อ่าน 43 นาที
แคทเทนินเบตา-1
แคทเทนินเบตา-1 หรือที่รู้จักกันในชื่อ β-แคทเทนิน ( beta - catenin) เป็น โปรตีน ที่ในมนุษย์ถูกสร้างขึ้นโดย ยีน CTNNB1
แคทเทนินเบตา-1
| ซีทีเอ็นบี1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | CTNNB1 , CTNNB, MRD19, อาร์มาดิลโล, แคทเทนินเบตา 1, EVR7, NEDSDV | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | โอมิม : 116806 ; เอ็มจีไอ : 88276 ; โฮโมโลยีน : 1434 ; GeneCards : CTNNB1 ; OMA : CTNNB1 - ออโธโลจี | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




แคทเทนินเบตา-1หรือที่รู้จักกันในชื่อβ-แคทเทนิน ( beta - catenin) เป็นโปรตีนที่ในมนุษย์ถูกสร้างขึ้นโดยยีนCTNNB1
β-catenin เป็นโปรตีน ที่มีหน้าที่สองอย่าง เกี่ยวข้องกับการควบคุมและการประสานงานของการยึดเกาะระหว่างเซลล์และการถอดรหัสยีนในมนุษย์ โปรตีน CTNNB1 ถูกเข้ารหัสโดยยีนCTNNB1 [ 5 ] [ 6 ] ในแมลงหวี่โปรตีนที่คล้ายคลึงกันเรียกว่าarmadillo β-catenin เป็นหน่วยย่อยของ โปรตีนคอมเพล็กซ์ cadherinและทำหน้าที่เป็นตัวส่งสัญญาณภายในเซลล์ในเส้นทางการส่งสัญญาณ Wnt [ 7 ] [ 8 ] [ 9 ] มันเป็นสมาชิกของ ตระกูลโปรตีน cateninและคล้ายคลึงกับγ-cateninหรือที่รู้จักกันในชื่อplakoglobin β-catenin มีการแสดงออกอย่างกว้างขวางในเนื้อเยื่อหลายชนิด ใน กล้ามเนื้อหัวใจ β-catenin จะอยู่บริเวณจุดเชื่อมต่อ adherensใน โครงสร้าง แผ่นดิสก์แทรกซึ่งมีความสำคัญต่อการเชื่อมต่อทางไฟฟ้าและทางกลระหว่างเซลล์กล้ามเนื้อหัวใจ ที่อยู่ติด กัน
การกลายพันธุ์ และการแสดงออกมากเกินไปของ β-catenin เกี่ยวข้องกับมะเร็งหลายชนิด รวมถึงมะเร็งตับ มะเร็งลำไส้ใหญ่มะเร็งปอดมะเร็งเต้านมมะเร็งรังไข่และมะเร็งเยื่อบุโพรงมดลูก [ 10 ] การเปลี่ยนแปลงตำแหน่งและการแสดงออกของ β-catenin เกี่ยวข้องกับโรคหัวใจ หลายรูปแบบ รวมถึงโรคกล้ามเนื้อหัวใจห้องล่างขยายตัว β-catenin ถูกควบคุมและทำลายโดยคอมเพล็กซ์ทำลาย β-cateninโดยเฉพาะอย่างยิ่ง โปรตีน adenomatous polyposis coli (APC) ซึ่งถูกเข้ารหัสโดย ยีนAPCที่ยับยั้งเนื้องอกดังนั้นการกลายพันธุ์ทางพันธุกรรมของ ยีน APCจึงมีความเชื่อมโยงอย่างมากกับมะเร็ง โดยเฉพาะอย่างยิ่งมะเร็งลำไส้ใหญ่ที่เกิดจากโรคติ่งเนื้อในลำไส้ใหญ่แบบครอบครัว (FAP)
การค้นพบ
β-catenin ถูกค้นพบครั้งแรกในช่วงต้นทศวรรษ 1990 ในฐานะส่วนประกอบของ คอมเพล็กซ์ การยึดเกาะเซลล์ ของสัตว์เลี้ยงลูกด้วยนม : โปรตีนที่รับผิดชอบในการยึดเกาะไซโตพลาสซึมของ แค ดเฮริน[ 11 ]แต่ในไม่ช้าก็พบว่าโปรตีนอาร์ มาดิลโลของแมลงหวี่ ซึ่งเกี่ยวข้องกับการเป็นตัวกลางของผลกระทบทางสัณฐานวิทยาของWingless/Wntนั้น มีความคล้ายคลึงกับ β-catenin ของสัตว์เลี้ยงลูกด้วยนม ไม่เพียงแต่ในโครงสร้างเท่านั้น แต่ยังรวมถึงหน้าที่ด้วย[ 12 ]ดังนั้น β-catenin จึงกลายเป็นหนึ่งในตัวอย่างแรกๆ ของmoonlighting : โปรตีนที่ทำหน้าที่ในเซลล์มากกว่าหนึ่งหน้าที่ที่แตกต่างกันอย่างสิ้นเชิง
โครงสร้าง
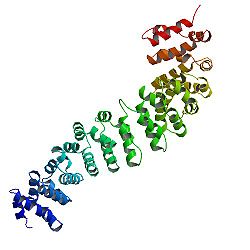
โครงสร้างโปรตีน

แกนกลางของ β-catenin ประกอบด้วยส่วนที่ซ้ำกัน ที่มีลักษณะเฉพาะหลาย ส่วน แต่ละส่วนมีความยาวประมาณ 40 กรดอะมิโน เรียกว่า armadillo repeats องค์ประกอบทั้งหมดเหล่านี้จะพับรวมกันเป็น โดเมนโปรตีนที่แข็งแรงเพียงโดเมนเดียวที่มีรูปร่างยาว เรียกว่า armadillo domain (ARM) armadillo repeat โดยเฉลี่ยประกอบด้วยอัลฟาเฮลิกซ์ 3 อัน ส่วนที่ซ้ำกันแรกของ β-catenin (ใกล้กับปลาย N-terminus) จะแตกต่างจากส่วนอื่นๆ เล็กน้อย เนื่องจากมีเฮลิกซ์ที่ยาวและมีรอยหักงอ ซึ่งเกิดจากการรวมกันของเฮลิกซ์ 1 และ 2 [ 13 ]เนื่องจากรูปร่างที่ซับซ้อนของส่วนที่ซ้ำกันแต่ละส่วน ARM domain ทั้งหมดจึงไม่ใช่แท่งตรง แต่มีความโค้งเล็กน้อย ทำให้เกิดพื้นผิวด้านนอก (นูน) และด้านใน (เว้า) พื้นผิวด้านในนี้ทำหน้าที่เป็นตำแหน่งการจับลิแกนด์สำหรับพันธมิตรปฏิสัมพันธ์ต่างๆ ของ ARM domain
ส่วนปลาย N และส่วนปลาย C ที่อยู่ไกลจากโดเมน ARM ไม่ได้มีโครงสร้างใดๆ ในสารละลายด้วยตัวมันเอง อย่างไรก็ตาม บริเวณ ที่ไม่มีระเบียบโดยเนื้อแท้ เหล่านี้ มีบทบาทสำคัญในการทำงานของ β-catenin บริเวณที่ไม่มีระเบียบที่ปลาย N ประกอบด้วยโมทีฟเชิงเส้นสั้นๆ ที่ได้รับการอนุรักษ์ไว้ ซึ่งรับผิดชอบในการจับกับTrCP1 (หรือที่รู้จักกันในชื่อ β-TrCP) E3 ubiquitin ligase – แต่เฉพาะเมื่อมันถูกฟอสฟอริเลตเท่านั้น ดังนั้น การสลายตัวของ β-catenin จึงเกิดขึ้นโดยส่วนปลาย N นี้ ในทางกลับกัน บริเวณปลาย C เป็นตัวกระตุ้นการถอดรหัส ที่แข็งแกร่ง เมื่อถูกดึงดูดไปยังDNAส่วนนี้ไม่ได้ไม่มีระเบียบอย่างสมบูรณ์: ส่วนหนึ่งของส่วนขยายปลาย C สร้างเกลียว ที่เสถียร ซึ่งอัดแน่นกับโดเมน ARM แต่ก็อาจมีส่วนร่วมกับพันธมิตรการจับที่แยกต่างหากได้เช่นกัน[ 14 ]องค์ประกอบโครงสร้างขนาดเล็กนี้ (HelixC) ปิดปลายปลาย C ของโดเมน ARM ป้องกันสารตกค้างที่ไม่ชอบน้ำ HelixC ไม่จำเป็นสำหรับการทำงานของ β-catenin ในการยึดเกาะระหว่างเซลล์ ในทางกลับกัน มันจำเป็นสำหรับการส่งสัญญาณ Wnt: อาจเพื่อดึงดูดโคแอคติเวเตอร์ต่างๆ เช่น 14-3-3zeta [ 15 ]อย่างไรก็ตาม คู่หูที่แน่นอนของมันในกลุ่มคอมเพล็กซ์การถอดรหัสทั่วไปยังคงไม่เป็นที่เข้าใจอย่างสมบูรณ์ และอาจเกี่ยวข้องกับผู้เล่นเฉพาะเนื้อเยื่อ[ 16 ]ที่น่าสังเกตคือ ส่วนปลาย C ของ β-catenin สามารถเลียนแบบผลกระทบของเส้นทาง Wnt ทั้งหมดได้ หากถูกเชื่อมเข้ากับโดเมนการจับ DNA ของปัจจัยการถอดรหัสLEF1 อย่างประดิษฐ์ [ 17 ]
พลาโคโกลบิน (เรียกอีกอย่างว่า γ-catenin) มีโครงสร้างที่คล้ายคลึงกับ β-catenin อย่างมาก ไม่เพียงแต่โดเมน ARM ของพวกมันจะคล้ายคลึงกันทั้งในด้านโครงสร้างและความสามารถในการจับกับลิแกนด์เท่านั้น แต่โมทีฟการจับ β-TrCP ที่ปลาย N-terminal ก็ยังได้รับการอนุรักษ์ไว้ในพลาโคโกลบินด้วย ซึ่งบ่งชี้ถึงบรรพบุรุษร่วมกันและการควบคุมร่วมกันกับ β-catenin [ 18 ]อย่างไรก็ตาม พลาโคโกลบินเป็นตัวกระตุ้นการถอดรหัสที่อ่อนแอมากเมื่อจับกับ DNA ซึ่งอาจเกิดจากความแตกต่างของลำดับปลาย C-terminal (พลาโคโกลบินดูเหมือนจะขาดโมทีฟตัวกระตุ้นการถอดรหัส ดังนั้นจึงยับยั้ง ยีนเป้าหมายของ วิถี Wntแทนที่จะกระตุ้น) [ 19 ]
การจับกับโดเมนอาร์มาดิลโล

ดังที่ได้กล่าวไว้ข้างต้นโดเมน ARMของ β-catenin ทำหน้าที่เป็นแพลตฟอร์มที่โมทีฟเชิงเส้น เฉพาะ สามารถจับได้ โมทีฟที่จับกับ β-catenin ซึ่งอยู่ในพันธมิตรที่มีโครงสร้างหลากหลาย มักจะไม่มีระเบียบในตัวเอง และโดยทั่วไปจะรับเอาโครงสร้างที่แข็งตัวเมื่อจับกับโดเมน ARM – ดังที่เห็นได้จากโมทีฟเชิงเส้นสั้นๆอย่างไรก็ตาม โมทีฟที่โต้ตอบกับ β-catenin ยังมีลักษณะเฉพาะหลายประการ ประการแรก พวกมันอาจมีความยาวถึงหรือเกิน 30 กรดอะมิโนและสัมผัสกับโดเมน ARM บนพื้นที่ผิวที่ใหญ่เกินไป คุณลักษณะที่ผิดปกติอีกประการหนึ่งของโมทีฟเหล่านี้คือระดับการฟอสโฟรีเลชัน ที่สูงบ่อยครั้ง เหตุการณ์การฟอสโฟรีเลชัน Ser / Thrดังกล่าวช่วยเพิ่มการจับของโมทีฟที่เกี่ยวข้องกับ β-catenin จำนวนมากกับโดเมน ARM อย่างมาก[ 20 ]
โครงสร้างของ β-catenin ที่ซับซ้อนกับโดเมนการจับ catenin ของ TCF ซึ่งเป็นพันธมิตรในการกระตุ้นการถอดรหัส ได้ให้แผนที่โครงสร้างเบื้องต้นว่าพันธมิตรการจับของ β-catenin สามารถสร้างปฏิสัมพันธ์ได้อย่างไร[ 21 ] โครงสร้างนี้แสดงให้เห็นว่าปลาย N ของ TCF ที่ไม่มีระเบียบนั้นปรับตัวให้เข้ากับสิ่งที่ดูเหมือนจะเป็นโครงสร้างที่แข็งแรง โดยมีลวดลายการจับที่ครอบคลุมการทำซ้ำของ β-catenin หลายครั้ง จุด "ร้อน" ของปฏิสัมพันธ์ที่มีประจุค่อนข้างแรงได้รับการกำหนด (ทำนายและได้รับการยืนยันในภายหลังว่าได้รับการอนุรักษ์ไว้สำหรับปฏิสัมพันธ์ β-catenin/E-cadherin) เช่นเดียวกับบริเวณที่ไม่ชอบน้ำซึ่งถือว่ามีความสำคัญในรูปแบบการจับโดยรวมและเป็นเป้าหมายของสารยับยั้งโมเลกุลขนาดเล็กที่มีศักยภาพในการรักษาโรคมะเร็งบางชนิด นอกจากนี้ การศึกษาต่อมายังแสดงให้เห็นถึงลักษณะเฉพาะอีกประการหนึ่ง คือ ความยืดหยุ่นในการจับของปลาย N ของ TCF กับ β-catenin [ 22 ] [ 23 ]โดเมน ARM ยังดึงดูดปัจจัยควบคุม เช่น คอมเพล็กซ์การปรับโครงสร้างโครมาตินที่ขึ้นอยู่กับ ATP ของ SWI/SNFซึ่งจับผ่านบริเวณที่ไม่เป็นระเบียบที่พบในหน่วยย่อยARID1A [ 24 ]
ในทำนองเดียวกัน เราพบE-cadherin ที่คุ้นเคย ซึ่งส่วนหางไซโตพลาสมิกสัมผัสกับโดเมน ARM ในลักษณะมาตรฐานเดียวกัน[ 25 ]โปรตีนโครงสร้าง axin (พาราโลกที่เกี่ยวข้องอย่างใกล้ชิดสองตัว คือaxin 1และaxin 2 ) มีโมทีฟการโต้ตอบที่คล้ายกันบนส่วนกลางที่ยาวและไม่เป็นระเบียบ[ 26 ]แม้ว่าโมเลกุลของ axin จะมีโมทีฟการดึงดูด β-catenin เพียงโมทีฟเดียว แต่ โปรตีน adenomatous polyposis coli (APC) ซึ่งเป็นคู่หูของมันมีโมทีฟดังกล่าว 11 โมทีฟเรียงต่อกันในแต่ละโปรโตเมอร์ จึงสามารถโต้ตอบกับโมเลกุล β-catenin หลายโมเลกุลพร้อมกันได้[ 27 ] เนื่องจากพื้นผิวของโดเมน ARM สามารถรองรับโมทีฟเปปไทด์ได้เพียงโมทีฟเดียวในแต่ละครั้ง โปรตีนเหล่านี้จึงแข่งขันกันเพื่อแย่งชิงโมเลกุล β-catenin ในเซลล์เดียวกัน การแข่งขันนี้เป็นกุญแจสำคัญในการทำความเข้าใจว่าเส้นทางการส่งสัญญาณ Wntทำงาน อย่างไร
อย่างไรก็ตาม ตำแหน่งการจับ "หลัก" บนโดเมน ARM ของ β-catenin นั้นไม่ใช่ตำแหน่งเดียว เฮลิกซ์แรกของโดเมน ARM สร้างช่องปฏิสัมพันธ์ระหว่างโปรตีนแบบพิเศษเพิ่มเติม: ซึ่งสามารถรองรับโมทีฟเชิงเส้นที่สร้างเฮลิกซ์ซึ่งพบในโคแอคติเวเตอร์BCL9 (หรือ BCL9Lที่เกี่ยวข้องอย่างใกล้ชิด) ซึ่งเป็นโปรตีนสำคัญที่เกี่ยวข้องกับการส่งสัญญาณ Wnt [ 28 ]แม้ว่ารายละเอียดที่แม่นยำจะไม่ชัดเจนนัก แต่ดูเหมือนว่า alpha-catenin จะใช้ตำแหน่งเดียวกันเมื่อ β-catenin อยู่ในตำแหน่งที่ adherens junctions [ 29 ]เนื่องจากช่องนี้แตกต่างจากตำแหน่งการจับ "หลัก" ของโดเมน ARM จึงไม่มีการแข่งขันระหว่าง alpha-catenin และ E-cadherin หรือระหว่าง TCF1 และ BCL9 ตามลำดับ[ 30 ]ในทางกลับกัน BCL9 และ BCL9L ต้องแข่งขันกับ α-catenin เพื่อเข้าถึงโมเลกุล β-catenin [ 31 ]
การทำงาน
บทบาทในฐานะปัจจัยการถอดรหัส
β-catenin สามารถเข้าสู่นิวเคลียสและทำหน้าที่เป็นปัจจัยการถอดรหัสได้ แม้ว่ากลไกที่แน่นอนสำหรับการเคลื่อนย้ายเข้าสู่นิวเคลียสนี้ยังอยู่ระหว่างการตรวจสอบ[ 32 ]เมื่ออยู่ในนิวเคลียสแล้ว β-catenin จะมีปฏิสัมพันธ์กับT-cell factor/Lymphoid enhancer factor (TCF/LEF) โดยเปลี่ยน TCF/LEF จากตัวยับยั้งเป็นตัวส่งเสริมการถอดรหัส นอกจากนี้ยังดึงดูดโปรตีนMediatorซึ่งจากนั้นจะดึงดูดRNA polymerase IIและปัจจัยการถอดรหัสทั่วไปอื่นๆ[ 33 ]เส้นทาง Wnt/β-catenin จะกระตุ้นการถอดรหัสของยีนหลายตัว โดยเฉพาะอย่างยิ่งยีนที่เข้ารหัสผลิตภัณฑ์ต่อไปนี้:
- ตัวขนส่งยาหลายชนิด ABC (ABCB1)
- Survivin (BIRC5)
- ไซคลิน ดี1 (CCND1)
- ปัจจัยการเจริญเติบโตของไฟโบรบลาสต์ 18 (FGF18)
- เมทริกซ์เมทัลโลโปรตีเนส-14 (MMP14)
- โปรโตออนโคยีน Myc (MYC)
ยีนจำนวนมากที่ถูกกระตุ้นโดยเส้นทาง β-catenin มีส่วนเกี่ยวข้องโดยตรงหรือโดยอ้อมกับการเจริญเติบโต การเพิ่มจำนวน และการอยู่รอดของเซลล์ ยีนเหล่านี้มีความสำคัญในกระบวนการเกิดตัวอ่อนและการเกิดเนื้องอก ทำให้บทบาทของ β-catenin ในฐานะปัจจัยการถอดรหัสเป็นที่น่าสนใจเป็นพิเศษสำหรับนักวิจัย[ 34 ]
การควบคุมการสลายตัวผ่านกระบวนการฟอสโฟรีเลชัน
ระดับเซลล์ของ β-catenin ส่วนใหญ่ถูกควบคุมโดยกระบวนการยูบิควิตินเนชันและการย่อยสลายโดยโปรตีโอโซมเอนไซม์ยูบิควิตินไลเกส E3 TrCP1 (หรือที่รู้จักกันในชื่อ β-TrCP) สามารถจดจำ β-catenin เป็นสารตั้งต้นได้ผ่านทางลำดับกรดอะมิโนสั้นๆ บนปลาย N-terminus ที่ไม่มีโครงสร้างที่แน่นอน อย่างไรก็ตาม ลำดับกรดอะมิโนนี้ (Asp-Ser-Gly-Ile-His-Ser) ของ β-catenin จำเป็นต้องได้รับการฟอสฟอริเลชันที่ซีรีนทั้งสองตำแหน่งเพื่อให้สามารถจับกับ β-TrCP ได้ การฟอสฟอริเลชันของลำดับกรดอะมิโนนี้กระทำโดยไกลโคเจนซินเทสไคเนส 3อัลฟาและเบตา (GSK3α และ GSK3β) GSK3 เป็นเอนไซม์ที่ทำงานอย่างต่อเนื่องและเกี่ยวข้องกับกระบวนการควบคุมที่สำคัญหลายอย่าง อย่างไรก็ตาม มีข้อกำหนดหนึ่งข้อคือ สารตั้งต้นของ GSK3 จำเป็นต้องได้รับการฟอสฟอริเลชันล่วงหน้าแล้วที่กรดอะมิโนสี่ตัวถัดไป (ทางด้าน C-terminus) จากตำแหน่งเป้าหมายที่แท้จริง ดังนั้นจึงจำเป็นต้องมี "ไคเนสเริ่มต้น" สำหรับการทำงานของมันด้วย ในกรณีของ β-catenin ไคเนสเริ่มต้นที่สำคัญที่สุดคือcasein kinase 1 (CKI) เมื่อสารตั้งต้นที่มีซีรีนและทรีโอนีนเป็นองค์ประกอบหลักได้รับการ "เตรียมพร้อม" แล้ว GSK3 สามารถ "เคลื่อนที่" ข้ามสารตั้งต้นนั้นจากปลาย C ไปยังปลาย N โดยทำการฟอสฟอริเลชันทุกๆ ซีรีนหรือทรีโอนีน ตัวที่ 4 ในแถวเดียวกัน กระบวนการนี้จะส่งผลให้เกิดการฟอสฟอริเลชันคู่ของโมทีฟการจดจำ β-TrCP ที่กล่าวถึงข้างต้นด้วยเช่นกัน
คอมเพล็กซ์การทำลายเบตา-แคทเทนิน

เพื่อให้GSK3เป็นไคเนส ที่มีประสิทธิภาพสูง บนซับสเตรต การฟอสโฟรีเลชันล่วงหน้าเพียงอย่างเดียวไม่เพียงพอ มีข้อกำหนดเพิ่มเติมอีกประการหนึ่ง: เช่นเดียวกับไมโทเจนแอคติเวตโปรตีนไคเนส (MAPK) ซับสเตรตจำเป็นต้องเชื่อมโยงกับเอนไซม์นี้ผ่านโมทีฟการเชื่อมต่อที่มีความสัมพันธ์สูงβ -catenin ไม่มีโมทีฟดังกล่าว แต่มีโปรตีนพิเศษตัวหนึ่งที่มี นั่นคือaxinยิ่งไปกว่านั้น โมทีฟการเชื่อมต่อ GSK3 ของมันอยู่ติดกับโมทีฟการจับ β-catenin โดยตรง[ 26 ]ด้วยวิธีนี้axin ทำหน้าที่เป็น โปรตีนโครงสร้างที่แท้จริงนำเอนไซม์ (GSK3) มารวมกับซับสเตรต (β-catenin) ให้อยู่ใกล้กัน
แต่ถึงกระนั้นaxinก็ไม่ได้ทำงานเพียงลำพัง ผ่าน โดเมน ควบคุมการส่งสัญญาณ G-protein (RGS) ที่ปลาย N-terminus มันจะดึงดูด โปรตีน adenomatous polyposis coli (APC) APCเปรียบเสมือน "ต้นคริสต์มาส" ขนาดใหญ่: ด้วยโมทีฟการจับ β-catenin จำนวนมาก ( โมเลกุล APCเพียงโมเลกุลเดียวมีโมทีฟดังกล่าวถึง 11 โมทีฟ[ 27 ] ) มันสามารถรวบรวมโมเลกุล β-catenin ได้มากที่สุดเท่าที่จะเป็นไปได้[ 35 ] APCสามารถโต้ตอบกับ โมเลกุล axin หลาย โมเลกุลพร้อมกันได้ เนื่องจากมีโมทีฟ SAMP (Ser-Ala-Met-Pro) สามโมทีฟเพื่อจับกับโดเมน RGSที่พบในaxinนอกจากนี้ axin ยังมีศักยภาพในการรวมตัวกันเป็นโอลิโกเมอร์ผ่านโดเมน DIX ที่ปลาย C-terminus ผลลัพธ์ที่ได้คือกลุ่มโปรตีนหลายโมเลกุลขนาดใหญ่ที่อุทิศให้กับการฟอสโฟรีเลชันของ β-catenin โดยทั่วไปคอมเพล็กซ์นี้เรียกว่าคอมเพล็กซ์การทำลายเบต้า-แคทเทนินแม้ว่าจะแตกต่างจาก กลไก โปรตีโอโซมที่รับผิดชอบในการย่อยสลายเบต้า-แคทเทนินก็ตาม[ 36 ]มันเพียงแค่ทำเครื่องหมายโมเลกุลเบต้า-แคทเทนินเพื่อการทำลายในภายหลัง
การส่งสัญญาณ Wnt และการควบคุมการทำลาย
ในเซลล์ที่อยู่ในสภาวะพัก โมเลกุล ของแอ็กซินจะรวมตัวกันเป็นโอลิโกเมอร์ผ่านโดเมน DIX ที่ปลาย C-terminus ซึ่งมีส่วนเชื่อมต่อสองส่วน ดังนั้นจึงสามารถสร้างโอลิโกเมอร์เชิงเส้นหรือแม้แต่พอลิเมอร์ภายในไซโตพลาสซึมของเซลล์ได้ โดเมน DIX นั้นมีเอกลักษณ์เฉพาะตัว โปรตีนอื่น ๆ ที่ทราบว่ามีโดเมน DIX มีเพียงDishevelledและDIXDC1 เท่านั้น (โปรตีน Dshตัวเดียวของแมลงหวี่สอดคล้องกับยีนพาราโลกัสสามยีน ได้แก่Dvl1 , Dvl2และDvl3ในสัตว์เลี้ยงลูกด้วยนม ) Dshจะเชื่อมต่อกับบริเวณไซโตพลาสซึมของ ตัวรับ Frizzledด้วยโดเมนPDZและDEP เมื่อ โมเลกุล WntจับกับFrizzledมันจะกระตุ้นลำดับเหตุการณ์ที่ไม่ค่อยเป็นที่รู้จัก ซึ่งส่งผลให้โดเมน DIX ของ dishevelled ถูกเปิดเผยและสร้างตำแหน่งการจับที่สมบูรณ์แบบสำหรับแอ็กซิน จากนั้น Axin จะถูกไทเทรตออกจากโครงสร้างโอลิโกเมอริกของมัน – คอมเพล็กซ์ทำลาย β-catenin – โดยDsh [ 37 ]เมื่อจับกับคอมเพล็กซ์ตัวรับแล้วAxinจะไม่สามารถจับกับ β-catenin และทำงานของ GSK3 ได้ ที่สำคัญคือ ส่วนของไซโตพลาสมิกของโปรตีน LRP5 และ LRP6 ที่เกี่ยวข้องกับ Frizzled มีลำดับของสารตั้งต้นเทียมของ GSK3 (Pro-Pro-Pro-Ser-Pro-x-Ser) ซึ่ง "เตรียมพร้อม" (ฟอสโฟรีเลตล่วงหน้า) โดยCKI อย่างเหมาะสม ราวกับว่าเป็นสารตั้งต้นที่แท้จริงของ GSK3 ตำแหน่งเป้าหมายเทียมเหล่านี้ยับยั้งการทำงานของ GSK3 อย่างมากในลักษณะการแข่งขัน[ 38 ] ด้วยวิธีนี้Axin ที่จับกับตัวรับ จะยกเลิกการเป็นตัวกลางในการฟอสโฟรีเลตของ β-catenin เนื่องจาก β-catenin ไม่ได้ถูกทำเครื่องหมายสำหรับการทำลายอีกต่อไป แต่ยังคงถูกผลิตต่อไป ความเข้มข้นของมันจึงจะเพิ่มขึ้น เมื่อระดับของ β-catenin สูงขึ้นจนอิ่มตัวในตำแหน่งการจับทั้งหมดในไซโตพลาสซึม มันก็จะเคลื่อนย้ายเข้าไปในนิวเคลียสด้วย เมื่อจับกับปัจจัยการถอดรหัสLEF1 , TCF1 , TCF2หรือTCF3แล้ว β-catenin จะบังคับให้ปัจจัยเหล่านั้นแยกตัวออกจากคู่หูเดิมคือโปรตีน Groucho ต่างจากGrouchoที่ดึงดูดตัวยับยั้งการถอดรหัส (เช่นฮิสโตน-ไลซีนเมทิลทรานสเฟอ เรส ) β-catenin จะจับ กับ ตัวกระตุ้นการถอดรหัสทำให้ยีนเป้าหมายทำงาน
บทบาทในการยึดเกาะระหว่างเซลล์

คอมเพล็กซ์ การยึดเกาะระหว่างเซลล์มีความสำคัญต่อการสร้างเนื้อเยื่อสัตว์ที่ซับซ้อน β-catenin เป็นส่วนหนึ่งของคอมเพล็กซ์โปรตีนที่สร้างจุดเชื่อมต่อแบบแอด เฮเรน ส์[ 39 ]คอมเพล็กซ์การยึดเกาะระหว่างเซลล์เหล่านี้จำเป็นต่อการสร้างและการบำรุงรักษา ชั้น เซลล์เยื่อบุผิวและสิ่งกีดขวาง ในฐานะที่เป็นส่วนประกอบของคอมเพล็กซ์ β-catenin สามารถควบคุมการเจริญเติบโตของเซลล์และการยึดเกาะระหว่างเซลล์ได้ นอกจากนี้ยังอาจรับผิดชอบในการส่งสัญญาณยับยั้งการสัมผัสที่ทำให้เซลล์หยุดการแบ่งตัวเมื่อแผ่นเยื่อบุผิวสมบูรณ์[ 40 ]คอมเพล็กซ์ E-cadherin – β-catenin – α-catenin เชื่อมโยงกับเส้นใยแอคติน อย่างอ่อน จุดเชื่อมต่อแบบแอดเฮเรนส์ต้องการ พลวัตของโปรตีนที่สำคัญเพื่อเชื่อมโยงกับโครงสร้างไซโตสเกเลตันของแอคติน[ 39 ] จึงทำให้เกิด การ ส่งสัญญาณเชิงกล [ 41 ] [ 42 ]
โปรตีนแคดเฮรินเป็นส่วนประกอบสำคัญของแอดเฮเรนส์จังก์ชัน แคดเฮรินสร้างโครงสร้างการเชื่อมต่อระหว่างเซลล์ที่เรียกว่าแอดเฮเรนส์จังก์ชันและเดสโมโซมแคดเฮรินสามารถมีปฏิสัมพันธ์แบบโฮโมฟิลิกผ่านโดเมนแคดเฮรินซ้ำภายนอกเซลล์ในลักษณะที่ขึ้นอยู่กับ Ca2+ ซึ่งสามารถยึดเซลล์เยื่อบุผิวที่อยู่ติดกันไว้ด้วยกัน ในขณะที่อยู่ในแอดเฮเรนส์จังก์ชัน แคดเฮรินจะดึงโมเลกุล β-catenin เข้ามาในบริเวณภายในเซลล์ β-catenin จะเชื่อมโยงกับโปรตีนที่มีพลวัตสูงอีกตัวหนึ่งคือα -cateninซึ่งจับกับเส้นใยแอคตินโดยตรง[ 43 ]สิ่งนี้เป็นไปได้เพราะ α-catenin และแคดเฮรินจับกับ β-catenin ที่ตำแหน่งที่แตกต่างกัน[ 44 ]ดังนั้นคอมเพล็กซ์ β-catenin – α-catenin จึงสามารถสร้างสะพานเชื่อมระหว่างแคดเฮรินและไซโตสเกเลตันของ แอคตินได้ [ 45 ]การจัดระเบียบของคอมเพล็กซ์แคดเฮริน-แคทเทนินยังได้รับการควบคุมเพิ่มเติมผ่านการฟอสโฟรีเลชันและเอนโดไซโทซิสของส่วนประกอบต่างๆ
บทบาทในการพัฒนา
β-Catenin มีบทบาทสำคัญในการชี้นำกระบวนการพัฒนาหลายอย่าง เนื่องจากสามารถจับกับปัจจัยการถอดรหัสและตัวควบคุมโครมาติน ได้โดยตรง [ 24 ]และสามารถควบคุมได้โดยสารนอกเซลล์ที่แพร่กระจายได้: Wnt โดยจะออกฤทธิ์ต่อตัวอ่อนในระยะแรกเพื่อเหนี่ยวนำให้เกิดการเปลี่ยนแปลงทั่วร่างกาย รวมถึงเซลล์แต่ละเซลล์ในระยะพัฒนาการที่ตามมา นอกจากนี้ยังควบคุมกระบวนการสร้างใหม่ทางสรีรวิทยาอีกด้วย
รูปแบบการพัฒนาตัวอ่อนในระยะเริ่มต้น
การส่งสัญญาณ Wnt และการแสดงออกของยีนที่ขึ้นอยู่กับ β-catenin มีบทบาทสำคัญในระหว่างการก่อตัวของส่วนต่างๆ ของร่างกายในตัวอ่อนระยะแรก ตัวอ่อนที่ได้รับการดัดแปลงทางทดลองที่ไม่แสดงโปรตีนนี้จะไม่สามารถพัฒนาเมโซเดอร์มและเริ่มต้น กระบวนการ แกสตรูเลชันได้[ 46 ] การกำหนดเอนโดเมโซเดอร์มของตัวอ่อนระยะแรกยังเกี่ยวข้องกับการกระตุ้นกิจกรรมการถอดรหัสที่ขึ้นอยู่กับ β-catenin โดยการเคลื่อนไหวทางสัณฐานวิทยาครั้งแรกของการเกิดตัวอ่อน ผ่านกระบวนการถ่ายทอดทางกล คุณลักษณะนี้พบได้ในสัตว์มีกระดูกสันหลังและสัตว์ขาปล้องที่มีสมมาตรสองด้าน และในสัตว์กลุ่มไนดาเรีย จึงมีการเสนอว่าคุณลักษณะนี้ได้รับการสืบทอดทางวิวัฒนาการมาจากการมีส่วนร่วมที่เป็นไปได้ในการกำหนดเอนโดเมโซเดอร์มของเมตาโซอาแรก[ 47 ] [ 48 ] [ 49 ] [ 50 ]
ในระหว่างระยะบลาสตูลาและแกสตรูลา เส้นทาง WntรวมถึงBMPและFGFจะกระตุ้นการสร้างแกนหน้า-หลัง ควบคุมตำแหน่งที่แม่นยำของร่องดั้งเดิม (การสร้างแกสตรูเลชันและเมโซเดอร์ม) รวมถึงกระบวนการสร้างระบบประสาท (การพัฒนาระบบประสาทส่วนกลาง) [ 51 ]
ใน เซลล์ไข่ ของ Xenopus β-catenin จะกระจายตัวอย่างเท่าเทียมกันในทุกส่วนของไข่ในตอนแรก แต่จะถูกกำหนดเป้าหมายสำหรับการยูบิควิตินเนชันและการย่อยสลายโดยคอมเพล็กซ์ทำลาย β-catenin การปฏิสนธิของไข่ทำให้เกิดการหมุนของชั้นคอร์เทกซ์ด้านนอก ทำให้กลุ่มของ โปรตีน FrizzledและDsh เคลื่อน เข้าใกล้บริเวณเส้นศูนย์สูตรมากขึ้น β-catenin จะถูกสะสมในบริเวณนั้นภายใต้อิทธิพลของเส้นทางการส่งสัญญาณ Wnt ในเซลล์ที่ได้รับส่วนของไซโตพลาซึมนี้ ในที่สุดมันจะเคลื่อนย้ายไปยังนิวเคลียสเพื่อจับกับTCF3เพื่อกระตุ้นยีนหลายตัวที่เหนี่ยวนำลักษณะเซลล์ด้านหลัง[ 52 ]การส่งสัญญาณนี้ส่งผลให้เกิดบริเวณของเซลล์ที่เรียกว่าเสี้ยวพระจันทร์สีเทา ซึ่งเป็นตัวจัดระเบียบการพัฒนาของตัวอ่อนแบบคลาสสิก หากบริเวณนี้ถูกผ่าตัดออกจากตัวอ่อน การเกิดแกสตรูเลชันจะไม่เกิดขึ้นเลย β-catenin ยังมีบทบาทสำคัญในการเหนี่ยวนำริมฝีปากบลาสโตพอร์ซึ่งเป็นจุดเริ่มต้นของการเกิดแกสตรูเลชัน[ 53 ]การยับยั้งการแปล GSK-3 โดยการฉีด antisense mRNA อาจทำให้เกิดช่องบลาสโตพอร์ที่สองและแกนร่างกายส่วนเกินได้ ผลกระทบที่คล้ายกันนี้อาจเกิดขึ้นจากการแสดงออกมากเกินไปของ β-catenin [ 54 ]
การแบ่งเซลล์แบบไม่สมมาตร
β-catenin ยังมีส่วนเกี่ยวข้องในการควบคุมชะตากรรมของเซลล์ผ่านการแบ่งเซลล์แบบไม่สมมาตรในสิ่งมีชีวิตต้นแบบC. elegansเช่นเดียวกับ เซลล์ไข่ ของ Xenopusซึ่งโดยพื้นฐานแล้วเป็นผลมาจากการกระจายตัวที่ไม่เท่ากันของDsh , Frizzled , axinและAPCในไซโตพลาสซึมของเซลล์แม่[ 55 ]
การต่ออายุสเต็มเซลล์
ผลลัพธ์ที่สำคัญที่สุดประการหนึ่งของการส่งสัญญาณ Wnt และระดับ β-catenin ที่สูงขึ้นในเซลล์บางชนิดคือการรักษาความสามารถในการเปลี่ยนแปลงเป็นเซลล์ ชนิดต่างๆ [ 51 ]ตัวอย่างเช่น อัตราของเซลล์ต้นกำเนิดในลำไส้ใหญ่ได้รับการรับรองโดยการสะสมของ β-catenin ซึ่งสามารถกระตุ้นได้โดยเส้นทาง Wnt [ 56 ]แรงทางกลจากการบีบตัวของลำไส้ใหญ่ที่มีความถี่สูงยังเกี่ยวข้องกับการรักษาระดับสมดุลของเซลล์ต้นกำเนิดในลำไส้ใหญ่โดยอาศัย β-catenin ผ่านกระบวนการถ่ายทอดทางกล คุณลักษณะนี้ได้รับการส่งเสริมในทางพยาธิวิทยาไปสู่การเพิ่มจำนวนเซลล์ที่มากเกินไปจนก่อให้เกิดเนื้องอกในเซลล์ที่แข็งแรงซึ่งถูกกดทับด้วยแรงดันเนื่องจากเซลล์เนื้องอกที่เพิ่มจำนวนมากเกินไปซึ่งมีการเปลี่ยนแปลงทางพันธุกรรม[ 57 ]
ในเซลล์ประเภทอื่นและระยะการพัฒนาต่างๆ β-catenin อาจส่งเสริมการเปลี่ยนแปลงไปเป็นเซลล์ชนิดต่างๆ โดยเฉพาะอย่างยิ่งเซลล์ในกลุ่ม เมโซเดอร์ม
การเปลี่ยนผ่านจากเซลล์เยื่อบุผิวไปเป็นเซลล์เนื้อเยื่อเกี่ยวพัน
β-catenin ยังทำหน้าที่เป็นมอร์โฟเจนในระยะหลังของการพัฒนาตัวอ่อน ร่วมกับTGF-βบทบาทสำคัญของ β-catenin คือการชักนำให้เกิดการเปลี่ยนแปลงทางสัณฐานวิทยาในเซลล์เยื่อบุผิว มันชักนำให้เซลล์เหล่านี้ละทิ้งการยึดเกาะที่แน่นหนาและมีลักษณะเป็น เมเซน ไคมอล ที่เคลื่อนไหวได้มากขึ้นและยึดเกาะกันอย่างหลวมๆ ในระหว่างกระบวนการนี้ เซลล์เยื่อบุผิวจะสูญเสียการแสดงออกของโปรตีนเช่นE-cadherin , Zonula occludens 1 (ZO1) และไซโตเคราตินในขณะเดียวกันพวกมันจะเปิดการแสดงออกของไวเมนติน , อัลฟา สมูท มัสเคิล แอคติน (ACTA2) และโปรตีนเฉพาะไฟโบรบลาสต์ 1 (FSP1) พวกมันยังผลิตส่วนประกอบของเมทริกซ์นอกเซลล์ เช่นคอลลาเจนชนิดที่ 1และไฟโบรเนกตินการกระตุ้นที่ผิดปกติของวิถี Wnt มีส่วนเกี่ยวข้องกับกระบวนการทางพยาธิวิทยา เช่น ไฟโบรซิสและมะเร็ง[ 58 ] ในการพัฒนาของกล้ามเนื้อหัวใจ β-catenin มีบทบาทสองเฟส ในขั้นต้น การกระตุ้น Wnt/β-catenin เป็นสิ่งจำเป็นสำหรับการกำหนดเซลล์มีเซนไคม์ให้กลายเป็นเซลล์หัวใจ อย่างไรก็ตาม ในระยะพัฒนาการที่ช้ากว่านั้น จำเป็นต้องลดระดับ β-catenin ลง[ 59 ] [ 60 ] [ 46 ]
การมีส่วนร่วมในสรีรวิทยาของหัวใจ
ในกล้ามเนื้อหัวใจ β-catenin จะสร้างคอมเพล็กซ์กับN-cadherinที่จุดเชื่อมต่อแบบ adherens junctionภายในโครงสร้างintercalated disc ซึ่งมีหน้าที่ในการเชื่อมต่อทางไฟฟ้าและทางกลของเซลล์หัวใจที่อยู่ติดกัน การศึกษาในแบบจำลองของเซลล์ กล้ามเนื้อหัวใจห้องล่าง ของหนูโตเต็มวัย แสดงให้เห็นว่าการปรากฏและการกระจายตัวของ β-catenin ถูกควบคุมตามพื้นที่และเวลาในระหว่างการเปลี่ยนแปลงสภาพของเซลล์เหล่านี้ในวัฒนธรรม โดยเฉพาะอย่างยิ่ง β-catenin เป็นส่วนหนึ่งของคอมเพล็กซ์ที่แตกต่างกันกับ N-cadherin และalpha-cateninซึ่งมีอยู่มากมายที่จุดเชื่อมต่อแบบ adherens junction ในระยะเริ่มต้นหลังจาก การแยก เซลล์กล้ามเนื้อหัวใจเพื่อการสร้างการสัมผัสระหว่างเซลล์ขึ้นใหม่[ 61 ]พบว่า β-catenin สร้างคอมเพล็กซ์กับemerinในเซลล์กล้ามเนื้อหัวใจที่จุดเชื่อมต่อแบบ adherens junction ภายใน intercalated disc และปฏิสัมพันธ์นี้ขึ้นอยู่กับการมีอยู่ของ ตำแหน่ง การฟอสโฟรีเลชั่น ของ GSK 3-betaบน β-catenin การกำจัดอีเมอรินทำให้ตำแหน่งของ β-catenin และสถาปัตยกรรมของแผ่นเชื่อมต่อระหว่างเซลล์โดยรวมเปลี่ยนแปลงไปอย่างมาก ซึ่งมีลักษณะคล้ายกับโรคกล้ามเนื้อหัวใจโต[ 62 ]
ในแบบจำลองสัตว์ของ โรค หัวใจหน้าที่ของ β-catenin ได้ถูกเปิดเผย ในแบบจำลองหนูตะเภาของภาวะลิ้นหัวใจเอออร์ติกตีบและภาวะ หัวใจห้องซ้ายโต β-catenin แสดงให้เห็นว่ามีการเปลี่ยนแปลงตำแหน่งภายในเซลล์จากแผ่นเชื่อมต่อระหว่างเซลล์ไปยังไซโตโซลแม้ว่าจะไม่มีการเปลี่ยนแปลงในปริมาณ β-catenin โดยรวมของเซลล์ก็ตามVinculinแสดงรูปแบบการเปลี่ยนแปลงที่คล้ายกัน N-cadherin ไม่แสดงการเปลี่ยนแปลง และไม่มีการเพิ่มขึ้นของplakoglobinที่แผ่นเชื่อมต่อระหว่างเซลล์เพื่อชดเชยในกรณีที่ไม่มี β-catenin [ 63 ]ในแบบจำลองแฮมสเตอร์ของโรคกล้ามเนื้อหัวใจและภาวะหัวใจล้มเหลว การยึดเกาะระหว่างเซลล์ไม่สม่ำเสมอและไม่เป็นระเบียบ และระดับการแสดงออกของ adherens junction/แผ่นเชื่อมต่อระหว่างเซลล์และ กลุ่ม นิวเคลียสของ β-catenin ลดลง[ 64 ]ข้อมูลเหล่านี้ชี้ให้เห็นว่าการสูญเสีย β-catenin อาจมีบทบาทในแผ่นเชื่อมต่อระหว่างเซลล์ที่เป็นโรคซึ่งเกี่ยวข้องกับภาวะกล้ามเนื้อหัวใจโตและภาวะหัวใจล้มเหลว ในแบบจำลองหนูที่เป็นโรคกล้ามเนื้อหัวใจขาดเลือดการถ่ายโอนยีนอะดีโนไวรัสของβ-catenin ที่ ไม่ สามารถฟอสโฟรีเลตได้ และทำงานอย่างต่อเนื่อง ช่วยลดขนาดของ MI กระตุ้น วงจรเซลล์และลดปริมาณอะพอพโทซิสในเซลล์กล้ามเนื้อหัวใจและไมโอไฟโบรบลาสต์ ของหัวใจ การค้นพบนี้สอดคล้องกับการแสดงออกที่เพิ่มขึ้นของโปรตีนที่ส่งเสริมการอยู่รอด เช่นsurvivinและBcl-2และปัจจัยการเจริญเติบโตของหลอดเลือดในขณะที่ส่งเสริมการแยกตัวของไฟโบรบลาสต์ ของหัวใจ ไปเป็นไมโอไฟโบรบลาสต์ การค้นพบเหล่านี้ชี้ให้เห็นว่า β-catenin สามารถส่งเสริมกระบวนการสร้างใหม่และการรักษาหลังเกิดภาวะกล้ามเนื้อหัวใจขาดเลือด[ 65 ]ใน แบบจำลองหนูที่ เป็นโรคหัวใจ ล้มเหลวจากความดัน โลหิตสูง โดยธรรมชาติ นักวิจัยตรวจพบการเคลื่อนย้ายของ β-catenin จากแผ่นเชื่อมต่อ/ ซาร์โคเลมมาไปยังนิวเคลียสซึ่งเห็นได้จากการลดลงของการแสดงออกของ β-catenin ในส่วนของโปรตีนเยื่อหุ้มเซลล์และการเพิ่มขึ้นในส่วนของนิวเคลียส นอกจากนี้ พวกเขายังพบว่าความสัมพันธ์ระหว่างไกลโคเจนซินเทสไคเนส-3βและ β-catenin อ่อนลง ซึ่งอาจบ่งชี้ถึงความเสถียรของโปรตีนที่เปลี่ยนแปลงไป โดยรวมแล้ว ผลลัพธ์ชี้ให้เห็นว่าการแปลตำแหน่งนิวเคลียสของ β-catenin ที่เพิ่มขึ้นอาจมีความสำคัญต่อความก้าวหน้าของภาวะหัวใจโต[ 66 ]
Regarding the mechanistic role of β-catenin in cardiac hypertrophy, transgenic mouse studies have shown somewhat conflicting results regarding whether upregulation of β-catenin is beneficial or detrimental.[67][68][69] A recent study using a conditional knockout mouse that either lacked β-catenin altogether or expressed a non-degradable form of β-catenin in cardiomyocytes reconciled a potential reason for these discrepancies. There appears to be strict control over the subcellular localization of β-catenin in cardiac muscle. Mice lacking β-catenin had no overt phenotype in the left ventricular myocardium; however, mice harboring a stabilized form of β-catenin developed dilated cardiomyopathy, suggesting that the temporal regulation of β-catenin by protein degradation mechanisms is critical for normal functioning of β-catenin in cardiac cells.[70] In a mouse model harboring knockout of a desmosomal protein, plakoglobin, implicated in arrhythmogenic right ventricular cardiomyopathy, the stabilization of β-catenin was also enhanced, presumably to compensate for the loss of its plakoglobin homolog. These changes were coordinate with Akt activation and glycogen synthase kinase 3β inhibition, suggesting once again that the abnormal stabilization of β-catenin may be involved in the development of cardiomyopathy.[71] Further studies employing a double knockout of plakoglobin and β-catenin showed that the double knockout developed cardiomyopathy, fibrosis and arrhythmias resulting in sudden cardiac death. Intercalated disc architecture was severely impaired and connexin 43-resident gap junctions were markedly reduced. Electrocardiogram measurements captured spontaneous lethal ventricular arrhythmias in the double transgenic animals, suggesting that the two catenins—β-catenin and plakoglobin—are critical and indispensable for mechanoelectrical coupling in cardiomyocytes.[72]
Clinical significance
Role in depression
ความสามารถของสมองของแต่ละบุคคลในการรับมือกับความเครียดได้อย่างมีประสิทธิภาพหรือไม่ และด้วยเหตุนี้จึงส่งผลต่อความเสี่ยงต่อภาวะซึมเศร้า ขึ้นอยู่กับ β-catenin ในสมองของแต่ละคน ตามการศึกษาที่ดำเนินการโดยIcahn School of Medicine at Mount Sinaiและตีพิมพ์เมื่อวันที่ 12 พฤศจิกายน 2014 ในวารสารNature [ 73 ] การส่งสัญญาณ β-catenin ที่สูงขึ้นจะเพิ่มความยืดหยุ่นในพฤติกรรม ในขณะที่การส่งสัญญาณ β - catenin ที่บกพร่องจะนำไปสู่ภาวะซึมเศร้าและการจัดการความเครียดที่ลดลง[ 73 ]
บทบาทในโรคหัวใจ
การเปลี่ยนแปลงรูปแบบการแสดงออกของ β-catenin เกี่ยวข้องกับโรคกล้ามเนื้อหัวใจห้องล่างขยายตัวในมนุษย์ โดยทั่วไปแล้วพบว่ามีการเพิ่มขึ้นของการแสดงออกของ β-catenin ในผู้ป่วยที่มีโรคกล้ามเนื้อหัวใจห้องล่าง ขยายตัว [ 74 ]ในการศึกษาเฉพาะเจาะจง ผู้ป่วยที่มีโรคกล้ามเนื้อหัวใจห้องล่างขยายตัวระยะสุดท้ายแสดงให้เห็น ระดับ mRNAและโปรตีนของตัวรับเอสโตรเจนอัลฟา (ER-alpha) เพิ่มขึ้นเกือบสองเท่า และปฏิสัมพันธ์ระหว่าง ER-alpha/beta-catenin ซึ่งมีอยู่ในแผ่นเชื่อมต่อระหว่างเซลล์ของหัวใจมนุษย์ที่ไม่เป็นโรคในกลุ่มควบคุมนั้นหายไป ซึ่งบ่งชี้ว่าการสูญเสียปฏิสัมพันธ์นี้ที่แผ่นเชื่อมต่อระหว่างเซลล์อาจมีบทบาทในการดำเนินไปของภาวะหัวใจล้มเหลว[ 75 ] β-catenin ร่วมกับ โปรตีน BCL9และ PYGO ประสานงานด้านต่างๆ ของการพัฒนาหัวใจ และการกลายพันธุ์ในBcl9หรือPygoในสิ่งมีชีวิตต้นแบบ เช่น หนูและปลาซีบราฟิช ทำให้เกิดฟีโนไทป์ที่คล้ายคลึงกับความผิดปกติของหัวใจพิการแต่กำเนิดใน มนุษย์มาก [ 76 ]
การมีส่วนร่วมในโรคมะเร็ง

β-catenin เป็นโปรโตออนโคยีน การกลายพันธุ์ของยีน นี้พบได้ทั่วไปในมะเร็งหลายชนิด ได้แก่มะเร็งตับชนิดปฐมภูมิมะเร็งลำไส้ใหญ่มะเร็งรังไข่ มะเร็งเต้านมมะเร็งปอด และกลิโอบลาสโตมามีการประมาณการว่าประมาณ 10% ของตัวอย่างเนื้อเยื่อทั้งหมดที่นำมาวิเคราะห์ลำดับจากมะเร็งทุกชนิดแสดงการกลายพันธุ์ในยีน CTNNB1 [ 77 ]การกลายพันธุ์ส่วนใหญ่เหล่านี้กระจุกตัวอยู่ในบริเวณเล็กๆ ของส่วนปลาย N ของ β-catenin: โมทีฟการจับ β-TrCP การกลายพันธุ์ที่ทำให้สูญเสียการทำงานของโมทีฟนี้ทำให้การยูบิควิตินิเลชันและการย่อยสลายของ β-catenin เป็นไปไม่ได้ ซึ่งจะทำให้ β-catenin เคลื่อนย้ายไปยังนิวเคลียสโดยไม่มีสิ่งกระตุ้นภายนอก และขับเคลื่อนการถอดรหัสของยีนเป้าหมายอย่างต่อเนื่อง ระดับ β-catenin ในนิวเคลียสที่เพิ่มขึ้นยังพบในมะเร็งเซลล์ฐาน (BCC) [ 78 ]มะเร็งเซลล์สความัสของศีรษะและลำคอ (HNSCC) มะเร็งต่อมลูกหมาก (CaP) [ 79 ] ไพโลแมทริกซ์มา (PTR) [ 80 ]และเมดุลโลบลาสโตมา (MDB) [ 81 ]ข้อสังเกตเหล่านี้อาจบ่งชี้หรือไม่บ่งชี้ถึงการกลายพันธุ์ในยีน β-catenin ก็ได้ ส่วนประกอบอื่นๆ ของวิถี Wnt ก็อาจทำงานผิดปกติได้เช่นกัน



Similar mutations are also frequently seen in the β-catenin recruiting motifs of APC. Hereditary loss-of-function mutations of APC cause a condition known as familial adenomatous polyposis. Affected individuals develop hundreds of polyps in their large intestine. Most of these polyps are benign in nature, but they have the potential to transform into deadly cancer as time progresses. Somatic mutations of APC in colorectal cancer are also not uncommon.[85] β-Catenin and APC are among the key genes (together with others, like K-Ras and SMAD4) involved in colorectal cancer development. The potential of β-catenin to change the previously epithelial phenotype of affected cells into an invasive, mesenchyme-like type contributes greatly to metastasis formation.
As a therapeutic target
เนื่องจาก β-catenin มีส่วนเกี่ยวข้องกับการพัฒนาของมะเร็ง การยับยั้ง β-catenin จึงยังคงได้รับความสนใจอย่างมาก แต่การกำหนดเป้าหมายไปยังบริเวณที่จับกับ β-catenin บนโดเมน armadillo นั้นไม่ใช่เรื่องง่าย เนื่องจากพื้นผิวของมันกว้างขวางและค่อนข้างเรียบ อย่างไรก็ตาม การจับกับ "จุดสำคัญ" ขนาดเล็กบนพื้นผิวนี้ก็เพียงพอแล้วสำหรับการยับยั้งที่มีประสิทธิภาพ ด้วยวิธีนี้ เพปไทด์แบบเกลียว "เย็บติด" ที่ได้มาจากโมทีฟการจับ β-catenin ตามธรรมชาติที่พบใน LEF1 จึงเพียงพอสำหรับการยับยั้งการถอดรหัสที่ขึ้นอยู่กับ β-catenin อย่างสมบูรณ์ เมื่อเร็วๆ นี้ สารประกอบโมเลกุลขนาดเล็กหลายชนิดได้รับการพัฒนาขึ้นเพื่อกำหนดเป้าหมายไปยังบริเวณที่มีประจุบวกสูงเดียวกันของโดเมน ARM (CGP049090, PKF118-310, PKF115-584 และ ZTM000990) นอกจากนี้ ระดับของ β-catenin ยังสามารถได้รับอิทธิพลโดยการกำหนดเป้าหมายไปยังส่วนประกอบต้นน้ำของวิถี Wnt รวมถึงคอมเพล็กซ์การทำลาย β-catenin ด้วย[ 86 ]ช่องยึดเกาะ N-terminal เพิ่มเติมยังมีความสำคัญต่อการกระตุ้นยีนเป้าหมาย Wnt (จำเป็นสำหรับการดึงดูด BCL9) บริเวณนี้ของโดเมน ARM สามารถกำหนดเป้าหมายทางเภสัชวิทยาได้โดยใช้กรดคาร์โนซิกเป็นต้น[ 87 ]บริเวณ "เสริม" นั้นเป็นเป้าหมายที่น่าสนใจอีกเป้าหมายหนึ่งสำหรับการพัฒนายา[ 88 ]แม้ว่าจะมีการวิจัยก่อนคลินิกอย่างเข้มข้น แต่ก็ยังไม่มีสารยับยั้ง β-catenin ที่พร้อมใช้งานเป็นตัวยาบำบัด อย่างไรก็ตาม สามารถตรวจสอบการทำงานของมันเพิ่มเติมได้โดยการน็อคดาวน์ด้วย siRNA โดยอาศัยการตรวจสอบความถูกต้องที่เป็นอิสระ[ 89 ]แนวทางการรักษาอีกวิธีหนึ่งสำหรับการลดการสะสมของ β-catenin ในนิวเคลียสคือการยับยั้งกาแลคติน-3 [ 90 ]สารยับยั้งกาแลคติน-3 GR-MD-02กำลังอยู่ระหว่างการทดลองทางคลินิกโดยใช้ร่วมกับยา ipilimumab ในขนาดที่ FDA อนุมัติในผู้ป่วยที่เป็นมะเร็งผิวหนังขั้นสูง[ 91 ]โปรตีนBCL9และBCL9Lได้รับการเสนอให้เป็นเป้าหมายในการรักษาโรคมะเร็งลำไส้ใหญ่ที่มีการส่งสัญญาณ Wnt มากเกินไป เนื่องจากการลบโปรตีนเหล่านี้ไม่รบกวนภาวะสมดุลปกติ แต่ส่งผลกระทบอย่างมากต่อพฤติกรรมการแพร่กระจาย[ 92 ]
บทบาทในกลุ่มอาการทารกในครรภ์ที่ได้รับแอลกอฮอล์
การทำให้ β-catenin ไม่เสถียรโดยเอทานอลเป็นหนึ่งในสองกลไกที่ทราบกันดีว่าการได้รับแอลกอฮอล์ทำให้เกิดกลุ่มอาการทารกในครรภ์ที่ได้รับแอลกอฮอล์ (อีกกลไกหนึ่งคือการขาดโฟเลตที่เกิดจากเอทานอล) เอทานอลนำไปสู่การทำให้ β-catenin ไม่เสถียรผ่านกลไกที่ขึ้นอยู่กับโปรตีน G โดยที่ Phospholipase Cβ ที่ถูกกระตุ้นจะไฮโดรไลซ์ phosphatidylinositol-(4,5)-bisphosphate ไปเป็น diacylglycerol และ inositol-(1,4,5)-trisphosphate inositol-(1,4,5)-trisphosphate ที่ละลายได้จะกระตุ้นให้แคลเซียมถูกปล่อยออกมาจากเอนโดพลาสมิกเรติคูลัม การเพิ่มขึ้นอย่างฉับพลันของแคลเซียมในไซโตพลาสซึมนี้จะกระตุ้น Ca2+/calmodulin-dependent protein kinase (CaMKII) CaMKII ที่ถูกกระตุ้นจะทำให้ β-catenin ไม่เสถียรผ่านกลไกที่ยังไม่เป็นที่เข้าใจอย่างชัดเจน แต่คาดว่าเกี่ยวข้องกับการฟอสโฟรีเลชันของ β-catenin โดย CaMKII โปรแกรมการถอดรหัส β-catenin (ซึ่งจำเป็นสำหรับการพัฒนาเซลล์ยอดประสาทตามปกติ) จึงถูกยับยั้ง ส่งผลให้เซลล์ยอดประสาทตายก่อนวัยอันควร[ 93 ]
ปฏิสัมพันธ์
มีการค้นพบว่า β-catenin มีปฏิกิริยากับ:
- APC , [ 94 ] [ 95 ] [ 96 ] [ 97 ] [ 98 ] [ 99 ] [ 100 ] [ 101 ]
- ARID1A , [ 24 ]
- AXIN1 , [ 102 ] [ 103 ]
- ตัวรับแอนโดรเจน [ 104 ] [ 105 ] [ 106 ] [ 107 ] [ 108 ] [ 109 ]
- CBY1 , [ 110 ]
- CDH1 , [ 25 ] [ 95 ] [ 111 ] [ 112 ] [ 113 ] [ 114 ] [ 115 ] [ 116 ] [ 117 ] [ 118 ] [ 119 ] [ 120 ] [ 121 ] [ 122 ] [ 123 ] [ 124 ] [ 125 ] [ 126 ] [ 127 ] [ 128 ] [ 129 ] [ 130 ] [ 131 ]
- CDH2 , [ 61 ] [ 132 ] [ 133 ]
- CDH3 , [ 130 ] [ 134 ]
- CDK5R1 , [ 135 ]
- ชุก , [ 136 ]
- CTNND1 , [ 95 ] [ 116 ]
- CTNNA1 , [ 112 ] [ 121 ] [ 137 ] [ 138 ] [ 139 ]
- EGFR , [ 116 ] [ 125 ] [ 140 ]
- เอเมอริน[ 141 ] [ 142 ]
- ESR1 [ 75 ]
- FHL2 , [ 143 ]
- GSK3B , [ 97 ] [ 144 ]
- HER2/นิว , [ 117 ] [ 140 ] [ 145 ]
- HNF4A , [ 108 ]
- IKK2 , [ 136 ]
- LEF1 [ 146 ] [ 147 ] [ 148 ] [ 149 ]รวมถึงแบบทรานส์เจนิก[ 150 ]
- MAGI1 , [ 126 ]
- MUC1 , [ 118 ] [ 151 ] [ 152 ] [ 153 ] [ 154 ] [ 155 ] [ 156 ]
- NR5A1 , [ 157 ] [ 158 ] [ 24 ]
- PCAF , [ 159 ]
- PHF17 , [ 160 ]
- พลาโคโกลบิน [ 95 ] [ 116 ]
- PTPN14 , [ 161 ]
- PTPRF , [ 117 ] [ 162 ]
- PTPRK (PTPkappa), [ 163 ]
- PTPRT (PTPrho), [ 164 ]
- PTPRU (PCP-2), [ 165 ] [ 166 ] [ 167 ]
- PSEN1 , [ 168 ] [ 169 ] [ 170 ]
- PTK7 [ 171 ]
- RuvB-like 1 , [ 172 ]
- SMAD7 , [ 146 ]
- SMARCA4 [ 173 ]
- SLC9A3R1 , [ 120 ]
- SWI/SNF , [ 24 ]
- USP9X , [ 174 ] และ
- วี-แคดเฮริน[ 175 ] [ 176 ]
- XIRP1 [ 177 ]
ดูเพิ่มเติม
อ่านเพิ่มเติม
- Kikuchi A (กุมภาพันธ์ 2000). "การควบคุมการส่งสัญญาณเบต้า-แคทเทนินในวิถี Wnt". Biochemical and Biophysical Research Communications . 268 (2): 243– 248. Bibcode : 2000BBRC..268..243K . doi : 10.1006/bbrc.1999.1860 . PMID 10679188 .
- Wilson PD (เมษายน 2544). "Polycystin: แง่มุมใหม่ของโครงสร้าง หน้าที่ และการควบคุม"วารสารของสมาคมโรคไตแห่งอเมริกา 12 ( 4): 834– 845. doi : 10.1681/ASN.V124834 . PMID 11274246 .
- Kalluri R, Neilson EG (ธันวาคม 2003). "การเปลี่ยนผ่านจากเยื่อบุผิวเป็นเนื้อเยื่อเกี่ยวพันและผลกระทบต่อการเกิดพังผืด"วารสารการวิจัยทางคลินิก 112 ( 12): 1776– 1784. doi : 10.1172/JCI20530 . PMC 297008 . PMID 14679171 .
- De Ferrari GV, Moon RT (ธันวาคม 2006). "การขึ้นลงของสัญญาณ Wnt ในความผิดปกติทางระบบประสาทที่พบได้บ่อย" Oncogene . 25 (57): 7545– 7553. doi : 10.1038/sj.onc.1210064 . hdl : 10533/178151 . PMID 17143299 . S2CID 35684619 .
ลิงก์ภายนอก
- beta+Catenin ที่ หัวข้อทางการ แพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
- "โปรตีนหลากหลายชนิดมีบทบาทในการควบคุมวิถีการส่งสัญญาณ Wnt/β-catenin แบบดั้งเดิม" (ดูข้อมูลเพิ่มเติมได้ที่ cancer.gov)
- "บทบาทของ β-catenin ในการส่งสัญญาณ การกำหนดชะตากรรมของเซลล์ และการเปลี่ยนสภาพเซลล์" ที่ nih.gov
- "นักวิจัยเสนอหลักฐานโดยตรงครั้งแรกเกี่ยวกับวิธีการที่โรคข้ออักเสบทำลายกระดูกอ่อน" ที่ rochester.edu
- ตำแหน่งจีโนมของยีน CTNNB1ในมนุษย์และรายละเอียดของยีนCTNNB1 ใน UCSC Genome Browser
บทความนี้ได้นำข้อความจากหอสมุดแห่งชาติสหรัฐอเมริกาด้านการแพทย์ มา ใช้ ซึ่งเป็นข้อมูลสาธารณะ
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ แคทเทนินเบตา-1
แคทเทนินเบตา-1 หรือที่รู้จักกันในชื่อ β-แคทเทนิน ( beta - catenin) เป็น โปรตีน ที่ในมนุษย์ถูกสร้างขึ้นโดย ยีน CTNNB1
การค้นพบ
β-catenin ถูกค้นพบครั้งแรกในช่วงต้นทศวรรษ 1990 ในฐานะส่วนประกอบของ คอมเพล็กซ์ การยึดเกาะเซลล์ ของสัตว์เลี้ยงลูกด้วยนม : โปรตีนที่รับผิดชอบในการยึดเกาะไซโตพลาสซึมของ แค ด เฮริน [ 11 ] แต่ในไม่ช้าก็พบว่าโปรตีน อาร์ มาดิลโลของแมลงหวี่...
โครงสร้างโปรตีน
แกนกลางของ β-catenin ประกอบด้วย ส่วนที่ซ้ำกัน ที่มีลักษณะเฉพาะหลาย ส่วน แต่ละส่วนมีความยาวประมาณ 40 กรดอะมิโน เรียกว่า armadillo repeats องค์ประกอบทั้งหมดเหล่านี้จะพับรวมกันเป็น โดเมนโปรตีน ที่แข็งแรงเพียงโดเมนเดียวที่มีรูปร่างยาว เรียกว่า armadillo domain...
การจับกับโดเมนอาร์มาดิลโล
ดังที่ได้กล่าวไว้ข้างต้น โดเมน ARM ของ β-catenin ทำหน้าที่เป็นแพลตฟอร์มที่ โมทีฟเชิงเส้น เฉพาะ สามารถจับได้ โมทีฟที่จับกับ β-catenin ซึ่งอยู่ในพันธมิตรที่มีโครงสร้างหลากหลาย มักจะ ไม่มีระเบียบ ในตัวเอง และโดยทั่วไปจะรับเอาโครงสร้างที่แข็งตัวเมื่อจับกับโดเมน...