ่ แม้ว่าหลังจากยุคไทรแอสสิกตอนปลาย บรรพบุรุษของสายพันธุ์ซินาปซิดใหม่โดยทั่วไปจะมีขนาดเล็กและเป็นสัตว์กินเนื้อทั่วไปที่ไม่เฉพาะเจาะจง[ 61 ]
ซินาปซิดที่เก่าแก่ที่สุดที่ระบุได้อย่างชัดเจนนั้นพบใน ยุค Moscovianของยุคคาร์บอนิเฟอรัสตอนปลาย ประมาณ 315-307 ล้านปีก่อน พบได้ทั้งในอเมริกาเหนือและยุโรป รวมถึงArchaeothyris และEchinerpeton (ทั้งคู่เป็น ophiaconodontids ) เช่นเดียวกับDendromaia ( varanopid ) แม้ว่า จะมีฟอสซิล Bashkirianที่อาจเก่ากว่า(~315-320 ล้านปีก่อน) ที่ไม่ชัดเจนซึ่งพบในJoggins Formationของแคนาดา[ 15 ] ซินาปซิด " pelycosaurian " ยุคแรกแพร่กระจายและมีความหลากหลาย กลายเป็นนักล่าบนบกที่โดดเด่นและสัตว์บกที่ใหญ่ที่สุดในช่วงปลายยุคคาร์บอนิเฟอรัสและต้นยุคเพอร์เมียนมีความยาวได้ถึง 6 เมตร (20 ฟุต) พวกมันแผ่ขยายออกไป มีขนาดใหญ่ อาจเป็นสัตว์เลือดเย็น และมีสมองขนาดเล็ก บางชนิด เช่นDimetrodonมีครีบขนาดใหญ่ที่อาจช่วยเพิ่มอุณหภูมิร่างกายได้ กลุ่ม ที่หลงเหลืออยู่เพียงไม่กี่กลุ่มยังคงอยู่รอดมาได้จนถึงช่วงปลายยุคเพอร์เมียน แต่เมื่อถึงกลางยุคเพอร์เมียนตอนปลาย กลุ่มทั้งหมดก็สูญพันธุ์ไปหรือวิวัฒนาการกลายเป็นผู้สืบทอดคือเทอราปซิด[ 62 ]

สัตว์กลุ่มเทอราปซิด (Therapsids) ซึ่งเป็นกลุ่มไซแนปซิด (Synapsids) ที่พัฒนาแล้ว ปรากฏขึ้นในช่วงยุคเพอร์เมียนตอนกลางและรวมถึงสัตว์บกขนาดใหญ่ที่สุดในยุคเพอร์เมียนตอนกลางและตอนปลายพวกมันมีทั้งสัตว์กินพืชและสัตว์กินเนื้อ ตั้งแต่สัตว์ขนาดเล็กเท่าหนู (เช่นโรเบอร์เทีย ) ไปจนถึงสัตว์กินพืชขนาดใหญ่และเทอะทะ น้ำหนักหนึ่งตันหรือมากกว่า (เช่นมอสชอปส์ ) หลังจากเจริญรุ่งเรืองมาหลายล้านปี สัตว์ที่ประสบความสำเร็จเหล่านี้ก็เกือบสูญพันธุ์ไปในการสูญพันธุ์ครั้งใหญ่ในยุคเพอร์เมียน-ไทรแอสสิกเมื่อประมาณ 250 ล้านปีก่อน ซึ่งเป็นการสูญพันธุ์ครั้งใหญ่ที่สุดที่รู้จักในประวัติศาสตร์โลกอาจเกี่ยวข้องกับ เหตุการณ์ภูเขาไฟ ไซบีเรียนแทร ปส์ มีเพียงเทอราปซิดไม่กี่ชนิดเท่านั้นที่ประสบความสำเร็จในสภาพแวดล้อมใหม่ของยุคไทรแอสสิก ตอนต้น พวกมันได้แก่ลิสโทรซอ รัส (Lystrosaurus) และไซโนกนาทัส (Cynognathus)ซึ่งชนิดหลังปรากฏขึ้นในภายหลังในยุคไทรแอสสิกตอนต้น อย่างไรก็ตาม พวกมันก็อยู่ร่วมกับอาร์โคซอร์ ยุคแรก (ซึ่งต่อมาได้วิวัฒนาการเป็นไดโนเสาร์ ) ที่เข้ามายึดครองพื้นที่สำคัญหลังจากเหตุการณ์การแบ่งเขตยุคสมิธเทียน-สปาเธียน อาร์โคซอร์บางชนิด เช่นยูปาร์คเคียมีขนาดเล็กและโครงสร้างไม่ซับซ้อน ในขณะที่บางชนิด เช่นเอริโทรซูคัสมีขนาดใหญ่เท่าหรือใหญ่กว่าเทอแรปซิดที่ใหญ่ที่สุด
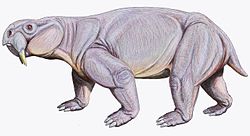
หลังจากการสูญพันธุ์ในยุคเพอร์เมียน ไซแนปซิดส์เหลือรอดอยู่ไม่เกินสามกลุ่ม[ 63 ]กลุ่มแรกประกอบด้วยเทโรเซฟาเลียนซึ่งมีชีวิตอยู่เพียง 20 ล้านปีแรกของยุคไทรแอสสิก กลุ่มที่สองเป็นสัตว์กินพืชที่มีจงอยปากเฉพาะทางที่รู้จักกันในชื่อไดไซโนดอนต์ (เช่นKannemeyeriidae ) ซึ่งมีสมาชิกบางตัวที่มีขนาดใหญ่ (หนักถึงหนึ่งตันหรือมากกว่า) และสุดท้ายคือไซโนดอนต์ที่กินเนื้อ กินพืช และกินแมลงที่มีลักษณะคล้ายสัตว์เลี้ยงลูกด้วยนมมากขึ้นเรื่อยๆ รวมถึงยูไซโนดอนต์จาก ยุค โอเลเนเคียนซึ่งตัวแทนในยุคแรกๆ คือไซโนกนาทัส

แตกต่างจากไดซิโนดอนต์ซึ่งมีขนาดใหญ่ ซิโนดอนต์กลับมีขนาดเล็กลงและมีลักษณะคล้ายสัตว์เลี้ยงลูกด้วยนมมากขึ้นเรื่อย ๆ ในช่วงยุคไทรแอสสิก แม้ว่าบางรูปแบบเช่นทรูซิโดซิโนดอนจะยังคงมีขนาดใหญ่ สัตว์เลี้ยงลูกด้วยนมกลุ่มแรกวิวัฒนาการมาจากซิโนดอนต์ในช่วงต้น ยุค นอเรียนของยุคไทรแอสสิกตอนปลาย ประมาณ 225 ล้านปีก่อน
ในลำดับวิวัฒนาการจากเทอแรปซิดยุคแรกไปสู่ไซโนดอนต์ ยูไซโนดอนต์ และสัตว์เลี้ยงลูกด้วยนม กระดูกขากรรไกรล่างหลัก หรือเดนทารี ได้เข้ามาแทนที่กระดูกข้างเคียง ดังนั้น ขากรรไกรล่างจึงค่อยๆ กลายเป็นกระดูกชิ้นใหญ่เพียงชิ้นเดียว โดยกระดูกขากรรไกรขนาดเล็กหลายชิ้นได้เคลื่อนย้ายเข้าไปในหูชั้นในทำให้เกิดการได้ยินที่ซับซ้อนขึ้น
บรรพบุรุษของสัตว์เลี้ยงลูกด้วยนมในยุคไทรแอสสิกและจูราสสิก รวมถึงญาติใกล้ชิดของพวกมัน มีอัตราการเผาผลาญสูง ซึ่งหมายความว่าพวกมันต้องบริโภคอาหาร (โดยทั่วไปคิดว่าเป็นแมลง) ในปริมาณที่มากกว่ามาก เพื่ออำนวยความสะดวกในการย่อยอาหาร อย่างรวดเร็ว ไซแนปซิดเหล่านี้จึงวิวัฒนาการการเคี้ยวและฟันที่พิเศษซึ่งช่วยในการเคี้ยว แขนขายังวิวัฒนาการให้เคลื่อนที่ใต้ลำตัวแทนที่จะไปด้านข้าง ทำให้พวกมันหายใจได้อย่างมีประสิทธิภาพมากขึ้นในระหว่างการเคลื่อนที่[ 64 ]สิ่งนี้ช่วยให้พวกมันสามารถรองรับความต้องการการเผาผลาญที่สูงขึ้นได้
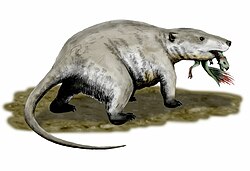
ไม่ว่าจะเกิดจากการเปลี่ยนแปลงสภาพภูมิอากาศ การเปลี่ยนแปลงของพืชพรรณ การแข่งขันทางนิเวศวิทยา หรือปัจจัยหลายอย่างรวมกัน สัตว์จำพวกไซโนดอนต์ขนาดใหญ่ที่เหลืออยู่ส่วนใหญ่ (อยู่ในวงศ์Traversodontidae ) และไดไซโนดอนต์ (อยู่ในวงศ์ Kannemeyeriidae) ได้สูญพันธุ์ไปแล้วใน ยุค ไรเทียนแม้กระทั่งก่อนเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในยุคไทรแอสสิก-จูราสสิกที่คร่าชีวิตอาร์โคซอร์ขนาดใหญ่ที่ไม่ใช่ไดโนเสาร์ ส่วนใหญ่ไป สัตว์จำพวก ไซแนปซิดในยุคมีโซโซอิกที่เหลืออยู่มีขนาดเล็ก ตั้งแต่ขนาดเท่าหนูชรูว์ไปจนถึงสัตว์เลี้ยงลูกด้วยนมคล้ายแบดเจอร์อย่าง Repenomamus

ในช่วงยุคจูราสสิกและครีเทเชียส ไซโนดอนต์ที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนมที่เหลืออยู่มีขนาดเล็ก เช่นไทรทิโลดอน (Tritylodon ) ไม่มีไซโนดอนต์ตัวใดใหญ่ไปกว่าแมว ไซโนดอนต์ส่วนใหญ่ในยุคจูราสสิกและครีเทเชียส กินพืช เป็นอาหารแม้ว่าบางชนิดจะกินเนื้อเป็นอาหาร ก็ตาม วงศ์ ไทรเท เลด อนทิดี (Tritheledontidae ) ซึ่งปรากฏตัวครั้งแรกในช่วงปลายยุคไทรแอสสิก เป็นสัตว์กินเนื้อและดำรงอยู่จนถึง ยุค จูราสสิกตอนกลางส่วนวงศ์ ไทรทิโลดอนทิดี (Tritylodontidae ) ปรากฏตัวครั้งแรกในช่วงเวลาเดียวกับไทรเทเลดอนต์ แต่เป็นสัตว์กินพืช กลุ่มนี้สูญพันธุ์ไปในช่วงปลายยุคครีเทเชียสตอนต้น โดยทั่วไปเชื่อกันว่าไดไซโนดอนต์สูญพันธุ์ไปในช่วงปลายยุคไทรแอสสิก แต่มีหลักฐานว่ากลุ่มนี้ยังคงมีชีวิตอยู่ โดยพบชิ้นส่วนกระดูกฟอสซิล 6 ชิ้นในหินยุคครีเทเชียสของรัฐควีนส์แลนด์ ประเทศออสเตรเลีย[ 65 ]หากเป็นจริง นั่นหมายความว่ามีสายพันธุ์ไดซิโนดอนต์ที่หลงเหลืออยู่ จำนวนมาก ในกอนด์วานาอย่างไรก็ตาม ฟอสซิลเหล่านี้ได้รับการอธิบายใหม่ในปี 2019 ว่ามีอายุ ในยุค ไพลสโตซีนและอาจเป็นของสัตว์มีถุงหน้าท้องไดโปรโตดอน ทิด[ 66 ]
ปัจจุบัน สัตว์เลี้ยงลูกด้วยนม ที่ยังมีชีวิตอยู่ประมาณ 5,500 ชนิด ประกอบด้วยทั้งสัตว์น้ำ ( วาฬและโลมา ) และสัตว์ปีก (ค้างคาว) รวมถึงสัตว์ที่ใหญ่ที่สุดเท่าที่เคยมีมา ( วาฬสีน้ำเงิน ) มนุษย์ก็เป็นสัตว์ในกลุ่มซินาปซิดเช่นกัน สัตว์เลี้ยง ลูกด้วย นมส่วนใหญ่เป็นสัตว์ออกลูกเป็นตัว ยกเว้นสัตว์ในกลุ่มโมโน ทรีม
ความสัมพันธ์
ด้านล่างนี้คือแผนภูมิวิวัฒนาการ ของไซแนปซิด ที่ได้รับการยอมรับมากที่สุดซึ่งแสดงให้เห็นสายบรรพบุรุษที่ยาว รวมถึง Mammalia และกลุ่มย่อยที่อยู่ฐานกว่า เช่น Theriodontia, Therapsida และ Sphenacodontia: [ 67 ] [ 68 ]
| ซินาปซิดา | |
ความไม่แน่นอนส่วนใหญ่ในแผนภูมิวิวัฒนาการของไดโนเสาร์กลุ่มซินาปซิดนั้น อยู่ในกลุ่มสมาชิกยุคแรกๆ รวมถึงรูปแบบที่เคยถูกจัดอยู่ในกลุ่มเพลิโคซอเรีย (Pelycosauria) มาก่อน ในการวิเคราะห์แผนภูมิวิวัฒนาการที่เก่าแก่ที่สุดครั้งหนึ่ง บริงค์แมนและอีเบิร์ธ (1983) ได้จัดให้วงศ์วาราโนปิเด (Varanopidae) อยู่ ร่วมกับ เคซาซอเรี ย (Caseasauria) ในฐานะกลุ่มที่แตกแขนงออกมาจากกลุ่มซินาปซิดในระดับพื้นฐานที่สุด ต่อมา ไรซ์ (1986) ได้แยกวงศ์วาราโนปิเดออกจากเคซาซอเรีย และจัดให้อยู่ในตำแหน่งที่พัฒนาแล้วมากกว่าบนลำต้นของแผนภูมิวิวัฒนาการ แม้ว่าการวิเคราะห์ส่วนใหญ่พบว่าเคซาซอเรียเป็นกลุ่มซินาปซิดพื้นฐานที่สุด แต่การวิเคราะห์ของเบนสัน (2012) กลับจัดให้กลุ่มที่มีวงศ์โอ ฟิ อาโคดอนทิเด (Ophiacodontidae ) และวาราโนปิเด (Varanopidae) เป็นกลุ่มซินาปซิดพื้นฐานที่สุด โดยมีเคซาซอเรียอยู่ในตำแหน่งที่พัฒนาแล้วมากกว่า เบนสันให้เหตุผลว่าการแก้ไขแผนภูมิวิวัฒนาการนี้เกิดจากการรวมลักษณะของโครงกระดูกส่วนอื่นๆ นอกเหนือจากกะโหลกศีรษะ (postcramental characteristics) เข้ามาในการวิเคราะห์ของเขาด้วย เมื่อรวมเฉพาะลักษณะกะโหลกหรือกะโหลกเท่านั้น Caseasauria ยังคงเป็นกลุ่ม synapsid พื้นฐานที่สุด ด้านล่างนี้คือแผนภูมิวิวัฒนาการที่ดัดแปลงมาจากการวิเคราะห์ของ Benson (2012): [ 69 ]
อย่างไรก็ตาม การตรวจสอบวิวัฒนาการของซินาปซิดพื้นฐานเมื่อไม่นานมานี้ ซึ่งรวมถึงเคซิดพื้นฐานและอีโอไทริดิดที่เพิ่งได้รับการอธิบายใหม่[ 70 ]ทำให้ Caseasauria กลับมาอยู่ในตำแหน่งที่เป็นพี่น้องกับซินาปซิดอื่นๆ ทั้งหมด Brocklehurst et al. (2016) [ 70 ]แสดงให้เห็นว่าลักษณะส่วนหลังกะโหลกหลายอย่างที่ Benson (2012) ใช้ในการรวม Caseasauria กับSphenacodontidaeและEdaphosauridaeนั้นไม่มีอยู่ในวัสดุส่วนหลังกะโหลกของอีโอไทริดิดที่เพิ่งค้นพบใหม่ ดังนั้นจึงได้มาจากการวิวัฒนาการแบบลู่เข้า
ดูเพิ่มเติม
- ยูริยาปซิดา – กลุ่มสัตว์เลื้อยคลานที่สูญพันธุ์ไปแล้ว
- รายชื่อสัตว์เลี้ยงลูกด้วยนมยุคก่อนประวัติศาสตร์
- รายชื่อไซแนปซิด
- การจำแนกประเภทสัตว์เลี้ยงลูกด้วยนม – อนุกรมวิธานของสัตว์เลี้ยงลูกด้วยนม
- ลำดับเหตุการณ์วิวัฒนาการของสิ่งมีชีวิต
- บรรพชีวินวิทยาของสัตว์มีกระดูกสันหลัง – การศึกษาทางวิทยาศาสตร์เกี่ยวกับสัตว์มีกระดูกสันหลังในยุคก่อนประวัติศาสตร์
หมายเหตุ
- ^กรีก : συν- ,โรมันไนซ์ : syn- ,แปลตรงตัวว่า ' ด้วยกัน' + ἁψίς ( apsís , 'ซุ้มประตู') > * συναψίδης ( synapsídes ) "มีซุ้มประตูที่เชื่อมติดกัน"; มีความหมายเหมือนกับเทอรอปซิดส์ (กรีก, "หน้าสัตว์ร้าย")
อ่านเพิ่มเติม
- โคลเบิร์ต, อีเอช (1969). วิวัฒนาการของสัตว์มีกระดูกสันหลัง (ฉบับที่ 2). นิวยอร์ก: จอห์น ไวลีย์ แอนด์ ซันส์ อิงค์ . ISBN 0-471-16466-6.
ลิงก์ภายนอก
- Synapsida - Pelycosauria - ที่ Palaeos
- ฟอสซิลสัตว์มีกระดูกสันหลังช่วงเปลี่ยนผ่าน - รวมถึงคำอธิบายของสกุลสำคัญ ในช่วง เปลี่ยนผ่านในลำดับวิวัฒนาการที่เชื่อมโยงไซแนปซิดดั้งเดิมกับสัตว์เลี้ยงลูกด้วยนม
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ซินาปซิดา
Synapsida [ a ] เป็นกลุ่ม สัตว์มีกระดูกสันหลัง สี่ขา ที่มีความหลากหลาย ซึ่งรวมถึง สัตว์เลี้ยงลูกด้วยนม ทั้งหมด และญาติที่สูญพันธุ์ไปแล้ว เป็นหนึ่งในสอง กลุ่ม หลัก ของกลุ่ม Amniota...
การจำแนกประเภทแบบลินเนียนและแบบคลาดิสติก
ในช่วงต้นศตวรรษที่ 20 เชื่อกันว่าไซแนปซิดเป็นหนึ่งในสี่กลุ่มย่อยหลักของ สัตว์เลื้อยคลาน อย่างไรก็ตาม ความคิดนี้ถูกหักล้างเมื่อตรวจสอบซากโครงกระดูกอย่างละเอียด เนื่องจากไซแนปซิดแตกต่างจากสัตว์เลื้อยคลานตรงที่มีช่องเปิดบริเวณขมับที่โดดเด่น ช่องเปิดเหล่านี้ใน...
ไซแนปซิดดั้งเดิมและไซแนปซิดขั้นสูง
ตามธรรมเนียมแล้ว ซินาปซิดจะถูกแบ่งออกเป็น กลุ่ม เทราปซิด ซึ่งเป็นกลุ่มซินาปซิดขั้นสูงและเป็นสาขาที่สัตว์เลี้ยงลูกด้วยนมวิวัฒนาการขึ้นมา และสัตว์เลี้ยงลูกด้วยนมดั้งเดิม (เดิมเรียกว่า เพลิโคซอร์ ) ซึ่งประกอบด้วยซินาปซิดอีก 6 วงศ์ที่ดั้งเดิมกว่า [ 22 ]...
การใช้คำว่า "synapsid" และ "therapsid" ในเชิงปฏิบัติและเชิงวิวัฒนาการ
ใน ระบบการตั้งชื่อทางวิวัฒนาการ คำเหล่านี้ถูกใช้แตกต่างออกไปเล็กน้อย เนื่องจากมีการรวมกลุ่มย่อยที่แตกแขนงออกมาด้วย บทความส่วนใหญ่ที่ตีพิมพ์ในช่วงศตวรรษที่ 21 ถือว่า "Pelycosaur" เป็นกลุ่มที่ไม่เป็นทางการของสมาชิกดั้งเดิม ในขณะที่ Therapsida...