อ่าน 29 นาที
โพลีพลอยดี
ภาวะ โพลีพลอยด์ คือ ภาวะที่ เซลล์ ของ สิ่งมีชีวิต มี โครโมโซม คู่ ( โฮโมล็อกัส ) มากกว่าสองคู่สิ่งมีชีวิตส่วนใหญ่ที่มี นิวเคลียส ( ยูคาริโอต ) จะเป็น ดิพลอยด์...
โพลีพลอยดี
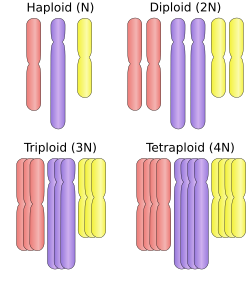
ภาวะ โพลีพลอยด์คือ ภาวะที่เซลล์ของสิ่งมีชีวิต มี โครโมโซม คู่ ( โฮโมล็อกัส ) มากกว่าสองคู่สิ่งมีชีวิตส่วนใหญ่ที่มีนิวเคลียส ( ยูคาริโอต ) จะเป็นดิพลอยด์หมายความว่าพวกมันมีโครโมโซมครบสองชุด ชุดหนึ่งจากพ่อและอีกชุดหนึ่งจากแม่ แต่ละชุดมีจำนวนโครโมโซมเท่ากัน และโครโมโซมจะจับคู่กันเป็นโครโมโซมโฮโมล็อกัส อย่างไรก็ตาม สิ่งมีชีวิตบางชนิดเป็นโพลีพลอยด์ ภาวะโพลีพลอยด์พบได้บ่อยในพืช ยูคาริโอตส่วนใหญ่มีเซลล์ร่างกาย แบบดิพลอยด์ แต่สร้างเซลล์สืบพันธุ์แบบแฮพลอยด์ (ไข่และอสุจิ) โดยกระบวนการไมโอซิสโมโนพลอยด์คือ สิ่งมีชีวิตที่มีโครโมโซมเพียงชุดเดียว และโดยทั่วไปคำนี้มักใช้กับเซลล์หรือสิ่งมีชีวิตที่ปกติเป็นดิพลอยด์เท่านั้น เช่น ตัวผู้ของผึ้งและ แมลงในอันดับ Hymenoptera อื่นๆ เป็นโมโนพลอยด์ แตกต่างจากสัตว์พืชและสาหร่าย หลายเซลล์ มีวงจรชีวิตที่มีสองรุ่นสลับกันไป รุ่นแก มีโท ไฟต์เป็นแฮพลอยด์และสร้างแกมีตโดย การแบ่งเซลล์แบบ ไมโทซิส ส่วนรุ่นสปอโร ไฟต์เป็นดิพลอยด์และสร้างสปอร์โดย การแบ่งเซลล์แบบ ไมโอซิส
โพลีพลอยดีเป็นผลมาจากการจำลองจีโนมทั้งหมดในระหว่างวิวัฒนาการของสปีชีส์ อาจเกิดขึ้นเนื่องจากการแบ่งเซลล์ ที่ผิดปกติ ไม่ว่าจะในระหว่างการแบ่งเซลล์แบบไมโทซิส หรือที่พบได้บ่อยกว่าคือความล้มเหลวในการแยกโครโมโซมในระหว่างการแบ่งเซลล์แบบไมโอซิส หรือจากการปฏิสนธิของไข่โดยอสุจิมากกว่าหนึ่งตัว[ 1 ]นอกจากนี้ยังสามารถชักนำให้เกิดโพลีพลอยดีในพืชและเซลล์เพาะเลี้ยงได้ด้วยสารเคมีบางชนิด สารที่รู้จักกันดีที่สุดคือคอลชิซีน ซึ่งสามารถทำให้โครโมโซมเพิ่มจำนวนเป็นสองเท่าได้ แม้ว่าการใช้สารนี้อาจมีผลกระทบอื่นๆ ที่ไม่ชัดเจนนักก็ตาม โอไรซาลิน ก็จะ ทำให้จำนวนโครโมโซมที่มีอยู่เพิ่มเป็นสองเท่าเช่นกัน
ภาวะโพลีพลอยดีทั้งตัวพบได้ยากในสัตว์เลี้ยงลูกด้วยนมอย่างไรก็ตาม เอนโดโพลีพลอยดี (เซลล์โพลีพลอยด์ในเนื้อเยื่อบางส่วนของสิ่งมีชีวิตที่เป็นดิพลอยด์) เกิดขึ้นในสัตว์เลี้ยงลูกด้วยนมด้วยความถี่สูงในอวัยวะต่างๆ เช่น สมอง ตับ หัวใจ และไขกระดูก[ 2 ]เอนโดโพลีพลอยดียังเกิดขึ้นในเซลล์ร่างกายของสัตว์ อื่นๆ เช่นปลาทอง [ 3 ]ปลาแซลมอนและซาลาแมนเดอร์และในอาณาจักรอื่นๆ ด้วย
โพลีพลอยดีเป็นเรื่องปกติในเฟิร์นและพืช ดอก (ดูHibiscus rosa-sinensis ) รวมถึงทั้ง สาย พันธุ์ ป่าและสายพันธุ์ ที่ปลูก ตัวอย่างเช่นข้าวสาลี หลังจาก การผสมข้ามพันธุ์และการดัดแปลงโดยมนุษย์มาหลายพันปี ก็มีสายพันธุ์ที่เป็นดิพลอยด์ (โครโมโซมสองชุด) เตตราพลอยด์ (โครโมโซมสี่ชุด) ซึ่งมีชื่อเรียกทั่วไปว่า ข้าว สาลีดูรัมหรือข้าวสาลีมาการอนี และเฮกซาพลอยด์ (โครโมโซมหกชุด) ซึ่งมีชื่อเรียกทั่วไปว่าข้าวสาลีขนมปัง พืชทางการเกษตรที่สำคัญหลายชนิดในสกุลBrassicaก็เป็นเตตราพลอยด์เช่นกันอ้อยอาจมีระดับพลอยดีสูงกว่าออกตาพลอยด์[ 4 ]
การ เกิดโพลีพลอยด์อาจเป็นกลไกของ การเกิดสปีชีส์ใหม่ในพื้นที่เดียวกัน เนื่องจากโพลีพลอยด์มักไม่สามารถผสมพันธุ์กับบรรพบุรุษดิพลอยด์ได้ ตัวอย่างเช่น พืชErythranthe peregrina การจัดลำดับดีเอ็นเอได้ยืนยันว่าสปีชีส์นี้มีต้นกำเนิดมาจาก E. × robertsiiซึ่งเป็นลูกผสมไตรพลอยด์ที่เป็นหมันระหว่างE. guttataและE. luteaซึ่งทั้งสองชนิดได้ถูกนำเข้ามาและแพร่กระจายในสหราชอาณาจักร ประชากรใหม่ของE. peregrinaเกิดขึ้นบนแผ่นดินใหญ่ของสกอตแลนด์และหมู่เกาะออร์กนีย์ผ่านการจำลองจีโนมจากประชากรท้องถิ่นของE. × robertsii [ 5 ] เนื่องจากมีการกลายพันธุ์ทางพันธุกรรมที่หายากE. peregrinaจึงไม่เป็นหมัน[ 6 ]
ในทางกลับกัน การเกิดโพลีพลอยด์ยังสามารถเป็นกลไกสำหรับ 'การเกิดสปีชีส์แบบย้อนกลับ' ได้อีกด้วย[ 7 ]ซึ่งทำให้การไหลของยีนเกิดขึ้นได้หลังจากการเกิดโพลีพลอยด์ แม้กระทั่งระหว่างสายพันธุ์ที่ก่อนหน้านี้ไม่มีการไหลของยีนในรูปแบบดิพลอยด์ รายละเอียดนี้ได้รับการอธิบายในระดับจีโนมในArabidopsis arenosaและArabidopsis lyrata [ 8 ] แต่ละสปีชีส์เหล่านี้ประสบกับเหตุการณ์ออโตโพลีพลอยด์อิสระ (โพลีพลอยด์ภายในสปีชีส์ ซึ่งจะกล่าวถึงต่อไป) ซึ่งทำให้เกิดการไหลของยีนระหว่างสปีชีส์ของอัลลีลที่ปรับตัวได้ ในกรณีนี้ทำให้สายพันธุ์โพลีพลอยด์รุ่นเยาว์แต่ละสายพันธุ์มีเสถียรภาพ[ 9 ]การถ่ายทอดยีนแบบปรับตัวได้ที่เกิดขึ้นจากโพลีพลอยด์ดังกล่าวอาจทำให้โพลีพลอยด์ทำหน้าที่เป็น 'ฟองน้ำอัลลีล' ซึ่งพวกมันสะสมความแปรผันทางจีโนมที่ซ่อนเร้นซึ่งอาจถูกนำมาใช้เมื่อเผชิญกับความท้าทายด้านสิ่งแวดล้อมในภายหลัง[ 7 ]
ศัพท์เฉพาะ
ประเภท

ชนิดของ โพลีพลอยด์จะถูกกำหนดชื่อตามจำนวนชุดโครโมโซมในนิวเคลียส โดยใช้ ตัวอักษรxแทนจำนวนโครโมโซมในแต่ละชุด:
- แฮพลอยด์ (หนึ่งชุด; 1 x ) ตัวอย่างเช่นมดไฟยุโรป ตัวผู้
- ดิพลอยด์ (สองชุด; 2 x ) ตัวอย่างเช่นมนุษย์
- ไตรพลอยด์ (สามชุด; 3 x ) เช่นดอกหญ้าฝรั่น ที่เป็นหมัน หรือแตงโมไร้เมล็ดซึ่งพบได้ทั่วไปในไฟลัมทาร์ดิกราดา[ 10 ]
- เตตราพลอยด์ (สี่ชุด; 4 x ) เช่นหนูวิสคาชาที่ราบ ปลาวงศ์ปลาแซลมอน[ 11 ]ฝ้ายGossypium hirsutum [ 12 ]
- พืชมีชุดโครโมโซม 5 ชุด (5 x ) ตัวอย่างเช่น ต้นเบิร์ชเคไน ( Betula kenaica )
- เฮ กซาพลอยด์ (หกชุด; 6 x ) เช่นข้าวสาลี บางสายพันธุ์ [ 13 ]กีวี[ 14 ]
- เฮปตาพลอยด์หรือเซปตาพลอยด์ (เจ็ดชุด; 7 x ) ตัวอย่างเช่นปลาสเตอร์เจียนไซบีเรีย ที่เลี้ยงไว้บางชนิด [ 15 ]
- อ็อกตาพลอยด์หรืออ็อกโทพลอยด์ (แปดชุด; 8x )ตัวอย่างเช่นAcipenser (สกุลปลาสเตอร์เจียน ) และดอกดาเลีย
- เดคาพลอยด์ (สิบชุดโครโมโซม; 10 เท่า ) ตัวอย่างเช่นสตรอว์เบอร์รี บางชนิด
- เฮนเดคาพลอยด์หรืออันเดคาพลอยด์ (สิบเอ็ดชุด; 11 x ) ตัวอย่างเช่นLepidium บางชนิด [ 16 ]และกุหลาบพันธุ์ต่างๆ
- โดเดคาพลอยด์หรือ ดู โอเดคาพลอยด์ (สิบสองชุด; 12 x ) เช่น พืชCelosia argenteaและSpartina anglica [ 17 ]หรือสัตว์สะเทินน้ำสะเทินบกXenopus ruwenzoriensis
- เตตราเตตราคอนตาพลอยด์ (สี่สิบสี่ชุด; 44 x ) เช่นหม่อนดำ[ 18 ]
การจำแนกประเภท
ออโตโพลีพลอยดี
ออโตโพลีพลอยด์ คือ โพลีพลอย ด์ ที่มีชุดโครโมโซมหลายชุดซึ่งได้มาจากกลุ่มสิ่งมีชีวิต เดียว
ตัวอย่างสองตัวอย่างของออโตโพลีพลอยด์ตามธรรมชาติ ได้แก่ พืชแบกรับTolmiea menzisii [ 19 ]และปลาสเตอร์เจียนขาวAcipenser transmontanum [ 20 ] กรณีส่วนใหญ่ของออโตโพลีพลอยด์เกิดจากการรวมตัวของแกมีตที่ไม่ลดจำนวน (2 n ) ซึ่งส่งผลให้เกิดลูกหลานที่เป็นไตรพลอยด์ ( n + 2 n = 3 n ) หรือเตตราพลอยด์ (2 n + 2 n = 4 n ) [ 21 ]ลูกหลานที่เป็นไตรพลอยด์มักจะเป็นหมัน (เช่นในปรากฏการณ์ของบล็อกไตรพลอยด์ ) แต่ในบางกรณีพวกมันอาจผลิตแกมีตที่ไม่ลดจำนวนในสัดส่วนสูงและช่วยในการสร้างเตตราพลอยด์ เส้นทางสู่เตตราพลอยด์นี้เรียกว่าสะพานไตรพลอยด์ [ 21 ] ไตรพลอยด์อาจคงอยู่ได้ผ่านการสืบพันธุ์แบบไม่อาศัยเพศในความเป็นจริง ออโตไตรพลอยด์ที่เสถียรในพืชมักเกี่ยวข้องกับระบบการผสมพันธุ์แบบอะโพมิกติก[ 22 ] ในระบบเกษตรกรรม ออ โตไตรพลอยดีสามารถทำให้ไม่มีเมล็ดได้ เช่นแตงโมและกล้วย[ 23 ] ไตรพลอยดียังถูกนำมาใช้ในการเลี้ยงปลาแซลมอนและปลาเทราต์เพื่อทำให้เป็นหมัน[ 24 ] [ 25 ]
ออโตโพลีพลอยด์เกิดขึ้นได้ยากจากการเพิ่มจำนวนจีโนมโซมาติกโดยธรรมชาติ ซึ่งพบได้ในการกลายพันธุ์ของตาแอปเปิล ( Malus domesticus ) [ 26 ] นี่เป็นเส้นทางที่พบบ่อยที่สุดของการเกิดโพลีพลอยด์ที่เหนี่ยวนำขึ้นโดยเทียม โดยใช้วิธีการต่างๆ เช่นการหลอมรวมโปรโตพลาสต์หรือการรักษาด้วยคอลชิซีนโอไรซาลินหรือสารยับยั้งไมโทซิสเพื่อขัดขวาง การแบ่ง ไมโทซิส ตามปกติ ซึ่งส่งผลให้เกิดเซลล์โพลีพลอยด์ กระบวนการนี้มีประโยชน์ในการปรับปรุงพันธุ์พืช โดยเฉพาะอย่างยิ่งเมื่อพยายามนำเชื้อพันธุ์ข้ามระดับพลอยด์[ 27 ]
ออโตโพลีพลอยด์มีชุด โครโมโซมที่เป็นโฮโมล็อกอย่างน้อยสามชุดซึ่งอาจนำไปสู่การจับคู่แบบมัลติวาเลนต์ในอัตราสูงระหว่างไมโอซิส (โดยเฉพาะในออโตโพลีพลอยด์ที่เพิ่งก่อตัวขึ้น หรือที่รู้จักกันในชื่อนีโอโพลีพลอยด์) และการลดลงของความสามารถในการสืบพันธุ์ที่เกี่ยวข้องเนื่องจากการผลิตแกเมตแอนยูพลอยด์[ 28 ]การคัดเลือกตามธรรมชาติหรือการคัดเลือกโดยมนุษย์เพื่อความสามารถในการสืบพันธุ์สามารถทำให้ไมโอซิสในออโตโพลีพลอยด์มีเสถียรภาพได้อย่างรวดเร็วโดยการฟื้นฟูการจับคู่แบบไบวาเลนต์ระหว่างไมโอซิส วิวัฒนาการปรับตัวอย่างรวดเร็วของกลไกไมโอซิส ส่งผลให้ระดับของมัลติวาเลนต์ลดลง (และด้วยเหตุนี้ไมโอซิสของออโตโพลีพลอยด์จึงมีเสถียรภาพ) ได้รับการบันทึกไว้ในArabidopsis arenosa [ 29 ]และArabidopsis lyrata [ 8 ]โดยมีอัลลีลปรับตัวเฉพาะของสายพันธุ์เหล่านี้ที่ใช้ร่วมกันระหว่างโพลีพลอยด์ที่วิวัฒนาการแล้วเท่านั้น[ 8 ] [ 30 ]
ความเหมือนกันในระดับสูงระหว่างโครโมโซมที่ซ้ำกันทำให้ออโตโพลีพลอยด์แสดง การถ่ายทอด ทางพันธุกรรมแบบโพลีโซมิก [ 31 ] ลักษณะนี้มักใช้เป็นเกณฑ์การวินิจฉัยเพื่อแยกแยะออโตโพลีพลอยด์ออกจากอัลโลโพลีพลอยด์ ซึ่งโดยทั่วไปจะแสดงการถ่ายทอดทางพันธุกรรมแบบไดโซมิกหลังจากที่พวกมันผ่านพ้นระยะนีโอโพลีพลอยด์ไปแล้ว[ 32 ]ในขณะที่สปีชีส์โพลีพลอยด์ส่วนใหญ่มีลักษณะเฉพาะอย่างชัดเจนว่าเป็นออโตโพลีพลอยด์หรืออัลโลโพลีพลอยด์ หมวดหมู่เหล่านี้แสดงถึงปลายสุดของสเปกตรัมของการเบี่ยงเบนระหว่างซับจีโนมของพ่อแม่ โพลีพลอยด์ที่อยู่ระหว่างสองขั้วนี้ ซึ่งมักเรียกว่าอัลโลโพลีพลอยด์แบบแบ่งส่วน อาจแสดงระดับการถ่ายทอดทางพันธุกรรมแบบโพลีโซมิกในระดับกลางที่แตกต่างกันไปตามโลคัส[ 33 ] [ 34 ]
ประมาณครึ่งหนึ่งของโพลีพลอยด์ทั้งหมดเชื่อว่าเป็นผลมาจากออโตโพลีพลอยดี[ 35 ] [ 36 ]แม้ว่าจะมีหลายปัจจัยที่ทำให้การประมาณสัดส่วนนี้ทำได้ยาก[ 37 ]
อัลโลโพลีพลอยดี
อัลโลโพลีพลอยด์ แอมฟิโพลีพลอยด์ หรือเฮเทอโรโพลีพลอยด์คือพืชโพลีพลอยด์ ที่มีโครโมโซมที่ได้มาจากสิ่งมีชีวิตสองกลุ่มหรือมากกว่านั้นที่แยกสายวิวัฒนาการออก จากกัน
เช่นเดียวกับในออโตโพลีพลอยดี กระบวนการนี้เกิดขึ้นเป็นหลักผ่านการรวมตัวของแกมีตที่ไม่ลดจำนวน (2n )ซึ่งสามารถเกิดขึ้นก่อนหรือหลังการผสมพันธุ์ในกรณีแรก แกมีตที่ไม่ลดจำนวนจากแต่ละแท็กซอนดิพลอยด์ หรือแกมีตที่ลดจำนวนจากแท็กซอนออโตเตตราพลอยด์สองแท็กซอน จะรวมกันเพื่อสร้างลูกหลานอัลโลโพลีพลอยด์ ในกรณีหลัง ลูกผสมF1 ดิ พลอยด์ หนึ่งตัวหรือมากกว่านั้น จะสร้างแกมีตที่ไม่ลดจำนวนซึ่งรวมกันเพื่อสร้างลูกหลานอัลโลโพลีพลอยด์ [38] การผสมพันธุ์ตามด้วยการจำลองจีโนมอาจเป็นเส้นทางที่พบได้บ่อยกว่าในการเกิดอัลโลโพลีพลอยดีเนื่องจากลูกผสม F1 ระหว่างแท็กซอนมักมีอัตราการสร้างแกมีตที่ไม่ลดจำนวนค่อนข้างสูง ความแตกต่างระหว่างจีโนมของแท็กซอนทั้งสองส่งผลให้เกิดการจับคู่ที่ผิดปกติระหว่าง โครโมโซม โฮโมโลกัสหรือการไม่แยกตัวระหว่างไมโอซิส[ 38 ]ในกรณีนี้ อัลโลโพลีพลอยดีสามารถฟื้นฟู การจับคู่ไมโอซิส แบบไบวาเลน ต์ปกติได้จริง ๆ โดยการให้โครโมโซมโฮโมโลกัสแต่ละตัวมีโฮโมล็อกของตัวเอง หากความแตกต่างระหว่างโครโมโซมโฮโมโลกัสเท่ากันในสองซับจีโนม สิ่งนี้ในทางทฤษฎีสามารถส่งผลให้เกิดการฟื้นฟูการจับคู่แบบไบวาเลนต์และการถ่ายทอดทางพันธุกรรมแบบดิโซมิกอย่างรวดเร็วหลังจากการเกิดอัลโลโพลีพลอยดี อย่างไรก็ตาม การจับคู่แบบมัลติวาเลนต์เป็นเรื่องปกติในอัลโลโพลีพลอยด์ที่เกิดขึ้นใหม่จำนวนมาก ดังนั้นจึงเป็นไปได้ว่าการทำให้ไมโอซิสมีเสถียรภาพส่วนใหญ่เกิดขึ้นอย่างค่อยเป็นค่อยไปผ่านการคัดเลือก[ 28 ] [ 32 ]
เนื่องจากการจับคู่ระหว่างโครโมโซมโฮโมโลจัสเกิดขึ้นได้ยากในอัลโลโพลีพลอยด์ที่เกิดขึ้นแล้ว พวกมันอาจได้รับประโยชน์จากเฮเทอโรไซโกซิตี คงที่ ของอัลลีลโฮโมโลจัส[ 39 ]ในบางกรณี เฮเทอโรไซโกซิตีดังกล่าวอาจมี ผล เฮเทอโรติกที่ เป็นประโยชน์ ไม่ว่าจะเป็นในแง่ของความเหมาะสมในบริบททางธรรมชาติหรือลักษณะที่พึงประสงค์ในบริบททางการเกษตร สิ่งนี้อาจอธิบายได้บางส่วนถึงความแพร่หลายของอัลโลโพลีพลอยดีใน หมู่พืชผล ข้าว สาลีขนมปังและทริติเคิลเป็นตัวอย่างของอัลโลโพลีพลอยด์ที่มีชุดโครโมโซมหกชุด ฝ้าย ถั่วลิสง และควินัวเป็นอัลโลเทตราพลอยด์ที่มีต้นกำเนิดหลายแหล่ง ในพืชตระกูล Brassicaceae สามเหลี่ยม U อธิบายความสัมพันธ์ระหว่าง Brassica ดิพลอยด์ทั่วไปสามชนิด ( B. oleracea , B. rapa และB. nigra ) และอัลโลเทตราพลอยด์สามชนิด ( B. napus , B. juncea และB. carinata )ที่ได้มาจากการผสมข้ามพันธุ์ระหว่างสายพันธุ์ดิพลอยด์ ความสัมพันธ์ที่คล้ายคลึงกันนี้มีอยู่ระหว่าง Tragopogon สามชนิดที่เป็นดิพลอยด์(T. dubius, T. pratensis และT. porrifolius )และTragopogonสองชนิดที่เป็นอัลโลเทตราพลอยด์ ( T. mirusและT. miscellus ) [ 40 ] รูปแบบที่ซับซ้อนของการวิวัฒนาการของอั ลโลโพลีพลอยด์ยังพบได้ในสัตว์ เช่น ในสกุลกบXenopus [ 41 ]
แอนยูพลอยด์
สิ่งมีชีวิตที่มีโครโมโซมหรือส่วนของโครโมโซมบางส่วนมีจำนวนน้อยกว่าหรือมากกว่าปกติเรียกว่าแอนยูพลอยด์ (มาจากคำภาษากรีกที่มีความหมายว่า "ไม่", "ดี" และ "พับ") แอนยูพลอยดีหมายถึงการเปลี่ยนแปลงจำนวนในส่วนหนึ่งของชุดโครโมโซม ในขณะที่โพลีพลอยดีหมายถึงการเปลี่ยนแปลงจำนวนในชุดโครโมโซมทั้งหมด[ 42 ]
เอนโดโพลีพลอยดี
โพลีพลอยดีเกิดขึ้นในเนื้อเยื่อบางส่วนของสิ่งมีชีวิตที่ปกติเป็นดิพลอยด์ เช่นเนื้อเยื่อตับ ของมนุษย์และ เนื้อเยื่อเอนโดสเปิร์ม ของพืช [ 43 ] [ 44 ]ซึ่งเรียกว่าเอนโดโพลี พลอยดี สปีชี ส์ที่มีเซลล์ที่ไม่มีนิวเคลียส นั่นคือโปรคาริโอตอาจเป็นโพลีพลอยด์ได้ ดังเช่นที่พบในแบคทีเรีย ขนาดใหญ่ Epulopiscium fishelsoni [ 45 ] ดังนั้นพลอยดีจึงถูกกำหนดโดยสัมพันธ์กับเซลล์
โมโนพลอยด์
โมโนพลอยด์คือสิ่งมีชีวิตที่มีโครโมโซมเพียงชุดเดียว และโดยทั่วไปคำนี้มักใช้กับเซลล์หรือสิ่งมีชีวิตที่ปกติเป็นดิพลอยด์เท่านั้น คำที่ใช้เรียกสิ่งมีชีวิตประเภทนี้โดยทั่วไปคือแฮพลอยด์
เงื่อนไขเชิงเวลา
นีโอโพลีพลอยดี
พืชที่มีชุดโครโมโซมหลายชุดซึ่งเพิ่งเกิดขึ้นใหม่
เมโซโพลีพลอยดี
นั่นคือโพลีพลอยด์ที่เกิดขึ้นในประวัติศาสตร์ช่วงหลังๆ ไม่ใหม่เท่ากับนีโอโพลีพลอยด์ และไม่เก่าเท่ากับพาเลโอโพลีพลอยด์ มันเป็นโพลีพลอยด์ที่มีอายุกลางๆ มักหมายถึงการเพิ่มจำนวนจีโนมทั้งหมดตามด้วยระดับไดพลอยด์ในระดับกลางๆ
โพลีพลอยดีโบราณ

การจำลองจีโนมโบราณน่าจะเกิดขึ้นในประวัติศาสตร์วิวัฒนาการของสิ่งมีชีวิตทั้งหมด เหตุการณ์การจำลองที่เกิดขึ้นเมื่อนานมาแล้วในประวัติศาสตร์ของสายพันธุ์วิวัฒนาการ ต่างๆ อาจตรวจจับได้ยากเนื่องจากการเกิดดิพลอยด์ ในภายหลัง (เช่น โพลีพลอยด์เริ่มมีพฤติกรรมทางไซโตเจเนติกส์เหมือนดิพลอยด์เมื่อเวลาผ่านไป) เนื่องจากการกลายพันธุ์และการแปลยีนค่อยๆ ทำให้โครโมโซมแต่ละคู่มีสำเนาหนึ่งที่ไม่เหมือนกับสำเนาอื่นๆ เมื่อเวลาผ่านไป เป็นเรื่องปกติที่สำเนาของยีนที่จำลองจะสะสมการกลายพันธุ์และกลายเป็นยีนเทียมที่ไม่ทำงาน[ 46 ]
ในหลายกรณี เหตุการณ์เหล่านี้สามารถอนุมานได้จากการเปรียบเทียบจีโนมที่เรียงลำดับ เท่านั้น ตัวอย่างของการจำลองจีโนมโบราณที่ไม่คาดคิดแต่ได้รับการยืนยันเมื่อเร็ว ๆ นี้ ได้แก่ยีสต์ขนมปัง ( Saccharomyces cerevisiae ), ผักกาดมัสตาร์ด/ผักกาดหอม ( Arabidopsis thaliana ), ข้าว ( Oryza sativa ) และการจำลองจีโนมทั้งหมดสองรอบ (สมมติฐาน2R ) ในบรรพบุรุษวิวัฒนาการ ยุคแรก ของสัตว์มีกระดูกสันหลัง (ซึ่งรวมถึง สายพันธุ์ มนุษย์ ) และอีกกรณีหนึ่งใกล้กับจุดกำเนิดของปลาเทเลออสท์ [ 47 ] พืชดอก ( Angiosperms ) มีภาวะโพลีพลอยดีในบรรพบุรุษของพวกมัน ยูคาริโอตทั้งหมดน่าจะเคยประสบกับเหตุการณ์โพลีพลอยดีในช่วงใดช่วงหนึ่งในประวัติศาสตร์วิวัฒนาการของพวกมัน
คำศัพท์อื่นๆ ที่คล้ายกัน
แคริโอไทป์
คาริโอไทป์คือชุดโครโมโซมลักษณะเฉพาะของสิ่งมี ชีวิต ยูคาริโอต[ 48 ] [ 49 ]การเตรียมและการศึกษาคาริโอไทป์เป็นส่วนหนึ่งของเซลล์วิทยาและโดยเฉพาะอย่างยิ่งเซลล์พันธุศาสตร์
แม้ว่าการจำลองและการถอดรหัสดีเอ็นเอในยูคาริโอต จะมีความเป็นมาตรฐานสูง แต่ก็ไม่สามารถกล่าวเช่นเดียวกันได้สำหรับคาริโอไทป์ของพวกมัน ซึ่งมีความแปรผันสูงระหว่างสปีชีส์ ทั้งในจำนวนโครโมโซมและในโครงสร้างโดยละเอียด แม้ว่าจะสร้างขึ้นจากโมเลกุลขนาดใหญ่ชนิดเดียวกันก็ตาม ในบางกรณี ยังมีความแปรผันอย่างมีนัยสำคัญแม้กระทั่งภายในสปีชีส์เดียวกัน ความแปรผันนี้เป็นพื้นฐานสำหรับการศึกษาหลายด้านในสิ่งที่อาจเรียกว่าเซลล์วิทยาเชิงวิวัฒนาการ
โครโมโซมโฮโมโลกัส
โครโมโซมโฮโมโลกัสคือโครโมโซมที่เกิดขึ้นจากการผสมข้ามสายพันธุ์และการเกิดอัลโลโพลีพลอยด์ซึ่งมีความสัมพันธ์แบบโฮโมโลกัสอย่างสมบูรณ์ในสายพันธุ์บรรพบุรุษ ตัวอย่างเช่นข้าวสาลีดูรัมเป็นผลมาจากการผสมข้ามสายพันธุ์ของพืชตระกูลหญ้าสองชนิดที่ มีชุดโครโมโซมสองชุด คือ Triticum urartuและ Aegilops speltoides บรรพบุรุษที่มีชุดโครโมโซมสองชุดนี้มีโครโมโซมสองชุด ชุดละ 7 โครโมโซม ซึ่งมีขนาดและยีนที่คล้ายคลึงกัน ข้าวสาลีดูรัมมีจีโนมแบบลูกผสมที่มีโครโมโซมสองชุดที่ได้มาจากTriticum urartuและโครโมโซมสองชุดที่ได้มาจากAegilops speltoidesโครโมโซมแต่ละคู่ที่ได้มาจาก พ่อแม่ Triticum urartuจะเป็นโฮโมโลกัสกับโครโมโซมคู่ตรงข้ามที่ได้มาจาก พ่อแม่ Aegilops speltoidesแม้ว่าโครโมโซมแต่ละคู่เองก็เป็นโฮโมโลกัสเช่นกัน
ตัวอย่าง
มนุษย์

ภาวะโพลีพลอยดีที่แท้จริงเกิดขึ้นได้ยากในมนุษย์ แม้ว่าเซลล์โพลีพลอยด์จะพบได้ใน เนื้อเยื่อ ที่มีการแยกแยะ สูง เช่นเนื้อเยื่อตับกล้ามเนื้อหัวใจ รก และไขกระดูก[ 50 ] [ 51 ]ภาวะแอนยูพลอยดีพบได้บ่อยกว่า
ภาวะโพลีพลอยดีเกิดขึ้นในมนุษย์ในรูปแบบของไตรพลอยดีซึ่งมีโครโมโซม 69 โครโมโซม (บางครั้งเรียกว่า 69, XXX) และเตตราพลอยดี ซึ่งมีโครโมโซม 92 โครโมโซม (บางครั้งเรียกว่า 92, XXXX) ไตรพลอยดี ซึ่งมักเกิดจากภาวะปฏิสนธิ หลายตัว เกิดขึ้นในประมาณ 2–3% ของการตั้งครรภ์ทั้งหมดของมนุษย์ และประมาณ 15% ของการแท้งบุตร การตั้งครรภ์ไตร พลอยดีส่วนใหญ่จบลงด้วยการ แท้งบุตร ผู้ที่รอดชีวิตจนครบกำหนดคลอดมักจะเสียชีวิตไม่นานหลังคลอด ในบางกรณี การรอดชีวิตหลังคลอดอาจยืดออกไปได้หากมี ภาวะมิกโซพลอยดี ซึ่งมีทั้ง เซลล์ ดิพลอยด์ และไตรพลอยด์ อยู่ร่วมกันมีรายงานหนึ่งกรณีของเด็กที่รอดชีวิตจนถึงอายุเจ็ดเดือนด้วยกลุ่มอาการไตรพลอยดีแบบสมบูรณ์ เขาไม่สามารถแสดงพัฒนาการทางจิตใจหรือร่างกายตามปกติในทารกแรกเกิด และเสียชีวิตจาก การติดเชื้อ Pneumocystis cariniiซึ่งบ่งชี้ถึงระบบภูมิคุ้มกันที่อ่อนแอ[ 52 ]
ภาวะไตรพลอยดีอาจเป็นผลมาจากภาวะไดจีนี (ชุดแฮพลอยด์ส่วนเกินมาจากมารดา) หรือภาวะไดแอนดรี (ชุดแฮพลอยด์ส่วนเกินมาจากบิดา) ภาวะไดแอนดรีส่วนใหญ่เกิดจากการจำลองชุดแฮพลอยด์ของบิดาจากอสุจิตัวเดียว แต่ก็อาจเป็นผลมาจากการปฏิสนธิของไข่ โดยอสุจิสองตัวได้เช่นกัน [ 53 ]ภาวะไดจีนีมักเกิดจากความล้มเหลวของการแบ่งไมโอซิสหนึ่งครั้งในระหว่างการสร้างไข่ ทำให้เกิดเซลล์ไข่ แบบดิพลอยด์ หรือความล้มเหลวในการขับโพ ลาร์ บอดี หนึ่งตัวออก จากเซลล์ไข่ ภาวะ ไดแอนดรีดูเหมือนจะพบได้มากในกรณี แท้งบุตรในระยะแรกในขณะที่ภาวะไดจีนีพบได้มากในไซโกตไตรพลอยด์ที่รอดชีวิตจนถึงระยะทารกในครรภ์[ 54 ]อย่างไรก็ตาม ในกรณีแท้งบุตรในระยะแรก ภาวะไดจีนีก็พบได้บ่อยกว่าในกรณีที่อายุครรภ์น้อยกว่า8 เดือน+อายุครรภ์ 1/2 สัปดาห์ หรือในกรณีที่มีตัวอ่อนอยู่ นอกจากนี้ยังมีฟีโนไทป์ ที่แตกต่างกันสองแบบ ในและทารก ในครรภ์ไตรพลอย ด์ซึ่งขึ้นอยู่กับที่มาของ ชุด แฮพลอยด์ ส่วนเกิน ในกรณีไดไจนี มักจะมี ทารกในครรภ์ที่เจริญเติบโตไม่ดี มีรูปร่างไม่สมมาตรมีภาวะต่อมหมวกไตฝ่อ อย่างเห็นได้ชัด และมีรกขนาดเล็กมาก[ 55 ]ในกรณี ไดแอนดรี จะเกิดครรภ์ไข่ปลาบางส่วน [ 53 ]ผลกระทบจากต้นกำเนิดของพ่อแม่เหล่านี้สะท้อนให้เห็นถึงผลกระทบของการ ประทับตราทางพันธุกรรม
ภาวะเทตราพลอยดีแบบสมบูรณ์นั้นวินิจฉัยได้ยากกว่าภาวะไตรพลอยดี แต่พบได้ใน 1-2% ของการแท้งบุตรในระยะเริ่มต้น อย่างไรก็ตาม เซลล์เทตราพลอยดีบางส่วนมักพบได้ในการวิเคราะห์โครโมโซมในการวินิจฉัยก่อนคลอดและโดยทั่วไปถือว่า 'ไม่เป็นอันตราย' ยังไม่ชัดเจนว่าเซลล์เทตราพลอยดีเหล่านี้มีแนวโน้มที่จะเกิดขึ้นระหว่าง การเพาะเลี้ยงเซลล์ ในหลอดทดลองหรือว่ามีอยู่ในเซลล์รกในร่างกาย ด้วยหรือไม่ อย่างไรก็ตาม มีรายงานทางคลินิกน้อยมากเกี่ยวกับการวินิจฉัยทารกในครรภ์/ทารกแรกเกิดที่มีภาวะโมเสกเทตราพลอยดี
ภาวะมิกโซพลอยดี (Mixoploidy)พบได้ค่อนข้างบ่อยในตัวอ่อนมนุษย์ก่อนการฝังตัว ซึ่งรวมถึงประชากรเซลล์ผสมระหว่างแฮพลอยด์/ดิพลอยด์ และดิพลอยด์/เตตราพลอยด์ ยังไม่เป็นที่ทราบแน่ชัดว่าตัวอ่อนเหล่านี้ไม่สามารถฝังตัวได้และจึงตรวจพบได้ยากในครรภ์ที่ดำเนินต่อไป หรือเป็นเพียงกระบวนการคัดเลือกที่เอื้อต่อเซลล์ดิพลอยด์
ปลา
ภาวะโพลีพลอยด์ตามธรรมชาติค่อนข้างพบได้ทั่วไปในปลา โดยได้รับการยืนยันแล้วในAcipenseriformes ( ปลาสเตอร์เจียนและปลาพาย ), Salmoniformes ( ปลาแซลมอนปลาเทราต์และญาติ) และCatostomidae (ปลาดูดอเมริกาเหนือ) สายพันธุ์ปลาโพลีพลอยด์ที่มีจำนวนชนิดมากที่สุดคือสมาชิกของCyprinidaeซึ่งประกอบด้วยปลาโพลีพลอยด์ประมาณ 400 ชนิด รวมทั้งสกุลโพลีพลอยด์โดยเฉพาะอีก 30 สกุล[ 56 ]
เหตุการณ์โพลีพลอยดีเกิดขึ้นภายในสายพันธุ์ต้นกำเนิดของปลาเทเลออสท์[ 47 ]
สัตว์อื่นๆ
ตัวอย่างในสัตว์มักพบได้บ่อยในสัตว์ไม่มีกระดูกสันหลัง[ 57 ]เช่นหนอนแบนปลิงและกุ้งน้ำเค็ม โพลีพลอยดียังพบได้ทั่วไปในสัตว์สะเทินน้ำสะเทินบก ตัวอย่างเช่น สกุลXenopus ซึ่งมีความสำคัญทางการแพทย์ มีหลายชนิดที่มีโครโมโซมมากถึง 12 ชุด (โดเดคาพลอยด์) [ 58 ]กิ้งก่าโพลีพลอยด์ก็พบได้ทั่วไปเช่นกัน ส่วนใหญ่เป็นหมันและสืบพันธุ์โดยการสืบพันธุ์แบบไม่อาศัยเพศบางชนิด เช่นLiolaemus chiliensisยังคงสืบพันธุ์แบบอาศัยเพศซาลาแมนเดอร์ ตุ่นโพลีพลอย ด์ (ส่วนใหญ่เป็นไตรพลอยด์) เป็นเพศเมียทั้งหมดและสืบพันธุ์โดยการขโมยสเปิร์ม[ 59 ] โดย "ขโมย" สเปิร์มจากตัวผู้ดิพลอยด์ของสายพันธุ์ที่เกี่ยวข้องเพื่อกระตุ้นการพัฒนาของไข่ แต่ไม่ได้รวมดีเอ็นเอของตัวผู้เข้าไปในลูกหลาน
แม้ว่าเนื้อเยื่อบางส่วนของสัตว์เลี้ยงลูกด้วยนม เช่นเซลล์ตับพาเรน ไคมัล จะเป็นโพลีพลอยด์ [ 60 ] [ 61 ]แต่ก็พบสัตว์เลี้ยงลูก ด้วยนม โพลีพลอยด์ได้น้อยมาก ซึ่งส่วน ใหญ่มักส่งผลให้ เสียชีวิตก่อนคลอด หนู ในวงศ์ Octodontidae ที่อาศัยอยู่ใน ทะเลทรายอันแห้งแล้งของอาร์เจนตินาซึ่งรู้จักกันในชื่อหนูวิสคาชาแห่งที่ราบ ( Tympanoctomys barrerae ) ได้รับการรายงานว่าเป็นข้อยกเว้นของ 'กฎ' นี้[ 62 ]อย่างไรก็ตาม การวิเคราะห์อย่างละเอียดโดยใช้สีโครโมโซมแสดงให้เห็นว่าT. barrerae มีโครโมโซมแต่ละคู่เพียงสองชุด ไม่ใช่สี่ชุดตามที่คาดไว้หากมันเป็นเตตราพลอยด์อย่างแท้จริง[ 63 ]หนูชนิดนี้ไม่ใช่หนูแต่เป็นญาติกับหนูตะเภาและชินชิลลา จำนวน ดิพลอยด์ "ใหม่" (2n ) ของมัน คือ 102 ดังนั้นเซลล์ของมันจึงมีขนาดใหญ่เป็นสองเท่าของขนาดปกติ ญาติที่ใกล้ชิดที่สุดของมันคือOctomys mimaxซึ่งเป็น หนูวิสคาชาแห่งเทือก แอนดีสในวงศ์เดียวกัน โดยมีจำนวนโครโมโซม 2n = 56 ดังนั้นจึงสันนิษฐานได้ว่า บรรพบุรุษที่มีลักษณะคล้าย Octomysได้ให้กำเนิดลูกหลานที่มีโครโมโซมสี่ชุด (กล่าวคือ2n = 4x = 112) ซึ่งด้วยจำนวนโครโมโซมที่เพิ่มขึ้นเป็นสองเท่า ทำให้ลูกหลานเหล่านี้แยกตัวออกจากพ่อแม่ในด้านการสืบพันธุ์
Har Swarup (1956) ได้ชักนำให้เกิดโพลีพลอยดีในปลาโดยใช้การบำบัดด้วยความเย็นจัดกับไข่ในช่วงเวลาใกล้เคียงกับการปฏิสนธิ ซึ่งทำให้เกิดเอ็มบริโอไตรพลอยด์ที่เจริญเติบโตได้สำเร็จ[ 64 ] [ 65 ]นอกจากนี้ยังพบว่าการช็อกด้วยความเย็นหรือความร้อนส่งผลให้เกิดเซลล์สืบพันธุ์ของสัตว์ครึ่งบกครึ่งน้ำที่ไม่ลดจำนวนลง แม้ว่าสิ่งนี้จะเกิดขึ้นในไข่มากกว่าในอสุจิก็ตาม[ 66 ] John Gurdon (1958) ได้ปลูกถ่ายนิวเคลียสที่สมบูรณ์จากเซลล์ร่างกายเพื่อสร้างไข่ดิพลอยด์ในกบXenopus (ซึ่งเป็นการต่อยอดจากงานของ Briggs และ King ในปี 1952) ที่สามารถพัฒนาไปจนถึงระยะลูกอ๊อดได้[ 67 ]นักวิทยาศาสตร์ชาวอังกฤษJBS Haldaneยกย่องงานนี้สำหรับศักยภาพในการประยุกต์ใช้ทางการแพทย์ และในการอธิบายผลลัพธ์ เขาเป็นหนึ่งในคนแรกๆ ที่ใช้คำว่า " โคลน " ในการอ้างอิงถึงสัตว์ ต่อมาชินยะ ยามานากะได้แสดงให้เห็นว่าเซลล์ที่โตเต็มวัยสามารถถูกตั้งโปรแกรมใหม่ให้กลายเป็นเซลล์ที่มีศักยภาพหลายอย่างได้ ซึ่งเป็นการขยายความเป็นไปได้ไปสู่เซลล์ที่ไม่ใช่เซลล์ต้นกำเนิด กอร์ดอนและยามานากะได้รับรางวัลโนเบลร่วมกันในปี 2012 จากผลงานนี้[ 67 ]
พืช
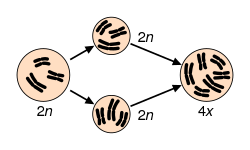
ภาวะโพลีพลอยดีพบได้บ่อยในพืช โดยบางการประมาณการระบุว่า 30–80% ของสายพันธุ์พืชที่ยังมีชีวิตอยู่เป็นโพลีพลอยดี และสายพันธุ์จำนวนมากแสดงหลักฐานของโพลีพลอยดีโบราณ ( พาลีโอโพลีพลอยดี ) ในจีโนมของพวกมัน[ 68 ] [ 69 ] [ 70 ] [ 71 ]การระเบิดครั้งใหญ่ของ ความหลากหลายของสายพันธุ์พืช ดอกดูเหมือนจะเกิดขึ้นพร้อมกับช่วงเวลาของการจำลองจีโนมโบราณที่หลายสายพันธุ์มีร่วมกัน[ 72 ]มีการยืนยันแล้วว่า 15% ของ เหตุการณ์ การเกิดสปี ชีส์ใหม่ของพืชดอกและ 31% ของเฟิร์น เกิดขึ้นพร้อมกับการเพิ่มขึ้นของพลอยดี[ 73 ]
พืชโพลีพลอยด์สามารถเกิดขึ้นเองตามธรรมชาติได้ด้วยกลไกหลายอย่าง รวมถึงความล้มเหลวของไมโอซิสหรือไมโทซิส และการรวมตัวของแกมีต ที่ไม่ลดจำนวน ( 2n ) [ 39 ]ทั้งออโตโพลีพลอยด์ (เช่น มันฝรั่ง[ 74 ] ) และอัลโลโพลีพลอยด์ (เช่น คาโนลา ข้าวสาลี และฝ้าย) สามารถพบได้ในพืชป่าและพืชที่ปลูกเลี้ยง
โพลีพลอยด์ส่วนใหญ่แสดงความแปรผันหรือสัณฐานวิทยาใหม่เมื่อเทียบกับสายพันธุ์พ่อแม่ ซึ่งอาจมีส่วนช่วยในกระบวนการเกิดสปีชีส์และการใช้ประโยชน์จากนิเวศวิทยา[ 69 ] [ 39 ]กลไกที่นำไปสู่ความแปรผันใหม่ในอัลโลโพลีพลอยด์ที่เกิดขึ้นใหม่อาจรวมถึงผลกระทบจากปริมาณยีน (อันเป็นผลมาจากสำเนาของเนื้อหาจีโนมที่มากขึ้น) การรวมตัวกันของลำดับชั้นการควบคุมยีนที่แตกต่างกัน การจัดเรียงโครโมโซมใหม่ และ การปรับโครงสร้าง เอพิเจเนติกส์ซึ่งทั้งหมดนี้ส่งผลต่อเนื้อหายีนและ/หรือระดับการแสดงออก[ 75 ] [ 76 ] [ 77 ] [ 78 ]การเปลี่ยนแปลงอย่างรวดเร็วเหล่านี้จำนวนมากอาจมีส่วนช่วยในการแยกตัวทางการสืบพันธุ์และการเกิดสปีชีส์ อย่างไรก็ตาม เมล็ดที่เกิดจากการผสมข้ามระหว่างโพลีพลอยด์ เช่น ระหว่างโพลีพลอยด์กับสายพันธุ์พ่อแม่ มักมีการพัฒนาของเอนโดสเปิร์มที่ผิดปกติซึ่งทำให้ความสามารถในการอยู่รอดลดลง[ 79 ] [ 80 ]จึงมีส่วนช่วยในการ เกิด สปีชีส์โพลีพลอยด์ พืชโพลีพลอยด์อาจผสมพันธุ์กับพืชดิพลอยด์และผลิต เมล็ดโพลีพลอยด์ได้ ดังที่สังเกตได้ในกลุ่มอะกามิกของCrepis [ 81 ]
พืชบางชนิดเป็นไตรพลอยด์ เนื่องจากกระบวนการแบ่งเซลล์แบบไมโอซิสถูกรบกวน พืชเหล่านี้จึงเป็นหมัน โดยพืชทุกต้นจะมีโครงสร้างทางพันธุกรรมเหมือนกันหมด ตัวอย่างเช่นต้นหญ้าฝรั่น ( Crocus sativus ) ซึ่งขยายพันธุ์โดยการปักชำเท่านั้น นอกจากนี้ ไม้พุ่มหายากมากจากแทสเมเนียอย่าง Lomatia tasmanicaก็เป็นพืชไตรพลอยด์ที่เป็นหมันเช่นกัน
มีต้นสน โพลีพลอยด์ที่เกิดขึ้นตามธรรมชาติ เพียง ไม่กี่ชนิด [ 82 ]ตัวอย่างหนึ่งคือต้นเรดวูดชายฝั่งSequoia sempervirensซึ่งเป็นเฮกซาพลอยด์ (6 x ) ที่มีโครโมโซม 66 โครโมโซม (2 n = 6 x = 66) แม้ว่าที่มาจะไม่ชัดเจนก็ตาม[ 83 ]
พืชน้ำ โดยเฉพาะพืชใบเลี้ยงเดี่ยวมีโพลีพลอยด์จำนวนมาก[ 84 ]
พืชผล
ประมาณร้อยละ 30 ของพืชผลเป็นโพลีพลอยด์ที่เกิดขึ้นเมื่อไม่นานมานี้ โดยพืชผลหลายชนิดยังแสดงหลักฐานของการเกิดการเพิ่มจำนวนโครโมโซมในอดีตด้วย เช่น สตรอว์เบอร์รี กล้วย ข้าวสาลี อ้อย และบรอกโคลี[ 85 ]
พืชโพลีพลอยด์อาจเกิดขึ้นเองตามธรรมชาติ หรืออาจถูกชักนำให้เกิดโพลีพลอยด์ได้โดยปัจจัยด้านสิ่งแวดล้อม ห้องปฏิบัติการ หรือสารเคมี (เช่น การใช้สารเคมีคอลชิซีน กับเมล็ด ) การชักนำให้เกิดโพลีพลอยด์เป็นเทคนิคที่ใช้กันทั่วไปเพื่อเอาชนะภาวะเป็นหมันของพืชลูกผสมในระหว่างการปรับปรุงพันธุ์พืช ตัวอย่างเช่นทริติเคิลเป็นลูกผสมระหว่างข้าวสาลี ( Triticum turgidum ) และข้าวไรย์ ( Secale cereale ) มันรวมเอาลักษณะที่ต้องการของพ่อแม่เข้าด้วยกัน แต่ลูกผสมเริ่มต้นเป็นหมัน หลังจากเกิดโพลีพลอยด์แล้ว ลูกผสมจะสามารถสืบพันธุ์ได้และสามารถขยายพันธุ์ต่อไปจนกลายเป็นทริติเคิลได้
พืชที่มีชุดโครโมโซมหลายชุดบางชนิดเป็นหมัน โดยเฉพาะอย่างยิ่งชุดโครโมโซมเลขคี่ เช่น ไตรพลอยดี และนี่อาจเป็นลักษณะที่พึงประสงค์ ตัวอย่างเช่น ผลไม้ไร้เมล็ดหลายชนิดไร้เมล็ดเนื่องจากมีชุดโครโมโซมหลายชุด พืชเหล่านี้ขยายพันธุ์โดยใช้วิธีการสืบพันธุ์แบบไม่อาศัยเพศ เช่นการต่อกิ่ง
ตัวอย่าง
- พืชไตรพลอยด์: แอปเปิ้ล บาง พันธุ์ (เช่นBelle de Boskoop , Jonagold , Mutsu , Ribston Pippin ), กล้วย , ส้ม , ขิง , แตงโม , [ 86 ]ดอกหญ้าฝรั่น , เนื้อมะพร้าวสีขาว
- พืชเทตราพลอยด์: แอ ปเปิ้ลบางสายพันธุ์, ข้าวสาลีดูรัมหรือข้าวสาลีมาการ อนี , ฝ้าย , มันฝรั่ง , คาโนลา / เรพซีด , ต้นหอม, ยาสูบ , ถั่วลิสง , ส้มคินโนว์ , เพลาร์โกเนียม
- พืชเฮกซาพลอยด์: ดอกเบญจมาศ , ข้าวสาลี ขนมปัง , ข้าว สาลีลูกผสม , ข้าวโอ๊ต , กีวี[ 14 ]
- พืชออกตาพลอยด์: สตรอว์เบอร์รี , ดาเลีย , แพนซี , อ้อย , โอคา ( Oxalis tuberosa ) [ 87 ]
- พืชโดเดคาพลอยด์: อ้อยลูกผสม บางชนิด [ 88 ]
พืชบางชนิดมีระดับพลอยดีที่หลากหลาย เช่น ดอกทิวลิปและดอกลิลลี่มักพบได้ทั้งแบบดิพลอยด์และไตรพลอยด์ดอกลิลลี่กลางวัน ( พันธุ์ Hemerocallis ) มีให้เลือกทั้งแบบดิพลอยด์และเตตราพลอยด์ และแอปเปิลและส้มคินโนว์อาจเป็นแบบดิพลอยด์ ไตรพลอยด์ หรือเตตราพลอยด์ก็ได้
เชื้อรา
นอกจากพืชและสัตว์แล้ว ประวัติวิวัฒนาการของเชื้อรา หลายชนิด ยังเต็มไปด้วยเหตุการณ์การเพิ่มจำนวนจีโนมทั้งหมดในอดีตและปัจจุบัน (ดู Albertin และ Marullo 2012 [ 89 ]สำหรับการทบทวน) ตัวอย่างของโพลีพลอยด์หลายชนิดเป็นที่รู้จัก:
- ออ โตโพลีพลอยด์: เชื้อราในน้ำสกุลAllomyces [ 90 ] สาย พันธุ์ Saccharomyces cerevisiaeบางสายพันธุ์ที่ใช้ในเบเกอรี่ [ 91 ]เป็นต้น
- อัลโลโพลีพลอยด์: Cyathus stercoreus ที่แพร่หลาย [ 92 ]ยีสต์ลาเกอร์อัลโลเทตราพลอยด์Saccharomyces pastorianus [ 93 ] ยีสต์ทำให้ไวน์เสียอัลโลไตรพลอยด์Dekkera bruxellensis [ 94 ]เป็นต้น
- พาเลโอโพลีพลอย ด์: เชื้อก่อโรคในมนุษย์Rhizopus oryzae [ 95 ]สกุลSaccharomyces [ 96 ] เป็นต้น
นอกจากนี้ โพลีพลอยดีมักเกี่ยวข้องกับการผสมข้ามพันธุ์และวิวัฒนาการแบบเครือข่ายซึ่งดูเหมือนจะแพร่หลายในกลุ่มเชื้อราหลายกลุ่ม อันที่จริงการเกิดสปีชีส์โฮโมพลอยด์ (การเกิดสปีชีส์ลูกผสมโดยไม่มีการเปลี่ยนแปลง จำนวน โครโมโซม ) ได้รับการพิสูจน์แล้วสำหรับเชื้อราบางสปีชีส์ (เช่นMicrobotryum violaceumในกลุ่ม Basidiomycota [ 97 ] )

สำหรับพืชและสัตว์ ลูกผสมของเชื้อราและโพลีพลอยด์แสดงการเปลี่ยนแปลงโครงสร้างและหน้าที่เมื่อเทียบกับบรรพบุรุษและคู่ดิพลอยด์ โดยเฉพาะอย่างยิ่ง ผลลัพธ์เชิงโครงสร้างและหน้าที่ของจีโนมโพลีพลอยด์ของ Saccharomycesสะท้อนให้เห็นถึงชะตากรรมวิวัฒนาการของจีโนมโพลีพลอยด์ของพืชอย่างน่าทึ่ง มีการอธิบายถึงการจัดเรียงโครโมโซมขนาดใหญ่[ 98 ]ซึ่งนำไปสู่โครโมโซมไคเมอริก[ 99 ]รวมถึงการดัดแปลงทางพันธุกรรมที่เฉพาะเจาะจงมากขึ้น เช่น การสูญเสียยีน[ 100 ]โฮโมอีลลีลของยีสต์อัลโลเทตราพลอยด์ S. pastorianusแสดงให้เห็นถึงการมีส่วนร่วมที่ไม่เท่ากันในทรานสคริปโตม [ 101 ] การ กระจายตัวของ ฟี โนไทป์ ยังพบได้หลังจากการเกิดโพลีพลอยด์และ/หรือการผสมพันธุ์ในเชื้อรา[ 102 ]ซึ่งเป็นเชื้อเพลิงสำหรับการคัดเลือกโดยธรรมชาติ และ การปรับตัวและการเกิดสปีชีส์ ในภายหลัง
โครมัลวีโอลาตา
กลุ่มสิ่งมีชีวิตยูคาริโอ ตอื่นๆได้ประสบกับเหตุการณ์โพลีพลอยด์หนึ่งครั้งหรือมากกว่านั้นในช่วงประวัติศาสตร์วิวัฒนาการของพวกมัน (ดู Albertin และ Marullo, 2012 [ 89 ]สำหรับการทบทวน) โอโอไมซีตซึ่งเป็นสมาชิกของเชื้อราที่ไม่ใช่เชื้อราแท้ มีตัวอย่างของสปีชีส์โพลีพลอยด์โบราณและโพลีพลอยด์หลายชนิด เช่น ภายในสกุลPhytophthora [ 103 ] ส ปีชีส์ของ สาหร่ายสีน้ำตาลบางชนิด( Fucales , Laminariales [ 104 ]และไดอะตอม[ 105 ] ) มีจีโนมโพลีพลอยด์ที่เห็นได้ชัด ใน กลุ่ม Alveolataสปีชีส์ที่โดดเด่นอย่างParamecium tetraureliaได้ผ่านการจำลองจีโนมทั้งหมดสามรอบติดต่อกัน[ 106 ]และได้สร้างตัวเองให้เป็นแบบจำลองหลักสำหรับการศึกษาโพลีพลอยด์โบราณ
แบคทีเรีย
แบคทีเรียDeinococcus radiodurans แต่ละตัวมีโครโมโซม 4-8 ชุด [ 107 ]การที่D. radiodurans สัมผัส กับ รังสี เอ็กซ์หรือการขาดน้ำอาจทำให้จีโนม ของมันแตกออก เป็นชิ้นส่วนเล็กๆ สุ่มๆ นับร้อย อย่างไรก็ตามD. radiodurans มีความต้านทานสูงต่อการสัมผัสเช่นนี้ กลไกที่ทำให้จีโนมได้รับการฟื้นฟูอย่างแม่นยำนั้นเกี่ยวข้องกับ การรวมตัวกันของโฮโมโลจัสที่ควบคุมโดย RecA และกระบวนการที่เรียกว่าการเชื่อมต่อสายที่ขึ้นอยู่กับการสังเคราะห์แบบขยาย (SDSA ) [ 108 ]
Azotobacter vinelandiiสามารถมีโครโมโซมได้มากถึง 80 ชุดต่อเซลล์ [ 109 ]อย่างไรก็ตาม สังเกตได้เฉพาะในวัฒนธรรมที่เติบโตอย่างรวดเร็วเท่านั้น ในขณะที่วัฒนธรรมที่เติบโตในอาหารเลี้ยงเชื้อสังเคราะห์ขั้นต่ำจะไม่เป็นโพลีพลอยด์ [ 110 ]
อาร์เคีย
อาร์เคียHalobacterium salinariumเป็นโพลีพลอยด์[ 111 ]และเช่นเดียวกับDeinococcus radioduransมีความทนทานสูงต่อการฉายรังสีเอ็กซ์และการขาดน้ำ ซึ่งเป็นสภาวะที่ทำให้เกิด การแตก ของดีเอ็นเอแบบสองสาย[ 112 ] แม้ว่าโครโมโซมจะแตกเป็นชิ้นเล็กชิ้นน้อยจำนวนมาก แต่โครโมโซมที่สมบูรณ์สามารถสร้างขึ้นใหม่ได้โดยใช้ชิ้นส่วนที่ซ้อนทับกัน กลไกนี้ใช้ โปรตีนที่จับกับดีเอ็นเอแบบสายเดี่ยวและน่าจะเป็นการซ่อมแซมแบบโฮโมโลจัสรีคอมบิเนชัน[ 113 ]
ดูเพิ่มเติม
- การเพิ่มจำนวนโครโมโซมเป็นสองชุด (Diploidization)
- จีโนมลูกผสมยูคาริโอต
- ระดับพลอยดี
- คอมเพล็กซ์โพลีพลอยด์
- โพลีโซมี
- การเงียบงันซึ่งกันและกัน
- ซิมแพทรี
อ่านเพิ่มเติม
- Snustad DP, Simmons MJ (2006). หลักการทางพันธุศาสตร์ (ฉบับที่ 4). โฮโบเคน รัฐนิวเจอร์ซีย์: John Wiley & Sons. ISBN 978-0-471-69939-2.
- โครงการ ริเริ่มจีโน มของArabidopsis (ธันวาคม 2000) " การวิเคราะห์ลำดับจีโนมของพืชดอก Arabidopsis thaliana" Nature 408 ( 6814 ): 796– 815. Bibcode : 2000Natur.408..796T doi : 10.1038/35048692 PMID 11130711
- Eakin GS, Behringer RR (ธันวาคม 2003). "การพัฒนาแบบเตตราพลอยด์ในหนู" . Developmental Dynamics . 228 (4): 751– 766. doi : 10.1002/dvdy.10363 . PMID 14648853 .
- Gaeta RT, Pires JC, Iniguez-Luy F, Leon E, Osborn TC (พฤศจิกายน 2550). "การเปลี่ยนแปลงทางจีโนมใน Brassica napus ที่สังเคราะห์ขึ้นใหม่และผลกระทบต่อการแสดงออกของยีนและฟีโนไทป์" The Plant Cell . 19 (11): 3403– 3417. Bibcode : 2007PlanC..19.3403G . doi : 10.1105/tpc.107.054346 . PMC 2174891 . PMID 18024568 .
- Gregory, T. Ryan; Mable, Barbara K. (2005). "Polyploidy in Animals". The Evolution of the Genome . หน้า 427–517 . doi : 10.1016/B978-012301463-4/50010-3 . ISBN 978-0-12-301463-4.
- Jaillon O, Aury JM, Brunet F, Petit JL, Stange - Thomann N, Mauceli E และคณะ (ตุลาคม 2547) "การจำลองจีโนมในปลาเทเลออส Tetraodon nigroviridis เผยให้เห็นโปรโตคาริโอไทป์ของสัตว์มีกระดูกสันหลังยุคแรก" Nature 431 ( 7011): 946– 957. Bibcode : 2004Natur.431..946J . doi : 10.1038/nature03025 . PMID 15496914 .
- Paterson AH, Bowers JE, Van de Peer Y, Vandepoele K (มีนาคม 2548). "การทำซ้ำจีโนมธัญพืชโบราณ" The New Phytologist . 165 (3): 658– 661. Bibcode : 2005NewPh.165..658P . doi : 10.1111/j.1469-8137.2005.01347.x . PMID 15720677 .
- Raes J, Vandepoele K, Simillion C, Saeys Y, Van de Peer Y (2003). "การตรวจสอบเหตุการณ์การทำซ้ำโบราณในจีโนมของ Arabidopsis"วารสารStructural and Functional Genomics 3 ( 1– 4 ): 117– 129. doi : 10.1023/A:1022666020026 . PMID 12836691 .
- Simillion C, Vandepoele K, Van Montagu MC, Zabeau M, Van de Peer Y (ตุลาคม 2545). "อดีตการทำซ้ำที่ซ่อนเร้นของ Arabidopsis thaliana" Proceedings of the National Academy of Sciences of the United States of America . 99 (21): 13627– 13632. Bibcode : 2002PNAS...9913627S . doi : 10.1073/pnas.212522399 . JSTOR 3073458 . PMC 129725 . PMID 12374856 .
- Soltis DE, Soltis PS , Schemske DW, Hancock JF, Thompson JN, Husband BC, Judd WS (2007). "Autopolyploidy ในพืชดอก: เราประเมินจำนวนชนิดต่ำเกินไปหรือไม่?" Taxon . 56 (1): 13– 30. JSTOR 25065732 .
- Soltis DE, Buggs RJ, Doyle JJ, Soltis PS (2010). "สิ่งที่เรายังไม่รู้เกี่ยวกับโพลีพลอยดี" Taxon . 59 (5): 1387– 1403. Bibcode : 2010Taxon..59.1387S . doi : 10.1002/tax.595006 .
- Taylor JS, Braasch I, Frickey T, Meyer A, Van de Peer Y (มีนาคม 2003). "การจำลองจีโนม ลักษณะร่วมของปลาครีบแข็ง 22,000 ชนิด" . Genome Research . 13 (3): 382– 390. doi : 10.1101/gr.640303 . PMC 430266 . PMID 12618368 .
- Tate, Jennifer A.; Soltis, Douglas E.; Soltis, Pamela S. (2005). "Polyploidy in Plants". The Evolution of the Genome . หน้า 371–426 . doi : 10.1016/B978-012301463-4/50009-7 . ISBN 978-0-12-301463-4.
- Van de Peer Y, Taylor JS, Meyer A (2003). "ปลาทั้งหมดเป็นโพลีพลอยด์โบราณหรือไม่?"วารสาร จีโนมิก ส์เชิงโครงสร้างและหน้าที่3 ( 1– 4): 65– 73. doi : 10.1023/A:1022652814749 . PMID 12836686 .
- Van de Peer Y (2004). "จีโนมของ Tetraodon ยืนยันผลการค้นพบ Takifugu: ปลาส่วนใหญ่เป็นโพลีพลอยด์โบราณ" . Genome Biology . 5 (12): 250. doi : 10.1186/gb-2004-5-12-250 . PMC 545788 . PMID 15575976 .
- Van De Peer, Yves; Meyer, Axel (2005). "การเพิ่มจำนวนยีนขนาดใหญ่และจีโนมโบราณ" วิวัฒนาการของจีโนมหน้า 329–368 . doi : 10.1016/B978-012301463-4/50008-5 . ISBN 978-0-12-301463-4.
- Wolfe KH, Shields DC (มิถุนายน 1997). "หลักฐานระดับโมเลกุลสำหรับการจำลองแบบโบราณของจีโนมยีสต์ทั้งหมด" Nature . 387 ( 6634): 708– 713. Bibcode : 1997Natur.387..708W . doi : 10.1038/42711 . PMID 9192896 .
- Wolfe KH (พฤษภาคม 2001). "โพลีพลอยด์เมื่อวานนี้และความลึกลับของการเกิดดิพลอยด์" Nature Reviews. Genetics . 2 (5): 333– 341. doi : 10.1038/35072009 . PMID 11331899 .
ลิงก์ภายนอก
- โพลีพลอยดี (Polyploidy) ในหนังสือชีววิทยาของคิมบอล (Kimball's Biology Pages)
- เว็บไซต์ Polyploidy portalเป็นโครงการที่ชุมชนสามารถแก้ไขได้ โดยมีข้อมูล งานวิจัย การศึกษา และบรรณานุกรมเกี่ยวกับโพลีพลอยดี
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ โพลีพลอยดี
ภาวะ โพลีพลอยด์ คือ ภาวะที่ เซลล์ ของ สิ่งมีชีวิต มี โครโมโซม คู่ ( โฮโมล็อกัส ) มากกว่าสองคู่สิ่งมีชีวิตส่วนใหญ่ที่มี นิวเคลียส ( ยูคาริโอต ) จะเป็น ดิพลอยด์...
การจำแนกประเภท
ออโตโพลีพลอยด์ คือ โพลีพลอย ด์ ที่มีชุดโครโมโซมหลายชุดซึ่งได้มาจาก กลุ่มสิ่งมีชีวิต เดียว
เงื่อนไขเชิงเวลา
พืชที่มีชุดโครโมโซมหลายชุดซึ่งเพิ่งเกิดขึ้นใหม่
คำศัพท์อื่นๆ ที่คล้ายกัน
คาริโอไทป์คือชุดโครโมโซมลักษณะเฉพาะของ สิ่งมี ชีวิต ยู คาริโอต [ 48 ] [ 49 ] การเตรียมและการศึกษาคาริโอไทป์เป็นส่วนหนึ่งของ เซลล์วิทยา และโดยเฉพาะอย่างยิ่ง เซลล์พันธุ ศาสตร์