อ่าน 38 นาที
การเผาผลาญ
เมตาบอลิซึม ( / m ə ˈ t æ b ə l ɪ z ə m / , จาก ภาษากรีก : μεταβολή metabolē , "การเปลี่ยนแปลง") หมายถึงชุดของ ปฏิกิริยาเคมี ที่จำเป็นต่อการดำรง ชีวิต ซึ่งเกิดขึ้นภายใน...
การเผาผลาญ

| ส่วนหนึ่งของชุดบทความเกี่ยวกับ |
| ชีวเคมี |
|---|
 |
|
เมตาบอลิซึม ( / m ə ˈ t æ b ə l ɪ z ə m / ,จากภาษากรีก : μεταβολή metabolē , "การเปลี่ยนแปลง") หมายถึงชุดของปฏิกิริยาเคมีที่จำเป็นต่อการดำรงชีวิตซึ่งเกิดขึ้นภายในสิ่งมีชีวิตหน้าที่หลักสามประการของเมตาบอลิซึม ได้แก่ การเปลี่ยนพลังงานในอาหารให้เป็นรูปแบบที่ใช้ได้สำหรับกระบวนการภายในเซลล์ การเปลี่ยนอาหารให้เป็นหน่วยสร้างของโมเลกุลขนาดใหญ่ ( พอลิเมอร์ชีวภาพ ) เช่นโปรตีนไขมันกรดนิวคลีอิกและคาร์โบไฮเดรตบางชนิดและการขับถ่ายของเสียจากกระบวนการเมตาบอลิ ซึม ปฏิกิริยาที่เร่งด้วย เอนไซม์เหล่านี้ช่วยให้สิ่งมีชีวิตเจริญเติบโต ขยายพันธุ์ รักษาโครงสร้างและตอบสนองต่อสิ่งแวดล้อม คำว่าเมตาบอลิซึมยังสามารถหมายถึงปฏิกิริยาเคมีทั้งหมดที่เกิดขึ้นในสิ่งมีชีวิต รวมถึงการย่อยอาหารและการขนส่งสารเข้าและระหว่างเซลล์ต่างๆ ด้วย ในความหมายที่กว้างขึ้น ชุดของปฏิกิริยาที่เกิดขึ้นภายในเซลล์เรียกว่าเมตาบอลิซึมระดับกลาง (หรือเมตาบอลิซึมขั้นกลาง)
ปฏิกิริยาเมตาบอลิซึมสามารถแบ่งออกเป็นสองประเภท คือแคตาโบลิก — การสลายสารประกอบ (ตัวอย่างเช่น การเปลี่ยนกลูโคสเป็นไพรูเวทโดยการหายใจระดับเซลล์ ) และอนาโบลิก — การสร้าง(การสังเคราะห์ทางชีวภาพ ) สารประกอบ (เช่น โปรตีน คาร์โบไฮเดรต ไขมัน และกรดนิวคลีอิก) โดยปกติแล้ว แคตาโบลิกจะปล่อยพลังงานออกมา และอนาโบลิกจะใช้พลังงาน
ปฏิกิริยาเคมีในกระบวนการเผาผลาญถูกจัดระเบียบเป็นวิถีการเผาผลาญซึ่งสารเคมีหนึ่งจะถูกเปลี่ยนไปเป็นสารเคมีอีกชนิดหนึ่งผ่านขั้นตอนต่างๆ โดยแต่ละขั้นตอนจะได้รับการอำนวยความสะดวกโดยเอนไซม์ เฉพาะ เอนไซม์มีความสำคัญอย่างยิ่งต่อกระบวนการเผาผลาญ เพราะช่วยให้สิ่งมีชีวิตสามารถขับเคลื่อนปฏิกิริยาที่ต้องการพลังงานและจะไม่เกิดขึ้นเองได้ โดยการเชื่อมโยง ปฏิกิริยา เหล่านั้นเข้ากับปฏิกิริยาที่เกิดขึ้นเองตามธรรมชาติซึ่งปลดปล่อยพลังงาน เอนไซม์ทำหน้าที่เป็นตัวเร่งปฏิกิริยา—ช่วยให้ปฏิกิริยาเกิดขึ้นได้เร็วขึ้น—และยังช่วยควบคุมอัตราของปฏิกิริยาการเผาผลาญ เช่น การตอบสนองต่อการเปลี่ยนแปลงใน สภาพแวดล้อม ของเซลล์หรือสัญญาณจากเซลล์อื่นๆ
ระบบการเผาผลาญของสิ่งมีชีวิตชนิดใดชนิดหนึ่งจะเป็นตัวกำหนดว่าสารใดที่สิ่งมีชีวิตนั้นจะพบว่ามีคุณค่าทางโภชนาการและสารใดที่เป็นพิษตัวอย่างเช่น โปรคา ริโอต บางชนิด ใช้ไฮโดรเจนซัลไฟด์ (H₂S) เป็นสารอาหาร แต่ก๊าซนี้เป็นพิษต่อสัตว์บางชนิด[ 1 ]เนื่องจากไฮโดรเจนซัลไฟด์เป็นก๊าซส่งสัญญาณสัตว์เลี้ยงลูกด้วยนมบางชนิดรวมถึงมนุษย์จึงผลิต H₂S ตามธรรมชาติในความเข้มข้นต่ำมาก ซึ่งทำหน้าที่ส่งสัญญาณและควบคุมที่สำคัญ[ 2 ]อัตราการเผาผลาญพื้นฐานของสิ่งมีชีวิตคือการวัดปริมาณพลังงานที่ใช้ไปในปฏิกิริยาเคมีทั้งหมดเหล่านี้
ลักษณะเด่นอย่างหนึ่งของการเผาผลาญคือความคล้ายคลึงกันของวิถีการเผาผลาญพื้นฐานในสิ่งมีชีวิตต่างชนิดกัน[ 3 ]ตัวอย่างเช่น ชุดของกรดคาร์บอกซิลิกซึ่งเป็นที่รู้จักกันดีว่าเป็นสารตัวกลางในวัฏจักรกรดซิตริกนั้นมีอยู่ในสิ่งมีชีวิตทุกชนิดที่รู้จัก โดยพบได้ในสิ่งมีชีวิตที่หลากหลาย ตั้งแต่แบคทีเรียเซลล์เดียว อย่าง Escherichia coli ( E. coli) ไปจนถึง สิ่งมีชีวิตหลายเซลล์ขนาดใหญ่เช่นช้าง[ 4 ] ความคล้ายคลึงกันในวิถีการเผาผลาญเหล่านี้น่าจะ เกิดจากการปรากฏตัวในช่วงต้นของวิวัฒนาการและการคงอยู่ของวิถีเหล่านี้น่าจะเกิดจากประสิทธิภาพของ วิถีเหล่านั้น [ 5 ] [ 6 ]ในโรคต่างๆ เช่นโรคเบาหวานชนิดที่ 2 กลุ่มอาการเมตาบอลิกและมะเร็งการเผาผลาญปกติจะถูกรบกวน[ 7 ]การเผาผลาญของเซลล์มะเร็งก็แตกต่างจากการเผาผลาญของเซลล์ปกติ และความแตกต่างเหล่านี้สามารถนำมาใช้เพื่อค้นหาเป้าหมายสำหรับการรักษาโรคมะเร็งได้[ 8 ]
สารชีวเคมีที่สำคัญ
โครงสร้างส่วนใหญ่ที่ประกอบขึ้นเป็นสัตว์ พืช และจุลินทรีย์นั้นสร้างขึ้นจากโมเลกุล พื้นฐานสี่ประเภท ได้แก่ กรด อะมิโนคาร์โบไฮเดรตกรดนิวคลีอิกและลิปิด (มักเรียกว่าไขมัน ) เนื่องจากโมเลกุลเหล่านี้มีความสำคัญต่อชีวิต ปฏิกิริยาเมตาบอลิซึมจึงมุ่งเน้นไปที่การสร้างโมเลกุลเหล่านี้ในระหว่างการสร้างเซลล์และเนื้อเยื่อ หรือการย่อยสลายและนำไปใช้เพื่อให้ได้พลังงานโดยการย่อยอาหาร สารชีวเคมีเหล่านี้สามารถรวมกันเพื่อสร้างพอลิเมอร์เช่นDNAและโปรตีนซึ่งเป็นโมเลกุลขนาดใหญ่ที่จำเป็นต่อชีวิต[ 9 ]
| ประเภทของโมเลกุล | ชื่อของรูปแบบ โมโนเม อร์ | ชื่อของรูปแบบ พอลิเมอร์ | ตัวอย่างของรูปแบบพอลิเมอร์ |
|---|---|---|---|
| กรดอะมิโน | กรดอะมิโน | โปรตีน (ประกอบด้วยพอลิเปปไทด์) | โปรตีนเส้นใยและโปรตีนทรงกลม |
| คาร์โบไฮเดรต | โมโนแซ็กคาไรด์ | โพลีแซ็กคาไรด์ | แป้งไกลโคเจนและเซลลูโลส |
| กรดนิวคลีอิก | นิวคลีโอไทด์ | โพลีนิวคลีโอไทด์ | ดีเอ็นเอและอาร์เอ็นเอ |
กรดอะมิโนและโปรตีน
โปรตีนประกอบด้วยกรดอะมิโนที่เรียงตัวเป็นสายตรงเชื่อมต่อกันด้วยพันธะเปปไทด์โปรตีนหลายชนิดเป็นเอนไซม์ที่เร่งปฏิกิริยาเคมีในกระบวนการเผาผลาญ โปรตีนอื่นๆ มีหน้าที่เชิงโครงสร้างหรือเชิงกล เช่น โปรตีนที่สร้างโครงร่างเซลล์ซึ่งเป็นระบบโครงสร้างที่ช่วยรักษารูปร่างของเซลล์[ 10 ]โปรตีนยังมีความสำคัญในการส่งสัญญาณของเซลล์การตอบสนองทางภูมิคุ้มกันการยึดเกาะของเซลล์การขนส่งแบบแอคทีฟผ่านเยื่อหุ้มเซลล์ และวงจรเซลล์[ 11 ]กรดอะมิโนยังมีส่วนช่วยในการเผาผลาญพลังงานของเซลล์โดยการจัดหาแหล่งคาร์บอนสำหรับการเข้าสู่รอบกรดซิตริก ( รอบกรดไตรคาร์บอกซิลิก ) [ 12 ]โดยเฉพาะอย่างยิ่งเมื่อแหล่งพลังงานหลัก เช่นกลูโคสขาดแคลน หรือเมื่อเซลล์ประสบกับความเครียดทางเมตาบอลิซึม[ 13 ]
ลิปิด
ลิปิดเป็นกลุ่มสารชีวเคมีที่มีความหลากหลายมากที่สุด การใช้งานเชิงโครงสร้างหลักของลิปิดคือการเป็นส่วนหนึ่งของเยื่อชีวภาพ ภายในและภายนอก เช่นเยื่อหุ้มเซลล์[ 11 ]พลังงานเคมีของ ลิปิดยังสามารถนำมาใช้ได้อีกด้วย ลิปิดประกอบด้วย โซ่ไฮโดรคาร์บอนยาวที่ไม่เป็นขั้วโดยมีบริเวณที่เป็นขั้วขนาดเล็กที่มีออกซิเจน ลิปิดมักถูกนิยามว่าเป็น โมเลกุลชีวภาพที่ไม่ ชอบน้ำหรือ แอม ฟิพาติกแต่จะละลายในตัวทำละลายอินทรีย์เช่นเอทานอลเบนซีนหรือคลอโรฟอร์ม[ 14 ]ไขมันเป็นกลุ่มสารประกอบขนาดใหญ่ที่ประกอบด้วยกรดไขมันและกลีเซอรอล โมเลกุลกลีเซอรอลที่เชื่อมต่อกับกรดไขมันสามโมเลกุลด้วยพันธะเอสเทอร์เรียกว่าไตรอะซิลกลีเซอไรด์[ 15 ]โครงสร้างพื้นฐานมีหลายรูปแบบรวมถึงโครงสร้างหลัก เช่นสฟิงโกซีนในสฟิงโกไมอีลินและ กลุ่ม ที่ชอบน้ำเช่นฟอสเฟตในฟอสโฟลิปิด สเตียรอยด์เช่น สเตอ รอลเป็นลิปิดอีกกลุ่มหลักหนึ่ง[ 16 ]
คาร์โบไฮเดรต
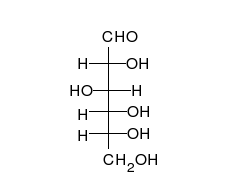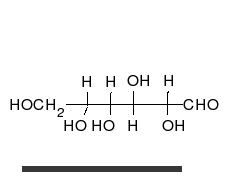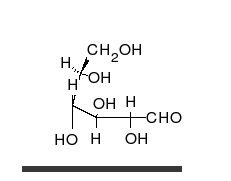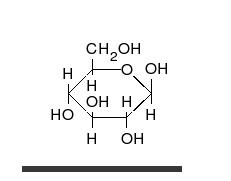
คาร์โบไฮเดรตเป็นอัลดีไฮด์หรือคีโตนที่มี หมู่ ไฮดรอก ซิลจำนวนมาก ติดอยู่ ซึ่งสามารถอยู่ในรูปของโซ่ตรงหรือวงแหวนได้ คาร์โบไฮเดรตเป็นโมเลกุลทางชีวภาพที่พบมากที่สุดและมีบทบาทมากมาย เช่น การเก็บและขนส่งพลังงาน ( แป้งไกลโคเจน) และเป็นส่วนประกอบโครงสร้าง ( เซลลูโลสในพืชไคตินในสัตว์) [ 11 ]หน่วยพื้นฐานของคาร์โบไฮเดรตเรียกว่าโมโนแซ็กคาไรด์ซึ่งรวมถึงกาแลคโตสฟรุกโตสและที่สำคัญที่สุดคือกลูโคสโมโนแซ็กคาไรด์สามารถเชื่อมต่อกันเพื่อสร้างพอลิแซ็กคาไรด์ได้ในรูปแบบที่แทบจะไม่มีที่สิ้นสุด[ 17 ]
นิวคลีโอไทด์
กรดนิวคลีอิกสองชนิด ได้แก่ DNA และRNAเป็นพอลิเมอร์ของนิวคลีโอไทด์แต่ละนิวคลีโอไทด์ประกอบด้วยฟอสเฟตที่ติดอยู่กับกลุ่มน้ำตาลไรโบสหรือดีออกซีไรโบ ส ซึ่งติดอยู่กับ เบสที่มีไนโตรเจนกรดนิวคลีอิกมีความสำคัญต่อการจัดเก็บและการใช้ข้อมูลทางพันธุกรรม และการตีความผ่านกระบวนการถอดรหัสและการสังเคราะห์โปรตีน[ 11 ]ข้อมูลนี้ได้รับการปกป้องโดยกลไกการซ่อมแซม DNA และแพร่กระจายผ่าน การจำลองแบบ DNA ไวรัสหลายชนิดมีจีโนม RNAเช่นHIVซึ่งใช้การถอดรหัสย้อนกลับเพื่อสร้างแม่แบบ DNA จากจีโนม RNA ของไวรัส[ 18 ] RNA ในไรโบไซม์เช่นสไปลโซโซมและไรโบโซมมีลักษณะคล้ายกับเอนไซม์ เนื่องจากสามารถเร่งปฏิกิริยาเคมีได้ นิวคลีโอไซด์แต่ละตัวถูกสร้างขึ้นโดยการติด นิวคลี โอเบส เข้า กับน้ำตาลไรโบส เบสเหล่านี้เป็น วงแหวน เฮเทอโรไซคลิกที่มีไนโตรเจน จัดเป็นพิวรีนหรือไพริมิดีน นิวคลีโอไทด์ยังทำหน้าที่เป็นโคเอนไซม์ในปฏิกิริยาการถ่ายโอนกลุ่มเมตาบอลิซึมด้วย[ 19 ]
โคเอนไซม์
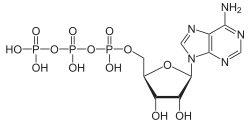
กระบวนการเผาผลาญเกี่ยวข้องกับปฏิกิริยาเคมีมากมาย แต่ส่วนใหญ่จัดอยู่ในปฏิกิริยาพื้นฐานไม่กี่ประเภทที่เกี่ยวข้องกับการถ่ายโอนหมู่ฟังก์ชันของอะตอมและพันธะภายในโมเลกุล[ 20 ]เคมีทั่วไปนี้ทำให้เซลล์สามารถใช้ตัวกลางการเผาผลาญจำนวนเล็กน้อยเพื่อขนส่งหมู่เคมีระหว่างปฏิกิริยาต่างๆ[ 19 ]ตัวกลางการถ่ายโอนหมู่เหล่านี้เรียกว่าโคเอนไซม์ปฏิกิริยาการถ่ายโอนหมู่แต่ละประเภทจะดำเนินการโดยโคเอนไซม์เฉพาะ ซึ่งเป็นสารตั้งต้นสำหรับชุดเอนไซม์ที่ผลิตมัน และชุดเอนไซม์ที่บริโภคมัน ดังนั้นโคเอนไซม์เหล่านี้จึงถูกสร้าง บริโภค และนำกลับมาใช้ใหม่อย่างต่อเนื่อง[ 21 ]
โคเอนไซม์หลักตัวหนึ่งคืออะดีโนซีนไตรฟอสเฟต (ATP) ซึ่งเป็นสกุลเงินพลังงานของเซลล์ นิ ว คลีโอไทด์ นี้ ใช้ในการถ่ายโอนพลังงานเคมีระหว่างปฏิกิริยาเคมีต่างๆ มี ATP อยู่ในเซลล์เพียงเล็กน้อย แต่เนื่องจากมีการสร้างใหม่อย่างต่อเนื่อง ร่างกายมนุษย์จึงสามารถใช้ ATP ได้ประมาณเท่ากับน้ำหนักตัวต่อวัน[ 21 ] ATP ทำหน้าที่เป็นสะพานเชื่อมระหว่างกระบวนการสลายและกระบวนการสร้าง กระบวนการสลายจะสลายโมเลกุล และกระบวนการสร้างจะนำโมเลกุลเหล่านั้นมารวมกัน ปฏิกิริยาการสลายจะสร้าง ATP และปฏิกิริยาการสร้างจะใช้ ATP นอกจากนี้ยังทำหน้าที่เป็นตัวนำกลุ่มฟอสเฟตในปฏิกิริยาฟอสโฟรีเลชัน[ 22 ]
วิตามินเป็นสารประกอบอินทรีย์ที่จำเป็นในปริมาณเล็กน้อยซึ่งไม่สามารถสร้างขึ้นในเซลล์ได้ ในโภชนาการของมนุษย์วิตามินส่วนใหญ่ทำหน้าที่เป็นโคเอนไซม์หลังจากได้รับการดัดแปลง ตัวอย่างเช่น วิตามินที่ละลายน้ำได้ทั้งหมดจะถูกฟอสโฟรีเลตหรือจับคู่กับนิวคลีโอไทด์เมื่อนำไปใช้ในเซลล์[ 23 ]นิโคตินาไมด์อะดีนีนไดนิวคลีโอไทด์ (NAD + ) ซึ่งเป็นอนุพันธ์ของวิตามินบี3 ( ไนอาซิน ) เป็นโคเอนไซม์ที่สำคัญซึ่งทำหน้าที่เป็นตัวรับไฮโดรเจน ดีไฮโดรจีเนสหลายร้อยชนิดแยกกันจะกำจัดอิเล็กตรอนออกจากสารตั้งต้นและลด NAD + ให้เป็น NADH รูปแบบที่ลดลงของโคเอนไซม์นี้จะเป็นสารตั้งต้นสำหรับ รีดักเทสใดๆในเซลล์ที่ต้องการถ่ายโอนอะตอมไฮโดรเจนไปยังสารตั้งต้น[ 24 ]นิโคตินาไมด์อะดีนีนไดนิวคลีโอไทด์มีอยู่ในสองรูปแบบที่เกี่ยวข้องกันในเซลล์ คือ NADH และ NADPH รูปแบบ NAD + /NADH มีความสำคัญมากกว่าในปฏิกิริยาแคตาโบลิก ในขณะที่ NADP + /NADPH ใช้ในปฏิกิริยาอะนาโบลิก[ 25 ]

แร่ธาตุและโคแฟคเตอร์
ธาตุอนินทรีย์มีบทบาทสำคัญในกระบวนการเผาผลาญ บางชนิดมีปริมาณมาก (เช่นโซเดียมและโพแทสเซียม) ในขณะ ที่บางชนิดทำงานที่ความเข้มข้นเพียงเล็กน้อย ประมาณ 99% ของน้ำหนักตัวของมนุษย์ประกอบด้วยธาตุคาร์บอนไนโตรเจนแคลเซียมโซเดียมคลอรีนโพแทสเซียมไฮโดรเจนฟอสฟอรัสออกซิเจนและกำมะถันสารประกอบอินทรีย์ ( โปรตีน ไขมัน และ คาร์โบไฮเดรต)ประกอบด้วยคาร์บอนและไนโตรเจนเป็นส่วนใหญ่ ออกซิเจนและไฮโดรเจนส่วนใหญ่จะอยู่ในรูปของน้ำ[ 26 ]
ธาตุอนินทรีย์ ที่มีอยู่มากมายทำหน้าที่เป็นอิเล็กโทรไลต์ไอออนที่สำคัญที่สุดคือโซเดียมโพแทสเซียมแคลเซียมแมกนีเซียมคลอไรด์ฟอสเฟตและไอออนอินทรีย์ไบคาร์บอเนตการรักษาระดับความเข้มข้นของไอออน ที่แม่นยำ ทั่วเยื่อหุ้มเซลล์ช่วยรักษาความดันออสโมติกและค่าpH [ 27 ]ไอออนยังมีความสำคัญต่อการทำงานของเส้นประสาทและกล้ามเนื้อ เนื่องจากศักยภาพการกระทำในเนื้อเยื่อเหล่านี้เกิดขึ้นจากการแลกเปลี่ยนอิเล็กโทรไลต์ระหว่างของเหลวนอก เซลล์ และของเหลวภายในเซลล์ ซึ่ง ก็คือ ไซโตซอล[ 28 ]อิเล็กโทรไลต์เข้าและออกจากเซลล์ผ่านโปรตีนในเยื่อหุ้มเซลล์ที่เรียกว่าช่องไอออนตัวอย่างเช่นการหดตัวของกล้ามเนื้อขึ้นอยู่กับการเคลื่อนที่ของแคลเซียม โซเดียม และโพแทสเซียมผ่านช่องไอออนในเยื่อหุ้มเซลล์และท่อที[ 29 ]
โลหะทรานซิชันมักพบเป็นธาตุติดตามในสิ่งมีชีวิต โดยสังกะสีและเหล็กเป็นธาตุที่มีปริมาณมากที่สุด[ 30 ]โคแฟคเตอร์โลหะจะจับกับตำแหน่งเฉพาะในโปรตีนอย่างแน่นหนา แม้ว่าโคแฟคเตอร์ของเอนไซม์จะสามารถเปลี่ยนแปลงได้ในระหว่างการเร่งปฏิกิริยา แต่ก็จะกลับคืนสู่สถานะเดิมเสมอเมื่อสิ้นสุดปฏิกิริยาที่ถูกเร่ง โลหะธาตุอาหารขนาดเล็กจะถูกดูดซึมเข้าสู่สิ่งมีชีวิตโดยตัวขนส่งเฉพาะและจับกับโปรตีนเก็บสะสม เช่นเฟอร์ริตินหรือเมทัลโลไทโอนีนเมื่อไม่ได้ใช้งาน[ 31 ] [ 32 ]
การสลายตัว
กระบวนการสลายสาร (Catabolism) คือชุดของกระบวนการเผาผลาญที่สลายโมเลกุลขนาดใหญ่ ซึ่งรวมถึงการสลายและออกซิไดซ์โมเลกุลอาหาร จุดประสงค์ของปฏิกิริยาสลายสารคือการให้พลังงานและส่วนประกอบที่จำเป็นสำหรับปฏิกิริยาสร้างสาร (Anabolic reactions) ซึ่งสร้างโมเลกุล[ 33 ]ลักษณะที่แท้จริงของปฏิกิริยาสลายสารเหล่านี้แตกต่างกันไปในแต่ละสิ่งมีชีวิต และสิ่งมีชีวิตสามารถจำแนกได้ตามแหล่งพลังงาน ไฮโดรเจน และคาร์บอน ( กลุ่มโภชนาการหลัก ) ดังแสดงในตารางด้านล่าง ออร์แกโนโทรฟใช้โมเลกุลอินทรีย์เป็นแหล่งของอะตอมไฮโดรเจนหรืออิเล็กตรอนในขณะที่ลิโทโทรฟใช้สารตั้งต้นอนินทรีย์ ในขณะที่โฟโตโทรฟเปลี่ยนแสงแดดเป็นพลังงานเคมี[ 34 ]เคโมโทรฟอาศัย ปฏิกิริยา รีดอก ซ์ ที่เกี่ยวข้องกับการถ่ายโอนอิเล็กตรอนจากโมเลกุลผู้ให้ที่ลดลง เช่นโมเลกุลอินทรีย์ไฮโดรเจนไฮโดรเจนซัลไฟด์หรือไอออนเหล็กไปยังออกซิเจนไนเตรตหรือซัลเฟตในสัตว์ ปฏิกิริยาเหล่านี้เกี่ยวข้องกับโมเลกุลอินทรีย์ที่ซับซ้อนซึ่งถูกย่อยสลายเป็นโมเลกุลที่เรียบง่ายกว่า เช่นคาร์บอนไดออกไซด์และน้ำ สิ่งมีชีวิต ที่สังเคราะห์แสงได้เช่น พืชและไซยาโนแบคทีเรียใช้ปฏิกิริยาการถ่ายโอนอิเล็กตรอนที่คล้ายกันเพื่อเก็บพลังงานที่ดูดซับจากแสงแดด[ 35 ]
| แหล่งพลังงาน | แสงแดด | รูปถ่าย- | -ถ้วยรางวัล | ||
| โมเลกุล | เคมีบำบัด- | ||||
| ผู้ให้ไฮโดรเจนหรืออิเล็กตรอน | สารประกอบอินทรีย์ | ออร์กาโน- | |||
| สารประกอบอนินทรีย์ | ลิโธ- | ||||
| แหล่งคาร์บอน | สารประกอบอินทรีย์ | เฮเทอโร- | |||
| สารประกอบอนินทรีย์ | อัตโนมัติ- | ||||
ชุดปฏิกิริยาแคตาโบลิกที่พบได้บ่อยที่สุดในสัตว์สามารถแบ่งออกเป็นสามขั้นตอนหลัก ในขั้นตอนแรก โมเลกุลอินทรีย์ขนาดใหญ่ เช่นโปรตีนโพลีแซ็กคาไรด์หรือไขมันจะถูกย่อยเป็นส่วนประกอบที่เล็กกว่าภายนอกเซลล์ ต่อมา โมเลกุลที่เล็กกว่าเหล่านี้จะถูกดูดซึมเข้าสู่เซลล์และเปลี่ยนเป็นโมเลกุลที่เล็กกว่า ซึ่งโดยทั่วไปคืออะเซทิลโคเอนไซม์เอ (acetyl-CoA) ซึ่งจะปล่อยพลังงานออกมาบางส่วน สุดท้าย หมู่แอเซทิลบน acetyl-CoA จะถูกออกซิไดซ์เป็นน้ำและคาร์บอนไดออกไซด์ในระหว่างวัฏจักรกรดซิตริกและ ห่วง โซ่การขนส่งอิเล็กตรอน ซึ่ง จะปล่อยพลังงานออกมามากขึ้นในขณะที่ลดโคเอนไซม์นิโคตินาไมด์อะดีนีนไดนิวคลีโอไทด์ (NAD + ) ให้เป็น NADH [ 33 ]
การย่อยอาหาร
เซลล์ไม่สามารถประมวลผลโมเลกุลขนาดใหญ่ได้โดยตรง โมเลกุลขนาดใหญ่ต้องถูกย่อยเป็นหน่วยที่เล็กกว่าก่อนจึงจะสามารถนำไปใช้ในการเผาผลาญของเซลล์ได้ เอนไซม์ประเภทต่างๆ ถูกนำมาใช้ในการย่อยพอลิเมอร์เหล่านี้เอนไซม์ย่อย เหล่านี้ ได้แก่โปรตีเอสที่ย่อยโปรตีนให้เป็นกรดอะมิโน และไกลโคไซด์ไฮโดรเลสที่ย่อยพอลิแซ็กคาไรด์ให้เป็นน้ำตาลโมโนแซ็กคาไรด์[ 37 ]
จุลินทรีย์จะหลั่งเอนไซม์ย่อยอาหารออกสู่สิ่งแวดล้อม[ 38 ] [ 39 ]ในขณะที่สัตว์จะหลั่งเอนไซม์เหล่านี้จากเซลล์เฉพาะในลำไส้ เท่านั้น รวมถึงกระเพาะอาหาร ตับอ่อนและต่อมน้ำลาย[ 40 ]จากนั้นกรดอะมิโนหรือน้ำตาลที่ถูกปล่อยออกมาจากเอนไซม์นอกเซลล์เหล่านี้จะถูกปั๊มเข้าไปในเซลล์โดยโปรตีนขนส่งแบบแอคทีฟ[ 41 ] [ 42 ]
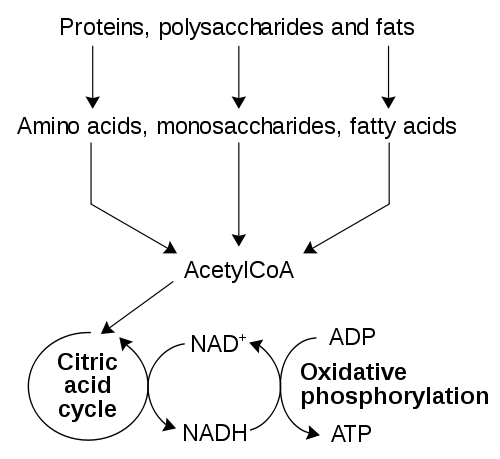
พลังงานจากสารประกอบอินทรีย์
การสลายคาร์โบไฮเดรตคือการสลายคาร์โบไฮเดรตให้เป็นหน่วยย่อยที่เล็กลง โดยปกติคาร์โบไฮเดรตจะถูกนำเข้าสู่เซลล์หลังจากที่ถูกย่อยเป็นโมโนแซ็กคาไรด์เช่นกลูโคสและฟรุกโตส [ 45 ] เมื่อเข้าไปภายในเซลล์แล้ว เส้นทางการสลายหลักคือไกลโคไลซิสซึ่งกลูโคสจะถูกเปลี่ยนเป็นไพรูเวท กระบวนการนี้สร้างโมเลกุล NADHที่นำพาพลังงานจาก NAD +และสร้างATPจากADPเพื่อใช้ในการขับเคลื่อนกระบวนการต่างๆ ภายในเซลล์[ 46 ]ไพรูเวทเป็นสารตัวกลางในเส้นทางการเผาผลาญหลายเส้นทาง แต่ส่วนใหญ่จะถูกเปลี่ยนเป็นอะเซทิล-CoAและป้อนเข้าสู่วัฏจักรกรดซิตริกซึ่งช่วยให้มีการผลิต ATP เพิ่มขึ้นโดยวิธีการฟอสโฟรีเลชันแบบออกซิเดชันการออกซิเดชันนี้ใช้ออกซิเจนโมเลกุลและปล่อยน้ำและคาร์บอนไดออกไซด์ซึ่งเป็นของเสีย เมื่อขาดออกซิเจน หรือเมื่อไพรูเวทถูกผลิตขึ้นเร็วกว่าที่วัฏจักรกรดซิตริกจะบริโภคได้ชั่วคราว (เช่น ในการออกกำลังกายอย่างหนัก) ไพรูเวทจะถูกเปลี่ยนเป็นแลคเตทโดยเอนไซม์แลคเตทดีไฮโดร จีเนส ซึ่งเป็นกระบวนการที่ออกซิไดซ์ NADH กลับไปเป็น NAD +เพื่อนำกลับมาใช้ใหม่ในกระบวนการไกลโคไลซิส ทำให้การผลิตพลังงานดำเนินต่อไปได้[ 47 ]ต่อมาแลคเตทจะถูกเปลี่ยนกลับไปเป็นไพรูเวทเพื่อผลิต ATP ในบริเวณที่ต้องการพลังงาน หรือกลับไปเป็นกลูโคสในวัฏจักรโคริเส้นทางทางเลือกสำหรับการสลายกลูโคสคือเส้นทางเพนโทสฟอสเฟตซึ่งผลิตพลังงานได้น้อยกว่า แต่สนับสนุนการสร้างสารชีวภาพ (การสังเคราะห์โมเลกุลชีวภาพ) เส้นทางนี้จะลดโคเอนไซม์NADP +ให้เป็น NADPH และผลิต สารประกอบ เพนโทสเช่นไรโบส 5-ฟอสเฟตสำหรับการสังเคราะห์โมเลกุลชีวภาพหลายชนิด เช่นนิวคลีโอไทด์และ กรดอะมิโนอะโร มาติก[ 48 ]

ไขมันจะถูกย่อยสลายด้วยไฮโดรไลซิสเป็นกรดไขมันอิสระและกลีเซอรอล กลีเซอรอลจะเข้าสู่ไกลโคไลซิส และกรดไขมันจะถูกย่อยสลายด้วยเบต้าออกซิเดชันเพื่อปลดปล่อยอะเซทิล-โคเอ ซึ่งจากนั้นจะถูกป้อนเข้าสู่วัฏจักรกรดซิตริก กรดไขมันจะปลดปล่อยพลังงานมากกว่าคาร์โบไฮเดรตเมื่อเกิดออกซิเดชัน สเตียรอยด์ยังถูกย่อยสลายโดยแบคทีเรียบางชนิดในกระบวนการที่คล้ายกับเบต้าออกซิเดชัน และกระบวนการย่อยสลายนี้เกี่ยวข้องกับการปลดปล่อยอะเซทิล-โคเอ โพรพิโอนิล-โคเอ และไพรูเวตในปริมาณมาก ซึ่งเซลล์สามารถนำไปใช้เป็นพลังงานได้ทั้งหมดM. tuberculosisยังสามารถเจริญเติบโตบนคอเลสเตอรอลไขมันเป็นแหล่งคาร์บอนเพียงอย่างเดียว และยีนที่เกี่ยวข้องกับเส้นทางการใช้คอเลสเตอรอลได้รับการยืนยันว่ามีความสำคัญในช่วงต่างๆ ของวงจรชีวิตการติดเชื้อของM. tuberculosis [ 49 ]
กรดอะมิโนถูกนำไปใช้ในการสังเคราะห์โปรตีนและโมเลกุลชีวภาพอื่นๆ หรือถูกออกซิไดซ์เป็นยูเรียและคาร์บอนไดออกไซด์เพื่อผลิตพลังงาน[ 50 ]เส้นทางการออกซิเดชันเริ่มต้นด้วยการกำจัดหมู่เอมีนโดยทรานส์อะมิเนส หมู่เอมีนจะถูกป้อนเข้าสู่วัฏจักรยูเรียเหลือโครงสร้างคาร์บอนที่ปราศจากหมู่เอมีนในรูปของกรดคีโต กรดคีโตหลายชนิดเหล่านี้เป็นสารตัวกลางในวัฏจักรกรดซิตริก ตัวอย่างเช่น α- คีโตกลูตาเรตที่เกิดจากการกำจัดหมู่เอมีนของกลูตาเมต [ 51 ] กรดอะมิโนที่สร้างกลูโคสยังสามารถเปลี่ยนเป็นกลูโคสได้ผ่านกระบวนการสร้างกลูโคสใหม่[ 52 ]
การแปลงพลังงาน
การฟอสโฟรีเลชันแบบออกซิเดทีฟ
ในกระบวนการฟอสโฟรีเลชันแบบออกซิเดชัน อิเล็กตรอนที่ถูกกำจัดออกจากโมเลกุลอินทรีย์ในบริเวณต่างๆ เช่น วัฏจักรกรดซิตริก จะถูกถ่ายโอนไปยังออกซิเจน และพลังงานที่ปล่อยออกมาจะถูกนำไปใช้ในการสร้าง ATP กระบวนการนี้เกิดขึ้นในยูคาริโอตโดยโปรตีนหลายชนิดในเยื่อหุ้มไมโทคอนเดรียที่เรียกว่าห่วงโซ่การขนส่งอิเล็กตรอนใน โปร คาริโอต โปรตีนเหล่านี้พบได้ใน เยื่อหุ้มชั้นในของเซลล์[ 53 ]โปรตีนเหล่านี้ใช้พลังงานจากโมเลกุลที่ลดลง เช่น NADH เพื่อ สูบโปรตอนข้ามเยื่อหุ้มเซลล์[ 54 ]

การสูบโปรตอนออกจากไมโตคอนเดรียทำให้เกิดความแตกต่างของความเข้มข้น ของโปรตอน ข้ามเยื่อหุ้มเซลล์และสร้างความลาดชันทางไฟฟ้าเคมี [ 55 ] แรงนี้จะผลักดันโปรตอนกลับเข้าไปในไมโตคอนเดรียผ่านฐานของเอนไซม์ที่เรียกว่าATP synthaseการไหลของโปรตอนทำให้หน่วยย่อยก้านหมุน ทำให้บริเวณที่ใช้งานของโดเมน synthase เปลี่ยนรูปร่างและฟอสโฟรีเลตอะดีโนซีนไดฟอสเฟตซึ่งเปลี่ยนเป็น ATP [ 21 ]
พลังงานจากสารประกอบอนินทรีย์
เคโม ลิโทโทรฟีเป็นเมตาบอลิซึมชนิดหนึ่งที่พบในโปรคาริโอตซึ่งได้รับพลังงานจากการออกซิเดชันของสารประกอบอนินทรีย์สิ่งมีชีวิตเหล่านี้สามารถใช้ไฮโดรเจน [ 56 ] สารประกอบ กำมะถันที่ลดลง(เช่นซัลไฟด์ไฮโดรเจนซัลไฟด์และไทโอซัลเฟต ) [ 1 ]เหล็กเฟอร์รัส (Fe(II)) [ 57 ]หรือแอมโมเนีย[ 58 ]เป็นแหล่งพลังงานรีดิวซ์ และได้รับพลังงานจากการออกซิเดชันของสารประกอบเหล่านี้[ 59 ]กระบวนการจุลินทรีย์เหล่านี้มีความสำคัญในวัฏจักรทางชีวธรณีเคมี ระดับโลก เช่นอะซิโตเจเนซิ ส ไน ตริฟิเคชันและดีไนตริฟิเคชันและมีความสำคัญต่อ ความอุดม สมบูรณ์ของดิน[ 60 ] [ 61 ]
พลังงานจากแสง
พืชไซยาโนแบคทีเรียแบคทีเรียสีม่วงแบคทีเรียกำมะถันสีเขียวและโปรติสต์บางชนิดสามารถดักจับพลังงานจากแสงแดดได้กระบวนการนี้มักเชื่อมโยงกับการเปลี่ยนคาร์บอนไดออกไซด์ให้เป็นสารประกอบอินทรีย์ ซึ่งเป็นส่วนหนึ่งของการสังเคราะห์แสง ซึ่งจะกล่าวถึงต่อไป ระบบการดักจับพลังงานและการตรึงคาร์บอนสามารถทำงานแยกกันได้ในโปรคาริโอต เช่น แบคทีเรียสีม่วงและแบคทีเรียกำมะถันสีเขียวสามารถใช้แสงแดดเป็นแหล่งพลังงาน ในขณะที่สลับระหว่างการตรึงคาร์บอนและการหมักสารประกอบอินทรีย์[ 62 ] [ 63 ]
ในสิ่งมีชีวิตหลายชนิด การจับพลังงานแสงอาทิตย์มีหลักการคล้ายคลึงกับการฟอสโฟรีเลชันแบบออกซิเดชัน เนื่องจากเกี่ยวข้องกับการเก็บพลังงานในรูปของความเข้มข้นของโปรตอน แรงขับเคลื่อนโปรตอนนี้จะขับเคลื่อนการสังเคราะห์ ATP [ 64 ]อิเล็กตรอนที่จำเป็นในการขับเคลื่อนห่วงโซ่การขนส่งอิเล็กตรอนนี้มาจากโปรตีนที่รวบรวมแสงที่เรียกว่าศูนย์ปฏิกิริยาการสังเคราะห์แสงศูนย์ปฏิกิริยาถูกจัดประเภทเป็นสองประเภทขึ้นอยู่กับลักษณะของรงควัตถุการสังเคราะห์แสงที่มีอยู่ โดยแบคทีเรียสังเคราะห์แสงส่วนใหญ่มีเพียงประเภทเดียว ในขณะที่พืชและไซยาโนแบคทีเรียมีสองประเภท[ 65 ]
ในพืช สาหร่าย และไซยาโนแบคทีเรียโฟโตซิสเต็ม IIใช้พลังงานแสงเพื่อกำจัดอิเล็กตรอนออกจากน้ำ ปล่อยออกซิเจนออกมาเป็นของเสีย จากนั้นอิเล็กตรอนจะไหลไปยังคอมเพล็กซ์ไซโตโครม b6fซึ่งใช้พลังงานของอิเล็กตรอนเหล่านั้นในการปั๊มโปรตอนข้าม เยื่อ ไทลาคอยด์ในคลอโรพลาสต์ [ 35 ] โปรตอนเหล่านี้จะเคลื่อนที่กลับผ่านเยื่อหุ้มเซลล์ในขณะที่ขับเคลื่อน ATP ซินเทสเช่นเดิม จากนั้นอิเล็กตรอนจะไหลผ่านโฟโตซิสเต็ม I และสามารถนำไปใช้ลดโคเอนไซม์ NADP +ได้[ 66 ]โคเอนไซม์นี้สามารถเข้าสู่รอบแคลวินหรือนำกลับมาใช้ใหม่เพื่อสร้าง ATP เพิ่มเติมได้[ 67 ]
อนาโบลิซึม
อนาโบลิซึมคือชุดของกระบวนการเผาผลาญเชิงสร้างสรรค์ โดยพลังงานที่ปล่อยออกมาจากแคตาโบลิซึมจะถูกนำมาใช้ในการสังเคราะห์โมเลกุลที่ซับซ้อน โดยทั่วไป โมเลกุลที่ซับซ้อนซึ่งประกอบเป็นโครงสร้างของเซลล์จะถูกสร้างขึ้นทีละขั้นตอนจากสารตั้งต้นที่เล็กกว่าและเรียบง่ายกว่า อนาโบลิซึมประกอบด้วยสามขั้นตอนพื้นฐาน ขั้นแรกคือการผลิตสารตั้งต้น เช่นกรดอะมิโนโมโนแซ็กคาไรด์ไอโซพรีนอยด์และนิวคลีโอไทด์ขั้นที่สองคือการกระตุ้นสารตั้งต้นเหล่านี้ให้เป็นรูปแบบที่ทำปฏิกิริยาได้โดยใช้พลังงานจาก ATP และขั้นที่สามคือการประกอบสารตั้งต้นเหล่านี้เข้าเป็นโมเลกุลที่ซับซ้อน เช่นโปรตีนโพลีแซ็กคาไร ด์ ลิปิดและกรดนิวคลีอิก[ 68 ]
กระบวนการสร้างสารในสิ่งมีชีวิตอาจแตกต่างกันไปตามแหล่งที่มาของโมเลกุลที่สร้างขึ้นในเซลล์ของพวกมัน สิ่งมีชีวิตที่สร้างอาหารเองได้เช่น พืช สามารถสร้างโมเลกุลอินทรีย์ที่ซับซ้อนในเซลล์ของพวกมัน เช่น โพลีแซ็กคาไรด์และโปรตีน จากโมเลกุลที่เรียบง่าย เช่นคาร์บอนไดออกไซด์และน้ำ ในทางกลับกัน สิ่งมีชีวิตที่ต้อง พึ่งพาอาหาร จากภายนอก จำเป็นต้องมีแหล่งของสารที่ซับซ้อนกว่า เช่น โมโนแซ็กคาไรด์และกรดอะมิโน เพื่อผลิตโมเลกุลที่ซับซ้อนเหล่านี้ สิ่งมีชีวิตยังสามารถจำแนกได้ตามแหล่งพลังงานขั้นสุดท้ายของพวกมัน: สิ่งมีชีวิตที่สร้างอาหารเองได้และสิ่งมีชีวิตที่ต้องพึ่งพาอาหารจากภายนอก ได้รับพลังงานจากแสง ในขณะที่สิ่งมีชีวิตที่สร้างอาหารเองได้และสิ่งมีชีวิตที่ต้องพึ่งพาอาหารจากภายนอก ได้รับพลังงานจากปฏิกิริยาออกซิเดชัน[ 68 ]
การตรึงคาร์บอน
การสังเคราะห์แสงคือการสังเคราะห์คาร์โบไฮเดรตจากแสงแดดและคาร์บอนไดออกไซด์ (CO2 )ในพืช ไซยาโนแบคทีเรีย และสาหร่าย การสังเคราะห์แสงแบบออกซิเจนจะแยกน้ำ โดยมีออกซิเจนเป็นของเสีย กระบวนการนี้ใช้ ATP และ NADPH ที่ผลิตโดยศูนย์ปฏิกิริยาการสังเคราะห์แสงดังที่ได้อธิบายไว้ข้างต้น เพื่อเปลี่ยน CO2 ให้เป็นกลีเซอเรต 3-ฟอสเฟตซึ่งสามารถเปลี่ยนเป็นกลูโคสได้ ปฏิกิริยาการตรึงคาร์บอนนี้ดำเนินการโดยเอนไซม์RuBisCOซึ่งเป็นส่วนหนึ่งของวัฏจักรแคลวิน-เบนสัน [ 69 ] การสังเคราะห์แสงในพืชมี 3 ประเภท ได้แก่การตรึงคาร์บอนแบบ C3 การตรึงคาร์บอนแบบ C4และการสังเคราะห์แสงแบบ CAMซึ่งแตกต่างกันตามเส้นทางที่คาร์บอนไดออกไซด์เข้าสู่วัฏจักรแคลวิน โดยพืช C3 จะตรึง CO2 โดยตรงในขณะที่การสังเคราะห์แสงแบบ C4 และ CAM จะรวม CO2 เข้ากับสารประกอบอื่นก่อน ซึ่งเป็นการปรับตัวเพื่อรับมือกับแสงแดดจัดและสภาพแห้งแล้ง[ 70 ]
ในโปรคาริโอต ที่สังเคราะห์ แสง กลไกการตรึงคาร์บอนมีความหลากหลายมากขึ้น ที่นี่ คาร์บอนไดออกไซด์สามารถถูกตรึงโดยวัฏจักรแคลวิน-เบนสันวัฏจักรกรดซิตริกแบบย้อนกลับ[ 71 ]หรือการคาร์บอกซิเลชันของอะเซทิล-โคเอ[ 72 ] [ 73 ]เคโมออโตโทรฟโปรคาริโอตยังตรึง CO 2ผ่านวัฏจักรแคลวิน-เบนสัน แต่ใช้พลังงานจากสารประกอบอนินทรีย์เพื่อขับเคลื่อนปฏิกิริยา[ 74 ]
คาร์โบไฮเดรตและไกลแคน
ในการสร้างคาร์โบไฮเดรต กรดอินทรีย์อย่างง่ายสามารถเปลี่ยนเป็นโมโนแซ็กคาไรด์เช่นกลูโคสแล้วนำไปใช้ในการประกอบพอลิแซ็กคาไรด์เช่นแป้งการสร้างกลูโคสจากสารประกอบต่างๆ เช่นไพรูเวต แลคเต ต กลีเซอ รอล กลีเซอเร ต3-ฟอสเฟตและกรดอะมิโนเรียกว่ากลูโคเนโอเจเน ซิส กลูโคเนโอเจเนซิสเปลี่ยนไพรูเวตเป็นกลูโคส-6-ฟอสเฟตผ่านตัวกลางหลายชุด ซึ่งหลายชุดใช้ร่วมกับไกลโคไลซิส [ 46 ] อย่างไรก็ตามเส้นทางนี้ไม่ใช่เพียงแค่ไกลโคไลซิสที่ดำเนินการย้อนกลับ เนื่องจากหลายขั้นตอนถูกเร่งปฏิกิริยาโดยเอนไซม์ที่ไม่ใช่ไกลโคไลซิส นี่เป็นสิ่งสำคัญเพราะช่วยให้การสร้างและการสลายตัวของกลูโคสได้รับการควบคุมแยกจากกัน และป้องกันไม่ให้ทั้งสองเส้นทางทำงานพร้อมกันในวงจรที่ไร้ประโยชน์ [ 75 ] [ 76 ]
แม้ว่าไขมันจะเป็นวิธีทั่วไปในการเก็บสะสมพลังงาน แต่ในสัตว์มีกระดูกสันหลังเช่น มนุษย์กรดไขมันในแหล่งสะสมเหล่านี้ไม่สามารถเปลี่ยนเป็นกลูโคสผ่านกระบวนการสร้างกลูโคสใหม่ได้เนื่องจากสิ่งมีชีวิตเหล่านี้ไม่สามารถเปลี่ยนอะเซทิล-โคเอเป็นไพรูเวตได้ พืชทำได้ แต่สัตว์ไม่มีกลไกเอนไซม์ที่จำเป็น[ 77 ]ด้วยเหตุนี้ หลังจากการอดอาหารเป็นเวลานาน สัตว์มีกระดูกสันหลังจำเป็นต้องสร้างคีโตนจากกรดไขมันเพื่อทดแทนกลูโคสในเนื้อเยื่อ เช่น สมองที่ไม่สามารถเผาผลาญกรดไขมันได้[ 78 ]ในสิ่งมีชีวิตอื่นๆ เช่น พืชและแบคทีเรีย ปัญหาการเผาผลาญนี้ได้รับการแก้ไขโดยใช้วัฏจักรไกลออกซิเลตซึ่งข้าม ขั้นตอน การดีคาร์บอกซิเลชันในวัฏจักรกรดซิตริกและอนุญาตให้เปลี่ยนอะเซทิล-โคเอเป็นออกซาโลอะซิเตตซึ่งสามารถนำไปใช้ในการผลิตกลูโคสได้[ 77 ] [ 79 ]นอกเหนือจากไขมันแล้ว กลูโคสจะถูกเก็บสะสมไว้ในเนื้อเยื่อส่วนใหญ่ในฐานะแหล่งพลังงานที่มีอยู่ในเนื้อเยื่อผ่านกระบวนการสร้างไกลโคเจน ซึ่งโดยปกติจะใช้เพื่อรักษาระดับกลูโคสในเลือด[ 80 ]
พอลิแซ็กคาไรด์และไกลแคนถูกสร้างขึ้นโดยการเติมโมโนแซ็กคาไรด์ตามลำดับโดยไกลโคซิลทรานสเฟอเรสจากตัวให้น้ำตาลฟอสเฟตที่ทำปฏิกิริยาได้ เช่นยูริดีนไดฟอสเฟตกลูโคส (UDP-Glc) ไปยัง หมู่ ไฮดรอกซิล ที่เป็นตัวรับ บนพอลิแซ็กคาไรด์ที่กำลังเติบโต เนื่องจากหมู่ไฮดรอกซิลใดๆ บนวงแหวนของสารตั้งต้นสามารถเป็นตัวรับได้ พอลิแซ็กคาไรด์ที่ผลิตได้จึงอาจมีโครงสร้างแบบเส้นตรงหรือแบบแตกแขนง[ 81 ]พอลิแซ็กคาไรด์ที่ผลิตได้อาจมีหน้าที่ทางโครงสร้างหรือทางเมตาบอลิซึมเอง หรือถูกถ่ายโอนไปยังลิปิดและโปรตีนโดยเอนไซม์โอลิโกแซ็กคาริลทรานสเฟอเรส[ 82 ] [ 83 ]
กรดไขมัน ไอโซพรีนอยด์ และสเตอรอล

กรดไขมันถูกสร้างขึ้นโดยเอนไซม์สังเคราะห์กรดไขมันที่ทำการพอลิเมอไรซ์แล้วลดหน่วยอะเซทิล-โคเอ โซ่อะซิลในกรดไขมันจะถูกขยายออกไปโดยวัฏจักรของปฏิกิริยาที่เพิ่มหมู่อะซิล ลดให้เป็นแอลกอฮอล์กำจัดน้ำออกเพื่อให้ได้ หมู่ แอลคีนแล้วลดอีกครั้งให้เป็น หมู่ แอลเคนเอนไซม์ในการสังเคราะห์กรดไขมันแบ่งออกเป็นสองกลุ่ม: ในสัตว์และเชื้อรา ปฏิกิริยาสังเคราะห์กรดไขมันทั้งหมดนี้ดำเนินการโดยโปรตีนประเภท I ที่มีหลายหน้าที่เพียงตัวเดียว[ 84 ]ในขณะที่ในพลาสติด ของพืช และแบคทีเรีย เอนไซม์ประเภท II ที่แยกกันจะทำแต่ละขั้นตอนในเส้นทาง[ 85 ] [ 86 ]
เทอร์พีนและไอโซพรีนอยด์เป็นลิปิดกลุ่มใหญ่ที่รวมถึงแคโรทีนอยด์และเป็นกลุ่มผลิตภัณฑ์ธรรมชาติจาก พืชที่ใหญ่ที่สุด [ 87 ]สารประกอบเหล่านี้สร้างขึ้นโดยการประกอบและการดัดแปลง หน่วย ไอโซพรีนที่ได้รับจากสารตั้งต้นที่ทำปฏิกิริยาได้ เช่น ไอโซเพนเทนิลไพโรฟอสเฟตและไดเมทิลอัลลิลไพโรฟอสเฟต [ 88 ] สารตั้งต้นเหล่านี้สามารถสร้างขึ้นได้หลายวิธี ในสัตว์และอาร์เคียเส้นทางเมวาโลเนตจะสร้างสารประกอบเหล่านี้จากอะเซทิล-โคเอ[ 89 ]ในขณะที่ในพืชและแบคทีเรียเส้นทางที่ไม่ใช่เมวาโลเนตจะใช้ไพรูเวตและ กลีเซอรั ลดีไฮด์ 3-ฟอสเฟตเป็นสารตั้งต้น[ 88 ] [ 90 ]ปฏิกิริยาสำคัญอย่างหนึ่งที่ใช้ผู้ให้ไอโซพรีนที่ถูกกระตุ้นเหล่านี้คือการสังเคราะห์สเตอรอลในที่นี้ หน่วยไอโซพรีนจะถูกรวมเข้าด้วยกันเพื่อสร้างสควาเลนจากนั้นพับขึ้นและสร้างเป็นวงแหวนเพื่อสร้างแลโนสเตอรอล[ 91 ]จากนั้นแลโนสเตอรอลสามารถแปลงเป็นสเตอรอลอื่นๆ เช่นคอเลสเตอรอลและเออร์โกสเตอรอลได้[ 91 ] [ 92 ]
โปรตีน
สิ่งมีชีวิตมีความสามารถในการสังเคราะห์กรดอะมิโนทั่วไป 20 ชนิดแตกต่างกัน แบคทีเรียและพืชส่วนใหญ่สามารถสังเคราะห์ได้ครบทั้ง 20 ชนิด แต่สัตว์เลี้ยงลูกด้วยนมสามารถสังเคราะห์ได้เพียง 11 กรดอะมิโนที่ไม่จำเป็น ดังนั้นกรดอะมิโนที่จำเป็นอีก 9 ชนิดจึง ต้องได้รับจากอาหาร[ 11 ]ปรสิตบางชนิดเช่น แบคทีเรียMycoplasma pneumoniaeขาดความสามารถในการสังเคราะห์กรดอะมิโนทั้งหมดและรับกรดอะมิโนโดยตรงจากโฮสต์[ 93 ]กรดอะมิโนทั้งหมดถูกสังเคราะห์จากสารตัวกลางในกระบวนการไกลโคไลซิส วงจรกรดซิตริก หรือวิถีเพนโทสฟอสเฟต ไนโตรเจนได้มาจากกลูตาเมตและกลูตา มี น การสังเคราะห์กรดอะมิโนที่ไม่จำเป็นขึ้นอยู่กับการสร้างกรดอัลฟา-คีโตที่เหมาะสม ซึ่งจากนั้นจะถูกทรานส์อะมิเนตเพื่อสร้างกรดอะมิโน[ 94 ]
กรดอะมิโนถูกสร้างเป็นโปรตีนโดยการเชื่อมต่อกันเป็นสายโซ่ของพันธะเปปไทด์โปรตีนแต่ละชนิดมีลำดับของกรดอะมิโนที่ไม่ซ้ำกัน ซึ่งก็คือโครงสร้างหลักของโปรตีนนั้น เช่นเดียวกับตัวอักษรที่สามารถนำมาประกอบกันเป็นคำได้หลากหลายแทบไม่มีที่สิ้นสุด กรดอะมิโนก็สามารถเชื่อมต่อกันในลำดับที่แตกต่างกันเพื่อสร้างโปรตีนได้หลากหลายชนิด โปรตีนถูกสร้างขึ้นจากกรดอะมิโนที่ถูกกระตุ้นโดยการเชื่อมต่อกับ โมเลกุล transfer RNAผ่าน พันธะ เอสเท อร์ สารตั้งต้น aminoacyl-tRNAนี้ถูกผลิตขึ้นใน ปฏิกิริยาที่ขึ้นอยู่กับ ATPซึ่งดำเนินการโดยaminoacyl tRNA synthetase [ 95 ] จากนั้น aminoacyl-tRNA นี้จะเป็นสารตั้งต้นสำหรับไรโบโซม ซึ่งจะเชื่อมต่อกรดอะมิโนเข้ากับสายโปรตีนที่ กำลังยืดออก โดยใช้ข้อมูลลำดับในmessenger RNA [ 96 ]
การสังเคราะห์และการกู้คืนนิวคลีโอไทด์
นิวคลีโอไทด์ถูกสร้างขึ้นจากกรดอะมิโน คาร์บอนไดออกไซด์ และกรดฟอร์มิกในวิถีทางที่ต้องใช้พลังงานเมตาบอลิซึมจำนวนมาก[ 97 ]ด้วยเหตุนี้ สิ่งมีชีวิตส่วนใหญ่จึงมีระบบที่มีประสิทธิภาพในการนำนิวคลีโอไทด์ที่สร้างขึ้นแล้วกลับมาใช้ใหม่[ 97 ] [ 98 ]พิวรีนถูกสังเคราะห์เป็นนิวคลีโอไซด์ (เบสที่ติดอยู่กับไรโบส ) [ 99 ]ทั้งอะดีนีนและกัวนีนถูกสร้างขึ้นจากนิวคลีโอไซด์ตั้งต้นอิโนซีนโมโนฟอสเฟต ซึ่งถูกสังเคราะห์โดยใช้อะตอมจากกรดอะมิโนไกลซีน กลูตามีนและกรดแอสปาร์ติกรวมถึงฟอร์เมตที่ถ่ายโอนมาจากโคเอนไซม์ เตตระ ไฮโดรโฟเลตในทางกลับกันไพริมิดีน ถูกสังเคราะห์จากเบส ออโรเทตซึ่งเกิดจากกลูตามีนและแอสปาร์เทต[ 100 ]
สารแปลกปลอมและการเผาผลาญรีดอกซ์
สิ่งมีชีวิตทุกชนิดต่างสัมผัสกับสารประกอบที่ไม่สามารถนำไปใช้เป็นอาหารได้ และหากสะสมอยู่ในเซลล์ก็จะเป็นอันตราย เนื่องจากไม่มีหน้าที่ในการเผาผลาญ สารประกอบที่อาจเป็นอันตรายเหล่านี้เรียกว่าซีโนไบโอติกส์ [ 101 ] ซีโนไบโอติกส์ เช่นยาสังเคราะห์สารพิษจากธรรมชาติและยาปฏิชีวนะจะถูกกำจัดพิษโดยเอนไซม์ที่ทำหน้าที่เผาผลาญซีโนไบโอติกส์ ในมนุษย์ เอนไซม์เหล่านี้ได้แก่ไซโตโครม P450 ออกซิเดส [ 102 ] ยูดีพี-กลูคูโรโนซิลทราน สเฟอเร ส[ 103 ]และกลูตาไธโอนเอส -ทรานสเฟอเรส[ 104 ]ระบบเอนไซม์นี้ทำงานในสามขั้นตอน ขั้นแรกจะออกซิไดซ์ซีโนไบโอติกส์ (ขั้นตอนที่ 1) จากนั้นจึงเชื่อมต่อกลุ่มที่ละลายน้ำได้เข้ากับโมเลกุล (ขั้นตอนที่ 2) ซีโนไบโอติกส์ที่ละลายน้ำได้ซึ่งได้รับการดัดแปลงแล้วสามารถถูกขับออกจากเซลล์ และในสิ่งมีชีวิตหลายเซลล์อาจถูกเผาผลาญต่อไปก่อนที่จะถูกขับออก (ขั้นตอนที่ 3) ในทางนิเวศวิทยาปฏิกิริยาเหล่านี้มีความสำคัญอย่างยิ่งในการย่อยสลายสารมลพิษโดยจุลินทรีย์และการบำบัดทางชีวภาพของดินที่ปนเปื้อนและคราบน้ำมัน[ 105 ]ปฏิกิริยาของจุลินทรีย์เหล่านี้หลายอย่างมีร่วมกับสิ่งมีชีวิตหลายเซลล์ แต่เนื่องจากความหลากหลายของชนิดของจุลินทรีย์ที่มากมาย สิ่งมีชีวิตเหล่านี้จึงสามารถจัดการกับสารแปลกปลอมได้หลากหลายกว่าสิ่งมีชีวิตหลายเซลล์ และสามารถย่อยสลายสารมลพิษอินทรีย์ที่คงทนเช่นสารประกอบออร์กาโนคลอรีน ได้ [ 106 ]
ปัญหาที่เกี่ยวข้องสำหรับสิ่งมีชีวิตแอโรบิกคือความเครียดจากออกซิเดชัน [ 107 ] ในที่นี้ กระบวนการต่างๆ รวมถึงการฟอสโฟรีเลชันแบบออกซิเดชันและการสร้างพันธะไดซัลไฟด์ระหว่างการพับโปรตีนจะสร้างอนุมูลออกซิเจนเช่นไฮโดรเจนเปอร์ออกไซด์ [ 108 ] สารออกซิแดนต์ที่เป็นอันตรายเหล่านี้จะถูกกำจัดโดย เมตาบอไลต์ ต้านอนุมูลอิสระเช่นกลูตาไธโอนและเอนไซม์ เช่นคาตาเลสและเพอร์ออกซิเดส[ 109 ] [ 110 ]
อุณหพลศาสตร์ของสิ่งมีชีวิต
สิ่งมีชีวิตต้องปฏิบัติตามกฎของอุณหพลศาสตร์ซึ่งอธิบายถึงการถ่ายเทความร้อนและงานกฎข้อที่สองของอุณหพลศาสตร์ระบุว่า ในระบบที่แยกตัวออกปริมาณเอนโทรปี (ความไม่เป็นระเบียบ) ไม่สามารถลดลงได้ แม้ว่าความซับซ้อนอันน่าทึ่งของสิ่งมีชีวิตจะดูเหมือนขัดแย้งกับกฎนี้ แต่ชีวิตก็เป็นไปได้ เนื่องจากสิ่งมีชีวิตทั้งหมดเป็นระบบเปิดที่แลกเปลี่ยนสสารและพลังงานกับสิ่งแวดล้อม ระบบสิ่งมีชีวิตไม่ได้อยู่ในสมดุลแต่เป็นระบบที่กระจายพลังงานซึ่งรักษาสถานะความซับซ้อนสูงโดยการทำให้เอนโทรปีของสิ่งแวดล้อมเพิ่มขึ้นมากขึ้น[ 111 ]การเผาผลาญของเซลล์บรรลุเป้าหมายนี้โดยการเชื่อมโยงกระบวนการสลายตัวที่เกิดขึ้นเองตามธรรมชาติเข้ากับกระบวนการสร้างที่ไม่เกิดขึ้นเองตามธรรมชาติ ใน แง่ ของอุณหพลศาสตร์การเผาผลาญรักษาความเป็นระเบียบโดยการสร้างความไม่เป็นระเบียบ[ 112 ]
กฎระเบียบและการควบคุม
เนื่องจากสภาพแวดล้อมของสิ่งมีชีวิตส่วนใหญ่มีการเปลี่ยนแปลงอยู่ตลอดเวลา ปฏิกิริยาของการเผาผลาญจึงต้องได้รับการควบคุม อย่างละเอียด เพื่อรักษาสภาวะคงที่ภายในเซลล์ ซึ่งเรียกว่าภาวะสมดุล[ 113 ] [ 114 ] การควบคุมการเผาผลาญยังช่วยให้สิ่งมีชีวิตสามารถตอบสนองต่อสัญญาณและมีปฏิสัมพันธ์กับสภาพแวดล้อมได้อย่าง กระตือรือร้น [ 115 ]แนวคิดสองประการที่เชื่อมโยงกันอย่างใกล้ชิดมีความสำคัญต่อการทำความเข้าใจว่าวิถีการเผาผลาญถูกควบคุมอย่างไร ประการแรก การควบคุมเอนไซม์ในวิถีการเผาผลาญคือการที่กิจกรรมของเอนไซม์เพิ่มขึ้นและลดลงเพื่อตอบสนองต่อสัญญาณ ประการที่สองการควบคุมที่เอนไซม์นี้กระทำคือผลกระทบที่การเปลี่ยนแปลงในกิจกรรมของเอนไซม์มีต่ออัตราโดยรวมของวิถีการเผาผลาญ ( การไหลผ่านวิถีการเผาผลาญ) [ 116 ]ตัวอย่างเช่น เอนไซม์อาจแสดงการเปลี่ยนแปลงกิจกรรมอย่างมาก (เช่น มีการควบคุมอย่างเข้มงวด) แต่ถ้าการเปลี่ยนแปลงเหล่านี้มีผลกระทบเพียงเล็กน้อยต่อการไหลของวิถีการเผาผลาญ เอนไซม์นี้ก็ไม่ได้เกี่ยวข้องกับการควบคุมวิถีการเผาผลาญ[ 117 ]

การควบคุมการเผาผลาญมีหลายระดับ ในการควบคุมภายใน วิถีการเผาผลาญจะควบคุมตัวเองเพื่อตอบสนองต่อการเปลี่ยนแปลงระดับของสารตั้งต้นหรือผลิตภัณฑ์ ตัวอย่างเช่น การลดลงของปริมาณผลิตภัณฑ์สามารถเพิ่มอัตราการไหลผ่านวิถีการเผาผลาญเพื่อชดเชยได้[ 116 ]การควบคุมประเภทนี้มักเกี่ยวข้องกับการควบคุมแบบอัลโลสเตอริกของกิจกรรมของเอนไซม์หลายตัวในวิถี การเผาผลาญ [ 119 ]การควบคุมภายนอกเกี่ยวข้องกับเซลล์ในสิ่งมีชีวิตหลายเซลล์ที่เปลี่ยนแปลงการเผาผลาญของตนเองเพื่อตอบสนองต่อสัญญาณจากเซลล์อื่น สัญญาณเหล่านี้มักอยู่ในรูปของสารสื่อประสาทที่ละลายน้ำได้ เช่นฮอร์โมนและปัจจัยการเจริญเติบโตและถูกตรวจจับโดยตัวรับ เฉพาะ บนพื้นผิวเซลล์[ 120 ]จากนั้นสัญญาณเหล่านี้จะถูกส่งผ่านภายในเซลล์โดยระบบตัวสื่อประสาทรองซึ่งมักเกี่ยวข้องกับการฟอสโฟรีเลชันของโปรตีน[ 121 ]
ตัวอย่างที่เข้าใจกันดีของการควบคุมภายนอกคือการควบคุมการเผาผลาญกลูโคสโดยฮอร์โมนอินซูลิน[ 122 ] อินซูลินถูกผลิตขึ้นเพื่อตอบสนองต่อระดับกลูโคสในเลือด ที่สูงขึ้น การจับตัวของฮอร์โมนกับตัวรับอินซูลินบนเซลล์จะกระตุ้นลำดับของโปรตีนไคเน ส ที่ทำให้เซลล์ดูดซึมกลูโคสและเปลี่ยนเป็นโมเลกุลสำหรับเก็บสะสม เช่น กรดไขมันและไกลโคเจน[ 123 ]การเผาผลาญไกลโคเจนถูกควบคุมโดยการทำงานของฟอสโฟริเลสซึ่งเป็นเอนไซม์ที่สลายไกลโคเจน และไกลโคเจนซินเทส ซึ่งเป็นเอนไซม์ที่สร้างไกลโคเจน เอนไซม์เหล่านี้ถูกควบคุมในลักษณะที่ผกผันกัน โดยการ ฟอสโฟริเลชันจะยับยั้งไกลโคเจนซินเทส แต่จะกระตุ้นฟอสโฟริเลส อินซูลินทำให้เกิดการสังเคราะห์ไกลโคเจนโดยการกระตุ้นโปรตีนฟอสฟาเทสและทำให้การฟอสโฟริเลชันของเอนไซม์เหล่านี้ลดลง[ 124 ]
วิวัฒนาการ

เส้นทางหลักของการเผาผลาญที่อธิบายไว้ข้างต้น เช่น ไกลโคไลซิสและวัฏจักรกรดซิตริก มีอยู่ในสิ่งมีชีวิตทั้งสามโดเมนและมีอยู่ในบรรพบุรุษร่วมสากลสุดท้าย [ 4 ] [ 125 ] เซลล์บรรพบุรุษสากลนี้เป็นโปรคาริโอตและน่าจะเป็นเมทาโนเจนที่มีการเผาผลาญกรดอะมิโน นิวคลีโอไทด์ คาร์โบไฮเดรต และไขมันอย่างกว้างขวาง[ 126 ] [ 127 ]การคงอยู่ของเส้นทางโบราณเหล่านี้ในระหว่างวิวัฒนาการ ในภายหลัง อาจเป็นผลมาจากการที่ปฏิกิริยาเหล่านี้เป็นทางออกที่ดีที่สุดสำหรับปัญหาการเผาผลาญเฉพาะของพวกมัน โดยเส้นทางต่างๆ เช่น ไกลโคไลซิสและวัฏจักรกรดซิตริกสร้างผลิตภัณฑ์สุดท้ายได้อย่างมีประสิทธิภาพสูงและในจำนวนขั้นตอนที่น้อยที่สุด[ 5 ] [ 6 ]เส้นทางแรกของการเผาผลาญโดยใช้เอนไซม์อาจเป็นส่วนหนึ่งของ การเผาผลาญ พิวรีนนิวคลีโอไทด์ ในขณะที่เส้นทางการเผาผลาญก่อนหน้านี้เป็นส่วนหนึ่งของโลก RNAโบราณ[ 128 ]
มีการเสนอแบบจำลองหลายแบบเพื่ออธิบายกลไกที่ทำให้เกิดวิวัฒนาการของวิถีเมตาบอลิซึมใหม่ ซึ่งรวมถึงการเพิ่มเอนไซม์ใหม่ตามลำดับลงในวิถีบรรพบุรุษสั้นๆ การทำซ้ำแล้วแยกตัวออกของวิถีทั้งหมด รวมถึงการดึงเอนไซม์ที่มีอยู่แล้วมาใช้และประกอบกันเป็นวิถีปฏิกิริยาใหม่[ 129 ]ความสำคัญสัมพัทธ์ของกลไกเหล่านี้ยังไม่ชัดเจน แต่การศึกษาทางจีโนมิกส์แสดงให้เห็นว่าเอนไซม์ในวิถีมีแนวโน้มที่จะมีบรรพบุรุษร่วมกัน ซึ่งบ่งชี้ว่าวิถีหลายๆ วิถีได้วิวัฒนาการทีละขั้นตอนด้วยฟังก์ชันใหม่ที่สร้างขึ้นจากขั้นตอนที่มีอยู่แล้วในวิถี[ 130 ]แบบจำลองทางเลือกมาจากการศึกษาที่ติดตามวิวัฒนาการของโครงสร้างโปรตีนในเครือข่ายเมตาบอลิซึม ซึ่งชี้ให้เห็นว่าเอนไซม์ถูกดึงมาใช้อย่างแพร่หลาย โดยยืมเอนไซม์เพื่อทำหน้าที่คล้ายกันในวิถีเมตาบอลิซึมที่แตกต่างกัน (เห็นได้ชัดในฐานข้อมูล MANET ) [ 131 ]กระบวนการดึงเอนไซม์เหล่านี้ส่งผลให้เกิดโมเสกของเอนไซม์ในเชิงวิวัฒนาการ[ 132 ]ความเป็นไปได้ประการที่สามคือบางส่วนของกระบวนการเผาผลาญอาจมีอยู่เป็น "โมดูล" ที่สามารถนำกลับมาใช้ใหม่ในเส้นทางต่างๆ และทำหน้าที่คล้ายกันกับโมเลกุลต่างๆ ได้[ 133 ]
นอกจากการวิวัฒนาการของวิถีการเผาผลาญใหม่แล้ว วิวัฒนาการยังอาจทำให้เกิดการสูญเสียหน้าที่การเผาผลาญได้อีกด้วย ตัวอย่างเช่น ในปรสิต บางชนิด กระบวนการเผาผลาญที่ไม่จำเป็นต่อการอยู่รอดจะหายไป และกรดอะมิโน นิวคลีโอไทด์ และคาร์โบไฮเดรตที่สร้างขึ้นแล้วอาจถูกเก็บเกี่ยวจากโฮสต์แทน [ 134 ] ความสามารถในการเผาผลาญที่ลดลงในลักษณะเดียวกันนี้พบได้ในสิ่งมีชีวิตที่อาศัยอยู่ร่วมกันแบบพึ่งพาอาศัยกัน[ 135 ]
การสืบสวนและการจัดการ

โดยทั่วไป การศึกษาเมตาบอลิซึมจะใช้ แนวทาง แบบลดทอนที่เน้นไปที่เส้นทางเมตาบอลิซึมเพียงเส้นทางเดียว การใช้สารติดตามกัมมันตรังสีในระดับสิ่งมีชีวิตทั้งหมด เนื้อเยื่อ และเซลล์นั้นมีคุณค่าอย่างยิ่ง เพราะสามารถกำหนดเส้นทางจากสารตั้งต้นไปสู่ผลิตภัณฑ์สุดท้ายได้โดยการระบุสารตัวกลางและผลิตภัณฑ์ที่ติดฉลากด้วยกัมมันตรังสี[ 136 ]จากนั้นจึงสามารถทำให้เอนไซม์ที่เร่งปฏิกิริยาเคมีเหล่านี้บริสุทธิ์และ ตรวจสอบ จลนศาสตร์และการตอบสนองต่อสารยับยั้งได้แนวทางคู่ขนานคือการระบุโมเลกุลขนาดเล็กในเซลล์หรือเนื้อเยื่อ ชุดโมเลกุลทั้งหมดเหล่านี้เรียกว่าเมตาโบโลมโดยรวมแล้ว การศึกษาเหล่านี้ให้มุมมองที่ดีเกี่ยวกับโครงสร้างและหน้าที่ของเส้นทางเมตาบอลิซึมแบบง่ายๆ แต่ไม่เพียงพอเมื่อนำไปใช้กับระบบที่ซับซ้อนกว่า เช่น เมตาบอลิซึมของเซลล์ทั้งหมด[ 137 ]
ภาพด้านขวาแสดงปฏิสัมพันธ์ระหว่างโปรตีนเพียง 43 ชนิดและเมตาบอไลต์ 40 ชนิด ซึ่งแสดงให้เห็นถึงความซับซ้อนของเครือข่ายเมตาบอลิซึม ในเซลล์ที่มีเอนไซม์แตกต่างกันหลายพันชนิด ลำดับของจีโนมให้รายการที่มีมากถึง 26,500 ยีน [ 138 ]อย่างไรก็ตาม ขณะนี้สามารถใช้ข้อมูลจีโนมนี้เพื่อสร้างเครือข่ายปฏิกิริยาทางชีวเคมีที่สมบูรณ์และสร้าง แบบจำลองทางคณิตศาสตร์แบบ องค์รวม มากขึ้น ซึ่งอาจอธิบายและทำนายพฤติกรรมของปฏิกิริยาเหล่านั้นได้ [ 139 ]แบบจำลองเหล่านี้มีประสิทธิภาพเป็นพิเศษเมื่อใช้ในการบูรณาการข้อมูลเส้นทางและเมตาบอไลต์ที่ได้จากวิธีการแบบดั้งเดิมเข้ากับข้อมูลการแสดงออกของยีนจากการศึกษาโปรตีโอมิกส์และไมโครอาร์เรย์ดีเอ็นเอ[ 140 ]ด้วยเทคนิคเหล่านี้ ขณะนี้ได้มีการสร้างแบบจำลองการเผาผลาญของมนุษย์ขึ้น ซึ่งจะชี้นำการค้นพบยาและการวิจัยทางชีวเคมีในอนาคต[ 141 ]ปัจจุบันแบบจำลองเหล่านี้ถูกนำมาใช้ในการวิเคราะห์เครือข่ายเพื่อจำแนกโรคของมนุษย์ออกเป็นกลุ่มที่ใช้โปรตีนหรือเมตาบอไลต์ร่วมกัน[ 142 ] [ 143 ]
เครือข่ายเมตาบอลิซึมของแบคทีเรียเป็นตัวอย่างที่โดดเด่นของ โครงสร้าง แบบโบว์ไท[ 144 ] [ 145 ] [ 146 ]ซึ่งเป็นสถาปัตยกรรมที่สามารถนำสารอาหารได้หลากหลายชนิดและผลิตผลิตภัณฑ์และโมเลกุลขนาดใหญ่ที่ซับซ้อนได้หลากหลายชนิดโดยใช้สกุลเงินกลางทั่วไปเพียงไม่กี่สกุล[ 147 ]
การประยุกต์ใช้ทางเทคโนโลยีที่สำคัญของข้อมูลนี้คือวิศวกรรมเมตาบอลิซึมในที่นี้ สิ่งมีชีวิต เช่นยีสต์พืช หรือแบคทีเรียจะถูกดัดแปลงพันธุกรรมเพื่อให้มีประโยชน์มากขึ้นในเทคโนโลยีชีวภาพสีแดงและช่วยในการผลิตยาเช่นยาปฏิชีวนะหรือสารเคมีอุตสาหกรรม เช่น1,3-โพรเพนไดออลและกรดชิกิมิก [ 148 ] [ 149 ] [ 150 ] การดัดแปลงพันธุกรรมเหล่านี้มักมีจุดมุ่งหมายเพื่อลดปริมาณพลังงานที่ใช้ในการผลิตผลิตภัณฑ์ เพิ่มผลผลิต และลดการผลิตของเสีย[ 151 ]
ประวัติศาสตร์
คำว่าเมตาบอลิซึมมาจาก คำภาษา กรีกโบราณ μεταβολή—"metabole" ซึ่งหมายถึง "การเปลี่ยนแปลง" ซึ่งมาจาก μεταβάλλειν—"metaballein" ซึ่งหมายถึง "การเปลี่ยนแปลง" [ 152 ]

ปรัชญากรีก
อริสโตเติลได้อธิบายรายละเอียดเกี่ยวกับมุมมองของเขาเกี่ยวกับการเผาผลาญอาหาร ไว้ในหนังสือ The Parts of Animals มากพอ ที่จะสร้างแบบจำลองการไหลแบบเปิดได้ เขาเชื่อว่าในแต่ละขั้นตอนของกระบวนการ วัสดุจากอาหารจะถูกเปลี่ยนแปลง โดยมีการปล่อยความร้อนออกมาในรูปของธาตุไฟ และสารตกค้างจะถูกขับออกมาในรูปของปัสสาวะ น้ำดี หรืออุจจาระ[ 153 ]
อิบนุ อัล-นาฟิสอธิบายกระบวนการเผาผลาญในงานเขียนของเขาในปี ค.ศ. 1260 ที่ชื่อว่าอัล-ริซาละฮ์ อัล-กามิลียะฮ์ ฟิล เซียรา อัล-นาบาวียะฮ์ (ตำราของกามิลเกี่ยวกับชีวประวัติของศาสดา) ซึ่งรวมถึงวลีต่อไปนี้ "ทั้งร่างกายและส่วนต่างๆ ของร่างกายอยู่ในสภาวะของการสลายตัวและการบำรุงเลี้ยงอย่างต่อเนื่อง ดังนั้นจึงหลีกเลี่ยงไม่ได้ที่จะมีการเปลี่ยนแปลงอย่างถาวร" [ 154 ]
การประยุกต์ใช้วิธีการทางวิทยาศาสตร์
ประวัติศาสตร์ของการศึกษาทางวิทยาศาสตร์เกี่ยวกับการเผาผลาญอาหารนั้นครอบคลุมหลายศตวรรษ และได้เปลี่ยนจากการตรวจสอบสัตว์ทั้งตัวในการศึกษายุคแรก มาเป็นการตรวจสอบปฏิกิริยาการเผาผลาญอาหารแต่ละอย่างในชีวเคมีสมัยใหม่การทดลอง ควบคุมครั้งแรก เกี่ยวกับการเผาผลาญอาหารในมนุษย์ได้รับการตีพิมพ์โดยSantorio Santorioในปี ค.ศ. 1614 ในหนังสือArs de statica medicina ของเขา เขาได้อธิบายวิธีการชั่งน้ำหนักตัวเองก่อนและหลังรับประทานอาหารนอนหลับทำงาน มีเพศสัมพันธ์ อดอาหาร ดื่ม และขับถ่าย เขาพบว่าอาหารส่วนใหญ่ที่เขารับประทานเข้าไปนั้นสูญเสียไปผ่านสิ่งที่เขาเรียกว่า " เหงื่อที่มองไม่เห็น " [ 155 ]

ในการศึกษาช่วงแรกๆ กลไกของกระบวนการเผาผลาญเหล่านี้ยังไม่ได้รับการระบุ และเชื่อกันว่าพลังชีวิต เป็นตัวขับเคลื่อนเนื้อเยื่อที่มีชีวิต [ 156 ]ในศตวรรษที่ 19 เมื่อศึกษาการหมักน้ำตาลเป็นแอลกอฮอล์โดยยีสต์หลุยส์ ปาสเตอร์สรุปว่าการหมักถูกเร่งปฏิกิริยาโดยสารภายในเซลล์ยีสต์ที่เขาเรียกว่า "สารหมัก" เขาเขียนว่า "การหมักแอลกอฮอล์เป็นกระบวนการที่สัมพันธ์กับชีวิตและการจัดระเบียบของเซลล์ยีสต์ ไม่ใช่กับการตายหรือการเน่าเปื่อยของเซลล์" [ 157 ] การค้นพบนี้ พร้อมกับการตีพิมพ์บทความเกี่ยวกับการสังเคราะห์ทางเคมีของ ยูเรียโดยฟรีดริช โวห์เลอร์ในปี 1828 ถือเป็นสารประกอบอินทรีย์ชนิดแรกที่เตรียมจากสารตั้งต้นอนินทรีย์ทั้งหมด[ 158 ]การสังเคราะห์ยูเรียของโวห์เลอร์แสดงให้เห็นว่าสารประกอบอินทรีย์สามารถสร้างขึ้นจากสารตั้งต้นอนินทรีย์ได้ ซึ่งเป็นการโต้แย้งทฤษฎีพลังชีวิตที่ครอบงำวิทยาศาสตร์ในช่วงต้นศตวรรษที่ 19 การวิเคราะห์สมัยใหม่ถือว่าความสำเร็จนี้เป็นพื้นฐานสำหรับการรวมเคมีอินทรีย์และอนินทรีย์เข้าด้วยกัน[ 159 ]
การค้นพบเอนไซม์ในช่วงต้นศตวรรษที่ 20 โดยEduard Buchnerได้แยกการศึกษาปฏิกิริยาเคมีของการเผาผลาญออกจากการศึกษาทางชีววิทยาของเซลล์ และเป็นจุดเริ่มต้นของชีวเคมี[ 160 ]ความรู้ทางชีวเคมีเติบโตอย่างรวดเร็วตลอดช่วงต้นศตวรรษที่ 20 หนึ่งในนักชีวเคมีสมัยใหม่ที่มีผลงานมากมายคือHans Krebs ซึ่งมีส่วนสำคัญอย่างมากใน การศึกษาการเผาผลาญ[ 161 ]เขาค้นพบวงจรยูเรีย และต่อมาได้ร่วมงานกับHans Kornberg ค้นพบ วงจรกรดซิตริกและวงจรไกลออกซิเลต[ 162 ] [ 163 ] [ 79 ]การวิจัยทางชีวเคมีสมัยใหม่ได้รับความช่วยเหลืออย่างมากจากการพัฒนาเทคนิคใหม่ๆ เช่นโครมาโทกรา ฟี สเปกโทร ส โก ปี NMR กล้องจุลทรรศน์อิเล็กตรอนและ การจำลอง พลศาสตร์โมเลกุลเทคนิคเหล่านี้ทำให้สามารถค้นพบและวิเคราะห์โมเลกุลและวิถีการเผาผลาญต่างๆ ในเซลล์ได้อย่างละเอียด[ 164 ]
ดูเพิ่มเติม
- เมตาบอลิซึมที่เกิดจากกิจกรรมของมนุษย์ – การหมุนเวียนของสารและพลังงานในสังคมมนุษย์
- สารต้านเมตาบอไลต์ – สารเคมีที่ยับยั้งการใช้เมตาบอไลต์
- ทฤษฎีแบตเตอรี่
- แคลอริเมตรี – การหาค่าการถ่ายเทความร้อนในระบบโดยการวัดคุณสมบัติอื่นๆ ของระบบนั้น
- ไมโครแคลอริเมตรีแบบไอโซเทอร์มอล – การวัดอัตราการไหลของความร้อนสุทธิเทียบกับเวลาที่ผ่านไป
- ความผิดปกติแต่กำเนิดของระบบเผาผลาญ – กลุ่มโรคทางพันธุกรรม
- สมมติฐานโลกเหล็ก-กำมะถัน – สถานการณ์สมมติเกี่ยวกับการกำเนิดของชีวิต ทฤษฎี "กระบวนการเผาผลาญมาก่อน" ในการกำเนิดของชีวิต
- ความผิดปกติทางเมตาบอลิซึม – โรคใด ๆ ที่ขัดขวางความสามารถของร่างกายในการประมวลผลและกระจายสารอาหาร
- ไมโครฟิสิโอเมตรี
- กลุ่มโภชนาการหลัก – กลุ่มของสิ่งมีชีวิต
- กระบวนการเผาผลาญขั้นพื้นฐาน – ปฏิกิริยาเคมีที่พัฒนาไปสู่กระบวนการเผาผลาญในปัจจุบัน
- การ วัดอัตราการหายใจ – การประมาณอัตราการเผาผลาญโดยการวัดปริมาณความร้อนที่เกิดขึ้น
- การเผาผลาญกระแสน้ำ
- กระบวนการเผาผลาญกำมะถัน – ชุดปฏิกิริยาเคมีที่เกี่ยวข้องกับกำมะถันในสิ่งมีชีวิต
- ผลกระทบทางความร้อนของอาหาร – การใช้พลังงานในการแปรรูปอาหาร
- เมตาบอลิซึมของเมือง – แบบจำลองการไหลเวียนของวัสดุและพลังงานในเมือง
- การเผาผลาญน้ำ – แง่มุมหนึ่งของภาวะสมดุลที่เกี่ยวข้องกับการควบคุมปริมาณน้ำในร่างกาย
- เมตาบอลิซึมล้น – ปรากฏการณ์ระดับเซลล์
- เมตาบอลิซึมของมะเร็ง
- Reactome – ฐานข้อมูลของวิถีทางชีวภาพ
- KEGG – แหล่งรวมฐานข้อมูลชีวสารสนเทศ
อ่านเพิ่มเติม
เบื้องต้น
- Rose S, Mileusnic R (1999). เคมีของชีวิต . สำนักพิมพ์ Penguin Press Science. ISBN 0-14-027273-9.
- Schneider EC, Sagan D (2005). สู่ความเย็น: การไหลของพลังงาน อุณหพลศาสตร์ และชีวิต . สำนักพิมพ์มหาวิทยาลัยชิคาโก. ISBN 0-226-73936-8.
- Lane N (2004). ออกซิเจน: โมเลกุลที่สร้างโลก . สหรัฐอเมริกา: สำนักพิมพ์มหาวิทยาลัยออกซ์ฟอร์ด. ISBN 0-19-860783-0.
ขั้นสูง
- Price N, Stevens L (1999). พื้นฐานของเอนไซม์วิทยา: ชีววิทยาของเซลล์และโมเลกุลของโปรตีนเร่งปฏิกิริยาสำนักพิมพ์มหาวิทยาลัยออกซ์ฟอร์ด ISBN 0-19-850229-X.
- Berg J, Tymoczko J, Stryer L (2002). ชีวเคมี . WH Freeman and Company. ISBN 0-7167-4955-6.
- Cox M, Nelson DL (2004). หลักการชีวเคมีของเลห์นิงเกอร์ . Palgrave Macmillan. ISBN 0-7167-4339-6.
- Brock TD , Madigan MR, Martinko J, Parker J (2002). ชีววิทยาของจุลินทรีย์ของ Brock . Benjamin Cummings. ISBN 0-13-066271-2.
- Da Silva JJ, Williams RJ (1991). เคมีชีวภาพของธาตุ: เคมีอนินทรีย์ของสิ่งมีชีวิต . สำนักพิมพ์แคลเรนดอน. ISBN 0-19-855598-9.
- Nicholls DG, Ferguson SJ (2002). ชีวพลังงาน . Academic Press Inc. ISBN 0-12-518121-3.
- Wood HG (กุมภาพันธ์ 1991). "ชีวิตด้วย CO หรือ CO2 และ H2 เป็นแหล่งคาร์บอนและพลังงาน" . FASEB Journal . 5 (2): 156– 63. doi : 10.1096/fasebj.5.2.1900793 . PMID 1900793 . S2CID 45967404 .
ลิงก์ภายนอก
ข้อมูลทั่วไป
- ชีวเคมีของการเผาผลาญ (จัดเก็บเมื่อวันที่ 8 มีนาคม 2548)
- Sparknotes SAT ชีวเคมีภาพรวมของชีวเคมี ระดับโรงเรียน
- เอกสารไฮเปอร์เท็กซ์บุ๊คชีววิทยาของ MIT ถูกเก็บถาวรเมื่อวันที่ 19 พฤษภาคม 2016 ที่คลังเก็บข้อมูลเว็บของโปรตุเกส คู่มือระดับปริญญาตรีเกี่ยวกับชีววิทยาระดับโมเลกุล
กระบวนการเผาผลาญของมนุษย์
- หัวข้อในชีวเคมีทางการแพทย์คู่มือเกี่ยวกับกระบวนการเผาผลาญในมนุษย์ ระดับโรงเรียน
- เว็บไซต์ชีวเคมีทางการแพทย์:แหล่งข้อมูลที่ครอบคลุมเกี่ยวกับกระบวนการเผาผลาญในร่างกายมนุษย์
ฐานข้อมูล
- แผนผังแสดงขั้นตอนการเผาผลาญอาหารที่ExPASy
- แผนภูมิเส้นทางการเผาผลาญของ IUBMB-Nicholson
- SuperCYP: ฐานข้อมูลเกี่ยวกับการเผาผลาญยาและไซโตโครมถูกเก็บถาวรเมื่อวันที่ 3 พฤศจิกายน 2011 ที่Wayback Machine
วิถีการเผาผลาญ
- ข้อมูลอ้างอิงเกี่ยวกับกระบวนการเผาผลาญ (Pathway) ถูกเก็บถาวรไว้เมื่อวันที่ 23 กุมภาพันธ์ 2552 ที่Wayback Machine
- วัฏจักรไนโตรเจนและการตรึงไนโตรเจนที่Wayback Machine (ดัชนีเก็บถาวร)
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การเผาผลาญ
เมตาบอลิซึม ( / m ə ˈ t æ b ə l ɪ z ə m / , จาก ภาษากรีก : μεταβολή metabolē , "การเปลี่ยนแปลง") หมายถึงชุดของ ปฏิกิริยาเคมี ที่จำเป็นต่อการดำรง ชีวิต ซึ่งเกิดขึ้นภายใน...
สารชีวเคมีที่สำคัญ
โครงสร้างส่วนใหญ่ที่ประกอบขึ้นเป็นสัตว์ พืช และจุลินทรีย์นั้นสร้างขึ้นจาก โมเลกุล พื้นฐานสี่ประเภท ได้แก่ กรด อะ มิโน คาร์โบไฮเดรต กรด นิวคลีอิก และ ลิปิด (มักเรียกว่า ไขมัน ) เนื่องจากโมเลกุลเหล่านี้มีความสำคัญต่อชีวิต...
กรดอะมิโนและโปรตีน
โปรตีนประกอบด้วย กรดอะมิโน ที่เรียงตัวเป็นสายตรงเชื่อมต่อกันด้วย พันธะเปปไทด์ โปรตีนหลายชนิดเป็น เอนไซม์ ที่ เร่ง ปฏิกิริยาเคมีในกระบวนการเผาผลาญ โปรตีนอื่นๆ มีหน้าที่เชิงโครงสร้างหรือเชิงกล เช่น โปรตีนที่สร้าง โครงร่างเซลล์...
ลิปิด
ลิปิดเป็นกลุ่มสารชีวเคมีที่มีความหลากหลายมากที่สุด การใช้งานเชิงโครงสร้างหลักของลิปิดคือการเป็นส่วนหนึ่งของ เยื่อชีวภาพ ภายในและภายนอก เช่นเยื่อ หุ้มเซลล์ [ 11 ] พลังงานเคมี ของ ลิปิดยังสามารถนำมาใช้ได้อีกด้วย ลิปิดประกอบด้วย โซ่ไฮโดรคาร์บอน...