อ่าน 24 นาที
กอร์โกนอปเซีย
กอร์โกนอปเซีย (มาจากภาษากรีก กอร์ กอน สัตว์ในตำนาน และ óps 'ลักษณะ') เป็น กลุ่ม สัตว์เลี้ยง ลูกด้วยนม ในกลุ่ม เทอแรปซิด ที่มีเขี้ยวแหลมคม ที่สูญพันธุ์ไปแล้ว จาก ยุคเพอร์เมียน...
กอร์โกนอปเซีย
| กอร์โกนอปเซียน ช่วงเวลา: ยุคเพอร์เมียนตอนกลาง - ยุคเพอร์เมียนตอนปลาย ( อาจเป็นบันทึกที่สิ้นสุดในยุคเพอร์เมียนตอนต้น) | |
|---|---|
 | |
| โครงกระดูกของInostrancevia alexandriที่Museo delle Scienzeเมืองเทรนโต ประเทศอิตาลี | |
| การจำแนกทางวิทยาศาสตร์ | |
| อาณาจักร: | แอนิมอลเลีย |
| ไฟลัม: | คอร์ดาต้า |
| กลุ่มสายพันธุ์ : | ซินาปซิดา |
| กลุ่มสายพันธุ์ : | เทราปซิดา |
| กลุ่มสายพันธุ์ : | เทอริโอดอนเทีย |
| กลุ่มสายพันธุ์ : | † กอร์โกนอปเซียซีลีย์ , 1894 |
| กลุ่มย่อย | |
กอร์โกนอปเซีย (มาจากภาษากรีกกอร์กอน สัตว์ในตำนาน และóps 'ลักษณะ') เป็นกลุ่ม สัตว์เลี้ยง ลูกด้วยนมในกลุ่มเทอแรปซิด ที่มีเขี้ยวแหลมคม ที่สูญพันธุ์ไปแล้ว จาก ยุคเพอร์เมียน ตอนกลางถึงตอนบนประมาณ 270 ถึง 252 ล้านปีก่อน ลักษณะเด่นคือ กะโหลกยาวและแคบ รวมถึงฟันเขี้ยว บนและบางครั้งล่างที่ยาว และฟันหน้าซึ่งน่าจะใช้เป็นอาวุธในการฟันและแทง ฟันหลังเขี้ยวโดยทั่วไปจะมีจำนวนน้อยลงหรือไม่มีเลย สำหรับการล่าเหยื่อขนาดใหญ่ พวกมันอาจใช้กลยุทธ์กัดแล้วถอย โดยซุ่มโจมตีและกัดเหยื่อจนอ่อนแรง จากนั้นติดตามเหยื่อในระยะที่ปลอดภัยก่อนที่บาดแผลจะทำให้เหยื่ออ่อนแรงลง เมื่อนั้นกอร์โกนอปเซียจะเข้าตะครุบเหยื่อและกัดเพื่อฆ่า พวกมันจะมีช่องปากที่กว้างมาก อาจเกิน 90 องศา โดยไม่ต้องขยับขากรรไกร
พวกมันมีขนาดใหญ่ขึ้นอย่างเห็นได้ชัดเมื่อเวลาผ่านไป โดยเติบโตจากกะโหลกขนาดเล็กที่มีความยาว 10–15 เซนติเมตร (4–6 นิ้ว) ในยุคเพอร์เมียนตอนกลาง ไปจนถึง ขนาดใหญ่โตคล้าย หมีที่มีความยาวถึง 60 เซนติเมตร (2 ฟุต) ในยุคเพอร์เมียนตอนปลาย กอร์โกนอปเซียนกลุ่มสุดท้ายRubidgeinaeเป็นกลุ่มที่แข็งแรงที่สุดในกลุ่มและสามารถกัดได้อย่างทรงพลังเป็นพิเศษ เชื่อกันว่ากอร์โกนอปเซียนอาศัยอยู่บนบก อย่างสมบูรณ์ และสามารถเดินด้วยท่าเดินกึ่งยืนตรง โดยมีระยะการเคลื่อนที่บนบกคล้ายกับจระเข้ ในปัจจุบัน พวกมันอาจมีความว่องไวกว่าเหยื่อของพวกมัน แต่คาดว่าน่าจะเป็นสัตว์เลือดอุ่น ที่อาศัยแรงเฉื่อย มากกว่า สัตว์เลือดอุ่นแบบ เอนโดเท อร์มิก ซึ่งแตกต่างจากเทโรเซฟาเลียนและไซโนดอนต์ ในยุคเดียวกัน ดังนั้นจึงอาจมีการเคลื่อนไหวค่อนข้างน้อยกว่า แม้ว่ากอร์โกนอปเซียนจะสามารถรักษาอุณหภูมิร่างกายให้สูงได้ แต่ก็ยังไม่แน่ชัดว่าพวกมันจะมีต่อมเหงื่อหรือขน (และโดยนัยคือหนวดและโครงสร้างที่เกี่ยวข้อง) หรือไม่ สมองของพวกมันมีลักษณะคล้ายคลึงกับ สมอง ของสัตว์เลื้อยคลาน ในปัจจุบัน มากกว่าสมองของสัตว์เลี้ยงลูกด้วยนมที่ ยังมีชีวิตอยู่ ส่วนใหญ่แล้วพวกมันอาจหากินในเวลากลางวันเป็นหลัก ( diurnally ) แม้ว่าบางชนิดอาจหากินในช่วงพลบค่ำหรือรุ่งเช้า ( creepuscular ) หรือหากินในเวลา กลางคืน (nocturnal) เชื่อกันว่าพวกมันมีสายตาแบบสองตา มีตาข้างขมับ ( parietal eye ) (ซึ่งตรวจจับแสงแดดและรักษาระบบจังหวะชีวิตประจำวัน ) มีประสาทสัมผัสการดมกลิ่นที่เฉียบคม มีอวัยวะรับกลิ่นใน โพรงจมูก ( vomeronasal organ ) ที่ทำงานได้ ("อวัยวะของเจคอบสัน") และอาจมีเยื่อแก้วหู ที่ยังพัฒนา ไม่ เต็มที่
กลุ่มเทอราปซิดหลักๆ ทั้งหมดได้วิวัฒนาการมาจากบรรพบุรุษ " เพลิโคซอร์ " (กลุ่มที่ไม่ชัดเจนนัก ซึ่งรวมถึงไซแนปซิด ทั้งหมด ที่ไม่ใช่เทอราปซิด) เมื่อ 275 ล้านปีก่อน การเข้ามาแทนที่เพลิโคซอร์ของเทอราปซิดเกิดขึ้นในช่วงยุคเพอร์เมียนตอนกลาง เนื่องจากโลกแห้งแล้งมากขึ้นเรื่อยๆ กอร์โกนอปเซียนกลายเป็นผู้ล่าสูงสุดในสภาพแวดล้อมของพวกมันหลังจากเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในยุคคาปิทาเนียนซึ่งทำให้ไดโนเซฟาเลียนและเทอโรเซฟาเลียนขนาดใหญ่บางชนิดสูญพันธุ์ไปหลังยุคเพอร์เมียนตอนกลาง แม้ว่าจะมีทวีปเดียวในยุคเพอร์เมีย น คือ พันเจีย แต่ กอร์โกนอปเซียนก็พบได้เพียงไม่กี่แห่งทั่วโลก พวกมันพบได้เป็นหลักในกลุ่ม หิน คารูซูเปอร์กรุ๊ปทางตอนใต้ของแอฟริกา (ส่วนใหญ่คือแอฟริกาใต้ แต่ก็พบในแทนซาเนีย แซมเบีย และมาลาวีด้วย) รวมถึงในไนเจอร์ รัสเซีย และจีน โดยอาจมีรายงานการพบในอินเดียด้วย สถานที่เหล่านี้เป็นพื้นที่กึ่งแห้งแล้งที่มีปริมาณน้ำฝนตามฤดูกาลสูง กอร์โกนอป เซียนที่เก่าแก่ที่สุดเท่าที่รู้จัก มีอายุย้อนไปถึงยุคเพอร์เมียนตอนกลาง มาจากเกาะมายอร์กา ของสเปน ทางตะวันตกของทะเลเมดิเตอร์เรเนียน สกุลของกอร์โกนอป เซียน ทั้งหมดมีลักษณะคล้ายคลึงกันมาก ดังนั้นหลายชนิดจึงได้รับการตั้งชื่อโดยอาศัยความแตกต่างที่ไม่ชัดเจนและอาจเกี่ยวข้องกับอายุ นับตั้งแต่การค้นพบในปลายศตวรรษที่ 19 และกลุ่มนี้ก็ได้รับการแก้ไขทางอนุกรมวิธานหลายครั้ง
กอร์โกนอปเซียนกลุ่มสุดท้ายสูญพันธุ์ไปในช่วงเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในยุคเพอร์เมียน-ไทรแอสสิกซึ่งเกิดขึ้นในช่วงปลายยุคเพอร์เมียน โดยกิจกรรมภูเขาไฟครั้งใหญ่ (ซึ่งจะก่อให้เกิดที่ราบสูงไซบีเรีย ) และการเพิ่มขึ้นอย่างมหาศาลของก๊าซเรือนกระจกส่งผลให้เกิดภาวะแห้งแล้งอย่างรวดเร็วเนื่องจากอุณหภูมิที่สูงขึ้นฝนกรดไฟป่าที่เกิดขึ้นบ่อยครั้งและการทำลายชั้นโอโซน ที่อาจเกิดขึ้นได้ แม้ว่าก่อนหน้านี้จะมีข้อเสนอแนะว่ากอร์โกนอปเซียนกลุ่มเล็ก ๆ บางชนิด เช่นไซโอโนซอรัสอาจมีชีวิตรอดมาจนถึงต้นยุคไทรแอสสิกแต่การวิเคราะห์ในภายหลังพบว่าข้อเสนอนี้ผิดพลาดแหล่งที่ อยู่อาศัยของสัตว์นักล่าขนาดใหญ่ ที่เคยเป็นของกอร์โกนอปเซียนนั้น จะถูกครอบครองโดยอาร์โคซอร์ (โดยเฉพาะจระเข้และไดโนเสาร์ ) ในยุคมีโซโซอิก
คำอธิบาย

กอร์โกนอปซิดในยุคเพอร์เมียนตอนต้นมีขนาดค่อนข้างเล็ก โดยมีความยาวกะโหลก 10–15 ซม. (4–6 นิ้ว) [ 1 ]ในขณะที่บางสกุลในยุคหลังมีขนาดใหญ่โตคล้ายหมี โดยสกุลที่ใหญ่ที่สุดคือInostranceviaมีความยาวถึง 3.5 ม. (11 ฟุต) และมีน้ำหนักตัว 300 กก. (660 ปอนด์) [ 2 ]อย่างไรก็ตาม กอร์โกนอปเซียนขนาดเล็กยังคงมีอยู่มากมายจนกระทั่งสูญพันธุ์ (แม้ว่าสายพันธุ์ขนาดเล็กอาจเป็นตัวอย่างวัยอ่อนของแท็กซาอื่นก็ตาม) [ 1 ]
เช่นเดียวกับเทอแรปซิดในยุคเพอร์เมียนอื่นๆ กอร์โกนอปเซียนได้พัฒนาลักษณะหลายอย่างของสัตว์เลี้ยงลูกด้วยนม ซึ่งอาจรวมถึงการเดินแบบพาราซาจิตัล (แขนขาอยู่ในแนวตั้งและเคลื่อนที่ขนานกับกระดูกสันหลัง) ตรงข้ามกับการเดินแบบกางขาของสัตว์ ครึ่ง บกครึ่งน้ำและไซแนปซิดในยุคก่อนหน้า การเปลี่ยนแปลงการเดินในเทอแรปซิดนี้อาจเกี่ยวข้องกับการลดขนาดหางและสูตรกระดูกนิ้ว[ 3 ] (จำนวนกระดูกต่อนิ้ว ซึ่งสำหรับกอร์โกนอปเซียนคือ 2, 3, 4, 5, 3 เหมือนสัตว์เลื้อยคลาน[ 4 ] ) การพัฒนาอื่นๆ ได้แก่ กระดูกคอร์ติคัลแบบแผ่นเส้นใยและฟันที่ฝังลึก[ 3 ]เช่นเดียวกับสัตว์เลื้อยคลาน กอร์โกนอปเซียนไม่มีเพดานปากรองที่แยกปากออกจากโพรงจมูก ทำให้ไม่สามารถเคี้ยวได้
กะโหลก
กายวิภาคของกอร์โกนอปเซียนแตกต่างกันน้อยมาก[ 5 ]หลายชนิดมีความแตกต่างกันเพียงเล็กน้อยในสัดส่วน และด้วยเหตุนี้ชนิดที่เล็กกว่าอาจเป็นลูกอ่อนของชนิดที่ใหญ่กว่าได้ ที่น่าสังเกตคือ กระดูกโว เมอร์ ที่ปลายจมูกจะแตกต่างกันไปในแต่ละชนิดในแง่ของระดับการขยายตัว รวมถึงตำแหน่ง ระดับการกางออก และรูปร่างของสันทั้ง 3 [ 1 ]โดยทั่วไปแล้วพวกมันจะมีกะโหลกที่ยาวและแคบ[ 4 ]ลูกอ่อน ของรูบิด เจียดูเหมือนจะมีจมูกที่กว้างกว่ายาว[ 6 ]ต่างจากยูเทอริโอดอนต์กระดูกท้ายทอย (ที่ด้านหลังของกะโหลก) เป็นรูปสี่เหลี่ยมผืนผ้า (กว้างกว่าสูง) และเว้า ตรงข้ามกับรูปสามเหลี่ยม[ 7 ] : 279

สมองของกอร์โกนอปเซียน เช่นเดียวกับเทอแรปซิดที่ไม่ใช่ สัตว์ เลี้ยงลูกด้วยนมชนิด อื่นๆ ขาดการขยายตัวของ นี โอคอร์เทกซ์มีสมองส่วนท้ายที่ค่อนข้างใหญ่เมื่อเทียบกับสมองส่วนหน้า มี เส้น ประสาทเอพิฟิเซียล ขนาดใหญ่ (พบในสิ่งมีชีวิตที่มีตาข้างขมับอยู่ด้านบนของศีรษะ) ต่อมใต้สมองที่ขยายใหญ่ขึ้นและรูปร่างโดยรวมที่ยาวเรียว โดยรวมแล้วคล้ายกับสมอง ของ สัตว์เลื้อยคลาน[ 8 ]กะโหลกศีรษะก็ค่อนข้างคล้ายสัตว์เลื้อยคลาน และมีขนาดเล็กกว่าและไม่หนาเท่ากับของสัตว์เลี้ยงลูกด้วยนม[ 7 ] : 280 ฟลอคคูลัสซึ่งเป็นกลีบของซีรีเบลลัมมีขนาดใหญ่ตามสัดส่วน และเกี่ยวข้องกับรีเฟล็กซ์เวสติบูโล-โอคูลาร์ (ซึ่งช่วยให้การมองคงที่ในขณะที่เคลื่อนศีรษะ) เมื่อพิจารณาจากทิศทางของท่อครึ่งวงกลมในหู (ซึ่งต้องวางตัวขนานกับพื้น) หัวของตัวอย่างกอร์โกนอปเซียน GPIT/RE/7124 จะเอียงไปข้างหน้าประมาณ 41° ทำให้การมองเห็นของดวงตาทั้งสองข้างซ้อนทับกันมากขึ้น และช่วยให้การมองเห็นแบบ สองตาดีขึ้น ซึ่งเป็นประโยชน์ต่อผู้ล่า ต่างจากสัตว์เลื้อยคลานหรือสัตว์เลี้ยงลูกด้วยนมท่อครึ่งวงกลมมีลักษณะแบนราบ อาจเป็นเพราะถูกคั่นอยู่ระหว่างกระดูกโอพิสโทติก ( กระดูก หูชั้นใน ) และกระดูกซูพราออกซิปิตัล[ 8 ]
ฟัน

เช่นเดียวกับสัตว์เลี้ยงลูกด้วยนมหลายชนิด กอร์โกนอปเซียนเป็นเฮเทอโรดอนต์ โดยมี ฟันหน้าฟันเขี้ยวและฟันหลังเขี้ยวที่ชัดเจนซึ่งมีลักษณะคล้ายคลึงกับฟันกรามและฟันกรามเล็ก[ 1 ]พวกมันมีฟันหน้าห้าซี่ในขากรรไกรบน (ส่วนใหญ่ ฟันสามซี่แรกมีขนาดเท่ากัน และสองซี่สุดท้ายสั้นกว่า) และสี่ซี่ในขากรรไกรล่าง[ 7 ] : 17–18
ในกอร์โกนอปเซียนส่วนใหญ่ ฟันหน้ามีขนาดใหญ่ และฟันเขี้ยวบนยาวเรียวคล้ายดาบ คล้ายกับแมวเขี้ยวดาบ ในยุคหลัง กอร์โกนอปเซียนบางชนิดมีฟันเขี้ยวบนยาวเป็นพิเศษ เช่นอินอสทรานเซเวียและบางชนิดมีแผ่นยื่นออกมาที่ขากรรไกรล่างเพื่อหุ้มปลายฟันเขี้ยวขณะที่ปากปิด ดาบโดยทั่วไปตีความได้ว่าใช้เป็นอาวุธสำหรับแทงหรือฟัน ซึ่งต้องใช้การอ้าปากที่กว้างมาก ฟันเขี้ยวทั้งบนและล่างของรูบิดเจียยาวเรียว และสัตว์ชนิดนี้จำเป็นต้องมีการอ้าปากที่กว้างกว่ามาก[ 9 ]รูปแบบรอยหยักของกอร์โกนอปเซียนคล้ายกับไดโนเสาร์เทอโรพอดมากกว่าซินาปซิดอื่นๆ[ 10 ]เพดานปากยังมีปุ่มและสันซึ่งมักจะมีฟันที่ใช้งานได้ ซึ่งอาจใช้ในการจับเหยื่อที่ดิ้นรน โดยเบี่ยงเบนแรงอันทรงพลังเหล่านี้ออกจากฟันเขี้ยวที่บอบบาง สันที่คล้ายกันนี้พบในHomotheriumซึ่งเป็น Machairodont [ 7 ] : 280 ฟันหลังเขี้ยวลดลงทั้งขนาดและจำนวนRubidgeines จำนวนมาก (Gorgonopsians รุ่นหลังสุด) ไม่มีฟันหลังเขี้ยวในขากรรไกรล่าง[ 11 ]และClelandinaไม่มีฟันหลังเขี้ยวเลย[ 6 ]
กอร์โกนอปเซียนเป็นสัตว์ที่มีฟันงอกหลายชุดและฟันจะงอกอย่างต่อเนื่องตลอดชีวิตของแต่ละตัว[ 12 ]เช่นเดียวกับเทอแรปซิดบางชนิด ในขณะที่มีเขี้ยวที่ใช้งานได้หนึ่งซี่ เขี้ยวอีกซี่หนึ่งก็จะงอกขึ้นมาแทนที่เมื่อมันหักไปโดยหลีกเลี่ยงไม่ได้ ด้านซ้ายและด้านขวาของขากรรไกรไม่จำเป็นต้องพร้อมกัน ดังนั้น ตัวอย่างเช่น เขี้ยวซี่แรกทางด้านซ้ายอาจใช้งานได้ในขณะที่เขี้ยวซี่แรกทางด้านขวายังคงงอกอยู่[ 12 ]วิธีการดังกล่าวอาจถูกนำมาใช้เพื่อให้มีเขี้ยวที่ใช้งานได้ครบชุดเสมอ เนื่องจากหากมีเขี้ยวเพียงซี่เดียวหรือไม่มีเขี้ยวเลยจะทำให้การล่าเหยื่อเป็นไปได้ยาก และการงอกฟันขนาดใหญ่เช่นนี้ต้องใช้เวลานาน ในทางกลับกัน เนื่องจากเขี้ยวที่ใช้งานได้มักจะพบในเบ้าฟันด้านหน้าสุด (แทนที่จะพบเท่าๆ กันในเบ้าฟันทั้งสองข้าง) จึงเป็นไปได้ว่าการเปลี่ยนเขี้ยวเกิดขึ้นเป็นจำนวนครั้งที่จำกัด และในที่สุดสัตว์ก็จะเหลือเขี้ยวที่ใช้งานได้ชุดเดียวถาวรในเบ้าฟันเหล่านี้ ในปี พ.ศ. 2527 นักบรรพชีวินวิทยาชาวอังกฤษDorisและKenneth Kermackเสนอว่าฟันเขี้ยวจะเติบโตจนมีขนาดเท่ากับกะโหลก และจะหักออกไปเรื่อยๆ จนกว่าสัตว์จะหยุดการเจริญเติบโต และกอร์โกนอปเซียนมีรูปแบบการเปลี่ยนฟันแบบจำกัดในระยะแรก ซึ่งพบได้ในสัตว์เลี้ยงลูกด้วยนมหลายชนิด รูปแบบการเปลี่ยนฟันของฟันซี่อื่นๆ ยังไม่ชัดเจน[ 9 ]ฟันหลังเขี้ยวจะถูกเปลี่ยนช้ากว่าฟันซี่อื่นๆ ซึ่งอาจเป็นเพราะไม่มีความสำคัญในเชิงหน้าที่[ 1 ]
กระดูกส่วนหลังกะโหลก

กระดูกสันหลังส่วนคอทั้งเจ็ดชิ้น(บริเวณคอ) มีขนาดเท่ากันหมด ยกเว้นชิ้นสุดท้ายซึ่งสั้นกว่าและอยู่ต่ำกว่า มีแอตลาส หนึ่งชิ้น และแอ็กซิส หนึ่ง ชิ้น เช่นเดียวกับแมวเขี้ยวเสือ คอยาวและมีกล้ามเนื้อที่พัฒนาอย่างดี ซึ่งจะเป็นประโยชน์อย่างยิ่งเมื่อเขี้ยวฝังอยู่ในตัวสัตว์ เช่นเดียวกับไซแนปซิดยุคแรกอื่นๆ กอร์โกนอปเซียนมีปุ่มกระดูกท้ายทอย เพียงปุ่มเดียว และข้อต่อของกระดูกสันหลังส่วนคอโดยรวมแล้วคล้ายกับสัตว์เลื้อยคลาน ทำให้ศีรษะสามารถเคลื่อนไหวไปด้านข้างได้ แต่จำกัดการเคลื่อนไหวขึ้นลง กระดูกสันหลังส่วนคอชิ้นสุดท้ายมีรูปร่างคล้ายกับกระดูกสันหลังส่วนอกมากกว่า[ 7 ] : 291–293
กระดูกสันหลังส่วนหลังมีรูปร่างคล้ายกระสวยและดูเหมือนกันหมด ส่วนยื่นของกระดูกสันหลังยื่นออกมาอย่างชันจากส่วนกลาง และมีสันแหลมคมอยู่ทั้งด้านหน้าและด้านหลัง ต่างจากยูเทอริโอดอนต์ กอร์โกนอปเซียนไม่มีกระดูกสันหลังส่วนเอวที่เห็นได้ชัดเจนอย่างไรก็ตามกระดูกสันหลังส่วนหลังที่เทียบเท่ากับชุดนั้นคล้ายกับกระดูกสันหลังส่วนเอวของแมวเขี้ยวเสือที่มีข้อต่อกระดูกโหนกแก้มที่ชัน ซึ่งมีประโยชน์ในการทำให้หลังส่วนล่างมั่นคง โดยเฉพาะอย่างยิ่งเมื่อจับเหยื่อที่ดิ้นรน[ 7 ] : 293–295
มีกระดูกสันหลังส่วนกระเบนเหน็บ 3 ชิ้น และกระดูกสันหลังชุดแรก ยึดติดกับ กระดูก เชิงกราน กระดูกเชิงกรานมีลักษณะคล้ายสัตว์เลื้อยคลาน โดยมี กระดูก สะโพก กระดูกก้นกบและกระดูกหัวหน่าว แยกออกจากกัน กระดูกต้นขาเป็นรูปตัว S เล็กน้อย สั้นแต่ยาวและเรียวกว่ากระดูกต้นแขนสำหรับคนส่วนใหญ่กระดูกหน้าแข้งและกระดูกน่องจะโค้งเข้าหากันอย่างมาก และกระดูกหน้าแข้งจะแข็งแรงกว่ากระดูกน่อง[ 7 ] : 295–299 ข้อต่อระหว่างข้อเท้าและกระดูกส้นเท้าอาจเคลื่อนไหวได้บ้าง นิ้วที่ห้าของทั้งมือและเท้าไม่ได้ยึดติดกับกระดูกข้อมือ / กระดูกข้อเท้าแต่เชื่อมต่อโดยตรงกับกระดูกปลายแขน /กระดูกส้นเท้า[ 13 ]
อนุกรมวิธาน
แหล่งฟอสซิล

ในปี พ.ศ. 2419 ซากดึกดำบรรพ์ของกอร์โกนอปเซียนชิ้นแรกถูกค้นพบในกลุ่มหินโบฟอร์ตของกลุ่มหินคารูซู เปอร์กรุ๊ปในแอฟริกาใต้ โดย ริชาร์ด โอเวนนักชีววิทยาและนักบรรพชีวินวิทยาเขาจัดประเภทฟอสซิลเป็นGorgonops torvusโดยการรวมคำว่าGorgon ของกรีก ซึ่งเป็นสัตว์ในตำนาน เข้ากับคำว่าóps ( ὄψ ) ซึ่งหมายถึง 'ลักษณะ' [ 14 ]ในแอฟริกา ยังพบกอร์โกนอปเซียนในหินโผล่ของคารูในหุบเขารูฮูฮูของแทนซาเนียหุบเขาอัปเปอร์ลูอังวาของแซมเบียและชิเวตาประเทศมาลาวี[ 7 ] : 7
กอร์โกนอปเซียนถูกค้นพบครั้งแรกในรัสเซียในช่วงทศวรรษ 1890 ที่แหล่งโซโคลกีบนแม่น้ำดวีนาเหนือ ภายใต้การดูแลของนักบรรพชีวินวิทยาชาวรัสเซียวลาดิมีร์ โปรโคโรวิช อมาลิท สกี ในสิ่งพิมพ์ที่ตีพิมพ์หลังการเสียชีวิต มันถูกอธิบายว่าเป็นInostrancevia alexandri (สกุลนี้สะกดว่าInostranzevia ) และเป็นหนึ่งในกอร์โกนอปเซียนที่รู้จักกันดีและมีขนาดใหญ่ที่สุด นับตั้งแต่นั้นมา มีการ อธิบายอนุกรมวิธานของรัสเซียเพิ่มเติมอีกเพียงไม่กี่ชนิด ได้แก่Pravoslavlevia , Sauroctonus , Viatkogorgon , SuchogorgonและNochnitsa [ 15 ]
กอร์โกนอปเซียนนั้นหายากอย่างเห็นได้ชัดนอกเหนือจากสองพื้นที่นี้[ 15 ]ในปี 1979 นักบรรพชีวินวิทยาชาวจีนหยาง จงเจี้ยนได้บรรยายถึงกอร์โกนอปเซียนของจีน " Wangwusaurus tayuensis " โดยอิงจากฟันจากชั้นหินJiyuan Formationใน ยุคเพอร์เมียนตอนปลาย [ 16 ]แต่ในปี 1981 นักบรรพชีวินวิทยาเดนิส ซิกอกโน-รัสเซลล์และ ไอ-หลิน ซุน พบว่าวัสดุที่กำหนดนั้นเป็นกลุ่มแบบสุ่ม ซึ่งมีเพียงสองชิ้นเท่านั้นที่มีความคล้ายคลึงกับกอร์โกนอปเซียนแม้เพียงเล็กน้อย[ 17 ] ในปี 2022 นักบรรพชีวินวิทยา จุน หลิว และ หวัน หยาง ได้ยืนยันการมีอยู่ของกอร์โกนอปเซียนในประเทศ จีนโดยอิงจากชุดฟอสซิลฟันที่ไม่สมบูรณ์ที่ค้นพบในแอ่งทูร์ปาน ซิ นเจียง[ 18 ]ในปี 2546 นักบรรพชีวินวิทยาชาวอินเดีย Sanghamitra Ray และ Saswati Bandyopadhyay ได้จัดกลุ่มชิ้นส่วนกะโหลกบางส่วนจากชั้นหินKundaram Formation ยุคเพอร์เมียนตอนปลาย ของแอ่ง Pranhita–Godavariให้เป็นกอร์โกนอปเซียนขนาดกลาง[ 19 ]แม้ว่าลักษณะของกอร์โกนอปเซียนจะได้รับการบันทึกไว้ในเทโรเซฟาเลียนบางชนิดแล้วก็ตาม[ 15 ]ในปี 2551 ชิ้นส่วนขากรรไกรบนและฟันเขี้ยวขนาดใหญ่และน่าจะเป็นของรูบิดจีนถูกระบุที่ชั้นหินMoradi Formation ยุคเพอร์เมียนตอนปลาย ในประเทศไนเจอร์ (หนึ่งในไม่กี่ชั้นหินยุคเพอร์เมียนตอนปลายในละติจูดต่ำที่มีสัตว์สี่ขา) และเป็นหลักฐานแรกของกอร์โกนอปเซียนในละติจูดต่ำ[ 20 ]กอร์โกนอปเซียนในละติจูดต่ำตัวที่สองได้รับการอธิบายในปี 2567 จากชั้นหินPort des Canonge Formationของเกาะมายอร์กาในทะเลเมดิเตอร์เรเนียนตะวันตก ที่น่าสังเกตคือ มันน่าจะเป็นกอร์โกนอปเซียนที่เก่าแก่ที่สุดเท่าที่เคยพบในบันทึกฟอสซิล โดยมีอายุขั้นต่ำก่อนยุค Wordian ตอนกลาง หรืออาจจะก่อนยุค Wordian ด้วยซ้ำ ซึ่งแสดงให้เห็นว่าการไม่มีกอร์โกนอปเซียนในละติจูดต่ำสะท้อนถึงการสุ่มตัวอย่างที่ไม่สม่ำเสมอมากกว่าที่จะเป็นการจำกัดเฉพาะในละติจูดสูง[ 21 ]
การจำแนกประเภท
เมื่อค้นพบ โอเวนสันนิษฐานว่ากอร์โกนอปส์และแท็กซาอื่นๆ อีกหลายชนิดที่เขาอธิบายจากกลุ่มคารูซูเปอร์กรุ๊ปเป็น สัตว์เลื้อยคลาน เลือดเย็นแม้ว่าจะมีฟันที่คล้ายกับสัตว์เลี้ยงลูกด้วยนมกินเนื้อก็ตาม เขาเสนอให้จัดจำแนกพวกมันทั้งหมดไว้ภายใต้ลำดับเทริโอโดนเทีย (ซึ่งเขาจัดไว้ในชั้นเรปทิเลีย) เขาตัดสินใจแบ่งเทริโอโดนเทียออกเป็นวงศ์ต่างๆโดยพิจารณาจากกายวิภาคของรูจมูก (กระดูกจมูก) — "โมโนนาริอาเลีย" สำหรับสัตว์ที่มีรูจมูกเปิดเพียงรูเดียวในกะโหลกศีรษะเช่นเดียวกับสัตว์เลี้ยงลูกด้วยนม "ไบนาริอาเลีย" สำหรับสัตว์ที่มีรูจมูกเปิดสองรูเช่นเดียวกับสัตว์เลื้อยคลาน และ "เทคตินาริอาเลีย" สำหรับกอร์โกนอปส์เนื่องจากรูจมูกของมันถูกปกคลุมด้วยหลังคากระดูกหนา[ 14 ] ( เทคตัสเป็นภาษาละตินแปลว่า "ปกคลุม มีหลังคา ปู") [ 22 ]ในปี พ.ศ. 2433 นักธรรมชาติวิทยาชาวอังกฤษริชาร์ด ไลเดกเกอร์ได้กำหนดให้กอร์โกนอปส์เป็นชนิดต้นแบบของวงศ์กอร์โกนอปซิดา[ b ] [ 23 ]แฮร์รี ซีลีย์นักบรรพชีวินวิทยาชาวอังกฤษในปี 1894 เชื่อว่าGorgonopsขาดช่องเปิดในกระดูกขมับ (temporal fenestra) ซึ่งเป็นลักษณะเฉพาะของ Theriodontia ดังนั้นจึงยกระดับ Gorgonopsidae เป็น Gorgonopsia ซึ่งแตกต่างจาก Theriodontia เขาจัดจำแนกวัสดุจากแอฟริกาใต้ทั้งหมดที่มีทั้งลักษณะของสัตว์เลื้อยคลานและสัตว์เลี้ยงลูกด้วยนมไว้ในอันดับ "Theriosuchia" และถือว่า Gorgonopsia และ Theriodontia เป็นอันดับย่อยของมัน[ 24 ]เฮนรี แฟร์ฟิลด์ ออสบอร์นนักบรรพชีวินวิทยาชาวอเมริกันได้ปรับปรุงการจำแนกประเภทของ Reptilia ใหม่ทั้งหมดในปี 1903 และตั้งกลุ่มหลักสองกลุ่มคือDiapsida และ Synapsida [ 25 ] และ ในปี 1905 โรเบิร์ต บรูมนักบรรพชีวินวิทยาชาวแอฟริกาใต้ได้สร้างกลุ่มที่สามคือTherapsidaเพื่อรวบรวม "สัตว์เลื้อยคลานที่มีลักษณะคล้ายสัตว์เลี้ยงลูกด้วยนม" รวมถึง Theriodontia เขายังท้าทายคำกล่าวอ้างของ Seeley และลดระดับGorgonopsกลับไปเป็น Theriodontia แต่เขาจัดให้อยู่ในกลุ่มย่อยTherocephalia ที่เขาสร้างขึ้นใหม่ โดยยุบ Gorgonopsia [ 26 ] ในปี พ.ศ. 2456 โดยเฉพาะอย่างยิ่งเมื่อพิจารณาจากกะโหลก G. torvusที่เกือบสมบูรณ์ซึ่งค้นพบโดยบาทหลวง John H. Whaits บรูมจึงคืนสถานะ Gorgonopsia อีกครั้ง[ 27 ]
จำนวนสกุลของแอฟริกาใต้เติบโตอย่างรวดเร็วในศตวรรษที่ 20 โดยมี Broom เป็นผู้นำหลัก ซึ่งงานวิจัยมากมายของเขาเกี่ยวกับเทอราปซิดใน Karoo ตั้งแต่เริ่มต้นอาชีพของเขาในประเทศในปี 1897 จนถึงการเสียชีวิตของเขาในปี 1951 นำไปสู่การบรรยายลักษณะของตัวอย่างต้นแบบของ กอร์โกนอปเซียน 57 ตัวอย่าง และ 29 สกุล แท็กซาของ Broom หลายรายการถูกยกเลิกในภายหลัง[ 28 ]นักวิจัยร่วมสมัยคนอื่นๆ จำนวนมากสร้างสปีชีส์หรือสกุลใหม่ทั้งหมดโดยอาศัยตัวอย่างเพียงตัวเดียว[ 7 ] : 57 ด้วยเหตุนี้ Gorgonopsia จึงเป็นหัวข้อของความวุ่นวายทางอนุกรมวิธาน และเป็นหนึ่งในกลุ่มซินาปซิดที่มีปัญหามากที่สุด เนื่องจากกายวิภาคของกะโหลกศีรษะแตกต่างกันเพียงเล็กน้อยในแต่ละแท็กซา หลายกลุ่มจึงถูกกำหนดโดยอาศัยความแตกต่างของสัดส่วนที่ไม่ชัดเจน รวมถึงสมาชิกที่เป็นที่รู้จักกันดีด้วยสปีชีส์ตามชื่อเรียกจะแตกต่างกันโดยส่วนใหญ่จากลักษณะที่ทราบกันว่ามีความแปรผันค่อนข้างมากขึ้นอยู่กับอายุของแต่ละบุคคล รวมถึงขนาดเบ้าตาความยาวจมูก และจำนวนฟันหลังเขี้ยว ดังนั้นจึงเป็นไปได้ที่บางกลุ่มอนุกรมวิธานจะเป็นชื่อพ้องกัน และแสดงถึงระยะการพัฒนาที่ แตกต่างกัน [ 29 ]
ความพยายามครั้งแรกในการจัดกลุ่มสายพันธุ์นี้ดำเนินการโดยนักสัตววิทยาชาวอังกฤษDavid Meredith Seares WatsonและนักบรรพชีวินวิทยาชาวอเมริกันAlfred Romerในปี 1956 ซึ่งได้แบ่งสายพันธุ์นี้ออกเป็น 20 วงศ์[ 30 ]โดยสมาชิก 3 วงศ์ (Burnetiidae, Hipposauridae และ Phthinosuchidae) ไม่ถือว่าเป็นกอร์โกนอปเซียนอีกต่อไป[ 6 ]ในปี 1970 และอีกครั้งในปี 1989 โดยส่วนใหญ่พิจารณาจากกลุ่มสิ่งมีชีวิตในแอฟริกา Sigogneau-Russell ได้ตีพิมพ์เอกสาร ทางวิชาการที่ครอบคลุม เกี่ยวกับกอร์โกนอปเซีย (โดยกำหนดให้เป็นอันดับย่อย ) และยอมรับเพียง 2 วงศ์ ได้แก่ Watongiidae และ Gorgonopidae [ 31 ] Watongiaถูกย้ายไปอยู่ในVaranopidaeในปี 2004 โดยนักบรรพชีวินวิทยาชาวแคนาดาRobert R. ReiszและMichel Laurin [ 32 ] Sigogneau-Russell ได้แบ่ง Gorgonopidae ออกเป็นสามวงศ์ย่อย ได้แก่ Gorgonopsinae, Rubidgeinae และ Inostranceviinae และลดจำนวนสกุลเหลือเพียงยี่สิบสามสกุล[ 31 ]ในปี 2002 นักบรรพชีวินวิทยาชาวรัสเซียMikhail Feodosievich Ivakhnenkoได้พิจารณาอนุกรมวิธานของรัสเซีย และถือว่า Gorgonopsia เป็นอันดับย่อย และจัดกลุ่มร่วมกับDinocephaliaไว้ในอันดับ "Gorgodontia" เขาแบ่ง Gorgonopsia ออกเป็นวงศ์ใหญ่ "Gorgonopioidea" (วงศ์ Gorgonopidae, Cyonosauridae และ Galesuchidae) และ "Rubidgeoidea" (วงศ์ Rubidgeidae, Phtinosuchidae และ Inostranceviidae) [ 33 ]ในปี 2550 นักชีววิทยา Eva VI Gebauer ในการทบทวน Gorgonopsia อย่างครอบคลุม (วิทยานิพนธ์ปริญญาเอกของเธอ) ได้ปฏิเสธแบบจำลองของ Ivakhnenko และเลือกใช้แบบจำลองของ Sigogneau-Russell แทน และยังลดจำนวนสกุลลงเหลือสิบสี่สกุล นอกเหนือจากสกุลของรัสเซีย ได้แก่Aloposaurus , Cyonosaurus , Aelurosaurus , Sauroctonus , Scylacognathus , Eoarctops , Gorgonops , Njalila (ได้รับการอธิบายอย่างเป็นทางการในปี 2569 [ 34 ] ), Lycaenops , Arctognathus , Aelurognathus , Sycosaurus , ClelandinaและRubidgea [ 7 ] : 57, 244 โดยทั่วไป แบบจำลองของ Sigogneau-Russell ได้รับการสนับสนุน แต่ยังไม่มีข้อสรุปที่ แน่ชัดว่าสกุลใดควรจัดอยู่ในวงศ์ย่อยใด[ 6]ในปี 2015 นักบรรพชีวินวิทยาชาวอเมริกัน Christian F. Kammerer และเพื่อนร่วมงานได้อธิบาย Eriphostoma ใหม่ (ซึ่งถูกระบุว่าเป็น theriodont ที่ไม่สามารถระบุชนิดได้) ว่าเป็น gorgonopsian [ 35 ]และรวมScylacognathusและEoarctopsไว้ในกลุ่ม เดียวกันในปีถัดมา [ 6 ]
แผนภูมิวิวัฒนาการ (แผนผังวงศ์ตระกูล) แรกของสมาชิกในสกุล Gorgonopsia ได้รับการตีพิมพ์ในปี 2016 โดยนักบรรพชีวินวิทยาชาวอเมริกัน Christian F. Kammerer ซึ่งได้ทำการศึกษา Rubidgeinae โดยเฉพาะ และได้บรรยายลักษณะของวงศ์ย่อยและเก้าชนิดที่เขากำหนดไว้ใหม่ (ลดจำนวนจากสามสิบหกชนิด) Kammerer ยังได้ฟื้นฟูDinogorgon , Leontosaurus , RuhuhucerberusและSmilesaurus ขึ้นมาใหม่ ด้วย Kammerer ไม่แน่ใจว่าLeontosaurus , Clelandina , DinogorgonและRubidgeaเป็นกลุ่มอนุกรมวิธานเดียวกันหรือไม่ (ซึ่งDinogorgonมีลำดับความสำคัญเหนือกว่า ) แต่เขาตัดสินใจจัดพวกมันทั้งหมดไว้ในเผ่า Rubidgeini เพื่อรอการตรวจสอบเพิ่มเติม[ 6 ]ในปี 2018 Kammerer และ Vladimir Masyutin นักบรรพชีวินวิทยาชาวรัสเซีย ได้ระบุสกุลใหม่Nochnitsaว่าเป็นกอร์โกนอปเซียนที่เก่าแก่ที่สุดเท่าที่รู้จัก และพบว่าแท็กซาของรัสเซียทั้งหมด (ยกเว้นViatkogorgonซึ่งอยู่ในกลุ่มนอก) ก่อตัวเป็นกลุ่มที่แยกออกจากแท็กซาของแอฟริกาโดยสิ้นเชิง[ 15 ]ในปี 2018 เช่นกัน นักบรรพชีววิทยา Eva-Maria Bendel, Kammerer และเพื่อนร่วมงานได้ฟื้นคืนชีพCynariops [ 1 ] ในปี 2022 Kammerer และ Bruce S. Rubidge นักบรรพชีวินวิทยา ได้อธิบายPhorcysจากแอฟริกาใต้[ 36 ] Macungo และเพื่อนร่วมงาน (2026) ได้อธิบายJirahgorgonว่าเป็นแท็กซาใหม่ของแอฟริกาใต้ที่มีความสัมพันธ์ใกล้ชิดกับPhorcysพวกเขายอมรับว่าสกุลทั้งสองนี้เป็นกลุ่มย่อยของกอร์โกนอปเซียนขนาดค่อนข้างใหญ่จากยุคเพอร์เมียนตอนกลาง ซึ่งแยกตัวออกจากกอร์โกนอปเซียนขนาดใหญ่ในยุคหลัง (เช่นInostranceviaและ rubidgeines) ที่พวกเขาตั้งชื่อเป็นวงศ์ใหม่ว่า Phorcyidae การวิเคราะห์ทางวิวัฒนาการของพวกเขาพบว่าสกุลทั้งสองนี้อยู่ในกลุ่มย่อยที่แตกแขนงในช่วงต้นของกอร์โกนอปเซียนแอฟริกา ซึ่งรวมถึงEriphostomaและGorgonopsด้วย[ 37 ]
| |||||||||||||||||||||
| วิวัฒนาการของ Gorgonopsia ตาม Macungo et al. (2026) [ 37 ] |
วิวัฒนาการ
โดยทั่วไปแล้ว Synapsida ถูกแบ่งออกเป็นสองกลุ่มหลักคือ " Pelycosauria " และกลุ่มที่พัฒนาต่อมาคือ Therapsida กลุ่มแรกประกอบด้วยสิ่งมีชีวิตเลือดเย็นที่มีการเดินแบบแผ่กว้างและคาดว่ามีการเผาผลาญต่ำกว่า ซึ่งวิวัฒนาการขึ้นในยุคคาร์บอนิเฟอรัสตอนบนในช่วงกลางถึงปลายศตวรรษที่ 20 นักบรรพชีวินวิทยาชาวอเมริกันEverett C. Olsonได้ศึกษาความหลากหลายของ Synapsida ใน ชั้นหิน San Angelo , FlowerpotและChickashaในยุคเพอร์เมียนตอนกลางของทวีปอเมริกาเหนือ และพบว่าความหลากหลายของ Pelycosaur ลดลงจากหกชนิดเหลือสามชนิดในชั้นหินเหล่านี้ และพวกมันอยู่ร่วมกับซากดึกดำบรรพ์ที่แตกหักหลายชิ้นซึ่งเขาตีความว่าเป็น Therapsida จากนั้นเขาเสนอว่าการเปลี่ยนแปลงทางวิวัฒนาการจากระดับ Pelycosaur ไปสู่ระดับ Therapsida เกิดขึ้นในช่วงยุคเพอร์เมียนตอนกลาง ( การสูญพันธุ์ของ Olson ) อย่างไรก็ตาม การจัดประเภทของ "Therapsida" เหล่านั้นและอายุของชั้นหินก็ถูกท้าทายในภายหลัง ดังนั้น ช่วงเวลาที่แน่นอนของการยึดครองของเทอราปซิดจึงไม่ชัดเจน แต่กลุ่มเทอราปซิดหลักทั้งหกกลุ่ม ( Biarmosuchia , Dinocephalia , Anomodontia , Gorgonopsia, Therocephalia และ Cynodontia) ได้วิวัฒนาการขึ้นเมื่อ 265 ล้านปีก่อนในช่วงยุค Wordian [ 38 ]
| ซินาปซิดา[ 38 ] |

ฟอสซิลกอร์โกนอปเซียนที่เก่าแก่ที่สุดในโลกที่ได้รับการยืนยันแล้ว มาจากชั้นหินปอร์ต เดส์ คาโนนจ์ (Port des Canonge Formation) บนเกาะมายอร์กาทางตะวันตกของทะเลเมดิเตอร์เรเนียน แม้ว่าการกำหนดอายุของหินในชั้นนี้จะไม่แม่นยำนัก แต่ การศึกษาทาง ธรณี แม่เหล็กวิทยา ละอองเรณูวิทยาและร่องรอย ฟอสซิล บ่งชี้ว่าอายุของมันอย่างน้อยที่สุดต้องอยู่ในช่วงต้นยุคเวิร์ดเดียน (Wordian) อย่างไรก็ตาม ตำแหน่ง ทางธรณีวิทยาของตัวฟอสซิลกอร์โกนอปเซียนเองและ ร่องรอย ฟอสซิล โดยรอบ อาจบ่งชี้ว่ามันมีอายุเก่าแก่กว่ามาก อาจเป็นยุคโรเดียน (Roadian ) ในช่วงต้นยุคกั ว ดาลูเปียน (Guadalupian ) (ยุคเพอร์เมียนตอนกลาง) หรือแม้กระทั่ง ยุคคุงกูเรียน ( Kungurian ) หรืออาร์ตินสเกียน (Artinskian) ของยุคซิซูราเลียน (Cisuralian ) (ยุคเพอร์เมียนตอนต้น) หากมีอายุเช่นนั้น มันจะไม่เพียงแต่เป็นฟอสซิลกอร์โกน อปเซียนที่เก่าแก่ที่สุดเท่าที่เคยพบเท่านั้น แต่ยังเป็นเทอแรปซิด (Therapsid) ที่เก่าแก่ที่สุดอีกด้วย แม้ว่าจะไม่สามารถระบุชนิดได้แน่ชัด แต่กายวิภาคของกอร์โกนอปเซียนแห่งมาลอร์กาชี้ให้เห็นว่ามันมีวิวัฒนาการมากกว่ากอร์โกนอปเซียนที่แยกตัวออกมาเร็วที่สุดที่ได้รับการยอมรับอย่างNochnitsaและสอดคล้องกับตำแหน่งภายในหรืออยู่นอกฐานของกลุ่ม "แอฟริกัน" และ "รัสเซีย" ซึ่งแสดงให้เห็นว่าไม่เพียงแต่กอร์โกนอปเซียนจะมีอยู่ใน Wordian ตอนต้น หากไม่ใช่ Cisuralian ตอนปลาย แต่การกระจายตัวของพวกมันก็ดำเนินไปอย่างดีแล้วในช่วงเวลานี้ และกลุ่มโดยรวมน่าจะกำเนิดขึ้นก่อนช่วงเวลานี้ นอกจากนี้ยังแสดงให้เห็นว่าความหลากหลายของกอร์โกนอปเซียน (และเทอแรปซิด) ในยุคแรกไม่ได้จำกัดอยู่เฉพาะละติจูดสูงของแพนเจีย ในเขตอบอุ่น ดังที่เคยเสนอไว้ก่อนหน้านี้จากการอ่านบันทึกฟอสซิลตามตัวอักษร แต่ยังรวมถึงละติจูดต่ำใกล้เส้นศูนย์สูตรของมหาทวีป (ซึ่งมาลอร์กาตั้งอยู่) ด้วย[ 21 ]
กอร์โกนอปเซียนที่เก่าแก่ที่สุดในละติจูดสูงทั้งสองชนิดมาจากกลุ่มหิน Karoo Sugergroup ของแอฟริกาใต้ ชนิดที่เก่ากว่าคือส่วนจมูกบางส่วนของสกุลขนาดใหญ่ที่ไม่สามารถระบุได้จากเขตEodicynodon Assemblage Zoneของแอ่ง Karoo ซึ่งมีอายุประมาณ Wordian ส่วนPhorcysซึ่งพบจากปลายสุดของ เขต Tapinocephalus Assemblage Zone ของ Karoo (มีอายุประมาณช่วง Wordian/Capitanian boundary) เป็นกอร์โกนอปเซียนที่เก่าแก่ที่สุดที่สามารถระบุได้ แม้ว่าจะแตกหักเป็นชิ้นเล็กชิ้นน้อย แต่คาดว่าทั้งสองชนิดจาก Karoo มีกะโหลกยาวประมาณ 30 เซนติเมตร (12 นิ้ว) [ 36 ]ซึ่งใหญ่กว่ากะโหลกของกอร์โกนอปเซียน Mallorcan ที่มีความยาวประมาณ 18 เซนติเมตร (7.1 นิ้ว) [ 21 ]และEriphostoma ที่มีอายุน้อยกว่าเล็กน้อยอย่างมีนัยสำคัญ ซึ่งแสดงให้เห็นว่าขนาดตัวของกอร์โกนอปเซียนก็มีความหลากหลายตั้งแต่ช่วงต้นของการวิวัฒนาการเช่นกัน[ 36 ]

ยุคเพอร์เมียนค่อยๆ แห้งแล้งลงเรื่อยๆ ในยุคคาร์บอนิเฟอรัสตอนบนและเพอร์เมียนตอนล่าง เพลิโคซอร์ดูเหมือนจะยังคงอาศัยอยู่ในแหล่งที่อยู่อาศัยที่เป็นหนองน้ำถ่านหินชื้นแฉะใกล้เส้นศูนย์สูตร (พบฟอสซิลภายในระยะ 10° จากทั้งสองด้านของเส้นศูนย์สูตรโบราณ) เลยจากนี้ไปจนถึงประมาณ 30° เป็นทะเลทรายกว้างใหญ่ที่ทอดยาวไปจนถึงชายฝั่ง แยกหนองน้ำออกจากเขตอบอุ่น ในยุคเพอร์เมียนตอนกลาง ป่าเขตร้อนได้เปลี่ยนไปเป็นระบบชื้น/แห้งตามฤดูกาล แต่หนองน้ำเชื่อมต่อกับเขตอบอุ่นผ่านทางช่องชายฝั่งตามแนวแพนเจียตะวันออก ทำให้เกิดการอพยพข้ามทวีปจากที่ปัจจุบันคือแอฟริกาใต้ไปยังที่ปัจจุบันคือรัสเซีย เทอราปซิดดูเหมือนจะวิวัฒนาการในภูมิทัศน์ที่ชื้น/แห้งตามฤดูกาลนี้ ขยายตัวไปถึงเขตอบอุ่นด้วย ในช่วงเวลานี้ ซินาปซิดเป็นสัตว์บกขนาดใหญ่เพียงชนิดเดียวในสภาพแวดล้อมนั้น และเพลิโคซอร์อาจไม่สามารถปรับตัวให้เข้ากับการแห้งแล้งได้ ในช่วงเวลาใกล้เคียงกับการสูญพันธุ์ของเพลิโคซอร์ เทราปซิดส์ประสบกับการแพร่กระจายทาง วิวัฒนาการครั้งใหญ่ (สัตว์กินเนื้อทั้งหมด) ซึ่งต่อเนื่องมาจนถึงยุคเพอร์เมียนตอนบน[ 38 ]
ตลอดช่วงยุคเพอร์เมียนตอนกลาง ไดโนเซฟาเลียนซึ่งมักมีขนาดใหญ่โตมโหฬารเป็นสัตว์เด่นในระบบนิเวศของพวกมัน โดยเทโรเซฟาเลียน ขั้นพื้นฐาน เข้ามาแทนที่กลุ่มนักล่าเขี้ยวเสือ[ 39 ] [ 36 ]พวกมันหายไปจากบันทึกฟอสซิลในช่วงเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในยุคคา ปิทาเนียน ซึ่งเกิดจากกิจกรรมภูเขาไฟที่ก่อให้เกิดหินภูเขาไฟเอ๋อเหมยซาน ในประเทศจีน สาเหตุที่แท้จริงของการสูญพันธุ์ยังไม่ชัดเจน แต่พวกมันถูกแทนที่ด้วยกอร์โกนอปเซียนและไดซิโนดอนต์ (ซึ่งเริ่มมีขนาดใหญ่ขึ้นอย่างมาก) และเทโรเซฟาเลียนที่มีขนาดเล็กกว่า[ 40 ]เชื่อกันว่าการสูญพันธุ์ของเทโรเซฟาเลียนขั้นพื้นฐานทำให้กอร์โกนอปเซียนมีความหลากหลาย มากขึ้น [ 36 ] [ 41 ]โดยการเข้ามาแทนที่น่าจะเกิดจากกอร์โกนอปเซียนสามารถล่าเหยื่อได้หลากหลายชนิดมากขึ้น[ 42 ]รูบิดจีนเป็นกอร์โกนอปเซียนที่มีวิวัฒนาการมากที่สุด และด้วยเหตุนี้จึงมีขนาดใหญ่ที่สุดและมีโครงสร้างที่แข็งแรงที่สุด[ 6 ]
บรรพชีววิทยา
กัด

กอร์โกนอปเซียนน่าจะเป็นนักล่าที่กระตือรือร้น รูบิดจีนมีกะโหลกที่แข็งแรงเป็นพิเศษในบรรดากอร์โกนอปเซียน เทียบได้กับกะโหลกของสัตว์นักล่าขนาดใหญ่ที่ใช้กะโหลกเป็นอาวุธหลัก เช่นโมซาซอร์หรือไดโนเสาร์เทอโรพอ ดบางชนิด กอร์โกนอปเซียนที่แข็งแรงน้อยกว่า มีเขี้ยวที่ยาวกว่าและแรงกัดที่อ่อนแอกว่ามาก เช่นสไมล์ซอรัสหรืออินอสทรานเซเวียน่าจะใช้เขี้ยวในการฟัน ซึ่งคล้ายกับแมวเขี้ยวเสือมากกว่า เขี้ยวหลังของเคลแลนดินาถูกแทนที่ด้วยสันเรียบ ซึ่งแตกต่างจากไดไซโนดอนต์ที่มีสันเคราติน คล้ายใบมีด และมันอาจจะล่าเหยื่อที่สามารถกลืนได้ทั้งตัวเป็นหลัก กลุ่มกอร์โกนอปเซียนอยู่ร่วมกันได้—มากถึงเจ็ดกลุ่มในเวลาเดียวกัน—และข้อเท็จจริงที่ว่ารูบิดจีนบางตัวมีเขี้ยวหลังในขณะที่บางตัวในยุคเดียวกันไม่มี แสดงให้เห็นว่าพวกมันมีการแบ่งส่วนนิเวศวิทยาและล่าเหยื่อที่แตกต่างกัน[ 6 ]
โดยทั่วไปแล้วเชื่อกันว่าเขี้ยวที่ยาวนั้นมีบทบาทสำคัญในกลยุทธ์การล่าของพวกมันข้อต่อขากรรไกร ของกอร์โกนอปเซียน เป็นข้อต่อคู่และประกอบด้วยกระดูกที่เคลื่อนไหวและหมุนได้ ซึ่งจะช่วยให้พวกมันอ้าปากได้กว้างมาก—อาจเกิน 90°—โดยไม่ต้องถอดข้อต่อขากรรไกรออก[ 43 ]ในทางกลับกัน มีการเสนอแนะ (ครั้งแรกในปี 2002 โดยนักชีววิทยา Blaire Van Valkenburgh และ Tyson Secco แม้ว่าจะอ้างอิงถึงแมวก็ตาม) ว่าเขี้ยวเสือวิวัฒนาการขึ้นเป็นหลักเนื่องจากการคัดเลือกทางเพศในรูปแบบของการแสดงออกใน การผสมพันธุ์ สิ่งนี้แสดงให้เห็นในกวางบางชนิดในปัจจุบัน แต่ยากที่จะทดสอบเนื่องจากขาดผู้ล่าซินาปซิดที่มีเขี้ยวเสือที่ยังมีชีวิตอยู่ ในแมวเขี้ยวเสือ เชื่อกันว่าสายพันธุ์ที่มีเขี้ยวยาว ("เขี้ยวแหลม") เป็นนักล่าแบบไล่ล่าในขณะที่สายพันธุ์ที่มีเขี้ยวสั้น ("เขี้ยวโค้ง") เชื่อกันว่าเป็น นักล่าแบบ ซุ่มโจมตี[ 44 ]ในบรรดาแมวเขี้ยวแหลมคม มีข้อเสนอแนะว่านักล่าเหล่านี้จะฆ่าเหยื่อด้วยการฟันที่คออย่างแม่นยำหลังจากจับเหยื่อได้ แต่กอร์โกนอปเซียนอาจไม่แม่นยำในการกัด เนื่องจากมีขากรรไกรและฟันแบบสัตว์เลื้อยคลาน แทนที่จะเป็นเช่นนั้น กอร์โกนอปเซียนอาจใช้กลยุทธ์กัดแล้วถอยหนี: นักล่าจะซุ่มโจมตีเหยื่อและกัดเหยื่ออย่างรุนแรงและทำให้เหยื่ออ่อนแอลง จากนั้นจะตามไปขณะที่เหยื่อพยายามหนี ก่อนที่จะตายเพราะบาดเจ็บ จากนั้นกอร์โกนอปเซียนจะกัดเพื่อฆ่า เนื่องจากฟันหลังเขี้ยวลดลงหรือไม่มีเลย เนื้อจึงถูกฉีกออกจากซากอย่างแรงและกลืนลงไปทั้งชิ้น[ 43 ]กลยุทธ์ "เจาะ-ดึง" นี้ยังถูกตั้งสมมติฐานว่าไดโนเสาร์เทอโรพอดก็ใช้เช่นกัน[ 10 ]

กอร์โกนอปเซียน เช่นเดียวกับสัตว์กินเนื้อยุคแรกอื่นๆ รวมถึงจระเข้ ส่วนใหญ่อาศัย "ระบบจลน์-เฉื่อย" (KI) ในการกัดเหยื่อ โดยที่กล้ามเนื้อปีกนกและกล้ามเนื้อขมับจะหนีบขากรรไกรปิดอย่างรวดเร็ว โดยใช้โมเมนตัมและพลังงานจลน์ของขากรรไกรและฟันในการจับเหยื่อ สัตว์กินเนื้อเลี้ยงลูกด้วยนม รวมถึงแมวเขี้ยวเสือ อาศัย "ระบบแรงดันสถิต" (SP) เป็นหลัก โดยที่กล้ามเนื้อขมับและกล้ามเนื้อบดเคี้ยวจะสร้างแรงกัดที่แข็งแกร่งเพื่อฆ่าเหยื่อ[ 11 ]กล้ามเนื้อขมับและกล้ามเนื้อบดเคี้ยวแยกออกจากกันเฉพาะในสัตว์เลี้ยงลูกด้วยนมเท่านั้น และกอร์โกนอปเซียนมีกล้ามเนื้อที่ทอดยาวจากด้านล่างของหลังคากะโหลกกลับไปยังกระดูกขมับ (ที่ด้านหลังของกะโหลก) และข้ามกระดูกแก้ม ส่วนที่ยึดกับแก้มจะทำให้กระดูกขากรรไกรคงที่และช่วยให้มันเคลื่อนที่ไปด้านข้างขณะปิด สิ่งนี้อาจมีความสำคัญมากในการกัด เนื่องจากกระดูกแก้มจะแข็งแรงขึ้นพร้อมกับฟันเขี้ยวที่ยาวขึ้น[ 7 ] : 278
กอร์โกนอปเซียนขนาดเล็ก เช่นไซโอโนซอรัส (ซึ่งอาจเป็นลูกของสายพันธุ์อื่น) มีกะโหลกและเขี้ยวที่เรียวบาง และอาจมีพฤติกรรมคล้ายกับหมาจิ้งจอกและสุนัขจิ้งจอก กอร์โกนอปเซียนขนาดใหญ่ เช่นกอร์โกนอปส์ มีจมูกยาวและแข็งแรง มีแก้มที่บานออกอย่างเห็นได้ชัด ซึ่งจะช่วยรองรับกระดูกปีกที่แข็งแรงและแรงกัดแบบ KI ที่ทรงพลัง อาร์คทอกนาทัสขนาดกลางมีกะโหลกเป็นรูปทรงกล่องและจมูกที่ทรงพลัง ซึ่งจะช่วยให้สามารถงอและบิดตัวได้อย่างมาก และมีการผสมผสานขององค์ประกอบการกัดทั้งแบบ KI และ SP กอร์โกนอปเซียนที่ใหญ่กว่านั้น เช่นอาร์คทอปส์มีจมูกที่สั้นกว่าและนูนกว่าเหมือนกับไดเมโทรดอน สเฟเนโคดอนต์ในยุคแรกๆ และจะสามารถงับขากรรไกรปิดได้อย่างรวดเร็วจากปากที่กว้าง (ซึ่งจำเป็นเนื่องจากมีเขี้ยวที่ยาว) Rubidgeinae ที่มีขนาดใหญ่กว่านั้นมีกะโหลกศีรษะที่แข็งแรงมาก สร้างขึ้นอย่างมั่นคง มีจมูกกว้าง แก้มยื่นออกมาอย่างเห็น ได้ชัด และฟันยาวมาก ดาบของRubidgea atroxยาวกว่าฟันของTyrannosaurus [ 11 ]
ต่างจากสัตว์กินเนื้อเลี้ยงลูกด้วยนม กอร์โกนอปเซียน (และเทอโรเซฟาเลียน) มีฟันหลังเขี้ยวลดลงหรือไม่มีเลย และขากรรไกรอาจไม่สามารถออกแรงเฉือนที่จำเป็นสำหรับการบดกระดูกให้เปิดออกเพื่อเข้าถึงไขกระดูกได้[ 45 ]ยังไม่เป็นที่แน่ชัดว่าไขกระดูกได้วิวัฒนาการขึ้นในไซแนปซิดยุคเพอร์เมียนแล้วหรือไม่ (ปลาและสัตว์สะเทินน้ำสะเทินบกหลายชนิดในปัจจุบันไม่มีไขกระดูก) แต่ในปี 2021 ได้มีการแสดงให้เห็นว่าสัตว์สะเทินน้ำสะเทินบก ยุคเพอร์เมียนตอนต้นอย่าง SeymouriaและDiscosauriscusน่าจะมีไขกระดูกที่สร้างเม็ดเลือดแดงในแขนขา[ 46 ]
การเคลื่อนที่
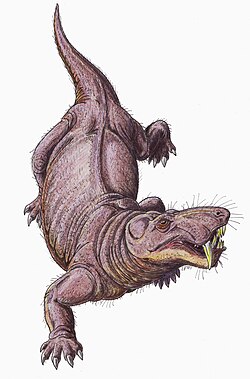
กอร์โกนอปเซียนถือว่าอาศัยอยู่บนบกอย่าง เคร่งครัด [ 47 ]เชื่อกันว่าพวกมันสามารถเคลื่อนที่ด้วยท่าเดินตั้งตรงคล้ายกับที่จระเข้ใช้ โดยวางแขนขาเกือบเป็นแนวตั้ง ตรงข้ามกับแนวนอนเหมือนท่าเดินกางขาของกิ้งก่าโพรงข้อต่อบนกระดูกสะบักเอียงไปทางหางอย่างมาก ดังนั้นแขนขาจึงมีการเคลื่อนไหวไปข้างหน้าอย่างจำกัด และอาจมีระยะก้าวสั้น กิ้งก่ามักจะขยับกระดูกสันหลังไปด้านข้างเพื่อเพิ่มระยะก้าว แต่ข้อต่อกระดูกสันหลังที่วางตัวในแนวตั้งมากกว่าในกอร์โกนอปเซียนจะทำให้กระดูกสันหลังแข็งและมั่นคงมากขึ้น ทำให้การเคลื่อนไหวเช่นนั้นเป็นไปได้ยาก[ 7 ] : 259–260
ข้อต่อหัวไหล่ของกอร์โกนอปเซียนมีโครงสร้างที่ผิดปกติอย่างมาก ส่วนหัวของกระดูกต้นแขนที่เชื่อมต่อกับหัวไหล่นั้นยาวกว่าเบ้ากระดูกข้อต่อหัวไหล่ (glenoid) ดังนั้นจึงไม่สามารถพอดีกับเบ้ากระดูกได้ ด้วยเหตุนี้ พวกมันจึงอาจเชื่อมต่อกันด้วยกระดูกอ่อนจำนวนมาก โดยที่กระดูกต้นแขนจะเคลื่อนที่แบบหมุนอยู่เหนือเบ้ากระดูกข้อต่อหัวไหล่ ในทางทฤษฎีแล้ว มุมระหว่างกระดูกต้นแขนและเบ้ากระดูกข้อต่อหัวไหล่จึงอาจมีตั้งแต่ 80 ถึง 145 องศาเมื่อมองจากด้านหน้าของสัตว์ หากมุมอยู่ทางด้านต่ำ ข้อต่อนี้จะค่อนข้างแข็งแรง ทำให้กล้ามเนื้อเดลทอยด์สามารถออกแรงได้มากผ่านแขนขาด้านหน้า เช่น เมื่อจับเหยื่อที่ดิ้นรน หรือยึดซากสัตว์ไว้ขณะฉีกเนื้อ หากกระดูกต้นแขนอยู่ในตำแหน่งมุมที่สูงขึ้น จะช่วยให้การเหยียดไปข้างหน้าและข้างหลัง (ตามแกนยาว) ดีขึ้น และทำให้ก้าวเดินยาวขึ้น ซึ่งมีประโยชน์ในการโจมตีหรือไล่ล่าในระยะสั้น กระดูกสะบักจะยื่นออกไปด้านข้างของสัตว์ (ยื่นออกไปด้านข้าง) และยังเป็นจุดยึดขนาดใหญ่สำหรับกล้ามเนื้อเดลตอยด์กล้ามเนื้อสแคปูโลฮิ วเมอรัลทั้งหมด มีจุดยึดที่พัฒนาอย่างแข็งแรง โดยเฉพาะอย่างยิ่งกล้ามเนื้อเดลตอยด์ เมื่อเหยียดแขนขาหน้ากล้ามเนื้อ เดลตอยด์อาจ ยกด้านหน้า (ขอบด้านหน้า) ของกระดูกฮิวเมอรัสขึ้น และกล้ามเนื้อโคราโคบราเคียลิสจะลดด้านหลัง (ขอบด้านหลัง) ลง เมื่อหดแขนขาหน้ากล้ามเนื้อเพคโทราลิสอาจดันขอบด้านหน้าลง และกล้ามเนื้อซับสแคปูลาริสจะดึงขอบด้านหลังขึ้น[ 7 ] : 260–264
ข้อต่อเชิงกรานมี ลักษณะเป็นข้อ ต่อแบบลูกบอลและเบ้า ตามปกติ หัวกระดูกต้นขาที่ค่อนข้างแบนนั้นในทางทฤษฎีแล้วสามารถเข้ากับเบ้าสะโพกได้ในมุมที่หลากหลาย ในปี 1982 นักบรรพชีวินวิทยา ทอม เอส. เคมป์ เสนอว่าเทอริโอดอนต์ยุคแรกๆ รวมถึงกอร์โกนอปเซียน สามารถวางกระดูกต้นขาได้ทั้งในมุมแนวนอนเมื่อเดินแบบกางขา และในมุมที่ตั้งฉากมากขึ้นเมื่อเดินแบบยืนตรง เขาเปรียบเทียบพฤติกรรมการเคลื่อนที่ของสิ่งมีชีวิตเหล่านี้กับจระเข้ ซึ่งใช้การเดินแบบกางขาในระยะทางสั้นๆ แต่เปลี่ยนเป็นการเดินแบบยืนตรงเมื่อวิ่งหรือเคลื่อนที่ในระยะทางไกล แม้ว่าสะโพกของตัวอย่างที่กำหนดหมายเลขเป็น GPIT/RE/7113 ดูเหมือนจะมีลักษณะทางกายวิภาคอยู่ระหว่างไดเมโทรดอนและสัตว์เลี้ยงลูกด้วยนม โดยที่กระดูกเชิงกรานขยายออกไปทางหัวมากกว่าทางหาง และกระดูกหัวหน่าวลดขนาดลงเล็กน้อย แต่กล้ามเนื้อพิวโบอิสคิโอเฟโมราลิส (กล้ามเนื้อขนาดใหญ่ที่มีเฉพาะในสัตว์เลื้อยคลานซึ่งทอดยาวจากกระดูกเชิงกรานไปยังกระดูกต้นขา) ยึดติดกับด้านล่างของกระดูกหัวหน่าวและ กระดูก อิสคิโอ อย่างกว้างขวาง ซึ่งจะช่วยให้สามารถสร้าง แรงดึงขา เข้าหาลำตัว ที่แข็งแรง (ดึงขาเข้าหาลำตัว) ซึ่งมีประโยชน์ในการเดินแบบกางขา นอกจากนี้ยังเป็นไปได้ว่ากอร์โกนอปเซียนใช้กล้ามเนื้อนี้เป็นหลักในขณะที่จับเหยื่อที่ดิ้นรน[ 7 ] : 264–270 หน้าแข้งค่อนข้างสั้นเมื่อเทียบกับกระดูกต้นขา ซึ่งบ่งชี้ว่ากอร์โกนอปเซียนไม่ได้ปรับตัวได้ดีสำหรับการวิ่งระยะทางไกล[ 7 ] : 298–299

ในส่วนของการวางเท้าบนพื้นนั้น กอร์โกนอปเซียนเป็นเทอแรปซิดยุคแรกเพียงกลุ่มเดียวที่มีลักษณะเอกตาแท็กโซนี (นิ้วสุดท้ายรับน้ำหนักมากที่สุด) โฮโมโพดี (รอยเท้าและรอยมือดูเหมือนกัน) และเซมิ-แพลนติกราดี (เท้าวางราบกับพื้นในระดับหนึ่ง) [ 48 ]การปรับตัวเหล่านี้อาจทำให้กอร์โกนอปเซียนว่องไวและคล่องแคล่วกว่าเหยื่อของพวกมัน[ 43 ]กอร์โกนอปเซียนมีนิ้วที่ค่อนข้างคล่องแคล่ว ซึ่งบ่งชี้ถึงความสามารถในการจับยึดทั้งมือและเท้า อาจเพื่อจับเหยื่อที่ดิ้นรนเพื่อป้องกันการรับน้ำหนักมากเกินไป และการแตกหักหรือฉีกขาดของเขี้ยวขณะที่กัดเหยื่อ[ 13 ]
ประสาทสัมผัส

ต่างจากยูเทอริโอดอนต์ แต่เหมือนกับสิ่งมีชีวิตเลือดเย็นบางชนิดในปัจจุบัน กอร์โกนอปเซียนทั้งหมดมีตาไพเนียลอยู่บนหัว ซึ่งใช้ในการตรวจจับแสงแดด (และด้วยเหตุนี้ อุณหภูมิที่เหมาะสมที่สุดในการทำกิจกรรม) เป็นไปได้ว่าเทอริโอดอนต์อื่นๆ สูญเสียสิ่งนี้ไปเนื่องจากการวิวัฒนาการของภาวะเลือดอุ่นเซลล์แกงลีออนเรตินาที่ไวต่อแสงในดวงตา—ควบคู่ไปกับการสูญเสียการมองเห็นสีและการเปลี่ยนไปใช้ชีวิตกลางคืน—หรือทั้งสองอย่าง[ 49 ] พฤติกรรม กลางคืนได้รับการสันนิษฐานมานานแล้วว่ามีต้นกำเนิดในสัตว์เลี้ยงลูกด้วยนม ( คอขวดกลางคืน ) แต่ขนาดเบ้าตาขนาดใหญ่และการมีวงแหวนสเคลราในไซแนปซิดยุคแรกๆ จำนวนมาก ซึ่งย้อนกลับไปไกลถึงยุคคาร์บอนิเฟอรัส บ่งชี้ว่าความสามารถในการออกไปในสภาพแสงน้อยนั้นวิวัฒนาการมาก่อนหน้านั้นมาก จากลักษณะเหล่านี้ ตัวอย่าง SAM-PK-K10034 อาจมีการมองเห็นแบบเมโซปิกและไซโอโนซอรัส อาจมีการมองเห็นแบบสโค โทปิกหรือโฟโตปิก[ 50 ]เส้นผ่านศูนย์กลางของวงแหวนสเคลอรัลของViatkogorgon ขนาดเล็ก มีขนาดใหญ่ตามสัดส่วน โดยมีเส้นผ่านศูนย์กลางภายใน 1.5 ซม. (0.6 นิ้ว) และเส้นผ่านศูนย์กลางภายนอก 2.3 ซม. (0.9 นิ้ว) เมื่อเทียบกับเส้นผ่านศูนย์กลาง 2.8 ซม. (1.1 นิ้ว) ของเบ้าตาเอง ซึ่งบ่งชี้ว่ามันออกหากินในเวลากลางคืนเป็นหลัก[ 15 ]ในบรรดากอร์โกนอปเซียนClelandina ในวงศ์ย่อยรูบิดจีน มีวงแหวนสเคลอรัลขนาดเล็กผิดปกติ ซึ่งบ่งชี้ว่ามันมีการมองเห็นแบบโฟโตปิกและออกหากิน ในเวลากลางวันอย่างเคร่งครัด Kammerer เสนอว่าการแบ่งส่วนนิเวศวิทยาในหมู่รูบิดจีน (เนื่องจากมีอนุกรมวิธานที่แตกต่างกันมากถึงเจ็ดชนิดที่อยู่ร่วมกันในพื้นที่) ส่วนหนึ่งเกิดขึ้นในรูปแบบของสายพันธุ์ต่างๆ ที่ออกหากินในเวลาที่แตกต่างกันของวัน แต่วงแหวนสเคลอรัลของClelandina เพียงชนิดเดียว ในวงศ์ย่อยนี้เท่านั้นที่ได้รับการระบุ ทำให้สมมติฐานนี้เป็นเพียงการคาดเดาอย่างมาก[ 6 ]
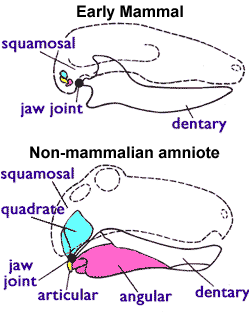
กอร์โกนอปเซียนมีโพรงจมูกค่อนข้างสั้น คล้ายกับเพลิโคซอร์ แต่มีสันยาวจำนวนมากอยู่ด้านหลังรูจมูกภายใน (ซึ่งเชื่อมต่อโพรงจมูกกับลำคอ) เนื่องจากอากาศที่หายใจเข้าไปจะไม่ผ่านสันเหล่านี้ จึงมักตีความว่าสันเหล่านี้เป็นกระดูกเทอร์บิเนตรับกลิ่น และจะทำให้กอร์โกนอปเซียนมีประสาทสัมผัสในการดมกลิ่นที่พัฒนาค่อนข้างสูง[ 51 ] กอร์ โกนอปเซียนมีอวัยวะโวเมอโรนาซัล ("อวัยวะของเจคอบสัน") ซึ่งเป็นส่วนหนึ่งของระบบรับกลิ่นเสริมซึ่งจะอยู่ที่ฐานของผนังกั้นจมูกแตกต่างจากไดไซโนดอนต์และเทอโรเซฟาเลียน ดูเหมือนจะมีท่อเชื่อมต่ออวัยวะกับปาก ซึ่งบ่งชี้ว่ามันทำงานได้ในกอร์โกนอปเซียน[ 52 ]
เทอริโอดอนต์ยุคแรก (รวมถึงกอร์โกนอปเซียน) อาจมีเยื่อแก้วหูซึ่งแตกต่างจากเพลิโคซอร์ในยุคก่อนหน้า โดยสังเกตได้จากการลดลงของการเชื่อมต่อระหว่างกระดูกควอดเรต (ที่ข้อต่อขากรรไกร) และกระดูกเทอริกอยด์ (ที่เพดานปาก ) ทำให้กระดูกควอดเรตสามารถสั่นได้อย่างอิสระในระดับหนึ่ง ซึ่งอาจช่วยให้สามารถตรวจจับเสียงที่เดินทางในอากาศที่มีแอมพลิจูดต่ำกว่า 1 มม. (0.039 นิ้ว) ได้ แต่เยื่อแก้วหูจะต้องได้รับการรองรับด้วยกระดูกอ่อนหรือเอ็นแทนที่จะเป็นกระดูก หากเป็นเช่นนั้นกระดูกโพสต์เดนทารี (ซึ่งในสัตว์เลี้ยงลูกด้วยนมยุคแรกประกอบเป็น กระดูก หูชั้นกลาง ) จะต้องแยกออกจากเดนทารี (กระดูกขากรรไกร) บันทึกฟอสซิลของกอร์โกนอปเซียนดูเหมือนจะบ่งชี้ว่าการเชื่อมต่อระหว่างโพสต์เดนทารีและเดนทารีลดลง อย่างไรก็ตาม เมื่อพิจารณาถึงความเชี่ยวชาญที่จำเป็นสำหรับการกัด สภาพของควอดเรตที่แยกตัวในกอร์โกนอปเซียนอาจอธิบายได้อีกอย่างหนึ่งว่าเป็นสเตร็ปโตสไตลี (ควอดเรตที่หมุนได้) เพื่อขยายช่องปากแทนที่จะอำนวยความสะดวกในการได้ยิน[ 3 ]
การควบคุมอุณหภูมิร่างกาย
มีการเปลี่ยนแปลงทางกายวิภาคครั้งสำคัญเกิดขึ้นระหว่างเพลิโคซอร์ในยุคแรกกับเทอแรปซิด ซึ่งสันนิษฐานว่าเกี่ยวข้องกับการเผาผลาญ ที่เพิ่มขึ้น และการกำเนิดของ ภาวะอุณหภูมิร่างกายคง ที่ (การรักษาระดับอุณหภูมิร่างกายให้สูง) วิวัฒนาการของเพดานปากรองและการแยกปากออกจากโพรงจมูกอาจเพิ่มประสิทธิภาพการระบายอากาศที่เกี่ยวข้องกับ กิจกรรม แอโรบิก ในระดับสูง กอร์โกนอปเซียนไม่มีเพดานปากรองที่เป็นกระดูก แต่เป็นไปได้ว่ามีเพดานปากรองที่เป็นเนื้อเยื่ออ่อน อย่างไรก็ตาม เพดานปากรองอาจช่วยในการกินอาหารปริมาณมากในคราวเดียวมากกว่าการระบายอากาศ การจัดระเบียบโครงกระดูกใหม่ (จากการเดินแบบกางขาไปเป็นการเดินแบบพาราซาจิตัล) สันนิษฐานว่าบ่งชี้ถึงการมีอยู่ของกระบังลมและดังนั้นจึงช่วยเพิ่มการระบายอากาศสำหรับกิจกรรมแอโรบิก แต่ก็อาจเป็นการเพิ่มความเร็วหรือความคล่องตัว ซึ่งไม่จำเป็นต้องเท่ากับกิจกรรมแอโรบิกที่เข้มข้น เช่นเดียวกับในจระเข้ กระดูกคอร์ติคัลแบบแผ่นเส้นใย ซึ่งเทอแรปซิดยุคแรกทั้งหมดมี บ่งชี้ถึงอัตราการเติบโตที่เพิ่มขึ้น แต่สิ่งนี้อาจไม่ได้เชื่อมโยงกับอัตราการเผาผลาญ[ 53 ]
โดยธรรมชาติแล้ว สัตว์เลื้อยคลานขนาดใหญ่ในปัจจุบันจะระบายความร้อนออกจากร่างกายในอัตราที่ช้ากว่าสัตว์เลื้อยคลานขนาดเล็ก และถูกจัดว่าเป็น "สัตว์เลือดอุ่นแบบเฉื่อย" แต่พวกมันยังคงรักษาอุณหภูมิร่างกายไว้ที่ระดับต่ำ 25–30 องศาเซลเซียส (77–86 องศาฟาเรนไฮต์) หากเทอราปซิดต้องการอุณหภูมิร่างกายที่สูงกว่า 35–40 องศาเซลเซียส (95–104 องศาฟาเรนไฮต์) พวกมันจะต้องเป็นสัตว์เลือดอุ่น (สร้างความร้อนในร่างกายเอง) หรือมีการควบคุมการสูญเสียความร้อนได้ดีกว่า (กล่าวคือ มีภาวะเลือดอุ่นที่ดีกว่า) การเดินแบบพาราซาจิตัลอาจช่วยในเรื่องนี้ เนื่องจากจะช่วยให้ส่วนใหญ่ของร่างกายอยู่เหนือพื้นดิน และช่วยให้เลือดไหลเวียนอยู่ในช่องท้องแทนที่จะต้องไหลเวียนผ่านแขนขา ซึ่งทั้งสองอย่างนี้จะช่วยลดการถ่ายเทความร้อนไปยังพื้นดินและรักษาอุณหภูมิแกนกลางของร่างกาย ให้คง ที่ หางที่สั้นลงยังช่วยลดพื้นที่ผิวทั้งหมดของสัตว์ ทำให้การสูญเสียความร้อนลดลงอีกด้วย[ 54 ]ในกลุ่มเทอแรปซิด มีเพียงยูเทอริโอดอนต์ (ไม่ใช่กอร์โกนอปเซียน) เท่านั้นที่มีเทอร์บิเนตจมูก สำหรับการหายใจ ซึ่งช่วยรักษาความชื้นขณะหายใจเอาอากาศเข้าไปเป็นจำนวนมาก และวิวัฒนาการของมันมักเกี่ยวข้องกับการเริ่มต้นของอัตราการบริโภคออกซิเจนแบบ "สัตว์เลี้ยงลูกด้วยนม" และต้นกำเนิดของเอนโดเทอร์มี[ 51 ]
หากกอร์โกนอปเซียนเป็นสัตว์เลือดอุ่นแบบเฉื่อย ก็ไม่ใช่เรื่องที่เป็นไปไม่ได้ที่พวกมันจะมีขน จมูกมักเต็มไปด้วยรูพรุน (รูเล็กๆ ที่มีหลอดเลือด) ซึ่งอาจบ่งชี้ถึงการมีผิวหนังที่หลวม (ตรงข้ามกับเกล็ด) ขนต่อมผิวหนัง ต่างๆ (เช่นต่อมเหงื่อ ) และหนวดอย่างไรก็ตาม สัตว์เลื้อยคลานบางชนิดมีรูปแบบของรูพรุนที่คล้ายกัน ซึ่งเกี่ยวข้องกับการพัฒนาของฟันมากกว่าผิวหนัง[ 55 ]การวิเคราะห์เกี่ยวกับกลไกทางชีวภาพของหูชั้นในพบว่ากอร์โกนอปเซียนน่าจะมีอุณหภูมิร่างกาย 23.7–28.6 °C (74.7–83.5 °F) ซึ่งบ่งชี้ว่ากลุ่มนี้เป็นสัตว์เลือดเย็น[ 56 ]
พยาธิวิทยาโบราณ


ด้านหน้าด้านข้างของกระดูกเรเดียส ซ้าย (กระดูกปลายแขน) ของตัวอย่างกอร์โกนอปเซียน NHCC LB396 แสดงให้เห็นรอยโรคกระดูกรูปวงกลม ซึ่งมีลักษณะเป็น หนาม แหลม ไม่สม่ำเสมอไปจนถึง แนวรัศมีที่ทำจากกระดูกชั้นนอก ล้อมรอบด้วยชั้นกระดูกใต้เยื่อหุ้มกระดูก บาง ๆ ซึ่งเติบโตอย่างรวดเร็วในช่วงฤดูการเจริญเติบโตเพียงฤดูเดียว ลักษณะนี้สอดคล้องกับภาวะเยื่อ หุ้ม กระดูกอักเสบซึ่งน่าจะเกิดจากภาวะเลือด ออกใต้ เยื่อหุ้มกระดูก สภาวะเฉพาะนี้รวมถึงอัตราการเติบโตที่รวดเร็วนั้นชวนให้นึกถึงสัตว์เลี้ยงลูกด้วยนมและไดโนเสาร์มากกว่าจระเข้หรือจิ้งจกมอนิเตอร์ในกลุ่มซินาปซิดยุคแรก พยาธิสภาพอื่น ๆ ที่พบมีเพียงโรคกระดูกอักเสบในกลุ่มเพลิโคซอร์หลายกลุ่ม[ 57 ]
ด้านริมฝีปาก (ด้านแก้ม) ของรากฟันเขี้ยวที่ใช้งานได้ของ RB382 มีรอยโรคมากถึง 8 รอย กระจุกตัวอยู่ตามแนวกึ่งกลางของฟัน ซึ่งมีลักษณะคล้ายฟันขนาดเล็กที่มีเนื้อฟันเนื้อเยื่อฟันและเคลือบฟันบาง ๆรอย โรค เหล่านี้มีรูปร่างเป็นวงกลมโดยประมาณ โดยมีเส้นผ่านศูนย์กลางแตกต่างกันไปตั้งแต่ 0.3–3.9 มม. (0.012–0.154 นิ้ว) แม้ว่าจะมีรูปร่างไม่เป็นวงกลมมากนักบริเวณกึ่งกลางของรากจนกระทั่งผ่านคอฟันลักษณะนี้สอดคล้องกับโรคโอโดนโตมา ในมนุษย์ ซึ่งเป็น เนื้องอกที่เกิดจากฟันชนิดที่พบบ่อยที่สุดซึ่งก่อนหน้านี้มีบันทึกฟอสซิลย้อนหลังไปเพียงไม่กี่ล้านปีเท่านั้น RB382 ที่มีอายุ 255 ล้านปี ถือเป็นกรณีโอโดนโตมาที่เก่าแก่ที่สุดเท่าที่รู้จัก[ 58 ]
จมูกของกอร์โกนอปเซียนตัวเต็มวัย SAM-PK-11490 จากสายพันธุ์กอร์โกนอปเซียนยุคเพอร์เมียนตอนกลางที่ไม่สามารถระบุชนิดได้ มีฟันฝังอยู่จากสัตว์ที่ไม่สามารถระบุชนิดได้ กระดูกมีแคลลัสเกิดขึ้นรอบๆ ฟัน แสดงว่าฟันหายดีแล้วและตัวนั้นรอดชีวิตจากการถูกโจมตี ฟันนั้นอาจมาจากผู้ล่า เช่น ไบอาร์โมซูเคียน เทโรเซฟาเลียน หรือกอร์โกนอปเซียนตัวอื่น หรืออาจเกิดจากการกัดกันเองที่ใบหน้า ซึ่งมักพบในสัตว์นักล่าที่อยู่รวมกันเป็นฝูง เช่น แมวใหญ่หรือจิ้งจกมอนิเตอร์ และมีการเสนอแนะไว้สำหรับสายพันธุ์ที่สูญพันธุ์ไปแล้วหลายสายพันธุ์ เช่น เทโรพอด สัตว์เลื้อยคลานในน้ำ และเสือเขี้ยวคม การกัดกันทางสังคมมีจุดประสงค์เพื่อแสดงอำนาจหรืออำนวยความสะดวกในการผสมพันธุ์ และหากถูกต้อง ก็แสดงว่ากอร์โกนอปเซียนยุคเพอร์เมียนตอนกลางอย่างน้อยบางชนิดเป็นสัตว์กินเนื้อที่อยู่รวมกันเป็นฝูง ฟันนั้นถูกมองข้ามไปในตอนแรก จึงไม่ชัดเจนว่าพยาธิสภาพนี้พบได้บ่อยแค่ไหน[ 59 ]
นิเวศวิทยาบรรพกาล
สภาพแวดล้อมโบราณ
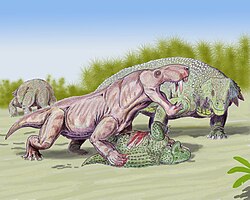
หลังจากการสูญพันธุ์ของไดโนเซฟาเลียนและ (ในแอฟริกาใต้) เทโรเซฟาเลียนพื้นฐานอย่างScylacosauridaeและLycosuchidaeกอร์โกนอปเซียนได้วิวัฒนาการจากรูปแบบขนาดเล็กและหายากไปเป็นนักล่าระดับสูงสุดขนาดใหญ่[ c ]ตลอดช่วงยุคเพอร์เมียนตอนกลางถึงตอนบน ในแอฟริกาใต้ ไดไซโนดอนต์เป็นสัตว์ที่พบได้บ่อยที่สุด ในขณะที่พาเรอาซอร์DeltavjatiaและScutosaurusมีจำนวนมากที่สุดในชั้นหินรัสเซียที่มีกอร์โกนอปเซียน[ 60 ]ในช่วงยุคเพอร์เมียนตอนบน กลุ่มโบฟอร์ตของแอฟริกาใต้เป็นทุ่งหญ้าสเตปป์กึ่งแห้งแล้งที่ มีอากาศหนาวเย็น มีแม่น้ำขนาดใหญ่ตามฤดูกาล ( ชั่วคราว ) และที่ราบน้ำท่วมถึงซึ่งระบายแหล่งน้ำที่อยู่ไกลออกไปทางเหนือสู่ทะเลคารู โดยมีบางครั้งที่เกิดน้ำท่วมฉับพลันหลังจากฝนตกหนักอย่างกะทันหัน[ 61 ] [ 62 ]การกระจายตัวของคาร์บอเนต สอดคล้องกับแหล่งสะสม แคลอรีในปัจจุบันซึ่งก่อตัวขึ้นในสภาพภูมิอากาศที่มีอุณหภูมิเฉลี่ย 16–20 °C (61–68 °F) และปริมาณน้ำฝนตามฤดูกาล 100–500 มม. (3.9–19.7 นิ้ว) [ 62 ]ชั้นหิน Salarevskian Formation ที่มีกอร์โกนอปเซียนในรัสเซียตะวันตกก็อาจถูกสะสมตัวในสภาพแวดล้อมกึ่งแห้งแล้งที่มีปริมาณน้ำฝนตามฤดูกาลสูง และมี พืช ที่ชอบความชื้นและ พืช ที่ชอบเกลือในพื้นที่ชายฝั่ง รวมถึงต้นสนที่ ทนแล้งได้ดีกว่า ในระดับความสูงที่สูงขึ้น[ 63 ]ชั้นหิน Moradi Formation เป็นทะเลทรายแห้งแล้ง ซึ่งส่วนใหญ่ถูกครอบงำโดยสัตว์เลื้อยคลานแคปโทไรนิดMoradisaurusและพาเรอาซอร์Bunostegos [ 20 ] มีต้นสนโวลเซียนและสภาพแวดล้อมถูกเปรียบเทียบกับทะเลทรายนามิบตอน ใน หรือแอ่งทะเลสาบ Eyre [ 64 ]
การสูญพันธุ์
ก่อนเหตุการณ์การสูญพันธุ์ในยุคเพอร์เมียน-ไทรแอสสิก กอร์โกนอปเซียนจำนวนมากเริ่มสูญพันธุ์ในแอฟริกาและยุโรปตะวันออก[ 65 ] [ 66 ] [ 67 ]ในยุโรปตะวันออก กอร์โกนอปเซียนสูญพันธุ์เนื่องจากไม่สามารถปรับตัวให้เข้ากับการเปลี่ยนแปลงของอุณหภูมิและความชื้นได้ หลังจากการสูญพันธุ์ของพวกมัน ช่องว่างของสัตว์นักล่าขนาดใหญ่ก็ถูกครอบครองโดยเทโรเซฟาเลียน ขนาดใหญ่ และ สัตว์ เลื้อยคลานอาร์โคซอริฟอร์มที่เข้ามาแทนที่ช่องว่างของสัตว์ นักล่าขนาดใหญ่ [ 65 ] [ 68 ] รูบิด จีนส่วนใหญ่ที่มีขนาดตัวใหญ่ได้ตายไปในช่วงต้นของเขตการรวมกลุ่มแดปโตเซฟาลัส รู บิดเจีย ซึ่งเป็นรูบิดจีนตัวสุดท้าย ได้สูญพันธุ์ไปในช่วงเหตุการณ์การเปลี่ยนแปลงที่แยกเขตย่อยล่างและบนออกจากกัน โดยถูกแทนที่ด้วยอินอสตอรันเซเวีย [ 66 ] กลุ่มกอร์โกนอปเซียนที่รู้จักกลุ่มสุดท้ายคืออินอสตอรันเซเวียและไซโอโนซอรัสในเขตการ รวมกลุ่ม แดปโตเซฟาลัส ตอนบน [ 67 ]ทั้งสองกลุ่มสิ่งมีชีวิตสูญพันธุ์ไปในช่วง PTEM ซึ่งเกิดจากภูเขาไฟระเบิดที่ก่อตัวเป็นที่ราบไซบีเรีย เป็นหลัก ก๊าซ เรือนกระจกที่พุ่งสูงขึ้นอย่างมากส่งผลให้เกิดภาวะแห้งแล้งอย่างรวดเร็วเนื่องจาก: อุณหภูมิพุ่งสูงขึ้น (สูงถึง 8–10 °C ที่เส้นศูนย์สูตร โดยมีอุณหภูมิเฉลี่ยที่เส้นศูนย์สูตรอยู่ที่ 32–35 °C หรือ 90–95 °F ในช่วงต้นยุคไทรแอสสิก) ฝนกรด (โดยมีค่า pHต่ำถึง 2 หรือ 3 ในช่วงที่มีการระเบิดและ 4 ทั่วโลก และการขาดแคลนป่าไม้ในช่วง 10 ล้านปีแรกของยุคไทรแอสสิก) ไฟป่าที่เกิด ขึ้นบ่อยครั้ง (แม้ว่าจะเกิดขึ้นค่อนข้างบ่อยอยู่แล้วตลอดช่วงยุคเพอร์เมียน) และการทำลายชั้นโอโซน ที่อาจเกิดขึ้น (อาจทำให้ การแผ่รังสี UV เพิ่มขึ้นชั่วคราว ถึง 400% ที่เส้นศูนย์สูตรและ 5000% ที่ขั้วโลก) [ 69 ]ในตอนแรก มีการคิดว่ากลุ่มอนุกรมวิธานขนาดเล็กบางกลุ่ม เช่นCyonosaurusมีชีวิตอยู่จนถึงยุคไทรแอสสิกตอนต้นแต่การวิเคราะห์ในภายหลังพบว่าเป็นความเข้าใจผิด อย่างไรก็ตาม นี่เป็นการพบกอร์โกนอปเซียนที่อายุน้อยที่สุดในแอ่งคารู ซึ่งบ่งชี้ว่ามันมีความทนทานต่อช่วงแรกของการสูญพันธุ์ครั้งใหญ่มากกว่า[ 68 ]ในกลุ่มเทอราปซิด เทอโรเซฟาเลียนขนาดเล็กและอะโนโมดอนต์กินพืชขนาดใหญ่สามารถข้ามพรมแดนเพอร์เมียน-ไทรแอสสิกได้ และมีชีวิตรอดจนถึง ยุคไทรแอสสิกตอน กลางและตอนบน ตามลำดับแต่มีเพียงไซโนดอนต์สายพันธุ์ขนาดเล็กเท่านั้นที่รอดชีวิตมาจนถึงยุคจูราสสิก ซึ่งลูกหลานของพวกมันจะรวมถึงสัตว์เลี้ยงลูกด้วยนมด้วย ช่องว่างในการล่าเหยื่อ ขนาดใหญ่ ที่กอร์โกนอปเซียนทิ้งไว้ในที่สุดก็ถูกเติมเต็มโดยอาร์โคซอร์ (รวมถึงจระเข้และไดโนเสาร์ ) ในช่วงต้นยุคไทรแอสสิก[ 70 ]
ดูเพิ่มเติม
หมายเหตุ
- ^ ce,สม cerebellum ; cnI,เส้นประสาทรับกลิ่น ; cnV +vcm—เส้นประสาทไตรเจมินัลและหลอดเลือดดำ capitis medialis ; cnVI,เส้นประสาท abducens ; cnVII,เส้นประสาทใบหน้า ; cnIX-XI, เส้นประสาท glossopharyngealและ vagoaccessory ; cnXII,เส้นประสาท hypoglossal ; en,เส้นประสาท epiphyseal ; fb,สมองส่วนหน้า ; fcl, flocculus ; ibic, สาขาภายในของหลอดเลือดแดง carotid ภายใน ; lob,หลอดรับกลิ่น ด้านซ้าย ; ob,หลอดรับกลิ่น ; ot, ทางเดินรับกลิ่น ; pg,ต่อมใต้สมอง ; pgll, กลีบข้างของต่อมใต้สมอง ; pf, ส่วนโค้งของพอนส์ ; rob, หลอดรับกลิ่นด้านขวา ; vc,ช่อง vidian ; vc=spa, ช่อง vidian ที่หลอดเลือดแดง sphenopalatineผ่าน vcd, vena capitis dorsalis
- ^ในการจำแนกประเภทล่าสุด ตระกูล Gorgonopsidae โดยทั่วไปจะถูกกำหนดขึ้นเพื่อไม่รวม ตัวแทน พื้นฐานของ Gorgonopsia [ 7 ] : 242–244
- ^การตีพิมพ์คำอธิบายอย่างเป็นทางการของสกุล Phorcysและ ตัวอย่าง ที่ไม่สามารถระบุชนิดได้ ที่คล้ายกันในปี 2022 แสดงให้เห็นว่ากอร์โกน อปเซียนบางชนิดมีขนาดใหญ่ขึ้นแล้วในช่วงยุคเพอร์เมียนตอนกลาง อย่างไรก็ตาม กอร์โกน อปเซียนขนาดใหญ่ เช่น Phorcysและเทอแรปซิด ขนาดใหญ่อื่นๆ อาจออกจากการแบ่งส่วนนิเวศวิทยาให้กับตัวแทนที่มีขนาดเล็กกว่าหลังจากการสูญพันธุ์ในยุคคาปิทาเนียน [ 36 ]
ลิงก์ภายนอก
 สื่อที่เกี่ยวข้องกับGorgonopsiaใน Wikimedia Commons
สื่อที่เกี่ยวข้องกับGorgonopsiaใน Wikimedia Commons ข้อมูลที่เกี่ยวข้องกับGorgonopsiaใน Wikispecies
ข้อมูลที่เกี่ยวข้องกับGorgonopsiaใน Wikispecies
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ กอร์โกนอปเซีย
กอร์โกนอปเซีย (มาจากภาษากรีก กอร์ กอน สัตว์ในตำนาน และ óps 'ลักษณะ') เป็น กลุ่ม สัตว์เลี้ยง ลูกด้วยนม ในกลุ่ม เทอแรปซิด ที่มีเขี้ยวแหลมคม ที่สูญพันธุ์ไปแล้ว จาก ยุคเพอร์เมียน...
คำอธิบาย
กอร์โกนอปซิดในยุคเพอร์เมียนตอนต้นมีขนาดค่อนข้างเล็ก โดยมีความยาวกะโหลก 10–15 ซม. (4–6 นิ้ว) [ 1 ] ในขณะที่บางสกุลในยุคหลังมีขนาดใหญ่โตคล้ายหมี โดยสกุลที่ใหญ่ที่สุดคือ Inostrancevia มีความยาวถึง 3.5 ม. (11 ฟุต) และมีน้ำหนักตัว 300 กก.
กะโหลก
กายวิภาคของกอร์โกนอปเซียนแตกต่างกันน้อยมาก [ 5 ] หลายชนิดมีความแตกต่างกันเพียงเล็กน้อยในสัดส่วน และด้วยเหตุนี้ชนิดที่เล็กกว่าอาจเป็นลูกอ่อนของชนิดที่ใหญ่กว่าได้ ที่น่าสังเกตคือ กระดูก โว เมอร์ ที่ปลายจมูกจะแตกต่างกันไปในแต่ละชนิดในแง่ของระดับการขยายตัว...
ฟัน
เช่นเดียวกับสัตว์เลี้ยงลูกด้วยนมหลายชนิด กอร์โกนอปเซียนเป็น เฮเทอโรดอนต์ โดยมี ฟันหน้า ฟัน เขี้ยว และฟันหลังเขี้ยวที่ชัดเจน ซึ่งมีลักษณะคล้ายคลึง กับ ฟันกราม และ ฟันกราม เล็ก [ 1 ] พวกมันมีฟันหน้าห้าซี่ในขากรรไกรบน (ส่วนใหญ่ ฟันสามซี่แรกมีขนาดเท่ากัน...