อ่าน 24 นาที
ซอโรโปดา
ซอโรโพดา ( / s ɔː ˈ r ɒ p ə d ə / ) ซึ่งสมาชิกเรียกว่า ซอโรพอด ( / ˈ s ɔːr ə p ɒ d z / ; [ 1 ] [ 2 ] มาจาก sauro- + -pod ; แปลตรงตัวว่า ' เท้า เหมือน กิ้งก่า ' ) เป็น กลุ่ม ของ...
ซอโรโปดา
| ซอโรพอด ช่วงเวลา: ต้นยุคจูราสสิก – ปลายยุคครีเทเชียส ( ซิเนมูเรียน – มาสทริชเชียน (อาจพบในช่วงปลายยุคไทรแอสสิก )) | |||||
|---|---|---|---|---|---|
 | |||||
| ไดโนเสาร์ซอโรพอด 6 ชนิด (จากซ้ายบนไปขวาล่าง): พาตาโกไททัน , บาโรซอรัส, จิราฟฟาไททัน, โอเมอิซอรัส, ชูโนซอรัสและอามาร์กาซอรัส | |||||
| การจำแนกทางวิทยาศาสตร์ | |||||
| อาณาจักร: | แอนิมอลเลีย | ||||
| ไฟลัม: | คอร์ดาต้า | ||||
| ระดับ: | สัตว์เลื้อยคลาน | ||||
| กลุ่มสายพันธุ์ : | ไดโนเสาร์ | ||||
| กลุ่มสายพันธุ์ : | ซอริสเชีย | ||||
| กลุ่มสายพันธุ์ : | † ซอโรโพโดมอร์ฟา | ||||
| กลุ่มสายพันธุ์ : | † ซอโรโพดิฟอร์ม | ||||
| กลุ่มสายพันธุ์ : | † บึงซอโรโพดา , 1878 | ||||
| กลุ่มย่อย | |||||
| |||||
| คำพ้องความหมาย | |||||
| |||||
ซอโรโพดา ( / s ɔː ˈ r ɒ p ə d ə / )ซึ่งสมาชิกเรียกว่าซอโรพอด ( / ˈ s ɔːr ə p ɒ d z / ; [ 1 ] [ 2 ]มาจากsauro- + -pod ; แปลตรงตัวว่า' เท้าเหมือน กิ้งก่า ' ) เป็นกลุ่มของไดโนเสาร์ซอริสเชียน ('สะโพกเหมือนกิ้งก่า') ซอโรพอดมีคอยาวมาก หางยาว หัวเล็ก (เมื่อเทียบกับส่วนอื่นๆ ของร่างกาย) และขาหนาเหมือนเสา 4 ขา พวกมันโดดเด่นในเรื่องขนาดมหึมาที่บางสายพันธุ์มี และกลุ่มนี้รวมถึงสัตว์ที่ใหญ่ที่สุดที่เคยอาศัยอยู่บนบก สกุลที่รู้จักกันดีได้แก่อลาโมซอรัส อะพาโทซอรัส อาร์เจนติ โนซอรัส บรา คิ โอ ซอรัส บ รอน โท ซอ รัส คามา ราซอรัส ดิโพล โด คั สเดรดนอตัสและมาเมนคิซอรัส[ 3 ] [ 4 ]
ไดโนเสาร์ซอโรพอดที่เก่าแก่ที่สุดที่ทราบแน่ชัดนั้นมาจาก ยุคจูราสสิ กตอนต้น[ 5 ] IsanosaurusและAntetonitrusเดิมทีถูกอธิบายว่าเป็นซอโรพอดในยุคไทรแอสสิก[ 6 ] [ 7 ]แต่อายุของพวกมัน และในกรณีของAntetonitrusสถานะซอโรพอดของมันก็ถูกตั้งคำถามในภายหลัง[ 8 ] [ 5 ] [ 9 ] อย่างไรก็ตาม รอยเท้าซอโรโพโดมอร์ฟที่คล้ายซอโรพอดจากFleming Fjord Formation ( กรีนแลนด์ ) อาจบ่งชี้ถึงการปรากฏตัวของกลุ่มนี้ในยุคไทรแอสสิกตอนปลาย[ 5 ]ในช่วงปลายยุคจูราสสิก (150 ล้านปีก่อน) ซอโรพอดได้แพร่กระจายไปทั่ว (โดยเฉพาะดิพลอโดซิดและแบรคิโอซอริเด ) ใน ช่วงปลายยุค ครีเทเชียสกลุ่มซอโรพอดกลุ่มหนึ่งคือไททาโนซอร์ได้เข้ามาแทนที่กลุ่มอื่น ๆ ทั้งหมดและมีการกระจายตัวเกือบจะทั่วโลก อย่างไรก็ตาม เช่นเดียวกับไดโนเสาร์ที่ไม่ใช่นกชนิดอื่นๆ ที่มีชีวิตอยู่ในช่วงเวลานั้น ไททาโนซอร์ก็สูญพันธุ์ไปในเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในยุคครีเทเชียส-พาลีโอจีน ซากดึกดำบรรพ์ของซอโรพอดถูกพบในทุกทวีป รวมถึงทวีปแอนตาร์กติกา[ 10 ] [ 11 ] [ 12 ] [ 13 ]
ชื่อซอโรพอด (Sauropoda) ถูกตั้งขึ้นโดยโอธเนียล ชาร์ลส์ มาร์ชในปี พ.ศ. 2321 และมาจากภาษากรีกโบราณซึ่งหมายถึง "เท้าของกิ้งก่า" [ 14 ]ซอโรพอดเป็นหนึ่งในกลุ่มไดโนเสาร์ที่รู้จักกันดีที่สุด และกลายเป็นส่วนสำคัญของวัฒนธรรมสมัยนิยมเนื่องจากขนาดที่ใหญ่โตของพวกมัน
การค้นพบฟอสซิลซอโรพอดที่สมบูรณ์นั้นหายากมาก หลายชนิด โดยเฉพาะอย่างยิ่งชนิดที่ใหญ่ที่สุด รู้จักกันเพียงจากกระดูกที่แยกออกจากกันเท่านั้น ตัวอย่างที่เกือบสมบูรณ์หลายชิ้นขาดหัว ปลายหาง และแขนขา
คำอธิบาย
ซอโรพอดเป็นสัตว์ กินพืช มัก เป็นสัตว์สี่ขาที่มีคอยาว[ 15 ] และมักมีฟันรูปช้อน (รูปทรงคล้ายไม้พาย: กว้างที่ปลาย แคบที่คอ) พวกมันมีหัวค่อนข้างเล็ก ลำตัวใหญ่โต และส่วนใหญ่มีหางยาว ขาหลังของพวกมันหนา ตรง และแข็งแรง ปลายขาเป็นเท้าคล้ายกระบองที่มีห้านิ้ว แม้ว่าจะมีเพียงสามนิ้วด้านใน (หรือในบางกรณีสี่นิ้ว) เท่านั้นที่มีกรงเล็บ ขาหน้าของพวกมันค่อนข้างเรียวและโดยทั่วไปปลายขาเป็นมือคล้ายเสาที่สร้างขึ้นเพื่อรองรับน้ำหนัก มักจะมีเพียงนิ้วหัวแม่มือเท่านั้นที่มีกรงเล็บ ภาพประกอบของซอโรพอดจำนวนมากขาดข้อเท็จจริงเหล่านี้ โดยแสดงภาพซอโรพอดอย่างไม่ถูกต้องว่ามีกีบหุ้มปลายนิ้วเท้าที่ไม่มีกรงเล็บ หรือมีกรงเล็บหรือกีบมากกว่าสามอันบนมือ กระดูกสันหลังส่วนหางส่วนต้นเป็นลักษณะเฉพาะของซอโรพอ ดอย่างยิ่ง [ 16 ]
ขนาด

ลักษณะเด่นที่สุดของซอโรพอดคือขนาดของมัน แม้แต่ซอโรพอดแคระ (อาจยาว 5 ถึง 6 เมตร (16 ถึง 20 ฟุต)) ก็ยังถูกนับว่าเป็นสัตว์ที่ใหญ่ที่สุดในระบบนิเวศ ของพวก มัน คู่แข่งที่แท้จริงในแง่ของขนาดคือโรร์ควอลเช่นวาฬสีน้ำเงิน แต่ต่างจากวาฬ ซอโรพอด เป็น สัตว์บกเป็นหลัก
โครงสร้างร่างกายของพวกมันไม่ได้แตกต่างกันมากนักเมื่อเทียบกับไดโนเสาร์ชนิดอื่น อาจเนื่องมาจากข้อจำกัดด้านขนาด แต่พวกมันก็แสดงให้เห็นถึงความหลากหลายอย่างมาก บางชนิด เช่นไดโพลโดซิดมีหางยาวมาก ซึ่งพวกมันอาจสามารถสะบัดเหมือนแส้เพื่อส่งสัญญาณหรือเพื่อขับไล่หรือทำร้ายผู้ล่า[ 17 ]หรือเพื่อสร้างเสียงระเบิด [ 18 ] [ 19 ] ซูเปอร์ซอรัสซึ่งมีความยาว 33 ถึง 34 เมตร (108 ถึง 112 ฟุต) [ 20 ]เป็นซอโรพอดที่ยาวที่สุดเท่าที่รู้จักจากซากที่ค่อนข้างสมบูรณ์ แต่ตัวอื่นๆ เช่น ไดโพลโดคัส ซึ่ง เป็นเจ้าของสถิติเดิม ก็มีความยาวมากเช่น กัน กระดูกสันหลังต้นแบบ (และปัจจุบันสูญหายไปแล้ว) ของ แอม ฟิโคเอเลียส แฟรกิลลิมัส (ปัจจุบันคือมาราปูนิซอรัส ) อาจมาจากสัตว์ที่มีความยาว 58 เมตร (190 ฟุต) [ 21 ]กระดูกสันหลังของมันจะยาวกว่าของวาฬสีน้ำเงินอย่างมาก อย่างไรก็ตาม งานวิจัยที่ตีพิมพ์ในปี 2015 คาดการณ์ว่าการประมาณขนาดของA. fragillimusอาจถูกประเมินสูงเกินไป[ 22 ]ไดโนเสาร์ที่ยาวที่สุดที่รู้จักจากฟอสซิลที่น่าเชื่อถือคือArgentinosaurus huinculensisโดยมีความยาวประมาณ 35 ถึง 36 เมตร (115 ถึง 118 ฟุต) ตามงานวิจัยล่าสุด[ 23 ] [ 24 ]อย่างไรก็ตาม ตัวอย่าง Barosaurus ยักษ์ BYU 9024 อาจมีขนาดใหญ่กว่านั้น โดยมีความยาวถึง 45–48 เมตร (148–157 ฟุต) [ 23 ] [ 24 ] [ 25 ] ไดโนเสาร์ ชนิดอื่นๆ เช่นbrachiosauridsมีความสูงมาก มีไหล่สูงและคอยาวมาก ซอโรพอดที่สูงที่สุดคือ ตัวอย่าง Barosaurus ยักษ์ ที่มีความสูง 22 เมตร (72 ฟุต) [ 23 ]เมื่อเปรียบเทียบกันแล้วยีราฟซึ่งเป็นสัตว์บกที่มีความสูงที่สุดในบรรดาสัตว์ที่มีชีวิตทั้งหมด มีความสูงเพียง 4.8 ถึง 5.6 เมตร (16 ถึง 18 ฟุต)
หลักฐานที่ดีที่สุดบ่งชี้ว่าไดโนเสาร์ที่มีขนาดใหญ่ที่สุดคือArgentinosaurus (65 ถึง 80 เมตริกตัน (72 ถึง 88 ตันสั้น) [ 26 ] [ 23 ] [ 24 ] ), Mamenchisaurus sinocanadorum (60 ถึง 80 เมตริกตัน หรือ 66 ถึง 88 ตันสั้น[ 24 ] ), ตัวอย่าง Barosaurus ยักษ์ (60-80+ เมตริกตัน[ 23 ] [ 24 ] [ 25 ] ) และPatagotitanกับPuertasaurus (50 ถึง 55 เมตริกตัน (49 ถึง 54 ตันยาว; 55 ถึง 61 ตันสั้น) [ 23 ] [ 24 ] ) ในขณะเดียวกัน 'เมกะซอโรพอด' เช่นบรูฮัทคาโยซอรัสได้รับการตรวจสอบมาเป็นเวลานานเนื่องจากการถกเถียงที่ขัดแย้งเกี่ยวกับความถูกต้อง แต่ภาพถ่ายล่าสุดที่ปรากฏขึ้นอีกครั้งในปี 2022 ได้ทำให้มันถูกต้องตามกฎหมาย[ 27 ]ทำให้สามารถประมาณการที่ทันสมัยมากขึ้นซึ่งอยู่ในช่วงระหว่าง 110 ถึง 170 เมตริกตัน (120 ถึง 190 ตันสั้น) ซึ่งมีขนาด เทียบเท่ากับ วาฬสีน้ำเงิน[ 28 ]น้ำหนักของAmphicoelias fragillimusถูกประมาณไว้ที่ 122.4 เมตริกตัน (120.5 ลองตัน; 134.9 ชอร์ตตัน) โดยมีความยาวสูงสุดเกือบ 60 เมตร (200 ฟุต) [ 21 ]แต่การวิจัยในปี 2015 โต้แย้งว่าการประมาณค่าเหล่านี้อิงจากไดโพลโดซิดมากกว่าเรบบาชิซอริเดที่ทันสมัยกว่า ซึ่งชี้ให้เห็นถึงความยาวที่สั้นกว่ามากที่ 35–40 เมตร (115–130 ฟุต) โดยมีมวลระหว่าง 80 ถึง 120 เมตริกตัน (79 ถึง 118 ลองตัน; 88 ถึง 132 ชอร์ตตัน) [ 22 ]การค้นพบเพิ่มเติมบ่งชี้ว่ามีหลายสายพันธุ์ที่อาจมีน้ำหนักถึงหรือเกิน 40 ตัน[ 29 ]สัตว์บกที่ใหญ่ที่สุดที่ยังมีชีวิตอยู่ในปัจจุบันคือช้างป่าซึ่งมีน้ำหนักไม่เกิน 10.4 เมตริกตัน (11.5 ชอร์ตตัน) [ 30 ]
ในบรรดาซอโรพอดที่เล็กที่สุด ได้แก่โอห์มเดโนซอรัส (ยาว 4 เมตร หรือ 13 ฟุต) ไททาโนซอรัส แคระ แมก ยาโรซอรัส (ยาว 6 เมตร หรือ 20 ฟุต) และบราคิโอซอริเดแคระยูโรพาซอรัสซึ่งมีความยาว 6.2 เมตร (20 ฟุต) เมื่อโตเต็มวัย[ 31 ]รูปร่างเล็กของมันน่าจะเป็นผลมาจากภาวะแคระแกร็นที่เกิดขึ้นในประชากรซอโรพอดที่แยกตัวอยู่บนเกาะในช่วงปลายยุคจูราสสิก ในพื้นที่ที่ปัจจุบันคือบริเวณลังเกนเบิร์กทางตอนเหนือของเยอรมนี [ 32 ] [ 33 ]บ ราคี ทราเคโลแพน ซอ โรพอดในกลุ่ม ไดโพลโดคอยด์ เป็นสมาชิกที่สั้นที่สุดในกลุ่มเนื่องจากคอที่สั้นผิดปกติ แตกต่างจากซอโรพอดอื่นๆ ที่คอสามารถยาวได้ถึงสี่เท่าของความยาวหลัง คอของบราคีทราเคโลแพน นั้น สั้นกว่ากระดูกสันหลัง
ฟอสซิลของไดโนเสาร์ที่อาจใหญ่ที่สุดเท่าที่เคยพบ (MOZ-Pv 1221) ถูกค้นพบในปี 2021 ในจังหวัดเนวเกน ทางตะวันตกเฉียงเหนือของปาตาโกเนีย ประเทศอาร์เจนตินา เชื่อกันว่าฟอสซิลเหล่านั้นมาจากไททาโนซอร์ ซึ่งเป็นหนึ่งในซอโรพอดที่ใหญ่ที่สุด[ 34 ] [ 29 ]
ในวันที่ 29 มีนาคม 2017 หรือก่อนหน้านั้นไม่นาน มีการค้นพบรอยเท้าซอโรพอดที่มีความยาวประมาณ 1.7 เมตร (5.6 ฟุต) ที่วาลมาดานีในภูมิภาคคิมเบอร์ลีย์ของรัฐเวสเทิร์นออสเตรเลีย[ 35 ]รายงานระบุว่าเป็นรอยเท้าที่ใหญ่ที่สุดเท่าที่เคยพบมา ในปี 2020 โมลินา-เปเรซและลาร์ราเมนดีประเมินขนาดของสัตว์ไว้ที่ 31 เมตร (102 ฟุต) และ 72 ตัน (79.4 ตันสั้น) โดยอิงจากรอยเท้าที่มีความยาว 1.75 เมตร (5.7 ฟุต) [ 23 ]
แขนขาและเท้า
ในฐานะสัตว์สี่ขา ขนาดใหญ่ ซอโรพอดได้พัฒนาแขนขาที่ "รับน้ำหนัก" (graviportal) พิเศษ เท้าหลังกว้างและยังคงมีกรงเล็บสามอันในสายพันธุ์ส่วนใหญ่[ 36 ]สิ่งที่ผิดปกติเป็นพิเศษเมื่อเทียบกับสัตว์อื่นๆ คือเท้าหน้า ( manus ) ที่ได้รับการดัดแปลงอย่างมาก เท้าหน้าของซอโรพอดนั้นแตกต่างจากเท้าหน้าของสัตว์สี่ขาขนาดใหญ่ในปัจจุบัน เช่นช้าง อย่างมาก แทนที่จะกางออกไปด้านข้างเพื่อสร้างเท้าที่กว้างเหมือนในช้าง กระดูก มือของซอโรพอดจะเรียงตัวเป็นคอลัมน์แนวตั้งอย่างสมบูรณ์ โดยมีกระดูกนิ้วที่ลดขนาดลงอย่างมาก (แม้ว่าจะไม่ชัดเจนว่าซอโรพอดที่ดั้งเดิมที่สุด เช่นวัลคาโนดอนและบาราปาซอรัสมีเท้าหน้าแบบนี้หรือไม่) [ 37 ]เท้าหน้าของยูซอโร พอดได้รับการดัดแปลงมาก จนนิ้วแต่ละนิ้วจะไม่สามารถมองเห็นได้ในขณะที่ยังมีชีวิตอยู่
การจัดเรียงของกระดูกฝ่าเท้าส่วนหน้า ( กระดูกฝ่ามือ ) ในยูซอโรพอดมีลักษณะเป็นรูปครึ่งวงกลม ดังนั้นรอยเท้าส่วนหน้าของซอโรพอดจึงมีรูปร่างคล้ายเกือกม้า แตกต่างจากช้าง หลักฐานรอยเท้าแสดงให้เห็นว่าซอโรพอดไม่มีแผ่นรองเนื้อนุ่มรองรับฝ่าเท้าส่วนหน้า ทำให้ฝ่าเท้าเว้า[ 37 ]กรงเล็บเดียวที่มองเห็นได้ในซอโรพอดส่วนใหญ่คือกรงเล็บนิ้วหัวแม่มือที่โดดเด่น (เกี่ยวข้องกับนิ้วที่ 1) ซอโรพอดเกือบทั้งหมดมีกรงเล็บดังกล่าว แม้ว่าจะไม่ทราบว่ามันทำหน้าที่อะไร กรงเล็บมีขนาดใหญ่ที่สุด (รวมถึงสูงและแบนด้านข้าง) ในดิพลอโดซิด และมีขนาดเล็กมากในบราคิโอซอริเด ซึ่งบางชนิดดูเหมือนจะสูญเสียกรงเล็บไปโดยสิ้นเชิงจากหลักฐานรอยเท้า[ 38 ]ไททาโนซอร์อาจสูญเสียกรงเล็บนิ้วหัวแม่มือไปโดยสิ้นเชิง (ยกเว้นรูปแบบในยุคแรก เช่นJanenschia )
ไททาโนซอร์เป็นสัตว์ที่ผิดปกติที่สุดในบรรดาซอโรพอด เนื่องจากตลอดประวัติศาสตร์ของกลุ่ม พวกมันไม่เพียงแต่สูญเสียกรงเล็บภายนอกเท่านั้น แต่ยังสูญเสียนิ้วเท้าด้านหน้าไปทั้งหมดด้วย ไททาโนซอร์ขั้นสูงไม่มีนิ้วหรือกระดูกนิ้ว และเดินได้เฉพาะบน "ตอ" รูปเกือกม้าที่ประกอบขึ้นจากกระดูกฝ่ามือรูปทรงกระบอก[ 39 ]
หลักฐานการพิมพ์จากโปรตุเกสแสดงให้เห็นว่า อย่างน้อยในซอโรพอดบางชนิด (น่าจะเป็นบราคิโอซอริเด) ส่วนล่างและด้านข้างของกระดูกฝ่าเท้าส่วนหน้าอาจถูกปกคลุมด้วยเกล็ดเล็กๆ ที่มีหนาม ซึ่งทิ้งร่องรอยไว้ในรอยพิมพ์[ 40 ]ในไททาโนซอร์ ปลายกระดูกฝ่ามือที่สัมผัสกับพื้นนั้นกว้างและเป็นรูปสี่เหลี่ยมจัตุรัสผิดปกติ และตัวอย่างบางชิ้นยังคงรักษาเนื้อเยื่ออ่อนที่ปกคลุมบริเวณนี้ไว้ ซึ่งบ่งชี้ว่าเท้าส่วนหน้าของสายพันธุ์เหล่านี้มีขอบที่บุด้วยวัสดุรองรับบางชนิด[ 39 ]
Matthew Bonnan [ 41 ] [ 42 ]ได้แสดงให้เห็นว่ากระดูกยาวของไดโนเสาร์ซอโรพอดเติบโตแบบไอโซเมตริกนั่นคือ รูปร่างแทบไม่เปลี่ยนแปลงเลยเมื่อซอโรพอดวัยเยาว์เติบโตเป็นตัวเต็มวัยขนาดมหึมา Bonnan แนะนำว่ารูปแบบการปรับขนาดที่แปลกประหลาดนี้ (สัตว์มีกระดูกสันหลังส่วนใหญ่แสดงการเปลี่ยนแปลงรูปร่างอย่างมีนัยสำคัญในกระดูกยาวที่เกี่ยวข้องกับการรองรับน้ำหนักที่เพิ่มขึ้น) อาจเกี่ยวข้องกับหลักการเดินบนไม้ค้ำ (ที่เสนอโดยนักวิทยาศาสตร์สมัครเล่น Jim Schmidt) ซึ่งขาที่ยาวของซอโรพอดตัวเต็มวัยช่วยให้พวกมันสามารถเดินทางได้ไกลอย่างง่ายดายโดยไม่ต้องเปลี่ยนกลไกโดยรวม
ถุงลม
เช่นเดียวกับ ไดโนเสาร์ซอริส เชีย นอื่นๆ (เช่นเทโรพอดรวมถึงนก) ซอโรพอดมีระบบถุงลมซึ่งเห็นได้จากรอยบุ๋มและโพรงกลวงในกระดูกสันหลัง ส่วนใหญ่ ที่ถูกรุกรานโดยถุงลม กระดูกกลวงที่มี อากาศ เป็นลักษณะเฉพาะของซอโรพอดทั้งหมด[ 43 ]ช่องว่างอากาศเหล่านี้ช่วยลดน้ำหนักโดยรวมของคอขนาดใหญ่ที่ซอโรพอดมี และระบบถุงลมโดยทั่วไปที่ช่วยให้มีการไหลของอากาศไปในทิศทางเดียวผ่านปอดที่แข็ง ทำให้ซอโรพอดได้รับออกซิเจนเพียงพอ[ 44 ]การปรับตัวนี้จะเป็นประโยชน์ต่อซอโรพอดโดยเฉพาะอย่างยิ่งในสภาวะที่มีออกซิเจนค่อนข้างต่ำในยุคจูราสสิกและยุคครีเทเชียสตอนต้น[ 45 ]
ลักษณะกลวงคล้ายนกของกระดูกซอโรพอดได้รับการยอมรับตั้งแต่ช่วงแรกของการศึกษาเกี่ยวกับสัตว์เหล่านี้ และในความเป็นจริง ตัวอย่างซอโรพอดอย่างน้อยหนึ่งตัวอย่างที่พบในศตวรรษที่ 19 ( Ornithopsis ) เดิมทีถูกระบุผิดว่าเป็นเทโรซอร์ (สัตว์เลื้อยคลานบินได้) เนื่องจากลักษณะนี้[ 46 ]
เกราะ

ซอโรพอดบางชนิดมีเกราะป้องกันมีบางสกุลที่มีกระบอง ขนาดเล็ก อยู่ที่หาง ตัวอย่างที่โดดเด่นคือชูโนซอรัสและไททาโนซอรัส หลายชนิด เช่นซัลตาซอรัสและแอมเพโลซอรัสมีแผ่น กระดูกเล็กๆ ปกคลุมบางส่วนของร่างกาย
ฟัน
การศึกษาวิจัยโดย Michael D'Emic และเพื่อนร่วมงานจากมหาวิทยาลัย Stony Brook พบว่าซอโรพอดมีวิวัฒนาการอัตราการเปลี่ยนฟันที่สูงเพื่อรองรับความอยากอาหารจำนวนมาก การศึกษาวิจัยชี้ให้เห็นว่าNigersaurusเปลี่ยนฟันแต่ละซี่ทุกๆ 14 วันCamarasaurusเปลี่ยนฟันแต่ละซี่ทุกๆ 62 วัน และDiplodocusเปลี่ยนฟันแต่ละซี่ทุกๆ 35 วัน[ 47 ]นักวิทยาศาสตร์พบว่าคุณสมบัติของฟันมีผลต่อระยะเวลาที่ฟันซี่ใหม่จะงอกฟันของCamarasaurus ใช้เวลานานในการงอกกว่าฟันของ Diplodocusเนื่องจากมีขนาดใหญ่กว่า[ 48 ]
นอกจากนี้ D'Emic และทีมงานของเขายังตั้งข้อสังเกตว่าความแตกต่างระหว่างฟันของซอโรพอดยังบ่งชี้ถึงความแตกต่างในด้านอาหาร ด้วย ไดโพลโดคั ส กินพืชที่อยู่ใกล้พื้นดิน ในขณะที่คามาราซอรัสกินใบไม้จากกิ่งบนและกิ่งกลาง ตามที่นักวิทยาศาสตร์กล่าว การแบ่งอาหารเฉพาะด้านช่วยให้ไดโนเสาร์กินพืชชนิดต่างๆ สามารถอยู่ร่วมกันได้[ 47 ] [ 48 ]
คอ
คอของซอโรพอดพบว่ามีความยาวมากกว่า 15 เมตร (49 ฟุต) ซึ่งยาวกว่าคอของยีราฟที่ยาวที่สุดในโลกถึงหกเท่า[ 44 ]ปัจจัยสำคัญหลายประการที่ทำให้คอยาวได้ขนาดนี้ ได้แก่ ขนาดตัวที่ใหญ่และท่าทางการเดินสี่ขาของไดโนเสาร์ ซึ่งเป็นฐานที่มั่นคงในการรองรับคอ และหัวของพวกมันก็วิวัฒนาการให้มีขนาดเล็กและเบามาก ทำให้สูญเสียความสามารถในการย่อยอาหารทางปาก การลดขนาดหัวให้เป็นเพียงเครื่องมือเก็บเกี่ยวพืชเข้าสู่ร่างกาย ทำให้ซอโรพอดต้องการพลังงานน้อยลงในการยกหัว จึงสามารถพัฒนาคอที่มีกล้ามเนื้อและเนื้อเยื่อเกี่ยวพันหนาแน่นน้อยลง ซึ่งช่วยลดมวลโดยรวมของคอลงอย่างมาก ทำให้สามารถยืดคอให้ยาวขึ้นได้อีก
ไดโนเสาร์ซอโรพอดมีโครงสร้างกระดูกที่ปรับตัวได้ดีมาก บางชนิดมี กระดูกสันหลังส่วนคอมากถึง 19 ชิ้นในขณะที่สัตว์เลี้ยงลูกด้วยนมเกือบทั้งหมดมีเพียง 7 ชิ้นเท่านั้น นอกจากนี้ กระดูกสันหลังแต่ละชิ้นยังยาวมากและมีช่องว่างจำนวนมากซึ่งเต็มไปด้วยอากาศ ระบบถุงลมที่เชื่อมต่อกับช่องว่างเหล่านี้ไม่เพียงแต่ช่วยลดน้ำหนักของคอที่ยาว แต่ยังช่วยเพิ่มการไหลเวียนของอากาศผ่านหลอดลม ช่วยให้พวกมันหายใจเอาอากาศได้เพียงพอ การวิวัฒนาการของกระดูกสันหลังที่ประกอบด้วยอากาศถึง 60% ทำให้ซอโรพอดสามารถลดปริมาณกระดูกที่หนาแน่นและหนักลงได้โดยไม่สูญเสียความสามารถในการหายใจเข้าลึกๆ เพื่อส่งออกซิเจนไปเลี้ยงทั่วร่างกาย[ 44 ]ตามที่ Kent Stevens กล่าว การสร้างแบบจำลองโครงกระดูกด้วยคอมพิวเตอร์จากกระดูกสันหลังแสดงให้เห็นว่าคอของซอโรพอดสามารถกวาดพื้นที่หาอาหารขนาดใหญ่ได้โดยไม่ต้องขยับตัว แต่ไม่สามารถหดกลับไปอยู่ในตำแหน่งที่สูงกว่าไหล่มากเพื่อสำรวจพื้นที่หรือเอื้อมไปถึงที่สูงขึ้นได้[ 49 ]
หน้าที่อีกอย่างหนึ่งที่เสนอไว้สำหรับคอยาวของซอโรพอดก็คือ ทำหน้าที่เป็นหม้อน้ำเพื่อรับมือกับความร้อนจำนวนมหาศาลที่เกิดจากมวลร่างกายขนาดใหญ่ของพวกมัน เมื่อพิจารณาว่ากระบวนการเผาผลาญจะต้องทำงานหนักมาก มันจึงย่อมสร้างความร้อนจำนวนมากเช่นกัน และการกำจัดความร้อนส่วนเกินนี้จึงเป็นสิ่งจำเป็นต่อการอยู่รอด[ 50 ] นอกจากนี้ยังมีการเสนอว่าคอยาวจะช่วยระบายความร้อนให้กับเส้นเลือดและหลอดเลือดแดงที่ไปเลี้ยงสมอง ป้องกันไม่ให้เลือดที่ร้อนเกินไปไปถึงศีรษะ อันที่จริงพบว่าอัตราการเผาผลาญที่เพิ่มขึ้นอันเป็นผลมาจากคอของซอโรพอดนั้นได้รับการชดเชยมากกว่าเล็กน้อยจากพื้นที่ผิวที่เพิ่มขึ้นซึ่งสามารถระบายความร้อนได้[ 51 ]
บรรพชีววิทยา
นิเวศวิทยา
การวิเคราะห์พื้นผิวการสึกหรอของฟัน (DMTA) ที่ดำเนินการกับซอโรพอดไททาโนซอริฟอร์มจากชั้นหินทามากาวะในยุคทูโรเนียน แสดงให้เห็นว่าซอโรพอดกินพืชที่อ่อนนุ่มกว่าเปลือกนอกของแมลงหรือเปลือกหอย โดยอาหารน่าจะประกอบด้วยเฟิร์นและพืชเมล็ดเปลือย ผลลัพธ์ DMTA ยังชี้ให้เห็นว่าซอโรพอดน่าจะเคี้ยวอาหารอย่างมีพลังมากกว่าเลพิโดซอร์ใน ปัจจุบัน [ 52 ]
เมื่อมีการค้นพบซอโรพอดเป็นครั้งแรก ขนาดอันใหญ่โตมโหฬารของพวกมันทำให้เหล่านักวิทยาศาสตร์หลายคนเปรียบเทียบพวกมันกับ วาฬในปัจจุบันงานวิจัยส่วนใหญ่ในช่วงศตวรรษที่ 19 และต้นศตวรรษที่ 20 สรุปว่าซอโรพอดมีขนาดใหญ่เกินกว่าจะทรงตัวบนบกได้ ดังนั้นพวกมันจึงต้องอาศัยอยู่ในน้ำเป็นหลักภาพวาดซอโรพอดส่วนใหญ่ในช่วงสามในสี่แรกของศตวรรษที่ 20 แสดงให้เห็นพวกมันจมอยู่ในน้ำทั้งหมดหรือบางส่วน[ 53 ]แนวคิดในยุคแรกนี้ถูกตั้งข้อสงสัยตั้งแต่ช่วงทศวรรษที่ 1950 เมื่อการศึกษาของ Kermack (1951) แสดงให้เห็นว่า หากสัตว์จมอยู่ในน้ำลึกหลายเมตร แรงดันจะมากพอที่จะทำให้ปอดและทางเดินหายใจยุบตัวจนถึงแก่ความตายได้[ 54 ]อย่างไรก็ตาม การศึกษาเกี่ยวกับนิเวศวิทยา ของซอโรพอดในยุคแรกนี้และงานวิจัยอื่นๆ มีข้อบกพร่องตรงที่ละเลยหลักฐานจำนวนมากที่แสดงให้เห็นว่าร่างกายของซอโรพอ ดมีถุงลม จำนวนมาก ในปี ค.ศ. 1878 นักบรรพชีวินวิทยา อี.ดี. โคป เคยเรียกโครงสร้างเหล่านี้ว่า "แผ่นลอย"
ตั้งแต่ช่วงทศวรรษ 1970 เป็นต้นมา ผลกระทบของถุงลมของซอโรพอดต่อวิถีชีวิตในน้ำที่คาดการณ์ไว้ของพวกมันเริ่มได้รับการสำรวจ นักบรรพชีวินวิทยา เช่น Coombs และ Bakker ใช้สิ่งนี้ รวมถึงหลักฐานจากตะกอนวิทยาและชีวกลศาสตร์เพื่อแสดงให้เห็นว่าซอโรพอดเป็นสัตว์บกเป็นหลัก ในปี 2004 DM Henderson ตั้งข้อสังเกตว่า เนื่องจากระบบถุงลมที่กว้างขวาง ซอโรพอดจึงลอยตัวได้และไม่สามารถจุ่มลำตัวลงไปใต้น้ำได้ทั้งหมด กล่าวอีกนัยหนึ่งคือ พวกมันจะลอยตัว และจะไม่ตกอยู่ในอันตรายจากการยุบตัวของปอดเนื่องจากแรงดันน้ำขณะว่ายน้ำ[ 53 ]
หลักฐานการว่ายน้ำของซอโรพอดมาจากร่องรอยฟอสซิลที่บางครั้งพบว่าเหลือเพียงรอยเท้าหน้า (manus) เท่านั้น เฮนเดอร์สันแสดงให้เห็นว่าร่องรอยดังกล่าวสามารถอธิบายได้ด้วยซอโรพอดที่มีแขนขาหน้ายาว (เช่นมาโครนาเรียน ) ที่ลอยอยู่ในน้ำตื้นพอสมควรซึ่งลึกพอที่จะทำให้ขาหลังที่สั้นกว่าไม่สัมผัสกับพื้น และใช้แขนขาหน้าในการพายไปข้างหน้า[ 53 ]อย่างไรก็ตาม เนื่องจากสัดส่วนของร่างกาย ซอโรพอดที่ลอยอยู่จึงไม่เสถียรและปรับตัวได้ไม่ดีสำหรับการอยู่ในน้ำเป็นเวลานาน รูปแบบการเคลื่อนที่ ในน้ำนี้ ประกอบกับความไม่เสถียร ทำให้เฮนเดอร์สันเรียกซอโรพอดในน้ำว่า "นักพายที่มึนงง" [ 53 ]
ดังนั้น แม้ว่าซอโรพอดจะไม่สามารถอาศัยอยู่ในน้ำได้ตามที่ปรากฏในภาพวาดทางประวัติศาสตร์ แต่ก็มีหลักฐานว่าพวกมันชอบที่อยู่อาศัยที่ชื้นแฉะและชายฝั่ง รอยเท้าของซอโรพอดมักพบตามแนวชายฝั่งหรือข้ามที่ราบน้ำท่วมถึง และฟอสซิลของซอโรพอดมักพบในสภาพแวดล้อมที่ชื้นแฉะหรือปะปนอยู่กับฟอสซิลของสิ่งมีชีวิตในทะเล[ 53 ]ตัวอย่างที่ดีของเรื่องนี้คือรอยเท้าขนาดใหญ่ ของ ซอโรพอดในยุคจูราสสิกที่พบใน แหล่งสะสม ของทะเลสาบในเกาะสกายของสกอตแลนด์[ 55 ]การศึกษาที่ตีพิมพ์ในปี 2021 ชี้ให้เห็นว่าซอโรพอดไม่สามารถอาศัยอยู่ในภูมิภาคขั้วโลกได้ การศึกษานี้ชี้ให้เห็นว่าพวกมันส่วนใหญ่ถูกจำกัดอยู่ในพื้นที่เขตร้อนและมีกระบวนการเผาผลาญที่แตกต่างจากไดโนเสาร์ชนิดอื่นมาก อาจอยู่ระหว่างสัตว์เลี้ยงลูกด้วยนมและสัตว์เลื้อยคลาน[ 56 ]การศึกษาใหม่ที่ตีพิมพ์โดย Taia Wyenberg-henzler ในปี 2022 ชี้ให้เห็นว่าซอโรพอดในอเมริกาเหนือลดลงเนื่องจากสาเหตุที่ไม่ทราบแน่ชัดเกี่ยวกับแหล่งที่อยู่อาศัยและการกระจายตัวในช่วงปลายยุคจูราสสิกและต้นยุคครีเทเชียส สาเหตุนี้ยังไม่ชัดเจน แต่มีการเสนอแนะว่ามีความคล้ายคลึงกันในแหล่งอาหารระหว่างอิกัวโนดอนเชียนฮาโดรซอรอยด์และซอโรพอด และอาจส่งผลให้เกิดการแข่งขันกัน อย่างไรก็ตาม สิ่งนี้ไม่สามารถอธิบายการลดลงของการกระจายตัวของซอโรพอดได้อย่างสมบูรณ์ เนื่องจากหากเกิดการกีดกันในการแข่งขัน จะทำให้การลดลงนั้นรวดเร็วกว่าที่ปรากฏในบันทึกฟอสซิลมาก ยิ่งไปกว่านั้น ต้องพิจารณาว่าการลดลงของซอโรพอดในอเมริกาเหนือเป็นผลมาจากการเปลี่ยนแปลงของพืชที่ซอโรพอดชอบกิน สภาพภูมิอากาศ หรือปัจจัยอื่นๆ หรือไม่ งานวิจัยเดียวกันนี้ยังเสนอแนะว่าอิกัวโนดอนเชียนและฮาโดรซอรอยด์ได้ใช้ประโยชน์จากแหล่งอาหารที่ว่างลงเมื่อไม่นานมานี้จากการลดลงของความหลากหลายของซอโรพอดในช่วงปลายยุคจูราสสิกและยุคครีเทเชียสในอเมริกาเหนือ[ 57 ]
การต้อนสัตว์และการดูแลลูก
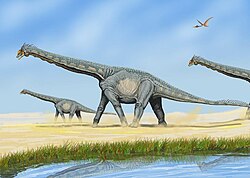
หลักฐานฟอสซิลจำนวนมาก ทั้งจากแหล่งกระดูกและรอยเท้า บ่งชี้ว่าซอโรพอดเป็นสัตว์ที่อยู่รวมกันเป็นฝูงอย่างไรก็ตาม องค์ประกอบของฝูงนั้นแตกต่างกันไปในแต่ละชนิด แหล่งกระดูกบางแห่ง เช่น แหล่งจากยุคจูราสสิกตอนกลางของอาร์เจนตินาดูเหมือนจะแสดงให้เห็นฝูงที่ประกอบด้วยสมาชิกที่มีอายุต่างกัน ปะปนกันระหว่างลูกอ่อนและตัวเต็มวัย อย่างไรก็ตาม แหล่งฟอสซิลและรอยเท้าอื่นๆ อีกหลายแห่งบ่งชี้ว่าซอโรพอดหลายชนิดเดินทางเป็นฝูงที่แยกตามอายุ โดยลูกอ่อนจะรวมฝูงแยกจากตัวเต็มวัย กลยุทธ์การรวมฝูงแบบแยกส่วนดังกล่าวพบได้ในสายพันธุ์ต่างๆ เช่นอลาโมซอรัสเบลลูซอรัสและไดโพลโดซิดบาง ชนิด [ 58 ]
ในการทบทวนหลักฐานเกี่ยวกับฝูงประเภทต่างๆ Myers และ Fiorillo พยายามอธิบายว่าเหตุใดซอโรพอดจึงมักรวมฝูงกันแยกกัน การศึกษาการสึกหรอของฟันในระดับจุลภาคแสดงให้เห็นว่าซอโรพอดวัยเยาว์มีอาหารที่แตกต่างจากซอโรพอดวัยผู้ใหญ่ ดังนั้นการรวมฝูงจึงไม่เกิดประโยชน์เท่ากับการรวมฝูงแยกกัน ซึ่งสมาชิกในฝูงแต่ละตัวสามารถหาอาหารได้อย่างประสานงานกัน ความแตกต่างของขนาดที่มากระหว่างวัยเยาว์และวัยผู้ใหญ่ก็อาจมีส่วนทำให้กลยุทธ์การหาอาหารและการรวมฝูงแตกต่างกันด้วย[ 58 ]

เนื่องจากการแยกตัวของลูกอ่อนและตัวเต็มวัยต้องเกิดขึ้นไม่นานหลังจากฟักไข่ และเมื่อรวมกับข้อเท็จจริงที่ว่าลูกซอโรพอดส่วนใหญ่น่าจะสามารถ ช่วยเหลือตัวเองได้ ตั้งแต่แรกเกิด Myers และ Fiorillo จึงสรุปว่าสายพันธุ์ที่มีฝูงแยกตามอายุจะไม่แสดงการดูแลจากพ่อแม่มากนัก[ 58 ]ในทางกลับกัน นักวิทยาศาสตร์ที่ศึกษาฝูงซอโรพอดที่มีอายุต่างกันได้แนะนำว่าสายพันธุ์เหล่านี้อาจดูแลลูกอ่อนเป็นเวลานานก่อนที่จะโตเต็มวัย[ 59 ]การศึกษาในปี 2014 ชี้ให้เห็นว่าระยะเวลาตั้งแต่การวางไข่จนถึงเวลาฟักไข่น่าจะอยู่ระหว่าง 65 ถึง 82 วัน[ 60 ]ยังไม่เป็นที่ทราบแน่ชัดว่าการแยกฝูงกับการรวมฝูงที่มีอายุต่างกันนั้นแตกต่างกันอย่างไรในกลุ่มซอโรพอดต่างๆ จำเป็นต้องค้นพบตัวอย่างเพิ่มเติมของพฤติกรรมการรวมฝูงจากสายพันธุ์ซอโรพอดอื่นๆ เพื่อตรวจหาแบบแผนการกระจายตัวที่เป็นไปได้[ 58 ]
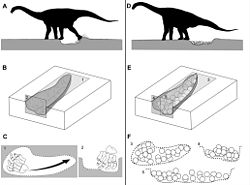
แหล่งทำรังหลายแห่งที่ค้นพบในอาร์เจนตินาและอินเดียมีไข่ฟอสซิลจำนวน 30-400 รังที่ได้รับการอนุรักษ์ไว้ ซึ่งเป็นหลักฐานแสดงถึงการดูแลลูกของแม่ซอโรพอด นักวิจัยแนะนำว่าซอโรพอดอาจตั้งถิ่นฐานในพื้นที่ทำรังใกล้กับกิจกรรมภูเขาไฟเพื่อการฟักไข่ด้วยความร้อนใต้พิภพ ซึ่งแม่จะรักษาความอบอุ่นให้ไข่ พฤติกรรมนี้คล้ายกับนกและสัตว์เลื้อยคลานในปัจจุบันที่ใช้วิธีเดียวกัน[ 61 ] [ 62 ] [ 63 ]
ท่าถีบ

ตั้งแต่ช่วงต้นประวัติศาสตร์ของการศึกษา นักวิทยาศาสตร์ เช่นออสบอร์นได้ตั้งสมมติฐานว่าซอโรพอดสามารถยืนขึ้นบนขาหลังได้ โดยใช้หางเป็น 'ขา' ที่สามของขาตั้งสามขา[ 64 ]โครงกระดูกจำลองของBarosaurus lentus ซึ่งเป็นไดโพลโด ซิด ที่ยืนขึ้นบนขาหลัง ณพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติอเมริกันเป็นตัวอย่างหนึ่งของสมมติฐานนี้ ในบทความปี 2005 รอธไชลด์และโมลนาร์ให้เหตุผลว่า หากซอโรพอดเคยยืนด้วยสองขาในบางครั้ง จะต้องมีหลักฐานของรอยแตกจากความเครียดใน 'มือ' ของแขนขาหน้า อย่างไรก็ตาม ไม่พบร่องรอยดังกล่าวหลังจากที่พวกเขาตรวจสอบโครงกระดูกซอโรพอดจำนวนมาก[ 65 ]

ไฮน์ริช มัลลิสัน (ในปี 2009) เป็นคนแรกที่ศึกษาศักยภาพทางกายภาพของซอโรพอดชนิดต่างๆ ในการยืนด้วยขาหลังสามขา มัลลิสันพบว่าลักษณะบางอย่างที่เคยเชื่อมโยงกับการปรับตัวในการยืนด้วยขาหลังนั้น แท้จริงแล้วไม่เกี่ยวข้อง (เช่น กระดูกสะโพกที่อยู่ห่างกันของไททาโนซอร์ ) หรืออาจเป็นอุปสรรคต่อการยืนด้วยขาหลัง ตัวอย่างเช่น ไททาโนซอร์มีกระดูกสันหลังที่ยืดหยุ่นผิดปกติ ซึ่งจะลดความมั่นคงในท่าทางแบบสามขาและจะทำให้กล้ามเนื้อต้องรับภาระมากขึ้น ในทำนองเดียวกัน เป็นไปได้ยากที่บราคิโอซอริเดจะยืนด้วยขาหลังได้ เนื่องจากจุดศูนย์ถ่วงของพวกมันอยู่ไปข้างหน้ามากกว่าซอโรพอดชนิดอื่นๆ ซึ่งจะทำให้ท่าทางดังกล่าวไม่มั่นคง[ 66 ]
ในทางกลับกัน ไดโพลโดซิดดูเหมือนจะปรับตัวได้ดีสำหรับการยืนสองขา ไดโพลโดซิดมีจุดศูนย์กลางมวลอยู่เหนือสะโพกโดยตรง ทำให้พวกมันทรงตัวได้ดีขึ้นบนสองขา ไดโพลโดซิดยังมีคอที่เคลื่อนไหวได้มากที่สุดในบรรดาซอโรพอด มีกระดูกเชิงกรานที่แข็งแรง และกระดูกสันหลังส่วนหางที่มีรูปร่างพิเศษที่ช่วยให้หางรับน้ำหนักได้ ณ จุดที่สัมผัสพื้น มัลลิสันสรุปว่าไดโพลโดซิดปรับตัวได้ดีกว่าช้าง ในการยืนสองขา ซึ่งช้างก็ทำเช่นนั้นเป็นครั้งคราวในป่า เขายังโต้แย้งว่ากระดูกหักจากความเครียดในป่าไม่ได้เกิดขึ้นจากพฤติกรรมในชีวิตประจำวัน[ 66 ]เช่น กิจกรรมที่เกี่ยวข้องกับการกินอาหาร (ขัดแย้งกับรอธส์ไชลด์และโมลนาร์) [ 65 ]
ท่าทางศีรษะและลำคอ

ยังไม่มีข้อสรุปที่แน่ชัดเกี่ยวกับวิธีการที่ไดโนเสาร์ซอโรพอดใช้ในการยกหัวและคอ รวมถึงท่าทางต่างๆ ที่พวกมันสามารถทำได้ในขณะมีชีวิต
มีการตั้งคำถามว่าคอยาวของซอโรพอดสามารถใช้ในการเล็มกินต้นไม้สูงได้หรือไม่ โดยอิงจากการคำนวณที่ชี้ให้เห็นว่าการสูบฉีดเลือดขึ้นไปที่หัวในท่าทางดังกล่าว[ 67 ]เป็นเวลานานจะใช้พลังงานประมาณครึ่งหนึ่งของพลังงานที่ร่างกายได้รับ[ 68 ]ยิ่งไปกว่านั้น การเคลื่อนเลือดขึ้นไปในระดับความสูงดังกล่าว—โดยไม่คำนึงถึงหัวใจเสริมที่สันนิษฐานไว้ในคอ[ 69 ] —จะต้องใช้หัวใจที่มีขนาดใหญ่กว่าวาฬที่มีขนาดใกล้เคียงกันถึง 15 เท่า[ 70 ]
ข้างต้นถูกนำมาใช้เพื่อโต้แย้งว่าคอยาวนั้นจะต้องอยู่ในแนวนอนมากกว่าหรือน้อยกว่า ซึ่งสันนิษฐานว่าจะช่วยให้สามารถกินพืชในพื้นที่กว้างได้โดยไม่ต้องเคลื่อนที่ไปมามากนัก ทำให้ประหยัดพลังงานได้มากสำหรับสัตว์ขนาดใหญ่เช่นนี้ ดังนั้นการสร้างภาพคอของDiplodocusและApatosaurus ขึ้น ใหม่จึงมักแสดงให้เห็นในท่าเกือบแนวนอน หรือที่เรียกว่า "ท่าที่เป็นกลาง ไม่เบี่ยงเบน" [ 71 ]
อย่างไรก็ตาม การวิจัยเกี่ยวกับสัตว์มีชีวิตแสดงให้เห็นว่าสัตว์มีกระดูกสันหลังสี่ขา เกือบทั้งหมดในปัจจุบัน จะงอส่วนฐานของคออย่างแหลมคมเมื่อตื่นตัว ซึ่งแสดงให้เห็นว่าการอนุมานใดๆ จากกระดูกเกี่ยวกับ "ท่าทางที่เป็นกลาง" ตามปกติ[ 71 ]นั้นไม่น่าเชื่อถืออย่างยิ่ง[ 72 ] [ 73 ] ในขณะเดียวกัน การสร้างแบบจำลองคอนก กระจอกเทศด้วยคอมพิวเตอร์ได้ก่อให้เกิดข้อสงสัยเกี่ยวกับความยืดหยุ่นที่จำเป็นสำหรับการกินหญ้าแบบอยู่กับที่[ 74 ] [ 75 ] [ 76 ]
ทางเดินและระบบการเคลื่อนที่



ร่องรอยเท้าของซอโรพอดและรอยเท้าฟอสซิล อื่นๆ (ที่รู้จักกันในชื่อ "อิคนิต") เป็นที่รู้จักจากหลักฐานมากมายที่พบในทวีปส่วนใหญ่ อิคนิตช่วยสนับสนุนสมมติฐานทางชีววิทยาอื่นๆ เกี่ยวกับซอโรพอด รวมถึงกายวิภาคของเท้าหน้าและเท้าหลังโดยทั่วไป (ดูแขนขาและเท้าด้านบน) โดยทั่วไป รอยเท้าจากเท้าหน้าจะมีขนาดเล็กกว่าเท้าหลังมาก และมักมีรูปร่างคล้ายพระจันทร์เสี้ยว บางครั้งอิคนิตยังคงร่องรอยของกรงเล็บไว้ และช่วยยืนยันว่ากลุ่มซอโรพอดกลุ่มใดที่สูญเสียกรงเล็บหรือแม้แต่นิ้วบนเท้าหน้า[ 77 ]
รอยเท้าซอโรพอดจากชั้นหินVillar del Arzobispo Formation ในยุค Berriasianตอนต้นของสเปน สนับสนุนพฤติกรรมรวมกลุ่มของฝูง รอยเท้าเหล่านี้อาจคล้ายกับSauropodichnus giganteusมากกว่า ichnogenera อื่นๆ แม้ว่าจะมีการเสนอแนะว่ามาจาก titanosauriform พื้นฐานก็ตาม รอยเท้ามีขนาดกว้าง และการจัดกลุ่มให้ใกล้เคียงกับSauropodichnusยังได้รับการสนับสนุนจากระยะห่างระหว่างมือกับเท้า รูปร่างของมือที่คล้ายเมล็ดถั่ว และรูปร่างของเท้าที่เป็นรูปสามเหลี่ยม ไม่สามารถระบุได้ว่ารอยเท้าของฝูงนั้นเกิดจากลูกอ่อนหรือตัวเต็มวัย เนื่องจากไม่มีการระบุอายุของแต่ละตัวจากรอยเท้าก่อนหน้านี้[ 78 ]

โดยทั่วไป รอยเท้าของซอโรพอดจะแบ่งออกเป็นสามประเภทตามระยะห่างระหว่างขาที่อยู่ตรงข้ามกัน ได้แก่ รอยเท้าแคบ รอยเท้าขนาดกลาง และรอยเท้ากว้าง ขนาดของรอยเท้าสามารถช่วยกำหนดได้ว่าขาของซอโรพอดชนิดต่างๆ นั้นอยู่ห่างกันมากน้อยเพียงใด และอาจส่งผลต่อวิธีการเดินของพวกมันอย่างไร[ 77 ]การศึกษาในปี 2004 โดย Day และเพื่อนร่วมงานพบว่าสามารถพบรูปแบบทั่วไปในกลุ่มของซอโรพอดขั้นสูง โดยแต่ละตระกูลของซอโรพอดจะมีลักษณะเฉพาะด้วยขนาดของรอยเท้า พวกเขาพบว่าซอโรพอดส่วนใหญ่นอกเหนือจากไททาโนซอร์มีขาที่แคบ โดยมีรอยประทับที่ชัดเจนของกรงเล็บนิ้วหัวแม่มือขนาดใหญ่บนเท้าหน้า รอยเท้าขนาดกลางที่มีรอยกรงเล็บบนเท้าหน้าอาจเป็นของบราคิโอซอริเดและไททาโนซอริฟอร์ม ดั้งเดิมอื่นๆ ซึ่งกำลังวิวัฒนาการให้มีขาที่อยู่ห่างกันมากขึ้น แต่ยังคงมีกรงเล็บอยู่ ไททาโนซอร์แท้ๆ ดั้งเดิมก็ยังคงมีกรงเล็บที่เท้าหน้า แต่ได้วิวัฒนาการให้มีขาที่กว้างขึ้นอย่างสมบูรณ์ ไททาโนซอร์ขั้นสูงยังคงมีแขนขาที่กว้าง โดยร่องรอยเท้าแสดงให้เห็นแขนขาที่กว้างและไม่มีกรงเล็บหรือนิ้วที่เท้าหน้า[ 79 ]
บางครั้งพบเพียงร่องรอยจากเท้าหน้าเท่านั้น Falkingham et al. [ 80 ]ใช้การจำลองด้วยคอมพิวเตอร์เพื่อแสดงให้เห็นว่าสิ่งนี้อาจเกิดจากคุณสมบัติของพื้นผิว ซึ่งต้องเหมาะสมเพื่อรักษาร่องรอยไว้[ 81 ]ความแตกต่างในพื้นที่ผิวของขาหลังและขาหน้า และด้วยเหตุนี้ แรงกดสัมผัสกับพื้นผิว อาจทำให้บางครั้งพบเพียงร่องรอยจากเท้าหน้าเท่านั้น
ชีวกลศาสตร์และความเร็ว

ในการศึกษาที่ตีพิมพ์ในวารสาร PLoS ONE เมื่อวันที่ 30 ตุลาคม 2013 โดยBill Sellers , Rodolfo Coria , Lee Margetts และคณะ ได้มีการสร้างแบบจำลองดิจิทัล ของArgentinosaurusเพื่อทดสอบการเคลื่อนที่ของมันเป็นครั้งแรก ก่อนการศึกษานี้ วิธีที่ใช้กันทั่วไปในการประมาณความเร็วคือการศึกษาโครงสร้าง กระดูก และร่องรอย เท้า โดยทั่วไป การศึกษาเกี่ยวกับโครงสร้างกระดูกและความเร็วของซอโรพอดมักมุ่งเน้นไปที่โครงกระดูกส่วนลำตัว ซึ่งมีลักษณะเฉพาะหลายอย่าง เช่น กระดูกอัลนา ที่ยื่นออกมา กระดูกเชิงกรานที่มีส่วนยื่น กว้าง กระดูกต้นขา ส่วนบน ที่เอียงเข้าด้านใน และกระดูกต้น ขาที่มีรูปร่างเป็นรูปไข่มาก ลักษณะเหล่านี้มีประโยชน์เมื่อพยายามอธิบายรูปแบบร่องรอยเท้าของสัตว์ที่เคลื่อนที่ตามแรงโน้มถ่วง เมื่อศึกษาร่องรอยเท้าเพื่อคำนวณความเร็วของซอโรพอด มีปัญหาอยู่บ้าง เช่น การให้ค่าประมาณเฉพาะการเดินบางแบบเท่านั้นเนื่องจากอคติในการเก็บรักษาและปัญหาด้านความแม่นยำอีกมากมาย[ 82 ]
เพื่อประเมินการเดินและความเร็วของอาร์เจนติโนซอรัสการศึกษาได้ทำการวิเคราะห์ระบบกล้ามเนื้อและกระดูก การวิเคราะห์ระบบกล้ามเนื้อและกระดูกก่อนหน้านี้มีเพียงอย่างเดียวคือการวิเคราะห์โฮมิโนอิดนกเทอร์เรอร์ และ ไดโนเสาร์ชนิดอื่นก่อนที่จะทำการวิเคราะห์ ทีมงานต้องสร้างโครงกระดูกดิจิทัลของสัตว์ดังกล่าว แสดงตำแหน่งที่มีการเรียงตัวของกล้ามเนื้อ ระบุตำแหน่งของกล้ามเนื้อและข้อต่อ และสุดท้ายคือการค้นหาคุณสมบัติของกล้ามเนื้อก่อนที่จะค้นหาการเดินและความเร็ว ผลการศึกษาด้านชีวกลศาสตร์เผยให้เห็นว่าอาร์เจนติโนซอรัสมีความสามารถทางกลไกที่ความเร็วสูงสุด 2 เมตร/วินาที (5 ไมล์ต่อชั่วโมง) เมื่อพิจารณาจากน้ำหนักตัวที่มากและความเครียดที่ข้อต่อของมันสามารถรับได้[ 83 ]ผลการวิจัยยังเผยให้เห็นอีกว่าสัตว์มีกระดูกสันหลังบนบกที่มีขนาดใหญ่กว่ามากอาจเป็นไปได้ แต่จะต้องมีการปรับโครงสร้างร่างกายอย่างมากและอาจต้องมีการเปลี่ยนแปลงพฤติกรรมที่เพียงพอเพื่อป้องกันการยุบตัวของข้อต่อ[ 82 ]
ขนาดร่างกาย

ซอโรพอดเป็นลูกหลานขนาดมหึมาของบรรพบุรุษที่มีขนาดเล็กอย่างน่าประหลาดใจ ไดโนเสาร์กลุ่มพื้นฐาน เช่นPseudolagosuchusและMarasuchusจากยุคไทรแอสสิกตอนกลางของอาร์เจนตินา มีน้ำหนักประมาณ 1 กิโลกรัม (2.2 ปอนด์) หรือน้อยกว่านั้น พวกมันวิวัฒนาการไปเป็นซอริสเคีย ซึ่งมีการเพิ่ม ขนาด โครงสร้าง อย่างรวดเร็ว แม้ว่าสมาชิกที่ดั้งเดิมกว่า เช่นEoraptor , Panphagia , Pantydraco , SaturnaliaและGuaibasaurusยังคงมีขนาดปานกลาง อาจต่ำกว่า 10 กิโลกรัม (22 ปอนด์) แม้แต่ในรูปแบบดั้งเดิมขนาดเล็กเหล่านี้ ก็ยังมีการเพิ่มขนาดที่เห็นได้ชัดในกลุ่มซอโรโพโดมอร์ฟ แม้ว่าซากดึกดำบรรพ์ในยุคนี้จะมีน้อย ทำให้การตีความยังเป็นการคาดเดาอยู่ มีตัวอย่างที่ชัดเจนของซอโรโพโดมอร์ฟขนาดเล็กที่พัฒนาแล้วคือแอนคิซอรัสซึ่งมีน้ำหนักต่ำกว่า 50 กก. (110 ปอนด์) แม้ว่ามันจะใกล้เคียงกับซอโรพอดมากกว่าเพลทซอรัสและริโอฮาซอรัสซึ่งมีน้ำหนักมากกว่า 1 ตัน (0.98 ตันยาว; 1.1 ตันสั้น) [ 50 ]
ซอโรพอดซึ่งวิวัฒนาการมาจากซอโรโพโดมอร์ฟนั้นมีขนาดใหญ่มาก ขนาดมหึมาของพวกมันน่าจะเป็นผลมาจากอัตราการเติบโตที่เพิ่มขึ้นซึ่งเป็นไปได้ด้วยภาวะเอนโดเทอร์มีเมตาโบลิกแบบเร็วซึ่งเป็นลักษณะที่วิวัฒนาการในซอโรโพโดมอร์ฟ เมื่อแยกออกเป็นซอโรพอดแล้ว ซอโรโพโดมอร์ฟก็ยังคงเติบโตอย่างต่อเนื่องจนมีขนาดใหญ่ขึ้น โดยซอโรพอดขนาดเล็ก เช่นบาราปาซอรัสและโคตาซอรัส ในยุคจูราสสิกตอนต้น ได้วิวัฒนาการไปเป็นรูปแบบที่ใหญ่กว่า เช่นมาเมนชิซอรัสและพาตาโกซอรัส ในยุคจูรา ส สิกตอนกลาง เพื่อตอบสนองต่อการเติบโตของซอโรพอด ผู้ล่าเทโรพอดของพวกมันก็เติบโตขึ้นเช่นกัน ดังที่แสดงให้เห็นโดยโคเอโลฟิซอยด์ขนาดเท่าอัลโลซอรัสจากเยอรมนี[ 50 ]
ขนาดในนีโอซอโรโพดา
นีโอซอโรโพดาเป็นกลุ่มไดโนเสาร์ที่มีขนาดตัวใหญ่ที่สุดเท่าที่เคยมีมา สันนิษฐานว่าข้อยกเว้นบางประการที่มีขนาดเล็กกว่านั้นเกิดจากภาวะแคระแกร็นบนเกาะหรือแรงกดดันทางนิเวศวิทยา อื่นๆ แม้ว่าจะมีแนวโน้มในไททาโนซอเรีย บางชนิด ที่มีขนาดเล็กลง ก็ตาม อย่างไรก็ตาม ไททาโนซอร์เป็นซอโรพอดที่ใหญ่ที่สุดเท่าที่เคยมีมา นอกจากไททาโนซอร์แล้ว ไดพลอโดคอยด์ก็มีขนาดใหญ่โตมโหฬารเช่นกัน ในขณะเดียวกัน กลุ่มไดพลอโดคอยด์ที่เรียกว่าไดเครโอซอริเดมีขนาดตัวเล็กถึงปานกลาง อย่างไรก็ตาม ไม่มีซอโรพอดตัวใดที่มีขนาดเล็กมาก แม้แต่ซอโรพอด "แคระ" ก็ยังมีขนาดใหญ่กว่า 500 กก. (1,100 ปอนด์) ซึ่งเป็นขนาดที่พบได้เพียงประมาณ 10% ของสัตว์เลี้ยงลูกด้วยนมทั้งหมด[ 50 ]
ยักษ์อิสระ
แม้โดยทั่วไปแล้วไดโนเสาร์กลุ่มซอโรพอดจะมีขนาดใหญ่ แต่ขนาดมหึมา (40 ตัน (39 ตันยาว; 44 ตันสั้น) หรือมากกว่านั้น) ก็เกิดขึ้นอย่างอิสระในหลายช่วงเวลาของการวิวัฒนาการ มีไดโนเสาร์ขนาดมหึมาจำนวนมากในยุคจูราสสิกตอนปลาย (โดยเฉพาะยุคคิมเมอริดเจียน ) เช่น ทูเรียซอรัส (Turiasaurus) ใน กลุ่มทูเรีย ซอรัส , มาเมนชิซอริเด (Mamenchisaurus)และ ซินเจียงไททัน ( Xinjiangtitan ) ในกลุ่มมาโพลโด คอยด์ (Diplodocoids ) เช่น ดิโพล โดคัส (Diplodocus) , อะพาโทซอรัส (Apatosaurus) , ซูเปอร์ซอรัส (Supersaurus)และ บารอซอรัส ( Barosaurus) ใน กลุ่มคามาราซอริเด(Camarasaurus)และบราคิโอ ซอริเด (Brachiosaurus)และ จิราฟฟาไททัน ( Giraffatitan ) ในกลุ่มบราคิโอซอริเด ตลอดช่วงต้นถึงปลายยุคครีเทเชียส ไดโนเสาร์ยักษ์อย่างซอโรโพไซดอน (Sauroposeidon) , พาราลิติทัน (Paralititan) , อาร์เจนติโนซอรัส (Argentinosaurus) , เพอร์ทาซอรัส (Puertasaurus) , แอนทาร์คโตซอรัส ( Antarctosaurus) , เดรดนอตั ส (Dreadnoughtus ) , โนโตโคลอสซัส (Notocolossus ) , ฟูทาโลนโกซอรัส (Futalognkosaurus ), พาตาโกไททัน(Patagotitan)และอะลาโมซอรัส (Alamosaurus)มีชีวิตอยู่ โดยทั้งหมดอาจเป็นไททาโนซอรัส หนึ่งในยักษ์ที่รู้จักกันน้อยคือHuanghetitan ruyangensisซึ่งรู้จักกันเฉพาะจากซี่โครงยาว 3 เมตร (9.8 ฟุต) เท่านั้น สายพันธุ์ยักษ์เหล่านี้มีชีวิตอยู่ในช่วงปลายยุคจูราสสิกถึงปลายยุคครีเทเชียส โดยปรากฏขึ้นอย่างอิสระในช่วงเวลา 85 ล้านปี[ 50 ]
ภาวะแคระแกร็นในซอโรพอด
ซอโรพอดแคระสองชนิดที่เป็นที่รู้จักกันดีบนเกาะ ได้แก่แมกยาโรซอรัส ในยุคครีเทเชีย ส (ซึ่งครั้งหนึ่งเคยมีการตั้งข้อสงสัยเกี่ยวกับสถานะความเป็นซอโรพอดแคระของมัน) และยูโรพาซอรัส ในยุคจูราสสิก ซึ่งทั้งสองชนิดมาจากยุโรป แม้ว่าซอโรพอดเหล่านี้จะมีขนาดเล็ก แต่หนทางเดียวที่จะพิสูจน์ได้ว่าพวกมันเป็นซอโรพอดแคระอย่างแท้จริงคือการศึกษาเนื้อเยื่อกระดูก การศึกษาโดยมาร์ติน แซนเดอร์และเพื่อนร่วมงานในปี 2549 ได้ตรวจสอบ ยูโร พาซอรัส โฮลเกรี จำนวน 11 ตัว โดยใช้เนื้อเยื่อกระดูก และแสดงให้เห็นว่าสายพันธุ์บนเกาะขนาดเล็กนี้วิวัฒนาการมาจากการลดลงของอัตราการเติบโตของกระดูกยาวเมื่อเทียบกับอัตราการเติบโตในสายพันธุ์บรรพบุรุษบนแผ่นดินใหญ่[ 84 ]ซอโรพอดแคระที่เป็นไปได้อีกสองชนิด ได้แก่ ราเพโทซอรัส ซึ่งอาศัยอยู่บนเกาะมาดากัสการ์เกาะโดดเดี่ยวในยุคครีเทเชียส และ แอมเพโลซอรัส ไททาโนซอร์ที่อาศัยอยู่บนคาบสมุทรไอบีเรียทางตอนใต้ของสเปนและฝรั่งเศสAmanziaจากสวิตเซอร์แลนด์อาจเป็นไดโนเสาร์แคระเช่นกัน แต่ยังไม่ได้รับการพิสูจน์[ 50 ]หนึ่งในกรณีสุดขั้วของภาวะแคระบนเกาะพบได้ในEuropasaurusซึ่งเป็นญาติของ Camarasaurus และ Brachiosaurus ที่มีขนาดใหญ่กว่ามากโดยมีความยาวเพียงประมาณ 6.2 เมตร (20 ฟุต) ซึ่งเป็นลักษณะเฉพาะของสายพันธุ์ เช่นเดียวกับสายพันธุ์แคระทั้งหมด อัตราการเติบโตที่ลดลงทำให้พวกมันมีขนาดเล็ก [ 31 ] [ 50 ] ไดโนเสาร์ซอโรพอดขนาดเล็กอีกกลุ่มหนึ่งคือ Ibirania ซึ่งเป็นไททาโนซอร์ในกลุ่ม Saltasauridae มีความยาว 5.7 เมตร (19 ฟุต) อาศัยอยู่ในสภาพแวดล้อมที่ไม่ใช่เกาะในบราซิลยุคครีเทเชียสตอนบน และเป็นตัวอย่างของภาวะแคระที่เกิดจากแรงกดดันทางนิเวศวิทยาอื่นๆ[ 85 ]
พยาธิวิทยาโบราณและปรสิตวิทยาโบราณ
โดยทั่วไปแล้ว ซอโรพอดมักไม่ค่อยพบร่องรอยการบาดเจ็บหรือสัญญาณของโรค แต่การค้นพบเมื่อไม่นานมานี้แสดงให้เห็นว่าพวกมันอาจประสบกับโรคดังกล่าวได้ ตัวอย่างไดโพลโดซิดจากชั้นหินมอร์ริสันที่เรียกว่า "ดอลลี่" ได้รับการอธิบายในปี 2022 โดยมีหลักฐานของการติดเชื้อทางเดินหายใจอย่างรุนแรง[ 86 ] [ 87 ]ซี่โครงซอโรพอดจากอำเภอหยุนหยาง มณฑลฉงชิงทางตะวันตกเฉียงใต้ของจีน แสดงหลักฐานการแตกหักของซี่โครงเนื่องจากกระดูกหักจากการบาดเจ็บ การติดเชื้อในกระดูก และภาวะกระดูกแข็งตัว[ 88 ]กระดูกหน้าแข้งซอโรพอดที่แสดงการแตกหักเบื้องต้นได้รับการอธิบายจากยุคจูราสสิกตอนกลางของอำเภอหยุนหยาง ทางตะวันตกเฉียงใต้ของจีน[ 89 ]
ฟอสซิล ไททาโนซอร์นาโนอิด Ibiraniaจากบราซิลบ่งชี้ว่าสิ่งมีชีวิตในสกุลต่างๆ มีความอ่อนแอต่อโรคต่างๆ เช่นโรคกระดูกอักเสบและการติดเชื้อปรสิต ตัวอย่างมาจากยุคครีเทเชียสตอนปลาย São José do Rio Preto Formation , Bauru Basinและได้รับการอธิบายในวารสาร Cretaceous Researchโดย Aureliano et al. (2021) [ 90 ]การตรวจสอบกระดูกของไททาโนซอร์เผยให้เห็นสิ่งที่ดูเหมือนจะเป็นหนอนเลือดปรสิตที่คล้ายกับ Paleoleishmania ในยุคก่อนประวัติศาสตร์ แต่มีขนาดใหญ่กว่า 10-100 เท่า ซึ่งดูเหมือนจะเป็นสาเหตุของโรคกระดูกอักเสบ ฟอสซิลนี้เป็นกรณีแรกที่ทราบกันดีของโรคกระดูกอักเสบรุนแรงที่เกิดจากหนอนเลือดในสัตว์ที่สูญพันธุ์ไปแล้ว [ 91 ] [ 92 ] [ 93 ]
ประวัติการค้นพบ
เศษซากฟอสซิลชิ้นแรกที่ได้รับการยอมรับว่าเป็นซอโรพอดนั้นมาจากประเทศอังกฤษและในตอนแรกถูกตีความในหลากหลายแง่มุม ความสัมพันธ์ของพวกมันกับไดโนเสาร์ชนิดอื่น ๆ ไม่ได้รับการยอมรับจนกระทั่งหลังจากการค้นพบครั้งแรกผ่านไปนานแล้ว


ฟอสซิลซอโรพอดชิ้นแรกที่ได้รับการอธิบายทางวิทยาศาสตร์คือฟันซี่เดียวที่รู้จักกันในชื่อที่ไม่ใช่ แบบ ลินเนียสว่าRutellum implicatum [ 94 ]ฟอสซิลนี้ได้รับการอธิบายโดยเอ็ดเวิร์ด ลูยด์ในปี 1699 แต่ในขณะนั้นยังไม่ได้รับการยอมรับว่าเป็นสัตว์เลื้อยคลานยุคก่อนประวัติศาสตร์ขนาดยักษ์[ 95 ]ไดโนเสาร์จะไม่ได้รับการยอมรับว่าเป็นกลุ่มจนกระทั่งอีกกว่าศตวรรษต่อมา
ริชาร์ด โอเวนได้ตีพิมพ์คำอธิบายทางวิทยาศาสตร์สมัยใหม่ครั้งแรกเกี่ยวกับซอโรพอดในปี พ.ศ. 2384 ในหนังสือและบทความที่ตั้งชื่อคาร์ดิโอดอนและเซทิโอซอรัสคาร์ดิโอดอนเป็นที่รู้จักจากฟันรูปหัวใจสองซี่ที่ผิดปกติ (ซึ่งเป็นที่มาของชื่อ) ซึ่งไม่สามารถระบุได้นอกเหนือจากข้อเท็จจริงที่ว่ามันมาจากสัตว์เลื้อยคลาน ขนาดใหญ่ ที่ ไม่เคยรู้จักมาก่อน เซทิโอซอรัสเป็นที่รู้จักจากซากที่ค่อนข้างดีกว่า แต่ก็ยังไม่สมบูรณ์ โอเวนคิดในขณะนั้นว่าเซทิโอซอรัสเป็นสัตว์เลื้อยคลานทะเลขนาดยักษ์ที่เกี่ยวข้องกับจระเข้ ในปัจจุบัน ดังนั้นจึงได้ชื่อนี้ ซึ่งหมายถึง "กิ้งก่าวาฬ" หนึ่งปีต่อมา เมื่อโอเวนตั้งชื่อไดโนเสาร์ว่า Dinosauriaเขาไม่ได้รวมเซทิโอซอรัสและคาร์ดิโอดอนไว้ในกลุ่มนั้น[ 96 ]
ในปี พ.ศ. 2393 Gideon Mantellตระหนักถึงลักษณะไดโนเสาร์ของกระดูกหลายชิ้นที่ Owen กำหนดให้เป็นCetiosaurus Mantell สังเกตเห็นว่ากระดูกขาประกอบด้วยโพรงไขกระดูกซึ่งเป็นลักษณะเฉพาะของสัตว์บก เขากำหนดให้ตัวอย่างเหล่านี้อยู่ในสกุล ใหม่ Pelorosaurusและจัดกลุ่มไว้ร่วมกับไดโนเสาร์ อย่างไรก็ตาม Mantell ยังคงไม่ยอมรับความสัมพันธ์กับCetiosaurus [ 46 ]
การค้นพบซอโรพอดครั้งต่อไปที่ได้รับการอธิบายและระบุผิดว่าเป็นอย่างอื่นที่ไม่ใช่ไดโนเสาร์คือชุดกระดูกสันหลังสะโพกที่อธิบายโดยแฮร์รี่ ซีลีย์ในปี พ.ศ. 2413 ซีลีย์พบว่ากระดูกสันหลังมีโครงสร้างที่เบามากเมื่อเทียบกับขนาด และมีช่องเปิดสำหรับถุงลม ( การเกิดโพรงอากาศ ) ถุงลมดังกล่าวในขณะนั้นเป็นที่รู้จักเฉพาะในนกและเทโรซอร์ เท่านั้น และซีลีย์พิจารณาว่ากระดูกสันหลังมาจากเทโรซอร์ เขาตั้งชื่อสกุลใหม่ว่าOrnithopsisหรือ "หน้านก" เนื่องจากเหตุนี้[ 46 ]
เมื่อฟิลลิปส์ได้บรรยาย ตัวอย่างที่สมบูรณ์มากขึ้นของ เซทิโอซอรัส ในปี พ.ศ. 2414 ในที่สุดเขาก็ยอมรับว่าสัตว์ชนิดนี้เป็นไดโนเสาร์ที่เกี่ยวข้องกับ เพโลโรซอรัส [ 97 ] อย่างไรก็ตามจนกระทั่งมีการบรรยายโครงกระดูกซอโรพอดที่เกือบสมบูรณ์ชุดใหม่จากสหรัฐอเมริกา (ซึ่งเป็นตัวแทนของอะพาโทซอรัสและคามาราซอรัส ) ในช่วงปลายปีนั้น ภาพรวมที่สมบูรณ์ของซอโรพอดจึงปรากฏขึ้น จอห์น เอ. ไรเดอร์ ศิลปินที่ได้รับการว่าจ้างจากนักบรรพชีวินวิทยา อีดี โคป ได้สร้างภาพจำลองโครงกระดูกซอโรพอดที่สมบูรณ์โดยประมาณขึ้น โดยอิงจากซากของคามาราซอรัสแม้ว่าลักษณะหลายอย่างจะยังไม่ถูกต้องหรือไม่สมบูรณ์ตามการค้นพบในภายหลังและการศึกษาทางชีวกลศาสตร์[ 98 ]นอกจากนี้ ในปี พ.ศ. 2420 ริชาร์ด ไลเดกเกอร์ได้ตั้งชื่อญาติอีกตัวหนึ่งของเซทิโอ ซอรัส ว่าไททาโนซอรัสโดยอิงจากกระดูกสันหลังที่แยกออกมา[ 46 ]

ในปี พ.ศ. 2321 ไดโนเสาร์ซอโรพอดที่สมบูรณ์ที่สุดถูกค้นพบและอธิบายโดยOthniel Charles Marshซึ่งตั้งชื่อมันว่าDiplodocusด้วยการค้นพบนี้ Marsh ยังได้สร้างกลุ่มใหม่เพื่อรวมDiplodocus , Cetiosaurusและญาติๆ ของพวกมันที่เพิ่มมากขึ้นเรื่อยๆ เพื่อแยกความแตกต่างจากกลุ่มไดโนเสาร์หลักอื่นๆ Marsh ตั้งชื่อกลุ่มนี้ว่า Sauropoda หรือ "เท้ากิ้งก่า" [ 46 ]
การจำแนกประเภท
คำจำกัดความ เชิงวิวัฒนาการแรกของ Sauropoda ได้รับการตีพิมพ์ในปี 1997 โดย Salgado และเพื่อนร่วมงาน พวกเขากำหนดกลุ่มนี้เป็นกลุ่มอนุกรมวิธานแบบอิงโหนดซึ่งประกอบด้วย " บรรพบุรุษร่วมที่ใกล้ที่สุดของVulcanodon karibaensisและ Eusauropoda และลูกหลานทั้งหมดของมัน" [ 99 ]ต่อมา มีการเสนอคำจำกัดความ แบบอิงลำต้น หลายแบบ รวมถึงแบบหนึ่งโดย Yates (2007) ซึ่งกำหนด Sauropoda ว่าเป็น "กลุ่มที่ครอบคลุมมากที่สุดที่รวมSaltasaurus loricatusแต่ไม่รวมMelanorosaurus readi " [ 100 ] [ 101 ]
ผู้สนับสนุนคำจำกัดความนี้ยังใช้ชื่อกลุ่มGravisauriaซึ่งกำหนดให้เป็นบรรพบุรุษล่าสุดของTazoudasaurus naimiและSaltasaurus loricatusและลูกหลานทั้งหมด[ 102 ]สำหรับกลุ่มที่เทียบเท่ากับ Sauropoda ตามที่ Salgado et al. กำหนด [ 103 ] กลุ่ม Gravisauria ได้ รับการตั้งชื่อโดยนักบรรพชีวินวิทยาชาวฝรั่งเศส Ronan Allain และ นักบรรพชีวินวิทยา ชาวโมร็อกโก Najat Aquesbi ในปี 2008 เมื่อ การวิเคราะห์ คลัดิสติกของไดโนเสาร์ที่ Allain ค้นพบTazoudasaurusส่งผลให้วงศ์Vulcanodontidaeกลุ่มนี้รวมถึงTazoudasaurusและVulcanodonและกลุ่มพี่น้อง Eusauropoda แต่ยังรวมถึงบางชนิดเช่นAntetonitrus , GongxianosaurusและIsanosaurusที่ไม่ได้อยู่ใน Vulcanodontidae แต่มีตำแหน่งพื้นฐานกว่าใน Sauropoda การนำ Sauropoda มาเปรียบเทียบกับกลุ่มที่มีวิวัฒนาการสูงกว่านี้ ซึ่งรวมถึง Vulcanodontidae และ Eusauropoda ในคำจำกัดความนั้นถือว่าสมเหตุสมผล โดยกำหนดให้เป็นกลุ่มที่เกิดจากบรรพบุรุษร่วมสุดท้ายของTazoudasaurusและSaltasaurus (Bonaparte และ Powell, 1980) และลูกหลานทั้งหมดของมัน Aquesbi กล่าวถึง synapomorphies สองประการ ซึ่งเป็นลักษณะร่วมที่มีวิวัฒนาการของ Gravisauria ได้แก่กระดูกสันหลังกว้างกว่าด้านข้างมากกว่าด้านหน้าและด้านหลัง และมี condyles femoris ที่ไม่สมมาตรที่ด้านล่างของกระดูกต้นขาก่อนหน้านี้ไม่คิดว่าสิ่งเหล่านี้เป็น synapomorphies ของ Eusauropoda แต่ Allian พบคุณสมบัติเหล่านี้ในTazoudasaurus เช่น กัน [ 104 ]
Gravisauria แยกตัวออกมาในช่วงต้นยุคจูราสสิกประมาณช่วงPliensbachianและToarcianเมื่อ 183 ล้านปีก่อน และ Aquesbi คิดว่านี่เป็นส่วนหนึ่งของการปฏิวัติครั้งใหญ่ในกลุ่มสัตว์ซึ่งรวมถึงการหายไปของProsauropoda , CoelophysoideaและThyreophora ขั้นพื้นฐาน ซึ่งพวกเขาเชื่อว่าเป็นผลมาจากการสูญพันธุ์ครั้งใหญ่ ทั่วโลก [ 104 ]
ความสัมพันธ์ทางวิวัฒนาการของไดโนเสาร์ซอโรพอดส่วนใหญ่มีเสถียรภาพมากขึ้นในช่วง ไม่ กี่ปีที่ผ่านมา แม้ว่าจะยังมีความไม่แน่นอนอยู่บ้าง เช่น การจัดวางตำแหน่งของEuhelopus , Haplocanthosaurus , JobariaและNemegtosauridae
แผนภูมิวิวัฒนาการหลังจากการวิเคราะห์ที่นำเสนอโดย Sander และเพื่อนร่วมงานในปี 2011 [ 50 ]
| †ซอโรโปดา | |
ดูเพิ่มเติม
ลิงก์ภายนอก
 สื่อที่เกี่ยวข้องกับซอโรโปดาในวิกิมีเดียคอมมอนส์
สื่อที่เกี่ยวข้องกับซอโรโปดาในวิกิมีเดียคอมมอนส์- Strauss, Bob (2008). "ซอโรพอด: ไดโนเสาร์ที่ใหญ่ที่สุดเท่าที่เคยมีมา" . about.com . ประเภทของไดโนเสาร์. เก็บถาวรจากต้นฉบับเมื่อ 2 กุมภาพันธ์ 2017 . สืบค้นเมื่อ7 กรกฎาคม 2008 .
- Rogers, KC ; Wilson, JA (2005). ไดโนเสาร์ซอโรพอด: วิวัฒนาการและบรรพชีววิทยา . เบิร์กลีย์, แคลิฟอร์เนีย: สำนักพิมพ์มหาวิทยาลัยแคลิฟอร์เนีย . ISBN 0-520-24623-3.
- Upchurch, P.; Barrett, PM; Dodson, P. (2004). "ซอโรโพดา". ในWeishampel, D. ; Dodson, P.; Osmólska, H. (บรรณาธิการ). ไดโนเสาร์ (ฉบับที่ 2). เบิร์กลีย์, แคลิฟอร์เนีย: สำนักพิมพ์มหาวิทยาลัยแคลิฟอร์เนีย . หน้า 259–322 .
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ซอโรโปดา
ซอโรโพดา ( / s ɔː ˈ r ɒ p ə d ə / ) ซึ่งสมาชิกเรียกว่า ซอโรพอด ( / ˈ s ɔːr ə p ɒ d z / ; [ 1 ] [ 2 ] มาจาก sauro- + -pod ; แปลตรงตัวว่า ' เท้า เหมือน กิ้งก่า ' ) เป็น กลุ่ม ของ...
คำอธิบาย
ซอโรพอดเป็น สัตว์ กินพืช มัก เป็นสัตว์สี่ขาที่ มีคอยาว [ 15 ] และมักมีฟันรูปช้อน (รูปทรงคล้ายไม้พาย: กว้างที่ปลาย แคบที่คอ) พวกมันมีหัวค่อนข้างเล็ก ลำตัวใหญ่โต และส่วนใหญ่มีหางยาว ขาหลังของพวกมันหนา ตรง และแข็งแรง ปลายขาเป็นเท้าคล้ายกระบองที่มีห้านิ้ว...
ขนาด
ลักษณะเด่นที่สุดของซอโรพอดคือขนาดของมัน แม้แต่ซอโรพอดแคระ (อาจยาว 5 ถึง 6 เมตร (16 ถึง 20 ฟุต)) ก็ยังถูกนับว่าเป็นสัตว์ที่ใหญ่ที่สุดใน ระบบนิเวศ ของพวก มัน คู่แข่งที่แท้จริงในแง่ของขนาดคือ โรร์ควอล เช่น วาฬสีน้ำเงิน แต่ต่างจากวาฬ ซอโรพอด เป็น สัตว์บก เป็นหลัก
แขนขาและเท้า
ในฐานะ สัตว์สี่ขา ขนาดใหญ่ ซอโรพอดได้พัฒนาแขนขาที่ "รับน้ำหนัก" (graviportal) พิเศษ เท้าหลังกว้างและยังคงมีกรงเล็บสามอันในสายพันธุ์ส่วนใหญ่ [ 36 ] สิ่งที่ผิดปกติเป็นพิเศษเมื่อเทียบกับสัตว์อื่นๆ คือเท้าหน้า ( manus ) ที่ได้รับการดัดแปลงอย่างมาก...