อ่าน 30 นาที
การแสดงออกของยีน
การแสดงออกของยีนคือกระบวนการที่ข้อมูลที่มีอยู่ในยีนถูกนำมาใช้เพื่อสร้างผลิตภัณฑ์ของยีนที่มีฟังก์ชันการทำงาน เช่นโปรตีนหรือ โมเลกุล RNA ที่มีฟังก์ชันการทำงาน...
การแสดงออกของยีน
การแสดงออกของยีนคือกระบวนการที่ข้อมูลที่มีอยู่ในยีนถูกนำมาใช้เพื่อสร้างผลิตภัณฑ์ของยีนที่มีฟังก์ชันการทำงาน เช่นโปรตีนหรือ โมเลกุล RNA ที่มีฟังก์ชันการทำงาน กระบวนการนี้เกี่ยวข้องกับหลายขั้นตอน รวมถึงการถอดรหัสลำดับของยีนเป็น RNA สำหรับยีนที่เข้ารหัสโปรตีน RNA นี้จะถูกแปล ต่อ ไปเป็นสายของกรดอะมิโนที่พับตัวเป็นโปรตีน ในขณะที่สำหรับยีนที่ไม่เข้ารหัส RNA ที่ได้นั้นเองก็มีบทบาทในเซลล์ การแสดงออกของยีนช่วยให้เซลล์สามารถใช้ข้อมูลทางพันธุกรรมในยีนเพื่อดำเนินฟังก์ชันทางชีวภาพได้หลากหลาย ในขณะที่ระดับการแสดงออกสามารถควบคุมได้เพื่อตอบสนองต่อความต้องการของเซลล์และการเปลี่ยนแปลงของสิ่งแวดล้อม แต่ยีนบางชนิดก็แสดงออกอย่างต่อเนื่องโดยมีการเปลี่ยนแปลงเพียงเล็กน้อย[ 1 ]
กลไก
การถอดเสียง

การสร้างสำเนา RNA จากสาย DNA เรียกว่าการถอดรหัส (transcription ) และดำเนินการโดยRNA polymeraseซึ่งจะเพิ่มไรโบนิวคลีโอไทด์ทีละตัวลงในสาย RNA ที่กำลังเติบโตตาม กฎ ความเสริมกันของเบสนิวคลีโอไทด์ RNA นี้จะเสริมกับสาย DNA แม่แบบ 3′ → 5′ [ 2 ]ยกเว้นว่าไทมีน (T) จะถูกแทนที่ด้วยยูราซิล (U) ใน RNA และอาจมีข้อผิดพลาดเกิดขึ้นได้
In bacteria, transcription is carried out by a single type of RNA polymerase, which needs to bind a DNA sequence called a Pribnow box with the help of the sigma factor protein (σ factor) to start transcription. In eukaryotes, transcription is performed in the nucleus by three types of RNA polymerases, each of which needs a special DNA sequence called the promoter and a set of DNA-binding proteins—transcription factors—to initiate the process (see regulation of transcription below). RNA polymerase I is responsible for transcription of ribosomal RNA (rRNA) genes. RNA polymerase II (Pol II) transcribes all protein-coding genes but also some non-coding RNAs (e.g., snRNAs, snoRNAs or long non-coding RNAs). RNA polymerase III transcribes 5S rRNA, transfer RNA (tRNA) genes, and some small non-coding RNAs (e.g., 7SK). Transcription ends when the polymerase encounters a sequence called the terminator.
mRNA processing
While transcription of prokaryotic protein-coding genes creates messenger RNA (mRNA) that is ready for translation into protein, transcription of eukaryotic genes leaves a primary transcript of RNA (pre-RNA), which first has to undergo a series of modifications to become a mature RNA. Types and steps involved in the maturation processes vary between coding and non-coding preRNAs; i.e. even though preRNA molecules for both mRNA and tRNA undergo splicing, the steps and machinery involved are different.[3] The processing of non-coding RNA is described below (non-coding RNA maturation).
The processing of pre-mRNA include 5′ capping, which is set of enzymatic reactions that add 7-methylguanosine (m7G) to the 5′ end of pre-mRNA and thus protect the RNA from degradation by exonucleases.[4] The m7G cap is then bound by cap binding complex heterodimer (CBP20/CBP80), which aids in mRNA export to cytoplasm and also protect the RNA from decapping.[5]
การดัดแปลงอีกอย่างหนึ่งคือการตัด 3′ และการเติมโพลีอะดีนีน [ 6 ] เกิดขึ้นหากมีลำดับสัญญาณการเติมโพลีอะดีนีน (5′- AAUAAA-3′) อยู่ในพรี-mRNA ซึ่งมักจะอยู่ระหว่างลำดับการเข้ารหัสโปรตีนและเทอร์มิเนเตอร์[ 7 ]พรี-mRNA จะถูกตัดก่อน จากนั้นจะมีการเติมอะดีนีน (A) ประมาณ 200 ตัวเพื่อสร้างหางโพลี(A) ซึ่งช่วยปกป้อง RNA จากการเสื่อมสภาพ[ 8 ]หางโพลี(A) จะถูกจับโดยโปรตีนที่จับกับโพลี(A) หลายตัว (PABPs)ซึ่งจำเป็นสำหรับการส่งออก mRNA และการเริ่มต้นการแปลใหม่[ 9 ]ในกระบวนการย้อนกลับของการกำจัดอะดีนีน หางโพลี(A) จะสั้นลงโดย เอ็กโซนิวคลีเอส CCR4-Not 3′-5′ ซึ่งมักนำไปสู่การสลายตัวของทรานสคริปต์ทั้งหมด[ 10 ]

การดัดแปลงที่สำคัญมากอย่างหนึ่งของพรีเอ็มอาร์เอ็นเอของยูคาริโอตคือการตัดต่อ อาร์เอ็นเอ พรีเอ็มอาร์เอ็นเอของยูคาริโอตส่วนใหญ่ประกอบด้วยส่วนสลับกันที่เรียกว่าเอ็กซอนและอินทรอน[ 11 ]ในระหว่างกระบวนการตัดต่อ คอมเพล็กซ์เร่งปฏิกิริยาของอาร์เอ็นเอ-โปรตีนที่เรียกว่าสไปโซโซม จะเร่งปฏิกิริยาการ ถ่ายโอนเอสเทอร์สองครั้งซึ่งจะกำจัดอินทรอนและปล่อยออกมาในรูปโครงสร้างแบบห่วง จากนั้นจึงตัดต่อเอ็กซอนที่อยู่ติดกันเข้าด้วยกัน[ 12 ]ในบางกรณี อินทรอนหรือเอ็กซอนบางส่วนอาจถูกกำจัดออกหรือคงไว้ในเอ็มอาร์เอ็นเอที่เจริญเต็มที่[ 13 ]การตัดต่อแบบทางเลือกนี้สร้างชุดของทรานสคริปต์ที่แตกต่างกันซึ่งมีต้นกำเนิดมาจากยีนเดียว เนื่องจากทรานสคริปต์เหล่านี้อาจถูกแปลเป็นโปรตีนที่แตกต่างกัน การตัดต่อจึงขยายความซับซ้อนของการแสดงออกของยีนยูคาริโอตและขนาดของโปรตีโอม ของสปีชี ส์[ 14 ]
การประมวลผล RNA อย่างกว้างขวางอาจเป็น ข้อได้ เปรียบเชิงวิวัฒนาการที่เกิดขึ้นได้จากนิวเคลียสของยูคาริโอต ในโปรคาริโอต การถอดรหัสและการแปลเกิดขึ้นพร้อมกัน ในขณะที่ในยูคาริโอตเยื่อหุ้มนิวเคลียสจะแยกกระบวนการทั้งสองออกจากกัน ทำให้มีเวลาสำหรับการประมวลผล RNA [ 15 ]
การเจริญเติบโตของ RNA ที่ไม่เข้ารหัส
ในสิ่งมีชีวิตส่วนใหญ่ยีนที่ไม่เข้ารหัส (ncRNA)จะถูกถอดรหัสเป็นสารตั้งต้นที่ต้องผ่านกระบวนการเพิ่มเติม ในกรณีของไรโบโซมอลอาร์เอ็นเอ (rRNA) มักจะถูกถอดรหัสเป็นพรี-rRNA ซึ่งประกอบด้วย rRNA หนึ่งตัวหรือมากกว่านั้น พรี-rRNA จะถูกตัดและดัดแปลง ( การเติมหมู่ เมทิลที่ตำแหน่ง 2′- Oและ การสร้าง ซูโดอูริดีน ) ที่ตำแหน่งเฉพาะโดยอาร์เอ็นเอขนาดเล็กประมาณ 150 ชนิดที่จำกัดอยู่ในนิวคลีโอลัส เรียกว่า snoRNA snoRNA จะรวมตัวกับโปรตีน เกิดเป็น snoRNP ในขณะที่ส่วนของ snoRNA จับคู่เบสกับอาร์เอ็นเอเป้าหมายและวางตำแหน่งการดัดแปลงที่ตำแหน่งที่แม่นยำ ส่วนของโปรตีนจะทำหน้าที่เร่งปฏิกิริยา ในยูคาริโอต โดยเฉพาะอย่างยิ่ง snoRNP ที่เรียกว่า RNase, MRP จะตัดพรี-rRNA 45Sออกเป็นrRNA 28S , 5.8Sและ18S rRNA และปัจจัยการประมวลผล RNA ก่อตัวเป็นกลุ่มก้อนขนาดใหญ่ที่เรียกว่านิวคลีโอลัส[ 16 ]
ในกรณีของ transfer RNA (tRNA) ตัวอย่างเช่น ลำดับ 5′ จะถูกกำจัดออกโดยRNase P [ 17 ] ในขณะที่ปลาย 3′ จะถูกกำจัดออกโดยเอนไซม์tRNase Z [ 18 ]และหาง 3′ CCA ที่ไม่มีแม่แบบจะถูกเพิ่มโดยนิวคลีโอไทด์ทรานสเฟอ เร ส[ 19 ]ในกรณีของmicro RNA (miRNA) miRNA จะถูกถอดรหัสเป็นทรานสคริปต์หลักหรือ pri-miRNA ก่อน โดยมีหมวกและหางโพลี-A และถูกประมวลผลเป็นโครงสร้างก้านห่วงสั้น ๆ 70 นิวคลีโอไทด์ที่เรียกว่า pre-miRNA ในนิวเคลียสของเซลล์โดยเอนไซม์DroshaและPashaหลังจากถูกส่งออกแล้ว มันจะถูกประมวลผลเป็น miRNA ที่โตเต็มที่ในไซโตพลาสซึมโดยการโต้ตอบกับเอนโดนิวคลีเอสDicerซึ่งยังเริ่มต้นการก่อตัวของคอมเพล็กซ์การยับยั้งที่เกิดจาก RNA (RISC)ซึ่งประกอบด้วยโปรตีน Argonaute
แม้แต่ snRNA และ snoRNA เองก็ยังต้องผ่านกระบวนการดัดแปลงหลายขั้นตอนก่อนที่จะกลายเป็นส่วนหนึ่งของคอมเพล็กซ์ RNP ที่ใช้งานได้[ 20 ]กระบวนการนี้เกิดขึ้นในนิวคลีโอพลาสม์หรือในช่องพิเศษที่เรียกว่าCajal bodies [ 21 ]เบสของพวกมันจะถูกเมทิลเลตหรือซูโดอริดิเนเลตโดยกลุ่มของRNA ขนาดเล็กที่เฉพาะเจาะจงกับ Cajal body (scaRNA)ซึ่งมีโครงสร้างคล้ายกับ snoRNA [ 22 ]
การแปล
สำหรับ RNA ที่ไม่มีรหัสบางชนิด RNA ที่เจริญเต็มที่คือผลิตภัณฑ์ยีนขั้นสุดท้าย[ 23 ]ในกรณีของ messenger RNA (mRNA) RNA นี้เป็นตัวนำข้อมูลที่เข้ารหัสสำหรับการสังเคราะห์โปรตีนหนึ่งตัวหรือมากกว่า mRNA ที่มีลำดับโปรตีนเพียงลำดับเดียว (พบได้ทั่วไปในยูคาริโอต) เรียกว่าmonocistronic ในขณะที่ mRNA ที่มีลำดับโปรตีนหลายลำดับ ( พบ ได้ทั่วไปในโปรคาริโอต) เรียกว่าpolycistronic
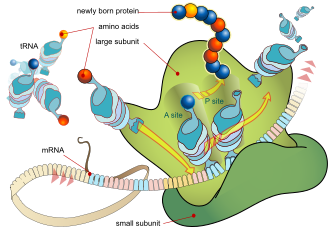
mRNA ทุกโมเลกุลประกอบด้วยสามส่วน ได้แก่ บริเวณที่ไม่ถูกแปลรหัส 5′ (5′UTR) บริเวณที่เข้ารหัสโปรตีนหรือกรอบการอ่านแบบเปิด (ORF) และบริเวณที่ไม่ถูกแปลรหัส 3′ (3′UTR) บริเวณที่เข้ารหัสจะบรรจุข้อมูลสำหรับการสังเคราะห์โปรตีนที่เข้ารหัสโดยรหัสพันธุกรรมเพื่อสร้างไตรเพล็ต แต่ละไตรเพล็ตของนิวคลีโอไทด์ในบริเวณที่เข้ารหัสเรียกว่าโคดอนและสอดคล้องกับไซต์การจับที่เสริมกับไตรเพล็ตแอนติโคดอนใน RNA ถ่ายโอน RNA ที่มีลำดับแอนติโคดอนเดียวกันจะบรรจุกรดอะมิโน ชนิดเดียวกันเสมอ จากนั้นกรดอะมิโนจะถูกเชื่อมต่อกันโดยไรโบโซมตามลำดับของไตรเพล็ตในบริเวณที่เข้ารหัส ไรโบโซมช่วยให้ RNA ถ่ายโอนจับกับ RNA สื่อสาร และนำกรดอะมิโนจาก RNA ถ่ายโอนแต่ละโมเลกุลมาสร้างเป็นโปรตีนที่ไม่มีโครงสร้าง[ 24 ] [ 25 ]โมเลกุล mRNA แต่ละโมเลกุลจะถูกแปลเป็นโมเลกุลโปรตีนจำนวนมาก โดยเฉลี่ยประมาณ 2800 โมเลกุลในสัตว์เลี้ยงลูกด้วยนม[ 26 ] [ 27 ]
ในโปรคาริโอต การแปลมักเกิดขึ้น ณ จุดของการถอดรหัส (ร่วมกับการถอดรหัส) โดยมักใช้ RNA ผู้ส่งสารที่ยังอยู่ในกระบวนการสร้าง ในยูคาริโอต การแปลสามารถเกิดขึ้นได้ในหลายบริเวณของเซลล์ ขึ้นอยู่กับว่าโปรตีนที่กำลังเขียนนั้นควรจะอยู่ที่ใด บริเวณหลักๆ คือไซโตพลาสซึมสำหรับโปรตีนไซโตพลาสซึมที่ละลายได้ และเยื่อหุ้มของเอนโดพลาสมิกเรติคูลัมสำหรับโปรตีนที่ต้องส่งออกจากเซลล์หรือแทรกเข้าไปในเยื่อหุ้ม เซลล์ โปรตีนที่ควรจะผลิตที่เอนโดพลาสมิกเรติคูลัมจะถูกจดจำในระหว่างกระบวนการแปล ซึ่งถูกควบคุมโดยอนุภาคการจดจำสัญญาณ ซึ่ง เป็นโปรตีนที่จับกับไรโบโซมและนำทางไปยังเอนโดพลาสมิกเรติคูลัมเมื่อพบเปปไทด์สัญญาณบนสายกรดอะมิโนที่กำลังเติบโต (เกิดใหม่) [ 28 ]
ระเบียบข้อบังคับ

Regulation of gene expression is the control of the amount and timing of appearance of the functional product of a gene. Control of expression is vital to allow a cell to produce the gene products it needs when it needs them; in turn, this gives cells the flexibility to adapt to a variable environment, external signals, damage to the cell, and other stimuli. More generally, gene regulation gives the cell control over all structure and function, and is the basis for cellular differentiation, morphogenesis and the versatility and adaptability of any organism.
Numerous terms are used to describe types of genes depending on how they are regulated; these include:
- A constitutive gene is a gene that is transcribed continually as opposed to a facultative gene, which is only transcribed when needed.
- A housekeeping gene is a gene that is required to maintain basic cellular function and so is typically expressed in all cell types of an organism. Examples include actin, GAPDH and ubiquitin. Some housekeeping genes are transcribed at a relatively constant rate and these genes can be used as a reference point in experiments to measure the expression rates of other genes.
- A facultative gene is a gene only transcribed when needed as opposed to a constitutive gene.
- An inducible gene is a gene whose expression is either responsive to environmental change or dependent on the position in the cell cycle.
Any step of gene expression may be modulated, from the DNA-RNA transcription step to post-translational modification of a protein. The stability of the final gene product, whether it is RNA or protein, also contributes to the expression level of the gene—an unstable product results in a low expression level. In general gene expression is regulated through changes[29] in the number and type of interactions between molecules[30] that collectively influence transcription of DNA[31] and translation of RNA.[32]
Some simple examples of where gene expression is important are:
- Control of insulin expression so it gives a signal for blood glucose regulation.
- X chromosome inactivation in female mammals to prevent an "overdose" of the genes it contains.
- ระดับการแสดงออกของไซคลิน ควบคุมการดำเนินไปตาม วงจรเซลล์ ของยูคาริโอ ต
การถอดรหัส

การควบคุมการถอดรหัสสามารถแบ่งออกเป็น 3 เส้นทางหลักที่มีอิทธิพล ได้แก่ พันธุกรรม (ปฏิสัมพันธ์โดยตรงของปัจจัยควบคุมกับยีน) การปรับเปลี่ยนปฏิสัมพันธ์ของปัจจัยควบคุมกับกลไกการถอดรหัส และเอพิเจเนติกส์ (การเปลี่ยนแปลงโครงสร้าง DNA ที่ไม่ใช่ลำดับซึ่งมีอิทธิพลต่อการถอดรหัส) [ 33 ] [ 34 ]

การโต้ตอบโดยตรงกับ DNA เป็นวิธีที่ง่ายที่สุดและตรงที่สุดที่โปรตีนจะเปลี่ยนแปลงระดับการถอดรหัส[ 35 ]ยีนมักจะมีตำแหน่งการจับโปรตีนหลายตำแหน่งรอบบริเวณการเข้ารหัสที่มีหน้าที่เฉพาะในการควบคุมการถอดรหัส[ 36 ]มีตำแหน่งการจับ DNA ที่ควบคุมหลายประเภทที่รู้จักกันในชื่อenhancers , insulators และ silencers [ 37 ] กลไกในการควบคุมการถอดรหัสมีความหลากหลาย ตั้งแต่การปิดกั้นตำแหน่งการจับที่สำคัญบน DNA สำหรับRNA polymeraseไปจนถึงการทำหน้าที่เป็นตัวกระตุ้นและส่งเสริมการถอดรหัสโดยการช่วยให้ RNA polymerase จับ[ 38 ]
กิจกรรมของปัจจัยการถอดรหัสจะถูกปรับเปลี่ยนเพิ่มเติมโดยสัญญาณภายในเซลล์ที่ทำให้เกิดการดัดแปลงโปรตีนหลังการแปล รวมถึงการฟอสโฟรีเลชันการอะเซทิเลชันหรือการไกลโคซิเลชัน [ 39 ] การเปลี่ยนแปลงเหล่านี้ส่งผลต่อความสามารถของปัจจัยการถอดรหัสในการจับกับ DNA โปรโมเตอร์โดยตรงหรือโดยอ้อม เพื่อดึงดูด RNA โพลีเมอเรส หรือเพื่อส่งเสริมการยืดตัวของโมเลกุล RNA ที่สังเคราะห์ขึ้นใหม่[ 40 ]
เยื่อหุ้มนิวเคลียสในยูคาริโอตช่วยให้สามารถควบคุมปัจจัยการถอดรหัสได้เพิ่มเติมโดยระยะเวลาที่ปัจจัยเหล่านั้นอยู่ในนิวเคลียส ซึ่งถูกควบคุมโดยการเปลี่ยนแปลงโครงสร้างที่ย้อนกลับได้และการจับกับโปรตีนอื่นๆ[ 41 ]สิ่งเร้าจากสิ่งแวดล้อมหรือสัญญาณจากต่อมไร้ท่อ[ 42 ]อาจทำให้เกิดการปรับเปลี่ยนโปรตีนควบคุม[ 43 ]กระตุ้นให้เกิดสัญญาณภายในเซลล์แบบต่อเนื่อง[ 44 ]ซึ่งส่งผลให้เกิดการควบคุมการแสดงออกของยีน
ปรากฏชัดว่ามีอิทธิพลอย่างมากของผลกระทบที่ไม่จำเพาะต่อลำดับ DNA ต่อการถอดรหัส[ 45 ]ผลกระทบเหล่านี้เรียกว่าepigeneticและเกี่ยวข้องกับโครงสร้างลำดับสูงของ DNA โปรตีนที่จับกับ DNA ที่ไม่จำเพาะต่อลำดับ และการดัดแปลงทางเคมีของ DNA [ 46 ]โดยทั่วไป ผลกระทบ epigenetic จะเปลี่ยนแปลงการเข้าถึง DNA ของโปรตีนและปรับเปลี่ยนการถอดรหัส[ 47 ]

ในยูคาริโอต โครงสร้างของโครมาตินซึ่งควบคุมโดยรหัสฮิสโตนจะควบคุมการเข้าถึง DNA ซึ่งส่งผลกระทบอย่างมากต่อการแสดงออกของยีนในบริเวณยูโครมาตินและเฮเทอโรโครมาติน[ 48 ]
ตัวเร่งปฏิกิริยา ปัจจัยการถอดรหัส คอมเพล็กซ์ตัวกลาง และลูป DNA
การแสดงออกของยีนในสัตว์เลี้ยงลูกด้วยนมถูกควบคุมโดยองค์ประกอบควบคุมแบบซิส หลายอย่าง รวมถึงโปรโมเตอร์หลักและองค์ประกอบที่อยู่ใกล้โปรโมเตอร์ซึ่งตั้งอยู่ใกล้กับตำแหน่งเริ่มต้นการ ถอดรหัส ของยีนต้นน้ำบน DNA (ไปทางบริเวณ 5' ของสายเซนส์ ) โมดูลควบคุมแบบซิสที่สำคัญอื่นๆ จะอยู่ในบริเวณ DNA ที่อยู่ห่างจากตำแหน่งเริ่มต้นการถอดรหัส ซึ่งรวมถึงตัวเร่งตัวยับยั้งตัวกั้นและองค์ประกอบเชื่อมโยง[ 49 ] ตัวเร่งและ ปัจจัยการถอดรหัสที่เกี่ยวข้องมีบทบาทสำคัญในการควบคุมการแสดงออกของยีน[ 50 ]
เอนแฮนเซอร์คือบริเวณจีโนมที่ควบคุมยีน เอนแฮนเซอร์ควบคุมโปรแกรมการแสดงออกของยีนเฉพาะเซลล์ โดยส่วนใหญ่มักจะวนผ่านระยะทางไกลๆ เพื่อเข้าใกล้โปรโมเตอร์ของยีนเป้าหมาย[ 51 ]เอนแฮนเซอร์หลายตัว ซึ่งแต่ละตัวมักจะอยู่ห่างจากยีนเป้าหมายหลายหมื่นหรือหลายแสนนิวคลีโอไทด์ จะวนไปยังโปรโมเตอร์ของยีนเป้าหมายและประสานงานกันเพื่อควบคุมการแสดงออกของยีน[ 51 ]
The illustration shows an enhancer looping around to come into proximity with the promoter of a target gene. The loop is stabilized by a dimer of a connector protein (e.g. dimer of CTCF or YY1). One member of the dimer is anchored to its binding motif on the enhancer and the other member is anchored to its binding motif on the promoter (represented by the red zigzags in the illustration).[52] Several cell function-specific transcription factors (among the about 1,600 transcription factors in a human cell)[53] generally bind to specific motifs on an enhancer.[54] A small combination of these enhancer-bound transcription factors, when brought close to a promoter by a DNA loop, govern transcription level of the target gene. Mediator (a complex usually consisting of about 26 proteins in an interacting structure) communicates regulatory signals from enhancer DNA-bound transcription factors directly to the RNA polymerase II (pol II) enzyme bound to the promoter.[55]
Enhancers, when active, are generally transcribed from both strands of DNA with RNA polymerases acting in two different directions, producing two eRNAs as illustrated in the figure.[56] An inactive enhancer may be bound by an inactive transcription factor. Phosphorylation of the transcription factor may activate it and that activated transcription factor may then activate the enhancer to which it is bound (see small red star representing phosphorylation of transcription factor bound to enhancer in the illustration).[57] An activated enhancer begins transcription of its RNA before activating transcription of messenger RNA from its target gene.[58]
DNA methylation and demethylation

การเมทิลเลชันของ DNAเป็นกลไกที่แพร่หลายในการมีอิทธิพลทางเอพิเจเนติกต่อการแสดงออกของยีน และพบได้ในแบคทีเรียและยูคาริโอตและมีบทบาทในการยับยั้งการถอดรหัสที่ถ่ายทอดได้และการควบคุมการถอดรหัส การเมทิลเลชันมักเกิดขึ้นบนไซโตซีน (ดูรูป) การเมทิลเลชันของไซโตซีนส่วนใหญ่เกิดขึ้นในลำดับไดนิวคลีโอไทด์ที่ไซโตซีนตามด้วยกัวนีน ซึ่งเป็นไซต์ CpGจำนวนไซต์ CpGในจีโนมของมนุษย์มีประมาณ 28 ล้านไซต์[ 59 ]ขึ้นอยู่กับชนิดของเซลล์ ประมาณ 70% ของไซต์ CpG มีไซโตซีนที่ถูกเมทิลเลชัน[ 60 ]
การเติมหมู่เมทิลลงบนไซโตซีนในดีเอ็นเอมีบทบาทสำคัญในการควบคุมการแสดงออกของยีน การเติมหมู่เมทิลลงบน CpG ในบริเวณโปรโมเตอร์ของยีนมักจะยับยั้งการถอดรหัสของยีน[ 61 ]ในขณะที่การเติมหมู่เมทิลลงบน CpG ในส่วนลำตัวของยีนจะเพิ่มการแสดงออก[ 62 ]เอนไซม์ TETมีบทบาทสำคัญในการกำจัดหมู่เมทิลออกจากไซโตซีนที่ถูกเติมหมู่เมทิล การกำจัดหมู่เมทิลลงบน CpG ในโปรโมเตอร์ของยีนโดย กิจกรรม ของเอนไซม์ TETจะเพิ่มการถอดรหัสของยีน[ 63 ]
การควบคุมหลังการถอดรหัส
ในยูคาริโอต ซึ่งการส่งออก RNA เป็นสิ่งจำเป็นก่อนที่จะสามารถแปลความหมายได้ การส่งออกนิวเคลียสถือเป็นการควบคุมการแสดงออกของยีนเพิ่มเติม การขนส่งทั้งหมดเข้าและออกจากนิวเคลียสเกิดขึ้นผ่านรูพรุนนิวเคลียส และการขนส่งถูกควบคุมโดย โปรตีนอิมพอร์ทินและเอ็กซ์พอร์ทินหลากหลายชนิด[ 64 ]
การแสดงออกของยีนที่เข้ารหัสโปรตีนจะเกิดขึ้นได้ก็ต่อเมื่อ mRNA ที่บรรจุรหัสนั้นมีชีวิตรอดได้นานพอที่จะถูกแปล[ 65 ]ในเซลล์ทั่วไป โมเลกุล RNA จะเสถียรได้ก็ต่อเมื่อได้รับการปกป้องจากการย่อยสลายโดยเฉพาะ[ 66 ]การย่อยสลาย RNA มีความสำคัญอย่างยิ่งในการควบคุมการแสดงออกในเซลล์ยูคาริโอต ซึ่ง mRNA ต้องเดินทางเป็นระยะทางไกลก่อนที่จะถูกแปล[ 67 ]ในยูคาริโอต RNA จะมีความเสถียรมากขึ้นด้วยการดัดแปลงหลังการถอดรหัสบางอย่าง โดยเฉพาะอย่างยิ่งหมวก 5′และหางโพลีอะดีนิเลต[ 9 ]
การย่อยสลาย mRNA โดยเจตนาไม่ได้ถูกใช้เป็นเพียงกลไกป้องกัน RNA จากภายนอก (โดยปกติมาจากไวรัส) แต่ยังใช้เป็นเส้นทางในการทำให้ mRNA ไม่ เสถียรอีกด้วย [ 68 ]หากโมเลกุล mRNA มีลำดับที่เสริมกับRNA รบกวนขนาดเล็กก็จะถูกกำหนดเป้าหมายให้ถูกทำลายผ่านทางเส้นทางการรบกวน RNA [ 69 ]
สามบริเวณหลักที่ไม่ได้รับการแปลและไมโครอาร์เอ็นเอ
บริเวณ 3′ -UTR (3′-UTR) ของเมสเซนเจอร์อาร์เอ็นเอ (mRNA) มักมีลำดับควบคุมที่ส่งผลต่อการแสดงออกของยีนหลังการถอดรหัส บริเวณ 3′-UTR เหล่านี้มักมีทั้งตำแหน่งการจับของไมโครอาร์เอ็นเอ (miRNA) และโปรตีนควบคุม[ 70 ]โดยการจับกับตำแหน่งเฉพาะภายใน 3′-UTR miRNA สามารถลดการแสดงออกของยีนของ mRNA ต่างๆ ได้โดยการยับยั้งการแปลหรือทำให้เกิดการย่อยสลายของทรานสคริปต์โดยตรง[ 71 ]นอกจากนี้ 3′-UTR อาจมีบริเวณตัวยับยั้งที่จับกับโปรตีนตัวยับยั้งที่ยับยั้งการแสดงออกของ mRNA [ 72 ]
3′-UTR มักมีองค์ประกอบตอบสนองไมโครอาร์เอ็นเอ (MREs) MREs คือลำดับที่ไมโครอาร์เอ็นเอจับกับ ซึ่งเป็นโมทีฟที่พบได้ทั่วไปใน 3′-UTRs ในบรรดาโมทีฟควบคุมทั้งหมดใน 3′-UTRs (เช่น รวมถึงบริเวณตัวยับยั้ง) MREs คิดเป็นประมาณครึ่งหนึ่งของโมทีฟทั้งหมด[ 73 ]
ณ ปี 2014 เว็บไซต์miRBase [ 74 ]ซึ่งเป็นคลังข้อมูลลำดับ และคำอธิบายประกอบของ miRNA ได้แสดงรายการ 28,645 รายการใน 233 สปีชีส์ทางชีวภาพ ในจำนวนนี้ miRNA จำนวน 1,881 รายการอยู่ในตำแหน่ง miRNA ของมนุษย์ที่ได้รับการอธิบายไว้ มีการคาดการณ์ว่า miRNA จะมีmRNA เป้าหมายโดยเฉลี่ยประมาณสี่ร้อยรายการ (ส่งผลต่อการแสดงออกของยีนหลายร้อยยีน) [ 75 ] Friedman et al. [ 75 ]ประมาณการว่าตำแหน่งเป้าหมายของ miRNA มากกว่า 45,000 ตำแหน่งภายใน 3′UTR ของ mRNA ของมนุษย์ได้รับการอนุรักษ์ไว้เหนือระดับพื้นหลัง และยีนที่เข้ารหัสโปรตีนของมนุษย์มากกว่า 60% อยู่ภายใต้แรงกดดันในการคัดเลือกเพื่อรักษาการจับคู่กับ miRNA
การทดลองโดยตรงแสดงให้เห็นว่า miRNA เพียงตัวเดียวสามารถลดความเสถียรของ mRNA ที่ไม่ซ้ำกันหลายร้อยตัวได้[ 76 ]การทดลองอื่นๆ แสดงให้เห็นว่า miRNA เพียงตัวเดียวอาจยับยั้งการผลิตโปรตีนหลายร้อยตัว แต่การยับยั้งนี้มักจะค่อนข้างอ่อน (น้อยกว่า 2 เท่า) [ 77 ] [ 78 ]
ผลกระทบของการควบคุมการแสดงออกของยีนโดย miRNA ที่ผิดปกติดูเหมือนจะมีความสำคัญในมะเร็ง[ 79 ]ตัวอย่างเช่น ในมะเร็งทางเดินอาหาร มีการระบุ miRNA จำนวน 9 ชนิดที่ มีการเปลี่ยนแปลง ทางเอพิเจเนติกส์และมีประสิทธิภาพในการลดการทำงานของเอนไซม์ซ่อมแซม DNA [ 80 ]
ผลกระทบของการควบคุมการแสดงออกของยีนที่ผิดปกติโดย miRNA ดูเหมือนจะมีความสำคัญในความผิดปกติทางจิตเวช เช่น โรคจิตเภท โรคอารมณ์สองขั้ว โรคซึมเศร้า โรคพาร์กินสัน โรคอัลไซเมอร์ และความผิดปกติในกลุ่มอาการออทิสติก[ 81 ] [ 82 ]
การแปล
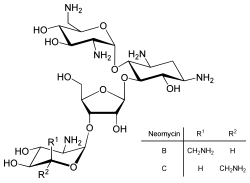
การควบคุมการแปลโดยตรงนั้นพบได้น้อยกว่าการควบคุมการถอดรหัสหรือความเสถียรของ mRNA แต่ก็มีการใช้บ้างเป็นครั้งคราว[ 83 ]การยับยั้งการแปลโปรตีนเป็นเป้าหมายหลักของสารพิษและยาปฏิชีวนะเพื่อให้สามารถฆ่าเซลล์ได้โดยการขัดขวางการควบคุมการแสดงออกของยีนตามปกติ[ 84 ]สารยับยั้งการสังเคราะห์โปรตีนได้แก่ ยาปฏิชีวนะนีโอไมซินและสารพิษริซิน[ 85 ]
การดัดแปลงหลังการแปล
การดัดแปลงหลังการแปล (PTMs) คือ การดัดแปลง โควาเลนต์ของโปรตีน เช่นเดียวกับการตัดต่อ RNA ซึ่งช่วยเพิ่มความหลากหลายของโปรตีโอมอย่างมีนัยสำคัญ การดัดแปลงเหล่านี้มักถูกเร่งปฏิกิริยาโดยเอนไซม์ นอกจากนี้ กระบวนการต่างๆ เช่น การเติมโควาเลนต์ไปยังหมู่ข้างเคียงของกรดอะมิโน มักจะสามารถย้อนกลับได้ด้วยเอนไซม์อื่นๆ อย่างไรก็ตาม บางกระบวนการ เช่นการตัดโครงสร้างหลักของโปรตีนด้วยโปรตีโอไลซิส จะไม่สามารถย้อนกลับได้[ 86 ]
PTMs play many important roles in the cell.[87] For example, phosphorylation is primarily involved in activating and deactivating proteins and in signaling pathways.[88] PTMs are involved in transcriptional regulation: an important function of acetylation and methylation is histone tail modification, which alters how accessible DNA is for transcription.[86] They can also be seen in the immune system, where glycosylation plays a key role.[89] One type of PTM can initiate another type of PTM, as can be seen in how ubiquitination tags proteins for degradation through proteolysis.[86] Proteolysis, other than being involved in breaking down proteins, is also important in activating and deactivating them, and in regulating biological processes such as DNA transcription and cell death.[90]
Measurement

Measuring gene expression is an important part of many life sciences, as the ability to quantify the level at which a particular gene is expressed within a cell, tissue or organism can provide a lot of valuable information. For example, measuring gene expression can:
- Identify viral infection of a cell (viral protein expression).
- Determine an individual's susceptibility to cancer (oncogene expression).
- Find if a bacterium is resistant to penicillin (beta-lactamase expression).
- Gene expression profiling evaluates a panel of genes to help understand the fundamental mechanism of a cell. This is increasingly used in cancer therapy to target specific chemotherapy. (SeeRNA-Seq and DNA microarray for details.)
Similarly, the analysis of the location of protein expression is a powerful tool, and this can be done on an organismal or cellular scale. Investigation of localization is particularly important for the study of development in multicellular organisms and as an indicator of protein function in single cells. Ideally, measurement of expression is done by detecting the final gene product (for many genes, this is the protein); however, it is often easier to detect one of the precursors, typically mRNA and to infer gene-expression levels from these measurements.
mRNA quantification
Levels of mRNA can be quantitatively measured by northern blotting, which provides size and sequence information about the mRNA molecules.[91] A sample of RNA is separated on an agarose gel and hybridized to a radioactively labeled RNA probe that is complementary to the target sequence.[92] The radiolabeled RNA is then detected by an autoradiograph.[93] Because the use of radioactive reagents makes the procedure time-consuming and potentially dangerous, alternative labeling and detection methods, such as digoxigenin and biotin chemistries, have been developed.[94] Perceived disadvantages of Northern blotting are that large quantities of RNA are required and that quantification may not be completely accurate, as it involves measuring band strength in an image of a gel.[95] On the other hand, the additional mRNA size information from the Northern blot allows the discrimination of alternately spliced transcripts.[96][97]
อีกแนวทางหนึ่งสำหรับการวัดปริมาณ mRNA คือ RT-qPCR ในเทคนิคนี้การถอดรหัสย้อนกลับจะตามด้วยPCR เชิงปริมาณการถอดรหัสย้อนกลับจะสร้างแม่แบบ DNA จาก mRNA ก่อน แม่แบบสายเดี่ยวนี้เรียกว่าcDNAจากนั้นแม่แบบ cDNA จะถูกขยายในขั้นตอนเชิงปริมาณ ซึ่งในระหว่างนั้นฟลูออเรสเซนซ์ที่ปล่อยออกมาจาก โพรบ ไฮบริดไดเซชัน ที่มีฉลาก หรือสีย้อมแทรกจะเปลี่ยนแปลงไปตามกระบวนการขยาย DNA [ 98 ]ด้วยเส้นโค้งมาตรฐานที่สร้างขึ้นอย่างระมัดระวัง qPCR สามารถให้การวัดที่แน่นอนของจำนวนสำเนาของ mRNA ดั้งเดิม โดยทั่วไปในหน่วยของสำเนาต่อนาโนลิตรของเนื้อเยื่อที่เป็นเนื้อเดียวกันหรือสำเนาต่อเซลล์[ 99 ] qPCR มีความไวสูงมาก (การตรวจจับโมเลกุล mRNA เพียงโมเลกุลเดียวเป็นไปได้ในทางทฤษฎี) แต่อาจมีราคาแพงขึ้นอยู่กับประเภทของตัวรายงานที่ใช้ โพรบโอลิโกนิวคลีโอไทด์ที่มีฉลากฟลูออเรสเซนซ์มีราคาแพงกว่าสีย้อมฟลูออเรสเซนซ์แทรกที่ไม่จำเพาะ[ 100 ]
สำหรับการสร้างโปรไฟล์การแสดงออกหรือการวิเคราะห์ยีนจำนวนมากในตัวอย่างเดียวด้วยความเร็วสูงอาจทำการPCR เชิงปริมาณ สำหรับยีนหลายร้อยยีนพร้อมกันในกรณีของอาร์เรย์ความหนาแน่นต่ำ [ 101 ]แนวทางที่สองคือไมโครอาร์เรย์แบบไฮบริดไดเซชันอาร์เรย์หรือ "ชิป" เดียวอาจมีโพรบเพื่อกำหนดระดับการถอดรหัสสำหรับยีนที่รู้จักทุกยีนในจีโนมของสิ่งมีชีวิตหนึ่งตัวหรือมากกว่า[ 102 ]หรืออาจใช้ เทคโนโลยี "แบบใช้แท็ก" เช่น การวิเคราะห์การแสดงออกของยีนแบบอนุกรม (SAGE) และRNA-Seqซึ่งสามารถให้การวัดความเข้มข้น ของ mRNA ที่แตกต่างกันในเซลล์ได้ [ 103 ]ข้อดีของวิธีการแบบใช้แท็กคือ "สถาปัตยกรรมแบบเปิด" ทำให้สามารถวัดการถอดรหัสใดๆ ได้อย่างแม่นยำ ไม่ว่าจะมีลำดับที่ทราบหรือไม่ทราบก็ตาม[ 104 ]การจัดลำดับรุ่นต่อไป (NGS) เช่นRNA-Seqเป็นอีกแนวทางหนึ่งที่สร้างข้อมูลลำดับจำนวนมหาศาลที่สามารถจับคู่กับจีโนมอ้างอิงได้ แม้ว่า NGS จะใช้เวลานาน มีราคาแพง และต้องใช้ทรัพยากรมาก แต่ก็สามารถระบุโพลีมอร์ฟิซึมของนิวคลีโอไทด์เดี่ยวตัวแปรการเชื่อมต่อ และยีนใหม่ได้ และยังสามารถใช้เพื่อสร้างโปรไฟล์การแสดงออกในสิ่งมีชีวิตที่มีข้อมูลลำดับน้อยหรือไม่มีเลย[ 105 ]
การหาปริมาณโปรตีน
สำหรับยีนที่เข้ารหัสโปรตีน ระดับการแสดงออกสามารถประเมินได้โดยตรงด้วยวิธีการหลายวิธี ซึ่งมีความคล้ายคลึงอย่างชัดเจนกับเทคนิคการหาปริมาณ mRNA
หนึ่งในวิธีการที่ใช้กันทั่วไปคือการทำเวสเทิร์นบลอตกับโปรตีนที่สนใจ[ 106 ]วิธีนี้ให้ข้อมูลเกี่ยวกับขนาดของโปรตีนนอกเหนือจากเอกลักษณ์ของโปรตีน ตัวอย่าง (มักจะ เป็น ไลเซต ของเซลล์ ) จะถูกแยกบนเจลโพลีอะคริลาไมด์ถ่ายโอนไปยังเมมเบรน แล้วตรวจสอบด้วยแอนติบอดีต่อโปรตีนที่สนใจ แอนติบอดีสามารถเชื่อมต่อกับฟลูออโรฟอร์หรือฮอร์สแรดิชเปอร์ออกซิเดสสำหรับการถ่ายภาพและ/หรือการหาปริมาณ ลักษณะการทดสอบแบบใช้เจลนี้ทำให้การหาปริมาณมีความแม่นยำน้อยลง แต่มีข้อดีคือสามารถระบุการดัดแปลงโปรตีนในภายหลังได้ เช่น การย่อยสลายโปรตีนหรือการยูบิควิตินเนชัน จากการเปลี่ยนแปลงขนาด
ความสัมพันธ์ระหว่าง mRNA และโปรตีน
แม้ว่าการถอดรหัสจะสะท้อนการแสดงออกของยีนโดยตรง แต่จำนวนสำเนาของโมเลกุล mRNA ไม่ได้มีความสัมพันธ์โดยตรงกับจำนวนโมเลกุลโปรตีนที่แปลจาก mRNA การหาปริมาณทั้งโปรตีนและ mRNA ช่วยให้สามารถเชื่อมโยงทั้งสองระดับได้ การควบคุมในแต่ละขั้นตอนของการแสดงออกของยีนสามารถส่งผลกระทบต่อความสัมพันธ์ได้ ดังที่แสดงให้เห็นในการควบคุมการแปล[ 27 ]หรือความเสถียรของโปรตีน[ 107 ]ปัจจัยหลังการแปล เช่น การขนส่งโปรตีนในเซลล์ที่มีขั้วสูง[ 108 ]ก็สามารถส่งผลต่อความสัมพันธ์ระหว่าง mRNA กับโปรตีนที่วัดได้เช่นกัน
การแปลเป็นภาษาท้องถิ่น

การวิเคราะห์การแสดงออกไม่ได้จำกัดอยู่เพียงแค่การหาปริมาณเท่านั้น แต่ยังสามารถระบุตำแหน่งได้ด้วย สามารถตรวจจับ mRNA ได้โดยใช้สาย mRNA เสริมที่มีการติดฉลากอย่างเหมาะสม และสามารถตรวจจับโปรตีนได้โดยใช้แอนติบอดีที่มีการติดฉลาก จากนั้นจึงนำตัวอย่างที่ตรวจสอบแล้วไปสังเกตด้วยกล้องจุลทรรศน์เพื่อระบุตำแหน่งของ mRNA หรือโปรตีนนั้น

โดยการแทนที่ยีนด้วยยีนเวอร์ชันใหม่ที่เชื่อมต่อกับโปรตีนเรืองแสงสีเขียวหรือสารที่คล้ายกัน การแสดงออกของยีนสามารถวัดได้โดยตรงในเซลล์ที่มีชีวิต วิธีนี้ทำได้โดยการถ่ายภาพโดยใช้กล้องจุลทรรศน์เรืองแสง การโคลนโปรตีนที่เชื่อมต่อกับ GFP เข้าไปในตำแหน่งดั้งเดิมในจีโนมโดยไม่ส่งผลกระทบต่อระดับการแสดงออกนั้นทำได้ยากมาก ดังนั้นวิธีนี้จึงมักไม่สามารถใช้ในการวัดการแสดงออกของยีนภายในเซลล์ได้ อย่างไรก็ตาม วิธีนี้ถูกนำมาใช้กันอย่างแพร่หลายในการวัดการแสดงออกของยีนที่ถูกนำเข้าไปในเซลล์โดยวิธีการประดิษฐ์ เช่น ผ่านเวกเตอร์การแสดงออกโดยการเชื่อมต่อโปรตีนเป้าหมายกับตัวรายงานเรืองแสง พฤติกรรมของโปรตีน รวมถึงตำแหน่งในเซลล์และระดับการแสดงออก สามารถเปลี่ยนแปลงได้อย่างมีนัยสำคัญ
การทดสอบอิมมูโนซอร์เบนต์แบบเชื่อมโยงเอนไซม์ (ELISA)ทำงานโดยใช้แอนติบอดีที่ตรึงอยู่บนแผ่นไมโครไทเตอร์เพื่อจับโปรตีนที่สนใจจากตัวอย่างที่เติมลงในหลุม โดยใช้แอนติบอดีตรวจจับที่เชื่อมต่อกับเอนไซม์หรือฟลูออโรฟอร์ ปริมาณโปรตีนที่จับได้สามารถวัดได้อย่างแม่นยำด้วย การตรวจ จับแบบฟลูออเรสเซนต์หรือ แบบ คัลเลอริเมตริกกระบวนการตรวจจับคล้ายคลึงกับเวสเทิร์นบลอตมาก แต่เนื่องจากไม่ต้องใช้ขั้นตอนเจล จึงสามารถวัดปริมาณได้อย่างแม่นยำยิ่งขึ้น
ดูเพิ่มเติม
- การทดสอบการแสดงออกของโมเลกุล AlloMap
- การคั่นหน้า
- อีพีซีเควกซ์
- แท็กลำดับที่แสดงออก
- แอตลาสการแสดงออก
- การวิเคราะห์การแสดงออกของยีน
- โครงสร้างยีน
- วิศวกรรมพันธุกรรม
- สิ่งมีชีวิตดัดแปลงพันธุกรรม
- รายชื่อฐานข้อมูลทางชีววิทยา
- รายชื่อยีนของมนุษย์
- ยีนที่สั่นไหว
- พารามิวเทชัน
- การผลิตโปรตีน
- การทำให้โปรตีนบริสุทธิ์
- ริโบโนมิกส์
- สันเขา
- เครื่องมือวิเคราะห์ลำดับดีเอ็นเอ
- การแตกตัวของการถอดรหัส
- สัญญาณรบกวนการถอดรหัส
- บันทึกการถอดเสียงของฟังก์ชันที่ไม่ทราบแน่ชัด
ลิงก์ภายนอก
- ฐานข้อมูลปัจจัยการถอดรหัสทางพันธุกรรมของพืช และแพลตฟอร์มข้อมูลและการวิเคราะห์การควบคุมการถอดรหัสทางพันธุกรรมของพืช
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การแสดงออกของยีน
การแสดงออกของยีนคือกระบวนการที่ข้อมูลที่มีอยู่ในยีนถูกนำมาใช้เพื่อสร้างผลิตภัณฑ์ของยีนที่มีฟังก์ชันการทำงาน เช่นโปรตีนหรือ โมเลกุล RNA ที่มีฟังก์ชันการทำงาน...
การถอดเสียง
การสร้างสำเนา RNA จากสาย DNA เรียกว่า การถอดรหัส (transcription ) และดำเนินการโดย RNA polymerase ซึ่งจะเพิ่มไร โบนิวคลีโอไทด์ ทีละตัวลงในสาย RNA ที่กำลังเติบโตตาม กฎ ความเสริมกัน ของเบสนิวคลีโอไทด์ RNA นี้จะ เสริม กับสาย DNA แม่แบบ 3′ → 5′ [ 2 ] ยกเว้นว่า...
mRNA processing
While transcription of prokaryotic protein-coding genes creates messenger RNA (mRNA) that is ready for translation into protein, transcription of eukaryotic genes leaves a primary transcript of RNA (pre-RNA), which first has to undergo a series of...
การเจริญเติบโตของ RNA ที่ไม่เข้ารหัส
ในสิ่งมีชีวิตส่วนใหญ่ ยีนที่ไม่เข้ารหัส (ncRNA) จะถูกถอดรหัสเป็นสารตั้งต้นที่ต้องผ่านกระบวนการเพิ่มเติม ในกรณีของไรโบโซมอลอาร์เอ็นเอ (rRNA) มักจะถูกถอดรหัสเป็นพรี-rRNA ซึ่งประกอบด้วย rRNA หนึ่งตัวหรือมากกว่านั้น พรี-rRNA จะถูกตัดและดัดแปลง ( การเติมหมู่...