อ่าน 32 นาที
ผลึกศาสตร์รังสีเอกซ์
การศึกษาโครงสร้างผลึกด้วยรังสีเอกซ์ เป็นวิทยาศาสตร์เชิงทดลองที่ใช้ในการหาโครงสร้างอะตอมและโมเลกุลของ ผลึก โดยโครงสร้างดังกล่าวจะทำให้ลำแสง รังสีเอกซ์ ที่ตกกระทบ เกิด...
ผลึกศาสตร์รังสีเอกซ์

การศึกษาโครงสร้างผลึกด้วยรังสีเอกซ์เป็นวิทยาศาสตร์เชิงทดลองที่ใช้ในการหาโครงสร้างอะตอมและโมเลกุลของผลึกโดยโครงสร้างดังกล่าวจะทำให้ลำแสงรังสีเอกซ์ ที่ตกกระทบ เกิดการเลี้ยวเบนไปในทิศทางเฉพาะ การวัดมุมและความเข้มของการเลี้ยวเบนของรังสีเอกซ์ ทำให้ นักผลึกศาสตร์สามารถสร้าง ภาพ สามมิติของความหนาแน่นของอิเล็กตรอนภายในผลึกและตำแหน่งของอะตอมรวมถึงพันธะเคมี ความไม่เป็นระเบียบ ของโครงสร้างผลึกและข้อมูลอื่นๆ ได้
การวิเคราะห์โครงสร้างด้วยรังสีเอกซ์มีความสำคัญอย่างยิ่งต่อการพัฒนาในหลายสาขาวิทยาศาสตร์ ในช่วงทศวรรษแรกของการใช้งาน วิธีนี้ใช้ในการกำหนดขนาดของอะตอม ความยาวและชนิดของพันธะเคมี และความแตกต่างในระดับอะตอมระหว่างวัสดุต่างๆ โดยเฉพาะอย่างยิ่งแร่ธาตุและโลหะผสมวิธีนี้ยังเปิดเผยโครงสร้างและหน้าที่ของโมเลกุลทางชีวภาพหลายชนิด รวมถึงวิตามินยาโปรตีนและกรดนิวคลีอิกเช่นดีเอ็นเอตลอดจนไวรัส การวิเคราะห์โครงสร้างด้วยรังสีเอกซ์ยังคงเป็นวิธีการหลักในการระบุโครงสร้างอะตอมของวัสดุและในการจำแนกวัสดุที่ดูคล้ายกันในการทดลองอื่นๆ โครงสร้างผลึกจากรังสีเอกซ์ยังสามารถช่วยอธิบาย คุณสมบัติ ทางอิเล็กทรอนิกส์หรือความยืดหยุ่น ที่ผิดปกติ ของวัสดุ ให้ความกระจ่างเกี่ยวกับปฏิกิริยาและกระบวนการทางเคมี หรือใช้เป็นพื้นฐานในการออกแบบยาเพื่อรักษาโรคต่างๆ
งานวิจัยสมัยใหม่เกี่ยวข้องกับหลายขั้นตอน ซึ่งทุกขั้นตอนล้วนมีความสำคัญ ขั้นตอนเบื้องต้น ได้แก่ การเตรียมตัวอย่างที่มีคุณภาพดี การบันทึกความเข้มของการเลี้ยวเบนอย่างระมัดระวัง และการประมวลผลข้อมูลเพื่อกำจัดสิ่งรบกวน จากนั้นจึงใช้วิธีการต่างๆ เพื่อประมาณโครงสร้างอะตอม ซึ่งโดยทั่วไปเรียกว่าวิธีการโดยตรง เมื่อได้ค่าประมาณเบื้องต้นแล้ว จะใช้เทคนิคการคำนวณเพิ่มเติม เช่น เทคนิคที่เกี่ยวข้องกับแผนที่ความแตกต่าง เพื่อให้ได้โครงสร้างที่สมบูรณ์ ขั้นตอนสุดท้ายคือการปรับตำแหน่งอะตอมเชิงตัวเลขให้สอดคล้องกับข้อมูลการทดลอง ซึ่งบางครั้งอาจได้รับความช่วยเหลือจาก การคำนวณ แบบ ab-initioในเกือบทุกกรณี โครงสร้างใหม่จะถูกบันทึกไว้ในฐานข้อมูลที่เปิดให้ชุมชนนานาชาติเข้าถึงได้
ประวัติศาสตร์


แม้ว่าผลึกจะได้รับการชื่นชมมานานแล้วในเรื่องความสม่ำเสมอและความสมมาตรแต่ก็ยังไม่มีการศึกษาทางวิทยาศาสตร์จนกระทั่งศตวรรษที่ 17 โยฮันเนส เคปเลอร์ตั้งสมมติฐานในงานของเขา เรื่อง Strena seu de Nive Sexangula (ของขวัญปีใหม่จากหิมะรูปหกเหลี่ยม) (1611) ว่าความสมมาตรแบบหกเหลี่ยมของผลึกเกล็ดหิมะเกิดจากการเรียงตัวอย่างเป็นระเบียบของอนุภาคน้ำทรงกลม[ 1 ]นักวิทยาศาสตร์ชาวเดนมาร์กนิโคลัส สเตโน (1669) เป็นผู้บุกเบิกการศึกษาเชิงทดลองเกี่ยวกับความสมมาตรของผลึก สเตโนแสดงให้เห็นว่ามุมระหว่างหน้าต่างๆ มีค่าเท่ากันในทุกตัวอย่างของผลึกชนิดใดชนิดหนึ่ง ( กฎความคงที่ของมุมระหว่างหน้า ) [ 2 ]เรเน จัสต์ เฮาย (1784) ค้นพบว่าทุกหน้าของผลึกสามารถอธิบายได้ด้วยรูปแบบการเรียงซ้อนอย่างง่ายของบล็อกที่มีรูปร่างและขนาดเดียวกัน ( กฎการลดลง ) ดังนั้น ในปี 1839 William Hallowes Millerจึงสามารถกำหนดป้ายกำกับที่ไม่ซ้ำกันให้กับแต่ละหน้าด้วยจำนวนเต็มขนาดเล็กสามตัว ซึ่งก็ คือดัชนี Millerซึ่งยังคงใช้ในการระบุหน้าผลึก การศึกษาของ Haüy นำไปสู่แนวคิดที่ว่าผลึกเป็นอาร์เรย์สามมิติปกติ ( แลตติส Bravais ) ของอะตอมและโมเลกุล โดย เซลล์หน่วยเดียวจะถูกทำซ้ำอย่างไม่มีที่สิ้นสุดตามทิศทางหลักสามทิศทาง ในศตวรรษที่ 19 แคตตาล็อกที่สมบูรณ์ของสมมาตรที่เป็นไปได้ของผลึกได้รับการจัดทำโดยJohan Hessel [ 3 ] Auguste Bravais [ 4 ] Evgraf Fedorov [ 5 ] Arthur Schönflies [ 6 ]และ (ในภายหลัง) William Barlow (1894) Barlow เสนอโครงสร้างผลึกหลาย แบบในช่วงปี 1880 ซึ่งได้รับการตรวจสอบความถูกต้องในภายหลังโดยการวิเคราะห์ผลึกด้วยรังสีเอกซ์[ 7 ]อย่างไรก็ตาม ข้อมูลที่มีอยู่ในช่วงทศวรรษ 1880 นั้นมีน้อยเกินไปที่จะยอมรับแบบจำลองของเขาว่าเป็นข้อสรุป
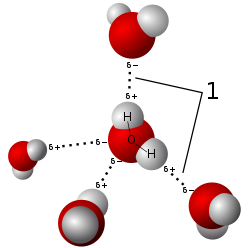
วิลเฮล์ม รอนต์เกนค้นพบรังสีเอ็กซ์ในปี พ.ศ. 2438 [ 8 ]นักฟิสิกส์ไม่แน่ใจในธรรมชาติของรังสีเอ็กซ์ แต่สงสัยว่ามันเป็นคลื่นของรังสีแม่เหล็กไฟฟ้าทฤษฎี ของแม็ กซ์ เวลล์เกี่ยวกับรังสี แม่เหล็กไฟฟ้าได้รับการยอมรับอย่างกว้างขวาง และการทดลองของชาร์ลส์ โกลเวอร์ บาร์คลาแสดงให้เห็นว่ารังสีเอ็กซ์แสดงปรากฏการณ์ที่เกี่ยวข้องกับคลื่นแม่เหล็กไฟฟ้า รวมถึงการโพลาไรเซชัน ตามขวาง และเส้นสเปกตรัมที่คล้ายกับที่สังเกตได้ในความยาวคลื่นที่มองเห็นได้ บาร์คลาสร้างสัญลักษณ์รังสีเอ็กซ์สำหรับเส้นสเปกตรัมที่คมชัด โดยสังเกตพลังงานสองระดับที่แยกจากกันในปี พ.ศ. 2452 ในตอนแรกตั้งชื่อว่า "A" และ "B" จากนั้นสันนิษฐานว่าอาจมีเส้นก่อนหน้า "A" เขาจึงเริ่มใช้ตัวอักษรเริ่มต้นที่ "K" [ 9 ] [ 10 ]การทดลองช่องเดี่ยวในห้องปฏิบัติการของอาร์โนลด์ ซอมเมอร์เฟลด์ชี้ให้เห็นว่ารังสีเอ็กซ์มีความยาวคลื่นประมาณ1 อังสตรอม[ 11 ]รังสีเอกซ์ไม่เพียงแต่เป็นคลื่นเท่านั้น แต่ยังมีคุณสมบัติของอนุภาคด้วย ทำให้ Sommerfeld ตั้งชื่อ สเปกตรัมต่อเนื่องว่า Bremsstrahlungเมื่ออิเล็กตรอนพุ่งชนวัสดุ[ 10 ] Albert Einsteinนำเสนอแนวคิดเรื่องโฟตอนในปี 1905 [ 12 ]แต่แนวคิดนี้ยังไม่ได้รับการยอมรับอย่างกว้างขวางจนกระทั่งปี 1922 [ 13 ] [ 14 ]เมื่อArthur Comptonยืนยันแนวคิดนี้โดยการกระเจิงของรังสีเอกซ์จากอิเล็กตรอน[ 15 ]คุณสมบัติคล้ายอนุภาคของรังสีเอกซ์ เช่น การแตกตัวเป็นไอออนของก๊าซ ทำให้William Henry Braggโต้แย้งในปี 1907 ว่ารังสีเอกซ์ไม่ใช่รังสีแม่เหล็กไฟฟ้า[ 16 ] [ 17 ] [ 18 ] [ 19 ] มุมมองของแบรกก์พิสูจน์แล้วว่าไม่เป็นที่นิยม และการสังเกตการเลี้ยวเบนของรังสีเอกซ์โดยแม็กซ์ ฟอน ลาเวในปี พ.ศ. 2455 [ 20 ]ยืนยันว่ารังสีเอกซ์เป็นรูปแบบหนึ่งของรังสีแม่เหล็กไฟฟ้า

แนวคิดที่ว่าผลึกสามารถใช้เป็นตะแกรงเลี้ยวเบนสำหรับรังสีเอกซ์ได้นั้น เกิดขึ้นในปี 1912 ในการสนทนาระหว่างพอล ปีเตอร์ อีวาลด์และแม็กซ์ ฟอน ลาเวในสวนอังกฤษที่เมืองมิวนิก อีวาลด์ได้เสนอแบบจำลองเรโซเนเตอร์ของผลึกสำหรับวิทยานิพนธ์ของเขา แต่แบบจำลองนี้ไม่สามารถตรวจสอบได้โดยใช้แสงที่มองเห็นได้เนื่องจากความยาวคลื่นนั้นมากกว่าระยะห่างระหว่างเรโซเนเตอร์มาก ฟอน ลาเว ตระหนักว่าจำเป็นต้องใช้รังสีแม่เหล็กไฟฟ้าที่มีความยาวคลื่นสั้นกว่า และเสนอว่ารังสีเอกซ์อาจมีความยาวคลื่นที่เทียบได้กับระยะห่างของหน่วยเซลล์ในผลึก ฟอน ลาเว ทำงานร่วมกับช่างเทคนิคสองคน คือวอลเตอร์ ฟรีดริชและพอล คิปปิง ผู้ช่วยของเขา เพื่อฉายลำแสงรังสีเอกซ์ผ่าน ผลึก คอปเปอร์ซัลเฟตและบันทึกการเลี้ยวเบนลงบนแผ่นฟิล์มถ่ายภาพหลังจากล้างฟิล์มแล้ว แผ่นฟิล์มแสดงจุดที่มีความคมชัดจำนวนมากเรียงตัวเป็นรูปแบบวงกลมตัดกันรอบจุดที่เกิดจากลำแสงกลาง ผลลัพธ์ถูกนำเสนอต่อสถาบันวิทยาศาสตร์และมนุษยศาสตร์แห่งบาวาเรียในเดือนมิถุนายน พ.ศ. 2455 ในชื่อ "Interferenz-Erscheinungen bei Röntgenstrahlen" (ปรากฏการณ์การแทรกสอดในรังสีเอกซ์) [ 20 ] [ 21 ]ฟอน ลาเว ได้พัฒนากฎที่เชื่อมโยงมุมการกระเจิงกับขนาดและการวางแนวของระยะห่างระหว่างหน่วยเซลล์ในผลึก ซึ่งทำให้เขาได้รับรางวัลโนเบลสาขาฟิสิกส์ในปี พ.ศ. 2457 [ 22 ]

หลังจากงานวิจัยบุกเบิกของ Von Laue สาขานี้ได้พัฒนาอย่างรวดเร็ว โดยเฉพาะอย่างยิ่งจากผลงานของนักฟิสิกส์William Lawrence BraggและWilliam Henry Bragg ผู้เป็นบิดา ในปี 1912–1913 Bragg ผู้เป็นบุตรชายได้พัฒนาBragg's lawซึ่งเชื่อมโยงการกระเจิงกับระนาบที่เว้นระยะห่างเท่าๆ กันภายในผลึก[ 8 ] [ 23 ] [ 24 ] [ 25 ] Bragg ทั้งพ่อและลูกได้รับรางวัลโนเบลสาขาฟิสิกส์ในปี 1915 จากผลงานด้านผลึกศาสตร์ โครงสร้างแรกๆ นั้นโดยทั่วไปแล้วจะเรียบง่าย เมื่อวิธีการคำนวณและวิธีการทดลองดีขึ้นในช่วงหลายทศวรรษต่อมา ก็เป็นไปได้ที่จะอนุมานตำแหน่งอะตอมที่เชื่อถือได้สำหรับการจัดเรียงอะตอมที่ซับซ้อนมากขึ้น
โครงสร้างแรกสุดเป็นผลึกอนินทรีย์และแร่ธาตุอย่างง่าย แต่แม้กระทั่งสิ่งเหล่านี้ก็ยังเผยให้เห็นกฎพื้นฐานของฟิสิกส์และเคมี โครงสร้างที่มีความละเอียดระดับอะตอมแรกที่ "ได้รับการแก้ไข" (เช่น กำหนด) ในปี 1914 คือโครงสร้างของเกลือแกง[ 26 ] [ 27 ] [ 28 ] การกระจายตัวของอิเล็กตรอนในโครงสร้างของเกลือแกงแสดงให้เห็นว่าผลึกไม่จำเป็นต้องประกอบด้วย โมเลกุล ที่ยึดเหนี่ยวกันด้วยพันธะโควาเลนต์และพิสูจน์การมีอยู่ของสารประกอบไอออนิก [ 29 ] โครงสร้างของเพชรได้รับการแก้ไขในปีเดียวกัน[ 30 ] [ 31 ]ซึ่งพิสูจน์การจัดเรียงแบบทรงสี่หน้าของพันธะเคมีและแสดงให้เห็นว่าความยาวของพันธะเดี่ยว C–C อยู่ที่ประมาณ 1.52 อังสตรอม โครงสร้างยุคแรกอื่นๆ ได้แก่ ทองแดง[ 32 ]แคลเซียมฟลูออไรด์ (CaF 2หรือที่รู้จักกันในชื่อฟลูออไรต์ ) แคลไซต์ (CaCO 3 ) และไพไรต์ (FeS 2 ) [ 33 ]ในปี พ.ศ. 2457; สปิเนล (MgAl 2 O 4 ) ในปี พ.ศ. 2458; [ 34 ] [ 35 ]รูป แบบ รูไทล์และอนาเทสของไทเทเนียมไดออกไซด์ (TiO 2 ) ในปี พ.ศ. 2459; [ 36 ]ไพโรครอยต์ (Mn(OH) 2 ) และขยายความไปถึงบรูไซต์ (Mg(OH) 2 ) ในปี พ.ศ. 2462 [ 37 ] [ 38 ]นอกจากนี้ ในปี พ.ศ. 2462 โซเดียมไนเตรต (NaNO 3 ) และซีเซียมไดคลอโรไอโอเดต( CsICl 2 ) ถูกกำหนดโดยRalph Walter Graystone Wyckoff [ 39 ]และ โครงสร้าง เวิร์ตไซต์(ZnS หกเหลี่ยม) ถูกกำหนดในปี พ.ศ. 2463 [ 40 ]
โครงสร้างของกราไฟต์ได้รับการไขปริศนาในปี พ.ศ. 2459 [ 41 ]โดยวิธีการเลี้ยวเบนของผงที่เกี่ยวข้อง[ 42 ]ซึ่งพัฒนาโดยปีเตอร์ เดบายและพอล เชอร์เรอร์และโดยอัลเบิร์ต ฮัลล์ อย่างอิสระ ในปี พ.ศ. 2460 [ 43 ]โครงสร้างของกราไฟต์ได้รับการกำหนดจากการเลี้ยวเบนของผลึกเดี่ยวในปี พ.ศ. 2467 โดยสองกลุ่ม อย่างอิสระ [ 44 ] [ 45 ]ฮัลล์ยังใช้วิธีผงเพื่อกำหนดโครงสร้างของโลหะต่างๆ เช่น เหล็ก[ 46 ]และแมกนีเซียม[ 47 ]
ผลกระทบต่อวิชาเคมี
การตกผลึกด้วยรังสีเอกซ์นำไปสู่ความเข้าใจที่ดีขึ้นเกี่ยวกับพันธะเคมีและปฏิสัมพันธ์ที่ไม่ใช่โคเวเลนต์การศึกษาเบื้องต้นเผยให้เห็นรัศมีทั่วไปของอะตอม และยืนยันแบบจำลองทางทฤษฎีของพันธะเคมีหลายแบบ เช่น พันธะเตตระเฮดรัลของคาร์บอนในโครงสร้างเพชร[ 30 ]พันธะออกตาเฮดรัลของโลหะที่พบในแอมโมเนียมเฮกซาคลอโรแพลทิเนต (IV) [ 48 ]และเรโซแนนซ์ที่พบในกลุ่มคาร์บอเนตแบบระนาบ[ 33 ]และในโมเลกุลอะโรมาติก[ 49 ] โครงสร้างของ เฮกซาเมทิลเบนซีนในปี 1928 ของKathleen Lonsdale [ 50 ]ได้สร้างสมมาตรหกเหลี่ยมของเบนซีนและแสดงให้เห็นความแตกต่างที่ชัดเจนในความยาวพันธะระหว่างพันธะ C–C แบบอะลิฟาติกและพันธะ C–C แบบอะโรมาติก การค้นพบนี้ทำให้เกิดแนวคิดเรื่องเรโซแนนซ์ระหว่างพันธะเคมี ซึ่งมีผลกระทบอย่างมากต่อการพัฒนาเคมี[ 51 ]ข้อสรุปของเธอได้รับการคาดการณ์ไว้ล่วงหน้าโดยWilliam Henry Braggซึ่งตีพิมพ์แบบจำลองของแนฟทาลีนและแอนทราซีนในปี พ.ศ. 2464 โดยอิงจากโมเลกุลอื่น ซึ่งเป็นรูปแบบแรกเริ่มของการแทนที่โมเลกุล[ 49 ] [ 52 ]
โครงสร้างแรกของสารประกอบอินทรีย์เฮกซาเมทิลีนเตตรามีนได้รับการแก้ไขในปี พ.ศ. 2466 [ 53 ] ตามมาอย่างรวดเร็วด้วยการศึกษา กรดไขมันสายยาวต่างๆซึ่งเป็นส่วนประกอบสำคัญของเยื่อชีวภาพ[ 54 ] [ 55 ] [ 56 ] [ 57 ] [ 58 ] [ 59 ] [ 60 ] [ 61 ] [ 62 ] ในช่วงทศวรรษ พ.ศ. 2473โครงสร้างของโมเลกุลขนาดใหญ่ที่มีความซับซ้อนแบบสองมิติเริ่มได้รับการแก้ไข ความก้าวหน้าที่สำคัญคือโครงสร้างของฟทาโลไซยานีน[ 63 ]ซึ่งเป็นโมเลกุลระนาบขนาดใหญ่ที่เกี่ยวข้องอย่างใกล้ชิดกับโมเลกุลพอร์ไฟริน ที่สำคัญใน ทางชีววิทยา เช่นฮีมคอร์รินและคลอโรฟิลล์
ในช่วงทศวรรษ 1920 วิคเตอร์ มอริตซ์ โกลด์ชมิดท์และต่อมาไลนัส พอลลิงได้พัฒนากฎเกณฑ์สำหรับการกำจัดโครงสร้างที่ไม่น่าจะเป็นไปได้ทางเคมี และสำหรับการกำหนดขนาดสัมพัทธ์ของอะตอม กฎเกณฑ์เหล่านี้ได้นำไปสู่โครงสร้างของบรูไคต์ (1928) และความเข้าใจเกี่ยวกับความเสถียรสัมพัทธ์ ของ ไทเทเนียมไดออกไซด์ในรูปรูไทล์บรูไคต์และอนาเทส
ระยะห่างระหว่างอะตอมที่เชื่อมต่อกันสองอะตอมเป็นการวัดที่ไวต่อความแข็งแรงของพันธะและลำดับของพันธะดังนั้น การศึกษาผลึกศาสตร์ด้วยรังสีเอกซ์จึงนำไปสู่การค้นพบพันธะประเภทแปลกใหม่ยิ่งขึ้นในเคมีอนินทรีย์เช่น พันธะคู่โลหะ-โลหะ[ 64 ] [ 65 ] [ 66 ]พันธะสี่เท่าโลหะ-โลหะ[ 67 ] [ 68 ] [ 69 ]และพันธะสามศูนย์สองอิเล็กตรอน [ 70 ] ผลึกศาสตร์ด้วยรังสีเอกซ์ หรือที่จริงแล้วคือ การทดลอง การกระเจิงคอมป์ตัน แบบไม่ยืดหยุ่น ยังได้ให้หลักฐานเกี่ยวกับลักษณะกึ่งโควาเลนต์ของพันธะไฮโดรเจน อีกด้วย [ 71 ]ในสาขาเคมีออร์กาโนเมทัลลิกโครงสร้างรังสีเอกซ์ของเฟอร์โรซีนได้เริ่มต้นการศึกษาทางวิทยาศาสตร์เกี่ยวกับสารประกอบแซนด์วิช [ 72 ] [ 73 ]ในขณะที่โครงสร้างของเกลือ Zeise กระตุ้นการวิจัยเกี่ยวกับ "พันธะย้อนกลับ" และสารประกอบ เชิงซ้อนโลหะ-ไพ[ 74 ] [ 75 ] [ 76 ] [ 77 ]ในที่สุด ผลึกศาสตร์รังสีเอกซ์ก็มีบทบาทสำคัญในการพัฒนาเคมีระดับเหนือโมเลกุลโดยเฉพาะอย่างยิ่งในการชี้แจงโครงสร้างของคราวน์อีเทอร์และหลักการของเคมีโฮสต์-เกสต์
ระบบ อนินทรีย์และออร์กาโนเมทัลลิกที่ซับซ้อนจำนวนมากได้รับการวิเคราะห์โดยใช้วิธีการผลึกเดี่ยว เช่นฟูล เลอรี นเมทัลโลพอร์ไฟรินและสารประกอบที่ซับซ้อนอื่นๆ การเลี้ยวเบนของผลึกเดี่ยวยังถูกนำมาใช้ในอุตสาหกรรมยาด้วย ฐานข้อมูลโครงสร้างเคมบริดจ์มีโครงสร้างมากกว่า 1,000,000 โครงสร้าง ณ เดือนมิถุนายน 2019 โดยโครงสร้างส่วนใหญ่เหล่านี้ได้รับการกำหนดโดยการวิเคราะห์ผลึกด้วยรังสีเอกซ์[ 78 ]
ผลกระทบต่อแร่ธาตุวิทยา

การประยุกต์ใช้ผลึกศาสตร์รังสีเอกซ์กับแร่ธาตุวิทยาเริ่มต้นด้วยโครงสร้างของแร่การ์เนตซึ่งถูกกำหนดโดยเมนเซอร์ในปี 1924 การศึกษาผลึกศาสตร์รังสีเอกซ์อย่างเป็นระบบของซิลิเกตได้ดำเนินการในช่วงทศวรรษที่ 1920 การศึกษานี้แสดงให้เห็นว่า เมื่อ อัตราส่วน Si / Oเปลี่ยนไป ผลึกซิลิเกตจะแสดงการเปลี่ยนแปลงอย่างมีนัยสำคัญในการจัดเรียงอะตอม มาชาตช์กีได้ขยายความเข้าใจเหล่านี้ไปยังแร่ธาตุที่อะลูมิเนียมเข้ามาแทนที่ อะตอม ซิลิคอนของซิลิเกต การประยุกต์ใช้ผลึกศาสตร์รังสีเอกซ์ครั้งแรกกับโลหะวิทยาก็เกิดขึ้นในช่วงกลางทศวรรษที่ 1920 เช่นกัน[ 80 ] [ 81 ] [ 82 ] [ 83 ] [ 84 ] [ 85 ]ที่โดดเด่นที่สุดคือโครงสร้างของโลหะผสม Mg 2 Sn ของไล นัส พอลลิง[ 86 ]นำไปสู่ทฤษฎีของเขาเกี่ยวกับความเสถียรและโครงสร้างของผลึกไอออนิกเชิงซ้อน[ 87 ]
เมื่อวันที่ 17 ตุลาคม พ.ศ. 2555 ยานสำรวจ Curiosityบนดาวอังคารที่ " Rocknest " ได้ทำการวิเคราะห์การเลี้ยวเบนของรังสีเอกซ์ของดินบนดาวอังคารเป็นครั้งแรกผลลัพธ์จากเครื่องวิเคราะห์ CheMin ของยานสำรวจเผยให้เห็นการมีอยู่ของแร่ธาตุหลายชนิด รวมถึงเฟลด์สปาร์ ไพรอกซีน และโอลิวีนและชี้ให้เห็นว่าดินบนดาวอังคารในตัวอย่างนั้นคล้ายกับ " ดินบะซอลต์ ที่ผุพัง " ของภูเขาไฟฮาวาย[ 79 ]

ผลกระทบต่อชีววิทยา
การศึกษาโครงสร้างผลึกด้วยรังสีเอกซ์ของโมเลกุลทางชีวภาพเริ่มต้นขึ้นด้วย ผลงานของ Dorothy Crowfoot Hodgkinผู้ซึ่งไขโครงสร้างของคอเลสเตอรอล (1937) เพนิซิลลิน (1946) และวิตามินบี12 (1956) ซึ่งทำให้เธอได้รับรางวัลโนเบลสาขาเคมีในปี 1964 ในปี 1969 เธอประสบความสำเร็จในการไขโครงสร้างของอินซูลินซึ่งเธอทำงานมานานกว่าสามสิบปี[ 88 ]
โครงสร้างผลึกของโปรตีน (ซึ่งมีรูปร่างไม่สม่ำเสมอและมีขนาดใหญ่กว่าคอเลสเตอรอลหลายร้อยเท่า) เริ่มได้รับการแก้ไขในช่วงปลายทศวรรษ 1950 โดยเริ่มจากโครงสร้างของ ไมโอโกลบินจาก วาฬสเปิร์มโดยเซอร์จอห์น โควดรีย์ เคนดรูว์ [ 89 ] ซึ่งทำให้เขาได้รับรางวัลโนเบลสาขาเคมี ร่วม กับแม็กซ์ เพรุตซ์ในปี 1962 [ 90 ]นับตั้งแต่ความสำเร็จนั้น โครงสร้างผลึกเอ็กซ์เรย์ของโปรตีน กรดนิวคลีอิก และโมเลกุลทางชีวภาพอื่นๆ ได้รับการกำหนดแล้ว 190,000 โครงสร้าง[ 91 ]วิธีการที่ใกล้เคียงที่สุดในแง่ของจำนวนโครงสร้างที่วิเคราะห์คือสเปกโทรสโกปีนิวเคลียร์แมกเนติกเรโซแนนซ์ (NMR)ซึ่งแก้ไขได้น้อยกว่าหนึ่งในสิบของจำนวนดังกล่าว[ 92 ]ผลึกศาสตร์สามารถแก้ไขโครงสร้างของโมเลกุลขนาดใหญ่ได้ตามต้องการ ในขณะที่ NMR ในสถานะสารละลายถูกจำกัดไว้ที่โมเลกุลขนาดเล็ก (น้อยกว่า)70 kDa ) การวิเคราะห์โครงสร้างผลึกด้วยรังสีเอกซ์ถูกนำมาใช้เป็นประจำเพื่อตรวจสอบว่ายาออกฤทธิ์กับโปรตีนเป้าหมายอย่างไร และการเปลี่ยนแปลงใดบ้างที่อาจช่วยปรับปรุงประสิทธิภาพได้[ 93 ]อย่างไรก็ตามโปรตีนเยื่อหุ้ม เซลล์ ยังคงเป็นเรื่องท้าทายในการตกผลึก เนื่องจากต้องใช้สารซักฟอกหรือสารทำให้เสียสภาพ อื่นๆ เพื่อละลายโปรตีนเหล่านั้นในสภาวะแยกเดี่ยว และสารซักฟอกดังกล่าวมักจะรบกวนการตกผลึก โปรตีนเยื่อหุ้มเซลล์เป็นส่วนประกอบขนาดใหญ่ของจีโนมและรวมถึงโปรตีนที่มีความสำคัญทางสรีรวิทยาอย่างมากหลายชนิด เช่นช่องไอออนและตัวรับ [ 94 ] [ 95 ] การแช่แข็งด้วยฮีเลียมถูกนำมาใช้เพื่อลดความเสียหายจากรังสีในผลึกโปรตีน[ 96 ]
วิธีการ
ภาพรวม

โดยทั่วไปมักใช้กรณีจำกัดสองกรณีของการตกผลึกด้วยรังสีเอกซ์ ได้แก่ "การตกผลึกโมเลกุลขนาดเล็ก" (ซึ่งรวมถึงของแข็งอนินทรีย์ต่อเนื่อง) และการตกผลึก "โมเลกุลขนาดใหญ่" การตกผลึกโมเลกุลขนาดเล็กมักเกี่ยวข้องกับผลึกที่มีอะตอมน้อยกว่า 100 อะตอมในหน่วยอสมมาตรโครงสร้างผลึกดังกล่าวโดยทั่วไปมีความละเอียดดีมากจนสามารถแยกแยะอะตอมได้เป็น "กลุ่ม" ของความหนาแน่นอิเล็กตรอนที่แยกจากกัน ในทางตรงกันข้าม การตกผลึกโมเลกุลขนาดใหญ่มักเกี่ยวข้องกับอะตอมหลายหมื่นอะตอมในเซลล์หน่วย โครงสร้างผลึกดังกล่าวโดยทั่วไปมีความละเอียดน้อยกว่า อะตอมและพันธะเคมีปรากฏเป็นท่อของความหนาแน่นอิเล็กตรอน แทนที่จะเป็นอะตอมที่แยกจากกัน โดยทั่วไป โมเลกุลขนาดเล็กก็ตกผลึกได้ง่ายกว่าโมเลกุลขนาดใหญ่ อย่างไรก็ตาม การตกผลึกด้วยรังสีเอกซ์ได้พิสูจน์แล้วว่าสามารถทำได้แม้กระทั่งกับไวรัสและโปรตีนที่มีอะตอมหลายแสนอะตอม ผ่านการสร้างภาพและเทคโนโลยีการตกผลึกที่ดีขึ้น[ 97 ]
เทคนิคการวิเคราะห์ผลึกเดี่ยวด้วยรังสีเอกซ์มีขั้นตอนพื้นฐานสามขั้นตอน ขั้นตอนแรก—และมักจะเป็นขั้นตอนที่ยากที่สุด—คือการได้ผลึกที่เหมาะสมของวัสดุที่กำลังศึกษา ผลึกควรมีขนาดใหญ่พอสมควร (โดยทั่วไปใหญ่กว่า 0.1 มม. ในทุกมิติ) มีองค์ประกอบที่บริสุทธิ์และมีโครงสร้างที่สม่ำเสมอ โดยไม่มีข้อบกพร่อง ภายในที่สำคัญ เช่น รอยแตกหรือการเกิดแฝด[ 98 ]
ในขั้นตอนที่สอง ผลึกจะถูกวางไว้ในลำแสงเอ็กซ์เรย์ที่มีความเข้มสูง ซึ่งโดยปกติจะเป็นความยาวคลื่นเดียว ( เอ็กซ์เรย์โมโนโครมาติก ) ทำให้เกิดรูปแบบการสะท้อนที่สม่ำเสมอ มุมและความเข้มของเอ็กซ์เรย์ที่เลี้ยวเบนจะถูกวัด โดยแต่ละสารประกอบจะมีรูปแบบการเลี้ยวเบนที่ไม่ซ้ำกัน[ 99 ]เมื่อผลึกหมุนไปทีละน้อย การสะท้อนก่อนหน้าจะหายไปและการสะท้อนใหม่จะปรากฏขึ้น ความเข้มของทุกจุดจะถูกบันทึกไว้ในทุกทิศทางของผลึก อาจต้องรวบรวมชุดข้อมูลหลายชุด โดยแต่ละชุดจะครอบคลุมการหมุนของผลึกมากกว่าครึ่งรอบเล็กน้อย และโดยทั่วไปจะมีค่าการสะท้อนหลายหมื่นค่า[ 100 ]
ในขั้นตอนที่สาม ข้อมูลเหล่านี้จะถูกรวมเข้าด้วยกันทางคอมพิวเตอร์พร้อมกับข้อมูลทางเคมีเสริมเพื่อสร้างและปรับปรุงแบบจำลองการจัดเรียงอะตอมภายในผลึก แบบจำลองการจัดเรียงอะตอมที่ปรับปรุงแล้วขั้นสุดท้าย—ซึ่งตอนนี้เรียกว่าโครงสร้างผลึก —มักจะถูกเก็บไว้ในฐานข้อมูลสาธารณะ[ 101 ]
การตกผลึก

แม้ว่าผลึกศาสตร์จะสามารถใช้เพื่อระบุลักษณะความไม่เป็นระเบียบในผลึกที่ไม่บริสุทธิ์หรือไม่สม่ำเสมอได้ แต่โดยทั่วไปแล้วผลึกศาสตร์ต้องการผลึกบริสุทธิ์ที่มีความสม่ำเสมอสูงเพื่อแก้โครงสร้างของการจัดเรียงอะตอมที่ซับซ้อน ผลึกบริสุทธิ์ที่มีความสม่ำเสมอสามารถหาได้จากวัสดุธรรมชาติหรือวัสดุสังเคราะห์ เช่น ตัวอย่างโลหะ แร่ธาตุ หรือวัสดุขนาดใหญ่อื่นๆ ความสม่ำเสมอของผลึกดังกล่าวสามารถปรับปรุงได้ด้วยการอบผลึก โมเลกุลขนาดใหญ่ [ 102 ] [ 103 ] [ 104 ]และวิธีการอื่นๆ อย่างไรก็ตาม ในหลายกรณี การได้ผลึกที่มีคุณภาพสำหรับการเลี้ยวเบนถือเป็นอุปสรรคสำคัญในการแก้โครงสร้างที่มีความละเอียดระดับอะตอม[ 105 ]
การศึกษาโครงสร้างผลึกของโมเลกุลขนาดเล็กและโมเลกุลขนาดใหญ่แตกต่างกันในขอบเขตของเทคนิคที่สามารถใช้ในการสร้างผลึกที่มีคุณภาพสำหรับการเลี้ยวเบนของรังสีเอกซ์ โมเลกุลขนาดเล็กโดยทั่วไปมีระดับความเป็นอิสระในการเปลี่ยนแปลงรูปร่างน้อย และสามารถตกผลึกได้ด้วยวิธีการที่หลากหลาย เช่น การตกตะกอนด้วยไอสารเคมีและการตกผลึกซ้ำในทางตรงกันข้าม โมเลกุลขนาดใหญ่โดยทั่วไปมีระดับความเป็นอิสระหลายระดับ และการตกผลึกของพวกมันจะต้องดำเนินการโดยรักษาสภาพโครงสร้างที่เสถียร ตัวอย่างเช่น โปรตีนและ โมเลกุล RNA ขนาดใหญ่ ไม่สามารถตกผลึกได้หากโครงสร้างระดับตติยภูมิของพวกมันคลี่ออกดังนั้น ขอบเขตของสภาวะการตกผลึกจึงจำกัดอยู่เฉพาะในสภาวะสารละลายที่โมเลกุลดังกล่าวคงสภาพพับอยู่

ผลึกโปรตีนมักจะเติบโตในสารละลาย วิธีที่พบได้บ่อยที่สุดคือการลดความสามารถในการละลายของโมเลกุลที่เป็นส่วนประกอบลงอย่างค่อยเป็นค่อยไป หากทำเร็วเกินไป โมเลกุลจะตกตะกอนออกจากสารละลาย กลายเป็นฝุ่นหรือเจลอสัณฐานที่ไร้ประโยชน์อยู่ที่ก้นภาชนะ การเติบโตของผลึกในสารละลายมีลักษณะเป็นสองขั้นตอน คือการเกิดนิวเคลียสของผลึกขนาดเล็ก (อาจมีเพียง 100 โมเลกุล) ตามด้วยการเติบโตของผลึกนั้น โดยในอุดมคติแล้วควรเป็นผลึกที่มีคุณภาพสำหรับการเลี้ยวเบน[ 106 ] [ 107 ]สภาวะของสารละลายที่เอื้อต่อขั้นตอนแรก (การเกิดนิวเคลียส) ไม่จำเป็นต้องเป็นสภาวะเดียวกันกับที่เอื้อต่อขั้นตอนที่สอง (การเติบโตในภายหลัง) สภาวะของสารละลายควรไม่เอื้อต่อขั้นตอนแรก (การเกิดนิวเคลียส) แต่ควรเอื้อต่อขั้นตอนที่สอง (การเติบโต) เพื่อให้เกิดผลึกขนาดใหญ่เพียงหนึ่งเดียวต่อหยด หากการเกิดนิวเคลียสเอื้อมากเกินไป จะเกิดผลึกขนาดเล็กจำนวนมากในหยดแทนที่จะเป็นผลึกขนาดใหญ่หนึ่งเดียว หากเอื้อน้อยเกินไป จะไม่มีผลึกเกิดขึ้นเลย แนวทางอื่นๆ เกี่ยวข้องกับการตกผลึกโปรตีนภายใต้น้ำมัน โดยที่สารละลายโปรตีนในน้ำจะถูกกระจายภายใต้น้ำมันเหลว และน้ำจะระเหยผ่านชั้นน้ำมัน น้ำมันชนิดต่างๆ มีค่าการซึมผ่านของการระเหยที่แตกต่างกัน ดังนั้นจึงทำให้เกิดการเปลี่ยนแปลงอัตราความเข้มข้นจากส่วนผสมของสารรับ/โปรตีนที่แตกต่างกัน[ 108 ]
เป็นการยากที่จะคาดการณ์สภาวะที่ดีสำหรับการเกิดนิวเคลียสหรือการเติบโตของผลึกที่มีระเบียบ[ 109 ]ในทางปฏิบัติ สภาวะที่เหมาะสมจะถูกระบุโดยการคัดกรองโดยจะเตรียมโมเลกุลจำนวนมาก และทดสอบสารละลายการตกผลึกที่หลากหลาย[ 110 ]โดยทั่วไปแล้วจะต้องทดลองสภาวะของสารละลายหลายร้อยหรือหลายพันสภาวะก่อนที่จะพบสภาวะที่ประสบความสำเร็จ สภาวะต่างๆ สามารถใช้กลไกทางกายภาพอย่างน้อยหนึ่งอย่างเพื่อลดความสามารถในการละลายของโมเลกุล ตัวอย่างเช่น บางสภาวะอาจเปลี่ยนค่า pH บางสภาวะมีเกลือในอนุกรม Hofmeisterหรือสารเคมีที่ลดค่าคงที่ไดอิเล็กตริกของสารละลาย และบางสภาวะมีพอลิเมอร์ขนาดใหญ่ เช่นโพลีเอทิลีนไกลคอล ที่ผลักโมเลกุลออกจากสารละลายด้วยผลกระทบทางเอนโทรปี นอกจากนี้ยังเป็นเรื่องปกติที่จะลองอุณหภูมิหลายๆ ระดับเพื่อกระตุ้นการตกผลึก หรือค่อยๆ ลดอุณหภูมิลงเพื่อให้สารละลายอิ่มตัวยิ่งยวด วิธีการเหล่านี้ต้องการโมเลกุลเป้าหมายจำนวนมาก เนื่องจากใช้ความเข้มข้นสูงของโมเลกุลที่จะตกผลึก เนื่องจากความยากลำบากในการได้รับโปรตีนเกรดการตกผลึกใน ปริมาณมาก ( มิลลิกรัม ) จึงได้มีการพัฒนาหุ่นยนต์ที่สามารถจ่ายหยดทดลองการตกผลึกได้อย่างแม่นยำ ซึ่งมีปริมาตรประมาณ 100 นาโนลิตรหมายความว่าใช้โปรตีนน้อยลง 10 เท่าต่อการทดลอง เมื่อเทียบกับการทดลองการตกผลึกที่จัดเตรียมด้วยมือ (ประมาณ 1 ไมโครลิตร ) [ 111 ]
มีหลายปัจจัยที่ทราบกันดีว่าสามารถยับยั้งการตกผลึกได้ โดยทั่วไปแล้ว ผลึกที่กำลังเติบโตจะถูกเก็บไว้ที่อุณหภูมิคงที่และป้องกันจากแรงกระแทกหรือการสั่นสะเทือนที่อาจรบกวนการตกผลึก สารเจือปนในโมเลกุลหรือในสารละลายสำหรับการตกผลึกมักเป็นอุปสรรคต่อการตกผลึก ความยืดหยุ่นของโครงสร้างในโมเลกุลก็มีแนวโน้มที่จะทำให้การตกผลึกเกิดขึ้นได้ยากขึ้นเช่นกัน เนื่องจากเอนโทรปี โมเลกุลที่มักจะรวมตัวกันเป็นเกลียวปกติมักจะไม่เต็มใจที่จะรวมตัวกันเป็นผลึก ผลึกอาจเสียหายได้จากการเกิดผลึกแฝดซึ่งอาจเกิดขึ้นเมื่อเซลล์หน่วยสามารถจัดเรียงตัวได้อย่างเหมาะสมในหลายทิศทาง แม้ว่าความก้าวหน้าล่าสุดในวิธีการคำนวณอาจช่วยให้สามารถแก้โครงสร้างของผลึกแฝดบางชนิดได้ก็ตาม เมื่อไม่สามารถตกผลึกโมเลกุลเป้าหมายได้ นักผลึกศาสตร์อาจลองอีกครั้งด้วยโมเลกุลที่มีการดัดแปลงเล็กน้อย แม้แต่การเปลี่ยนแปลงเล็กน้อยในคุณสมบัติของโมเลกุลก็อาจนำไปสู่ความแตกต่างอย่างมากในพฤติกรรมการตกผลึกได้
สำหรับโปรตีน พบว่าสิ่งเจือปนบางครั้งอาจช่วยส่งเสริมและบางครั้งอาจขัดขวางการเติบโตของผลึก การสั่นด้วยเสียงที่ได้ยินบางครั้งก็ได้ผล ปัญหาเหล่านี้อาจเกี่ยวข้องกับ ลักษณะ การแยกเฟสของการตกผลึกของโปรตีน[ 112 ]
การเก็บรวบรวมข้อมูล
การติดตั้งคริสตัล
ผลึกจะถูกติดตั้งเพื่อการวัดเพื่อให้สามารถยึดไว้ในลำแสงเอ็กซ์เรย์และหมุนได้ มีวิธีการติดตั้งหลายวิธี ในอดีต ผลึกจะถูกบรรจุลงในหลอดแก้วขนาดเล็กพร้อมกับสารละลายตกผลึก ( ของเหลวแม่ ) ผลึกของโมเลกุลขนาดเล็กมักจะติดด้วยน้ำมันหรือกาวกับเส้นใยแก้วหรือห่วง ซึ่งทำจากไนลอนหรือพลาสติกและติดอยู่กับแท่งแข็ง ผลึกโปรตีนจะถูกตักขึ้นด้วยห่วง จากนั้นแช่แข็งอย่างรวดเร็วด้วยไนโตรเจนเหลว[ 113 ] การแช่แข็งนี้ช่วยลดความเสียหายจากรังสีเอ็กซ์เรย์ เช่นเดียวกับการเคลื่อนที่ทางความร้อน ( ปรากฏการณ์เดบาย-วอลเลอร์) อย่างไรก็ตาม ผลึกโปรตีนที่ไม่ได้รับการบำบัดมักจะแตกหากแช่แข็งอย่างรวดเร็ว ดังนั้นโดยทั่วไปจึงต้องแช่ในสารละลายป้องกันการแข็งตัวก่อนการแช่แข็ง[ 114 ]การแช่ล่วงหน้านี้อาจทำให้ผลึกแตก ทำให้เสียหายสำหรับการศึกษาผลึกศาสตร์ โดยทั่วไป สภาวะการแช่แข็งที่ประสบความสำเร็จจะถูกระบุโดยการลองผิดลองถูก
หลอดแคปิลลารีหรือห่วงจะถูกติดตั้งบนเครื่องวัดมุม (goniometer ) ซึ่งช่วยให้สามารถวางตำแหน่งได้อย่างแม่นยำภายในลำแสงเอ็กซ์เรย์และหมุนได้ เนื่องจากทั้งผลึกและลำแสงมักมีขนาดเล็กมาก ผลึกจึงต้องอยู่ตรงกลางลำแสงด้วยความแม่นยำประมาณ 25 ไมโครเมตร ซึ่งทำได้โดยใช้กล้องที่โฟกัสไปที่ผลึก เครื่องวัดมุมที่พบได้บ่อยที่สุดคือ "เครื่องวัดมุมแคปปา" (kappa goniometer) ซึ่งมีมุมการหมุนสามมุม ได้แก่ มุม ω ซึ่งหมุนรอบแกนที่ตั้งฉากกับลำแสง มุม κ ซึ่งหมุนรอบแกนที่ทำมุมประมาณ 50° กับแกน ω และสุดท้าย มุม φ รอบแกนของห่วง/หลอดแคปิลลารี เมื่อมุม κ เป็นศูนย์ แกน ω และ φ จะอยู่ในแนวเดียวกัน การหมุนแบบ κ ช่วยให้การติดตั้งผลึกสะดวกยิ่งขึ้น เนื่องจากแขนที่ติดตั้งผลึกสามารถเหวี่ยงออกไปทางนักผลึกศาสตร์ได้ การสั่นสะเทือนที่เกิดขึ้นระหว่างการเก็บข้อมูล (ที่กล่าวถึงด้านล่าง) เกี่ยวข้องเฉพาะแกน ω เท่านั้น เครื่องวัดมุมแบบเก่าคือเครื่องวัดมุมสี่วง และเครื่องมือที่เกี่ยวข้อง เช่น เครื่องวัดมุมหกวง
บันทึกความคิดสะท้อน

ความเข้มสัมพัทธ์ของการสะท้อนแสงให้ข้อมูลเพื่อกำหนดการจัดเรียงของโมเลกุลภายในผลึกในรายละเอียดระดับอะตอม ความเข้มของการสะท้อนแสงเหล่านี้สามารถบันทึกได้ด้วยฟิล์มถ่ายภาพตัวตรวจจับพื้นที่ (เช่นตัวตรวจจับพิกเซล ) หรือด้วย เซ็นเซอร์ภาพแบบ CCD ( charge-coupled device ) ยอดที่มุมเล็กๆ สอดคล้องกับข้อมูลความละเอียดต่ำ ในขณะที่ยอดที่มุมสูงแสดงถึงข้อมูลความละเอียดสูง ดังนั้นจึงสามารถกำหนดขีดจำกัดสูงสุดของความละเอียดของโครงสร้างได้จากภาพไม่กี่ภาพแรก สามารถกำหนดมาตรวัดคุณภาพการเลี้ยวเบนบางอย่างได้ในขั้นตอนนี้ เช่นความเป็นโมเสกของผลึกและความไม่เป็นระเบียบโดยรวม ซึ่งสังเกตได้จากความกว้างของยอด นอกจากนี้ยังสามารถวินิจฉัยความผิดปกติบางอย่างของผลึกที่ทำให้ไม่เหมาะสมสำหรับการแก้โครงสร้างได้อย่างรวดเร็วในขั้นตอนนี้
จุดข้อมูลเพียงชุดเดียวไม่เพียงพอที่จะสร้างภาพผลึกทั้งหมดขึ้นมาใหม่ได้ มันเป็นเพียงส่วนเล็กๆ ของชุดข้อมูลสามมิติทั้งหมด ในการรวบรวมข้อมูลที่จำเป็นทั้งหมด ผลึกจะต้องถูกหมุนทีละขั้นจนครบ 180° โดยบันทึกภาพในทุกขั้นตอน อันที่จริงแล้ว ต้องหมุนมากกว่า 180° เล็กน้อยเพื่อให้ครอบคลุมปริภูมิผกผันเนื่องจากความโค้งของทรงกลมอีวาลด์อย่างไรก็ตาม หากผลึกมีความสมมาตรสูง อาจบันทึกภาพในช่วงมุมที่แคบกว่า เช่น 90° หรือ 45° ได้ ควรเปลี่ยนแกนการหมุนอย่างน้อยหนึ่งครั้ง เพื่อหลีกเลี่ยงการเกิด "จุดบอด" ในปริภูมิผกผันใกล้กับแกนการหมุน โดยทั่วไปแล้ว จะโยกผลึกเล็กน้อย (ประมาณ 0.5–2°) เพื่อจับภาพในปริภูมิผกผันที่กว้างขึ้น
อาจจำเป็นต้องใช้ชุดข้อมูลหลายชุดสำหรับ วิธี การกำหนดเฟส บาง วิธี ตัวอย่างเช่น การกำหนดเฟส การกระจายตัวผิดปกติแบบหลายความยาวคลื่นจำเป็นต้องบันทึกการกระเจิงอย่างน้อยสามความยาวคลื่น (และโดยปกติสี่ความยาวคลื่น เพื่อความซ้ำซ้อน) ของรังสีเอกซ์ที่เข้ามา ผลึกเดี่ยวอาจเสื่อมสภาพมากเกินไปในระหว่างการเก็บรวบรวมชุดข้อมูลหนึ่งชุดเนื่องจากความเสียหายจากรังสี ในกรณีเช่นนี้ จะต้องเก็บรวบรวมชุดข้อมูลจากผลึกหลายชิ้น[ 115 ]
สมมาตรของผลึก หน่วยเซลล์ และการปรับขนาดภาพ
ชุดรูปแบบการเลี้ยวเบนสองมิติที่บันทึกไว้ ซึ่งแต่ละรูปแบบสอดคล้องกับการวางแนวผลึกที่แตกต่างกัน จะถูกแปลงเป็นชุดสามมิติ การประมวลผลข้อมูลเริ่มต้นด้วยการจัดทำดัชนีการสะท้อน ซึ่งหมายถึงการระบุขนาดของเซลล์หน่วยและจุดสูงสุดของภาพใดที่สอดคล้องกับตำแหน่งใดในปริภูมิผกผัน ผลพลอยได้ของการจัดทำดัชนีคือการกำหนดสมมาตรของผลึก กล่าวคือ กลุ่มปริภูมิของมันกลุ่มปริภูมิบางกลุ่มสามารถกำจัดได้ตั้งแต่เริ่มต้น ตัวอย่างเช่น ไม่สามารถสังเกตสมมาตรการสะท้อนในโมเลกุลไครัลได้ ดังนั้นจึงอนุญาตให้มีเพียง 65 กลุ่มปริภูมิจาก 230 กลุ่มที่เป็นไปได้สำหรับโมเลกุลโปรตีนซึ่งเกือบทั้งหมดเป็นไครัล การจัดทำดัชนีโดยทั่วไปทำได้โดยใช้รูทีนการจัดทำดัชนีอัตโนมัติ[ 116 ]เมื่อกำหนดสมมาตรแล้ว ข้อมูลจะถูกรวมเข้าด้วยกัน กระบวนการ นี้จะแปลงภาพหลายร้อยภาพที่มีการสะท้อนแสงหลายพันจุดให้เป็นไฟล์เดียว ซึ่งประกอบด้วย (อย่างน้อยที่สุด) บันทึกค่าดัชนีมิลเลอร์ของการสะท้อนแสงแต่ละจุด และค่าความเข้มของการสะท้อนแสงแต่ละจุด (ในขั้นตอนนี้ ไฟล์มักจะรวมถึงการประมาณค่าความคลาดเคลื่อนและการวัดความไม่สมบูรณ์ (ส่วนใดของการสะท้อนแสงที่กำหนดถูกบันทึกไว้ในภาพนั้น))
ชุดข้อมูลทั้งหมดอาจประกอบด้วยภาพแยกกันหลายร้อยภาพที่ถ่ายจากทิศทางการวางตัวที่แตกต่างกันของผลึก จะต้องรวมและปรับขนาดภาพเหล่านี้โดยใช้จุดสูงสุดที่ปรากฏในภาพสองภาพขึ้นไป ( การรวม ) และการปรับขนาดเพื่อให้ได้มาตราส่วนความเข้มที่สม่ำเสมอ การเพิ่มประสิทธิภาพมาตราส่วนความเข้มมีความสำคัญอย่างยิ่ง เนื่องจากความเข้มสัมพัทธ์ของจุดสูงสุดเป็นข้อมูลสำคัญที่ใช้ในการกำหนดโครงสร้าง เทคนิคการเก็บข้อมูลทางผลึกศาสตร์แบบซ้ำๆ และความสมมาตรสูงของวัสดุผลึกทำให้เครื่องวัดการเลี้ยวเบนบันทึกการสะท้อนที่สมมาตรกันหลายครั้ง สิ่งนี้ช่วยให้สามารถคำนวณค่าR-factor ที่เกี่ยวข้องกับความสมมาตร ซึ่งเป็นดัชนีความน่าเชื่อถือโดยพิจารณาจากความคล้ายคลึงกันของความเข้มที่วัดได้ของการสะท้อนที่สมมาตรกัน[ 117 ]ดังนั้นจึงสามารถประเมินคุณภาพของข้อมูลได้
ระยะเริ่มต้น
ความเข้มของ "จุด" การเลี้ยวเบนแต่ละจุดเป็นสัดส่วนกับค่าสัมบูรณ์ยกกำลังสองของแฟกเตอร์โครงสร้างแฟกเตอร์โครงสร้างเป็นจำนวนเชิงซ้อนที่มีข้อมูลที่เกี่ยวข้องกับทั้งแอมพลิจูดและเฟสของคลื่นเพื่อให้ได้แผนที่ความหนาแน่นอิเล็กตรอน ที่ตีความได้ จำเป็นต้องทราบทั้งแอมพลิจูดและเฟส (แผนที่ความหนาแน่นอิเล็กตรอนช่วยให้นักผลึกศาสตร์สร้างแบบจำลองเริ่มต้นของโมเลกุลได้) ไม่สามารถบันทึกเฟสได้โดยตรงในระหว่างการทดลองการเลี้ยวเบน: นี่คือสิ่งที่เรียกว่าปัญหาเฟสการประมาณค่าเฟสเริ่มต้นสามารถทำได้หลายวิธี:
- การกำหนดเฟสแบบ Ab initioหรือวิธีโดยตรง – วิธีนี้มักเป็นวิธีที่เลือกใช้สำหรับโมเลกุลขนาดเล็ก (<1000 อะตอมที่ไม่ใช่ไฮโดรเจน) และได้ถูกนำมาใช้ประสบความสำเร็จในการแก้ปัญหาเฟสสำหรับโปรตีนขนาดเล็ก หากความละเอียดของข้อมูลดีกว่า 1.4 Å (140 pm)วิธีโดยตรงเพื่อรับข้อมูลเฟสได้ โดยใช้ประโยชน์จากความสัมพันธ์ของเฟสที่ทราบระหว่างกลุ่มการสะท้อนบางกลุ่ม [ 118 ] [ 119 ]
- การแทนที่โมเลกุล – หากทราบโครงสร้างที่เกี่ยวข้อง สามารถใช้เป็นแบบจำลองการค้นหาในการแทนที่โมเลกุลเพื่อกำหนดทิศทางและตำแหน่งของโมเลกุลภายในเซลล์หน่วย เฟสที่ได้ด้วยวิธีนี้สามารถใช้สร้างแผนที่ความหนาแน่นอิเล็กตรอนได้ [ 120 ]
- การกระเจิงของรังสีเอกซ์แบบผิดปกติ ( MADหรือSAD phasing ) – สามารถสแกนความยาวคลื่นของรังสีเอกซ์ผ่านขอบการดูดกลืน [ a ]ของอะตอม ซึ่งจะเปลี่ยนการกระเจิงในลักษณะที่ทราบได้ โดยการบันทึกชุดการสะท้อนที่สมบูรณ์ที่ความยาวคลื่นสามค่าที่แตกต่างกัน (ต่ำกว่ามาก สูงกว่ามาก และตรงกลางของขอบการดูดกลืน) จะสามารถหาโครงสร้างย่อยของอะตอมที่เกิดการเลี้ยวเบนแบบผิดปกติ และด้วยเหตุนี้จึงสามารถหาโครงสร้างของโมเลกุลทั้งหมดได้ วิธีที่นิยมที่สุดในการรวมอะตอมที่เกิดการกระเจิงแบบผิดปกติเข้ากับโปรตีนคือการแสดงออกของโปรตีนในเมไทโอนีนออโซโทรฟ (โฮสต์ที่ไม่สามารถสังเคราะห์เมไทโอนีนได้) ในอาหารเลี้ยงเชื้อที่อุดมไปด้วยซีลีโนเมไทโอนีน ซึ่งมี อะตอม ของซีลีเนียมจากนั้นสามารถทำการทดลองการกระจายตัวแบบผิดปกติหลายความยาวคลื่น (MAD) รอบขอบการดูดกลืน ซึ่งควรจะให้ตำแหน่งของหมู่เมไทโอนีนใดๆ ภายในโปรตีน ทำให้ได้เฟสเริ่มต้น [ 121 ]
- วิธีการอะตอมหนัก ( การแทนที่ไอโซมอร์ฟัสหลายตัว ) – หากสามารถนำอะตอมโลหะที่มีความหนาแน่นของอิเล็กตรอนเข้าไปในผลึกได้ สามารถใช้ วิธีการโดยตรงหรือวิธีการในปริภูมิแพตเตอร์สันเพื่อกำหนดตำแหน่งของอะตอมเหล่านั้นและเพื่อให้ได้เฟสเริ่มต้น สามารถนำอะตอมหนักดังกล่าวเข้าไปได้โดยการแช่ผลึกในสารละลายที่มีอะตอมหนัก หรือโดยการตกผลึกร่วม (การปลูกผลึกในขณะที่มีอะตอมหนักอยู่) เช่นเดียวกับการหาเฟสการกระจายตัวแบบผิดปกติหลายความยาวคลื่น การเปลี่ยนแปลงในแอมพลิจูดการกระเจิงสามารถตีความเพื่อให้ได้เฟส แม้ว่านี่จะเป็นวิธีการดั้งเดิมที่ใช้ในการแก้โครงสร้างผลึกโปรตีน แต่ส่วนใหญ่ถูกแทนที่ด้วยการหาเฟสการกระจายตัวแบบผิดปกติหลายความยาวคลื่นด้วยซีลีโนเมไทโอนีนแล้ว[ 120 ]
การสร้างแบบจำลองและการปรับปรุงขั้นตอน


เมื่อได้เฟสเริ่มต้นแล้ว สามารถสร้างแบบจำลองเบื้องต้นได้ ตำแหน่งอะตอมในแบบจำลองและค่า Debye-Waller factor (หรือB -factor ซึ่งคำนึงถึงการเคลื่อนที่ทางความร้อนของอะตอม) สามารถปรับแต่งให้เข้ากับข้อมูลการเลี้ยวเบนที่สังเกตได้ โดยในอุดมคติแล้วจะได้ชุดเฟสที่ดีขึ้น จากนั้นจึงสามารถปรับแบบจำลองใหม่ให้เข้ากับแผนที่ความหนาแน่นอิเล็กตรอนใหม่ และดำเนินการปรับแต่งในรอบต่อๆ ไป กระบวนการวนซ้ำนี้จะดำเนินต่อไปจนกว่าความสัมพันธ์ระหว่างข้อมูลการเลี้ยวเบนและแบบจำลองจะสูงสุด ความสอดคล้องจะวัดได้จากค่าR -factorซึ่งกำหนดโดย

โดยที่Fคือแฟกเตอร์โครงสร้างเกณฑ์คุณภาพที่คล้ายกันคือR freeซึ่งคำนวณจากชุดย่อย (~10%) ของการสะท้อนแสงที่ไม่ได้รวมอยู่ในการปรับปรุงโครงสร้าง ปัจจัย R ทั้งสอง ขึ้นอยู่กับความละเอียดของข้อมูล โดยทั่วไปแล้วR freeควรมีค่าประมาณความละเอียดในหน่วยอังสตรอมหารด้วย 10 ดังนั้นชุดข้อมูลที่มีความละเอียด 2 Å ควรให้ค่าR free สุดท้าย ประมาณ 0.2 คุณลักษณะของพันธะเคมี เช่น สเตอริโอเคมี พันธะไฮโดรเจน และการกระจายของความยาวพันธะและมุม เป็นมาตรวัดเสริมของคุณภาพแบบจำลอง ในการสร้างแบบจำลองแบบวนซ้ำ มักจะพบอคติเฟสหรืออคติแบบจำลอง เนื่องจากค่าประมาณเฟสมาจากแบบจำลอง แผนที่ที่คำนวณแต่ละรอบจึงมักแสดงความหนาแน่นในทุกที่ที่แบบจำลองมีความหนาแน่น โดยไม่คำนึงว่าจะมีความหนาแน่นจริงหรือไม่ ปัญหานี้สามารถบรรเทาได้ด้วยการถ่วงน้ำหนักความน่าจะเป็นสูงสุดและการตรวจสอบโดยใช้แผนที่ละเว้น[ 124 ]
อาจเป็นไปไม่ได้ที่จะสังเกตอะตอมทุกตัวในหน่วยอสมมาตร ในหลายกรณีความไม่เป็นระเบียบทางผลึกศาสตร์ทำให้แผนที่ความหนาแน่นของอิเล็กตรอนเบลอ อะตอมที่มีการกระเจิงแสงอ่อนๆ เช่น ไฮโดรเจน มักจะมองไม่เห็น นอกจากนี้ ยังเป็นไปได้ที่อะตอมเดียวจะปรากฏหลายครั้งในแผนที่ความหนาแน่นของอิเล็กตรอน เช่น หากหมู่ข้างเคียงของโปรตีนมีโครงสร้างที่อนุญาตได้หลายแบบ (<4) ในกรณีอื่นๆ นักผลึกศาสตร์อาจตรวจพบว่าโครงสร้างพันธะโควาเลนต์ที่อนุมานได้สำหรับโมเลกุลนั้นไม่ถูกต้องหรือเปลี่ยนแปลงไป ตัวอย่างเช่น โปรตีนอาจถูกตัดหรือได้รับการดัดแปลงหลังการแปลรหัสซึ่งไม่ได้รับการตรวจพบก่อนการตกผลึก
ความผิดปกติ
ความท้าทายทั่วไปในการปรับปรุงโครงสร้างผลึกเกิดจากความไม่เป็นระเบียบทางผลึกศาสตร์ ความไม่เป็นระเบียบสามารถเกิดขึ้นได้หลายรูปแบบ แต่โดยทั่วไปเกี่ยวข้องกับการอยู่ร่วมกันของสปีชีส์หรือคอนฟอร์เมชันสองชนิดขึ้นไป การไม่รู้จักความไม่เป็นระเบียบส่งผลให้เกิดการตีความที่ผิดพลาด ข้อผิดพลาดจากการสร้างแบบจำลองความไม่เป็นระเบียบที่ไม่เหมาะสมแสดงให้เห็นได้จากสมมติฐานที่ลดทอนของ ไอโซเมริซึม การยืดพันธะ[ 125 ] ความไม่เป็นระเบียบถูกสร้างแบบจำลองโดยสัมพันธ์กับจำนวนประชากรสัมพัทธ์ของส่วนประกอบ ซึ่งมักจะมีเพียงสองส่วน และเอกลักษณ์ของส่วนประกอบเหล่านั้น ในโครงสร้างของโมเลกุลขนาดใหญ่และไอออน ตัวทำละลายและไอออนตรงข้ามมักไม่เป็นระเบียบ
การวิเคราะห์ข้อมูลเชิงคำนวณประยุกต์
ปัจจุบันการใช้ระเบียบวิธีคำนวณในการวิเคราะห์ข้อมูลการเลี้ยวเบนรังสีเอกซ์ของผงวัสดุเป็นที่แพร่หลายมากขึ้น โดยทั่วไปจะเปรียบเทียบข้อมูลจากการทดลองกับไดอะแกรมการเลี้ยวเบนที่จำลองขึ้นจากโครงสร้างแบบจำลอง โดยคำนึงถึงพารามิเตอร์ของเครื่องมือ และปรับปรุงพารามิเตอร์โครงสร้างหรือโครงสร้างจุลภาคของแบบจำลองโดยใช้อัลกอริทึมการลดค่าต่ำสุดแบบกำลังสองน้อยที่สุด เครื่องมือส่วนใหญ่ที่มีอยู่ซึ่งอนุญาตให้ระบุเฟสและการปรับปรุงโครงสร้าง นั้นใช้พื้นฐานจากวิธี Rietveld [ 126 ] [ 127 ]โดยบางส่วนเป็นซอฟต์แวร์แบบเปิดและฟรี เช่น FullProf Suite [ 128 ] [ 129 ] Jana2006 [ 130 ] MAUD [ 131 ] [ 132 ] [ 133 ] Rietan [ 134 ] GSAS [ 135 ]เป็นต้น ในขณะที่บางส่วนมีให้ใช้งานภายใต้ใบอนุญาตเชิงพาณิชย์ เช่น Diffrac.Suite TOPAS [ 136 ] Match! [ 137 ]เป็นต้น เครื่องมือเหล่านี้ส่วนใหญ่ยังอนุญาตให้ ทำการปรับปรุง Le Bail (เรียกอีกอย่างว่าการจับคู่โปรไฟล์) นั่นคือ การปรับปรุงพารามิเตอร์ของเซลล์โดยอิงจากตำแหน่งยอด Bragg และโปรไฟล์ยอด โดยไม่คำนึงถึงโครงสร้างผลึกวิทยาด้วยตัวมันเอง เครื่องมือรุ่นใหม่กว่าช่วยให้สามารถปรับปรุงข้อมูลทั้งโครงสร้างและโครงสร้างจุลภาคได้ เช่น โปรแกรม FAULTS ที่รวมอยู่ใน FullProf Suite [ 138 ]ซึ่งช่วยให้สามารถปรับปรุงโครงสร้างที่มีข้อบกพร่องในระนาบ (เช่น ข้อบกพร่องในการเรียงซ้อน การเกิดแฝด การเจริญเติบโตแทรกซ้อน)
การตกตะกอนของโครงสร้าง
เมื่อแบบจำลองโครงสร้างของโมเลกุลเสร็จสมบูรณ์แล้ว มักจะมีการบันทึกไว้ในฐานข้อมูลทางผลึกศาสตร์เช่นฐานข้อมูลโครงสร้างเคมบริดจ์ (สำหรับโมเลกุลขนาดเล็ก) ฐานข้อมูลโครงสร้างผลึกอนินทรีย์ (ICSD) (สำหรับสารประกอบอนินทรีย์) หรือธนาคารข้อมูลโปรตีน (สำหรับโปรตีนและบางครั้งกรดนิวคลีอิก) โครงสร้างจำนวนมากที่ได้จากโครงการเชิงพาณิชย์ส่วนตัวในการตกผลึกโปรตีนที่มีความสำคัญทางการแพทย์นั้น ไม่ได้ถูกบันทึกไว้ในฐานข้อมูลทางผลึกศาสตร์สาธารณะ
บทบาทของสตรีในการศึกษาผลึกศาสตร์ด้วยรังสีเอกซ์
ผู้หญิงจำนวนหนึ่งเป็นผู้บุกเบิกในด้านผลึกศาสตร์รังสีเอกซ์ในช่วงเวลาที่พวกเธอถูกกีดกันจากสาขาวิทยาศาสตร์กายภาพอื่นๆ ส่วนใหญ่[ 139 ]
แคธลีน ลอนส์เดลเป็นนักศึกษาผู้ช่วยวิจัยของวิลเลียม เฮนรี แบรกก์ซึ่งมีนักศึกษาผู้ช่วยวิจัยหญิง 11 คน จากทั้งหมด 18 คน เธอเป็นที่รู้จักทั้งในด้านงานวิจัยเชิงทดลองและเชิงทฤษฎี ลอนส์เดลเข้าร่วมทีมวิจัยด้านผลึกศาสตร์ของแบรกก์ที่สถาบันวิทยาศาสตร์แห่งราชวงศ์ในลอนดอนในปี 1923 และหลังจากแต่งงานและมีบุตร เธอก็กลับไปทำงานกับแบรกก์ในฐานะนักวิจัย เธอได้ยืนยันโครงสร้างของวงแหวนเบนซีน ดำเนินการศึกษาเกี่ยวกับเพชร เป็นหนึ่งในสองผู้หญิงคนแรกที่ได้รับเลือกเข้าสู่ราชสมาคมในปี 1945 และในปี 1949 ได้รับการแต่งตั้งให้เป็นศาสตราจารย์หญิงคนแรกด้านเคมีและหัวหน้าภาควิชาผลึกศาสตร์ที่มหาวิทยาลัยคอลเลจลอนดอน[ 140 ]ลอนส์เดลสนับสนุนการมีส่วนร่วมของสตรีในวงการวิทยาศาสตร์มากขึ้นเสมอ และกล่าวไว้ในปี 1970 ว่า “ประเทศใดก็ตามที่ต้องการใช้ประโยชน์จากนักวิทยาศาสตร์และนักเทคโนโลยีที่มีศักยภาพทั้งหมดอย่างเต็มที่ สามารถทำได้ แต่ไม่ควรคาดหวังว่าจะได้ผู้หญิงมาง่ายๆ เหมือนกับที่ได้ผู้ชาย ... ดังนั้น การเสนอแนะว่าประเทศใดก็ตามที่ต้องการให้สตรีที่แต่งงานแล้วกลับมาประกอบอาชีพด้านวิทยาศาสตร์อีกครั้ง เมื่อลูกๆ ของเธอไม่ต้องการการปรากฏตัวของเธออีกต่อไป ควรจัดเตรียมมาตรการพิเศษเพื่อส่งเสริมให้เธอทำเช่นนั้น จึงเป็นการมองโลกในแง่ดีเกินไป” [ 141 ] ในช่วงเวลานี้ ลอนส์เดลได้เริ่มทำงานร่วมกับวิลเลียม ที. แอสเบอรี ในการจัดทำตารางกลุ่มอวกาศ 230 ตาราง ซึ่งได้รับการตีพิมพ์ในปี 1924 และกลายเป็นเครื่องมือสำคัญสำหรับนักผลึกศาสตร์

ในปี 1932 โดโรธี ฮอดจ์กินเข้าร่วมห้องปฏิบัติการของนักฟิสิกส์ จอห์น เดสมอนด์ เบอร์นัล ซึ่งเป็นอดีตนักศึกษาของแบร็ก ที่เคมบริดจ์ สหราชอาณาจักร เธอและเบอร์นัลได้ถ่ายภาพเอ็กซ์เรย์โปรตีนผลึกเป็นครั้งแรก ฮอดจ์กินยังมีบทบาทสำคัญในการก่อตั้งสหภาพคริสตัลโลกราฟีระหว่างประเทศเธอได้รับรางวัลโนเบลสาขาเคมีในปี 1964 จากผลงานการใช้เทคนิคเอ็กซ์เรย์เพื่อศึกษาโครงสร้างของเพนิซิลลิน อินซูลิน และวิตามินบี 12 งานวิจัยเกี่ยวกับเพนิซิลลินของเธอเริ่มต้นในปี 1942 ในช่วงสงคราม และเกี่ยวกับวิตามินบี 12 ในปี 1948 แม้ว่ากลุ่มวิจัยของเธอจะค่อยๆ เติบโตขึ้น แต่จุดสนใจหลักของพวกเขาคือการวิเคราะห์เอ็กซ์เรย์ของผลิตภัณฑ์จากธรรมชาติ เธอเป็นสตรีชาวอังกฤษเพียงคนเดียวที่เคยได้รับรางวัลโนเบลในสาขาวิทยาศาสตร์

โรซาลินด์ แฟรงคลินทำงานเกี่ยวกับการถ่ายภาพรังสีเอกซ์ของเส้นใยดีเอ็นเอ ซึ่งพิสูจน์แล้วว่าเป็นกุญแจสำคัญในการค้นพบโครงสร้างเกลียวคู่ของเจมส์ วัตสันและฟรานซิส คริก ซึ่งทำให้ทั้งคู่ได้รับรางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ในปี 1962 วัตสันเปิดเผยในบันทึกอัตชีวประวัติเกี่ยวกับการค้นพบโครงสร้างของดีเอ็นเอ เรื่อง The Double Helix [ 142 ] ว่าเขาได้ใช้ภาพถ่ายรังสีเอกซ์ของแฟรงคลินโดยไม่ได้รับอนุญาตจากเธอ แฟรงคลินเสียชีวิตด้วยโรคมะเร็งเมื่อ อายุได้ 30 กว่าปี ก่อนที่วัตสันจะได้รับรางวัลโนเบล แฟรงคลินยังได้ทำการศึกษาโครงสร้างที่สำคัญของคาร์บอนในถ่านหินและกราไฟต์ รวมถึงไวรัสของพืชและสัตว์ด้วย
อิซาเบลลา คาร์ลจากห้องปฏิบัติการวิจัยกองทัพเรือสหรัฐฯ ได้พัฒนาวิธีการทดลองเพื่อทำความเข้าใจทฤษฎีทางคณิตศาสตร์ของผลึกศาสตร์ งานของเธอช่วยเพิ่มความเร็วและความแม่นยำในการวิเคราะห์ทางเคมีและชีวการแพทย์ แต่มีเพียงสามีของเธอ เจโรม เท่านั้นที่ได้รับรางวัลโนเบลสาขาเคมีร่วมกับเฮอร์เบิร์ต ฮอปต์แมน ในปี 1985 "สำหรับความสำเร็จอันโดดเด่นในการพัฒนาวิธีการโดยตรงสำหรับการกำหนดโครงสร้างผลึก" อย่างไรก็ตาม หน่วยงานที่มอบรางวัลอื่นๆ ก็ได้มอบรางวัลต่างๆ ให้แก่อิซาเบลลาในฐานะบุคคลที่มีชื่อเสียงเช่นกัน
ผู้หญิงได้เขียนตำราและงานวิจัยจำนวนมากในสาขาผลึกศาสตร์รังสีเอกซ์ เป็นเวลาหลายปีที่ Lonsdale เป็นบรรณาธิการของInternational Tables for Crystallographyซึ่งให้ข้อมูลเกี่ยวกับโครงสร้างผลึก สมมาตร และกลุ่มพื้นที่ รวมถึงข้อมูลทางคณิตศาสตร์ ฟิสิกส์ และเคมีเกี่ยวกับโครงสร้างOlga Kennardจากมหาวิทยาลัยเคมบริดจ์ก่อตั้งและบริหารCambridge Crystallographic Data Centreซึ่งเป็นแหล่งข้อมูลโครงสร้างโมเลกุลขนาดเล็กที่เป็นที่ยอมรับในระดับสากล ตั้งแต่ปี 1965 จนถึงปี 1997 Jenny Pickworth Gluskerนักวิทยาศาสตร์ชาวอังกฤษ เป็นผู้ร่วมเขียน Crystal Structure Analysis: A Primer [ 143 ]ซึ่งตีพิมพ์ครั้งแรกในปี 1971 และ ณ ปี 2010 อยู่ในฉบับที่สามEleanor Dodsonนักชีววิทยาที่เกิดในออสเตรเลีย ซึ่งเริ่มต้นจากการเป็นช่างเทคนิคของ Dorothy Hodgkin เป็นผู้ริเริ่มหลักเบื้องหลังCCP4โครงการคอมพิวเตอร์ร่วมมือที่ปัจจุบันแบ่งปันเครื่องมือซอฟต์แวร์มากกว่า 250 รายการกับนักผลึกศาสตร์โปรตีนทั่วโลก
รางวัลโนเบลที่เกี่ยวข้องกับผลึกศาสตร์รังสีเอกซ์
| ปี | ผู้ได้รับรางวัล | รางวัล | เหตุผล |
|---|---|---|---|
| 1914 | แม็กซ์ ฟอน ลาเว | ฟิสิกส์ | "สำหรับการค้นพบการเลี้ยวเบนของรังสีเอ็กซ์โดยผลึก" [ 144 ] |
| 1915 | วิลเลียม เฮนรี แบร็ก | ฟิสิกส์ | "สำหรับบริการของพวกเขาในการวิเคราะห์โครงสร้างผลึกโดยใช้รังสีเอ็กซ์" [ 145 ] |
| วิลเลียม ลอว์เรนซ์ แบร็ก | |||
| พ.ศ. 2505 | แม็กซ์ เอฟ. เพรุตซ์ | เคมี | "สำหรับการศึกษาโครงสร้างของโปรตีนทรงกลม " [ 146 ] |
| จอห์น ซี. เคนดรูว์ | |||
| พ.ศ. 2505 | เจมส์ ดิวอี้ วัตสัน | ยา | "สำหรับการค้นพบของพวกเขาเกี่ยวกับโครงสร้างโมเลกุลของกรดนิวคลีอิกและความสำคัญของการถ่ายโอนข้อมูลในวัสดุที่มีชีวิต" [ 147 ] |
| ฟรานซิส แฮร์รี่ คอมป์ตัน คริก | |||
| มอริซ ฮิวจ์ เฟรเดอริค วิลกินส์ | |||
| พ.ศ. 2507 | โดโรธี ฮอดจ์กิน | เคมี | "สำหรับ การกำหนด โครงสร้างของสารชีวเคมีที่สำคัญโดยใช้เทคนิคเอ็กซ์เรย์ " [ 148 ] |
| พ.ศ. 2515 | สแตนฟอร์ด มัวร์ | เคมี | "เพื่อการมีส่วนร่วมของพวกเขาในการทำความเข้าใจความเชื่อมโยงระหว่างโครงสร้างทางเคมีและกิจกรรมเร่งปฏิกิริยาของศูนย์กลางการทำงานของ โมเลกุล ไรโบเอนไซม์ " [ 149 ] |
| วิลเลียม เอช. สไตน์ | |||
| พ.ศ. 2519 | วิลเลียม เอ็น. ลิปส์คอมบ์ | เคมี | "สำหรับการศึกษาโครงสร้างของโบเรนที่ให้ความกระจ่างเกี่ยวกับปัญหาของพันธะเคมี" [ 150 ] |
| พ.ศ. 2528 | เจอโรม คาร์ล | เคมี | "สำหรับความสำเร็จอันโดดเด่นของพวกเขาในการพัฒนาวิธีการโดยตรงสำหรับการกำหนดโครงสร้างผลึก" [ 151 ] |
| เฮอร์เบิร์ต เอ. ฮอปต์แมน | |||
| 1988 | โยฮันน์ ไดเซนโฮเฟอร์ | เคมี | "เพื่อการกำหนดโครงสร้างสามมิติของศูนย์ปฏิกิริยาการสังเคราะห์แสง " [ 152 ] |
| ฮาร์ทมุต มิเชล | เคมี | ||
| โรเบิร์ต ฮูเบอร์ | เคมี | ||
| พ.ศ. 2540 | จอห์น อี. วอล์คเกอร์ | เคมี | "เพื่อการอธิบายกลไกเอนไซม์ที่อยู่เบื้องหลังการสังเคราะห์อะดีโนซีนไตรฟอสเฟต (ATP)" [ 153 ] |
| 2003 | โรเดอริค แม็คคินนอน | เคมี | "สำหรับการค้นพบเกี่ยวกับช่องในเยื่อหุ้มเซลล์ [...] สำหรับการศึกษาโครงสร้างและกลไกของช่องไอออน " [ 154 ] |
| ปีเตอร์ อาเกร | "สำหรับการค้นพบเกี่ยวกับช่องทางในเยื่อหุ้มเซลล์ [...] สำหรับการค้นพบช่องทางน้ำ " [ 154 ] | ||
| 2006 | โรเจอร์ ดี. คอร์นเบิร์ก | เคมี | "สำหรับการศึกษาพื้นฐานโมเลกุลของการถอดรหัสยูคาริโอต " [ 155 ] |
| 2009 | เอดา อี. โยนาธ | เคมี | "สำหรับการศึกษาโครงสร้างและหน้าที่ของไรโบโซม " [ 156 ] |
| โทมัส เอ. สไตทซ์ | |||
| เวนคัตรามัน รามาคริชนัน | |||
| 2012 | ไบรอัน โคบิลก้า | เคมี | "สำหรับการศึกษาตัวรับที่เชื่อมโยงกับโปรตีน G " [ 157 ] |
ดูเพิ่มเติม
- แถบบีเวอร์ส-ลิปสัน
- การเลี้ยวเบนของแบร็ก
- ฐานข้อมูลผลึกศาสตร์
- กลุ่มจุดผลึกศาสตร์
- แผนที่แสดงความหนาแน่นที่แตกต่างกัน
- การเลี้ยวเบนของอิเล็กตรอน
- การเลี้ยวเบนรังสีเอกซ์แบบกระจายพลังงาน
- พารามิเตอร์ Flack
- การเลี้ยวเบนแบบตกกระทบเฉียง
- ขีดจำกัดเฮนเดอร์สัน
- ปีสากลแห่งผลึกศาสตร์
- รูปแบบความหนาแน่นหลายขั้ว
- การเลี้ยวเบนของนิวตรอน
- การเลี้ยวเบนของผง
- พีทิโคกราฟี
- สมการเชอร์เรอร์
- การศึกษาโครงสร้างผลึกด้วยเลเซอร์เฟมโตวินาทีแบบอนุกรม
- การกระเจิงรังสีเอกซ์มุมเล็ก (SAXS)
- การกำหนดโครงสร้าง
- รังสีเอกซ์ความเร็วสูงพิเศษ
- การตกผลึกของไวรัส
- การกระเจิงรังสีเอกซ์มุมกว้าง (WAXS)
- การเลี้ยวเบนของรังสีเอกซ์
หมายเหตุ
- ^ขอบการดูดกลืนนี้เดิมทีทราบมาจากสเปกโทรสโกปีการดูดกลืน รังสี เอ็กซ์ดู "การกระเจิงผิดปกติของรังสีเอ็กซ์" skuld.bmsc.washington.eduเก็บถาวรจากต้นฉบับเมื่อวันที่ 29 สิงหาคม 2549สำหรับคำแนะนำเกี่ยวกับการกระเจิงที่ผิดปกติ
อ่านเพิ่มเติม
ตารางสากลสำหรับผลึกศาสตร์
- Hahn T ,บรรณาธิการ (2002). ตารางสากลสำหรับผลึกศาสตร์ เล่ม A สมมาตรของกลุ่มอวกาศ (ฉบับที่ 5). ดอร์เดรชท์: สำนักพิมพ์ Kluwer Academic Publishersสำหรับสหภาพผลึกศาสตร์ระหว่างประเทศ ISBN 0-7923-6590-9.
- Rossmann MG, Arnold E, บรรณาธิการ (2001). ตารางสากลสำหรับผลึกศาสตร์ เล่ม F ผลึกศาสตร์ของโมเลกุลทางชีวภาพดอร์เดรชท์: สำนักพิมพ์ Kluwer Academic Publishers สำหรับสหภาพผลึกศาสตร์สากลISBN 0-7923-6857-6.
- Hahn T, บรรณาธิการ (1996). ตารางสากลสำหรับผลึกศาสตร์ ฉบับย่อสำหรับการสอน เล่ม A สมมาตรของกลุ่มอวกาศ (ฉบับที่ 4). ดอร์เดรชท์: สำนักพิมพ์ Kluwer Academic Publishers สำหรับสหภาพผลึกศาสตร์ระหว่างประเทศISBN 0-7923-4252-6.
หนังสือรวมบทความ
- Carter Jr CW, Sweet RM, บรรณาธิการ (1997). การตกผลึกของโมเลกุลขนาดใหญ่, ตอนที่ A (วิธีการทางเอนไซม์วิทยา, เล่ม 276) . ซานดิเอโก: สำนักพิมพ์ Academic Press. ISBN 0-12-182177-3.
- Carter Jr CW, Sweet RM, บรรณาธิการ (1997). การตกผลึกของโมเลกุลขนาดใหญ่, ตอนที่ B (วิธีการทางเอนไซม์วิทยา, เล่ม 277) . ซานดิเอโก: สำนักพิมพ์ Academic Press. ISBN 0-12-182178-1.
- Ducruix A, Giegé R, บรรณาธิการ (1999). การตกผลึกของกรดนิวคลีอิกและโปรตีน: แนวทางปฏิบัติ (ฉบับที่ 2). อ็อกซ์ฟอร์ด: สำนักพิมพ์มหาวิทยาลัยอ็อกซ์ฟอร์ด. ISBN 0-19-963678-8.
ตำราเรียน
- Birkholz M, Fewster PF, Genzel C (2005). "บทที่ 1: หลักการของการเลี้ยวเบนรังสีเอกซ์"การวิเคราะห์ฟิล์มบางโดยการกระเจิงรังสีเอกซ์ไวน์ไฮม์: Wiley-VCH. ISBN 978-3-527-31052-4– ผ่านทาง ResearchGate
- Blow D (2002). โครงร่างของผลึกศาสตร์สำหรับนักชีววิทยา . อ็อกซ์ฟอร์ด: สำนักพิมพ์มหาวิทยาลัยอ็อกซ์ฟอร์ด. ISBN 0-19-851051-9.
- Burns G, Glazer AM (1990). กลุ่มอวกาศสำหรับนักวิทยาศาสตร์และวิศวกร (ฉบับที่ 2). บอสตัน: Academic Press, Inc. ISBN 0-12-145761-3.
- Clegg W (1998). การหาโครงสร้างผลึก (Oxford Chemistry Primer) . Oxford: Oxford University Press. ISBN 0-19-855901-1.
- Cullity BD (1978). องค์ประกอบของการเลี้ยวเบนรังสีเอกซ์ (ฉบับที่ 2). เรดดิง รัฐแมสซาชูเซตส์: สำนักพิมพ์แอดดิสัน-เวสลีย์ISBN 0-534-55396-6.
- Drenth J (1999). หลักการของการตกผลึกโปรตีนด้วยรังสีเอกซ์ . นิวยอร์ก: Springer-Verlag. ISBN 0-387-98587-5.
- Giacovazzo C (1992). พื้นฐานของผลึกศาสตร์ . อ็อกซ์ฟอร์ด: สำนักพิมพ์มหาวิทยาลัยอ็อกซ์ฟอร์ด. ISBN 0-19-855578-4.
- Glusker JP, Lewis M, Rossi M (1994). การวิเคราะห์โครงสร้างผลึกสำหรับนักเคมีและนักชีววิทยา . นิวยอร์ก: VCH Publishers. ISBN 0-471-18543-4.
- Massa W (2004). การกำหนดโครงสร้างผลึก . เบอร์ลิน: Springer. ISBN 3-540-20644-2.
- McPherson A (1999). การตกผลึกของโมเลกุลชีวภาพขนาดใหญ่ . โคลด์สปริงฮาร์เบอร์, นิวยอร์ก: สำนักพิมพ์ห้องปฏิบัติการโคลด์สปริงฮาร์เบอร์. ISBN 0-87969-617-6.
- McPherson A (2003). บทนำสู่ผลึกศาสตร์ของโมเลกุลขนาดใหญ่ . John Wiley & Sons. ISBN 0-471-25122-4.
- McRee DE (1993). การศึกษาโครงสร้างผลึกโปรตีนเชิงปฏิบัติ . ซานดิเอโก: Academic Press. ISBN 0-12-486050-8.
- O'Keeffe M, Hyde BG (1996). โครงสร้างผลึก; เล่ม 1 รูปแบบและสมมาตรวอชิงตัน ดี.ซี.: สมาคมแร่ธาตุวิทยาแห่งอเมริกา, ชุดเอกสารทางวิชาการISBN 0-939950-40-5.
- Rhodes G (2000). Crystallography Made Crystal Clear (PDF) . ซานดิเอโก: Academic Press. ISBN 0-12-587072-8จัดเก็บ(PDF)จากต้นฉบับเมื่อ 2021-10-08 เรียกดูเมื่อ2007-09-16 – ผ่านทาง UW-Eau Claire, Chem 406, Fall 2005
- Rupp B (2009). ผลึกศาสตร์ชีวโมเลกุล: หลักการ การปฏิบัติ และการประยุกต์ใช้ในชีววิทยาโครงสร้างนิวยอร์ก: Garland Science. ISBN 978-0-8153-4081-2.
- Warren BE (1969). การเลี้ยวเบนของรังสีเอ็กซ์ . นิวยอร์ก: Dover Publications. ISBN 0-486-66317-5.
- Zachariasen WH (1945). ทฤษฎีการเลี้ยวเบนของรังสีเอกซ์ในผลึก . นิวยอร์ก: Dover Publications. LCCN 67026967 .
การวิเคราะห์ข้อมูลเชิงคำนวณประยุกต์
- Young RA, บรรณาธิการ (1993). วิธีการของรีทเวลด์ . อ็อกซ์ฟอร์ด: สำนักพิมพ์มหาวิทยาลัยอ็อกซ์ฟอร์ดและสหภาพคริสตัลโลกราฟีระหว่างประเทศ. ISBN 0-19-855577-6.
ประวัติศาสตร์
- Bijvoet MJ , Burgers WG, Hägg G, บรรณาธิการ (1969). เอกสารยุคแรกเกี่ยวกับการเลี้ยวเบนของรังสีเอกซ์โดยผลึกเล่มที่ 1. อูเทรคต์: จัดพิมพ์โดย A. Oosthoek's Uitgeversmaatschappij NV ให้แก่สหภาพคริสตัลโลกราฟีนานาชาติ
- Bijvoet JM, Burgers WG, Hägg G, eds (1972) บทความต้นเกี่ยวกับการเลี้ยวเบนของรังสีเอกซ์ด้วยคริสตัล ฉบับที่ ครั้งที่สอง Utrecht: เผยแพร่สำหรับ International Union of Crystallography โดย A. Oosthoek's Uitgeversmaatschappij NV
- Bragg WL, Phillips DC, Lipson H (1992). การพัฒนาการวิเคราะห์ด้วยรังสีเอกซ์ . นิวยอร์ก: Dover. ISBN 0-486-67316-2.
- Ewald PPและคณะ (บรรณาธิการ) (1962). ห้าสิบปีแห่งการเลี้ยวเบนรังสีเอก ซ์ . อูเทรคต์: จัดพิมพ์โดย A. Oosthoek's Uitgeversmaatschappij NV ให้แก่สหภาพคริสตัลโลกราฟีนานาชาติdoi : 10.1007/978-1-4615-9961-6 . ISBN 978-1-4615-9963-0.
{{cite book}}:ปัญหาความไม่เข้ากันของหมายเลข ISBN / วันที่ ( ขอความช่วยเหลือ ) - Ewald PP (บรรณาธิการ). "50 ปีแห่งการเลี้ยวเบนของรังสีเอ็กซ์"สหภาพคริสตัลโลกราฟีระหว่างประเทศ. เก็บถาวรจากต้นฉบับเมื่อ 2008-03-23 . สืบค้น เมื่อ 2006-12-11 .
พิมพ์ซ้ำในรูปแบบ pdf สำหรับการประชุม IUCr ครั้งที่ XVIII, กลาสโกว์, สก็อตแลนด์
- ฟรีดริช ดับเบิลยู (1922) "ดาย เกชิคเทอ เดอร์ อัฟฟินดุง เดอร์ เรินต์เกนสตราห์อินเทอร์เฟเรนเซน " ดายนาตูร์วิสเซนชาฟเทิน10 (16): 363. Bibcode : 1922NW.....10..363F . ดอย : 10.1007/BF01565289 . S2CID 28141506 . เก็บถาวรจากต้นฉบับเมื่อ 2020-03-29 . สืบค้นเมื่อ2019-12-03 .
- Lonsdale K (1949). ผลึกและรังสีเอ็กซ์ . นิวยอร์ก: D. van Nostrand.
ลิงก์ภายนอก
บทช่วยสอน
- การเรียนรู้ผลึกศาสตร์
- แนะนำแบบง่ายๆ ไม่ซับซ้อน ไม่เน้นศัพท์เทคนิค
- ชุดวิดีโอ "The Crystallography Collection"จากสถาบัน Royal Institution
- เอกสาร "การตกผลึกของโมเลกุลขนาดเล็ก" ( PDF ) บนเว็บไซต์ของสถาบันเทคโนโลยีอิลลินอยส์
- สหภาพคริสตัลโลกราฟีระหว่างประเทศ
- ผลึกศาสตร์เบื้องต้น
- บทเรียนเชิงโต้ตอบเกี่ยวกับแฟกเตอร์โครงสร้างซึ่งสาธิตคุณสมบัติของรูปแบบการเลี้ยวเบนของผลึก 2 มิติ
- หนังสือภาพเกี่ยวกับการแปลงฟูริเยร์แสดงความสัมพันธ์ระหว่างผลึกและรูปแบบการเลี้ยวเบนในแบบ 2 มิติ
- บันทึกการบรรยายเรื่องผลึกศาสตร์รังสีเอกซ์และการหาโครงสร้างจัดเก็บไว้เมื่อวันที่ 15 ตุลาคม 2017 ที่Wayback Machine
- การบรรยายออนไลน์เรื่อง วิธีการกระเจิงรังสีเอกซ์สมัยใหม่สำหรับการวิเคราะห์วัสดุระดับนาโนโดย ริชาร์ด เจ. มาตี
- ไทม์ไลน์เชิงโต้ตอบด้านผลึกศาสตร์ เก็บถาวร เมื่อวัน ที่ 30 มิถุนายน 2021 ที่Wayback MachineจากRoyal Institution
ฐานข้อมูลหลัก
- ฐานข้อมูลผลึกศาสตร์แบบเปิด (COD)
- ฐานข้อมูลโปรตีน ( PDB )
- ฐานข้อมูลกรดนิวคลีอิก (Nucleic Acid Databank) ถูกเก็บถาวรเมื่อวันที่ 14 กรกฎาคม 2018 ที่Wayback Machine (NDB)
- ฐานข้อมูลโครงสร้างเคมบริดจ์ ( CSD )
- ฐานข้อมูลโครงสร้างผลึกอนินทรีย์ (Inorganic Crystal Structure Database ) ถูกเก็บถาวรเมื่อวันที่ 3 มีนาคม 2012 ที่Wayback Machine ( ICSD )
- ฐานข้อมูลการตกผลึกของโมเลกุลชีวภาพขนาดใหญ่ (BMCD)
ฐานข้อมูลอนุพันธ์
- ผลรวม PDB
- โปรทีโอพีเดีย – สารานุกรมสามมิติแบบร่วมมือของโปรตีนและโมเลกุลอื่นๆ
- RNABase
- ฐานข้อมูล HIC-Up ของลิแกนด์ PDB ถูกเก็บถาวรเมื่อวันที่ 8 สิงหาคม 2020 ที่Wayback Machine
- ฐานข้อมูลการจำแนกโครงสร้างโปรตีน
- การจำแนกโครงสร้างโปรตีน CATH
- รายชื่อโปรตีนที่อยู่ภายในเยื่อหุ้มเซลล์ซึ่งมีโครงสร้างสามมิติที่ทราบแล้วเก็บถาวรเมื่อวันที่ 11 เมษายน 2554 ที่Wayback Machine
- ฐานข้อมูลการวางตัวของโปรตีนในเยื่อหุ้มเซลล์
การตรวจสอบโครงสร้าง
- ชุดตรวจสอบโครงสร้าง MolProbity
- โปรเอสเอ-เว็บ
- NQ-Flipper (ตรวจสอบโรตาเมอร์ที่ไม่พึงประสงค์ของกรดอะมิโนแอสปาราจีนและกลูตามีน)
- เซิร์ฟเวอร์ DALI (ระบุโปรตีนที่คล้ายกับโปรตีนที่กำหนด)
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ผลึกศาสตร์รังสีเอกซ์
การศึกษาโครงสร้างผลึกด้วยรังสีเอกซ์ เป็นวิทยาศาสตร์เชิงทดลองที่ใช้ในการหาโครงสร้างอะตอมและโมเลกุลของ ผลึก โดยโครงสร้างดังกล่าวจะทำให้ลำแสง รังสีเอกซ์ ที่ตกกระทบ เกิด...
ประวัติศาสตร์
แม้ว่าผลึกจะได้รับการชื่นชมมานานแล้วในเรื่องความสม่ำเสมอและ ความสมมาตร แต่ก็ยังไม่มีการศึกษาทางวิทยาศาสตร์จนกระทั่งศตวรรษที่ 17 โยฮันเนส เคปเลอร์ ตั้งสมมติฐานในงานของเขา เรื่อง Strena seu de Nive Sexangula (ของขวัญปีใหม่จากหิมะรูปหกเหลี่ยม) (1611)...
ผลกระทบต่อวิชาเคมี
การตกผลึกด้วยรังสีเอกซ์นำไปสู่ความเข้าใจที่ดีขึ้นเกี่ยวกับ พันธะเคมี และ ปฏิสัมพันธ์ที่ไม่ใช่โคเวเลนต์ การศึกษาเบื้องต้นเผยให้เห็นรัศมีทั่วไปของอะตอม และยืนยันแบบจำลองทางทฤษฎีของพันธะเคมีหลายแบบ เช่น พันธะเตตระเฮดรัลของคาร์บอนในโครงสร้างเพชร [ 30 ]...
ผลกระทบต่อแร่ธาตุวิทยา
การประยุกต์ใช้ผลึกศาสตร์รังสีเอกซ์กับ แร่ธาตุวิทยา เริ่มต้นด้วยโครงสร้างของ แร่การ์เนต ซึ่งถูกกำหนดโดยเมนเซอร์ในปี 1924 การศึกษาผลึกศาสตร์รังสีเอกซ์อย่างเป็นระบบของ ซิลิเกต ได้ดำเนินการในช่วงทศวรรษที่ 1920 การศึกษานี้แสดงให้เห็นว่า เมื่อ อัตราส่วน Si / O...