อ่าน 18 นาที
การมองเห็นในปลา
การมองเห็นเป็นระบบประสาทสัมผัส ที่สำคัญสำหรับ ปลาส่วนใหญ่ดวงตา ของ ปลามีลักษณะคล้ายกับดวงตาของสัตว์มีกระดูกสันหลังบนบก เช่นนกและสัตว์เลี้ยงลูกด้วยนมแต่มีเลนส์ที่มีรูปทรงกลม...
การมองเห็นในปลา

การมองเห็นเป็นระบบประสาทสัมผัส ที่สำคัญสำหรับ ปลาส่วนใหญ่ดวงตา ของ ปลามีลักษณะคล้ายกับดวงตาของสัตว์มีกระดูกสันหลังบนบก เช่นนกและสัตว์เลี้ยงลูกด้วยนมแต่มีเลนส์ที่มีรูปทรงกลม มากกว่า นกและสัตว์เลี้ยงลูกด้วยนม (รวมถึงมนุษย์) โดยปกติจะปรับโฟกัสโดยการเปลี่ยนรูปร่างของเลนส์ แต่ปลาโดยปกติจะปรับโฟกัสโดยการขยับเลนส์ให้เข้าใกล้หรือห่างจากเรตินาเรตินาของปลาโดยทั่วไปมีทั้งเซลล์แท่งและเซลล์กรวย (สำหรับ การมองเห็น ในที่มืดและที่สว่าง ) และปลาส่วนใหญ่ สามารถมองเห็นสีได้ ปลาบางชนิดสามารถมองเห็นรังสีอัลตราไวโอเลตและบางชนิดไวต่อแสงโพลาไรซ์
ในบรรดาปลาที่ไม่มีขากรรไกรปลาแลมเพรย์[ 1 ] มีดวงตาที่พัฒนาแล้ว ในขณะที่ปลาแฮกฟิชมีเพียงจุดรับแสงแบบดั้งเดิม[ 2 ]บรรพบุรุษของปลาแฮกฟิชในปัจจุบัน ซึ่งเชื่อกันว่าเป็นโปรโตเวอร์เทเบรต[ 3 ]เห็นได้ชัดว่าถูกผลักดันไปยังน้ำลึกและมืดมาก ซึ่งพวกมันมีความเสี่ยงต่อผู้ล่าที่มีสายตาน้อยกว่า และการมีจุดรับแสงแบบนูนเป็นประโยชน์ เพราะสามารถรวบรวมแสงได้มากกว่าแบบแบนหรือเว้า การมองเห็นของปลาแสดงให้เห็นถึงการปรับ ตัวทางวิวัฒนาการ ให้เข้ากับสภาพแวดล้อมทางสายตา ตัวอย่างเช่นปลาทะเลน้ำลึกมีดวงตาที่เหมาะสมกับสภาพแวดล้อมที่มืด
น้ำในฐานะสภาพแวดล้อมทางสายตา


ปลาและสัตว์น้ำอื่นๆ อาศัยอยู่ในสภาพแวดล้อมที่มีแสงแตกต่างจากสัตว์บกน้ำดูดซับแสงดังนั้นเมื่อความลึกเพิ่มขึ้น ปริมาณแสงที่มีอยู่จะลดลงอย่างรวดเร็ว คุณสมบัติทางแสงของน้ำยังส่งผลให้มี การดูดซับแสงที่ มีความยาวคลื่น ต่างกัน ในระดับที่แตกต่างกัน ตัวอย่างเช่น แสงที่มองเห็นได้ซึ่งมีความยาวคลื่นยาว (เช่น สีแดง สีส้ม) จะถูกดูดซับในน้ำมากกว่าแสงที่มีความยาวคลื่นสั้นกว่า (สีเขียว สีน้ำเงิน) แสงอัลตราไวโอเลต (ซึ่งมีความยาวคลื่นสั้นกว่าสีม่วง) สามารถทะลุทะลวงได้ลึกกว่าสเปกตรัมของแสงที่มองเห็นได้[ 5 ]นอกจากคุณสมบัติทั่วไปของน้ำเหล่านี้แล้ว แหล่งน้ำต่างๆ อาจดูดซับแสงที่มีความยาวคลื่นต่างกันเนื่องจากปริมาณเกลือและ/หรือสารเคมีในน้ำที่แตกต่างกัน
น้ำมีประสิทธิภาพมากในการดูดซับแสงที่เข้ามา ดังนั้นปริมาณแสงที่ทะลุผ่านมหาสมุทรจึงลดลงอย่างรวดเร็ว (ถูกลดทอน) ตามความลึก ในน้ำทะเลใส ที่ความลึก 1 เมตร พลังงานแสงอาทิตย์ที่ตกกระทบผิวมหาสมุทรจะเหลืออยู่เพียง 45% ที่ความลึก 10 เมตร จะเหลือแสงอยู่เพียง 16% และที่ความลึก 100 เมตร จะเหลือแสงเพียง 1% ของแสงเดิม ไม่มีแสงทะลุผ่านเกิน 1000 เมตร[ 6 ]
นอกจากการลดทอนโดยรวมแล้ว มหาสมุทรยังดูดซับความยาวคลื่นแสงที่แตกต่างกันในอัตราที่แตกต่างกัน ความยาวคลื่นที่ปลายสุดของสเปกตรัมที่มองเห็นได้จะถูกลดทอนเร็วกว่าความยาวคลื่นตรงกลาง ความยาวคลื่นที่ยาวกว่าจะถูกดูดซับก่อน ในน้ำทะเลใส สีแดงจะถูกดูดซับที่ระดับความลึก 10 เมตร สีส้มที่ระดับความลึกประมาณ 40 เมตร และสีเหลืองจะหายไปก่อนระดับความลึก 100 เมตร ความยาวคลื่นที่สั้นกว่าจะทะลุทะลวงได้ไกลกว่า โดยแสงสีน้ำเงินและสีเขียวจะไปถึงระดับความลึกที่ลึกที่สุด[ 6 ]นี่คือเหตุผลที่สิ่งต่างๆ ปรากฏเป็นสีน้ำเงินใต้น้ำ: การรับรู้สีของดวงตาขึ้นอยู่กับความยาวคลื่นของแสงที่ดวงตาได้รับ วัตถุปรากฏเป็นสีแดงต่อสายตาเพราะมันสะท้อนแสงสีแดงและดูดซับสีอื่นๆ ดังนั้นสีเดียวที่ไปถึงดวงตาคือสีแดง สีน้ำเงินเป็นสีของแสงเพียงสีเดียวที่มีอยู่ที่ระดับความลึกใต้น้ำ ดังนั้นจึงเป็นสีเดียวที่สามารถสะท้อนกลับไปยังดวงตาได้ และทุกสิ่งจึงมีสีน้ำเงินเจือปนอยู่ใต้น้ำ วัตถุสีแดงที่ระดับความลึกจะไม่ปรากฏเป็นสีแดงเพราะไม่มีแสงสีแดงให้สะท้อนจากวัตถุ วัตถุในน้ำจะปรากฏเป็นสีจริงเฉพาะบริเวณใกล้ผิวน้ำที่ยังมีคลื่นแสงทุกความยาวคลื่นอยู่ หรือหากมีการให้คลื่นแสงความยาวคลื่นอื่น ๆ ด้วยวิธีเทียม เช่น การส่องสว่างวัตถุด้วยไฟฉายดำน้ำ[ 6 ]
โครงสร้างและหน้าที่


ดวงตาของปลาโดยทั่วไปคล้ายคลึงกับดวงตาของสัตว์มีกระดูกสันหลังอื่นๆ โดยเฉพาะอย่างยิ่งสัตว์สี่ขา (สัตว์ครึ่งบกครึ่งน้ำ สัตว์เลื้อยคลาน นก และสัตว์เลี้ยงลูกด้วยนม ซึ่งทั้งหมดวิวัฒนาการมาจากบรรพบุรุษที่เป็นปลา) แสงเข้าสู่ดวงตาที่กระจกตาผ่านรูม่านตาไปยังเลนส์ปลาส่วนใหญ่ดูเหมือนจะมีขนาดรูม่านตาคงที่ แต่ ปลา ฉลามและปลากระเบนมีม่านตา ที่เป็นกล้ามเนื้อ ซึ่งช่วยให้สามารถปรับขนาดเส้นผ่านศูนย์กลางของรูม่านตาได้ รูปร่างของรูม่านตาแตกต่างกันไป และอาจเป็นวงกลมหรือเป็นร่อง[ 5 ]
เลนส์โดยทั่วไปมีรูปทรงกลม แต่ในบางชนิดอาจมีรูปวงรีเล็กน้อย เมื่อเปรียบเทียบกับสัตว์มีกระดูกสันหลังบนบก เลนส์ของปลาโดยทั่วไปจะมีความหนาแน่นและมีรูปทรงกลมมากกว่า ในสภาพแวดล้อมทางน้ำดัชนีหักเหของกระจกตาและน้ำโดยรอบจะไม่แตกต่างกันมากนัก (เมื่อเทียบกับอากาศบนบก) ดังนั้นเลนส์จึงต้องทำหน้าที่หักเหแสงเป็นส่วนใหญ่[ 7 ]เนื่องมาจาก " การไล่ระดับ ดัชนีหักเหภายในเลนส์ — ซึ่งเป็นไปตามที่คาดหวังจากทฤษฎีทางแสง" [ 8 ]เลนส์ทรงกลมของปลาจึงสามารถสร้างภาพที่คมชัดปราศจากความคลาดเคลื่อนทรงกลมได้[ 7 ]
เมื่อแสงผ่านเลนส์แล้ว แสงจะถูกส่งผ่านตัวกลางที่เป็นของเหลวโปร่งใสจนกระทั่งถึงเรตินาซึ่งมีเซลล์รับแสง อยู่ เช่นเดียวกับสัตว์มีกระดูกสันหลังชนิดอื่นๆ เซลล์รับแสงจะอยู่บนชั้นใน ดังนั้นแสงจึงต้องผ่านชั้นของเซลล์ประสาท อื่นๆ ก่อนที่จะไปถึงเซลล์รับแสง เรตินาประกอบด้วยเซลล์รูปแท่งและเซลล์รูปกรวย[ 5 ] มีความคล้ายคลึงกันระหว่างดวงตาของปลาและสัตว์มีกระดูกสันหลังชนิดอื่นๆ โดยปกติแล้ว แสงจะเข้าสู่ดวงตาของปลาทางกระจกตาและผ่านรูม่านตาเพื่อไปถึงเลนส์ ปลาส่วนใหญ่มีรูม่านตาขนาดคงที่ ในขณะที่ปลาบางชนิดมีม่านตาที่เป็นกล้ามเนื้อซึ่งช่วยให้สามารถปรับขนาดเส้นผ่านศูนย์กลางของรูม่านตาได้
ดวงตาของปลาจะมีเลนส์ทรงกลมมากกว่าสัตว์มีกระดูกสันหลังบนบกชนิดอื่นๆ การปรับโฟกัสในสัตว์เลี้ยงลูกด้วยนมและนกมักทำได้โดยการเปลี่ยนรูปร่างของเลนส์ตา ในขณะที่ในปลาจะทำได้โดยการเคลื่อนเลนส์ให้ห่างจากหรือใกล้กับเรตินามากขึ้น เรตินาของปลาโดยทั่วไปจะมีทั้งเซลล์แท่งและเซลล์กรวยที่รับผิดชอบการมองเห็นในที่มืดและในที่สว่าง ปลาส่วนใหญ่สามารถมองเห็นสีได้ มีบางชนิดที่สามารถมองเห็นรังสีอัลตราไวโอเลตได้ ในขณะที่บางชนิดมีความไวต่อแสงโพลาไรซ์[ 9 ]
เรตินาของปลาประกอบด้วยเซลล์รูปแท่งที่ให้ความไวในการมองเห็นสูงในสภาพแสงน้อย และเซลล์รูปกรวยที่ให้ความละเอียดเชิงเวลาและเชิงพื้นที่สูงกว่าเซลล์รูปแท่ง ซึ่งทำให้สามารถมองเห็นสีได้โดยการเปรียบเทียบการดูดกลืนแสงระหว่างเซลล์รูปกรวยชนิดต่างๆ[ 10 ]ตามที่ Marshall และคณะกล่าวไว้ สัตว์ส่วนใหญ่ในแหล่งที่อยู่อาศัยในทะเลไม่มีหรือมีการมองเห็นสีที่ค่อนข้างง่าย อย่างไรก็ตาม การมองเห็นสีในมหาสมุทรมีความหลากหลายมากกว่าบนบก ซึ่งส่วนใหญ่เป็นผลมาจากสภาพแวดล้อมที่มีแสงและพฤติกรรมเกี่ยวกับสีที่แตกต่างกันอย่างมาก[ 11 ]
เรตินา
ภายในเรตินาเซลล์แท่งให้ความไวในการมองเห็นสูง (โดยแลกกับความคมชัด ) และใช้ในสภาพแสงน้อย เซลล์กรวยให้ความละเอียดเชิงพื้นที่และเวลาที่สูงกว่าเซลล์แท่ง และทำให้สามารถมองเห็นสีได้โดยการเปรียบเทียบการดูดกลืนแสงระหว่างเซลล์กรวยชนิดต่างๆ ซึ่งมีความไวต่อความยาวคลื่นที่แตกต่างกัน อัตราส่วนของเซลล์แท่งต่อเซลล์กรวยขึ้นอยู่กับระบบนิเวศของปลาแต่ละชนิดเช่นปลาที่หากินในเวลากลางวันในน้ำใสจะมีเซลล์กรวยมากกว่าปลาที่อาศัยอยู่ในสภาพแวดล้อมที่มีแสงน้อย การมองเห็นสีมีประโยชน์มากกว่าในสภาพแวดล้อมที่มีความยาวคลื่นให้เลือกหลากหลายเช่นใกล้ผิวน้ำในน้ำใส มากกว่าในน้ำลึกที่มีเพียงช่วงความยาวคลื่นแคบๆ เท่านั้น[ 5 ]
การกระจายตัวของเซลล์รับแสงทั่วเรตินาไม่สม่ำเสมอ บางบริเวณมีความหนาแน่นของเซลล์รูปกรวยสูงกว่า เช่น (ดูที่โฟเวีย ) ปลาอาจมีสองหรือสามบริเวณที่เชี่ยวชาญด้านความคมชัดสูง (เช่น สำหรับการจับเหยื่อ) หรือความไว (เช่น จากแสงสลัวที่มาจากด้านล่าง) การกระจายตัวของเซลล์รับแสงอาจเปลี่ยนแปลงไปตามกาลเวลาในระหว่างการพัฒนาของแต่ละตัว โดยเฉพาะอย่างยิ่งในกรณีที่สายพันธุ์มักจะเคลื่อนย้ายระหว่างสภาพแวดล้อมแสงที่แตกต่างกันในระหว่างวงจรชีวิต (เช่น จากน้ำตื้นไปน้ำลึก หรือจากน้ำจืดไปมหาสมุทร) [ 5 ]หรือเมื่อสเปกตรัมอาหารเปลี่ยนแปลงไปพร้อมกับการเจริญเติบโตของปลา ดังเช่นที่พบในปลาไอซ์ฟิชแอนตาร์กติกChampsocephalus gunnari [ 12 ]
บางชนิดมีtapetumซึ่งเป็นชั้นสะท้อนแสงที่สะท้อนแสงที่ผ่านเรตินาให้กลับผ่านเข้าไปอีกครั้ง ช่วยเพิ่มความไวในสภาวะแสงน้อย เช่น ในสัตว์กลางคืนและสัตว์ทะเลลึก โดยให้โอกาสโฟตอนถูกจับโดยตัวรับแสงได้อีกครั้ง[ 7 ] อย่างไรก็ตาม สิ่งนี้มาพร้อมกับข้อเสียคือความละเอียดลดลง บางชนิดสามารถปิด tapetum ได้อย่างมีประสิทธิภาพในสภาวะแสงสว่าง โดยใช้ชั้นเม็ดสีเข้มปกคลุมไว้เมื่อจำเป็น[ 5 ]
จอประสาทตาใช้ออกซิเจนจำนวนมากเมื่อเทียบกับเนื้อเยื่ออื่นๆ ส่วนใหญ่ และมีเลือดที่มีออกซิเจนเพียงพอหล่อเลี้ยงเพื่อให้มั่นใจว่าการทำงานเป็นไปอย่างเหมาะสม[ 5 ]
ที่พัก
การปรับโฟกัสคือกระบวนการที่ดวงตาของสัตว์มีกระดูกสันหลังปรับโฟกัสไปที่วัตถุเมื่อวัตถุเคลื่อนเข้าใกล้หรือไกลออกไป ในขณะที่นกและสัตว์เลี้ยงลูกด้วยนมปรับโฟกัสโดยการทำให้เลนส์ตาเสียรูป ปลาและสัตว์ครึ่งบก ครึ่งน้ำ มักจะปรับโฟกัสโดยการเคลื่อนเลนส์ให้เข้าใกล้หรือไกลจากเรตินา[ 5 ]พวกมันใช้กล้ามเนื้อพิเศษที่เปลี่ยนระยะห่างของเลนส์จากเรตินา ในปลาที่มีกระดูกกล้ามเนื้อนี้เรียกว่าretractor lentisและจะคลายตัวเมื่อมองใกล้ ในขณะที่ปลาที่มีกระดูกอ่อน กล้ามเนื้อนี้เรียกว่าprotractor lentisและจะคลายตัวเมื่อมองไกล ดังนั้นปลาที่มีกระดูกจึงปรับโฟกัสเมื่อมองไกลโดยการเคลื่อนเลนส์ให้เข้าใกล้เรตินา ในขณะที่ปลาที่มีกระดูกอ่อนปรับโฟกัสเมื่อมองใกล้โดยการเคลื่อนเลนส์ให้ไกลจากเรตินา[ 13 ] [ 14 ] [ 15 ]
การปรับภาพนิ่ง

มีความจำเป็นต้องมีกลไกบางอย่างที่ช่วยรักษาเสถียรภาพของภาพในระหว่างการเคลื่อนไหวศีรษะอย่างรวดเร็ว ซึ่งทำได้โดยรีเฟล็กซ์เวสติบูโล-โอคูลาร์ซึ่งเป็นการเคลื่อนไหวของดวงตา แบบ รีเฟล็กซ์ ที่ช่วยรักษาเสถียรภาพของภาพบนเรตินาโดยการสร้างการเคลื่อนไหวของดวงตาในทิศทางตรงกันข้ามกับการเคลื่อนไหวของศีรษะ จึงช่วยรักษาภาพไว้ที่กึ่งกลางของสนามการมองเห็น ตัวอย่างเช่น เมื่อศีรษะเคลื่อนไปทางขวา ดวงตาจะเคลื่อนไปทางซ้าย และในทางกลับกัน รีเฟล็กซ์เวสติบูโล-โอคูลาร์ของมนุษย์เป็นการเคลื่อนไหวของดวงตา แบบรีเฟล็ ก ซ์ ที่ช่วยรักษาเสถียรภาพของภาพบนเรตินาในระหว่างการเคลื่อนไหวของศีรษะโดยการสร้างการเคลื่อนไหวของดวงตาในทิศทางตรงกันข้ามกับการเคลื่อนไหวของศีรษะ จึงช่วยรักษาภาพไว้ที่กึ่งกลางของสนามการมองเห็น ในทำนองเดียวกัน ปลาจะมีรีเฟล็กซ์เวสติบูโล-โอคูลาร์ที่ช่วยรักษาเสถียรภาพของภาพบนเรตินาเมื่อมันขยับหาง[ 16 ] ในสัตว์หลายชนิด รวมถึงมนุษย์หูชั้นในทำหน้าที่เป็นอะนาล็อกทางชีวภาพของ มาตรวัด ความเร่งในระบบรักษาเสถียรภาพภาพของกล้อง เพื่อรักษาเสถียรภาพของภาพโดยการขยับดวงตาเมื่อตรวจพบการหมุนของศีรษะ สัญญาณยับยั้งจะถูกส่งไปยังกล้ามเนื้อนอกลูกตาข้างหนึ่ง และสัญญาณกระตุ้นจะถูกส่งไปยังกล้ามเนื้ออีกข้างหนึ่ง ผลที่ได้คือการเคลื่อนไหวของดวงตาเพื่อชดเชย การเคลื่อนไหวของดวงตาของมนุษย์โดยทั่วไปจะล่าช้ากว่าการเคลื่อนไหวของศีรษะไม่เกิน 10 มิลลิวินาที[ 17 ]
แผนภาพทางด้านขวาแสดง วงจร การสะท้อนกลับของระบบเวสติบูลาร์ต่อการเคลื่อนไหวของดวงตาใน แนวนอน ใน ปลา ที่มีกระดูกแข็งและปลาที่มีกระดูกอ่อน
- "Goldfish" แสดงให้เห็นรีเฟล็กซ์เวสติบูโล-โอคูลาร์สามเซลล์ประสาทหลักที่เชื่อมโยงท่อครึ่งวงกลมแนวนอนกับแอ็บดิวเซนส์ฝั่งตรงข้าม (ABD) และมอเตอร์นิวรอนMRฝั่งเดียวกัน[ 18 ]
- "Flatfish" แสดงให้เห็นว่าหลังจากการเคลื่อนที่ของระบบทรงตัว 90° เทียบกับแกนการมองเห็น (metamorphosis) การเคลื่อนไหวของดวงตาเพื่อชดเชยจะเกิดขึ้นโดยการเปลี่ยนทิศทางสัญญาณจากท่อแนวนอนไปยังเซลล์ประสาทสั่งการในแนวตั้งและแนวเฉียง[ 19 ] [ 20 ]
- ใน "Shark" เซลล์ประสาทคลองแนวนอน/ลำดับที่สองจะฉายไปยังเซลล์ประสาทสั่งการ ABD และ MR ฝั่งตรงข้าม รวมถึงเซลล์ประสาท AI ฝั่งเดียวกัน 1° เซลล์ประสาท เวสติบูลาร์ ลำดับแรก ; ATD เส้นทางขึ้น ของDeiter [ 20 ]
อัลตราไวโอเลต
การมองเห็นของปลาถูกควบคุมโดยรงควัตถุการมองเห็นสี่ชนิดที่ดูดซับความยาวคลื่นแสงต่างๆ รงควัตถุแต่ละชนิดสร้างขึ้นจากโครโมฟอร์และโปรตีนทรานส์เมมเบรนที่เรียกว่าออปซิน การกลายพันธุ์ในออปซินทำให้เกิดความหลากหลายในการมองเห็น รวมถึงความแปรผันในการดูดซับความยาวคลื่น[ 21 ]การกลายพันธุ์ของออปซินบนรงควัตถุ SWS-1 ทำให้สัตว์มีกระดูกสันหลังบางชนิดสามารถดูดซับแสง UV (≈360 นาโนเมตร) ได้ ดังนั้นพวกมันจึงสามารถมองเห็นวัตถุที่สะท้อนแสง UV ได้[ 22 ]ปลาหลากหลายสายพันธุ์ได้พัฒนาและรักษาลักษณะการมองเห็นนี้ไว้ตลอดวิวัฒนาการ ซึ่งบ่งชี้ว่ามันเป็นประโยชน์ การมองเห็น UV อาจเกี่ยวข้องกับการหาอาหาร การสื่อสาร และการเลือกคู่ครอง
ทฤษฎีหลักเกี่ยวกับการคัดเลือกเชิงวิวัฒนาการของการมองเห็นรังสียูวีในปลาบางชนิดเกิดจากบทบาทสำคัญในการเลือกคู่ครอง การทดลองทางพฤติกรรมแสดงให้เห็นว่าปลาหมอสีแอฟริกันใช้สัญญาณภาพในการเลือกคู่ครอง โดยทั่วไปแหล่งเพาะพันธุ์ของพวกมันอยู่ในน้ำตื้นที่มีความใสสูงและแสงยูวีสามารถส่องผ่านได้ ปลาหมอสีแอฟริกันตัวผู้ส่วนใหญ่มีสีฟ้าซึ่งสะท้อนแสงยูวี ตัวเมียสามารถเลือกคู่ครองสายพันธุ์เดียวกันได้อย่างถูกต้องเมื่อมีสัญญาณภาพที่สะท้อนแสงเหล่านี้อยู่ ซึ่งแสดงให้เห็นว่าการตรวจจับแสงยูวีมีความสำคัญต่อการเลือกคู่ครองที่ถูกต้อง[ 23 ]รูปแบบสีสะท้อนแสงยูวียังช่วยเพิ่มความน่าดึงดูดใจของตัวผู้ในปลาหางนกยูงและปลาหนามสามแฉกในการทดลอง ปลาหางนกยูงตัวเมียใช้เวลาตรวจสอบตัวผู้ที่มีสีสะท้อนแสงยูวีมากกว่าตัวผู้ที่มีสีที่ถูกปิดกั้นการสะท้อนแสงยูวีอย่างมีนัยสำคัญ[ 24 ]ในทำนองเดียวกัน ปลาหนามสามแฉกตัวเมียชอบตัวผู้ที่มองเห็นในสเปกตรัมเต็มรูปแบบมากกว่าตัวผู้ที่มองเห็นในตัวกรองที่ปิดกั้นรังสียูวี[ 25 ]ผลลัพธ์เหล่านี้ชี้ให้เห็นอย่างชัดเจนถึงบทบาทของการตรวจจับ UV ในการคัดเลือกทางเพศ และด้วยเหตุนี้จึงส่งผลต่อความเหมาะสมในการสืบพันธุ์ บทบาทที่โดดเด่นของการตรวจจับแสง UV ในการเลือกคู่ของ ปลา ทำให้ลักษณะดังกล่าวคงอยู่ได้ตลอดเวลา การมองเห็น UV อาจเกี่ยวข้องกับการหาอาหารและพฤติกรรมการสื่อสารอื่นๆ ด้วย
ปลาหลายชนิดสามารถมองเห็น รังสี อัลตราไวโอเลตในช่วงสเปกตรัมที่ไกลกว่าสีม่วงได้[ 26 ]
บางครั้งการมองเห็นด้วยรังสีอัลตราไวโอเลตจะใช้เฉพาะในช่วงใดช่วงหนึ่งของวงจรชีวิตของปลาเท่านั้น ตัวอย่างเช่นปลาเทราต์สีน้ำตาล วัยอ่อน อาศัยอยู่ในน้ำตื้นซึ่งพวกมันใช้การมองเห็นด้วยรังสีอัลตราไวโอเลตเพื่อเพิ่มความสามารถในการตรวจจับแพลงก์ตอนสัตว์เมื่อพวกมันโตขึ้น พวกมันจะย้ายไปยังน้ำที่ลึกกว่าซึ่งมีแสงอัลตราไวโอเลตน้อย[ 22 ]
ปลา แดมเซลสองแถบ Dascyllus reticulatus มีสีที่สะท้อนรังสีอัลตราไวโอเลตซึ่งดูเหมือนว่าจะใช้เป็นสัญญาณเตือนภัยสำหรับปลาชนิดเดียวกัน[ 27 ] ปลา ล่าเหยื่อไม่สามารถมองเห็นสิ่งนี้ได้หากการมองเห็นของพวกมันไม่ไวต่อรังสีอัลตราไวโอเลต มีหลักฐานเพิ่มเติมสำหรับมุมมองนี้ว่าปลาบางชนิดใช้รังสีอัลตราไวโอเลตเป็น "ช่องทางการสื่อสารลับที่มีความแม่นยำสูงซึ่งซ่อนจากผู้ล่า" ในขณะที่ปลาชนิดอื่นใช้รังสีอัลตราไวโอเลตเพื่อส่งสัญญาณทางสังคมหรือทางเพศ[ 5 ] [ 28 ]
แสงโพลาไรซ์
การจะระบุว่าปลาไวต่อ แสงโพลาไรซ์ หรือไม่ นั้นไม่ใช่เรื่องง่ายแม้ว่าจะดูเหมือนเป็นไปได้ในกลุ่มสิ่งมีชีวิตหลายกลุ่มก็ตาม มีการพิสูจน์อย่างชัดเจนแล้วในปลาแอนโชวี่ [ 29 ] ความ สามารถในการตรวจจับแสงโพลาไรซ์อาจให้ความคมชัดและ/หรือข้อมูลทิศทางที่ดีกว่าสำหรับปลาที่อพยพ แสงโพลาไรซ์มีมากที่สุดในตอนรุ่งเช้าและพลบค่ำ[ 5 ]แสงโพลาไรซ์ที่สะท้อนจากเกล็ดปลาอาจทำให้ปลาตัวอื่นตรวจจับปลาตัวนั้นได้ดีขึ้นเมื่อเทียบกับพื้นหลังที่กระจายแสง[ 30 ]และอาจให้ข้อมูลที่เป็นประโยชน์แก่ปลาที่อยู่รวมกันเป็นฝูงเกี่ยวกับความใกล้ชิดและการวางตัวของพวกมันเมื่อเทียบกับปลาตัวอื่นๆ ที่อยู่ใกล้เคียง[ 31 ]การทดลองบางอย่างบ่งชี้ว่า การใช้โพลาไรซ์ทำให้ปลาบางชนิดสามารถปรับการมองเห็นของพวกมันเพื่อให้มองเห็นเหยื่อได้ไกลเป็นสองเท่าของระยะปกติ[ 9 ]
กรวยคู่
ปลาส่วนใหญ่มีโคนคู่ซึ่งเป็นเซลล์โคนคู่ที่เชื่อมต่อกัน สมาชิกแต่ละตัวของโคนคู่อาจมีการดูดซับสูงสุดที่แตกต่างกัน และหลักฐานเชิงพฤติกรรมสนับสนุนแนวคิดที่ว่าโคนแต่ละประเภทในโคนคู่สามารถให้ข้อมูลแยกกันได้ (กล่าวคือ สัญญาณจากสมาชิกแต่ละตัวของโคนคู่ไม่จำเป็นต้องรวมกัน) [ 32 ]
การปรับตัวให้เข้ากับถิ่นที่อยู่





ปลาที่อาศัยอยู่ในน้ำผิวดินลึกประมาณ 200 เมตร หรือที่เรียกว่าปลาอีพิเพลาจิกอาศัยอยู่ในเขตที่มีแสงแดดส่องถึง ซึ่งผู้ล่าที่ใช้สายตาจะใช้ระบบการมองเห็นที่ออกแบบมาค่อนข้างเป็นไปตามที่คาดไว้ แต่ถึงกระนั้นก็อาจมีการปรับตัวที่ผิดปกติได้ปลาสี่ตาจะมีตาที่ยกสูงขึ้นเหนือส่วนบนของหัวและแบ่งออกเป็นสองส่วนที่แตกต่างกัน เพื่อให้สามารถมองเห็นทั้งใต้น้ำและเหนือน้ำได้ในเวลาเดียวกัน จริงๆ แล้วปลาสี่ตามีเพียงสองตา แต่ดวงตาของพวกมันได้รับการปรับให้เข้ากับวิถีชีวิตที่อาศัยอยู่บนผิวน้ำเป็นพิเศษ ดวงตาจะอยู่บนส่วนบนของหัว และปลาจะลอยอยู่บนผิวน้ำโดยมีเพียงครึ่งล่างของแต่ละตาอยู่ใต้น้ำ สองส่วนนี้ถูกแบ่งออกด้วยแถบเนื้อเยื่อ และดวงตามีรูม่านตา 2 รู ซึ่งเชื่อมต่อกันด้วยส่วนหนึ่งของม่านตาครึ่งบนของดวงตาได้รับการปรับให้เข้ากับการมองเห็นในอากาศ ครึ่งล่างสำหรับการมองเห็นในน้ำ[ 35 ]เลนส์ของดวงตาจะเปลี่ยนความหนาจากบนลงล่างเพื่อชดเชยความแตกต่างของดัชนีการหักเหของอากาศกับน้ำ ปลาเหล่านี้ใช้เวลาส่วนใหญ่อยู่ที่ผิวน้ำ อาหารของพวกมันส่วนใหญ่ประกอบด้วยแมลงบนบกที่หาได้ที่ผิวน้ำ[ 36 ]
ปลาเมโซเพลาจิกอาศัยอยู่ในน้ำลึก ในเขตแสงสลัวที่ระดับความลึกถึง 1,000 เมตร ซึ่งปริมาณแสงแดดที่มีอยู่ไม่เพียงพอที่จะสนับสนุนการสังเคราะห์แสงปลาเหล่านี้ปรับตัวให้มีชีวิตที่กระฉับกระเฉงภายใต้สภาพแสงน้อย ส่วนใหญ่เป็นนักล่าที่ใช้สายตาและมีดวงตาขนาดใหญ่ ปลาที่อาศัยอยู่ในน้ำลึกบางชนิดมีดวงตาเป็นท่อที่มีเลนส์ขนาดใหญ่และมีเซลล์รูปแท่งที่มองขึ้นด้านบนเท่านั้น ซึ่งทำให้มองเห็นแบบสองตาและมีความไวต่อสัญญาณแสงขนาดเล็กมาก[ 37 ]การปรับตัวนี้ทำให้การมองเห็นในระยะสุดท้ายดีขึ้นโดยแลกกับการมองเห็นด้านข้าง และช่วยให้นักล่าสามารถมองเห็นปลาหมึกหมึกกระดองและปลาขนาดเล็กที่ปรากฏเป็นเงาตัดกับความมืดด้านบนได้ เพื่อการมองเห็นที่ไวขึ้นในที่แสงน้อยปลาบางชนิดมีตัวสะท้อนแสงอยู่ด้านหลังเรตินาปลาไฟฉายมีสิ่งนี้รวมถึงอวัยวะเรือง แสง ซึ่งพวกมันใช้ร่วมกันเพื่อตรวจจับแสงสะท้อนจากดวงตาของปลาตัวอื่น[ 38 ] [ 39 ] [ 40 ]
ใน ระดับความลึกที่ลึกลงไปอีก 1,000 เมตร จะพบปลาบาธิเพลาจิกที่ระดับความลึกนี้ มหาสมุทรมืดสนิท และปลาเหล่านี้อยู่นิ่ง ปรับตัวให้ใช้พลังงานน้อยที่สุดในถิ่นที่อยู่ที่มีอาหารน้อยมากและไม่มีแสงแดดการเรืองแสงทางชีวภาพเป็นแสงเดียวที่มีในระดับความลึกเหล่านี้ การขาดแสงนี้หมายความว่าสิ่งมีชีวิตต้องพึ่งพาประสาทสัมผัสอื่นนอกเหนือจากการมองเห็น ดวงตาของพวกมันมีขนาดเล็กและอาจใช้งานไม่ได้เลย[ 41 ] [ 42 ]
ปลาแบนสามารถพบได้ที่ก้นมหาสมุทร ปลาแบนเป็น ปลาที่อาศัยอยู่ บนพื้นทะเล มีแรงลอยตัวเป็นลบ จึงสามารถพักอยู่บนพื้นทะเลได้ แม้ว่าปลาแบนจะอาศัยอยู่บนพื้นทะเล แต่โดยทั่วไปแล้วพวกมันไม่ใช่ปลาทะเลน้ำลึก แต่พบได้ส่วนใหญ่ในปากแม่น้ำและบนไหล่ทวีป เมื่อลูกปลา แบน ฟักออกมา พวกมันจะมีรูปร่างยาวและสมมาตรเหมือนปลาที่มีกระดูก ทั่วไป ลูกปลาไม่ได้อาศัยอยู่บนพื้นทะเล แต่ลอยอยู่ในทะเลเหมือนแพลงก์ตอน ในที่สุดพวกมันก็จะเริ่มเปลี่ยนแปลงรูปร่างไปเป็นตัวเต็มวัย ตาข้างหนึ่งจะเคลื่อนตัวข้ามส่วนบนของหัวไปยังอีกด้านหนึ่งของลำตัว ทำให้ปลาตาบอดไปข้างหนึ่ง ลูกปลาจะสูญเสียถุงลมและหนาม และจมลงสู่ก้นทะเล โดยวางด้านที่ตาบอดไว้บนพื้นผิวด้านล่าง[ 43 ]ริชาร์ด ดอว์กินส์อธิบายสิ่งนี้ว่าเป็นตัวอย่างของการปรับตัวเชิงวิวัฒนาการ
...ปลาที่มีกระดูกโดยทั่วไปมักมีแนวโน้มที่จะแบนราบในแนวตั้ง... ดังนั้นจึงเป็นเรื่องธรรมชาติที่เมื่อบรรพบุรุษของ [ปลาแบน] ลงไปอยู่ที่ก้นทะเล พวกมันจึงนอนตะแคงข้างหนึ่ง...แต่สิ่งนี้ทำให้เกิดปัญหาที่ว่าตาข้างหนึ่งจะมองลงไปในทรายตลอดเวลาและแทบจะไม่มีประโยชน์เลย ในวิวัฒนาการ ปัญหานี้ได้รับการแก้ไขโดยที่ตาข้างล่าง 'เคลื่อน' ไปอยู่ด้านบน[ 44 ]
![ปลาทะเลน้ำลึกส่วนใหญ่ไม่สามารถมองเห็นแสงสีแดงได้ ปลาปากหลวมสต็อปไลท์น้ำลึกสร้างแสงชีวภาพสีแดงเพื่อให้สามารถล่าเหยื่อด้วยลำแสงที่มองไม่เห็นอย่างมีประสิทธิภาพ[45]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b2/Malacosteus.JPG/250px-Malacosteus.JPG) ปลาทะเลน้ำลึกส่วนใหญ่ไม่สามารถมองเห็นแสงสีแดงได้ ปลาปากหลวมสต็อปไลท์ น้ำลึก สร้างแสงชีวภาพสีแดงเพื่อให้สามารถล่าเหยื่อด้วยลำแสงที่มองไม่เห็น[ 45 ]
ปลาทะเลน้ำลึกส่วนใหญ่ไม่สามารถมองเห็นแสงสีแดงได้ ปลาปากหลวมสต็อปไลท์ น้ำลึก สร้างแสงชีวภาพสีแดงเพื่อให้สามารถล่าเหยื่อด้วยลำแสงที่มองไม่เห็น[ 45 ]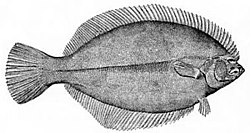 เมื่อลูกปลาแบนโตขึ้น ตาข้างหนึ่งจะหมุนไปอีกด้านหนึ่ง เพื่อให้ปลาสามารถพักพิงบนพื้นทะเลได้
เมื่อลูกปลาแบนโตขึ้น ตาข้างหนึ่งจะหมุนไปอีกด้านหนึ่ง เพื่อให้ปลาสามารถพักพิงบนพื้นทะเลได้ ปลาเพลสยุโรปเป็นปลาแบนที่มีตาโปน ดังนั้นเมื่อมันฝังตัวลงในทรายเพื่อพรางตัว มันก็ยังสามารถมองเห็นได้
ปลาเพลสยุโรปเป็นปลาแบนที่มีตาโปน ดังนั้นเมื่อมันฝังตัวลงในทรายเพื่อพรางตัว มันก็ยังสามารถมองเห็นได้
![ปลาทะเลน้ำลึกส่วนใหญ่ไม่สามารถมองเห็นแสงสีแดงได้ ปลาปากหลวมสต็อปไลท์น้ำลึกสร้างแสงชีวภาพสีแดงเพื่อให้สามารถล่าเหยื่อด้วยลำแสงที่มองไม่เห็นอย่างมีประสิทธิภาพ[45]](https://upload.wikimedia.org/wikipedia/commons/thumb/b/b2/Malacosteus.JPG/250px-Malacosteus.JPG)


เหยื่อมักจะมีตาอยู่ด้านข้างของหัวเพื่อให้มองเห็นได้กว้างขึ้น เพื่อหลีกเลี่ยงผู้ล่า ผู้ล่ามักจะมีตาอยู่ด้านหน้าของหัวเพื่อให้รับรู้ความลึกได้ ดีขึ้น [ 46 ] [ 47 ]ผู้ล่าที่อาศัยอยู่ก้นทะเล เช่นปลาแบนมีตาเรียงตัวกันเพื่อให้มองเห็นสิ่งที่อยู่เหนือพวกมันได้แบบสองตาขณะที่พวกมันนอนอยู่บนพื้น
การระบายสี
ปลาได้วิวัฒนาการวิธีการใช้สีสัน อย่างซับซ้อน ตัวอย่างเช่นปลาเหยื่อมีวิธีการใช้สีสันเพื่อทำให้ผู้ล่าที่ใช้สายตามองเห็นได้ยากขึ้น ในปลาที่อาศัยอยู่ในทะเลเปิดการปรับตัวเหล่านี้ส่วนใหญ่เกี่ยวข้องกับการลดขนาดของเงา ซึ่งเป็นรูปแบบหนึ่งของการพรางตัววิธีหนึ่งในการทำเช่นนี้คือการลดพื้นที่เงาโดยการบีบอัดด้านข้างของลำตัว อีกวิธีหนึ่งซึ่งเป็นรูปแบบหนึ่งของการพรางตัวเช่นกัน คือการใช้สีตัดกันในกรณีของปลาที่อาศัยอยู่ในผิวน้ำและการใช้สีตัดกันในกรณีของปลาที่อาศัยอยู่ในชั้นกลางน้ำการใช้สีตัดกันทำได้โดยการระบายสีปลาด้วยเม็ดสีที่เข้มกว่าที่ด้านบนและเม็ดสีที่อ่อนกว่าที่ด้านล่างในลักษณะที่สีเข้ากับพื้นหลัง เมื่อมองจากด้านบน บริเวณหลังที่เข้มกว่าของสัตว์จะกลมกลืนกับความมืดของน้ำด้านล่าง และเมื่อมองจากด้านล่าง บริเวณท้องที่อ่อนกว่าจะกลมกลืนกับแสงแดดจากผิวน้ำ การส่องสว่างย้อนกลับเกิดขึ้นผ่านการเรืองแสงทางชีวภาพโดยการผลิตแสงจากอวัยวะเรืองแสง ด้านท้อง โดยมีเป้าหมายเพื่อให้ความเข้มของแสงจากด้านล่างของปลาตรงกับความเข้มของแสงจากพื้นหลัง[ 48 ]
ปลาเบนทิกซึ่งพักอยู่บนพื้นทะเลจะซ่อนตัวโดยการขุดลงไปในทรายหรือหลบเข้าไปในซอกหลืบ หรือพรางตัวโดยการกลมกลืนกับพื้นหลังหรือทำให้ดูเหมือนหินหรือสาหร่ายทะเล[ 49 ]
แม้ว่าเครื่องมือเหล่านี้อาจมีประสิทธิภาพในฐานะกลไกการหลีกเลี่ยงผู้ล่า แต่ก็ยังเป็นเครื่องมือที่มีประสิทธิภาพสำหรับผู้ล่าเองด้วยเช่นกัน ตัวอย่างเช่นฉลามโคมไฟท้องกำมะหยี่ น้ำลึก ใช้การพรางตัวเพื่อซ่อนตัวจากเหยื่อ[ 50 ]
 ปลาที่อาศัยอยู่บนผิวน้ำ เช่นปลาทูน่าครีบฟ้าแอตแลนติก ตัวนี้ มักจะมีสีเงินแซมอยู่ทั่วตัว
ปลาที่อาศัยอยู่บนผิวน้ำ เช่นปลาทูน่าครีบฟ้าแอตแลนติก ตัวนี้ มักจะมีสีเงินแซมอยู่ทั่วตัว ปลาผีเสื้อสี่ตามีตาปลอมอยู่ที่ส่วนท้ายลำตัวทำให้ผู้ล่าสับสนว่าส่วนไหนคือส่วนหน้าของปลา
ปลาผีเสื้อสี่ตามีตาปลอมอยู่ที่ส่วนท้ายลำตัวทำให้ผู้ล่าสับสนว่าส่วนไหนคือส่วนหน้าของปลา ปลาจอห์น ดอรี่มีจุดคล้ายตาขนาดใหญ่ตรงกลางลำตัว ซึ่งทำให้เหยื่อสับสน
ปลาจอห์น ดอรี่มีจุดคล้ายตาขนาดใหญ่ตรงกลางลำตัว ซึ่งทำให้เหยื่อสับสน



ปลาบางชนิดยังแสดงจุดคล้ายตาปลอม อีก ด้วย ปลาผีเสื้อสี่ตาได้ชื่อมาจากจุดสีดำขนาดใหญ่ที่ส่วนท้ายของแต่ละข้างของลำตัว จุดนี้ล้อมรอบด้วยวงแหวนสีขาวสว่างคล้ายตา มีแถบสีดำแนวตั้งบนหัวพาดผ่านตาจริง ทำให้มองเห็นได้ยาก[ 51 ]สิ่งนี้อาจทำให้ผู้ล่าคิดว่าปลาตัวใหญ่กว่าที่เป็นจริง และสับสนระหว่างส่วนท้ายกับส่วนหน้า สัญชาตญาณแรกของปลาผีเสื้อเมื่อถูกคุกคามคือการหนี ทำให้จุดคล้ายตาปลอมอยู่ใกล้ผู้ล่ามากกว่าหัว ผู้ล่าส่วนใหญ่เล็งไปที่ดวงตา และจุดคล้ายตาปลอมนี้หลอกผู้ล่าให้เชื่อว่าปลาจะหนีโดยใช้หางนำหน้า
ปลา จอห์นดอรี่เป็นปลาชายฝั่งที่อาศัยอยู่ทั้ง บนบกและในน้ำ มีลำตัวแบนราบสูง ลำตัวบางมากจนแทบมองไม่เห็นจากด้านหน้า นอกจากนี้ยังมีจุดสีดำขนาดใหญ่ที่ด้านข้างทั้งสองข้าง ซึ่งใช้ในการส่ง "ดวงตาอาฆาต" หากมีอันตรายเข้ามาใกล้ ดวงตาขนาดใหญ่ที่ด้านหน้าของหัวช่วยให้มันมองเห็นภาพสองระยะและรับรู้ความลึกที่จำเป็นต่อการจับเหยื่อจุดคล้ายดวงตาที่ด้านข้างลำตัวของปลาจอห์นดอรี่ยังทำให้เหยื่อสับสน จากนั้นจึงดูดเหยื่อเข้าไปในปาก[ 52 ]
บาร์เรลอายส์


| วิดีโอภายนอก | |
|---|---|
ปลาบาร์เรลอายส์เป็นวงศ์ของปลาขนาดเล็กรูปร่างแปลกตาที่อาศัยอยู่ในทะเลลึก ชื่อของมันมาจากดวงตาทรงกระบอกคล้ายถังที่มักจะหันขึ้นด้านบนเพื่อตรวจจับเงาของเหยื่อ[ 53 ] [ 54 ]ปลาบาร์เรลอายส์มีดวงตาขนาดใหญ่ที่ยืดหดได้ซึ่งเด่นชัดและยื่นออกมาจากกะโหลกศีรษะ ดวงตาเหล่านี้มักจะมองขึ้นด้านบน แต่ในบางชนิดก็สามารถหมุนไปข้างหน้าได้ ดวงตาของพวกมันมีเลนส์ขนาดใหญ่และเรตินาที่มี เซลล์รูปแท่งจำนวนมากเป็นพิเศษและมีความหนาแน่นของโรดอปซิน(เม็ดสี "สีม่วงสำหรับการมองเห็น") สูง ไม่มีเซลล์รูปกรวย [ 53 ]
ปลาชนิดบาร์เรลอาย ( Macropinna microstoma ) มีโดมป้องกันโปร่งใสอยู่เหนือหัว คล้ายกับโดมเหนือห้องนักบินของเครื่องบิน ซึ่งสามารถมองเห็นเลนส์ตาได้ โดมนี้แข็งแรงและยืดหยุ่นได้ และคาดว่าช่วยปกป้องดวงตาจากเนมาโตซิสต์ (เซลล์ที่ทำให้เกิดอาการแสบ) ของไซโฟโนฟอร์ซึ่งเชื่อกันว่าปลาบาร์เรลอายขโมยอาหารมา จากไซโฟโนฟอร์ [ 53 ] [ 54 ] [ 55 ]
ปลากระบอกตาชนิดอื่น เช่นปลาปากสีน้ำตาล (brownsnout spookfish ) เป็นสัตว์มีกระดูกสันหลังเพียงชนิดเดียวที่ทราบกันว่าใช้กระจกเงาแทนเลนส์ในการโฟกัสภาพในดวงตา[ 56 ] [ 57 ]เป็นเรื่องผิดปกติที่มันใช้ทั้งระบบการหักเหและการสะท้อนแสงในการมองเห็น ดวงตาหลักที่เป็นท่อมีส่วนที่บวมรูปไข่ด้านข้างเรียกว่าไดเวอร์ติคูลัมซึ่งแยกออกจากดวงตาโดยส่วนใหญ่ด้วยเซปตัมเรตินาบุอยู่เกือบทั้งภายในดวงตา และมี ช่องเปิด กระจกตา 2 ช่อง ช่องหนึ่งหันขึ้นและอีกช่องหนึ่งหันลง ซึ่งยอมให้แสงเข้าสู่ดวงตาหลักและไดเวอร์ติคูลัมตามลำดับ ดวงตาหลักใช้เลนส์ในการโฟกัสภาพเช่นเดียวกับปลาชนิดอื่น อย่างไรก็ตาม ภายในไดเวอร์ติคูลัม แสงจะถูกสะท้อนและโฟกัสไปที่เรตินาโดยกระจกเงาโค้งแบบผสมที่ได้มาจากทาพีทัมของเรตินาซึ่งประกอบด้วยแผ่นสะท้อนแสงขนาดเล็กหลายชั้นที่อาจทำจากผลึกกัวนี น โครงสร้างแยกส่วนของดวงตาปลาสปุ๊กฟิชปากสีน้ำตาลช่วยให้ปลาสามารถมองเห็นทั้งด้านบนและด้านล่างได้ในเวลาเดียวกัน นอกจากนี้ ระบบกระจกยังเหนือกว่าเลนส์ในการรวบรวมแสง เป็นไปได้ว่าดวงตาหลักทำหน้าที่ตรวจจับวัตถุที่เป็นเงาตัดกับแสงแดด ในขณะที่ส่วนยื่นทำหน้าที่ตรวจจับ แสงวาบ ชีวเรืองแสงจากด้านข้างและด้านล่าง[ 56 ]
ฉลาม

ดวงตาของฉลามคล้ายคลึงกับดวงตาของสัตว์มีกระดูกสันหลังชนิดอื่น ๆ รวมถึงเลนส์กระจกตาและเรตินา ที่คล้ายคลึงกัน แม้ว่าการมองเห็นของพวกมันจะปรับตัวได้ดีกับ สภาพแวดล้อม ทางทะเลด้วยความช่วยเหลือของเนื้อเยื่อที่เรียกว่าtapetum lucidumเนื้อเยื่อนี้อยู่ด้านหลังเรตินาและสะท้อนแสงกลับไปยังเรตินา ทำให้มองเห็นได้ชัดเจนขึ้นในน้ำมืด ประสิทธิภาพของเนื้อเยื่อนี้แตกต่างกันไป โดยฉลามบางชนิดมี การปรับตัวให้เข้า กับชีวิตกลางคืน ได้ดีกว่า ฉลามหลายชนิดสามารถหดและขยายรูม่านตาได้เหมือนมนุษย์ ซึ่งปลาเทเลออสท์ไม่สามารถทำได้ ฉลามมีเปลือกตา แต่พวกมันไม่กระพริบตาเพราะน้ำโดยรอบช่วยทำความสะอาดดวงตา เพื่อปกป้องดวงตา ฉลามบางชนิดมีเยื่อหุ้มตาชั้นใน (nictitating membrane) เยื่อหุ้มนี้จะคลุมดวงตาขณะล่าเหยื่อและเมื่อฉลามถูกโจมตี อย่างไรก็ตาม ฉลามบางชนิด รวมถึงฉลามขาว ( Carcharodon carcharias ) ไม่มีเยื่อหุ้มนี้ แต่จะกลอกตาไปด้านหลังเพื่อป้องกันดวงตาเมื่อโจมตีเหยื่อ ความสำคัญของการมองเห็นในพฤติกรรมการล่าเหยื่อของฉลามยังคงเป็นที่ถกเถียงกันอยู่ บางคนเชื่อว่า การรับรู้ ทางไฟฟ้าและเคมีมีความสำคัญมากกว่า ในขณะที่บางคนชี้ไปที่เยื่อหุ้มตาชั้นในว่าเป็นหลักฐานว่าการมองเห็นมีความสำคัญ สันนิษฐานได้ว่าฉลามจะไม่ปกป้องดวงตาของมันหากดวงตาไม่สำคัญ การใช้การมองเห็นอาจแตกต่างกันไปตามสายพันธุ์และสภาพน้ำ ขอบเขตการมองเห็นของฉลามสามารถสลับระหว่าง การมองเห็น แบบตาเดียวและแบบสามมิติได้ตลอดเวลา[ 58 ] การศึกษา ไมโครสเปกโตรโฟโตเมตรีของฉลาม 17 สายพันธุ์พบว่า 10 สายพันธุ์มีเฉพาะเซลล์รับแสงรูปแท่งและไม่มีเซลล์รูปกรวยในเรตินาทำให้พวกมันมองเห็นได้ดีในเวลากลางคืนแต่ทำให้ตาบอดสีสายพันธุ์ที่เหลืออีกเจ็ดสายพันธุ์นอกจากเซลล์รูปแท่งแล้วยังมีเซลล์รับแสงรูปกรวย ชนิดเดียว ที่ไวต่อสีเขียว และมองเห็นได้เฉพาะเฉดสีเทาและเขียวเท่านั้น จึงเชื่อกันว่าตาบอดสีอย่างมีประสิทธิภาพ การศึกษานี้บ่งชี้ว่าความแตกต่างของวัตถุกับพื้นหลังมากกว่าสีอาจมีความสำคัญต่อการตรวจจับวัตถุ[ 59 ] [ 60 ] [ 61 ]
ตัวอย่างอื่นๆ



ปลาขนาดเล็กมัก รวม ฝูงกันเพื่อความปลอดภัย การรวมฝูงนี้มีข้อดีในด้านการมองเห็น ทั้งจากการทำให้ปลาล่าเหยื่อ สับสน และจากการมีดวงตาหลายดวงสำหรับฝูงที่มองว่าเป็นกลุ่มเดียวกัน “ผลกระทบจากการทำให้ผู้ล่าสับสน” มาจากแนวคิดที่ว่าผู้ล่าจะแยกแยะเหยื่อแต่ละตัวออกจากกลุ่มได้ยาก เนื่องจากเป้าหมายที่เคลื่อนไหวจำนวนมากทำให้เกิดการรับรู้มากเกินไปในช่องทางการมองเห็นของผู้ล่า[ 62 ] “ปลาที่รวมฝูงกันจะมีขนาดและสีเงินเหมือนกัน ดังนั้นจึงเป็นเรื่องยากสำหรับผู้ล่าที่เน้นการมองเห็นที่จะแยกแยะปลาแต่ละตัวออกจากฝูงปลาที่บิดตัวและกระพริบแสง และมีเวลาเพียงพอที่จะจับเหยื่อก่อนที่มันจะหายไปในฝูง” [ 63 ] “ผลกระทบจากดวงตาหลายดวง” มาจากแนวคิดที่ว่าเมื่อขนาดของกลุ่มเพิ่มขึ้น งานในการสแกนสภาพแวดล้อมเพื่อหาผู้ล่าสามารถกระจายไปยังปลาหลายตัวได้การทำงานร่วมกันเป็นกลุ่มน่าจะทำให้มีความระมัดระวังมากขึ้น[ 64 ] [ 65 ]
โดยปกติปลาเป็นสัตว์เลือดเย็น มีอุณหภูมิร่างกายเท่ากับน้ำโดยรอบ อย่างไรก็ตามปลาล่าเหยื่อ ในมหาสมุทรบางชนิด เช่นปลาดาบและปลาฉลามและปลาทูน่า บาง ชนิด สามารถเพิ่มอุณหภูมิให้กับบางส่วนของร่างกายเมื่อล่าเหยื่อในน้ำลึกและเย็น ปลาดาบที่มีสายตาดีเยี่ยมใช้ระบบทำความร้อนที่เกี่ยวข้องกับกล้ามเนื้อ ซึ่งเพิ่มอุณหภูมิในดวงตาและสมองได้มากถึง 15 องศาเซลเซียส การเพิ่มอุณหภูมิของเรตินาช่วยปรับปรุงอัตราการตอบสนองของดวงตาต่อการเปลี่ยนแปลงของการเคลื่อนไหวอย่างรวดเร็วของเหยื่อได้มากถึงสิบเท่า[ 66 ] [ 67 ] [ 68 ]
ปลาบางชนิดมี แสง สะท้อนจากดวงตา[ 69 ]แสงสะท้อนจากดวงตาเป็นผลมาจากชั้นรวบรวมแสงในดวงตาที่เรียกว่าtapetum lucidumซึ่งสะท้อนแสงสีขาว ไม่พบในมนุษย์ แต่สามารถมองเห็นได้ในสัตว์ชนิดอื่น เช่น กวางที่ถูกไฟหน้ารถส่อง แสงสะท้อนจากดวงตาช่วยให้ปลาสามารถมองเห็นได้ดีในสภาพแสงน้อย รวมถึงในน้ำขุ่น (น้ำมีสีหรือน้ำกระเพื่อม) ทำให้พวกมันได้เปรียบเหยื่อ การมองเห็นที่ดียิ่งขึ้นนี้ช่วยให้ปลาสามารถอาศัยอยู่ในบริเวณที่ลึกกว่าในมหาสมุทรหรือทะเลสาบได้ โดยเฉพาะอย่างยิ่งปลาวอลอาย น้ำจืด ได้รับการตั้งชื่อเช่นนั้นเพราะแสงสะท้อนจากดวงตาของมัน[ 70 ]
ปลา หลายชนิดใน วงศ์ Loricariidaeซึ่งเป็นวงศ์ของปลาแค ท ฟิช มีม่านตา ที่ดัดแปลง เรียกว่าม่านตาโอเมก้าส่วนบนของม่านตาจะโค้งลงมาเป็นรูปห่วงที่สามารถขยายและหดตัวได้ เรียกว่า ม่านตาปิด เมื่อระดับแสงสูง รูม่านตาจะลดขนาดเส้นผ่านศูนย์กลางลง และห่วงจะขยายออกไปปิดตรงกลางรูม่านตา ทำให้เกิดส่วนที่ส่งผ่านแสงเป็นรูปพระจันทร์เสี้ยว[ 71 ]ลักษณะนี้ได้ชื่อมาจากความคล้ายคลึงกับอักษรกรีกโอเมก้า (Ω) ที่กลับหัว ต้นกำเนิดของโครงสร้างนี้ยังไม่เป็นที่ทราบแน่ชัด แต่มีการเสนอแนะว่าการทำให้โครงร่างของดวงตาที่มองเห็นได้ชัดเจนนั้นดูไม่ชัดเจน ช่วยในการพรางตัวในสัตว์ที่มีลายจุดจำนวนมาก[ 71 ]
ระบบประสาทรับรู้ระยะทาง

ระบบการมองเห็นเป็นระบบรับรู้ระยะทางที่ให้ข้อมูลแก่ปลาเกี่ยวกับตำแหน่งหรือวัตถุที่อยู่ไกลออกไปโดยไม่จำเป็นต้องให้ปลาสัมผัสโดยตรง ระบบรับรู้ระยะทางดังกล่าวมีความสำคัญ เพราะช่วยให้สามารถสื่อสารกับปลาตัวอื่น และให้ข้อมูลเกี่ยวกับตำแหน่งของอาหารและผู้ล่า รวมถึงการหลีกเลี่ยงสิ่งกีดขวางหรือการรักษาตำแหน่งในฝูงปลาตัวอย่างเช่น ปลาบางชนิดที่อยู่รวมกันเป็นฝูงจะมี "เครื่องหมายแสดงการรวมฝูง" อยู่ที่ด้านข้าง เช่น แถบสีที่เด่นชัดซึ่งเป็นเครื่องหมายอ้างอิงและช่วยให้ปลาที่อยู่ใกล้เคียงสามารถตัดสินตำแหน่งสัมพัทธ์ของพวกมันได้[ 73 ]แต่ระบบการมองเห็นไม่ใช่ระบบเดียวที่สามารถทำหน้าที่ดังกล่าวได้ ปลาที่อยู่รวมกันเป็นฝูงบางชนิดยังมีเส้นข้างลำตัวที่ทอดยาวไปตามลำตัว เส้นข้างลำตัวนี้ช่วยให้ปลาสามารถรับรู้การเปลี่ยนแปลงของความดันน้ำและความปั่นป่วนที่อยู่ติดกับลำตัวได้ โดยใช้ข้อมูลนี้ ปลาที่อยู่รวมกันเป็นฝูงสามารถปรับระยะห่างจากปลาที่อยู่ใกล้เคียงได้หากพวกมันเข้ามาใกล้เกินไปหรือออกไปไกลเกินไป[ 73 ]
ระบบการมองเห็นในปลาได้รับการเสริมด้วยระบบรับรู้ชนิดอื่นที่มีฟังก์ชันที่เทียบเคียงได้หรือเสริมกัน ปลาบางชนิดตาบอดและต้องพึ่งพาระบบรับรู้ทางเลือกอื่นโดยสิ้นเชิง[ 74 ]ประสาทสัมผัสอื่นๆ ที่สามารถให้ข้อมูลเกี่ยวกับตำแหน่งหรือวัตถุที่อยู่ไกลออกไปได้ ได้แก่การได้ยินและการ หาตำแหน่งด้วยเสียงสะท้อน การรับรู้ทาง ไฟฟ้าการรับรู้ทางแม่เหล็กและการรับรู้ทางเคมี ( กลิ่นและรส ) ตัวอย่างเช่นปลาดุกมีตัวรับสารเคมีทั่วทั้งตัว ซึ่งหมายความว่าพวกมัน "รับรส" ทุกสิ่งที่สัมผัสและ "ดมกลิ่น" สารเคมีใดๆ ในน้ำ "ในปลาดุกการรับรสมีบทบาทสำคัญในการกำหนดทิศทางและตำแหน่งของอาหาร" [ 75 ]
ปลากระดูกอ่อน (ฉลาม ปลากระเบน และปลาคิเมร่า) ใช้การรับรู้สนามแม่เหล็ก พวกมันมี ตัว รับไฟฟ้า พิเศษ ที่เรียกว่าแอมพูลลาของลอเรนซินีซึ่งตรวจจับการเปลี่ยนแปลงเล็กน้อยของศักย์ไฟฟ้า ตัวรับเหล่านี้ตั้งอยู่ตามปากและจมูกของปลา ทำงานตามหลักการที่ว่าสนามแม่เหล็กที่เปลี่ยนแปลงตามเวลาซึ่งเคลื่อนที่ผ่านตัวนำจะเหนี่ยวนำให้เกิดศักย์ไฟฟ้าที่ปลายทั้งสองข้างของตัวนำ แอมพูลลายังอาจช่วยให้ปลาตรวจจับการเปลี่ยนแปลงของอุณหภูมิน้ำได้อีกด้วย[ 76 ] [ 77 ]เช่นเดียวกับในนก การรับรู้สนามแม่เหล็กอาจให้ข้อมูลที่ช่วยให้ปลาสร้างแผนที่เส้นทางการอพยพได้[ 78 ]
ดูเพิ่มเติม
- ตาของสัตว์ขาปล้อง
- อัตราส่วนของ Matthiessen
- ตาหอย
- ตาข้างขมับ
- ตาแบบง่ายในสัตว์ไม่มีกระดูกสันหลัง
- ระบบการมองเห็น
หมายเหตุ
- ^ Meyer-Rochow, V. Benno; Stewart, Duncan (1996). "การทบทวนโครงสร้างระดับจุลภาคของดวงตาในระยะตัวอ่อนและหลังตัวอ่อนของปลาแลมเพรย์ (ไซโคลสโตมาตา) โดยเน้นเป็นพิเศษที่ Geotria australis (สีเทา)" Microscopy Research and Technique . 35 (6): 431– 444. doi : 10.1002/(SICI)1097-0029(19961215)35:6<431::AID-JEMT3>3.0.CO;2-L . PMID 9016447 . S2CID 22940203 .
- ^ Lamb, Trevor D.; Collin, Shaun P.; Pugh, Edward N. (2007). "วิวัฒนาการของดวงตาของสัตว์มีกระดูกสันหลัง: ออปซิน, โฟโตรีเซปเตอร์, เรตินา และถ้วยตา" Nature Reviews Neuroscience 8 ( 12): 960– 976. doi : 10.1038/nrn2283 . ISSN 1471-003X . PMC 3143066 . PMID 18026166 . ดูเพิ่มเติมได้ในงานวิจัยเรื่อง "ต้นกำเนิดของดวงตาของสัตว์มีกระดูกสันหลัง" โดย Lamb และคณะ ปี 2008
- ^ Trevor D. Lamb; Shaun P. Collin; Edward N. Pugh Jr (2007). "วิวัฒนาการของดวงตาของสัตว์มีกระดูกสันหลัง: ออปซิน, โฟโตรีเซปเตอร์, เรตินา และถ้วยตา" Nature Reviews Neuroscience 8 ( 12): 960– 976. doi : 10.1038/nrn2283 . PMC 3143066 . PMID 18026166 .
- ^ Ocean Explorer NOAA. อัปเดตล่าสุด: 26 สิงหาคม 2553
- ^ a b c d e f g h i j k Helfman et al. 2009 , หน้า 84–87.
- ^ a b c Webb, Paul (2019) Introduction to Oceanographyบทที่ 6.5 แสง Rebus Community มหาวิทยาลัย Roger Williams ตำราเรียนแบบเปิด
 เนื้อหาถูกคัดลอกจากแหล่งข้อมูลนี้ ซึ่งเผยแพร่ภายใต้Creative Commons Attribution 4.0 International License
เนื้อหาถูกคัดลอกจากแหล่งข้อมูลนี้ ซึ่งเผยแพร่ภายใต้Creative Commons Attribution 4.0 International License - ^ a b c d Land, MF; Nilsson, D. (2012). ดวงตาของสัตว์ . สำนักพิมพ์มหาวิทยาลัยออกซ์ฟอร์ด. ISBN 9780199581146.
- ^ Wehner, R (2005). "สรีรวิทยาประสาทสัมผัส: ดวงตาไร้สมอง" ( PDF) Nature . 435 (7039): 157– 159. Bibcode : 2005Natur.435..157W . doi : 10.1038/435157a . PMID 15889076 . S2CID 4408533 .
- ^ a b Novales Flamarique, Iñigo (2019). "พฤติกรรมการว่ายน้ำปรับการมองเห็นแบบโพลาไรเซชันของปลาเพื่อเพิ่มระยะการมองเห็นเหยื่อเป็นสองเท่า" Scientific Reports . 9 (1): 944. Bibcode : 2019NatSR...9..944N . doi : 10.1038/s41598-018-37632-1 . PMC 6353921 . PMID 30700806 .
- ^ Ingram, Norianne T.; Sampath, Alapakkam P.; Fain, Gordon L. (2016). "ทำไมเซลล์รูปแท่งจึงไวต่อแสงมากกว่าเซลล์รูปกรวย?"วารสารสรีรวิทยา 594 ( 19): 5415– 5426. doi : 10.1113/JP272556 . PMC 5043029 . PMID 27218707 .
- ^ Marshall, Justin; Carleton, Karen L.; Cronin, Thomas (2015). "การมองเห็นสีในสิ่งมีชีวิตในทะเล" Current Opinion in Neurobiology . 34 : 86– 94. doi : 10.1016/j.conb.2015.02.002 . PMID 25725325 . S2CID 20978931 .
- ^ Miyazaki, T; Iwamu, T; Meyer-Rochow, VB (2011). "ตำแหน่งของบริเวณศูนย์กลางเรตินาเปลี่ยนแปลงไปตามอายุใน Champsocephalus gunnari (Channichthyidae) ปลาล่าเหยื่อจากน่านน้ำชายฝั่งแอนตาร์กติกา" Polar Biology . 34 (8): 1117– 1123. Bibcode : 2011PoBio..34.1117M . doi : 10.1007/s00300-011-0969-2 . S2CID 19066809 .
- ^ Schwab, IR; Hart, N (2006). "มากกว่าขาวดำ" . British Journal of Ophthalmology . 90 (4): 406. doi : 10.1136/bjo.2005.085571 . PMC 1857009 . PMID 16572506 .
- ^ Schwab, Ivan R. (2012)พยานแห่งวิวัฒนาการ: วิวัฒนาการของดวงตาหน้า 82 สำนักพิมพ์มหาวิทยาลัยออกซ์ฟอร์ด ISBN 9780195369748.
- ^ Khorramshahia, O; Schartaua, JM; Krögera, RHH (2008). "ระบบที่ซับซ้อนของเอ็นและกล้ามเนื้อช่วยยึดเลนส์ตาไว้ในดวงตาของปลากระดูกแข็ง (teleosts)" . Vision Research . 48 (13): 1503– 1508. doi : 10.1016/j.visres.2008.03.017 . PMID 18471852 . S2CID 17757889 .
- ^ Singh HR และ Khanna SS (1994)ความก้าวหน้าทางชีววิทยาของปลาหน้า 235 สำนักพิมพ์ Hindustan ISBN 978-81-7075-029-1.
- ^ Barnes, GR (1 กุมภาพันธ์ 1979). "การทำงานของระบบเวสติบูโล-โอคูลาร์ระหว่างการเคลื่อนไหวศีรษะและดวงตาที่ประสานกันเพื่อรับเป้าหมายภาพ"วารสารสรีรวิทยา 287 ( 1): 127– 147. doi : 10.1113/jphysiol.1979.sp012650 . PMC 1281486 . PMID 311828 .
- ^ Graf, Werner; Spencer, Robert; Baker, Harriet; Baker, Robert (1 พฤษภาคม 1997). "เส้นทางเวสติบูลาร์ที่กระตุ้นและยับยั้งไปยังนิวเคลียสของมอเตอร์นอกลูกตาในปลาทอง" วารสารประสาทสรีรวิทยา 77 ( 5): 2765– 2779. doi : 10.1152/jn.1997.77.5.2765 . PMID 9163391 . S2CID 13004673 .
- ^ Graf, W.; Baker, R. (1 ตุลาคม 1985). "ปฏิกิริยาตอบสนองของระบบเวสติบูลาร์ต่อการเคลื่อนไหวของดวงตาในปลาแบนโตเต็มวัย II. การเชื่อมต่อระหว่างระบบเวสติบูลาร์และระบบเคลื่อนไหวของดวงตา" วารสารประสาทสรีรวิทยา 54 (4): 900– 916. doi : 10.1152/jn.1985.54.4.900 . PMID 4067626 .
- ^ a b Graf, Werner; Spencer, Robert; Baker, Harriet; Baker, Robert (1 กันยายน 2544). "ปฏิกิริยาตอบสนองของระบบเวสติบูลาร์ต่อดวงตาของปลาแบนโตเต็มวัย III. รูปแบบการกระตุ้นและการยับยั้งแบบผกผันเฉพาะสายพันธุ์" วารสารประสาทสรีรวิทยา 86 ( 3): 1376– 1388. doi : 10.1152/jn.2001.86.3.1376 . PMID 11535684 .
- ^ Yokoyama, Shozo; Yokoyama, Ruth (พฤศจิกายน 1996). "วิวัฒนาการเชิงปรับตัวของตัวรับแสงและเม็ดสีการมองเห็นในสัตว์มีกระดูกสันหลัง". Annual Review of Ecology and Systematics . 27 (1): 543– 567. Bibcode : 1996AnRES..27..543Y . doi : 10.1146/annurev.ecolsys.27.1.543 .
- ^ a b Shi, Yongsheng; Yokoyama, Shozo (8 กรกฎาคม 2546). "การวิเคราะห์ระดับโมเลกุลของความสำคัญเชิงวิวัฒนาการของการมองเห็นรังสีอัลตราไวโอเลตในสัตว์มีกระดูกสันหลัง" . Proceedings of the National Academy of Sciences . 100 (14): 8308– 8313. Bibcode : 2003PNAS..100.8308S . doi : 10.1073/pnas.1532535100 . PMC 166225 . PMID 12824471 .
- ^ Carleton, Karen L; Hárosi, Ferenc I; Kocher, Thomas D (เมษายน 2543). "เม็ดสีการมองเห็นของปลาหมอสีแอฟริกัน: หลักฐานการมองเห็นรังสีอัลตราไวโอเลตจากไมโครสเปกโตรโฟโตเมตรีและลำดับดีเอ็นเอ" . Vision Research . 40 (8): 879– 890. doi : 10.1016/S0042-6989(99)00238-2 . PMID 10720660 . S2CID 5420659 .
- ^ Kodric-Brown, Astrid; Johnson, Sally C. (1 กุมภาพันธ์ 2545). "รูปแบบการสะท้อนแสงอัลตราไวโอเลตของปลาหางนกยูงตัวผู้ช่วยเพิ่มความน่าดึงดูดใจต่อปลาหางนกยูงตัวเมีย" พฤติกรรมสัตว์63 (2): 391– 396. doi : 10.1006/anbe.2001.1917 . S2CID 53172856 .
- ^ Rick, Ingolf P.; Modarressie, Ricarda; Bakker, Theo CM (กุมภาพันธ์ 2549). "ความยาวคลื่น UV มีผลต่อการเลือกคู่ของปลาหนามสามแฉกเพศเมีย". พฤติกรรมสัตว์ . 71 (2): 307– 313. doi : 10.1016/j.anbehav.2005.03.039 . S2CID 937644 .
- ^ Jacobs, Gerald H. (สิงหาคม 1992). "การมองเห็นรังสีอัลตราไวโอเลตในสัตว์มีกระดูกสันหลัง" . American Zoologist . 32 (4): 544– 554. doi : 10.1093/icb/32.4.544 .
- ^ Losey, George S (1 สิงหาคม 2546). "หน้าที่การพรางตัวและการสื่อสารของสี UV-visible ในปลาแดมเซลแนวปะการังสองชนิด Dascyllus aruanus และ D.reticulatus *". พฤติกรรมสัตว์ 66 ( 2): 299– 307. doi : 10.1006/anbe.2003.2214 . S2CID 140204848 .
- ^ Siebeck, Ulrike E.; Parker, Amira N.; Sprenger, Dennis; Mäthger, Lydia M.; Wallis, Guy (มีนาคม 2010). "ปลาแนวปะการังชนิดหนึ่งที่ใช้ลวดลายอัลตราไวโอเลตเพื่อการจดจำใบหน้าแบบลับๆ" . Current Biology . 20 (5): 407– 410. Bibcode : 2010CBio...20..407S . doi : 10.1016/j.cub.2009.12.047 . PMID 20188557 . S2CID 3743161 .
- ↑ Horváth G และ Varjú D (2004) แสงโพลาไรซ์ในการมองเห็นของสัตว์: รูปแบบโพลาไรเซชันในธรรมชาติ p. 294, สปริงเกอร์.ไอเอสบีเอ็น 978-3-540-40457-6.
- ^ Denton, EJ ; Nichol, JAC (1965). "การโพลาไรซ์ของแสงที่สะท้อนจากพื้นผิวสีเงินของAlburnus alburnus " (PDF) . J. Mar. Biol. Assoc. U. K . 150 (3): 78– 94. Bibcode : 1965JMBUK..45..705D . doi : 10.1017/S0025315400016532 . เก็บถาวรจากต้นฉบับ(PDF)เมื่อวันที่ 15 เมษายน 2012 . สืบค้นเมื่อ28 กันยายน 2011 .
- ^ Rowe, DM; Denton, EJ (1997). "พื้นฐานทางกายภาพของการสื่อสารแบบสะท้อนกลับระหว่างปลา โดยเฉพาะอย่างยิ่งปลาแมคเคอเรลม้าTrachurus trachurus " Phil . Trans. R. Soc. Lond. B. 352 ( 1353): 531– 549. Bibcode : 1997RSPTB.352..531R . doi : 10.1098/rstb.1997.0037 . PMC 1691948 .
- ^ Pignatelli, V.; Champ, C.; Marshall, J.; Vorobyev, M. (2010). "กรวยคู่ใช้สำหรับการแยกแยะสีในปลาแนวปะการังRhinecanthus aculeatus " . Biology Letters . 6 (4): 537– 539. doi : 10.1098/rsbl.2009.1010 . PMC 2936199 . PMID 20129950 .
- ^ Froese, Rainer ; Pauly, Daniel (บรรณาธิการ). " Gigantura chuni " . FishBase . ฉบับตุลาคม 2010.
- ^ Froese, Rainer ; Pauly, Daniel (บรรณาธิการ). " Dissostichus mawsoni " . FishBase . ฉบับเดือนสิงหาคม 2552.
- ^เนลสัน, โจเซฟ, เอส. (2006). ปลาแห่งโลก . จอห์น ไวลีย์ แอนด์ ซันส์ อิงค์. ISBN 978-0-471-25031-9.
{{cite book}}: CS1 maint: multiple names: authors list ( link ) - ↑ ฟรอส, ไรเนอร์ ; พอลี, แดเนียล (บรรณาธิการ). " อานาเบิ้ลส์ อานาเบิ้ลส์ " . ฟิชเบส . ฉบับเดือนมีนาคม 2550
- ^ Moyle & Cech 2004 , หน้า 585.
- ^ Morin, James G.; Harrington, Anne; Nealson, Kenneth; Krieger, Neil; Baldwin, Thomas O.; Hastings, JW (1975). "แสงสว่างสำหรับทุกเหตุผล: ความหลากหลายในพฤติกรรมของปลาไฟฉาย" Science . 190 (4209): 74– 76. Bibcode : 1975Sci...190...74M . doi : 10.1126/science.190.4209.74 . S2CID 83905458 .
- ^ McCosker, John E. (มีนาคม 1977). "ปลาเรืองแสง". Scientific American . 236 (3): 106– 114. Bibcode : 1977SciAm.236c.106M . doi : 10.1038/scientificamerican0377-106 . JSTOR 24953941. PMID 841297 .
- ^แพ็กซ์ตัน, จอห์น อาร์. (1998). แพ็กซ์ตัน, เจ.อาร์.; เอชไมเยอร์, ดับเบิลยู.เอ็น. (บรรณาธิการ). สารานุกรมปลา . ซานดิเอโก: สำนักพิมพ์วิชาการ. หน้า 162. ISBN 978-0-12-547665-2.
- ^ Ryan P "สิ่งมีชีวิตในทะเลลึก: เขตบาธิเพลาจิก" เก็บ ถาวร เมื่อวันที่ 21 ธันวาคม 2008 ที่ Wayback Machine Te Ara – สารานุกรมแห่งนิวซีแลนด์ปรับปรุงเมื่อวันที่ 21 กันยายน 2007
- ^ Moyle & Cech 2004 , หน้า 587.
- ↑ชาปลู, ฟรองซัวส์ และ อามาโอกะ, คูนิโอ (1998) Paxton, JR & Eschmeyer, WN (สหพันธ์) สารานุกรมปลา . ซานดิเอโก: สำนักพิมพ์วิชาการ. xxx. ไอเอสบีเอ็น 978-0-12-547665-2.
- ^ ดอว์กินส์, ริชาร์ด (1991). ช่างทำนาฬิกาตาบอด . ลอนดอน: สำนักพิมพ์เพนกวิน. หน้า 92. ISBN 978-0-14-014481-9.
- ^ Kenaley, CP (2007). "การแก้ไขสกุลปลาปากหลวมไฟจราจรMalacosteus (Teleostei: Stomiidae: Malacosteinae) พร้อมคำอธิบายชนิดใหม่จากซีกโลกใต้เขตอบอุ่นและมหาสมุทรอินเดีย" Copeia . 2007 (4): 886– 900. doi : 10.1643/0045-8511(2007)7[886:ROTSLG]2.0.CO;2 . S2CID 1038874 .
- ^ "สัตว์กินเนื้อ" . กระทรวงมหาดไทยสหรัฐอเมริกา สำนักงานจัดการที่ดิน 14 ธันวาคม 2009. เก็บถาวรจากต้นฉบับเมื่อ 14 มิถุนายน 2011. สืบค้นเมื่อ28 มีนาคม 2011 .
- ^ Boroditsky, Lera (24 มิถุนายน 1999). "แสงและดวงตา: บันทึกการบรรยาย" . บันทึกการบรรยาย . สแตนฟอร์ด . เก็บถาวรจากต้นฉบับเมื่อ 5 กรกฎาคม 2010 . สืบค้นเมื่อ11 พฤษภาคม 2010 .
- ^ การพราง ตัวด้วยสี (Countershading) BBC: วิทยาศาสตร์และธรรมชาติสืบค้นเมื่อ 28 กันยายน 2011
- ^คู่มือเตรียมความพร้อมสำหรับเพื่อนและศัตรูที่น่าสงสัยสถาบันวิจัยทางทะเลลองบีช
- ^ Claes, Julien M.; Aksnes, Dag L.; Mallefet, Jérôme (พฤษภาคม 2010). "นักล่าลึกลับแห่งฟยอร์ด: การพรางตัวด้วยการส่องสว่างย้อนกลับในฉลาม (Etmopterus spinax)". Journal of Experimental Marine Biology and Ecology . 388 ( 1– 2): 28– 32. Bibcode : 2010JEMBE.388...28C . doi : 10.1016/j.jembe.2010.03.009 .
- ^ FishBase Froese, Rainer ; Pauly, Daniel (บรรณาธิการ). " Chaetodon capistratus " . FishBase . ฉบับเดือนกรกฎาคม 2552.
- ^ Walrond, Carl (2006)ปลาชายฝั่ง - ปลาที่อาศัยอยู่บนพื้นทะเลเปิดเก็บถาวรเมื่อวันที่ 14 มกราคม 2013 ที่ Wayback Machine , Te Ara: สารานุกรมแห่งนิวซีแลนด์เข้าถึงเมื่อวันที่ 28 พฤษภาคม 2019
- ^ a b c Robison, Bruce H.; Reisenbichler, Kim R. (18 ธันวาคม 2008). "Macropinna microstoma และความขัดแย้งของดวงตาท่อ". Copeia . 2008 (4): 780– 784. doi : 10.1643/CG-07-082 . S2CID 85768623 .
- ^ a b Robison, Bruce H.; Reisenbichler, Kim R. (18 ธันวาคม 2008). "Macropinna microstoma และความขัดแย้งของดวงตาท่อ". Copeia . 2008 (4): 780– 784. doi : 10.1643/CG-07-082 . JSTOR 25512162 . S2CID 85768623 . ProQuest 207224476 .
- "นักวิจัยไขปริศนาปลาทะเลลึกที่มีดวงตาเป็นท่อและหัวโปร่งใส"สถาบันวิจัยสัตว์น้ำมอนเทอเรย์เบย์ (ข่าวประชาสัมพันธ์) 23 กุมภาพันธ์ 2552
- ^ Froese, Rainer ; Pauly, Daniel (บรรณาธิการ). " Macropinna microstoma " . FishBase . ฉบับเดือนกันยายน 2011.
- ^ a b Wagner, Hans-Joachim; Douglas, Ron H.; Frank, Tamara M.; Roberts, Nicholas W.; Partridge, Julian C. (มกราคม 2009). "ดวงตาของสัตว์มีกระดูกสันหลังแบบใหม่ที่ใช้ทั้งเลนส์หักเหและเลนส์สะท้อนแสง" Current Biology . 19 (2): 108– 114. Bibcode : 2009CBio...19..108W . doi : 10.1016/j.cub.2008.11.061 . PMID 19110427 . S2CID 18680315 .
- ^ผู้สื่อข่าว ลูอิส สมิธ ด้านสิ่งแวดล้อม (8 มกราคม 2552) "ปลาที่มีสี่ตา สามารถมองทะลุความมืดมิดของทะเลลึกได้"เดอะไทมส์
{{cite news}}: CS1 maint: multiple names: authors list ( link ) - ^ Martin, R. Aidan. "Vision and a Carpet of Light" . ReefQuest Centre for Shark Research . สืบค้นเมื่อ22 สิงหาคม 2552 .
- ↑ฮาร์ต, นาธาน สก็อตต์; ธีสส์, ซูซาน มิเชล; ฮาราบุช, เบลค คริสติน; คอลลิน, ชอน แพทริค (มีนาคม 2554) "หลักฐานทางไมโครสเปกโตรโฟโตเมตริกสำหรับสีเดียวของโคนในฉลาม" นาตูร์วิสเซ่นชาฟเทิน . 98 (3): 193– 201. Bibcode : 2011NW.....98..193H . ดอย : 10.1007/s00114-010-0758-8 . PMID21212930 . S2CID 30148811 .
- "ฉลามตาบอดสี ผลการศึกษาใหม่พบ" Australian Geographic . 19 มกราคม 2011.
- ^กิลล์, วิคตอเรีย (18 มกราคม 2011). "ฉลามอาจตาบอดสี" . บีบีซี นิวส์. สืบค้นเมื่อ19 มกราคม 2011 .
- ↑ฮาร์ต, นาธาน สก็อตต์; ธีสส์, ซูซาน มิเชล; ฮาราบุช, เบลค คริสติน; คอลลิน, ชอน แพทริค (มีนาคม 2554) "หลักฐานทางไมโครสเปกโตรโฟโตเมตริกสำหรับสีเดียวของโคนในฉลาม" นาตูร์วิสเซ่นชาฟเทิน . 98 (3): 193– 201. Bibcode : 2011NW.....98..193H . ดอย : 10.1007/s00114-010-0758-8 . PMID21212930 . S2CID 30148811 .
- ^ Milinski, Manfred; Heller, Rolf (ตุลาคม 1978). "อิทธิพลของผู้ล่าต่อพฤติกรรมการหาอาหารที่เหมาะสมที่สุดของปลาหนาม (Gasterosteus aculeatus L.)" Nature . 275 (5681): 642– 644. Bibcode : 1978Natur.275..642M . doi : 10.1038/275642a0 . S2CID 4184043 .
- ^ Moyle & Cech 2004 , หน้า .
- ^ Roberts, Gilbert (พฤษภาคม 1996). "เหตุใดความระมัดระวังของแต่ละบุคคลจึงลดลงเมื่อขนาดกลุ่มเพิ่มขึ้น" พฤติกรรมสัตว์ 51 ( 5): 1077– 1086. doi : 10.1006/anbe.1996.0109 . S2CID 53202810 .
- ^ Lima, Steven L. (มกราคม 1995). "กลับสู่พื้นฐานของการเฝ้าระวังป้องกันผู้ล่า: ผลกระทบของขนาดกลุ่ม" พฤติกรรมสัตว์ 49 ( 1): 11– 20. doi : 10.1016/0003-3472(95)80149-9 . S2CID 53205760 .
- ^ Fritsches, Kerstin A.; Brill, Richard W.; Warrant, Eric J. (มกราคม 2548). "ดวงตาที่อบอุ่นให้การมองเห็นที่เหนือกว่าในปลาดาบ" . Current Biology . 15 (1): 55– 58. Bibcode : 2005CBio...15...55F . doi : 10.1016/j.cub.2004.12.064 . PMID 15649365 . S2CID 14070646 .
- ^ฮอปกิน, ไมเคิล (10 มกราคม 2548). "ปลาดาบให้ความร้อนกับดวงตาเพื่อการมองเห็นที่ดีขึ้น" Nature : news050110–2. doi : 10.1038/news050110-2 .
- ↑เฮลฟ์แมน และคณะ 2552 , หน้า 95–97.
- ^ Somiya, H (1980). "ปลาที่มีแสงสะท้อนในตา: สัณฐานวิทยาเชิงหน้าที่ของ Tapetum Lucidum ชนิดกัวนีน" Marine Ecology Progress Series . 2 : 9– 26. Bibcode : 1980MEPS....2....9S . doi : 10.3354/meps002009 .
- ^ Johnson JA และ Esser R (2009) " http://www.fishculturesection.org/Aquanotes/pdf/Aq_App_Note_1_April_2009.pdf เก็บถาวรเมื่อ 1 พฤษภาคม 2011 ที่ Wayback Machineการเพาะเลี้ยงปลาวอลอาย – การปรับตัวให้กินอาหารในที่มืด"สมาคมการประมงอเมริกัน เอกสารประกอบการประยุกต์ใช้ด้านการเพาะเลี้ยงสัตว์น้ำ
- ^ a b Douglas, Ron H.; Collin, Shaun P.; Corrigan, Julie (15 พฤศจิกายน 2002). "ดวงตาของปลาดุกปากดูด (Loricariidae, วงศ์ย่อย Hypostomus): การตอบสนองของรูม่านตา ความคลาดเคลื่อนทรงกลมตามยาวของเลนส์ และลักษณะทางภูมิศาสตร์ของเรตินา"วารสารชีววิทยาเชิงทดลอง 205 ( 22): 3425– 3433. doi : 10.1242/jeb.205.22.3425 . PMID 12364396 .
- ^ Yoshizawa, Masato; Yamamoto, Yoshiyuki; O'Quin, Kelly E; Jeffery, William R (ธันวาคม 2012). "วิวัฒนาการของพฤติกรรมปรับตัวและตัวรับความรู้สึกส่งเสริมการถดถอยของดวงตาในปลาถ้ำตาบอด" . BMC Biology . 10 (1): 108. doi : 10.1186/1741-7007-10-108 . PMC 3565949 . PMID 23270452 .
- ^ a b Bone & Moore 2008 , หน้า 418–422.
- ^โบน แอนด์ มัวร์ 2008 , หน้า 311.
- ^ Atema, Jelle (1980) "ประสาทสัมผัสทางเคมี สัญญาณทางเคมี และพฤติกรรมการกินอาหารในปลา"หน้า 57–101 ใน: Bardach, JEพฤติกรรมของปลาและการนำไปใช้ในการจับและเพาะเลี้ยงปลาศูนย์ปลาโลก ISBN 978-971-02-0003-0.
- ^ Fields, R. Douglas; Fields, Kyle D.; Fields, Melanie C. (ตุลาคม 2550). "เจลเซมิคอนดักเตอร์ในอวัยวะรับความรู้สึกของฉลาม?" . Neuroscience Letters . 426 (3): 166– 170. doi : 10.1016/j.neulet.2007.08.064 . PMC 2211453 . PMID 17904741 .
- ^ Brown, Brandon R. (มีนาคม 2010). "การตอบสนองต่ออุณหภูมิในอิเล็กโทรเซนเซอร์และแรงดันไฟฟ้าความร้อนในอิเล็กโทรไลต์" . Journal of Biological Physics . 36 (2): 121– 134. doi : 10.1007/s10867-009-9174-8 . PMC 2825305 . PMID 19760113 .
- ^ Johnsen, Sönke; Lohmann, Kenneth J. (กันยายน 2548). "ฟิสิกส์และประสาทชีววิทยาของการรับรู้สนามแม่เหล็ก" Nature Reviews Neuroscience . 6 (9): 703– 712. doi : 10.1038/nrn1745 . PMID 16100517 . S2CID 13996233 .
อ่านเพิ่มเติม
- อาร์เธอร์, โจเซฟ; นิโคล, โคลิน; โซมิยะ, ฮิโรอากิ (1989). ดวงตาของปลา . สำนักพิมพ์แคลเรนดอน. ISBN 978-0-19-857195-7.
- Douglas, RH & Djamgoz, M. (บรรณาธิการ) (1990) ระบบการมองเห็นของปลาสำนักพิมพ์ Chapman and Hall, 526 หน้า
- แลมบ์, เทรเวอร์ ดี. (14 มิถุนายน 2011). "วิวัฒนาการของดวงตา". Scientific American . 305 (1): 64– 69. Bibcode : 2011SciAm.305f..64L . doi : 10.1038/scientificamerican0711-64 .
- Land, Michael F และ Nilsson, Dan-Eric (2012) Animal Eyesสำนักพิมพ์ Oxford University Press ISBN 9780199581146.
- Lamb, Trevor D.; Collin, Shaun P.; Pugh, Edward N. (ธันวาคม 2007). "วิวัฒนาการของดวงตาของสัตว์มีกระดูกสันหลัง: ออปซิน, โฟโตรีเซปเตอร์, เรตินา และถ้วยตา" Nature Reviews . Neuroscience . 8 (12): 960– 976. doi : 10.1038/nrn2283 . PMC 3143066. PMID 18026166 .
- "ติดตามความเคลื่อนไหวของการวิวัฒนาการ" Phys.org 3ธันวาคม 2007
- Nilsson, DE; Pelger, S (22 เมษายน 1994). "การประเมินอย่างมองโลกในแง่ร้ายเกี่ยวกับเวลาที่จำเป็นสำหรับการวิวัฒนาการของดวงตา". Proceedings of the Royal Society of London. Series B: Biological Sciences . 256 (1345): 53– 58. Bibcode : 1994RSPSB.256...53N . doi : 10.1098/rspb.1994.0048 . PMID 8008757. S2CID 13061351 .
- เบอร์ลินสกี, เดวิด (2002) ดาร์วินเจอคู่ปรับแล้วหรือ? เก็บถาวรเมื่อวันที่ 4 มีนาคม 2016 ที่Wayback Machineหน้า 34, The Vexing Eye (จดหมาย). บทวิจารณ์ , 1 ธันวาคม 2002
- นิลส์สัน, แดน-อี. "ระวังวิทยาศาสตร์เทียม: การตอบโต้การโจมตีของเดวิด เบอร์ลินสกีต่อการคำนวณของผมเกี่ยวกับระยะเวลาที่ดวงตาจะวิวัฒนาการ" . Talk Reason . เก็บถาวรจากต้นฉบับเมื่อ 21 พฤษภาคม 2019 . สืบค้นเมื่อ28 เมษายน 2013 .
- Meyer-Rochow, Victor Benno; Coddington, Paul Edward (2003). "ดวงตาและการมองเห็นของปลาCheimarrichthys foster VON HAAST (1874) ในนิวซีแลนด์: เนื้อเยื่อวิทยา เคมีแสง และสรีรวิทยาไฟฟ้า" ใน Val, Adalberto Luís; Kapoor, BG (บรรณาธิการ). การปรับตัวของปลา . สำนักพิมพ์ Science Publishers. หน้า 337–383 . ISBN 978-1-57808-249-0.
- "วิวัฒนาการของดวงตา" – วิดีโอเกี่ยวกับแบบจำลอง Nilsson-Pelger (เลื่อนลงด้านล่าง)
บรรณานุกรม
- Marshall, Justin; Carleton, Karen L; Cronin, Thomas (ตุลาคม 2015). "การมองเห็นสีในสิ่งมีชีวิตในทะเล". Current Opinion in Neurobiology . 34 : 86–94 . doi : 10.1016/j.conb.2015.02.002 . PMID 25725325. S2CID 20978931 .
- Kamijo, Makiko; Kawamura, Mayuko; Fukamachi, Shoji (พฤษภาคม 2018). "การสูญเสียยีนออปซินสีแดงทำให้การแยกเพศระหว่างปลาเมดากะที่มีสีผิวต่างกันลดลง" กระบวนการทางพฤติกรรม150 : 25– 28. doi : 10.1016/j.beproc.2018.02.006 . PMID 29447852 . S2CID 4239046 .
ลิงก์ภายนอก
- เปรียบเทียบระบบการมองเห็นของปลาและมนุษย์ – สไลด์นำเสนอ
- ปลาสามารถมองเห็นน้ำได้หรือไม่? - AboutFishTank
- รูปภาพสัตว์เลี้ยง Axolotl ถูกเก็บถาวรเมื่อวันที่ 12 พฤศจิกายน 2024 ที่Wayback Machine
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การมองเห็นในปลา
การมองเห็นเป็นระบบประสาทสัมผัส ที่สำคัญสำหรับ ปลาส่วนใหญ่ดวงตา ของ ปลามีลักษณะคล้ายกับดวงตาของสัตว์มีกระดูกสันหลังบนบก เช่นนกและสัตว์เลี้ยงลูกด้วยนมแต่มีเลนส์ที่มีรูปทรงกลม...
น้ำในฐานะสภาพแวดล้อมทางสายตา
ปลาและสัตว์น้ำอื่นๆ อาศัยอยู่ในสภาพแวดล้อมที่มีแสงแตกต่างจากสัตว์บก น้ำ ดูดซับ แสง ดังนั้นเมื่อความลึกเพิ่มขึ้น ปริมาณแสงที่มีอยู่จะลดลงอย่างรวดเร็ว คุณสมบัติทางแสงของน้ำยังส่งผลให้มี การดูดซับแสงที่ มีความยาวคลื่น ต่างกัน ในระดับที่แตกต่างกัน ตัวอย่างเช่น...
โครงสร้างและหน้าที่
ดวงตาของปลาโดยทั่วไปคล้ายคลึงกับดวงตาของสัตว์มีกระดูกสันหลังอื่นๆ โดยเฉพาะอย่างยิ่งสัตว์ สี่ขา (สัตว์ครึ่งบกครึ่งน้ำ สัตว์เลื้อยคลาน นก และสัตว์เลี้ยงลูกด้วยนม ซึ่งทั้งหมดวิวัฒนาการมาจากบรรพบุรุษที่เป็นปลา) แสงเข้าสู่ดวงตาที่ กระจกตา ผ่าน รูม่านตา ไปยัง เลนส์...
เรตินา
ภายในเรตินา เซลล์แท่ง ให้ความไวในการมองเห็นสูง (โดยแลกกับ ความคมชัด ) และใช้ในสภาพแสงน้อย เซลล์กรวยให้ความละเอียดเชิงพื้นที่และเวลาที่สูงกว่าเซลล์แท่ง และทำให้สามารถมองเห็นสีได้โดยการเปรียบเทียบการดูดกลืนแสงระหว่างเซลล์กรวยชนิดต่างๆ...