อ่าน 15 นาที
การแปล (ชีววิทยา)
การแปลรหัสพันธุกรรม เป็นกระบวนการใน เซลล์ชีวภาพ ที่ สร้าง โปรตีน โดย ใช้ โมเลกุล RNA เป็นแม่แบบ โปรตีนที่สร้างขึ้นเป็นลำดับของ กรดอะมิโน ที่กำหนดโดยลำดับของ นิวคลีโอไทด์ ใน RNA...
การแปล (ชีววิทยา)

| ส่วนหนึ่งของชุดบทความเกี่ยวกับ |
| ชีววิทยา |
|---|
 |
|
การแปลรหัสพันธุกรรมเป็นกระบวนการในเซลล์ชีวภาพ ที่ สร้างโปรตีนโดย ใช้ โมเลกุลRNA เป็นแม่แบบ โปรตีนที่สร้างขึ้นเป็นลำดับของ กรดอะมิโนที่กำหนดโดยลำดับของนิวคลีโอไทด์ใน RNA โดยจะพิจารณานิวคลีโอไทด์ครั้งละสามตัว แต่ละกลุ่มสามตัวจะทำให้เกิดกรดอะมิโนเฉพาะหนึ่งชนิดในโปรตีนที่สร้างขึ้น การจับคู่ระหว่างกลุ่มนิวคลีโอไทด์กับกรดอะมิโนเรียกว่ารหัสพันธุกรรมการแปลรหัสพันธุกรรมดำเนินการโดยกลุ่มของ RNA และโปรตีนที่ทำหน้าที่เรียกว่าไรโบโซมกระบวนการทั้งหมดนี้เรียกว่าการแสดงออกของยีน
ในการแปลรหัส mRNA จะถูกถอดรหัสในไรโบโซมซึ่งอยู่นอกนิวเคลียส เพื่อสร้าง สาย กรดอะมิโน เฉพาะ หรือพอลิเปปไทด์พอลิเปปไทด์จะพับตัวเป็น โปรตีน ที่ทำงานได้และทำหน้าที่ในเซลล์ พอลิเปปไทด์ยังสามารถเริ่มพับตัวได้ในระหว่างการสังเคราะห์โปรตีน[ 1 ]ไรโบโซมช่วยอำนวยความสะดวกในการถอดรหัสโดยการเหนี่ยวนำให้ลำดับแอนติโคดอนของ tRNA ที่ เป็นคู่กัน จับกับ โคดอนของ mRNA tRNA จะนำกรดอะมิโนเฉพาะที่เชื่อมต่อกันเป็นพอลิเปปไทด์เมื่อ mRNA ผ่านและถูก "อ่าน" โดยไรโบโซม ขั้นตอนการแปลรหัสมีสามขั้นตอน ได้แก่ การเริ่มต้น การยืดตัว และการสิ้นสุด
กลไกพื้นฐาน

กระบวนการพื้นฐานของการผลิตโปรตีนคือการเพิ่มกรดอะมิโนทีละตัวที่ปลายของสายโพลีเปปไทด์ที่กำลังก่อตัว การดำเนินการนี้ทำโดย ไร โบโซม[ 2 ]ไรโบโซมประกอบด้วยสองหน่วยย่อย ในยูคาริโอตจะมีหน่วยย่อยขนาดเล็ก (40S)และหน่วยย่อยขนาดใหญ่ (60S)หน่วยย่อยเหล่านี้จะมารวมกันก่อนการแปล mRNA เป็นโปรตีน เพื่อให้ได้ตำแหน่งสำหรับการแปลและการผลิตโพลีเปปไทด์[ 3 ]การเลือกชนิดของกรดอะมิโนที่จะเพิ่มนั้นถูกกำหนดโดย โมเลกุลของ สารส่งสัญญาณ RNA (mRNA) กรดอะมิโนแต่ละตัวที่เพิ่มเข้ามาจะจับคู่กับลำดับย่อยสามนิวคลีโอไทด์ของ mRNA สำหรับแต่ละไตรเพล็ตที่เป็นไปได้ กรดอะมิโนที่สอดคล้องกันจะได้รับการยอมรับ กรดอะมิโนที่เพิ่มเข้ามาในสายโซ่จะจับคู่กับไตรเพล็ตนิวคลีโอไทด์ที่ต่อเนื่องกันใน mRNA ด้วยวิธีนี้ ลำดับของนิวคลีโอไทด์ในสาย mRNA แม่แบบจะกำหนดลำดับของกรดอะมิโนในสายกรดอะมิโนที่สร้างขึ้น[ 4 ] การเพิ่มกรดอะมิโนเกิดขึ้นที่ปลาย Cของเปปไทด์ ดังนั้นการแปลจึงกล่าวได้ว่าเป็นการแปลแบบอะมีนไปยังคาร์บอกซิล[ 5 ]
เอ็ ม อาร์เอ็น เอ (mRNA) ทำหน้าที่ ขนส่งข้อมูล ทางพันธุกรรมที่เข้ารหัสอยู่ในรูปของลำดับนิวคลีโอไทด์จากโครโมโซมไปยังไรโบโซม ไรโบนิวคลีโอไทด์เหล่านี้จะถูก "อ่าน" โดยกลไกการแปลรหัสทางพันธุกรรมในลำดับของกลุ่มนิวคลี โอไทด์ สามตัวที่เรียกว่าโคดอนแต่ละกลุ่มนิวคลีโอไทด์สามตัวนั้นจะเข้ารหัสกรดอะมิโน เฉพาะชนิดหนึ่ง
โมเลกุลของไรโบโซมจะแปลรหัสนี้ไปเป็นลำดับกรดอะมิโนที่เฉพาะเจาะจง ไรโบโซมเป็นโครงสร้างหลายหน่วยย่อยที่ประกอบด้วยไรโบโซมอลอาร์เอ็นเอ (rRNA) และโปรตีน มันคือ "โรงงาน" ที่กรดอะมิโนถูกประกอบเข้าด้วยกันเป็นโปรตีน
ทรานสเฟอร์อาร์เอ็นเอ (tRNA) เป็นสายอาร์เอ็นเอขนาดเล็กที่ไม่มีรหัส (74–93 นิวคลีโอไทด์) ซึ่งทำหน้าที่ขนส่งกรดอะมิโนไปยังไรโบโซม ชุดยีนของ tRNA มีความหลากหลายอย่างมากในแต่ละสปีชีส์ โดยแบคทีเรียบางชนิดมีระหว่าง 20 ถึง 30 ยีน ในขณะที่ยูคาริโอตที่ซับซ้อนอาจมีหลายพันยีน[ 6 ] tRNA มีไซต์สำหรับยึดกรดอะมิโน และไซต์ที่เรียกว่าแอนติโคดอน แอนติโคดอนเป็นไตรเพล็ตของอาร์เอ็นเอที่เสริมกับไตรเพล็ตของเอ็มอาร์เอ็นเอที่เข้ารหัสกรดอะมิโนที่ เป็นเป้าหมาย
เอนไซม์ อะมิโนเอซิลทีอาร์เอ็นเอซินเทส ( ) ทำหน้าที่เร่งปฏิกิริยาการเชื่อมต่อระหว่างทีอาร์เอ็นเอเฉพาะกับกรดอะมิโน ที่ลำดับแอ นติโคดอนของทีอาร์เอ็นเอต้องการ ผลิตภัณฑ์ของปฏิกิริยานี้คืออะ มิ โนเอซิลทีอาร์เอ็นเอ กรดอะมิโนจะเชื่อมต่อด้วยหมู่คาร์บอกซิลกับ 3' OH ของทีอาร์เอ็นเอด้วยพันธะเอสเทอร์ เมื่อทีอาร์เอ็นเอมีกรดอะมิโนเชื่อมต่ออยู่ ทีอาร์เอ็นเอจะถูกเรียกว่า "ชาร์จ" อะมิโนเอซิลทีอาร์เอ็นเอซินเทสที่จับคู่ทีอาร์เอ็นเอกับกรดอะมิโนผิด สามารถสร้างอะมิโนเอซิลทีอาร์เอ็นเอที่ชาร์จผิด ซึ่งอาจส่งผลให้กรดอะมิโนที่ตำแหน่งนั้นในโปรตีนไม่เหมาะสม การ "แปลรหัสผิดพลาด" [ 7 ]ของรหัสพันธุกรรมนี้เกิดขึ้นตามธรรมชาติในระดับต่ำในสิ่งมีชีวิตส่วนใหญ่ แต่สภาพแวดล้อมของเซลล์บางอย่างทำให้การถอดรหัส mRNA ที่อนุญาตเพิ่มขึ้น ซึ่งบางครั้งก็เป็นประโยชน์ต่อเซลล์
ไรโบโซมมีตำแหน่งจับกับ tRNA สองตำแหน่ง ได้แก่ ตำแหน่งอะมิโนเอซิล (ย่อว่า A) และตำแหน่งเปปทิดิล/ตำแหน่งออก (ย่อว่า P/E) สำหรับ mRNA นั้น ทั้งสามตำแหน่งจะเรียงตัวในแนว5'ถึง3' EPA เนื่องจากไรโบโซมเคลื่อนที่ไปทางปลาย 3' ของ mRNA ตำแหน่ง Aจับกับ tRNA ที่เข้ามาซึ่งมีโคดอนที่เข้าคู่กันบน mRNA ตำแหน่ง P/Eจับกับ tRNA ที่มีสายโพลีเปปไทด์ที่กำลังเติบโต เมื่ออะมิโนเอซิล-tRNA จับกับโคดอนที่ตรงกันบน mRNA ในตอนแรก มันจะอยู่ในตำแหน่ง A จากนั้น พันธะเปปไทด์จะเกิดขึ้นระหว่างกรดอะมิโนของ tRNA ในตำแหน่ง A และกรดอะมิโนของ tRNA ที่มีประจุในตำแหน่ง P/E สายโพลีเปปไทด์ที่กำลังเติบโตจะถูกถ่ายโอนไปยัง tRNA ในตำแหน่ง A การเคลื่อนย้ายเกิดขึ้น ทำให้ tRNA เคลื่อนไปยังตำแหน่ง P/E โดยที่ไม่มีกรดอะมิโนอยู่ด้วย tRNA ที่อยู่ในไซต์ A ซึ่งตอนนี้มีโซ่โพลีเปปไทด์อยู่แล้ว จะถูกย้ายไปยังไซต์ P/E และtRNA ที่ไม่มีประจุจะออกไป และ tRNA อะมิโนเอซิลอีกตัวจะเข้าไปในไซต์ A เพื่อทำซ้ำกระบวนการ[ 8 ]
หลังจากที่กรดอะมิโนตัวใหม่ถูกเพิ่มเข้าไปในสายโซ่ และหลังจากที่ tRNA ถูกปล่อยออกจากไรโบโซมและเข้าสู่ไซโตซอล พลังงานที่ได้จากการไฮโดรไลซิสของ GTP ที่จับกับทรานสโลเคสEEF2 จะเคลื่อนไรโบโซมลงไปหนึ่งโคดอนไปทางปลาย 3'พลังงานที่จำเป็นสำหรับการแปลโปรตีนนั้นมีนัยสำคัญ สำหรับโปรตีนที่มี กรดอะมิโน nตัว จำนวนพันธะฟอสเฟตพลังงานสูงที่จำเป็นในการแปลโปรตีนนั้นคือ 4 n -1 [ 9 ]อัตราการแปลโปรตีนนั้นแตกต่างกัน โดยจะสูงกว่าอย่างมีนัยสำคัญในเซลล์โปรคาริโอต (มากถึง 17–21 หน่วยกรดอะมิโนต่อวินาที) มากกว่าในเซลล์ยูคาริโอต (มากถึง 6–9 หน่วยกรดอะมิโนต่อวินาที) [ 10 ]
เวที
การแปลดำเนินไปในสี่ขั้นตอน ได้แก่ การเริ่มต้น การยืดออก การสิ้นสุด และการรีไซเคิล[ 11 ]
การเริ่มต้น
การเริ่มต้นเกี่ยวข้องกับหน่วยย่อยขนาดเล็กของไรโบโซมที่จับกับปลาย 5' ของ mRNA โดยอาศัยปัจจัยการเริ่มต้น [ 12 ] ไรโบโซมและปัจจัยที่เกี่ยวข้องจะประกอบกันและจับกับ mRNA tRNA ตัวแรกจะถูกยึดติดที่โคดอนเริ่มต้นกระบวนการนี้ถูกกำหนดให้เป็นแบบขึ้นอยู่กับแคป ซึ่งไรโบโซมจะจับที่แคป 5' ในตอนแรกแล้วจึงเคลื่อนที่ไปยังโคดอนหยุด หรือแบบไม่ขึ้นอยู่กับแคป ซึ่งไรโบโซมจะไม่จับกับแคป 5' ในตอนแรก แคป 5' จะถูกเพิ่มเมื่อพรี-mRNA ที่เกิดขึ้นใหม่มีความยาวประมาณ 20 นิวคลีโอไทด์[ 13 ]
การเริ่มต้นที่ขึ้นอยู่กับแคป
การเริ่มต้นการแปลมักเกี่ยวข้องกับการโต้ตอบของโปรตีนสำคัญบางชนิดปัจจัยเริ่มต้นกับแท็กพิเศษที่ผูกติดกับปลาย 5' ของโมเลกุล mRNA ซึ่งก็คือ5' capรวมถึง5' UTR ด้วย โปรตีนเหล่านี้จะจับกับหน่วยย่อยไร โบโซมขนาดเล็ก (40S) และยึด mRNA ไว้ในตำแหน่ง[ 14 ]
eIF3เกี่ยวข้องกับหน่วยย่อยไรโบโซม 40S และมีบทบาทในการป้องกันไม่ให้หน่วยย่อยไรโบโซมขนาดใหญ่ (60S) จับตัวก่อนกำหนด eIF3 ยังมีปฏิสัมพันธ์กับคอมเพล็กซ์eIF4Fซึ่งประกอบด้วยปัจจัยเริ่มต้นอีกสามตัว ได้แก่eIF4A , eIF4EและeIF4G eIF4G เป็นโปรตีนโครงสร้างที่เชื่อมโยงโดยตรงกับทั้ง eIF3 และส่วนประกอบอีกสองตัวeIF4Eเป็นโปรตีนที่จับกับแคป การจับแคปโดย eIF4E มักถูกพิจารณาว่าเป็นขั้นตอนที่จำกัดอัตราของการเริ่มต้นแบบขึ้นอยู่กับแคป และความเข้มข้นของ eIF4E เป็นจุดควบคุมการแปลรหัส ไวรัสบางชนิดจะตัดส่วนหนึ่งของ eIF4G ที่จับกับ eIF4E ออกไป ทำให้การแปลรหัสแบบขึ้นอยู่กับแคปไม่สามารถแย่งชิงกลไกของโฮสต์ไปสนับสนุนข้อความของไวรัส (แบบไม่ขึ้นอยู่กับแคป) ได้eIF4Aเป็น RNA เฮลิเคสที่ขึ้นอยู่กับ ATP ซึ่งช่วยไรโบโซมโดยการแก้ไขโครงสร้างทุติยภูมิบางอย่างที่เกิดขึ้นตามแนวการถอดรหัส mRNA ผลการศึกษาชีววิทยาเชิงโครงสร้างล่าสุดยังระบุว่าโปรตีน eIF4A ตัวที่สองสามารถเชื่อมโยงกับคอมเพล็กซ์การเริ่มต้นได้พร้อมกัน โดยเฉพาะอย่างยิ่งการโต้ตอบกับ eIF3 [ 15 ] [ 16 ]โปรตีนที่จับกับโพลี(A) (PABP) ยังเชื่อมโยงกับ คอมเพล็กซ์ eIF4Fผ่านทาง eIF4G และจับกับหางโพลี-Aของโมเลกุล mRNA ของยูคาริโอตส่วนใหญ่ โปรตีนนี้มีส่วนเกี่ยวข้องในการทำให้เกิดวงกลมของ mRNA ในระหว่างการแปล[ 17 ]
คอมเพล็กซ์เริ่มต้นก่อนการสังเคราะห์โปรตีน 43S (43S PIC) พร้อมด้วยโปรตีนแฟคเตอร์จะเคลื่อนที่ไปตามสาย mRNA ไปยังปลาย 3' ในกระบวนการที่เรียกว่า 'การสแกน' เพื่อไปถึงรหัสเริ่มต้น (โดยทั่วไปคือ AUG) ในยูคาริโอตและอาร์เคียกรดอะมิโนที่เข้ารหัสโดยรหัสเริ่มต้นคือเมไทโอนีนtRNA เริ่มต้นที่มีเมไทโอนีน (Met-tRNA i Met )จะถูกนำไปยังตำแหน่ง P ของหน่วยย่อยไรโบโซมขนาดเล็กโดยแฟคเตอร์เริ่มต้นยูคาริโอต 2 (eIF2)มันจะไฮโดรไลซ์ GTP และส่งสัญญาณให้แฟคเตอร์หลายตัวแยกตัวออกจากหน่วยย่อยไรโบโซมขนาดเล็ก ซึ่งในที่สุดจะนำไปสู่การรวมตัวของหน่วยย่อยขนาดใหญ่ (หรือ หน่วยย่อย 60S ) จากนั้นไรโบโซมที่สมบูรณ์ ( 80S ) จะเริ่มการยืดตัวของการแปลรหัส
การควบคุมการสังเคราะห์โปรตีนได้รับอิทธิพลบางส่วนจากการฟอสโฟรีเลชันของeIF2 (ผ่านทางซับยูนิต α) ซึ่งเป็นส่วนหนึ่งของสารประกอบเชิงซ้อนสามส่วน eIF2-GTP-Met-tRNA i Met (eIF2-TC) เมื่อ eIF2 จำนวนมากถูกฟอสโฟรีเลชัน การสังเคราะห์โปรตีนจะถูกยับยั้ง ซึ่งเกิดขึ้นภายใต้ภาวะขาดแคลนกรดอะมิโนหรือหลังจากการติดเชื้อไวรัส อย่างไรก็ตาม ปัจจัยเริ่มต้นนี้เพียงส่วนน้อยเท่านั้นที่ถูกฟอสโฟรีเลชันตามธรรมชาติ ตัวควบคุมอีกตัวหนึ่งคือ4EBPซึ่งจับกับปัจจัยเริ่มต้นeIF4Eและยับยั้งการโต้ตอบกับeIF4Gจึงป้องกันการเริ่มต้นแบบขึ้นอยู่กับแคป เพื่อต่อต้านผลกระทบของ 4EBP ปัจจัยการเจริญเติบโตจะฟอสโฟรีเลต 4EBP ลดความสัมพันธ์ของมันกับ eIF4E และอนุญาตให้มีการสังเคราะห์โปรตีน
ในขณะที่การสังเคราะห์โปรตีนได้รับการควบคุมทั่วโลกโดยการปรับการแสดงออกของปัจจัยเริ่มต้นที่สำคัญ รวมถึงจำนวนของไรโบโซม mRNA แต่ละตัวสามารถมีอัตราการแปลที่แตกต่างกันได้เนื่องจากการมีอยู่ขององค์ประกอบลำดับควบคุม สิ่งนี้ได้รับการพิสูจน์แล้วว่ามีความสำคัญในหลายบริบท รวมถึงไมโอซิสของยีสต์และการตอบสนองต่อเอทิลีนในพืช นอกจากนี้ งานวิจัยล่าสุดในยีสต์และมนุษย์ชี้ให้เห็นว่าความแตกต่างทางวิวัฒนาการในลำดับควบคุมซิสสามารถส่งผลกระทบต่อการควบคุมการแปลได้[ 18 ] นอกจากนี้ เฮลิเคส RNA เช่นDHX29และDed1/DDX3มีส่วนร่วมในกระบวนการเริ่มต้นการแปล โดยเฉพาะอย่างยิ่งสำหรับ mRNA ที่มี 5'UTR ที่มีโครงสร้าง[ 19 ] [ 20 ]
การเริ่มต้นที่ไม่ขึ้นกับแคป
ตัวอย่างที่ได้รับการศึกษามากที่สุดของการเริ่มต้นการแปลที่ไม่ขึ้นกับแคปในยูคาริโอตคือการใช้ไซต์ทางเข้าไรโบโซมภายใน (IRES) ซึ่งแตกต่างจากการแปลที่ขึ้นกับแคป การแปลที่ไม่ขึ้นกับแคปไม่จำเป็นต้องใช้แคป 5' เพื่อเริ่มต้นการสแกนจากปลาย 5' ของ mRNA จนถึงโคดอนเริ่มต้น ไรโบโซมสามารถกำหนดตำแหน่งไปยังไซต์เริ่มต้นได้โดยการจับโดยตรง ปัจจัยการเริ่มต้น และ/หรือ ITAFs (ปัจจัยที่ออกฤทธิ์ข้าม IRES) โดยไม่ต้องสแกน5' UTR ทั้งหมด วิธีการแปลนี้มีความสำคัญในสภาวะที่ต้องการการแปล mRNA เฉพาะในระหว่างความเครียดของเซลล์ เมื่อการแปลโดยรวมลดลง ตัวอย่างเช่น ปัจจัยที่ตอบสนองต่ออะพอพโทซิสและการตอบสนองที่เกิดจากความเครียด[ 21 ]
การยืดตัว
การยืดสายขึ้นอยู่กับปัจจัยการยืดสายในตอนท้ายของขั้นตอนเริ่มต้น mRNA จะถูกจัดวางตำแหน่งเพื่อให้สามารถแปลรหัสพันธุกรรมถัดไปได้ในระหว่างขั้นตอนการยืดสายของการสังเคราะห์โปรตีน tRNA ตัวเริ่มต้นจะเข้าครอบครองตำแหน่ง P ในไรโบโซม และตำแหน่ง A พร้อมที่จะรับอะมิโนเอซิล-tRNA ในระหว่างการยืดสาย กรดอะมิโนแต่ละตัวจะถูกเพิ่มเข้าไปในสายโพลีเปปไทด์ที่เกิดขึ้นใหม่ในไมโครไซเคิลสามขั้นตอน ขั้นตอนในไมโครไซเคิลนี้คือ (1) การจัดวางอะมิโนเอซิล-tRNA ที่ถูกต้องในตำแหน่ง A ของไรโบโซม ซึ่งถูกนำเข้ามาในตำแหน่งนั้นโดย eEF1 (2) การสร้างพันธะเปปไทด์ และ (3) การเลื่อน mRNA ไปหนึ่งรหัสพันธุกรรมเมื่อเทียบกับไรโบโซมด้วยความช่วยเหลือของ eEF2 ต่างจากแบคทีเรียซึ่งการเริ่มต้นการแปลเกิดขึ้นทันทีที่ปลาย 5' ของ mRNA ถูกสังเคราะห์ ในยูคาริโอต การเชื่อมโยงที่แน่นแฟ้นระหว่างการถอดรหัสและการแปลนั้นเป็นไปไม่ได้ เพราะการถอดรหัสและการแปลเกิดขึ้นในส่วนที่แยกจากกันของเซลล์ ( นิวเคลียสและไซโตพลาสซึม ) สารตั้งต้น mRNA ของยูคาริโอตจะต้องได้รับการประมวลผลในนิวเคลียส (เช่น การเติมหมวกการเติมโพลีอะดีนีนการตัดต่อ) ในไรโบโซมก่อนที่จะถูกส่งออกไปยังไซโตพลาสซึมเพื่อการแปล การแปลยังอาจได้รับผลกระทบจากการหยุดชั่วคราวของไรโบโซม ซึ่งสามารถกระตุ้นการโจมตีของเอนโดนิวคลีโอไลติกต่อ tRNA ซึ่งเป็นกระบวนการที่เรียกว่าการสลายตัวของ mRNA ที่ไม่เกิดขึ้น การหยุดชั่วคราวของไรโบโซมยังช่วยในการพับตัวของพอลิเปปไทด์ที่เกิดขึ้นใหม่บนไรโบโซมระหว่างการแปล และทำให้การแปลโปรตีนล่าช้าในขณะที่กำลังเข้ารหัส tRNA ซึ่งสามารถกระตุ้นการเลื่อนเฟรมของไรโบโซมได้[ 22 ]
tRNA ตัวสุดท้ายที่ได้รับการตรวจสอบโดยหน่วยย่อยไรโบโซมขนาดเล็ก ( การปรับตัว ) จะถ่ายโอนกรดอะมิโนไปยังหน่วยย่อยไรโบโซมขนาดใหญ่ซึ่งจะจับกับ tRNA ตัวก่อนหน้า ( การถ่ายโอนหมู่เปปไทด์ ) จากนั้นไรโบโซมจะเคลื่อนไปยังรหัส mRNA ถัดไปเพื่อดำเนินการต่อ ( การเคลื่อนย้าย ) ทำให้เกิดสายโซ่กรดอะมิโนขึ้น
ในการแปลรหัสทางพันธุกรรมของแบคทีเรียและอาร์เคียการแปลรหัสเกิดขึ้นในไซโตซอล ซึ่งไรโบโซมจะจับกับเอ็มอาร์เอ็นเอ ในยูคาริ โอต การแปลรหัสสามารถเกิดขึ้นได้ทั้งในไซโตพลาสซึมและข้ามเยื่อหุ้มของเอนโดพลาสมิกเรติคูลัมผ่านกระบวนการที่เรียกว่าการเคลื่อนย้ายร่วมกับการแปลรหัส (co-translational translocation ) ในการเคลื่อนย้ายร่วมกับการแปลรหัส คอมเพล็กซ์ไรโบโซม-เอ็มอาร์เอ็นเอทั้งหมดจะจับกับเยื่อหุ้มชั้นนอกของเอนโดพลาสมิกเรติคูลัมแบบหยาบ (ER) และโปรตีนใหม่จะถูกสังเคราะห์และปล่อยเข้าไปใน ER โพลีเปปไทด์ที่สร้างขึ้นใหม่สามารถถูกหลั่งออกไป ทันทีหรือเก็บไว้ภายใน ER เพื่อการขนส่ง โดยเวสิเคิลและการหลั่งออกนอกเซลล์ ในอนาคต
RNA หลายชนิดที่ถูกถอดรหัสเช่น tRNA, ribosomal RNA และ small nuclear RNA จะไม่ถูกแปลเป็นโปรตีน
ยาปฏิชีวนะหลายชนิดออกฤทธิ์โดยการยับยั้งการสังเคราะห์โปรตีน ได้แก่อะนิโซไมซินไซโคลเฮกซิไม ด์ คลอแรม เฟนิ คอล เตตราไซค ลิ น สเตรปโต ไม ซิ น อิริโทรไมซิ นและพูโรไม ซิน ไรโบโซมของโปรคาริโอตมีโครงสร้างแตกต่างจากไรโบโซมของยูคาริโอต ดังนั้นยาปฏิชีวนะจึงสามารถออกฤทธิ์จำเพาะต่อการติดเชื้อ แบคทีเรียได้ โดยไม่ทำลายเซลล์ของ โฮสต์ ยูคาริโอต
การเลิกจ้าง
การยุติการยืดตัวขึ้นอยู่กับปัจจัยการปลดปล่อยeRF1ซึ่งจดจำรหัสหยุดทั้งสามตัว เมื่อถึงรหัสหยุด การยุติของพอลิเปปไทด์จะเกิดขึ้น ไรโบโซมจะถูกแยกส่วน และพอลิเปปไทด์ที่สมบูรณ์จะถูกปล่อยออกมาeRF3เป็น GTPase ที่ขึ้นอยู่กับไรโบโซมซึ่งช่วยให้ eRF1 ปล่อยพอลิเปปไทด์ที่สมบูรณ์ออกมา จีโนมของมนุษย์เข้ารหัสยีนบางส่วนที่มีรหัสหยุด mRNA ที่รั่วไหลอย่างน่าประหลาดใจ: ในยีนเหล่านี้ การยุติการแปลไม่มีประสิทธิภาพเนื่องจากเบส RNA พิเศษในบริเวณใกล้เคียงกับรหัสหยุด การยุติที่รั่วไหลในยีนเหล่านี้ทำให้เกิดการอ่านผ่านการแปลได้มากถึง 10% ของรหัสหยุดของยีนเหล่านี้ ยีนบางส่วนเหล่านี้เข้ารหัสโดเมนโปรตีน ที่ใช้งานได้ในส่วนขยายการอ่านผ่านเพื่อให้ ไอโซฟอร์มโปรตีนใหม่สามารถเกิดขึ้นได้ กระบวนการนี้เรียกว่า 'การอ่านผ่านการแปลที่ใช้งานได้' [ 23 ]
เมื่อไซต์ A ของไรโบโซมถูกครอบครองโดยรหัสหยุด (UAA, UAG หรือ UGA) บน mRNA ทำให้เกิดโครงสร้างหลักของโปรตีน tRNA มักจะไม่สามารถจดจำหรือจับกับรหัสหยุดได้ แต่รหัสหยุดจะกระตุ้นการจับของโปรตีนปัจจัยปลดปล่อย[ 24 ] (RF1 และ RF2) ซึ่งกระตุ้นการแยกส่วนของคอมเพล็กซ์ไรโบโซม/mRNA ทั้งหมดโดยการไฮโดรไลซิสของสายโพลีเปปไทด์จากศูนย์เปปทิดิลทรานสเฟอเรส[ 2 ]ของไรโบโซม[ 25 ] ยาหรือลำดับโมทีฟพิเศษบน mRNA สามารถเปลี่ยนโครงสร้างของไรโบโซมเพื่อให้ tRNA ที่ใกล้เคียงจับกับรหัสหยุดแทนที่จะเป็นปัจจัยปลดปล่อย ในกรณีดังกล่าวของ 'การอ่านผ่านการแปล' การแปลจะดำเนินต่อไปจนกว่าไรโบโซมจะพบรหัสหยุดถัดไป[ 23 ]
การรีไซเคิล
เมื่อโปรตีนที่สังเคราะห์ถูกปล่อยออกมา ไรโบโซมที่ซับซ้อนจะแยกตัวออกเป็นหน่วยย่อยเพื่อนำกลับมาใช้ใหม่[ 11 ]
ข้อผิดพลาดในการแปล
แม้ว่าโดยทั่วไปแล้วไรโบโซมจะถูกพิจารณาว่าเป็นเครื่องจักรที่แม่นยำและต่อเนื่อง แต่กระบวนการแปลรหัสก็อาจเกิดข้อผิดพลาดได้ ซึ่งอาจนำไปสู่การสังเคราะห์โปรตีนที่ผิดพลาดหรือการยุติการแปลรหัสก่อนกำหนด ไม่ว่าจะเป็นเพราะ tRNA จับคู่กับโคดอนที่ผิด หรือเพราะ tRNA จับคู่กับกรดอะมิโนที่ผิด[ 26 ]อัตราข้อผิดพลาดในการสังเคราะห์โปรตีนได้รับการประมาณไว้ที่ระหว่าง 1 ใน 10⁵ ถึง 1 ใน 10³ ของกรดอะมิโนที่ใส่ผิด ขึ้นอยู่กับเงื่อนไขการทดลอง[ 27 ]ในทางกลับกัน อัตราการยุติการแปลรหัสก่อนกำหนดได้รับการประมาณไว้ที่ระดับ 10⁻⁴ เหตุการณ์ต่อโคดอนที่แปลรหัส[ 28 ] [ 29 ]
ระเบียบข้อบังคับ
การแปลรหัสพันธุกรรมเป็นหนึ่งในกระบวนการที่ใช้พลังงานมากที่สุดในเซลล์ ดังนั้นจึงมีการควบคุมอย่างเข้มงวด กลไกมากมายได้วิวัฒนาการขึ้นเพื่อควบคุมและกำกับการแปลรหัสพันธุกรรมทั้งในยูคาริโอตและโปรคาริโอตการควบคุมการแปลรหัสพันธุกรรมสามารถส่งผลกระทบต่ออัตราโดยรวมของการสังเคราะห์โปรตีน ซึ่งมีความสัมพันธ์อย่างใกล้ชิดกับสภาวะการเผาผลาญและการเพิ่มจำนวนของเซลล์
เพื่อศึกษาถึงกระบวนการนี้ นักวิทยาศาสตร์ได้ใช้วิธีการที่หลากหลาย เช่น ชีววิทยาเชิงโครงสร้าง เคมีวิเคราะห์ (โดยใช้แมสสเปกโทรเมตรี) การถ่ายภาพการแปล mRNA ของรีพอร์เตอร์ (ซึ่งการแปล mRNA เชื่อมโยงกับผลลัพธ์ เช่น การเรืองแสงหรือการเปล่งแสง) การตรวจจับผ่านการรวมกรดอะมิโนกัมมันตรังสี และวิธีการที่ใช้การจัดลำดับรุ่นต่อไป[ 30 ] นอกจากนี้ยังสามารถใช้ วิธีการอื่นๆ เช่นการทดสอบ toeprintingเพื่อกำหนดตำแหน่งของไรโบโซมของ mRNA เฉพาะในหลอดทดลอง และร่องรอยของโปรตีนอื่นๆ ที่ควบคุมการแปล เพื่อเจาะลึกเข้าไปในกระบวนการที่ซับซ้อนนี้ นักวิทยาศาสตร์มักใช้เทคนิคที่เรียกว่าการทำโปรไฟล์ไรโบโซม[ 31 ]วิธีนี้ช่วยให้นักวิจัยสามารถถ่ายภาพทรานสเลโทม ซึ่งแสดงให้เห็นว่าส่วนใดของ mRNA กำลังถูกแปลเป็นโปรตีนโดยไรโบโซมในเวลาใดเวลาหนึ่ง การทำโปรไฟล์ไรโบโซมให้ข้อมูลเชิงลึกที่มีค่าเกี่ยวกับพลวัตของการแปล เผยให้เห็นปฏิสัมพันธ์ที่ซับซ้อนระหว่างลำดับยีน โครงสร้าง mRNA และการควบคุมการแปล[ 18 ]การขยายแนวคิดนี้ การวิเคราะห์โปรไฟล์ไรโบโซมระดับเซลล์เดี่ยว เป็นเทคนิคที่ช่วยให้สามารถศึกษาขั้นตอนการแปลที่ความละเอียดระดับเซลล์แต่ละเซลล์ได้[ 32 ]การวิเคราะห์โปรไฟล์ไรโบโซมระดับเซลล์เดี่ยวได้เปิดเผยว่าความแตกต่างทางพันธุกรรมและการแสดงออกของ mRNA ที่เกิดขึ้นตามมานั้นสามารถส่งผลกระทบต่ออัตราการแปลในลักษณะเฉพาะของ RNA ได้เช่นกัน การวิเคราะห์โปรไฟล์ไรโบโซมระดับเซลล์เดี่ยวมีศักยภาพที่จะช่วยให้เข้าใจธรรมชาติที่ไม่เป็นเนื้อเดียวกันของเซลล์ได้ดียิ่งขึ้น นำไปสู่ความเข้าใจที่ละเอียดอ่อนยิ่งขึ้นว่าการควบคุมการแปลสามารถส่งผลกระทบต่อพฤติกรรมของเซลล์ สถานะการเผาผลาญ และการตอบสนองต่อสิ่งเร้าหรือสภาวะต่างๆ ได้อย่างไร
การแทนที่กรดอะมิโน
ในบางเซลล์กรดอะมิโน บางชนิด อาจพร่องไปและส่งผลต่อประสิทธิภาพการแปล ตัวอย่างเช่นเซลล์ T ที่ถูกกระตุ้น จะหลั่งอินเตอร์เฟรอน-γซึ่งกระตุ้นให้เกิด การขาดแคลน ทริปโตเฟน ภายในเซลล์ โดยการเพิ่มการทำงานของ เอนไซม์ อินโดลามีน 2,3-ไดออกซิเจเนส 1 (IDO1) แม้ว่าทริปโตเฟนจะพร่องไป แต่การสังเคราะห์โปรตีนแบบ in-frame ยังคงดำเนินต่อไปได้แม้จะมีรหัสทริป โตเฟน อยู่ โดยทำได้โดยการใส่ฟีนิลอะลานีนแทนทริปโตเฟน เปปไทด์ที่ได้เรียกว่า "สารทดแทน W>F" สารทดแทน W>F เหล่านี้มีจำนวนมากในมะเร็ง บาง ชนิดและมีความเกี่ยวข้องกับการแสดงออกของ IDO1 ที่เพิ่มขึ้น ในทางฟังก์ชัน สารทดแทน W>F สามารถทำให้การทำงานของโปรตีน บกพร่องได้ [ 33 ]
ความสำคัญทางคลินิก
การควบคุมการแปลมีความสำคัญต่อการพัฒนาและการอยู่รอดของมะเร็งเซลล์มะเร็งมักจะต้องควบคุมขั้นตอนการแปลของการแสดงออกของยีน แม้ว่าจะยังไม่เข้าใจอย่างถ่องแท้ว่าเหตุใดการแปลจึงถูกกำหนดเป้าหมายมากกว่าขั้นตอนอื่นๆ เช่นการถอดรหัสในขณะที่เซลล์มะเร็งมักจะมีปัจจัยการแปลที่เปลี่ยนแปลงทางพันธุกรรม แต่โดยทั่วไปแล้วเซลล์มะเร็งมักจะปรับเปลี่ยนระดับของปัจจัยการแปลที่มีอยู่[ 34 ]เส้นทางการส่งสัญญาณมะเร็งที่สำคัญหลายเส้นทาง รวมถึงเส้นทางRAS–MAPK , PI3K/AKT/mTOR , MYC และWNT–β-catenin จะทำการตั้งโปรแกรม จีโนม ใหม่ ผ่านการแปล[ 35 ]เซลล์มะเร็งยังควบคุมการแปลเพื่อปรับตัวให้เข้ากับความเครียดของเซลล์ ในระหว่างความเครียด เซลล์จะแปล mRNA ที่สามารถบรรเทาความเครียดและส่งเสริมการอยู่รอด ตัวอย่างเช่น การแสดงออกของAMPKในมะเร็งหลายชนิด การกระตุ้นของมันจะกระตุ้นกระบวนการต่อเนื่องที่ในที่สุดจะช่วยให้มะเร็งหลีกเลี่ยง การตายของเซลล์แบบโปรแกรม ( apoptosis ) ที่เกิดจากการขาดสารอาหารได้ การบำบัดมะเร็งในอนาคตอาจเกี่ยวข้องกับการขัดขวางกลไกการแปลของเซลล์เพื่อต่อต้านผลกระทบที่เกิดขึ้นภายหลังของมะเร็ง[ 34 ]
การสร้างแบบจำลองทางคณิตศาสตร์ของการแปล
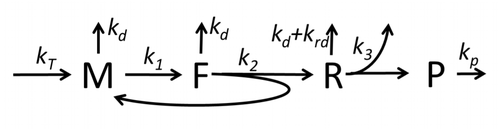
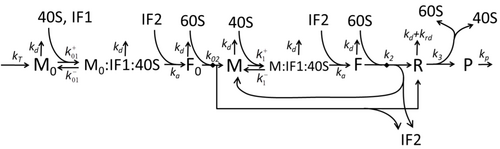
คำอธิบายกระบวนการถอดเสียงและแปลความหมาย โดยกล่าวถึงเฉพาะกระบวนการพื้นฐานที่สุด ประกอบด้วย:
- การผลิตโมเลกุล mRNA (รวมถึงกระบวนการตัดต่อ)
- การเริ่มต้นของโมเลกุลเหล่านี้ด้วยความช่วยเหลือของปัจจัยเริ่มต้น (เช่น การเริ่มต้นอาจรวมถึงขั้นตอนการสร้างวงกลม แม้ว่าจะไม่จำเป็นเสมอไปก็ตาม)
- การเริ่มต้นการแปลรหัสพันธุกรรม โดยการดึงหน่วยย่อยไรโบโซมขนาดเล็กเข้ามา
- การประกอบไรโบโซมให้สมบูรณ์
- การยืดตัว (เช่น การเคลื่อนที่ของไรโบโซมไปตาม mRNA พร้อมกับการสร้างโปรตีน)
- การสิ้นสุดการแปล
- การเสื่อมสภาพของโมเลกุล mRNA
- การสลายตัวของโปรตีน
กระบวนการสร้างกรดอะมิโนเพื่อสร้างโปรตีนในการแปลรหัสเป็นหัวข้อของแบบจำลองทางฟิสิกส์ต่างๆ มาเป็นเวลานาน โดยเริ่มจากแบบจำลองจลนศาสตร์โดยละเอียดแบบแรก เช่น[ 37 ]หรือแบบจำลองอื่นๆ ที่คำนึงถึงแง่มุมสุ่มของการแปลรหัสและใช้การจำลองด้วยคอมพิวเตอร์ แบบจำลองการสังเคราะห์โปรตีนที่อิงตามจลนศาสตร์ทางเคมีจำนวนมากได้รับการพัฒนาและวิเคราะห์ในช่วงสี่ทศวรรษที่ผ่านมา[ 38 ] [ 39 ]นอกเหนือจากจลนศาสตร์ทางเคมีแล้ว รูปแบบการสร้างแบบจำลองต่างๆ เช่นTotally Asymmetric Simple Exclusion Process [ 39 ] Probabilistic Boolean Networks , Petri Netsและmax-plus algebra ได้ถูกนำมาใช้เพื่อสร้างแบบจำลองจลนศาสตร์โดยละเอียดของการ สังเคราะห์โปรตีนหรือบางขั้นตอน แบบจำลองพื้นฐานของการสังเคราะห์โปรตีนที่คำนึงถึงกระบวนการ 'พื้นฐาน' ทั้งแปดได้รับการพัฒนา[ 36 ]ตามแบบแผนที่ว่า " แบบจำลองที่ มีประโยชน์ นั้นเรียบง่ายและขยายได้" [ 40 ]แบบจำลองที่ง่ายที่สุดM0แสดงโดยกลไกจลนศาสตร์ปฏิกิริยา (รูป M0) โดยทั่วไปจะรวมถึง 40S, 60S และปัจจัยเริ่มต้น (IF) ที่จับกัน (รูปที่ M1') และขยายเพิ่มเติมเพื่อรวมผลของไมโครอาร์เอ็นเอต่อการสังเคราะห์โปรตีน[ 41 ]โมเดลส่วนใหญ่ในลำดับชั้นนี้สามารถแก้ได้ด้วยวิธีวิเคราะห์ โซลูชันเหล่านี้ใช้เพื่อดึง 'ลายเซ็นจลนศาสตร์' ของกลไกเฉพาะต่างๆ ของการควบคุมการสังเคราะห์
รหัสพันธุกรรม
นอกจากนี้ยังสามารถแปลได้ทั้งด้วยมือ (สำหรับลำดับสั้นๆ) หรือด้วยคอมพิวเตอร์ (หลังจากตั้งโปรแกรมให้เหมาะสมก่อน ดูหัวข้อด้านล่าง) ซึ่งช่วยให้นักชีววิทยาและนักเคมีสามารถวาดลำดับกรดอะมิโนหลักของโปรตีนที่เข้ารหัสลงบนกระดาษได้
ขั้นแรก แปลงเบส DNA แต่ละตัวให้เป็นเบส RNA ที่เป็นคู่ตรงข้าม (โปรดทราบว่าคู่ตรงข้ามของ A คือ U) ดังแสดงด้านล่าง โปรดสังเกตว่าสาย DNA ที่เป็นแม่แบบคือสายที่ใช้ในการสร้าง RNA ส่วนสาย DNA อีกสายหนึ่งจะเหมือนกับ RNA แต่มีไทมีนแทนยูราซิล
DNA -> RNA A -> U ที -> เอ ซี -> จี จี -> ซี A=T-> A=U
จากนั้นแบ่ง RNA ออกเป็นกลุ่มสามเบส (กลุ่มละสามเบส) โปรดสังเกตว่ามี "หน้าต่าง" การแปลหรือเฟรมการอ่าน 3 เฟรม ขึ้นอยู่กับว่าคุณเริ่มอ่านรหัสจากจุดใด สุดท้าย ใช้ตารางในหัวข้อ รหัสพันธุกรรมเพื่อแปลงข้อมูลข้างต้นเป็นสูตรโครงสร้างตามที่ใช้ในวิชาเคมี
วิธีนี้จะให้โครงสร้างขั้นต้นของโปรตีน อย่างไรก็ตามโปรตีนมักจะพับตัว โดยขึ้นอยู่กับส่วนที่ชอบน้ำและไม่ชอบน้ำตามสายโซ่โครงสร้างขั้นที่สองมักจะยังพอคาดเดาได้ แต่โครงสร้างขั้นที่สาม ที่ถูกต้องนั้น มักจะยากมากที่จะระบุได้ เพื่อให้ได้โครงสร้างสามมิติที่แม่นยำและการปฏิสัมพันธ์ของอะตอม จึง มีการใช้ ชีววิทยาโครงสร้างและวิธี การทางชีวฟิสิกส์ อื่นๆ อีกหลายวิธี
ในขณะที่ลักษณะอื่นๆ เช่น โครงสร้างสามมิติ หรือที่เรียกว่าโครงสร้างระดับตติยภูมิ ของโปรตีนนั้น สามารถทำนายได้โดยใช้อัลกอริทึมที่ซับซ้อนเท่านั้น แต่ลำดับกรดอะมิโน หรือที่เรียกว่าโครงสร้างระดับปฐมภูมิ สามารถกำหนดได้จากลำดับกรดนิวคลีอิกเพียงอย่างเดียว โดยใช้ตารางการแปลงค่าช่วย
วิธีการนี้อาจไม่ได้ให้องค์ประกอบของกรดอะมิโนที่ถูกต้องของโปรตีน โดยเฉพาะอย่างยิ่งหาก มีการนำ กรดอะมิโน ที่ไม่เป็นไปตามแบบแผน เช่นซีลีโนซิสเทอีนเข้ามาในโปรตีน ซึ่งถูกกำหนดโดยรหัสหยุดแบบปกติร่วมกับลำดับแฮร์พินที่อยู่ถัดไป (ลำดับการแทรกซีลีโนซิสเทอีน หรือ SECIS)
มีโปรแกรมคอมพิวเตอร์มากมายที่สามารถแปลงลำดับดีเอ็นเอ/อาร์เอ็นเอไปเป็นลำดับโปรตีนได้ โดยปกติแล้วจะใช้รหัสพันธุกรรมมาตรฐาน แต่มีโปรแกรมเพียงไม่กี่โปรแกรมที่สามารถจัดการกับกรณี "พิเศษ" ทั้งหมดได้ เช่น การใช้รหัสเริ่มต้นทางเลือกซึ่งมีความสำคัญทางชีววิทยา ตัวอย่างเช่น รหัสเริ่มต้นทางเลือกที่หายากอย่าง CTG จะเข้ารหัสโปรตีนเมไทโอนีนเมื่อใช้เป็นรหัสเริ่มต้น และเข้ารหัสโปรตีนลิวซีนในตำแหน่งอื่นๆ ทั้งหมด
ตัวอย่าง: ตารางการแปลแบบย่อสำหรับรหัสพันธุกรรมมาตรฐาน (จากเว็บเพจ NCBI Taxonomy) [ 42 ]
AAs = FFLLSSSSYY**CC*WLLLLPPPPHHQQRRRRIIIMTTTTNNKKSSRRVVVVAAAADDEEGGGG เริ่ม = ---M---------------M---------------M---------------------------- ฐาน1 = TTTTTTTTTTTTTTTTCCCCCCCCCCCCCCCCAAAAAAAAAAAAAAAAGGGGGGGGGGGGGGGG Base2 = TTTTCCCCAAAAGGGGTTTTCCCCAAAAGGGGTTTTCCCCAAAAGGGGTTTTCCCCAAAAGGGG ฐาน 3 = TCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAG
แถว "Starts" แสดงรหัสเริ่มต้นสามแบบ ได้แก่ UUG, CUG และ AUG ซึ่งพบได้บ่อยมาก นอกจากนี้ยังแสดงกรดอะมิโนตัวแรกเมื่อตีความว่าเป็นรหัสเริ่มต้น ซึ่งในกรณีนี้คือเมไทโอนีนทั้งหมด
ตารางการแปล
แม้แต่เมื่อทำงานกับลำดับยูคาริโอตทั่วไป เช่น จีโนม ของยีสต์ก็มักมีความต้องการที่จะสามารถใช้ตารางการแปลทางเลือกได้ โดยเฉพาะอย่างยิ่งสำหรับการแปลยีนไมโทคอนเดรีย ปัจจุบันตารางการแปลต่อไปนี้ได้รับการกำหนดโดย กลุ่มอนุกรมวิธาน ของ NCBIสำหรับการแปลลำดับในGenBank : [ 42 ]
- รหัสมาตรฐาน
- รหัสไมโตคอนเดรียของสัตว์มีกระดูกสันหลัง
- รหัสไมโตคอนเดรียของยีสต์
- รหัส ไมโทคอนเดรียของรา โปรโตซัว และสัตว์กลุ่มซีลันเทอเรต และรหัสของไมโคพลาสมา/สไปโรพลาสมา
- รหัสไมโตคอนเดรียของสัตว์ไม่มีกระดูกสันหลัง
- รหัสนิวเคลียร์ ของซีลิเอต ดาซิคลาเดเซียน และเฮกซามิตา
- รหัสคิเนโตพลาสต์
- รหัสไมโทคอนเดรียของสัตว์ทะเลกลุ่มเอ คิโนเดอร์มและหนอนแบน
- รหัสนิวเคลียร์ของยูโพลติด
- รหัสพลาสติด ของแบคทีเรีย อาร์เคีย และพืช
- รหัสนิวเคลียร์ทางเลือกของยีสต์
- รหัสไมโตคอนเดรียของแอสซิเดียน
- รหัสไมโทคอนเดรียทางเลือกของหนอนแบน
- รหัสนิวเคลียร์ของBlepharisma
- รหัสไมโทคอนเดรีย ของคลอโรไฟเซียน
- รหัส ไมโทคอนเดรียของพยาธิใบไม้
- รหัสไมโตคอนเดรียของScenedesmus obliquus
- รหัสไมโทคอนเดรียของThraustochytrium
- รหัสไมโทคอนเดรีย ของPterobranchia
- กลุ่มผู้สมัคร SR1 และรหัสแบคทีเรีย Gracilibacteria
- รหัสนิวเคลียร์ของPachysolen tannophilus
- รหัสนิวเคลียร์ที่ไร้ประโยชน์
- รหัสนิวเคลียร์คอนดิลอสโตมา
- รหัสนิวเคลียร์เมโซดิเนียม
- รหัสนิวเคลียร์เพริทริช
- รหัสนิวเคลียร์ของบลาสโตคริธิเดีย
- รหัสไมโทคอนเดรียของ Cephalodiscidae
ดูเพิ่มเติม
อ่านเพิ่มเติม
- Champe PC, Harvey RA, Ferrier DR (2004). Lippincott's Illustrated Reviews: Biochemistry (ฉบับที่ 3). Hagerstwon, MD: Lippincott Williams & Wilkins. ISBN 0-7817-2265-9.
- Cox M, Nelson DR, Lehninger AL (2005). หลักการชีวเคมีของเลห์นิงเกอร์ (ฉบับที่ 4). ซานฟรานซิสโก...: WH Freeman. ISBN 0-7167-4339-6.
- Malys N, McCarthy JE (มีนาคม 2011). "การเริ่มต้นการแปล: สามารถคาดการณ์ความแปรผันในกลไกได้" . Cellular and Molecular Life Sciences . 68 (6): 991– 1003. doi : 10.1007/s00018-010-0588-z . PMC 11115079 . PMID 21076851 . S2CID 31720000 .
ลิงก์ภายนอก
- คอลเลกชันแอนิเมชั่นเซลล์เสมือนจริง: ขอแนะนำการแปล
- เครื่องมือแปล (จากลำดับดีเอ็นเอหรืออาร์เอ็นเอ)
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การแปล (ชีววิทยา)
การแปลรหัสพันธุกรรม เป็นกระบวนการใน เซลล์ชีวภาพ ที่ สร้าง โปรตีน โดย ใช้ โมเลกุล RNA เป็นแม่แบบ โปรตีนที่สร้างขึ้นเป็นลำดับของ กรดอะมิโน ที่กำหนดโดยลำดับของ นิวคลีโอไทด์ ใน RNA...
กลไกพื้นฐาน
กระบวนการพื้นฐานของ การผลิตโปรตีน คือการเพิ่มกรดอะมิโนทีละตัวที่ปลายของสายโพลีเปปไทด์ที่กำลังก่อตัว การดำเนินการนี้ทำโดย ไร โบ โซม [ 2 ] ไรโบโซมประกอบด้วยสองหน่วยย่อย ในยูคาริโอตจะมีหน่วยย่อย ขนาดเล็ก (40S) และ หน่วยย่อยขนาดใหญ่ (60S)...
เวที
การแปลดำเนินไปในสี่ขั้นตอน ได้แก่ การเริ่มต้น การยืดออก การสิ้นสุด และการรีไซเคิล [ 11 ]
การเริ่มต้น
การเริ่มต้นเกี่ยวข้องกับหน่วยย่อยขนาดเล็กของไรโบโซมที่จับกับปลาย 5' ของ mRNA โดยอาศัย ปัจจัยการเริ่มต้น [ 12 ] ไร โบโซมและปัจจัยที่เกี่ยวข้องจะประกอบกันและจับกับ mRNA tRNA ตัวแรกจะถูกยึดติดที่ โคดอนเริ่มต้น กระบวนการนี้ถูกกำหนดให้เป็นแบบขึ้นอยู่กับแคป...