อ่าน 86 นาที
วิวัฒนาการของมนุษย์
โฮโมเซเปียนส์เป็นสายพันธุ์ที่แตกต่างกันของ วงศ์ โฮมินิดในกลุ่มไพรเมตซึ่งรวมถึงลิงใหญ่ทั้งหมดตลอดประวัติศาสตร์วิวัฒนาการมนุษย์ค่อยๆ พัฒนาลักษณะต่างๆ...
วิวัฒนาการของมนุษย์

โฮโมเซเปียนส์เป็นสายพันธุ์ที่แตกต่างกันของ วงศ์ โฮมินิดในกลุ่มไพรเมตซึ่งรวมถึงลิงใหญ่ทั้งหมด[ 2 ]ตลอดประวัติศาสตร์วิวัฒนาการมนุษย์ค่อยๆ พัฒนาลักษณะต่างๆ เช่นการเดินสองขาความคล่องแคล่วและภาษาที่ซับซ้อน [ 3 ] มนุษย์ยุคใหม่ผสมพันธุ์กับมนุษย์ยุคโบราณ [ 4 ]ซึ่งบ่งชี้ว่าวิวัฒนาการของพวกเขาไม่ได้เป็นเส้นตรง แต่เป็นแบบใยแมงมุม[ 5 ] [ 6 ] [ 7 ] [ 8 ]การศึกษาต้นกำเนิดของมนุษย์เกี่ยวข้องกับหลายสาขาวิทยาศาสตร์ รวมถึงมานุษยวิทยาทางกายภาพและ วิวัฒนาการ บรรพชีวินวิทยาและพันธุศาสตร์สาขานี้ยังเป็นที่รู้จักในชื่อ anthropogeny , anthropogenesisและ anthropogony [ 9 ] [ 10 ]โดยสองคำหลังบางครั้งใช้เพื่ออ้างถึงหัวข้อที่เกี่ยวข้องกับ hominization
Primates diverged from other mammals about 85 million years ago, in the Late Cretaceous period, with their earliest fossils appearing over 55 mya, during the Paleocene.[11] Primates produced successive clades leading to the ape superfamily, which gave rise to the hominid and the gibbon families; these diverged some 15–20 mya. African and Asian hominids (including orangutans) diverged about 14 mya. Hominins (including the Australopithecine and Panina subtribes) parted from the Gorillini tribe between 8 and 9 mya; Australopithecine (including the extinct biped ancestors of humans) separated from the Pan genus (containing chimpanzees and bonobos) 4–7 mya.[12] The Homo genus is evidenced by the appearance of H. habilis over 2 mya,[a] while anatomically modern humans emerged in Africa approximately 300,000 years ago.
Before Homo
Early evolution of primates
The evolutionary history of primates can be traced back 65 million years.[13][14][15][16][17] The details of the origins and early evolution of primates are however still unknown to a large extent due to scarcity of fossil evidence. One of the oldest known primate-like mammal species, the Plesiadapis, came from North America;[18][19][20][21][22][23] another, Archicebus, came from China.[24] Other such early primates include Altiatlasius and Algeripithecus, which were found in Northern Africa.[25][26] Other similar basal primates were widespread in Eurasia and Africa during the tropical conditions of the Paleocene and Eocene.

David R. Begun ตั้งทฤษฎีว่าไพรเมตยุคแรกเจริญรุ่งเรืองในยูเรเซียก่อน และสายพันธุ์ที่นำไปสู่ลิงแอฟริกันและมนุษย์ รวมถึงDryopithecusได้อพยพลงใต้จากยุโรปหรือเอเชียตะวันตกไปยังแอฟริกา[ 27 ]อย่างไรก็ตาม บรรพบุรุษที่เป็นไปได้ของไพรเมตในยุคแรก เช่นAltiatlasius เป็นที่ทราบกัน ว่าอาศัยอยู่ในทวีปแอฟริกามานานกว่าDryopithecus มาก [ 25 ]
ประชากรไพรเมตเขตร้อนที่ยังคงหลงเหลืออยู่ ซึ่งพบเห็นได้อย่างสมบูรณ์ที่สุดในชั้น ฟอสซิลยุคอีโอซีนตอนบนและ โอลิโกซีน ตอนล่างสุดของแอ่ง ฟาอียุมทางตะวันตกเฉียงใต้ของกรุงไคโรเป็นต้นกำเนิดของไพรเมตทุกสายพันธุ์ที่ยังมีชีวิตอยู่ในปัจจุบัน รวมถึงลีเมอร์แห่งมาดากัสการ์ลอริสแห่งเอเชียตะวันออกเฉียงใต้กาลาโกหรือ "บุชเบบี้" แห่งแอฟริกา และแอนโทรปอยด์ซึ่งได้แก่พลาทิไรน์หรือลิงโลกใหม่ คาทาร์ไรน์หรือลิงโลกเก่า และลิงใหญ่ รวมถึงมนุษย์และโฮมินิดอื่นๆ
แคทาร์ไรน์ที่เก่าแก่ที่สุดเท่าที่รู้จักคือKamoyapithecusจากยุคโอลิโกซีนตอนบนสุดที่ Eragaleit ในหุบเขาเกรตริฟต์ ตอนเหนือ ในเคนยา มีอายุราว 24 ล้านปีก่อน[ 28 ]เชื่อกันว่าบรรพบุรุษของมันคือสายพันธุ์ที่เกี่ยวข้องกับAegyptopithecus , PropliopithecusและParapithecusจาก Faiyum เมื่อประมาณ 35 ล้านปีก่อน[ 29 ]ในปี 2010 Saadaniusได้รับการอธิบายว่าเป็นญาติใกล้ชิดของบรรพบุรุษร่วมสุดท้ายของ แคทาร์ไรน์ กลุ่มมงกุฎและมีอายุโดยประมาณ 29–28 ล้านปีก่อน ซึ่งช่วยเติมเต็มช่องว่าง 11 ล้านปีในบันทึกฟอสซิล[ 30 ]

ในช่วงต้นสมัยไมโอซีน เมื่อประมาณ 22 ล้านปีก่อน ลิงคาตาร์ไรน์ดั้งเดิมหลายชนิด ที่ปรับตัวให้เข้า กับ การอาศัยอยู่บนต้นไม้จากแอฟริกาตะวันออก บ่งชี้ถึงประวัติศาสตร์อันยาวนานของการวิวัฒนาการและกระจายตัวมาก่อน ฟอสซิลเมื่อ 20 ล้านปีก่อน รวมถึงชิ้นส่วนที่เชื่อว่าเป็นของVictoriapithecusซึ่งเป็นลิงโลกเก่าที่เก่าแก่ที่สุด ในบรรดาสกุลที่เชื่อว่าอยู่ใน สายพันธุ์ ลิงใหญ่เมื่อประมาณ 13 ล้านปีก่อน ได้แก่Proconsul , Rangwapithecus , Dendropithecus , Limnopithecus , Nacholapithecus , Equatorius , Nyanzapithecus , Afropithecus , HeliopithecusและKenyapithecusซึ่งทั้งหมดมาจากแอฟริกาตะวันออก
การค้นพบซากดึกดำบรรพ์ของ โฮมินอยด์ที่ไม่ใช่กลุ่มเซอร์โคพิเทซิดทั่วไปใน ยุคไมโอซีนตอนกลางจากแหล่งที่อยู่ห่างไกลกัน เช่นโอตาวิพิเทคัสจากแหล่งสะสมในถ้ำในนามิเบีย และเพียโรลาพิเทคัสและดรายโอพิเทคัสจากฝรั่งเศส สเปน และออสเตรีย เป็นหลักฐานแสดงให้เห็นถึงความหลากหลายของรูปแบบต่างๆ ทั่วทวีปแอฟริกาและลุ่มน้ำเมดิเตอร์เรเนียนในช่วงสภาพภูมิอากาศที่ค่อนข้างอบอุ่นและคงที่ของยุคไมโอซีนตอนต้นและตอนกลางโอรีโอพิเท คัส ซึ่ง เป็นโฮมินอยด์ที่ อายุน้อยที่สุดใน ยุคไมโอซีนพบในแหล่งถ่านหินในอิตาลีซึ่งมีอายุประมาณ 9 ล้านปี
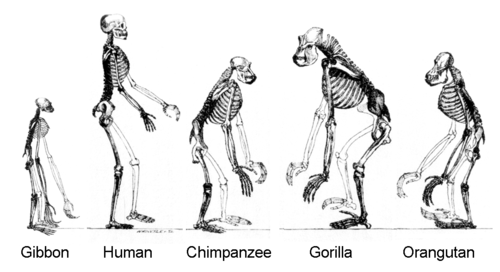
หลักฐานทางโมเลกุลบ่งชี้ว่าสายพันธุ์ของชะนีแยกตัวออกจากสายพันธุ์ของลิงใหญ่เมื่อประมาณ 18–12 ล้านปีก่อน และสายพันธุ์ของอุรังอุตัง (วงศ์ย่อยPonginae ) [ b ]แยกตัวออกจากลิงใหญ่อื่นๆ เมื่อประมาณ 12 ล้านปีก่อน ไม่มีฟอสซิลใดที่บันทึกบรรพบุรุษของชะนีได้อย่างชัดเจน ซึ่งอาจมีต้นกำเนิดมาจากประชากรโฮมินอยด์ในเอเชียตะวันออกเฉียงใต้ที่ยังไม่เป็นที่รู้จัก แต่ฟอสซิลของอุรังอุตังยุคแรกอาจแสดงโดยSivapithecusจากอินเดียและGriphopithecusจากตุรกี ซึ่งมีอายุประมาณ 10 ล้านปีก่อน[ 31 ]
วงศ์ย่อย Hominidae วงศ์ย่อยHomininae (โฮมินิดแอฟริกา) แยกตัวออกจาก Ponginae (อุรังอุตัง) เมื่อประมาณ 14 ล้านปีก่อน โฮมินิน (รวมถึงมนุษย์และเผ่าย่อย Australopithecine และPanina ) แยกตัวออกจาก เผ่า Gorillini (กอริลลา) ระหว่าง 8 ถึง 9 ล้านปีก่อน Australopithecine (รวมถึงบรรพบุรุษสองขาที่สูญพันธุ์ไปแล้วของมนุษย์) แยกตัวออกจาก สกุล Pan (ซึ่งประกอบด้วยชิมแปนซีและโบโนโบ) 4–7 ล้านปีก่อน[ 12 ]สกุลHomoมีหลักฐานจากการปรากฏตัวของH. habilisเมื่อกว่า 2 ล้านปี ก่อน [ a ] ในขณะที่มนุษย์ยุคใหม่ที่มีลักษณะทางกายวิภาคเหมือนปัจจุบันถือกำเนิดขึ้นในแอฟริกาเมื่อประมาณ 300,000 ปีก่อน
การแยกสายวิวัฒนาการของมนุษย์จากลิงใหญ่ชนิดอื่นๆ

ฟอสซิลของ Nakalipithecusที่พบในเคนยาอาจเป็นตัวอย่างของสายพันธุ์ที่ใกล้เคียงกับบรรพบุรุษร่วมสุดท้ายของกอริลลา ชิมแปนซี และมนุษย์ หลักฐานทางโมเลกุลชี้ให้เห็นว่าระหว่าง 8 ถึง 4 ล้านปีก่อน กอริลลาและชิมแปนซี (สกุล Pan ) ได้แยกตัวออกจากสายพันธุ์ที่นำไปสู่มนุษย์ ดีเอ็นเอของมนุษย์มีความเหมือนกับดีเอ็นเอของชิมแปนซีประมาณ 98.4% เมื่อเปรียบเทียบความแตกต่างของนิวคลีโอไทด์เดี่ยว (ดูพันธุศาสตร์วิวัฒนาการของมนุษย์ ) อย่างไรก็ตาม บันทึกฟอสซิลของกอริลลาและชิมแปนซีมีจำกัด ทั้งการเก็บรักษาที่ไม่ดี – ดินในป่าฝนมีแนวโน้มที่จะเป็นกรดและละลายกระดูก – และอคติในการสุ่มตัวอย่างอาจเป็นปัจจัยที่ทำให้เกิดปัญหานี้
โฮมินินกลุ่มอื่นๆ อาจปรับตัวให้เข้ากับสภาพแวดล้อมที่แห้งแล้งกว่านอกเขตเส้นศูนย์สูตร และที่นั่นพวกเขาได้พบกับละมั่ง ไฮยีน่า สุนัข หมู ช้าง ม้า และสัตว์อื่นๆ เขตเส้นศูนย์สูตรหดตัวลงหลังจากประมาณ 8 ล้านปีก่อน และมีหลักฐานฟอสซิลน้อยมากสำหรับการแยกสายพันธุ์—ซึ่งเชื่อว่าเกิดขึ้นในช่วงเวลานั้น—ของสายพันธุ์โฮมินินออกจากสายพันธุ์ของกอริลลาและชิมแปนซี ฟอสซิลที่เก่าแก่ที่สุดที่บางคนอ้างว่าอยู่ในสายพันธุ์มนุษย์คือSahelanthropus tchadensis (7 ล้านปีก่อน) และOrrorin tugenensis (6 ล้านปีก่อน) ตามด้วยArdipithecus (5.5–4.4 ล้านปีก่อน) ซึ่งมีสายพันธุ์Ar. kadabbaและAr. ramidus

มีการโต้แย้งในการศึกษาประวัติชีวิตของAr. ramidusว่าสายพันธุ์นี้ให้หลักฐานสำหรับการปรับตัวทางกายวิภาคและพฤติกรรมในโฮมินินยุคแรกเริ่ม ซึ่งแตกต่างจากสายพันธุ์ลิงใหญ่ที่ยังมีชีวิตอยู่[ 33 ]การศึกษานี้แสดงให้เห็นถึงความสัมพันธ์ระหว่างสัณฐานวิทยาของกะโหลกศีรษะของAr. ramidus กับของทารกและชิมแปนซีวัยรุ่น ซึ่งชี้ให้เห็นว่าสายพันธุ์นี้วิวัฒนาการ สัณฐานวิทยาของกะโหลกศีรษะและใบหน้าแบบเด็กหรือแบบเพโดมอร์ฟิก ผ่านการแยก ตัวของวิถีการเจริญเติบโตแบบเฮเทอโรโครนิก นอกจากนี้ยังมีการโต้แย้งว่าสายพันธุ์นี้ให้การสนับสนุนแนวคิดที่ว่าโฮมินินยุคแรกเริ่ม คล้ายกับโบโนโบ ( Pan paniscus ) ซึ่งเป็นสายพันธุ์ที่ไม่ก้าวร้าวมากนักในสกุลPanอาจวิวัฒนาการผ่านกระบวนการทำให้เชื่องด้วยตนเอง ด้วยเหตุนี้ ผู้เขียนจึงโต้แย้งกับสิ่งที่เรียกว่า "แบบจำลองอ้างอิงชิมแปนซี" [ 34 ]โดยเสนอว่าไม่สามารถใช้พฤติกรรมทางสังคมและการผสมพันธุ์ของชิมแปนซี ( Pan troglodytes ) ในแบบจำลองวิวัฒนาการทางสังคมของโฮมินินยุคแรกได้อีกต่อไป เมื่อแสดงความคิดเห็นเกี่ยวกับการไม่มีลักษณะทางกายภาพของสุนัขที่ก้าวร้าวในAr. ramidusและนัยยะที่สิ่งนี้มีต่อวิวัฒนาการของจิตวิทยาสังคมของโฮมินิน พวกเขาเขียนว่า:
แน่นอนว่าAr. ramidusแตกต่างจากโบโนโบอย่างมาก โดยโบโนโบยังคงมีระบบลับคมเขี้ยวที่ใช้งานได้ อย่างไรก็ตาม ข้อเท็จจริงที่ว่าAr. ramidusมีลักษณะทางเพศที่ลดลงร่วมกับโบโนโบ และมีรูปร่างคล้ายเด็กมากกว่าเมื่อเทียบกับชิมแปนซี บ่งชี้ว่าการปรับตัวด้านพัฒนาการและสังคมที่เห็นได้ชัดในโบโนโบอาจเป็นประโยชน์ในการสร้างภาพใหม่เกี่ยวกับจิตวิทยาสังคมและทางเพศของโฮมินินยุคแรกในอนาคต อันที่จริง แนวโน้มไปสู่การดูแลของแม่ที่เพิ่มขึ้น การเลือกคู่ครองของเพศเมีย และการเลี้ยงตัวเองอาจมีความแข็งแกร่งและละเอียดอ่อนกว่าในAr. ramidusมากกว่าที่เราเห็นในโบโนโบ[ 33 ] : 128
ผู้เขียนโต้แย้งว่า การปรับตัวพื้นฐานหลายอย่างของมนุษย์นั้นวิวัฒนาการขึ้นในระบบนิเวศป่าไม้โบราณของแอฟริกาในช่วงปลายสมัยไมโอซีนและต้นสมัยพลิโอซีนดังนั้น พวกเขาจึงโต้แย้งว่ามนุษย์อาจไม่ได้วิวัฒนาการมาจากบรรพบุรุษที่มีลักษณะคล้ายชิมแปนซีอย่างที่เคยเชื่อกันมา นี่แสดงให้เห็นว่าการปรับตัวของมนุษย์ยุคใหม่หลายอย่างเป็น ลักษณะ ที่สืบทอดมาจากวิวัฒนาการระดับลึก และพฤติกรรมและรูปร่างของชิมแปนซีอาจวิวัฒนาการขึ้นหลังจากแยกตัวออกจากบรรพบุรุษร่วมกันกับมนุษย์แล้ว
สกุลออสตราโลพิเทคัส

สกุลAustralopithecusวิวัฒนาการขึ้นในแอฟริกาตะวันออกเมื่อประมาณ 4 ล้านปีก่อน ก่อนที่จะแพร่กระจายไปทั่วทวีปและสูญพันธุ์ไปในที่สุดเมื่อ 2 ล้านปีก่อน ในช่วงเวลานั้นมีออสตราโลพิเทคัสหลายรูปแบบอาศัยอยู่ รวมถึงAustralopithecus anamensis , A. afarensis , A. sedibaและA. africanusยังคงมีการถกเถียงกันในหมู่นักวิชาการว่าโฮมินิดบางชนิดในแอฟริกาในช่วงเวลานั้น เช่นP. robustusและP. boiseiจัดอยู่ในสกุลเดียวกันหรือไม่ หากเป็นเช่นนั้น พวกมันจะถูกพิจารณาว่าเป็น "ออสตราโลพิเทคัสที่แข็งแรง" ในขณะที่ชนิดอื่นๆ จะถูกพิจารณาว่าเป็น "ออสตราโลพิเทคัสที่บอบบาง" อย่างไรก็ตาม หากสายพันธุ์เหล่านี้จัดอยู่ในสกุลของตนเองจริง ๆ พวกมันอาจได้รับชื่อใหม่ว่า Paranthropus
- Australopithecus (4–1.8 Ma) โดยมีสายพันธุ์ A. anamensis , A. afarensis , A. africanus , A. bahrelghazali , A. garhiและ A. sediba ;
- Kenyanthropus (3–2.7 ล้านปี) โดยมีสายพันธุ์ K. platyops ;
- Paranthropus (3–1.2 Ma) โดยมีสายพันธุ์ P. aethiopicus , P. boiseiและ P. โรบัสตัส
มี การอ้างว่ามีการค้นพบ สายพันธุ์ใหม่ที่เสนอชื่อAustralopithecus deyiremedaซึ่งมีชีวิตอยู่ในช่วงเวลาเดียวกันกับA. afarensis มีการถกเถียงกันว่า A. deyiremedaเป็นสายพันธุ์ใหม่หรือเป็นA. afarensis [ 35 ] Australopithecus prometheusหรือที่รู้จักกันในชื่อLittle Footเพิ่งได้รับการกำหนดอายุไว้ที่ 3.67 ล้านปีโดยใช้เทคนิคการหาอายุแบบใหม่ ทำให้สกุลAustralopithecusมีอายุเก่าแก่เท่ากับafarensis [ 36 ] เมื่อพิจารณาจากนิ้วหัวแม่เท้าที่สามารถงอได้ที่พบใน Little Foot ดูเหมือนว่าตัวอย่างนี้จะเป็นนักปีนป่ายที่ดี เชื่อกันว่าเนื่องจากมีสัตว์นักล่าในเวลากลางคืนในภูมิภาคนี้ เขาจึงสร้างแท่นทำรังในเวลากลางคืนบนต้นไม้ในลักษณะเดียวกับชิมแปนซีและกอริลลา
วิวัฒนาการของสกุลHomo
−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — | ( O. praegens ) ( O. tugenensis ) ( อาร์. รามิดัส ) H. habilis ( H. rudolfensis )( Au. garhi ) H. erectus ( H. บรรพบุรุษ )( H. ergaster )( Au. sediba ) |
| ||||||||||||||||||||||||||
( ล้านปีก่อน ) | ||||||||||||||||||||||||||||
ตัวแทนที่เก่าแก่ที่สุดที่ได้รับการบันทึกไว้ของสกุลHomoคือขากรรไกร Lediซึ่งมีอายุ 2.75 [ 37 ] [ 38 ] – 2.8 ล้านปีก่อน (mya) [ 39 ] [ 40 ]และอาจเป็นสายพันธุ์ที่เก่าแก่ที่สุดที่มีหลักฐานยืนยันการใช้เครื่องมือหิน สมองของโฮมินินยุคแรกเหล่านี้มีขนาดใกล้เคียงกับสมองของชิมแปนซี แม้ว่าจะมีการเสนอแนะว่านี่เป็นช่วงเวลาที่ยีนSRGAP2 ของมนุษย์ เพิ่มจำนวนเป็นสองเท่า ทำให้เกิดการเชื่อมต่อของคอร์เทกซ์ส่วนหน้าอย่างรวดเร็วยิ่งขึ้น ในช่วงหนึ่งล้านปีต่อมา กระบวนการเพิ่มขนาดสมอง อย่างรวดเร็ว ได้เกิดขึ้น และด้วยการปรากฏตัวของHomo erectusและHomo ergasterในบันทึกฟอสซิล ความจุของกะโหลกศีรษะจึงเพิ่มขึ้นเป็นสอง เท่าเป็น 850 cm³ [ 41 ] (การเพิ่มขึ้นของขนาดสมองของมนุษย์ดังกล่าวเทียบเท่ากับการที่แต่ละรุ่นมีเซลล์ประสาท มากกว่า พ่อแม่ถึง 125,000 เซลล์) เชื่อกันว่าH. erectusและH. ergasterเป็นกลุ่มแรกที่ใช้ไฟและเครื่องมือที่ซับซ้อน และเป็นกลุ่มแรกของสายพันธุ์โฮมินินที่ออกจากแอฟริกา แพร่กระจายไปทั่วแอฟริกา เอเชีย และยุโรป ระหว่าง1.3 ถึง 1.8 ล้านปีก่อน
ตามทฤษฎีต้นกำเนิดแอฟริกาเมื่อเร็ว ๆ นี้ มนุษย์ยุคใหม่วิวัฒนาการในแอฟริกาโดยอาจมาจากH. heidelbergensis , H. rhodesiensisหรือH. antecessorและอพยพออกจากทวีปเมื่อประมาณ 50,000 ถึง 100,000 ปีที่แล้ว โดยค่อย ๆ แทนที่ประชากรท้องถิ่นของH. erectus , โฮมินินเดนิโซวา , H. floresiensis , H. luzonensisและH. neanderthalensisซึ่งบรรพบุรุษของพวกเขาได้ออกจากแอฟริกาในการอพยพครั้งก่อน[ 42 ] [ 43 ] [ 44 ] [ 45 ] [ 46 ]โฮโมเซเปียนส์โบราณซึ่งเป็นบรรพบุรุษของมนุษย์ยุคใหม่ที่มีลักษณะทางกายวิภาค วิวัฒนาการในยุคหินเก่าตอนกลางระหว่าง 400,000 ถึง 250,000 ปีที่แล้ว[ 47 ] [ 48 ] [ 49 ] หลักฐาน ดีเอ็นเอล่าสุดชี้ให้เห็นว่าแฮพลอไทป์ หลายชนิดที่มีต้นกำเนิด จากนีแอนเดอร์ทัลมีอยู่ในประชากรที่ไม่ใช่ชาวแอฟริกันทั้งหมด และนีแอนเดอร์ทัลและโฮมินินอื่นๆ เช่น เดนิโซแวน อาจมีส่วนร่วมในจีโนม มากถึง 6% ของ มนุษย์ในปัจจุบัน ซึ่งบ่งชี้ถึงการผสมพันธุ์ข้ามสายพันธุ์ที่จำกัดระหว่างสายพันธุ์เหล่านี้ [ 50 ] [ 51 ] [ 52 ] ตามที่นักมานุษยวิทยาบางคนกล่าว การเปลี่ยนผ่านไปสู่ความทันสมัยทางพฤติกรรมด้วยการพัฒนาวัฒนธรรมเชิงสัญลักษณ์ ภาษา และเทคโนโลยีหิน เฉพาะทาง เกิดขึ้นเมื่อประมาณ 50,000 ปีที่แล้ว (จุดเริ่มต้นของยุคหินเก่าตอนบน ) [ 53 ]แม้ว่าคนอื่นๆ จะชี้ให้เห็นหลักฐานของการเปลี่ยนแปลงทีละน้อยในช่วงเวลาที่ยาวนานกว่าในยุคหินเก่าตอนกลาง[ 54 ]
Homo sapiens เป็น สปีชีส์เดียว ที่ยังมีชีวิตอยู่ ของสกุลHomoในขณะที่บาง สปีชีส์ของ Homo (ที่สูญพันธุ์ไปแล้ว) อาจเป็นบรรพบุรุษของHomo sapiensแต่หลายสปีชีส์ หรืออาจจะส่วนใหญ่ น่าจะเป็น "ญาติ" ที่แยกตัวออกจากสายพันธุ์บรรพบุรุษของโฮมินิน[ 55 ] [ 56 ]ยังไม่มีข้อสรุปที่แน่ชัดว่ากลุ่มใดควรได้รับการพิจารณาว่าเป็นสปีชีส์ที่แยกต่างหาก และกลุ่มใดควรเป็นสปีชีส์ย่อย ซึ่งอาจเป็นเพราะฟอสซิลมีน้อย หรือเนื่องจากความแตกต่างเล็กน้อยที่ใช้ในการจำแนกสปีชีส์ในสกุลHomo [ 56 ] ทฤษฎีปั๊มน้ำซาฮารา (ซึ่งอธิบายถึงทะเลทรายซาฮารา ที่"เปียก"ซึ่งสามารถสัญจรผ่านได้เป็นบางครั้ง ) เป็นคำอธิบายที่เป็นไปได้ประการหนึ่งของการอพยพและการแยกสปีชีส์เป็นระยะ ๆใน สกุลHomo
จากหลักฐานทางโบราณคดีและบรรพชีวินวิทยา ทำให้สามารถอนุมานถึงแนวทางการบริโภคอาหารในสมัยโบราณ[ 57 ]ของ สายพันธุ์ Homo ต่างๆ ได้ในระดับหนึ่ง และศึกษาบทบาทของอาหารในการวิวัฒนาการทางกายภาพและพฤติกรรมภายใน Homo [ 58 ] [ 59 ] [ 60 ] [ 61 ] [ 62 ]
นักมานุษยวิทยาและนักโบราณคดีบางคนเชื่อในทฤษฎีภัยพิบัติโตบาซึ่งระบุว่าการระเบิดครั้งใหญ่ของทะเลสาบโตบาบนเกาะสุมาตราในอินโดนีเซียเมื่อราว 70,000 ปีก่อน ทำให้เกิดภาวะอดอยากทั่วโลก[ 63 ]ส่งผลให้มนุษย์ส่วนใหญ่เสียชีวิตและเกิดภาวะคอขวดของประชากรซึ่งส่งผลต่อการถ่ายทอดทางพันธุกรรมของมนุษย์ทุกคนในปัจจุบัน[ 64 ]อย่างไรก็ตาม หลักฐานทางพันธุกรรมและโบราณคดีสำหรับเรื่องนี้ยังคงเป็นที่ถกเถียงกันอยู่[ 65 ]การศึกษาทางพันธุกรรมในปี 2023 ชี้ให้เห็นว่าภาวะคอขวดของประชากร มนุษย์ที่คล้ายกัน ซึ่งมีผู้รอดชีวิตระหว่าง 1,000 ถึง 100,000 คน เกิดขึ้น "เมื่อราว 930,000 ถึง 813,000 ปีก่อน ... กินเวลานานประมาณ 117,000 ปี และทำให้บรรพบุรุษของมนุษย์เกือบจะสูญพันธุ์" [ 66 ] [ 67 ]
H. habilisและH. gautengensis
โฮโม ฮาบิลิสมีชีวิตอยู่ตั้งแต่ประมาณ 2.8 [ 40 ]ถึง 1.4 ล้านปีก่อน สายพันธุ์นี้วิวัฒนาการในแอฟริกาใต้และแอฟริกาตะวันออกในช่วงปลายไพลโอซีนหรือต้นไพลสโตซีนประมาณ 2.5–2 ล้านปีก่อน เมื่อมันแยกตัวออกจากออสตราโลพิเทคัสด้วยการพัฒนาของฟันกรามที่เล็กลงและสมองที่ใหญ่ขึ้น มันเป็นหนึ่งในโฮมินินกลุ่มแรกๆ ที่รู้จักกัน มันสร้างเครื่องมือจากหินและอาจรวมถึงกระดูกสัตว์ด้วย ซึ่งนำไปสู่ชื่อโฮโมฮาบิลิส (ภาษาละติน 'มนุษย์ผู้มีฝีมือ') ที่ผู้ค้นพบ หลุยส์ ลีคีย์ ตั้งให้ นักวิทยาศาสตร์บางคนเสนอให้ย้ายสายพันธุ์นี้จากโฮโมไปเป็นออสตราโลพิเทคัสเนื่องจากสัณฐานวิทยาของโครงกระดูกของมันปรับตัวให้เข้ากับการอาศัยอยู่บนต้นไม้มากกว่าการเดินด้วยสองขาเหมือนโฮมินินในยุคหลัง[ 68 ]
ในเดือนพฤษภาคม พ.ศ. 2553 มีการค้นพบสายพันธุ์ใหม่Homo gautengensisในแอฟริกาใต้[ 69 ]
H. rudolfensisและH. georgicus
นี่คือชื่อสายพันธุ์ที่เสนอสำหรับฟอสซิลจากช่วงประมาณ 1.9–1.6 ล้านปีก่อน ซึ่งความสัมพันธ์กับHomo habilisยังไม่ชัดเจน
- Homo rudolfensisหมายถึงกะโหลกศีรษะที่ไม่สมบูรณ์เพียงชิ้นเดียวจากเคนยา นักวิทยาศาสตร์เสนอว่านี่เป็นตัวอย่างของHomo habilisแต่ยังไม่ได้รับการยืนยัน[ 70 ]ปัจจุบันเชื่อกันว่าH. rudolfensisเป็นสายพันธุ์โฮมินิดยุคแรก ควบคู่ไปกับH. erectusและH. habilis [ 71 ]
- Homo georgicusจากจอร์เจียอาจเป็นรูปแบบกลางระหว่าง Homo habilisและ Homo erectus [ 72 ] หรือ เป็นสายพันธุ์ย่อยของ Homo erectus [ 73 ]
เอช. เออร์แกสเตอร์และเอช. อิเร็กตัส

The first fossils of Homo erectus were discovered by Dutch physician Eugene Dubois in 1891 on the Indonesian island of Java. He originally named the material Anthropopithecus erectus (1892–1893, considered at this point as a chimpanzee-like fossil primate) and Pithecanthropus erectus (1893–1894, changing his mind as of based on its morphology, which he considered to be intermediate between that of humans and apes).[74] Years later, in the 20th century, the German physician and paleoanthropologistFranz Weidenreich (1873–1948) compared in detail the characters of Dubois' Java Man, then named Pithecanthropus erectus, with the characters of the Peking Man, then named Sinanthropus pekinensis. Weidenreich concluded in 1940 that because of their anatomical similarity with modern humans it was necessary to gather all these specimens of Java and China in a single species of the genus Homo, the species H. erectus.[75][76]
Homo erectus lived from about 1.8 Ma to about 108,000 years ago.[77] This population appears to have died out when the savannah corridors closed, and tropical jungle took over.;[78] however, nearby H. floresiensis survived it. The early phase of H. erectus, from 1.8 to 1.25 Ma, is considered by some to be a separate species, H. ergaster, or as H. erectus ergaster, a subspecies of H. erectus. Many paleoanthropologists now use the term Homo ergaster for the non-Asian forms of this group, and reserve H. erectus only for those fossils that are found in Asia and meet certain skeletal and dental requirements which differ slightly from H. ergaster.
ในแอฟริกาในช่วงต้นสมัยไพลสโตซีน 1.5–1 ล้านปีก่อนเชื่อกันว่า ประชากรบางกลุ่มของ โฮโม ฮาบิลิส มีวิวัฒนาการสมองที่ใหญ่ขึ้นและสร้างเครื่องมือหินที่ซับซ้อนมากขึ้น ความแตกต่างเหล่านี้และอื่นๆ เพียงพอสำหรับนักมานุษยวิทยาที่จะจัดจำแนกพวกเขาเป็นสายพันธุ์ใหม่ โฮโม อิเร็กตัสในแอฟริกา[ 79 ]สายพันธุ์นี้อาจใช้ไฟในการปรุงเนื้อสัตว์ ด้วย ริชาร์ด แรงแฮม ตั้งข้อสังเกตว่าโฮโมดูเหมือนจะอาศัยอยู่บนพื้นดิน มีลำไส้สั้นลง ฟันเล็กลง และ "สมอง [บวม] จนมีขนาดใหญ่ในปัจจุบัน ซึ่งสิ้นเปลืองเชื้อเพลิงอย่างน่ากลัว" [ 80 ]และตั้งสมมติฐานว่าการควบคุมไฟและการปรุงอาหาร ซึ่งปลดปล่อยคุณค่าทางโภชนาการที่เพิ่มขึ้น เป็นการปรับตัวที่สำคัญที่แยกโฮโม ออก จากออสตราโลพิเทคัสที่นอนบนต้นไม้[ 81 ]
H. cepranensisและH. บรรพบุรุษ
มีการเสนอว่าสายพันธุ์เหล่านี้เป็นสายพันธุ์ที่อยู่ระหว่างH. erectusและH. heidelbergensis
- H. antecessorเป็นที่รู้จักจากฟอสซิลจากสเปนและอังกฤษที่มีอายุ 1.2 ล้านปีถึง 500,000 ปี[ 82 ] [ 83 ] ฟอสซิล กระดูกสะบัก ของ H. antecessorสองชิ้นถูกค้นพบที่ถ้ำ Gran Dolina ในสเปน นักวิจัยได้เปรียบเทียบกระดูกสะบักที่พบกับฟอสซิลอื่นๆ ของชิมแปนซี มนุษย์ยุคใหม่ และมนุษย์ถ้ำ พวกเขาพบว่า ฟอสซิลของ H. antecessorมีความคล้ายคลึงกับมนุษย์ยุคใหม่ในด้านพัฒนาการมากกว่า ซึ่งสอดคล้องกับผลการค้นพบก่อนหน้านี้เกี่ยวกับกะโหลกของH. antecessorซึ่งกระดูกกะโหลกของพวกมันแสดงลักษณะที่ทั้งโฮมินินยุคกลางไพลสโตซีนของจีนและมนุษย์ยุคใหม่มีร่วมกัน ผลการค้นพบเหล่านี้ชี้ให้เห็นว่าH. cepranensisอาจเป็นบรรพบุรุษร่วมสุดท้ายระหว่างH. sapiens , H. neanderthalensisและ Denisovans แม้ว่าจะต้องมีการวิจัยเพิ่มเติมเพื่อตรวจสอบสมมติฐานนี้ก็ตาม[ 84 ]การศึกษาในปี 2020 ระบุว่าH. antecessorซึ่งมีโครงสร้างใบหน้าคล้ายกับH. sapiensจัดอยู่ในสกุลHomoในขณะที่โครงสร้างกะโหลกศีรษะของนีแอนเดอร์ทาลเป็นโครงสร้างที่พัฒนามาจาก H. sapiens [ 85 ]
- H. cepranensisหมายถึงกะโหลกศีรษะชิ้นเดียวจากอิตาลี ซึ่งคาดว่ามีอายุราว 800,000 ปี [ 86 ]ในปี 1994 นักวิจัยในอิตาลีค้นพบชิ้นส่วนกะโหลกศีรษะ หลังจากประกอบเข้าด้วยกัน รูปร่างที่โดดเด่นทำให้แยกชิ้นส่วนเหล่านี้ออกจากสายพันธุ์โฮมินิดอื่นๆ ส่งผลให้นักวิจัยเชื่อว่าเป็นส่วนหนึ่งของสายพันธุ์โฮมินิดที่ไม่เคยรู้จักมาก่อน พวกเขาตั้งชื่อว่า H. cepranensis sp. nov. ตามชื่อสถานที่ที่พบ (Campo Grande of Ceprano) เมื่อเปรียบเทียบกะโหลกศีรษะกับสายพันธุ์โฮมินิดอื่นๆ ที่รู้จัก พวกเขาพบว่า กระดูก ของ H. cepranensis sp. nov. แตกต่างกันในด้านความหนา ความยาว และมุมของกระดูก ซึ่งนำไปสู่ข้อสรุปว่า H. cepranensis sp. nov. เป็นสายพันธุ์โฮมินิดที่แตกต่างออกไป [ 87 ]
เอช. ไฮเดลเบอร์เกนซิส

H. heidelbergensis ("มนุษย์ไฮเดลเบิร์ก") มีชีวิตอยู่ตั้งแต่ประมาณ 800,000 ถึงประมาณ 300,000 ปีที่แล้ว มีการเสนอชื่อนี้ว่าHomo sapiens heidelbergensisหรือHomo sapiens paleohungaricus [ 88 ]
ในปี ค.ศ. 1907 คนงานเหมืองในเยอรมนีค้นพบขากรรไกรล่างของเมาเออร์ ซึ่งเป็นขากรรไกรดึกดำบรรพ์ที่เก่าแก่มาก การวิเคราะห์ขากรรไกรโดยออตโต โชเทนแซค นำไปสู่การจัดให้ขากรรไกรนี้เป็นของสายพันธุ์ใหม่ คือH. heidelbergensisความถูกต้องของสายพันธุ์นี้ในฐานะสายพันธุ์ของตัวเองและบรรพบุรุษของมนุษย์นีแอนเดอร์ทัลนั้นเป็นที่ถกเถียงกัน ในปี ค.ศ. 2009 มีการศึกษาเพื่อประเมินขากรรไกรล่างของเมาเออร์อีกครั้งโดยเปรียบเทียบกับซากดึกดำบรรพ์อื่นๆ เพื่อพิจารณาว่าH. heidelbergensisอาจอยู่ในระดับวิวัฒนาการของมนุษย์ในระดับใด หรือแม้แต่ว่ามันมีความถูกต้องในฐานะสายพันธุ์ที่แยกต่างหากหรือไม่ จากการประเมินใหม่นี้ นักวิจัยสรุปว่าH. heidelbergensisน่าจะเป็นสายพันธุ์แอฟริกา-ยุโรปที่เป็นบรรพบุรุษร่วมกันของทั้งมนุษย์นีแอนเดอร์ทัลและH. sapiensแม้ว่าจะเป็นไปได้เช่นกันว่าH. heidelbergensisเป็นสายพันธุ์ยุโรปที่เป็นบรรพบุรุษของมนุษย์นีแอนเดอร์ทัลเท่านั้น ในขณะที่สายพันธุ์อื่นจากแอฟริกาเป็นบรรพบุรุษของH. sapiens [ 89 ]
H. rhodesiensisและกะโหลกศีรษะ Gawis
- ในปี พ.ศ. 2464 มีการค้นพบกะโหลกศีรษะในถ้ำระหว่างการสำรวจเหมืองแร่ในพื้นที่ซึ่งปัจจุบันรู้จักกันในชื่อประเทศแซมเบียคาดว่ามีอายุ 300,000–125,000 ปี[ 90 ] นักวิจัยส่วนใหญ่ในปัจจุบันจัดให้มนุษย์โร ดีเซียอยู่ในกลุ่มHomo heidelbergensisแม้ว่าจะมีการเสนอชื่อเรียกอื่นๆ เช่นHomo sapiens โบราณ และHomo sapiens rhodesiensis ก็ตาม ฟอสซิลนี้ได้ขยายแนวคิดเกี่ยวกับวิวัฒนาการของH. sapiens การประมาณการในปัจจุบันระบุว่า H. sapiensถือกำเนิดขึ้นเมื่อประมาณ 500,000 ปีที่แล้ว[ 91 ]
- ในเดือนกุมภาพันธ์ พ.ศ. 2549 มีการค้นพบฟอสซิลกะโหลกศีรษะกาวิสซึ่งอาจเป็นสายพันธุ์ที่อยู่ระหว่างH. erectusและH. sapiensหรืออาจเป็นจุดสิ้นสุดของการวิวัฒนาการ กะโหลกศีษะจากกาวิส ประเทศเอธิโอเปีย เชื่อว่ามีอายุ 500,000–250,000 ปี มีเพียงรายละเอียดโดยสรุปเท่านั้นที่ทราบ และผู้ค้นพบยังไม่ได้เผยแพร่การศึกษาที่ผ่านการตรวจสอบโดยผู้เชี่ยวชาญ ลักษณะใบหน้าของมนุษย์กาวิสบ่งชี้ว่าเป็นสายพันธุ์ที่อยู่ระหว่างกลางหรือเป็นตัวอย่างของเพศหญิง "มนุษย์โบโด" [ 92 ]
มนุษย์นีแอนเดอร์ทาลและมนุษย์เดนิโซวัน

Homo neanderthalensisหรือเรียกอีกอย่างว่าHomo sapiens neanderthalensis [ 93 ]อาศัยอยู่ในยุโรปและเอเชียตั้งแต่ 400,000 [ 94 ]ถึงประมาณ 28,000 ปีที่แล้ว[ 95 ] มีความแตกต่างทางกายวิภาคที่ชัดเจนหลายประการระหว่างมนุษย์ยุคใหม่( AMH) และตัวอย่างนีแอนเดอร์ทัล ซึ่งหลายอย่างเกี่ยวข้องกับการปรับตัวที่เหนือกว่าของนีแอนเดอร์ทัลต่อสภาพแวดล้อมที่หนาวเย็น อัตราส่วนพื้นผิวต่อปริมาตรของนีแอนเดอร์ทัลยังต่ำกว่าใน ประชากร อินูอิต ยุคใหม่ ซึ่งบ่งชี้ถึงการกักเก็บความร้อนในร่างกายที่เหนือกว่า
นอกจากนี้ มนุษย์นีแอนเดอร์ทาลยังมีสมองที่ใหญ่กว่าอย่างเห็นได้ชัด ดังที่แสดงให้เห็นจากแบบจำลองสมองภายใน ซึ่งทำให้เกิดข้อสงสัยเกี่ยวกับความด้อยกว่าทางสติปัญญาของพวกเขาเมื่อเทียบกับมนุษย์ยุคใหม่ อย่างไรก็ตาม มวลร่างกายที่มากกว่าของมนุษย์นีแอนเดอร์ทาลอาจต้องการมวลสมองที่ใหญ่กว่าเพื่อการควบคุมร่างกาย[ 96 ]นอกจากนี้ การวิจัยล่าสุดโดย Pearce, Stringerและ Dunbar ได้แสดงให้เห็นถึงความแตกต่างที่สำคัญในโครงสร้างของสมอง ขนาดที่ใหญ่กว่าของช่องเบ้าตาและกลีบสมองส่วนท้ายทอย ของมนุษย์นีแอนเดอร์ทาล บ่งชี้ว่าพวกเขามีความสามารถในการมองเห็นที่ดีกว่ามนุษย์ยุคใหม่ ซึ่งมีประโยชน์ในสภาพแสงที่สลัวกว่าของยุโรปในยุคน้ำแข็ง
มนุษย์นีแอนเดอร์ทาลอาจมีสมองที่มีความจุน้อยกว่าสำหรับการทำงานทางสังคมการอนุมานขนาดกลุ่มสังคมจากปริมาตรภายในกะโหลกศีรษะ (ลบด้วยขนาดกลีบสมองส่วนท้ายทอย) ชี้ให้เห็นว่ากลุ่มมนุษย์นีแอนเดอร์ทาลอาจมีจำนวนจำกัดเพียง 120 คน เมื่อเทียบกับความสัมพันธ์ที่เป็นไปได้ 144 แบบสำหรับมนุษย์ยุคใหม่[ 97 ]กลุ่มสังคมที่ใหญ่ขึ้นอาจหมายความว่ามนุษย์ยุคใหม่มีความเสี่ยงน้อยลงต่อการผสมพันธุ์ในหมู่ญาติ การค้าขายในพื้นที่ที่กว้างขึ้น (ได้รับการยืนยันในการกระจายของเครื่องมือหิน) และการแพร่กระจายนวัตกรรมทางสังคมและเทคโนโลยีที่รวดเร็วยิ่งขึ้น ปัจจัยทั้งหมดนี้อาจมีส่วนทำให้โฮโมเซเปียนส์ ยุคใหม่ เข้ามาแทนที่ประชากรมนุษย์นีแอนเดอร์ทาลเมื่อ 28,000 ปีก่อน[ 96 ]
หลักฐานก่อนหน้านี้จากการจัดลำดับดีเอ็นเอไมโทคอนเดรียชี้ให้เห็นว่าไม่มีการถ่ายทอดยีนอย่างมีนัยสำคัญระหว่างH. neanderthalensisและH. sapiensและทั้งสองเป็นสายพันธุ์ที่แยกจากกันซึ่งมีบรรพบุรุษร่วมกันเมื่อประมาณ 660,000 ปีที่แล้ว[ 98 ] [ 99 ] [ 100 ]อย่างไรก็ตาม การจัดลำดับจีโนมของนีแอนเดอร์ทัลในปี 2010 ระบุว่านีแอนเดอร์ทัลได้ผสมพันธุ์กับมนุษย์ยุคใหม่ที่มีลักษณะทางกายวิภาคเหมือนมนุษย์ปัจจุบันเมื่อประมาณ 45,000-80,000 ปีที่แล้ว ซึ่งเป็นช่วงเวลาที่มนุษย์ยุคใหม่อพยพออกจากแอฟริกา แต่ก่อนที่พวกเขาจะกระจายไปทั่วยุโรป เอเชีย และที่อื่นๆ[ 101 ]การจัดลำดับพันธุกรรมของโครงกระดูกมนุษย์อายุ 40,000 ปีจากโรมาเนียแสดงให้เห็นว่า 11% ของจีโนมเป็นนีแอนเดอร์ทัล ซึ่งหมายความว่าบุคคลนั้นมีบรรพบุรุษเป็นนีแอนเดอร์ทัลเมื่อ 4–6 รุ่นก่อน[ 102 ]นอกเหนือจากการผสมพันธุ์ข้ามสายพันธุ์ในตะวันออกกลางก่อนหน้านี้ แม้ว่าประชากรโรมาเนียที่ผสมพันธุ์ข้ามสายพันธุ์นี้ดูเหมือนจะไม่ใช่บรรพบุรุษของมนุษย์ยุคใหม่ แต่การค้นพบนี้บ่งชี้ว่ามีการผสมพันธุ์ข้ามสายพันธุ์เกิดขึ้นซ้ำแล้วซ้ำเล่า[ 103 ]
มนุษย์ยุคใหม่ที่ไม่ใช่ชาวแอฟริกันทั้งหมดมีดีเอ็นเอประมาณ 1% ถึง 4% (หรือ 1.5% ถึง 2.6% ตามข้อมูลล่าสุด) ที่ได้มาจากนีแอนเดอร์ทาล[ 104 ] [ 101 ] [ 105 ]การค้นพบนี้สอดคล้องกับการศึกษาล่าสุดที่ระบุว่าการแยกตัวของอัลลีลของมนุษย์บางส่วนมีอายุย้อนไปถึงหนึ่งล้านปี แม้ว่าการตีความนี้จะถูกตั้งคำถามก็ตาม[ 106 ] [ 107 ]นีแอนเดอร์ทาลและโฮโมเซเปียนส์ยุคใหม่ (AMH Homo sapiens)อาจอยู่ร่วมกันในยุโรปได้นานถึง 10,000 ปี ในช่วงเวลานั้นประชากร AMH เพิ่มจำนวนขึ้นอย่างมาก มีจำนวนมากกว่านีแอนเดอร์ทาลอย่างมหาศาล และอาจเอาชนะนีแอนเดอร์ทาลได้ด้วยจำนวนที่มากกว่า[ 108 ]
ในปี 2551 นักโบราณคดีที่ทำงานในถ้ำเดนิโซวาในเทือกเขาอัลไตของไซบีเรียได้ค้นพบชิ้นส่วนกระดูกเล็กๆ จากนิ้วที่ห้าของสมาชิกวัยเยาว์ของมนุษย์อีกสายพันธุ์หนึ่ง คือชาวเดนิโซวา[ 109 ]วัตถุโบราณ รวมถึงกำไล ที่ขุดพบในถ้ำในระดับเดียวกันนั้น ได้รับ การหาอายุด้วย คาร์บอน พบว่ามีอายุประมาณ 40,000 ปีก่อนคริสตกาล เนื่องจากดีเอ็นเอรอดชีวิตอยู่ในชิ้นส่วนฟอสซิลเนื่องจากสภาพอากาศที่เย็นของถ้ำเดนิโซวา จึงได้ทำการจัดลำดับทั้งดีเอ็นเอไมโทคอนเดรียและดีเอ็นเอนิวเคลียร์[ 50 ] [ 110 ]
แม้ว่าจุดแยกของ mtDNA จะอยู่ในช่วงเวลาที่ยาวนานอย่างไม่คาดคิด[ 111 ]แต่ลำดับจีโนมทั้งหมดกลับชี้ให้เห็นว่าชาวเดนิโซแวนอยู่ในสายพันธุ์เดียวกันกับชาวนีแอนเดอร์ทาล โดยทั้งสองสายพันธุ์แยกตัวออกจากกันไม่นานหลังจากที่สายพันธุ์ของพวกเขาแยกออกจากสายพันธุ์ที่ให้กำเนิดมนุษย์ยุคใหม่[ 50 ]เป็นที่ทราบกันว่ามนุษย์ยุคใหม่เคยอาศัยอยู่ร่วมกับชาวนีแอนเดอร์ทาลในยุโรปและตะวันออกใกล้เป็นเวลานานกว่า 40,000 ปี[ 112 ]และการค้นพบนี้ทำให้เกิดความเป็นไปได้ว่าชาวนีแอนเดอร์ทาล ชาวเดนิโซแวน และมนุษย์ยุคใหม่อาจเคยอยู่ร่วมกันและผสมพันธุ์กันได้ การมีอยู่ของสายพันธุ์ที่ห่างไกลนี้ทำให้ภาพของมนุษยชาติในช่วงปลายยุคไพลสโตซีน มีความซับซ้อนมากขึ้น กว่าที่เคยคิดไว้[ 110 ] [ 113 ] นอกจากนี้ยังพบหลักฐานว่าดีเอ็นเอของชาว เมลานีเซียนยุคใหม่บางส่วนมากถึง 6% มาจากชาวเดนิโซแวน ซึ่งบ่งชี้ว่ามีการผสมพันธุ์กันอย่างจำกัดในเอเชียตะวันออกเฉียงใต้[ 114 ] [ 115 ]
อัลลีลที่เชื่อว่ามีต้นกำเนิดมาจากนีแอนเดอร์ทาลและเดนิโซแวนได้รับการระบุที่ตำแหน่งทางพันธุกรรมหลายแห่งในจีโนมของมนุษย์ยุคใหม่นอกทวีปแอฟริกาแฮพลอไทป์ของแอนติเจนเม็ดเลือดขาวของมนุษย์ (HLA) จากเดนิโซแวนและนีแอนเดอร์ทาลคิดเป็นมากกว่าครึ่งหนึ่งของอัลลีล HLA ของชาวเอเชียและยุโรปยุคใหม่[ 52 ]ซึ่งบ่งชี้ถึงการคัดเลือกเชิงบวกที่แข็งแกร่งสำหรับ อัลลีล ที่ถ่ายทอดเข้ามา เหล่านี้ คอรีน ซิโมเนติ จากมหาวิทยาลัยแวนเดอร์บิลต์ในแนชวิลล์และทีมของเธอพบจากบันทึกทางการแพทย์ของผู้คนเชื้อสายยุโรป 28,000 คนว่าการมีอยู่ของส่วนดีเอ็นเอของนีแอนเดอร์ทาลอาจเกี่ยวข้องกับอัตราการเกิดภาวะซึมเศร้าที่สูงขึ้น[ 116 ]
การถ่ายทอดยีนจากประชากรนีแอนเดอร์ทาลไปยังมนุษย์ยุคใหม่ไม่ได้เป็นไปในทิศทางเดียวเสมอไป เซอร์จิ คาสเตลลาโน จากสถาบันแม็กซ์พลังค์เพื่อมานุษยวิทยาเชิงวิวัฒนาการรายงานในปี 2016 ว่าในขณะที่จีโนมของเดนิโซแวนและนีแอนเดอร์ทาลมีความสัมพันธ์กันมากกว่าที่จะมีความสัมพันธ์กับเรา จีโนมของนีแอนเดอร์ทาลไซบีเรียกลับแสดงความคล้ายคลึงกับยีนของมนุษย์ยุคใหม่มากกว่าประชากรนีแอนเดอร์ทาลในยุโรป ซึ่งบ่งชี้ว่าประชากรนีแอนเดอร์ทาลได้ผสมพันธุ์กับมนุษย์ยุคใหม่เมื่อประมาณ 100,000 ปีที่แล้ว อาจจะในแถบตะวันออกใกล้[ 117 ]
การศึกษาเกี่ยวกับเด็กนีแอนเดอร์ทาลที่ยิบรอลตาร์แสดงให้เห็นจากพัฒนาการของสมองและการงอกของฟันว่าเด็กนีแอนเดอร์ทาลอาจเติบโตเร็วกว่าโฮโมเซเปียนส์[ 118 ]
เอช. ฟลอเรเซียนซิส

H. floresiensisซึ่งมีชีวิตอยู่ระหว่างประมาณ 190,000 ถึง 50,000 ปีก่อนปัจจุบัน (BP) ได้รับฉายาว่าฮอบบิทเนื่องจากขนาดที่เล็ก ซึ่งอาจเป็นผลมาจากภาวะแคระแกร็นบนเกาะ[ 119 ] H. floresiensisน่าสนใจทั้งในด้านขนาดและอายุ โดยเป็นตัวอย่างของสายพันธุ์ล่าสุดของสกุลHomoที่แสดงลักษณะเฉพาะที่แตกต่างจากมนุษย์ยุคใหม่ กล่าวอีกนัยหนึ่งH. floresiensisมีบรรพบุรุษร่วมกันกับมนุษย์ยุคใหม่ แต่แยกตัวออกจากสายพันธุ์มนุษย์ยุคใหม่และมีวิวัฒนาการที่แตกต่างออกไป การค้นพบหลักคือโครงกระดูกที่เชื่อว่าเป็นของผู้หญิงอายุประมาณ 30 ปี พบในปี 2003 และมีอายุประมาณ 18,000 ปี หญิงที่ยังมีชีวิตอยู่นั้นคาดว่ามีความสูงหนึ่งเมตร โดยมีปริมาตรสมองเพียง 380 cm³ (ถือว่าเล็กสำหรับชิมแปนซีและน้อยกว่าหนึ่งในสามของ ค่าเฉลี่ย ของมนุษย์ที่ 1400 cm³ ) [ 119 ]
อย่างไรก็ตาม มีการถกเถียงกันอย่างต่อเนื่องว่าH. floresiensisเป็นสายพันธุ์ที่แยกต่างหากจริงหรือไม่[ 120 ]นักวิทยาศาสตร์บางคนเชื่อว่าH. floresiensisเป็นH. sapiens ยุคใหม่ ที่มีภาวะแคระแกร็นผิดปกติ[ 121 ]สมมติฐานนี้ได้รับการสนับสนุนบางส่วน เนื่องจากมนุษย์ยุคใหม่บางคนที่อาศัยอยู่บน เกาะ ฟลอเรสซึ่งเป็นเกาะในอินโดนีเซียที่พบโครงกระดูกนั้น เป็นคนแคระซึ่งเมื่อรวมกับภาวะแคระแกร็นผิดปกติ อาจส่งผลให้มนุษย์มีขนาดเล็กกว่าปกติอย่างมาก ข้อโต้แย้งสำคัญอีกประการหนึ่งเกี่ยวกับH. floresiensisในฐานะสายพันธุ์ที่แยกต่างหากคือ โครงกระดูกนี้ถูกพบพร้อมกับเครื่องมือที่เกี่ยวข้องกับH. sapiensเท่านั้น[ 121 ]
อย่างไรก็ตาม สมมติฐานเรื่องภาวะแคระแกร็นจากพยาธิสภาพนั้นไม่สามารถอธิบายลักษณะทางกายวิภาค เพิ่มเติม ที่แตกต่างจากมนุษย์ยุคใหม่ (ไม่ว่าจะป่วยหรือไม่) แต่คล้ายคลึงกับสมาชิกโบราณของสกุลของเรา นอกเหนือจากลักษณะกะโหลกศีรษะแล้ว ลักษณะเหล่านี้ยังรวมถึงรูปร่างของกระดูกในข้อมือ แขนท่อนล่าง ไหล่ เข่า และเท้า นอกจากนี้ สมมติฐานนี้ยังไม่สามารถอธิบายการค้นพบตัวอย่างจำนวนมากของบุคคลที่มีลักษณะเดียวกันนี้ ซึ่งบ่งชี้ว่าลักษณะเหล่านี้พบได้ทั่วไปในประชากรจำนวนมาก และไม่ได้จำกัดอยู่เพียงบุคคลเดียว[ 120 ]
ในปี 2016 มีการค้นพบฟอสซิลฟันและขากรรไกรบางส่วนจากโฮมินินที่สันนิษฐานว่าเป็นบรรพบุรุษของH. floresiensis [ 122 ]ที่Mata Mengeซึ่งอยู่ห่างจาก Liang Bua ประมาณ 74 กม. (46 ไมล์) ฟอสซิลเหล่านี้มีอายุประมาณ 700,000 ปี[ 123 ]และนักโบราณคดีชาวออสเตรเลีย Gerrit van den Bergh ตั้งข้อสังเกตว่าฟอสซิลเหล่านี้มีขนาดเล็กกว่าฟอสซิลในยุคหลัง[ 124 ]
เอช. ลูโซเนนซิส
ตัวอย่างจำนวนเล็กน้อยจากเกาะลูซอนซึ่งมีอายุระหว่าง 50,000 ถึง 67,000 ปีที่ผ่านมา ได้รับการกำหนดโดยผู้ค้นพบเมื่อไม่นานมานี้ โดยพิจารณาจากลักษณะทางทันตกรรม ให้เป็นสายพันธุ์มนุษย์ใหม่H. luzonensis [ 125 ]
เอช.เซเปียนส์

H. sapiens (คำคุณศัพท์sapiensมาจากภาษาละติน แปลว่า "ฉลาด" หรือ "มีสติปัญญา") เกิดขึ้นในแอฟริกาเมื่อราว 300,000 ปีก่อน โดยน่าจะสืบเชื้อสายมาจากH. heidelbergensisหรือสายพันธุ์ที่เกี่ยวข้อง[ 126 ] [ 127 ]ในเดือนกันยายน พ.ศ. 2562 นักวิทยาศาสตร์ได้รายงานการกำหนดรูปร่างกะโหลกเสมือนจริงของบรรพบุรุษร่วมสุดท้ายของมนุษย์ยุคใหม่ ( H. sapiens ) โดยใช้คอมพิวเตอร์ โดยอิงจาก ภาพสแกน CT จำนวน 260 ภาพ ซึ่งเป็นตัวแทนของมนุษย์ยุคใหม่กลุ่มแรกสุด และเสนอแนะว่ามนุษย์ยุคใหม่ถือกำเนิดขึ้นระหว่าง 260,000 ถึง 350,000 ปีก่อน ผ่านการรวมตัวของประชากรใน แอฟริกา ตะวันออกและแอฟริกาใต้[ 128 ] [ 129 ]
ระหว่าง 400,000 ปีก่อนและช่วงระหว่างยุคน้ำแข็งครั้งที่สองในยุคไพลสโตซีนตอนกลางประมาณ 250,000 ปีก่อน แนวโน้มของการขยายปริมาตรภายในกะโหลกศีรษะและการพัฒนาเทคโนโลยีเครื่องมือหินได้พัฒนาขึ้น ซึ่งเป็นหลักฐานสำหรับการเปลี่ยนผ่านจากH. erectusไปเป็นH. sapiensหลักฐานโดยตรงชี้ให้เห็นว่ามีการอพยพของH. erectus ออกจากแอฟริกาจากนั้นมีการแยกสายพันธุ์ของH. sapiensออกจากH. erectusในแอฟริกา การอพยพครั้งต่อมา (ทั้งภายในและภายนอกแอฟริกา) ในที่สุดก็เข้ามาแทนที่H. erectus ที่กระจัดกระจายไปก่อนหน้านี้ ทฤษฎีการอพยพและต้นกำเนิดนี้มักถูกเรียกว่า "สมมติฐานต้นกำเนิดเดียวเมื่อไม่นานมานี้" หรือทฤษฎี "ออกจากแอฟริกา" H. sapiens ผสมพันธุ์กับมนุษย์โบราณทั้งในแอฟริกาและยูเรเซีย โดยเฉพาะในยูเรเซียกับนีแอนเดอร์ทาลและเดนิโซแวน[ 50 ] [ 114 ]
ทฤษฎีภัยพิบัติโทบาซึ่งตั้งสมมติฐานเกี่ยวกับคอขวดประชากรของH. sapiensเมื่อประมาณ 70,000 ปีก่อน[ 130 ]เป็นที่ถกเถียงกันตั้งแต่ข้อเสนอแรกในช่วงทศวรรษ 1990 และในช่วงทศวรรษ 2010 ก็แทบไม่มีการสนับสนุนเลย[ 131 ]ความแปรผันทางพันธุกรรมของมนุษย์ที่โดดเด่นเกิดขึ้นจากผลของผู้ก่อตั้งการผสมผสานแบบโบราณและ แรงกดดันทาง วิวัฒนาการ ล่าสุด
การเปลี่ยนแปลงทางกายวิภาค
นับตั้งแต่โฮโมเซเปียนส์แยกตัวออกจากบรรพบุรุษร่วมสุดท้ายที่ใช้ร่วมกับชิมแปนซีวิวัฒนาการของมนุษย์มีลักษณะเฉพาะด้วยการเปลี่ยนแปลงทางสัณฐานวิทยาการพัฒนาสรีรวิทยาพฤติกรรมและสิ่งแวดล้อม หลายประการ [ 10 ]วิวัฒนาการทางสิ่งแวดล้อม (วัฒนธรรม) ที่ค้นพบในภายหลังในช่วงไพลสโตซีนมีบทบาทสำคัญในวิวัฒนาการของมนุษย์ที่สังเกตได้จากการเปลี่ยนแปลงของมนุษย์ระหว่างระบบการดำรงชีพ[ 132 ] [ 10 ]การปรับตัวที่สำคัญที่สุดเหล่านี้ ได้แก่ การเดินสองขา ขนาดสมองที่ใหญ่ขึ้นการเจริญเติบโตที่ ยาวนานขึ้น (การตั้งครรภ์และวัยทารก) และความแตกต่างทางเพศ ที่ลดลง ความสัมพันธ์ระหว่างการเปลี่ยนแปลงเหล่านี้เป็นหัวข้อของการถกเถียงอย่างต่อเนื่อง[ 133 ]การเปลี่ยนแปลงทางสัณฐานวิทยาที่สำคัญอื่นๆ ได้แก่ วิวัฒนาการของการจับที่ทรงพลังและแม่นยำซึ่งเป็นการเปลี่ยนแปลงที่เกิดขึ้นครั้งแรกในH. erectus [ 134 ]
การเดินสองขา

การเดินสองขา ( Bipedalism ) เป็นการปรับตัวขั้นพื้นฐานของโฮมินิด และถือเป็นสาเหตุหลักที่อยู่เบื้องหลังการเปลี่ยนแปลงโครงกระดูกหลายอย่างที่พบในโฮมินิดที่เดินสองขาทั้งหมด บันทึกฟอสซิลเกี่ยวกับการเดินสองขาตั้งแต่ทศวรรษ 1990 ระบุว่าโฮมินินยุคแรกสุดถือกำเนิดขึ้นในช่วงปลายยุคไมโอซีน[ 135 ]โฮมินินยุคแรกสุดที่สันนิษฐานว่าเป็นการเดินสองขาแบบดั้งเดิมนั้น ถือว่าเป็นSahelanthropus [ 136 ]หรือOrrorinซึ่งทั้งสองสายพันธุ์ถือกำเนิดขึ้นเมื่อประมาณ 6 ถึง 7 ล้านปีก่อน สัตว์ที่เดินด้วยข้อเท้าและไม่เดินสองขา เช่นกอริลลาและชิมแปนซี แยกสายวิวัฒนาการออกจากโฮมินินในช่วงเวลาเดียวกัน ดังนั้นSahelanthropusหรือOrrorinอาจเป็นบรรพบุรุษร่วมสุดท้ายของเราArdipithecusซึ่งเป็นสัตว์ที่เดินสองขาอย่างสมบูรณ์ ถือกำเนิดขึ้นเมื่อประมาณ 5.6 ล้านปีก่อน[ 137 ]
สัตว์สองขาในยุคแรกวิวัฒนาการไปเป็นออสตราโลพิเทคัสและต่อมาก็วิวัฒนาการเป็นสกุลโฮโมมีทฤษฎีหลายอย่างเกี่ยวกับคุณค่าของการปรับตัวของการเดินสองขา เป็นไปได้ว่าการเดินสองขาเป็นที่นิยมเพราะทำให้มือว่างสำหรับการเอื้อมและถืออาหาร ประหยัดพลังงานระหว่างการเคลื่อนที่[ 138 ]ทำให้สามารถวิ่งและล่าสัตว์ในระยะทางไกลได้ ให้ขอบเขตการมองเห็นที่ดีขึ้น และช่วยหลีกเลี่ยงภาวะอุณหภูมิร่างกายสูงเกินไปโดยการลดพื้นที่ผิวที่สัมผัสกับแสงแดดโดยตรง คุณสมบัติเหล่านี้ล้วนเป็นประโยชน์ต่อการเจริญเติบโตในสภาพแวดล้อมทุ่งหญ้าสะวันนาและป่าไม้ใหม่ที่เกิดขึ้นจากการยกตัวของหุบเขาแอฟริกาตะวันออกเมื่อเทียบกับถิ่นที่อยู่อาศัยที่เป็นป่าทึบก่อนหน้านี้[ 138 ] [ 139 ] [ 140 ]การศึกษาในปี 2007 ให้การสนับสนุนสมมติฐานที่ว่าการเดินสองขาวิวัฒนาการขึ้นเพราะใช้พลังงานน้อยกว่าการเดินสี่ขาด้วยข้อเท้า[ 141 ] [ 142 ]อย่างไรก็ตาม การศึกษาล่าสุดชี้ให้เห็นว่าการเดินสองขาโดยปราศจากความสามารถในการใช้ไฟจะไม่สามารถทำให้เกิดการแพร่กระจายไปทั่วโลกได้[ 143 ]การเปลี่ยนแปลงในการเดินนี้ทำให้ขายาวขึ้นตามสัดส่วนเมื่อเทียบกับความยาวของแขน ซึ่งสั้นลงเนื่องจากไม่จำเป็นต้องใช้การโหนกิ่งไม้การเปลี่ยนแปลงอีกอย่างหนึ่งคือรูปร่างของนิ้วหัวแม่เท้า การศึกษาล่าสุดชี้ให้เห็นว่าออสตราโลพิเทคัสยังคงใช้ชีวิตอยู่บนต้นไม้บางส่วนอันเป็นผลมาจากการรักษานิ้วหัวแม่เท้าที่สามารถจับยึดได้ ซึ่งค่อยๆ หายไปในฮาบิไลน์
ในทางกายวิภาค วิวัฒนาการของการเดินสองขามาพร้อมกับการเปลี่ยนแปลงโครงกระดูกจำนวนมาก ไม่เพียงแต่ที่ขาและกระดูกเชิงกรานเท่านั้น แต่ยังรวมถึงกระดูกสันหลังเท้า ข้อเท้า และกะโหลกศีรษะ ด้วย [ 144 ]กระดูกต้นขาได้วิวัฒนาการไปอยู่ในตำแหน่งที่เอียงเล็กน้อยเพื่อย้ายจุดศูนย์ถ่วงไปทางจุดศูนย์กลางทางเรขาคณิตของร่างกาย ข้อเข่าและข้อเท้ามีความแข็งแรงมากขึ้นเพื่อรองรับน้ำหนักที่เพิ่มขึ้นได้ดีขึ้น เพื่อรองรับน้ำหนักที่เพิ่มขึ้นบนกระดูกสันหลังแต่ละข้อในท่าตั้งตรง กระดูกสันหลังของมนุษย์จึงมีรูปร่างเป็นตัว S และกระดูกสันหลังส่วนเอวก็ สั้นลงและกว้างขึ้น ที่เท้า นิ้วหัวแม่เท้าเคลื่อนไปอยู่ในแนวเดียวกับนิ้วเท้าอื่นๆ เพื่อช่วยในการเคลื่อนที่ไปข้างหน้า แขนและปลายแขนสั้นลงเมื่อเทียบกับขาทำให้วิ่งได้ง่ายขึ้น รูเปิดกะโหลกศีรษะเคลื่อนไปอยู่ใต้กะโหลกศีรษะและไปทางด้านหน้ามากขึ้น[ 145 ]
The most significant changes occurred in the pelvic region, where the long downward facing iliac blade was shortened and widened as a requirement for keeping the center of gravity stable while walking;[31] bipedal hominids have a shorter but broader, bowl-like pelvis due to this. A drawback is that the birth canal of bipedal apes is smaller than in knuckle-walking apes, though there has been a widening of it in comparison to that of australopithecine and modern humans, thus permitting the passage of newborns due to the increase in cranial size. This is limited to the upper portion, since further increase can hinder normal bipedal movement.[146]
The shortening of the pelvis and smaller birth canal evolved as a requirement for bipedalism and had significant effects on the process of human birth, which is much more difficult in modern humans than in other primates. During human birth, because of the variation in size of the pelvic region, the fetal head must be in a transverse position (compared to the mother) during entry into the birth canal and rotate about 90 degrees upon exit.[147] The smaller birth canal became a limiting factor to brain size increases in early humans and prompted a shorter gestation period leading to the relative immaturity of human offspring, who are unable to walk much before 12 months and have greater neoteny, compared to other primates, who are mobile at a much earlier age.[140] The increased brain growth after birth and the increased dependency of children on mothers had a major effect upon the female reproductive cycle,[148] and the more frequent appearance of alloparenting in humans when compared with other hominids.[149] Delayed human sexual maturity also led to the evolution of menopause with one explanation, the grandmother hypothesis, providing that elderly women could better pass on their genes by taking care of their daughter's offspring, as compared to having more children of their own.[150][151]
Encephalization


ในที่สุดเผ่าพันธุ์มนุษย์ก็พัฒนาสมองให้มีขนาดใหญ่กว่าไพรเมตอื่นๆ มาก โดยทั่วไปแล้วมนุษย์ยุคใหม่จะมีขนาด 1,330 cm³ (81 cu in) ซึ่งใหญ่กว่าสมองของชิมแปนซีหรือกอริลลาเกือบสามเท่า[ 154 ] หลังจากช่วงเวลาที่สมองหยุดนิ่งกับAustralopithecus anamensisและArdipithecusซึ่งเป็นสายพันธุ์ที่มีสมองขนาดเล็กกว่าอันเป็นผลมาจากการเดินสองขา[ 155 ]รูปแบบของการเพิ่มขนาดสมองก็เริ่มต้นขึ้นกับHomo habilisซึ่งมีสมองขนาด 600 cm³ ( 37 cu in) ใหญ่กว่าสมองของชิมแปนซีเล็กน้อย วิวัฒนาการนี้ยังคงดำเนินต่อไปในHomo erectusที่มีขนาด 800–1,100 cm³ ( 49–67 cu in) และถึงจุดสูงสุดในนีแอนเดอร์ทาลที่มีขนาด 1,200–1,900 cm³ ( 73–116 cu in) ซึ่งใหญ่กว่าHomo sapiens ยุคใหม่เสีย อีก การเพิ่มขนาดของสมองนี้เกิดขึ้นในช่วงการเจริญเติบโตของสมอง หลังคลอด ซึ่งเกินกว่าลิงชนิดอื่น ๆ มาก ( heterochrony ) นอกจากนี้ยังช่วยให้ มนุษย์วัยเด็ก สามารถเรียนรู้ทางสังคมและ เรียน รู้ภาษาได้ เป็นเวลานาน โดยเริ่มตั้งแต่เมื่อ 2 ล้านปีก่อน การเพิ่มขนาดของสมองอาจเกิดจากการพึ่งพาอาหารที่มีแคลอรีสูงและหาได้ยาก[ 156 ]
นอกจากนี้ การเปลี่ยนแปลงในโครงสร้างของสมองมนุษย์อาจมีความสำคัญมากกว่าการเพิ่มขนาดเสียอีก[ 157 ] [ 158 ] [ 159 ] [ 58 ]กะโหลกฟอสซิลแสดงให้เห็นว่าขนาดสมองของมนุษย์ยุคแรกอยู่ในช่วงเดียวกับมนุษย์ยุคปัจจุบันเมื่อ 300,000 ปีก่อน แต่มีรูปร่างสมองแบบปัจจุบันเมื่อประมาณ 100,000 ถึง 35,000 ปีก่อน[ 160 ]

กลีบขมับซึ่งมีศูนย์กลางสำหรับการประมวลผลภาษาได้เพิ่มขึ้นอย่างไม่สมส่วน เช่นเดียวกับคอร์เทกซ์ส่วนหน้าซึ่งเกี่ยวข้องกับการตัดสินใจที่ซับซ้อนและการควบคุมพฤติกรรมทางสังคม[ 154 ]การเพิ่มขนาดของสมองมีความเชื่อมโยงกับการเพิ่มขึ้นของแป้ง[ 57 ]และเนื้อสัตว์[ 161 ] [ 162 ]ในอาหาร อย่างไรก็ตาม การศึกษาแบบเมตาในปี 2022 ได้ตั้งคำถามถึงบทบาทของเนื้อสัตว์[ 163 ]ปัจจัยอื่นๆ ได้แก่ การพัฒนาการทำอาหาร[ 164 ]และมีการเสนอว่าสติปัญญาเพิ่มขึ้นเพื่อตอบสนองต่อความจำเป็นที่เพิ่มขึ้นในการแก้ปัญหาทางสังคมเมื่อสังคมมนุษย์มีความซับซ้อนมากขึ้น[ 165 ]การเปลี่ยนแปลงในสัณฐานวิทยาของกะโหลกศีรษะ เช่น ขากรรไกรล่างที่เล็กลงและการยึดติดของกล้ามเนื้อขากรรไกรล่าง ทำให้มีพื้นที่มากขึ้นสำหรับสมองที่จะเติบโต[ 166 ]
การเพิ่มขึ้นของปริมาตรของนีโอคอร์เทกซ์ยังรวมถึงการเพิ่มขนาดของซีรีเบลลัม อย่างรวดเร็ว ด้วย หน้าที่ของมันโดยทั่วไปเกี่ยวข้องกับการทรงตัวและการควบคุมการเคลื่อนไหวอย่างละเอียด แต่เมื่อไม่นานมานี้พบว่าเกี่ยวข้องกับการพูดและการรับรู้ลิงใหญ่ รวมถึงโฮมินิด มีซีรีเบลลัมที่เด่นชัดกว่าเมื่อเทียบกับนีโอคอร์เทกซ์มากกว่าไพรเมตชนิดอื่นๆ มีการเสนอแนะว่าเนื่องจากหน้าที่ในการควบคุมการรับรู้และการเคลื่อนไหว และการเรียนรู้การเคลื่อนไหวของกล้ามเนื้อที่ซับซ้อน ซีรีเบลลัมอาจเป็นพื้นฐานของการปรับตัวทางเทคโนโลยีของมนุษย์ รวมถึงเงื่อนไขเบื้องต้นของการพูด[ 167 ] [ 168 ] [ 169 ] [ 170 ]
เป็นการยากที่จะระบุข้อได้เปรียบในการอยู่รอดในทันทีของการเพิ่มขนาดสมอง เนื่องจากความเปลี่ยนแปลงของสมองที่สำคัญจากHomo erectusไปสู่Homo heidelbergensisไม่ได้มาพร้อมกับการเปลี่ยนแปลงทางเทคโนโลยีที่สำคัญ มีการเสนอแนะว่าการเปลี่ยนแปลงส่วนใหญ่เป็นด้านสังคมและพฤติกรรม รวมถึงความสามารถในการเห็นอกเห็นใจที่เพิ่มขึ้น[ 171 ] [ 172 ]การเพิ่มขนาดของกลุ่มสังคม[ 165 ] [ 173 ] [ 174 ]และความยืดหยุ่นทางพฤติกรรมที่เพิ่มขึ้น[ 175 ]มนุษย์มีความพิเศษในความสามารถในการรับข้อมูลผ่านการส่งต่อทางสังคมและปรับใช้ข้อมูลนั้น[ 176 ]สาขาใหม่ของการวิวัฒนาการทางวัฒนธรรมศึกษาการเปลี่ยนแปลงทางสังคมและวัฒนธรรมของมนุษย์จากมุมมองเชิงวิวัฒนาการ[ 177 ]

ความแตกต่างทางเพศ
การลดลงของความแตกต่างทางเพศในมนุษย์นั้นเห็นได้ชัดเจนจากการลดลงของฟันเขี้ยวของเพศ ชาย เมื่อเทียบกับลิงชนิดอื่น (ยกเว้นชะนี) และการลดลงของสันคิ้วและความแข็งแรงโดยทั่วไปของเพศชาย การเปลี่ยนแปลงทางสรีรวิทยาที่สำคัญอีกประการหนึ่งที่เกี่ยวข้องกับเพศในมนุษย์คือวิวัฒนาการของ ภาวะ เป็นสัดแบบซ่อนเร้นมนุษย์เป็นโฮมินอยด์เพียงกลุ่มเดียวที่เพศหญิงมีภาวะเจริญพันธุ์ตลอดทั้งปีและไม่มีสัญญาณพิเศษใด ๆ ของภาวะเจริญพันธุ์ที่ร่างกายสร้างขึ้น (เช่น การบวมของอวัยวะเพศหรือการเปลี่ยนแปลงที่ชัดเจนในความพร้อมในการผสมพันธุ์ในช่วงเป็นสัด) [ 190 ]
ถึงกระนั้น มนุษย์ยังคงรักษาความแตกต่างทางเพศในระดับหนึ่งไว้ได้ ทั้งในเรื่องการกระจายตัวของขนตามร่างกายและไขมันใต้ผิวหนัง รวมถึงขนาดโดยรวม โดยเพศชายมีขนาดใหญ่กว่าเพศหญิงประมาณ 15% [ 191 ]การเปลี่ยนแปลงเหล่านี้โดยรวมได้รับการตีความว่าเป็นผลมาจากการให้ความสำคัญกับการผูกพันเป็นคู่ มากขึ้น ซึ่งอาจเป็นทางออกสำหรับความต้องการการลงทุนจากพ่อแม่ที่เพิ่มขึ้นเนื่องจากช่วงวัยทารกของลูกนั้นยาวนาน[ 192 ]
ตรงข้ามกับกระดูกอัลนาร์

การสัมผัสระหว่างนิ้วหัวแม่มือและปลายนิ้วก้อยของมือข้างเดียวกัน (ulnar opposition) เป็นลักษณะเฉพาะของสกุลHomo [ 193 ]ซึ่งรวมถึงมนุษย์นีแอนเดอร์ทาล มนุษย์ ยุคใหม่จาก Sima de los Huesosและมนุษย์ยุคใหม่[ 194 ] [ 195 ]ในไพรเมตชนิดอื่น นิ้วหัวแม่มือจะสั้นและไม่สามารถแตะนิ้วก้อยได้[ 194 ] การสัมผัส ระหว่างนิ้วหัวแม่มือและปลายนิ้วก้อยช่วยให้มือมนุษย์สามารถจับได้อย่างแม่นยำและแข็งแรง ซึ่งเป็นพื้นฐานของการใช้มืออย่างชำนาญทุกรูปแบบ
การเปลี่ยนแปลงอื่นๆ
การเปลี่ยนแปลงอื่นๆ อีกหลายประการได้บ่งบอกถึงวิวัฒนาการของมนุษย์ รวมถึงการพึ่งพาการมองเห็นมากกว่าการดมกลิ่น ( หลอดรับกลิ่น ลดลงอย่างมาก ) ระยะเวลาพัฒนาการในวัยเด็กที่ยาวนานขึ้นและการพึ่งพาของทารกที่สูงขึ้น[ 196 ]ลำไส้ที่เล็กลงและฟันที่เล็กและเรียงตัวไม่ตรงกัน การเผาผลาญพื้นฐานที่เร็วขึ้น[ 197 ]การสูญเสียขนตามร่างกาย[ 198 ] ความหนาแน่น ของต่อมเหงื่อเอคครีน ที่ เพิ่ม ขึ้นถึงสิบเท่าเมื่อเทียบกับ ไพรเมตแคทารินอื่นๆ[ 199 ]แต่มนุษย์ใช้น้ำน้อยกว่าชิมแปนซีและกอริลลา 30% ถึง 50% ต่อวัน[ 200 ]การนอนหลับแบบ REMมากขึ้นแต่การนอนหลับโดยรวมน้อยลง[ 201 ]การเปลี่ยนแปลงรูปร่างของส่วนโค้งของฟันจากรูปตัวยูเป็นรูปพาราโบลา การพัฒนาของคาง (พบเฉพาะในโฮโมเซเปียนส์ ) กระบวนการสไตลอยด์และกล่องเสียง ที่ลงมาต่ำ ลง เมื่อมือและแขนของมนุษย์ปรับตัวให้เข้ากับการสร้างเครื่องมือและถูกใช้ในการปีนป่ายน้อยลง กระดูกสะบักก็เปลี่ยนแปลงไปด้วย ผลข้างเคียงคือทำให้บรรพบุรุษของมนุษย์สามารถขว้างสิ่งของด้วยแรง ความเร็ว และความแม่นยำที่มากขึ้น[ 202 ]
การใช้เครื่องมือ




การใช้เครื่องมือได้รับการตีความว่าเป็นสัญญาณของสติปัญญา และมีการตั้งทฤษฎีว่าการใช้เครื่องมืออาจกระตุ้นวิวัฒนาการของมนุษย์ในบางแง่มุม โดยเฉพาะอย่างยิ่งการขยายตัวอย่างต่อเนื่องของสมองมนุษย์[ 204 ]บรรพชีวินวิทยายังไม่สามารถอธิบายการขยายตัวของอวัยวะนี้ในช่วงหลายล้านปีได้ แม้ว่าจะใช้พลังงานสูงมากก็ตาม สมองของมนุษย์ยุคใหม่ใช้พลังงานโดยเฉลี่ยประมาณ 13 วัตต์ (260 กิโลแคลอรีต่อวัน) ซึ่งคิดเป็นหนึ่งในห้าของการใช้พลังงานขณะพักของร่างกาย[ 205 ]การใช้เครื่องมือที่เพิ่มขึ้นจะช่วยให้สามารถล่าสัตว์เพื่อบริโภคเนื้อสัตว์ที่มีพลังงานสูง และช่วยให้สามารถแปรรูปพืชที่มีพลังงานสูงได้มากขึ้น นักวิจัยจึงเสนอว่าโฮมินินยุคแรกจึงอยู่ภายใต้แรงกดดันทางวิวัฒนาการเพื่อเพิ่มความสามารถในการสร้างและใช้เครื่องมือ[ 206 ]
เป็นการยากที่จะระบุได้อย่างแน่ชัดว่ามนุษย์ยุคแรกเริ่มใช้เครื่องมือเมื่อใด เนื่องจากยิ่งเครื่องมือเหล่านี้มีความดั้งเดิมมากเท่าไร (เช่น หินที่มีขอบคม) ก็ยิ่งยากที่จะตัดสินว่าสิ่งเหล่านั้นเป็นวัตถุธรรมชาติหรือสิ่งประดิษฐ์ของมนุษย์[ 204 ]มีหลักฐานบางอย่างที่บ่งชี้ว่าออสตราโลพิเทคัส (4 ล้านปีก่อน) อาจใช้กระดูกที่แตกหักเป็นเครื่องมือแต่เรื่องนี้ยังเป็นที่ถกเถียงกันอยู่[ 207 ]
สิ่งมีชีวิตหลายชนิดสร้างและใช้เครื่องมือแต่สกุลมนุษย์เป็นสกุลที่ครองความเป็นใหญ่ในด้านการสร้างและใช้เครื่องมือที่ซับซ้อนกว่า เครื่องมือที่เก่าแก่ที่สุดที่รู้จักคือเศษหินจากเวสต์เทอร์คานา ประเทศเคนยา ซึ่งมีอายุย้อนไปถึง 3.3 ล้านปีก่อน[ 208 ]เครื่องมือหินที่เก่าแก่รองลงมามาจากโกนา ประเทศเอธิโอเปียและถือเป็นจุดเริ่มต้นของเทคโนโลยีโอลโดวัน เครื่องมือเหล่านี้มีอายุย้อนไปถึงประมาณ 2.6 ล้านปีก่อน[ 209 ]พบฟอสซิลโฮโมใกล้กับเครื่องมือโอลโดวันบางชิ้นและระบุอายุไว้ที่ 2.3 ล้านปี ซึ่งบ่งชี้ว่าบางที สายพันธุ์ โฮโมอาจสร้างและใช้เครื่องมือเหล่านี้จริง ๆ นี่เป็นความเป็นไปได้ แต่ยังไม่ใช่หลักฐานที่แน่ชัด[ 210 ]กระบวนการสไตลอยด์ของกระดูกฝ่ามือชิ้นที่สามช่วยให้กระดูกมือล็อกเข้ากับกระดูกข้อมือ ทำให้สามารถออกแรงกดที่ข้อมือและมือได้มากขึ้นจากนิ้วหัวแม่มือและนิ้วอื่น ๆ ที่จับสิ่งของได้ ซึ่งช่วยให้มนุษย์มีความคล่องแคล่วและแข็งแรงในการสร้างและใช้เครื่องมือที่ซับซ้อน ลักษณะทางกายวิภาคที่เป็นเอกลักษณ์นี้แยกมนุษย์ออกจากลิงและไพรเมตที่ไม่ใช่มนุษย์อื่นๆ และไม่พบในฟอสซิลมนุษย์ที่มีอายุมากกว่า 1.8 ล้านปี[ 211 ]
เบอร์นาร์ด วูด ตั้งข้อสังเกตว่าParanthropusอาศัยอยู่ร่วมกับ สายพันธุ์ Homo ยุคแรก ในบริเวณ "Oldowan Industrial Complex" ในช่วงเวลาเดียวกันโดยประมาณ แม้ว่าจะไม่มีหลักฐานโดยตรงที่ระบุว่าParanthropusเป็นผู้ผลิตเครื่องมือ แต่กายวิภาคของพวกเขาก็เป็นหลักฐานทางอ้อมที่บ่งชี้ถึงความสามารถของพวกเขาในด้านนี้ นักมานุษยวิทยาบรรพกาลส่วนใหญ่เห็นพ้องต้องกันว่า สายพันธุ์ Homo ยุคแรก เป็นผู้รับผิดชอบ เครื่องมือ Oldowanส่วนใหญ่ที่พบ พวกเขาโต้แย้งว่าเมื่อพบเครื่องมือ Oldowan ส่วนใหญ่ร่วมกับฟอสซิลมนุษย์Homoมักจะอยู่ด้วยเสมอ แต่Paranthropusไม่ได้อยู่ด้วย[ 210 ]
ในปี พ.ศ. 2537 แรนดัล ซัสแมน ใช้กายวิภาคของนิ้วหัวแม่มือที่สามารถงอได้เป็นพื้นฐานในการโต้แย้งว่าทั้ง สายพันธุ์ โฮโมและพารานโทรปัสต่างก็เป็นผู้สร้างเครื่องมือ เขาเปรียบเทียบกระดูกและกล้ามเนื้อของนิ้วหัวแม่มือของมนุษย์และชิมแปนซี พบว่ามนุษย์มีกล้ามเนื้อ 3 มัดที่ชิมแปนซีไม่มี นอกจากนี้ มนุษย์ยังมีกระดูกฝ่ามือที่หนากว่าและมีหัวที่กว้างกว่า ทำให้สามารถจับสิ่งของได้อย่างแม่นยำกว่ามือของชิมแปนซี ซัสแมนตั้งสมมติฐานว่ากายวิภาคของนิ้วหัวแม่มือที่สามารถงอได้ของมนุษย์ในปัจจุบันเป็นการตอบสนองเชิงวิวัฒนาการต่อความต้องการที่เกี่ยวข้องกับการสร้างและการจัดการเครื่องมือ และทั้งสองสายพันธุ์ต่างก็เป็นผู้สร้างเครื่องมือ[ 210 ]
การเปลี่ยนผ่านสู่ความทันสมัยเชิงพฤติกรรม
นักมานุษยวิทยาอธิบายพฤติกรรมของมนุษย์ยุคใหม่ว่ารวมถึงลักษณะทางวัฒนธรรมและพฤติกรรม เช่น ความเชี่ยวชาญด้านเครื่องมือ การใช้เครื่องประดับและรูปภาพ (เช่น ภาพวาดในถ้ำ) การจัดระเบียบพื้นที่อยู่อาศัย พิธีกรรม (เช่น ของขวัญในหลุมศพ) เทคนิคการล่าสัตว์เฉพาะทาง การสำรวจพื้นที่ทางภูมิศาสตร์ที่ไม่เอื้ออำนวย และ เครือข่ายการแลกเปลี่ยน สินค้าตลอดจนลักษณะทั่วไปอื่นๆ เช่น ภาษาและการคิดเชิงสัญลักษณ์ที่ซับซ้อน การถกเถียงยังคงดำเนินต่อไปว่า "การปฏิวัติ" นำไปสู่มนุษย์ยุคใหม่ ("บิ๊กแบงของจิตสำนึกของมนุษย์") หรือว่าวิวัฒนาการเป็นไปอย่างค่อยเป็นค่อยไปมากกว่า[ 54 ]
จนกระทั่งประมาณ 50,000–40,000 ปีที่แล้ว การใช้เครื่องมือหินดูเหมือนจะพัฒนาไปทีละขั้น แต่ละช่วง ( H. habilis , H. ergaster , H. neanderthalensis ) แสดงถึงเทคโนโลยีใหม่ ตามมาด้วยการพัฒนาอย่างช้าๆ จนกระทั่งถึงช่วงต่อไป ปัจจุบันนักมานุษยวิทยาโบราณกำลังถกเถียงกันว่า สายพันธุ์ Homo เหล่านี้ มีพฤติกรรมของมนุษย์ยุคใหม่บางส่วนหรือทั้งหมดหรือไม่ พวกเขาดูเหมือนจะอนุรักษ์นิยมทางวัฒนธรรม โดยรักษาเทคโนโลยีและรูปแบบการหาอาหารแบบเดิมไว้เป็นเวลานานมาก
เมื่อราว 50,000 ปีก่อนคริสตกาลวัฒนธรรมมนุษย์เริ่มพัฒนาอย่างรวดเร็ว การเปลี่ยนผ่านไปสู่ความทันสมัยทางพฤติกรรมนั้น บางคนเรียกว่า " การก้าวกระโดดครั้งใหญ่ " [ 212 ]หรือ "การปฏิวัติยุคหินเก่าตอนบน" [ 213 ]เนื่องจากการปรากฏตัวอย่างฉับพลันในบันทึกทางโบราณคดีของสัญญาณที่โดดเด่นของพฤติกรรมสมัยใหม่และการล่าสัตว์ใหญ่[ 214 ]หลักฐานของความทันสมัยทางพฤติกรรมที่เก่าแก่กว่านั้นก็มีอยู่ในแอฟริกาเช่นกัน โดยมีหลักฐานที่เก่ากว่าของภาพนามธรรม กลยุทธ์การดำรงชีพที่กว้างขึ้น เครื่องมือและอาวุธที่ซับซ้อนมากขึ้น และพฤติกรรม "สมัยใหม่" อื่นๆ และนักวิชาการหลายคนได้โต้แย้งเมื่อเร็วๆ นี้ว่าการเปลี่ยนผ่านไปสู่ความทันสมัยเกิดขึ้นเร็วกว่าที่เคยเชื่อกัน[ 54 ] [ 215 ] [ 216 ] [ 217 ]
นักวิชาการคนอื่นๆ พิจารณาว่าการเปลี่ยนแปลงนั้นค่อยเป็นค่อยไปมากกว่า โดยสังเกตว่าลักษณะบางอย่างได้ปรากฏขึ้นแล้วในกลุ่มโฮโมเซเปียนส์ โบราณในแอฟริกา เมื่อ 300,000–200,000 ปีก่อน[ 218 ] [ 219 ] [ 220 ] [ 221 ] [ 222 ]หลักฐานล่าสุดชี้ให้เห็นว่าประชากรอะบอริจินออสเตรเลียแยกตัวออกจากประชากรแอฟริกาเมื่อ 75,000 ปีก่อน และพวกเขาเดินทางทางทะเลเป็นระยะทาง 160 กิโลเมตร (99 ไมล์) เมื่อ 60,000 ปีก่อน ซึ่งอาจลดความสำคัญของการปฏิวัติยุคหินเก่าตอนบนลง[ 223 ]
มนุษย์ยุคใหม่เริ่มฝังศพผู้ตาย ทำเสื้อผ้าจากหนังสัตว์ ล่าสัตว์ด้วยเทคนิคที่ซับซ้อนมากขึ้น (เช่น การใช้กับดักหลุมหรือการไล่สัตว์ลงจากหน้าผา) และวาดภาพในถ้ำ[ 224 ]เมื่อวัฒนธรรมมนุษย์ก้าวหน้าขึ้น ประชากรกลุ่มต่างๆ ได้คิดค้นเทคโนโลยีที่มีอยู่แล้วขึ้นมาใหม่ สิ่งประดิษฐ์ต่างๆ เช่น เบ็ดตกปลา กระดุม และเข็มกระดูก แสดงให้เห็นถึงความหลากหลายทางวัฒนธรรม ซึ่งไม่เคยพบเห็นมาก่อน 50,000 ปีก่อน โดยทั่วไปแล้ว ประชากร H. neanderthalensis รุ่นเก่า จะไม่มีความหลากหลายในด้านเทคโนโลยี แม้ว่า กลุ่มสิ่งประดิษฐ์ Chatelperronianจะถูกพบว่าเป็นการเลียนแบบเทคโนโลยีAurignacian ของ H. sapiens โดยมนุษย์นีแอนเดอร์ทัลก็ตาม [ 225 ]
วิวัฒนาการของมนุษย์ในปัจจุบันและที่กำลังดำเนินอยู่
ประชากร มนุษย์ยุคใหม่ทางกายวิภาคยังคงวิวัฒนาการต่อไป เนื่องจากได้รับผลกระทบจากการคัดเลือกโดยธรรมชาติและการเปลี่ยนแปลงทางพันธุกรรมแม้ว่าแรงกดดันในการคัดเลือกในบางลักษณะ เช่น ความต้านทานต่อโรคฝีดาษ จะลดลงในยุคปัจจุบัน แต่มนุษย์ก็ยังคงอยู่ภายใต้การคัดเลือกโดยธรรมชาติสำหรับลักษณะอื่นๆ อีกมากมาย บางส่วนเกิดจากแรงกดดันด้านสิ่งแวดล้อมที่เฉพาะเจาะจง ในขณะที่บางส่วนเกี่ยวข้องกับการเปลี่ยนแปลงวิถีชีวิตนับตั้งแต่การพัฒนาการเกษตร (10,000 ปีที่แล้ว) การขยายตัวของเมือง (5,000 ปีที่แล้ว) และการพัฒนาอุตสาหกรรม (250 ปีที่แล้ว) มีการโต้แย้งว่าวิวัฒนาการของมนุษย์ได้เร่งตัวขึ้นนับตั้งแต่การพัฒนาการเกษตรเมื่อ 10,000 ปีที่แล้วและอารยธรรมเมื่อประมาณ 5,000 ปีที่แล้ว ส่งผลให้เกิดความแตกต่างทางพันธุกรรมอย่างมากระหว่างประชากรมนุษย์ในปัจจุบันที่แตกต่างกัน[ 226 ]และงานวิจัยล่าสุดระบุว่าสำหรับบางลักษณะ การพัฒนาและนวัตกรรมของวัฒนธรรมมนุษย์ได้ผลักดันรูปแบบใหม่ของการคัดเลือกที่อยู่ร่วมกับ และในบางกรณีได้เข้ามาแทนที่การคัดเลือกโดยธรรมชาติเป็นส่วนใหญ่[ 227 ]

สิ่งที่เห็นได้ชัดเจนเป็นพิเศษคือความแตกต่างในลักษณะภายนอก เช่นผมหยิกแบบแอฟริกาหรือวิวัฒนาการล่าสุดของผิวขาวและผมสีบลอนด์ในบางประชากร ซึ่งเป็นผลมาจากความแตกต่างของสภาพภูมิอากาศ แรงกดดันจากการคัดเลือกที่รุนแรงเป็นพิเศษส่งผลให้มนุษย์ปรับตัวเข้ากับที่สูงได้โดยมีความแตกต่างกันในแต่ละประชากรที่แยกตัวออกมา การศึกษาพื้นฐานทางพันธุกรรมแสดงให้เห็นว่าบางอย่างพัฒนาขึ้นเมื่อไม่นานมานี้ โดยชาวทิเบตมีวิวัฒนาการมาตลอด 3,000 ปีจนมีสัดส่วนของอัลลีลEPAS1ที่ปรับตัวเข้ากับที่สูงได้สูง
วิวัฒนาการอื่นๆ เกี่ยวข้องกับโรคประจำถิ่น : การมีอยู่ของมาลาเรียทำให้เกิดการคัดเลือกคุณลักษณะเซลล์เคียว ( รูปแบบ เฮเทอโรไซกัสของยีนเซลล์เคียว) ในขณะที่หากไม่มีมาลาเรีย ผลกระทบต่อสุขภาพของโรคโลหิตจางเซลล์เคียวจะคัดเลือกต่อต้านคุณลักษณะนี้ ตัวอย่างเช่น ประชากรที่มีความเสี่ยงต่อโรคคุรุซึ่ง เป็นโรคร้ายแรงที่ทำให้ ร่างกายอ่อนแอ มีการแสดงออกของ ยีน โปรตีนพรีออน G127V ที่เป็นตัวแปรภูมิคุ้มกันมากกว่าอัลลีลที่ไม่สร้างภูมิคุ้มกันอย่างมีนัยสำคัญ ความถี่ของตัวแปรทางพันธุกรรม นี้ เกิดจากการอยู่รอดของบุคคลที่มีภูมิคุ้มกัน[ 229 ] [ 230 ]แนวโน้มที่รายงานบางอย่างยังคงไม่สามารถอธิบายได้และเป็นหัวข้อของการวิจัยอย่างต่อเนื่องในสาขาใหม่ของเวชศาสตร์วิวัฒนาการ: กลุ่มอาการรังไข่หลายถุง (PCOS) ลดความสามารถในการเจริญพันธุ์ ดังนั้นจึงคาดว่าจะอยู่ภายใต้การคัดเลือกเชิงลบที่รุนแรงมาก แต่ความแพร่หลายของโรคนี้ในประชากรมนุษย์บ่งชี้ถึงแรงกดดันการคัดเลือกที่ต่อต้าน ตัวตนของแรงกดดันนั้นยังคงเป็นหัวข้อของการถกเถียงกันอยู่[ 231 ]
วิวัฒนาการของมนุษย์ที่เกี่ยวข้องกับการเกษตรในช่วงไม่นานมานี้รวมถึงความต้านทานทางพันธุกรรมต่อโรคติดเชื้อที่ปรากฏในประชากรมนุษย์โดยการข้ามกำแพงสายพันธุ์จากสัตว์เลี้ยง[ 232 ]เช่นเดียวกับการเปลี่ยนแปลงในการเผาผลาญเนื่องจากการเปลี่ยนแปลงในอาหาร เช่นการคงอยู่ของแลคเตส
วิวัฒนาการที่ขับเคลื่อนด้วยวัฒนธรรมสามารถท้าทายความคาดหวังของการคัดเลือกโดยธรรมชาติได้: ในขณะที่ประชากรมนุษย์ประสบกับแรงกดดันบางอย่างที่ผลักดันให้เกิดการคัดเลือกเพื่อผลิตบุตรในวัยที่อายุน้อยลง การเกิดขึ้นของการคุมกำเนิดที่มีประสิทธิภาพ การศึกษาที่สูงขึ้น และบรรทัดฐานทางสังคมที่เปลี่ยนแปลงไปได้ผลักดันการคัดเลือกที่สังเกตได้ในทิศทางตรงกันข้าม[ 233 ]อย่างไรก็ตาม การคัดเลือกที่ขับเคลื่อนด้วยวัฒนธรรมไม่จำเป็นต้องทำงานขัดแย้งหรือต่อต้านการคัดเลือกโดยธรรมชาติเสมอไป: ข้อเสนอบางประการเพื่ออธิบายอัตราการขยายตัวของสมองมนุษย์ในปัจจุบันที่สูง บ่งชี้ถึงการป้อนกลับชนิดหนึ่ง ซึ่งประสิทธิภาพการเรียนรู้ทางสังคมที่เพิ่มขึ้นของสมองจะส่งเสริมการพัฒนาทางวัฒนธรรม ซึ่งในทางกลับกันจะส่งเสริมประสิทธิภาพที่มากขึ้น ซึ่งผลักดันการพัฒนาทางวัฒนธรรมที่ซับซ้อนมากขึ้นซึ่งต้องการประสิทธิภาพที่มากขึ้นไปอีก และเป็นเช่นนี้เรื่อยไป[ 234 ]วิวัฒนาการที่ขับเคลื่อนด้วยวัฒนธรรมมีข้อดีตรงที่นอกเหนือจากผลกระทบทางพันธุกรรมแล้ว ยังสามารถสังเกตได้จากบันทึกทางโบราณคดีด้วย: การพัฒนาเครื่องมือหินตลอดช่วงยุคหินเก่าเชื่อมโยงกับการพัฒนาทางปัญญาที่ขับเคลื่อนด้วยวัฒนธรรมในรูปแบบของการได้มาซึ่งทักษะที่ได้รับการสนับสนุนจากวัฒนธรรมและการพัฒนาเทคโนโลยีที่ซับซ้อนมากขึ้นเรื่อยๆ และความสามารถทางปัญญาในการสร้างสรรค์สิ่งเหล่านั้น[ 235 ]
ในยุคปัจจุบัน นับตั้งแต่มีการพัฒนาอุตสาหกรรม มีแนวโน้มบางประการที่สังเกตได้ เช่น ภาวะหมดประจำเดือนเกิดขึ้นช้าลง[ 236 ]แนวโน้มอื่นๆ ที่รายงานดูเหมือนจะรวมถึงระยะเวลาการสืบพันธุ์ของมนุษย์ที่ยาวนานขึ้น และการลดลงของระดับคอเลสเตอรอล น้ำตาลในเลือด และความดันโลหิตในประชากรบางกลุ่ม[ 236 ]
ประวัติการศึกษา
ก่อนยุคของดาร์วิน
ชื่อHomoของสกุลทางชีววิทยาที่มนุษย์เป็นสมาชิกอยู่นั้นมา จาก ภาษาละตินแปลว่า 'มนุษย์' [ e ]เดิมทีคาร์ล ลินเนียส เป็นผู้เลือกชื่อนี้ ในระบบการจำแนกประเภทของเขา[ f ]คำว่าhuman ในภาษาอังกฤษ มาจากhumanus ในภาษาละติน ซึ่งเป็นรูปคำคุณศัพท์ของhomo คำว่า homoในภาษาละตินมาจากรากศัพท์อินโด-ยุโรป* dhghemหรือ 'โลก' [ 237 ]ลินเนียสและนักวิทยาศาสตร์คนอื่นๆ ในสมัยของเขายังถือว่าลิงใหญ่เป็นญาติใกล้ชิดที่สุดของมนุษย์โดยพิจารณาจากความคล้ายคลึงกันทางสัณฐานวิทยาและกายวิภาคศาสตร์[ 238 ]
ดาร์วิน
ความเป็นไปได้ในการเชื่อมโยงมนุษย์กับลิงยุคก่อนโดยสืบเชื้อสายนั้นชัดเจนขึ้นหลังจากปี 1859 เมื่อชาร์ลส์ ดาร์วินตี พิมพ์หนังสือ On the Origin of Speciesซึ่งเขาได้โต้แย้งถึงแนวคิดเรื่องวิวัฒนาการของสายพันธุ์ใหม่จากสายพันธุ์ก่อนหน้า หนังสือของดาร์วินไม่ได้กล่าวถึงคำถามเรื่องวิวัฒนาการของมนุษย์ โดยกล่าวเพียงว่า "ความกระจ่างจะถูกเปิดเผยเกี่ยวกับต้นกำเนิดของมนุษย์และประวัติศาสตร์ของเขา" [ 239 ]
การถกเถียงครั้งแรกเกี่ยวกับธรรมชาติของวิวัฒนาการของมนุษย์เกิดขึ้นระหว่างโทมัส เฮนรี ฮักซ์ลีย์และริชาร์ด โอเวน ฮัก ซ์ลีย์โต้แย้งว่ามนุษย์มีวิวัฒนาการมาจากลิง โดยยกตัวอย่างความคล้ายคลึงและความแตกต่างระหว่างมนุษย์กับลิงชนิดอื่นๆ โดยเฉพาะอย่างยิ่งในหนังสือEvidence as to Man's Place in Nature ที่ตีพิมพ์ในปี 1863 ผู้สนับสนุนยุคแรกของดาร์วินหลายคน (เช่นอัลเฟรด รัสเซล วอลเลซและชาร์ลส์ ไลเอล ) ในตอนแรกไม่เห็นด้วยว่าต้นกำเนิดของความสามารถทางจิตและความรู้สึกทางศีลธรรมของมนุษย์สามารถอธิบายได้ด้วยการคัดเลือกโดยธรรมชาติแม้ว่าต่อมาจะมีการเปลี่ยนแปลง ดาร์วินนำทฤษฎีวิวัฒนาการและการคัดเลือกทางเพศมาใช้กับมนุษย์ในหนังสือThe Descent of Man, and Selection in Relation to Sex ที่ตีพิมพ์ในปี 1871 [ 240 ]
ฟอสซิลชิ้นแรก
ปัญหาสำคัญในศตวรรษที่ 19 คือการขาดแคลนฟอสซิลตัวกลาง ซากของมนุษย์นีแอนเดอร์ทาลถูกค้นพบในเหมืองหินปูนในปี 1856 สามปีก่อนการตีพิมพ์หนังสือOn the Origin of Speciesและฟอสซิลของมนุษย์นีแอนเดอร์ทาลก็ถูกค้นพบในยิบรอลตาร์ก่อนหน้านั้น แต่เดิมมีการอ้างว่าซากเหล่านั้นเป็นซากของมนุษย์ยุคใหม่ที่ป่วยด้วยโรคบางชนิด[ 241 ]แม้ว่าEugène Dubois จะค้นพบ สิ่งที่ปัจจุบันเรียกว่าHomo erectusที่Trinilเกาะชวา ในปี 1891 แต่ก็ต้องรอจนกระทั่งปี 1920 เมื่อมีการค้นพบฟอสซิลดังกล่าวในแอฟริกา ทำให้เริ่มมีการสะสมสายพันธุ์ตัวกลางมากขึ้น[ 242 ]ในปี 1925 Raymond Dartได้บรรยายถึงAustralopithecus africanus [ 243 ] ตัวอย่างต้นแบบคือ Taung Childซึ่งเป็นทารก Australopithecine ที่ถูกค้นพบในถ้ำ ซากศพของเด็กที่พบนั้นประกอบด้วยกะโหลกศีรษะขนาดเล็กที่ได้รับการอนุรักษ์ไว้อย่างดีเยี่ยม และแบบ จำลอง สมอง ที่หล่อขึ้น ภายใน
แม้ว่าสมองจะมีขนาดเล็ก (410 ลูกบาศก์เซนติเมตร)แต่รูปร่างของมันกลมมน ซึ่งแตกต่างจากสมองของชิมแปนซีและกอริลลา และคล้ายกับสมองของมนุษย์ยุคใหม่มากกว่า นอกจากนี้ ตัวอย่างยังแสดงให้เห็นฟันเขี้ยวที่ สั้น และตำแหน่งของช่องเปิดกะโหลกศีรษะ (รูในกะโหลกศีรษะที่กระดูกสันหลังเข้าไป) เป็นหลักฐานของ การเดิน สองขาลักษณะทั้งหมดเหล่านี้ทำให้ดาร์ทเชื่อว่าเด็กทาอุงเป็นบรรพบุรุษของมนุษย์ที่เดินสองขา เป็นรูปแบบการเปลี่ยนผ่านระหว่างลิงและมนุษย์
ฟอสซิลแอฟริกาตะวันออก

ในช่วงทศวรรษ 1960 และ 1970 มีการค้นพบฟอสซิลหลายร้อยชิ้นในแอฟริกาตะวันออก ในบริเวณหุบเขาโอลดูไวและทะเลสาบเทอร์คานาการค้นหาเหล่านี้ดำเนินการโดยครอบครัวลีคีย์ โดยมีหลุยส์ ลีคีย์ และ แมรี ลีคีย์ภรรยาของเขาและต่อมาริชาร์ดลูกชาย และเมฟ ลูกสะใภ้ซึ่งเป็นนักล่าฟอสซิลและนักมานุษยวิทยาบรรพกาล ได้เข้าร่วมด้วย จากแหล่งฟอสซิลของโอลดูไวและทะเลสาบเทอร์คานา พวกเขาได้รวบรวมตัวอย่างของมนุษย์ยุคแรกเริ่ม ได้แก่ ออสตราโลพิเทคัสและ สายพันธุ์ โฮโมและแม้กระทั่งโฮโมอิเร็ก ตั ส
การค้นพบเหล่านี้ตอกย้ำว่าแอฟริกาเป็นแหล่งกำเนิดของมนุษยชาติ ในช่วงปลายทศวรรษ 1970 และทศวรรษ 1980 เอธิโอเปียกลายเป็นจุดสนใจใหม่ของมานุษยวิทยาบรรพกาลหลังจากที่"ลูซี่"ฟอสซิลที่สมบูรณ์ที่สุดของสายพันธุ์Australopithecus afarensisถูกค้นพบในปี 1974 โดยDonald Johansonใกล้กับHadarใน ภูมิภาค สามเหลี่ยม Afar ที่เป็น ทะเลทราย ทางตอนเหนือของเอธิโอเปีย แม้ว่าตัวอย่างจะมีสมองขนาดเล็ก แต่กระดูกเชิงกรานและกระดูกขาเกือบจะเหมือนกับของมนุษย์ยุคใหม่ แสดงให้เห็นอย่างแน่นอนว่าโฮมินินเหล่านี้เดินตัวตรง[ 244 ]ลูซี่ถูกจัดเป็นสายพันธุ์ใหม่Australopithecus afarensisซึ่งเชื่อว่ามีความสัมพันธ์ใกล้ชิดกับสกุลHomoในฐานะบรรพบุรุษโดยตรง หรือเป็นญาติใกล้ชิดของบรรพบุรุษที่ไม่รู้จัก มากกว่าโฮมินิดหรือโฮมินินอื่น ๆ ที่รู้จักในช่วงเวลาแรกเริ่มนี้[ 245 ] (ตัวอย่างนี้ได้รับฉายาว่า "ลูซี่" ตามชื่อเพลง " Lucy in the Sky with Diamonds " ของ วงเดอะบีทเทิลส์ซึ่งถูกเปิดเสียงดังและซ้ำๆ ในค่ายระหว่างการขุดค้น) [ 246 ] ต่อมา พื้นที่สามเหลี่ยมอาฟาร์ ได้ค้นพบฟอสซิลโฮมินินอีกมากมาย โดยเฉพาะอย่างยิ่งฟอสซิลที่ทีมของ ทิม ดี. ไวท์ค้นพบหรือบรรยายไว้ในช่วงทศวรรษ 1990 ซึ่งรวมถึงArdipithecus ramidusและA. kadabba [ 247 ]
ในปี 2013 โครงกระดูกฟอสซิลของHomo nalediซึ่งเป็นสายพันธุ์โฮมินินที่สูญพันธุ์ไปแล้วซึ่งจัดอยู่ในสกุลHomo (อย่างไม่เป็นทางการ) ถูกค้นพบใน ระบบ ถ้ำ Rising Star ซึ่งเป็นแหล่งโบราณคดีในภูมิภาค Cradle of Humankindของแอฟริกาใต้ในจังหวัดGauteng ใกล้กับ โจฮันเนสเบิร์ก [ 248 ] [ 249 ] ณเดือนกันยายน 2015 มีการขุดค้นฟอสซิลของบุคคลอย่างน้อยสิบห้าคน รวมเป็นจำนวน 1,550 ตัวอย่างจากถ้ำ[ 249 ]สายพันธุ์นี้มีลักษณะเด่นคือมวลร่างกายและความสูงคล้ายกับประชากรมนุษย์ที่มีขนาดเล็ก ปริมาตร กะโหลกศีรษะภายใน ที่เล็กกว่า คล้ายกับAustralopithecusและสัณฐานวิทยาของกะโหลกศีรษะ (รูปทรงกะโหลก) คล้ายกับ สายพันธุ์ Homo ยุคแรก กายวิภาคของโครงกระดูกผสมผสานลักษณะดั้งเดิมที่รู้จักจากAustralopithecinesกับลักษณะที่รู้จักจากโฮมินินยุคแรก บุคคลเหล่านี้แสดงให้เห็นถึงร่องรอยของการถูกทิ้งอย่างจงใจภายในถ้ำในช่วงเวลาใกล้เสียชีวิต ฟอสซิลเหล่านี้มีอายุราว 250,000 ปีที่แล้ว[ 250 ]ดังนั้นจึงไม่ใช่บรรพบุรุษแต่ร่วมสมัยกับการปรากฏตัวครั้งแรกของมนุษย์ยุคใหม่ที่มี สมองขนาดใหญ่ขึ้น [ 251 ]
การปฏิวัติทางพันธุกรรม
การปฏิวัติทางพันธุกรรมในการศึกษาวิวัฒนาการของมนุษย์เริ่มต้นขึ้นเมื่อVincent SarichและAllan Wilsonวัดความแรงของปฏิกิริยาข้ามภูมิคุ้มกันของอัลบูมินในซีรั่มเลือด ระหว่างสิ่งมีชีวิตสองคู่ รวมถึงมนุษย์และลิงแอฟริกัน (ชิมแปนซีและกอริลลา) [ 252 ]ความแรงของปฏิกิริยาสามารถแสดงออกมาในเชิงตัวเลขเป็นระยะทางภูมิคุ้มกัน ซึ่งเป็นสัดส่วนกับจำนวนความแตกต่างของกรดอะมิโนระหว่างโปรตีนที่เหมือนกันในสายพันธุ์ต่างๆ โดยการสร้างเส้นโค้งการสอบเทียบของ ID ของคู่สายพันธุ์ที่มีเวลาการแยกสายพันธุ์ที่ทราบในบันทึกฟอสซิล ข้อมูลสามารถใช้เป็นนาฬิกาโมเลกุลเพื่อประมาณเวลาการแยกสายพันธุ์ของคู่สายพันธุ์ที่มีบันทึกฟอสซิลที่ไม่สมบูรณ์หรือไม่ทราบ
ในบทความสำคัญของพวกเขาในปี 1967 ในวารสารScience Sarich และ Wilson ได้ประมาณเวลาการแยกสายพันธุ์ของมนุษย์และลิงไว้ที่สี่ถึงห้าล้านปีที่แล้ว[ 252 ]ในขณะที่การตีความมาตรฐานของบันทึกฟอสซิลระบุว่าการแยกสายพันธุ์นี้เกิดขึ้นเมื่ออย่างน้อย 10 ถึง 30 ล้านปีที่แล้ว การค้นพบฟอสซิลในภายหลัง โดยเฉพาะอย่างยิ่ง "Lucy" และการตีความใหม่ของวัสดุฟอสซิลที่เก่ากว่า โดยเฉพาะอย่างยิ่งRamapithecusแสดงให้เห็นว่าการประมาณอายุที่น้อยกว่านั้นถูกต้องและยืนยันวิธีการอัลบูมิน
ความก้าวหน้าในการจัดลำดับดีเอ็นเอโดยเฉพาะดีเอ็นเอไมโทคอนเดรีย (mtDNA) และดีเอ็นเอโครโมโซม Y (Y-DNA) ทำให้เข้าใจต้นกำเนิดของมนุษย์ได้ดียิ่งขึ้น[ 139 ] [ 253 ] [ 254 ]การประยุกต์ใช้ หลักการ นาฬิกาโมเลกุลได้ ปฏิวัติการศึกษาวิวัฒนาการระดับโมเลกุล
จากการแยกตัวของมนุษย์จากอุรังอุตังเมื่อประมาณ 10 ถึง 20 ล้านปีก่อน การศึกษาในอดีตโดยใช้แบบจำลองนาฬิกาโมเลกุลชี้ให้เห็นว่ามีการกลายพันธุ์ประมาณ 76 ครั้งต่อรุ่นที่ไม่ได้รับการถ่ายทอดจากพ่อแม่ไปยังลูกหลานของมนุษย์ หลักฐานนี้สนับสนุนช่วงเวลาการแยกตัวระหว่างโฮมินินและชิมแปนซีที่กล่าวไว้ข้างต้น อย่างไรก็ตาม การศึกษาในปี 2012 ในไอซ์แลนด์เกี่ยวกับเด็ก 78 คนและพ่อแม่ของพวกเขาชี้ให้เห็นอัตราการกลายพันธุ์เพียง 36 ครั้งต่อรุ่น ข้อมูลนี้ขยายช่วงเวลาการแยกตัวระหว่างมนุษย์และชิมแปนซีไปสู่ช่วงเวลาที่เร็วกว่า 7 ล้านปีก่อน ( Ma ) การวิจัยเพิ่มเติมกับลูกหลานของประชากรชิมแปนซีป่า 226 ตัวในแปดสถานที่ชี้ให้เห็นว่าชิมแปนซีสืบพันธุ์เมื่ออายุเฉลี่ย 26.5 ปี ซึ่งบ่งชี้ว่าการแยกตัวของมนุษย์จากชิมแปนซีเกิดขึ้นระหว่าง 7 ถึง 13 ล้านปีก่อน และข้อมูลเหล่านี้ชี้ให้เห็นว่าArdipithecus (4.5 ล้านปี), Orrorin (6 ล้านปี) และSahelanthropus (7 ล้านปี) อาจอยู่ในสายพันธุ์ โฮมินิด และอาจเป็นไปได้ว่าการแยกสายพันธุ์เกิดขึ้นนอกภูมิภาค รอยแยกแอฟริกาตะวันออก
นอกจากนี้ การวิเคราะห์ยีนของทั้งสองสายพันธุ์ในปี 2549 ยังให้หลักฐานว่า แม้บรรพบุรุษของมนุษย์จะเริ่มแยกตัวออกจากชิมแปนซีแล้ว การผสมพันธุ์ข้ามสายพันธุ์ระหว่าง "มนุษย์ยุคดึกดำบรรพ์" และ "ชิมแปนซียุคดึกดำบรรพ์" ก็ยังคงเกิดขึ้นอย่างสม่ำเสมอเพียงพอที่จะเปลี่ยนแปลงยีนบางส่วนในกลุ่มยีน ใหม่ ได้
- การเปรียบเทียบจีโนมของมนุษย์และชิมแปนซีครั้งใหม่ชี้ให้เห็นว่า หลังจากที่ทั้งสองสายพันธุ์แยกจากกันแล้ว พวกมันอาจเริ่มผสมพันธุ์กัน... ข้อค้นพบหลักคือโครโมโซม Xของมนุษย์และชิมแปนซีดูเหมือนจะแยกสายพันธุ์กันเมื่อประมาณ 1.2 ล้านปีก่อน เมื่อเทียบกับโครโมโซมอื่นๆ
ผลการวิจัยชี้ให้เห็นว่า:
- ในความเป็นจริงแล้วมีการแยกสายพันธุ์ระหว่างมนุษย์และชิมแปนซีสองครั้ง โดยครั้งแรกตามมาด้วยการผสมพันธุ์ระหว่างประชากรทั้งสอง และจากนั้นก็มีการแยกสายพันธุ์ครั้งที่สอง ข้อเสนอแนะเกี่ยวกับการผสมข้ามสายพันธุ์ทำให้บรรดานักมานุษยวิทยาโบราณตกใจ แต่พวกเขาก็ยังคงให้ความสำคัญกับข้อมูลทางพันธุกรรมใหม่นี้อย่างจริงจัง[ 255 ]
การค้นหาโฮมินินยุคแรกสุด
ในช่วงทศวรรษ 1990 ทีมงานนักมานุษยวิทยาบรรพกาลหลายทีมได้ทำงานทั่วทวีปแอฟริกาเพื่อค้นหาหลักฐานการแยกสายพันธุ์ครั้งแรกสุดของโฮมินินจากลิงใหญ่ ในปี 1994 มีฟ ลีคีย์ ค้นพบออสตราโลพิเทคัส อนาเมนซิส การค้นพบนี้ถูกบดบังด้วยการค้นพบอาร์ดิ พิเทคัส รามิดัสของทิม ดี. ไวท์ ในปี 1995 ซึ่งทำให้บันทึกฟอสซิลย้อนกลับไปได้ถึง4.2 ล้านปีก่อน
ในปี 2000 มาร์ติน พิกฟอร์ดและบริจิตต์ เซนูต์ค้นพบ ซากดึกดำบรรพ์ของ มนุษย์ยุคก่อนประวัติศาสตร์ที่เดินสองขา อายุ 6 ล้านปี ใน เนินเขาตูเก น ประเทศเคนยา ซึ่งพวกเขาตั้งชื่อว่า ออร์โรริน ตูเกเนนซิส (Orrorin tugenensis ) และในปี 2001 ทีมที่นำโดยมิเชล บรูเนต์ค้นพบกะโหลกของ ซาเฮลันโทรปัส ทชาเดนซิส ( Sahelanthropus tchadensis ) ซึ่งมีอายุประมาณ7.2ล้านปีและบรูเนต์ให้เหตุผลว่ากะโหลกนี้เป็นสัตว์ที่เดินสองขา ดังนั้นจึงเป็นโฮมินิด (หรือก็คือ โฮมินิน) ( ดู Hominidae ; คำว่า "hominids" และ hominins)
การกระจายตัวของมนุษย์


{kind=link}
{kind=link}
นักมานุษยวิทยาในช่วงทศวรรษ 1980 มีความเห็นที่แตกต่างกันเกี่ยวกับรายละเอียดบางประการของอุปสรรคในการสืบพันธุ์และการแพร่กระจายของการอพยพของสกุลHomoต่อมา พันธุศาสตร์ได้ถูกนำมาใช้เพื่อตรวจสอบและแก้ไขปัญหาเหล่านี้ ตามทฤษฎีปั๊มซาฮาราหลักฐานชี้ให้เห็นว่าสกุลHomoได้อพยพออกจากแอฟริกาอย่างน้อยสามครั้งและอาจถึงสี่ครั้ง (เช่นHomo erectus , Homo heidelbergensisและสองหรือสามครั้งสำหรับHomo sapiens ) หลักฐานล่าสุดชี้ให้เห็นว่าการแพร่กระจายเหล่านี้มีความเกี่ยวข้องอย่างใกล้ชิดกับช่วงเวลาที่ผันผวนของการเปลี่ยนแปลงสภาพภูมิอากาศ[ 260 ]
หลักฐานล่าสุดชี้ให้เห็นว่ามนุษย์อาจออกจากแอฟริกาเร็วกว่าที่เคยคิดไว้ถึงครึ่งล้านปี ทีมร่วมฝรั่งเศส-อินเดียได้ค้นพบสิ่งประดิษฐ์ของมนุษย์ในเนินเขา Siwalk ทางเหนือของนิวเดลี ซึ่งมีอายุย้อนหลังไปอย่างน้อย 2.6 ล้านปี ซึ่งเร็วกว่าการค้นพบสกุลHomo ที่เก่าแก่ที่สุดก่อนหน้านี้ ที่Dmanisiในจอร์เจียซึ่งมีอายุ 1.85 ล้านปี แม้จะเป็นที่ถกเถียงกัน แต่เครื่องมือที่พบในถ้ำของจีนก็สนับสนุนข้อสันนิษฐานที่ว่ามนุษย์ใช้เครื่องมือมาตั้งแต่ 2.48 ล้านปีก่อน[ 261 ]สิ่งนี้ชี้ให้เห็นว่าประเพณีเครื่องมือ "สับ" ของเอเชียที่พบในชวาและจีนตอนเหนืออาจออกจากแอฟริกาก่อนการปรากฏตัวของขวานมือ Acheulian
การแพร่กระจายของมนุษย์ยุคใหม่(Homo sapiens)
จนกระทั่งมีหลักฐานทางพันธุกรรม ก็มีแบบจำลองหลักสองแบบสำหรับการแพร่กระจายของมนุษย์ยุคใหม่สมมติฐานแบบหลายภูมิภาคเสนอว่าสกุลHomoประกอบด้วยประชากรที่เชื่อมต่อกันเพียงกลุ่มเดียวดังเช่นในปัจจุบัน (ไม่ใช่สายพันธุ์ที่แยกจากกัน) และวิวัฒนาการของมันเกิดขึ้นทั่วโลกอย่างต่อเนื่องในช่วงสองสามล้านปีที่ผ่านมา แบบจำลองนี้เสนอโดยMilford H. Wolpoffใน ปี 1988 [ 262 ] [ 263 ]ในทางตรงกันข้าม แบบจำลอง "ออกจากแอฟริกา" เสนอว่าH. sapiens ยุคใหม่ แยกสายพันธุ์ในแอฟริกาเมื่อไม่นานมานี้ (นั่นคือประมาณ 200,000 ปีที่แล้ว) และการอพยพผ่านยูเรเซีย ในเวลาต่อมาส่งผลให้สายพันธุ์ Homoอื่นๆ ถูกแทนที่เกือบทั้งหมดแบบจำลองนี้ได้รับการพัฒนาโดยChris Stringerและ Peter Andrews [ 264 ] [ 265 ]
การจัดลำดับ mtDNA และ Y-DNA ที่เก็บตัวอย่างจากประชากรพื้นเมืองหลากหลายกลุ่มเผยให้เห็นข้อมูลบรรพบุรุษที่เกี่ยวข้องกับมรดกทางพันธุกรรมของทั้งชายและหญิง และเสริมสร้างทฤษฎี "ออกจากแอฟริกา" และลดทอนมุมมองของวิวัฒนาการหลายภูมิภาค[ 266 ]ความแตกต่างของแผนภูมิพันธุกรรมที่จัดเรียงกันนั้นถูกตีความว่าสนับสนุนต้นกำเนิดเดียวเมื่อไม่นานมานี้[ 267 ]
"Out of Africa" จึงได้รับการสนับสนุนอย่างมากจากการวิจัยโดยใช้ดีเอ็นเอไมโทคอนเดรียเพศหญิงและโครโมโซม Y เพศชาย หลังจากวิเคราะห์แผนผังลำดับวงศ์ตระกูลที่สร้างขึ้นโดยใช้ mtDNA 133 ชนิด นักวิจัยสรุปว่าทั้งหมดสืบเชื้อสายมาจากบรรพบุรุษหญิงชาวแอฟริกันที่เรียกว่าMitochondrial Eve "Out of Africa" ยังได้รับการสนับสนุนจากข้อเท็จจริงที่ว่าความหลากหลายทางพันธุกรรมของไมโทคอนเดรียสูงที่สุดในกลุ่มประชากรแอฟริกัน[ 268 ]
การศึกษาวิจัยเกี่ยวกับความหลากหลายทางพันธุกรรมของชาวแอฟริกันอย่างกว้างขวาง ซึ่งนำโดยSarah Tishkoffพบว่าชาวซานมีความหลากหลายทางพันธุกรรมมากที่สุดในบรรดาประชากร 113 กลุ่มที่นำมาศึกษา ทำให้พวกเขากลายเป็นหนึ่งใน 14 "กลุ่มประชากรบรรพบุรุษ" การวิจัยยังระบุแหล่งกำเนิดที่เป็นไปได้ของการอพยพของมนุษย์ยุคใหม่ในแอฟริกาตะวันตกเฉียงใต้ ใกล้กับชายแดนชายฝั่งของนามิเบียและ แอง โกลา[ 269 ]หลักฐานฟอสซิลไม่เพียงพอสำหรับนักโบราณคดีRichard Leakeyที่จะยุติข้อถกเถียงเกี่ยวกับสถานที่ที่มนุษย์ยุคใหม่ปรากฏตัวครั้งแรกในแอฟริกา[ 270 ]การศึกษาแฮปโลกรุ๊ปในดีเอ็นเอโครโมโซม Yและดีเอ็นเอไมโทคอนเดรียส่วนใหญ่สนับสนุนต้นกำเนิดจากแอฟริกาเมื่อไม่นานมานี้[ 271 ]หลักฐานทั้งหมดจากดีเอ็นเอออโตโซมส่วนใหญ่ยังสนับสนุนต้นกำเนิดจากแอฟริกาเมื่อไม่นานมานี้ อย่างไรก็ตาม หลักฐานของการผสมผสานทางพันธุกรรมในมนุษย์ยุคใหม่ทั้งในแอฟริกาและต่อมาทั่วทั้งยูเรเซีย ได้รับการเสนอแนะจากงานวิจัยหลายชิ้นเมื่อไม่นานมานี้[ 272 ]
การจัดลำดับจีโนมของมนุษย์นีแอนเดอร์ทาล[ 104 ]และมนุษย์เดนิโซวัน[ 50 ] เมื่อเร็วๆ นี้แสดงให้เห็นว่ามีการผสมผสานกับประชากรเหล่านี้เกิดขึ้น กลุ่มมนุษย์ยุคใหม่ทั้งหมดที่อยู่นอกทวีปแอฟริกามี อัลลีล ของ มนุษย์นีแอนเดอร์ทาล 1–4% หรือ (ตามการวิจัยล่าสุด) ประมาณ 1.5–2.6% ในจีโนมของพวกเขา[ 105 ]และชาวเมลานีเซีย บางกลุ่ม มีอัลลีลของมนุษย์เดนิโซวันเพิ่มอีก 4–6% ผลลัพธ์ใหม่เหล่านี้ไม่ได้ขัดแย้งกับแบบจำลอง "ออกจากแอฟริกา" ยกเว้นในการตีความที่เข้มงวดที่สุด แม้ว่าจะทำให้สถานการณ์ซับซ้อนมากขึ้นก็ตาม หลังจากฟื้นตัวจากภาวะคอขวดทางพันธุกรรมที่นักวิจัยบางคนคาดการณ์ว่าอาจเชื่อมโยงกับภัยพิบัติภูเขาไฟโทบากลุ่มเล็กๆ กลุ่มหนึ่งได้ออกจากแอฟริกาและผสมพันธุ์กับมนุษย์นีแอนเดอร์ทาล อาจจะในตะวันออกกลาง บนที่ราบสเตปป์ยูเรเซีย หรือแม้แต่ในแอฟริกาเหนือ ก่อนที่พวกเขาจะจากไป ลูกหลานของพวกเขาซึ่งส่วนใหญ่ยังคงมีเชื้อสายแอฟริกันได้แพร่กระจายไปทั่วโลก ส่วนหนึ่งผสมพันธุ์กับเดนิโซวัน อาจจะในเอเชียตะวันออกเฉียงใต้ ก่อนที่จะอพยพไปยังเมลานีเซีย[ 114 ] แฮปโลไทป์ HLAที่มีต้นกำเนิดจากนีแอนเดอร์ทัลและเดนิโซวาได้รับการระบุในประชากรยูเรเซียและโอเชียเนียในปัจจุบัน[ 52 ] ยีน EPAS1ของเดนิโซวันยังพบในประชากรทิเบตด้วย[ 273 ]การศึกษาจีโนมมนุษย์โดยใช้การเรียนรู้ของเครื่องจักรได้ระบุถึงการมีส่วนร่วมทางพันธุกรรมเพิ่มเติมในยูเรเซียจากประชากรบรรพบุรุษ "ที่ไม่รู้จัก" ซึ่งอาจเกี่ยวข้องกับสายพันธุ์นีแอนเดอร์ทัล-เดนิโซวัน[ 274 ]

ยังคงมีทฤษฎีที่แตกต่างกันอยู่ว่ามีการอพยพจากแอฟริกาเพียงครั้งเดียวหรือหลายครั้ง แบบจำลองการกระจายตัวหลายครั้งเกี่ยวข้องกับทฤษฎีการกระจายตัวทางใต้[ 275 ] [ 276 ] [ 277 ]ซึ่งได้รับการสนับสนุนมากขึ้นในช่วงไม่กี่ปีที่ผ่านมาจากหลักฐานทางพันธุกรรม ภาษาศาสตร์ และโบราณคดี ในทฤษฎีนี้ มีการกระจายตัวตามแนวชายฝั่งของมนุษย์ยุคใหม่จากแหลมแอฟริกาข้ามช่องแคบบาบเอลมันดิบไปยังเยเมนในช่วงที่ระดับน้ำทะเลต่ำกว่าเมื่อประมาณ 70,000 ปีที่แล้ว กลุ่มนี้ช่วยตั้งถิ่นฐานในเอเชียตะวันออกเฉียงใต้และโอเชียเนีย ซึ่งอธิบายถึงการค้นพบแหล่งโบราณสถานของมนุษย์ยุคแรกในพื้นที่เหล่านี้เร็วกว่าในเลแวนต์มาก[ 275 ]ดูเหมือนว่ากลุ่มนี้จะพึ่งพาแหล่งทรัพยากรทางทะเลเพื่อความอยู่รอด
สตีเฟน ออปเพนไฮเมอร์เสนอว่าคลื่นมนุษย์ลูกที่สองอาจกระจายตัวผ่านโอเอซิสในอ่าวเปอร์เซียและเทือกเขาซากรอสไปยังตะวันออกกลาง หรืออาจเดินทางข้ามคาบสมุทรไซนายเข้าสู่เอเชียหลังจาก 50,000 ปีก่อนคริสตกาลไม่นาน ส่งผลให้เกิดประชากรมนุษย์ส่วนใหญ่ในยูเรเซีย มีการเสนอแนะว่ากลุ่มที่สองนี้อาจมีเทคโนโลยีเครื่องมือล่าสัตว์ขนาดใหญ่ที่ซับซ้อนกว่าและพึ่งพาแหล่งอาหารชายฝั่งน้อยกว่ากลุ่มแรก หลักฐานส่วนใหญ่เกี่ยวกับการขยายตัวของกลุ่มแรกน่าจะถูกทำลายโดยระดับน้ำทะเลที่สูงขึ้นในช่วงปลายของยุคน้ำแข็งสูงสุดแต่ละ ครั้ง [ 275 ]แบบจำลองการกระจายตัวหลายครั้งขัดแย้งกับการศึกษาที่ระบุว่าประชากรของยูเรเซียและประชากรของเอเชียตะวันออกเฉียงใต้และโอเชียเนียล้วนสืบเชื้อสายมาจากสายพันธุ์ ดีเอ็นเอไมโทคอนเดรีย L3 เดียวกัน ซึ่งสนับสนุนการอพยพครั้งเดียวออกจากแอฟริกาที่ก่อให้เกิดประชากรที่ไม่ใช่แอฟริกันทั้งหมด[ 278 ]
จากข้อมูลอายุช่วงต้นของชาวอาวริญญาเชียนแห่งบาโดชานอิหร่าน โอปเพนไฮเมอร์เสนอว่าการแพร่กระจายครั้งที่สองนี้อาจเกิดขึ้นในช่วงที่มีฝนตกชุกราว 50,000 ปีก่อนปัจจุบัน โดยวัฒนธรรมการล่าสัตว์ใหญ่ของมนุษย์ยุคใหม่ได้แพร่กระจายขึ้นไปตามเทือกเขาซากรอส นำพาจีโนมของมนุษย์ยุคใหม่จากโอมาน ข้ามอ่าวเปอร์เซีย ขึ้นเหนือไปยังอาร์เมเนียและอนาโตเลีย โดยมีสายพันธุ์หนึ่งเดินทางลงใต้ไปยังอิสราเอลและไซเรนิเซีย[ 214 ]
หลักฐานทางพันธุกรรมล่าสุดชี้ให้เห็นว่าประชากรที่ไม่ใช่ชาวแอฟริกันในปัจจุบันทั้งหมด รวมถึงประชากรในยูเรเซียและโอเชียเนีย สืบเชื้อสายมาจากคลื่นเดียวที่อพยพออกจากแอฟริกาเมื่อประมาณ 65,000 ถึง 50,000 ปีที่แล้ว[ 279 ] [ 280 ] [ 281 ]
หลักฐาน
หลักฐานที่ใช้เป็นพื้นฐานในการอธิบายวิวัฒนาการของมนุษย์ในเชิงวิทยาศาสตร์นั้นมาจากหลายสาขาของวิทยาศาสตร์ธรรมชาติแหล่งความรู้หลักเกี่ยวกับกระบวนการวิวัฒนาการนั้นโดยทั่วไปมาจากบันทึกฟอสซิล แต่ตั้งแต่มีการพัฒนาพันธุศาสตร์ในช่วงทศวรรษ 1970 การวิเคราะห์ดีเอ็นเอก็เข้ามามีบทบาทสำคัญไม่แพ้กัน การศึกษาเกี่ยวกับการเจริญเติบโต การสืบเชื้อสายและโดยเฉพาะอย่างยิ่งชีววิทยาการพัฒนาเชิงวิวัฒนาการของทั้งสัตว์มีกระดูกสันหลังและสัตว์ไม่มีกระดูกสันหลังนั้นให้ข้อมูลเชิงลึกมากมายเกี่ยวกับวิวัฒนาการของสิ่งมีชีวิตทั้งหมด รวมถึงวิวัฒนาการของมนุษย์ด้วย การศึกษาเฉพาะเจาะจงเกี่ยวกับต้นกำเนิดและชีวิตของมนุษย์คือมานุษยวิทยาโดยเฉพาะอย่างยิ่งมานุษยวิทยาบรรพกาล ซึ่งมุ่งเน้นการศึกษาประวัติศาสตร์ยุคก่อนประวัติศาสตร์ของ มนุษย์ [ 282 ]
หลักฐานจากพันธุศาสตร์

ญาติที่ใกล้ชิดที่สุดของมนุษย์ในปัจจุบันคือโบโนโบและชิมแปนซี (ทั้งสองสกุลPan ) และกอริลลา (สกุลGorilla ) [ 283 ]จากการจัดลำดับจีโนมของทั้งมนุษย์และชิมแปนซี ณ ปี 2012 ประมาณการความคล้ายคลึงกันระหว่างลำดับดีเอ็นเอของทั้งสองชนิดอยู่ระหว่าง 95% ถึง 99% [ 283 ] [ 284 ] [ 34 ]นอกจากนี้ยังเป็นที่น่าสังเกตว่าหนูมีดีเอ็นเอที่ทำงานร่วมกับมนุษย์ประมาณ 97.5% [ 285 ] โดยใช้เทคนิคที่เรียกว่านาฬิกาโมเลกุลซึ่งประมาณเวลาที่จำเป็นสำหรับการสะสมของการกลายพันธุ์ที่แตกต่างกันระหว่างสองสายพันธุ์ สามารถคำนวณวันที่โดยประมาณสำหรับการแยกสายพันธุ์ได้
ชะนี (วงศ์ Hylobatidae) และอุรังอุตัง (สกุลPongo ) เป็นกลุ่มแรกที่แยกตัวออกจากสายพันธุ์ที่นำไปสู่โฮมินิน รวมถึงมนุษย์ ตามมาด้วยกอริลลา (สกุลGorilla ) และในที่สุดก็คือชิมแปนซี (สกุลPan ) บางคนกำหนดวันที่แยกสายพันธุ์ระหว่างโฮมินินและชิมแปนซีไว้ระหว่าง4 ถึง 8ล้านปีก่อนนั่นคือในช่วงปลายยุคไมโอซีน [ 286 ] [ 287 ] [ 288 ] [ 289 ] อย่างไรก็ตามการเกิดสปีชีส์ใหม่ดูเหมือนจะยืดเยื้อผิดปกติ การแยกสายพันธุ์ครั้งแรกเกิดขึ้นในช่วงระหว่าง7 ถึง 13ล้านปีก่อนแต่การผสมข้ามสายพันธุ์อย่างต่อเนื่องทำให้การแยกสายพันธุ์ไม่ชัดเจนและทำให้การแยกสายพันธุ์อย่างสมบูรณ์ล่าช้าออกไปหลายล้านปี แพตเตอร์สัน (2006) กำหนดวันที่การแยกสายพันธุ์ครั้งสุดท้ายไว้ที่5 ถึง 6ล้านปีก่อน[ 290 ]
หลักฐานทางพันธุกรรมยังถูกนำมาใช้เพื่อเปรียบเทียบสายพันธุ์ภายในสกุลHomoโดยศึกษาการถ่ายทอดยีนระหว่างมนุษย์ยุคใหม่ตอนต้นกับมนุษย์นีแอนเดอร์ทาลและเพื่อเพิ่มความเข้าใจเกี่ยวกับรูปแบบการอพยพและการแยกสายพันธุ์ของมนุษย์ยุคแรก โดยการเปรียบเทียบส่วนต่างๆ ของจีโนมที่ไม่ได้อยู่ภายใต้การคัดเลือกโดยธรรมชาติและซึ่งสะสมการกลายพันธุ์ในอัตราที่ค่อนข้างคงที่ ทำให้สามารถสร้างแผนผังลำดับวงศ์ตระกูลทางพันธุกรรมที่รวมเอาสายพันธุ์มนุษย์ทั้งหมดตั้งแต่บรรพบุรุษร่วมสุดท้ายได้
ทุกครั้งที่การกลายพันธุ์บางอย่าง ( การเปลี่ยนแปลงนิวคลีโอไทด์เดี่ยว ) ปรากฏขึ้นในบุคคลและถูกส่งต่อไปยังลูกหลานของเขาหรือเธอ จะเกิดกลุ่มแฮปโลกรุ๊ปขึ้น ซึ่งรวมถึงลูกหลานทั้งหมดของบุคคลนั้นที่จะมียีนกลายพันธุ์นั้นด้วย โดยการเปรียบเทียบดีเอ็นเอ ไมโทคอนเด รียซึ่งสืทอดมาจากมารดาเท่านั้น นักพันธุศาสตร์ได้สรุปว่าบรรพบุรุษร่วมเพศหญิงคนสุดท้ายที่มีเครื่องหมายทางพันธุกรรมพบในมนุษย์ยุคใหม่ทุกคน หรือที่เรียกว่าอีฟแห่งไมโทคอนเด รีย น่าจะมีชีวิตอยู่เมื่อประมาณ 200,000 ปีที่แล้ว
พันธุศาสตร์วิวัฒนาการของมนุษย์ศึกษาว่าจีโนมของมนุษย์แตกต่างกันอย่างไรในแต่ละบุคคล อดีตวิวัฒนาการที่ก่อให้เกิดจีโนมเหล่านั้น และผลกระทบในปัจจุบัน ความแตกต่างระหว่างจีโนมมีนัยสำคัญและการประยุกต์ใช้ในด้านมานุษยวิทยาการแพทย์ และนิติวิทยาศาสตร์ ข้อมูลทางพันธุกรรมสามารถให้ข้อมูลเชิงลึกที่สำคัญเกี่ยวกับวิวัฒนาการของมนุษย์ คะแนนโพลีจีนิกจากดีเอ็นเอโบราณแสดงให้เห็นถึงการคัดเลือกแบบมีทิศทางสำหรับลักษณะต่างๆ รวมถึงวิวัฒนาการของสติปัญญาของมนุษย์ในบางช่วงเวลา[ 291 ]
ในเดือนพฤษภาคม พ.ศ. 2566 นักวิทยาศาสตร์รายงานเส้นทางการวิวัฒนาการของมนุษย์ที่ซับซ้อนกว่าที่เคยเข้าใจมาก่อน จากการศึกษาพบว่ามนุษย์วิวัฒนาการมาจากสถานที่และช่วงเวลาต่างๆ ในแอฟริกา แทนที่จะมาจากสถานที่และช่วงเวลาเดียว[ 292 ] [ 293 ]
หลักฐานจากบันทึกฟอสซิล


มีหลักฐานฟอสซิลน้อยมากสำหรับการแยกสายพันธุ์ของกอริลลา ชิมแปนซี และโฮมินิน[ 294 ]ฟอสซิลที่เก่าแก่ที่สุดที่ถูกเสนอให้เป็นสมาชิกของสายพันธุ์โฮมินิน ได้แก่Sahelanthropus tchadensisซึ่งมีอายุ7 ล้านปีก่อนOrrorin tugenensisซึ่งมีอายุ5.7 ล้านปีก่อนและArdipithecus kadabbaซึ่งมีอายุ5.6 ล้านปีก่อนแต่ละชนิดเหล่านี้ถูกกล่าวอ้างว่าเป็น บรรพบุรุษที่ เดินสองขาของโฮมินินในยุคต่อมา แต่ในแต่ละกรณี ข้อกล่าวอ้างเหล่านี้ก็ถูกโต้แย้ง นอกจากนี้ยังเป็นไปได้ว่าหนึ่งหรือมากกว่านั้นของสายพันธุ์เหล่านี้เป็นบรรพบุรุษของลิงแอฟริกันสายพันธุ์อื่น หรือเป็นตัวแทนของบรรพบุรุษร่วมกันระหว่างโฮมินินและลิงสายพันธุ์อื่น
คำถามเกี่ยวกับความสัมพันธ์ระหว่างซากดึกดำบรรพ์ยุคแรกเหล่านี้กับสายพันธุ์โฮมินินจึงยังคงต้องหาคำตอบต่อไป จากสายพันธุ์ยุคแรกเหล่านี้ ออสตราโลพิเทคัสถือกำเนิดขึ้นเมื่อประมาณ4 ล้านปีก่อนและแยกออกเป็นสอง สาขา คือ สาขา ที่แข็งแรง (เรียกอีกอย่างว่าParanthropus ) และสาขาที่บอบบาง ซึ่งหนึ่งในนั้น (อาจจะเป็น A. garhi ) น่าจะกลายเป็นบรรพบุรุษของสกุลHomoสายพันธุ์ออสตราโลพิเทคัสที่มีซากดึกดำบรรพ์มากที่สุดคือAustralopithecus afarensisโดยมีซากดึกดำบรรพ์มากกว่า 100 ตัวอย่าง พบตั้งแต่ทางตอนเหนือของเอธิโอเปีย (เช่น "ลูซี่" ที่มีชื่อเสียง) ไปจนถึงเคนยา และแอฟริกาใต้ ซากดึกดำบรรพ์ของออสตราโลพิเทคัสที่แข็งแรง เช่นA. robustus (หรืออีกชื่อหนึ่งคือParanthropus robustus ) และA./P. boiseiมีอยู่มากมายในแอฟริกาใต้ ณ แหล่งโบราณคดีต่างๆ เช่นKromdraaiและSwartkransและบริเวณรอบทะเลสาบ Turkanaในเคนยา
สมาชิกที่เก่าแก่ที่สุดของสกุลHomoคือHomo habilisซึ่งวิวัฒนาการเมื่อประมาณ2.8 ล้านปีก่อน [ 40 ] H. habilisเป็นสายพันธุ์แรกที่เรามีหลักฐานยืนยันการใช้เครื่องมือหิน พวกเขาพัฒนา เทคโนโลยีหิน Oldowanซึ่งตั้งชื่อตามหุบเขา Olduvai ที่พบตัวอย่างแรก นักวิทยาศาสตร์บางคนพิจารณาว่าHomo rudolfensisซึ่งเป็นกลุ่มฟอสซิลที่มีขนาดตัวใหญ่กว่าและมีรูปร่างคล้ายกับ ฟอสซิล H. habilis ดั้งเดิม เป็นสายพันธุ์ที่แยกต่างหาก ในขณะที่คนอื่นๆ พิจารณาว่าเป็นส่วนหนึ่งของH. habilisซึ่งเป็นเพียงการแปรผันภายในสายพันธุ์ หรืออาจเป็นความแตกต่างทางเพศสมองของโฮมินินยุคแรกเหล่านี้มีขนาดใกล้เคียงกับสมองของชิมแปนซี และการปรับตัวหลักของพวกเขาคือการเดินสองขาเพื่อปรับตัวให้เข้ากับการดำรงชีวิตบนบก
ในช่วงหนึ่งล้านปีต่อมา กระบวนการพัฒนาสมองได้เริ่มต้นขึ้น และเมื่อโฮโมอิเร็ กตัสปรากฏตัว ในบันทึกฟอสซิล (ประมาณ 1.9 ล้านปีก่อน ) ความจุของ กะโหลกศีรษะก็เพิ่มขึ้นเป็นสองเท่า โฮโมอิเร็กตัสเป็นโฮมินินกลุ่มแรกที่อพยพออกจากแอฟริกา และระหว่าง1.8 ถึง 1.3ล้านปีก่อนสายพันธุ์นี้ได้แพร่กระจายไปทั่วแอฟริกา เอเชีย และยุโรป ประชากรโฮโมอิเร็กตัส กลุ่มหนึ่ง ซึ่งบางครั้งก็ถูกจัดเป็นสายพันธุ์แยกต่างหากคือโฮโมเออร์แกสเตอร์ ยัง คงอยู่ในแอฟริกาและวิวัฒนาการไปเป็นโฮโมเซเปียนส์เชื่อกันว่าโฮโมอิเร็กตัสและโฮโมเออร์แกสเตอร์เป็นสายพันธุ์แรกที่ใช้ไฟและเครื่องมือที่ซับซ้อน ในยูเรเซีย โฮโมอิเร็กตัส ได้วิวัฒนาการไปเป็นสายพันธุ์ต่างๆ เช่นโฮโมแอนทีเซสเซอร์โฮโมไฮเดลเบอร์เกนซิสและโฮโมนีแอนเดอร์ทาเลนซิส ฟอสซิลที่เก่าแก่ที่สุดของมนุษย์ยุคใหม่ที่มีลักษณะทางกายวิภาคเหมือนปัจจุบันนั้นมาจากยุคหินเก่าตอนกลาง ประมาณ 300,000-200,000 ปีที่แล้ว เช่น ซากเฮอร์โตและโอโมในเอธิโอเปีย ซาก เจเบล อิรุดในโมร็อกโก และซากฟลอริสบาดในแอฟริกาใต้ ส่วนฟอสซิลในยุคต่อมาจากถ้ำสคูลในอิสราเอลและยุโรปตอนใต้เริ่มต้นเมื่อประมาณ 90,000 ปีที่แล้ว ( 0.09 ล้านปีที่แล้ว )
เมื่อมนุษย์ยุคใหม่แพร่กระจายออกจากแอฟริกา พวกเขาได้พบกับโฮมินินอื่นๆ เช่นโฮโมนีแอนเดอร์ทาเลนซิสและเดนิโซแวน ซึ่งอาจวิวัฒนาการมาจากประชากรของโฮโมอิเร็กตัสที่ออกจากแอฟริกาเมื่อประมาณ2 ล้านปีก่อนลักษณะของการปฏิสัมพันธ์ระหว่างมนุษย์ยุคแรกกับสายพันธุ์พี่น้องเหล่านี้เป็นแหล่งที่มาของข้อโต้แย้งมายาวนาน คำถามคือมนุษย์เข้ามาแทนที่สายพันธุ์ก่อนหน้านี้หรือไม่ หรือว่าพวกเขามีความคล้ายคลึงกันมากพอที่จะผสมพันธุ์กันได้ ซึ่งในกรณีนี้ประชากรก่อนหน้านี้อาจมีส่วนช่วยในการถ่ายทอดสารพันธุกรรมให้กับมนุษย์ยุคใหม่[ 295 ] [ 296 ]
การอพยพออกจากแอฟริกานี้คาดว่าเริ่มต้นเมื่อประมาณ 70,000–50,000 ปีก่อนและมนุษย์ยุคใหม่ได้แพร่กระจายไปทั่วโลกในเวลาต่อมา โดยเข้ามาแทนที่โฮมินินรุ่นก่อนๆ ไม่ว่าจะด้วยการแข่งขันหรือการผสมข้ามสายพันธุ์ พวกเขาอาศัยอยู่ในยูเรเซียและโอเชียเนียเมื่อ 40,000 ปีก่อน และในทวีปอเมริกาเมื่ออย่างน้อย 14,500 ปีก่อน[ 297 ]
การผสมพันธุ์ข้ามสายพันธุ์

The hypothesis of interbreeding, also known as hybridization, admixture or hybrid-origin theory, has been discussed ever since the discovery of Neanderthal remains in the 19th century.[298] The linear view of human evolution began to be abandoned in the 1970s as different species of humans were discovered that made the linear concept increasingly unlikely. In the 21st century with the advent of molecular biology techniques and computerization, whole-genome sequencing of Neanderthal and human genome were performed, confirming recent admixture between different human species.[104] In 2010, evidence based on molecular biology was published, revealing unambiguous examples of interbreeding between archaic and modern humans during the Middle Paleolithic and early Upper Paleolithic. It has been demonstrated that interbreeding happened in several independent events that included Neanderthals and Denisovans, as well as several unidentified hominins.[299] Today, approximately 2% of DNA from all non-African populations (including Europeans, Asians, and Oceanians) is Neanderthal,[104] with traces of Denisovan heritage.[300] Also, 4–6% of modern Melanesian genetics are Denisovan.[300] Comparisons of the human genome to the genomes of Neanderthals, Denisovans and apes can help identify features that set modern humans apart from other hominin species. In a 2016 comparative genomics study, a Harvard Medical School/UCLA research team made a world map on the distribution and made some predictions about where Denisovan and Neanderthal genes may be impacting modern human biology.[301][302]
For example, comparative studies in the mid-2010s found several traits related to neurological, immunological,[303] developmental, and metabolic phenotypes, that were developed by archaic humans to European and Asian environments and inherited to modern humans through admixture with local hominins.[304][305]
แม้ว่าเรื่องราววิวัฒนาการของมนุษย์มักจะเป็นที่ถกเถียงกัน แต่การค้นพบหลายอย่างตั้งแต่ปี 2010 แสดงให้เห็นว่าวิวัฒนาการของมนุษย์ไม่ควรถูกมองว่าเป็นเพียงความก้าวหน้าเชิงเส้นตรงหรือแบบแตกแขนง แต่เป็นการผสมผสานของสายพันธุ์ที่เกี่ยวข้อง[ 50 ] [ 6 ] [ 7 ] [ 8 ]ในความเป็นจริง การวิจัยทางจีโนมิกส์แสดงให้เห็นว่าการผสมข้ามสายพันธุ์ระหว่างสายพันธุ์ที่แตกต่างกันอย่างมากเป็นกฎ ไม่ใช่ข้อยกเว้น ในวิวัฒนาการของมนุษย์[ 5 ]ยิ่งไปกว่านั้น มีการโต้แย้งว่าการผสมข้ามสายพันธุ์เป็นแรงผลักดันที่สำคัญในการกำเนิดของมนุษย์ยุคใหม่[ 5 ]
เครื่องมือหิน
มีการค้นพบเครื่องมือหินครั้งแรกเมื่อประมาณ 2.6 ล้านปีก่อน เมื่อโฮมินินในแอฟริกาตะวันออกใช้ เครื่องมือที่เรียกว่า เครื่องมือแกน ซึ่ง เป็น เครื่องมือสับที่ทำจากแกนกลมที่ถูกแยกออกด้วยการตีแบบง่ายๆ[ 306 ]นี่เป็นจุดเริ่มต้นของยุคหินเก่าหรือยุคหิน โบราณ โดยถือว่าจุดสิ้นสุดของยุคหินเก่าคือจุดสิ้นสุดของยุคน้ำแข็ง ครั้งสุดท้าย เมื่อประมาณ 10,000 ปีก่อน ยุคหินเก่าแบ่งออกเป็นยุคหินเก่าตอนต้น (ยุคหินยุคแรก) ซึ่งสิ้นสุดเมื่อประมาณ 350,000–300,000 ปีก่อนยุคหินเก่าตอนกลาง (ยุคหินยุคกลาง) จนถึง 50,000–30,000 ปีก่อน และยุคหินเก่าตอนปลาย (ยุคหินยุคหลัง) 50,000–10,000 ปีก่อน
นักโบราณคดีที่ทำงานในหุบเขาเกรตริฟต์ในเคนยาได้ค้นพบเครื่องมือหินที่เก่าแก่ที่สุดในโลก ซึ่งมีอายุราว 3.3 ล้านปี เครื่องมือเหล่านี้มีอายุมากกว่าเครื่องมือหินจากเอธิโอเปียที่เคยครองตำแหน่งนี้มาก่อนถึง 700,000 ปี[ 208 ] [ 307 ] [ 308 ] [ 309 ]
ช่วงเวลาตั้งแต่ 700,000 ถึง 300,000 ปีก่อน เรียกอีกอย่างว่ายุคอะเชอูเลียนเมื่อมนุษย์ยุคเออร์แกสเตอร์ (หรืออีเร็กตัส ) สร้างขวาน หินขนาดใหญ่ จากหินเหล็กไฟและควอตไซต์ในตอนแรกค่อนข้างหยาบ (อะเชอูเลียนตอนต้น) ต่อมา " ปรับแต่ง " ด้วยการตีเพิ่มเติมที่ละเอียดกว่าที่ด้านข้างของเศษหิน หลังจาก 350,000 ปีก่อน เทคนิคเลอวาลลัวส์ที่ละเอียดกว่าได้รับการพัฒนาขึ้น ซึ่งเป็นการตีต่อเนื่องกันหลายครั้ง โดยใช้ในการสร้างเครื่องขูด เครื่องหั่น ("ราคลอยร์") เข็ม และเข็มแบน[ 306 ]ในที่สุด หลังจากประมาณ 50,000 ปีก่อน เครื่องมือหินเหล็กไฟที่ละเอียดและเฉพาะทางมากขึ้นก็ถูกสร้างขึ้นโดยมนุษย์นีแอนเดอร์ทาลและโครแม็กนอง ผู้อพยพ (มีด ใบมีด ที่ตัก) เครื่องมือที่ทำจากกระดูกยังถูกสร้างขึ้นโดยH. sapiensในแอฟริกาเมื่อ 90,000–70,000 ปีก่อน[ 215 ] [ 310 ]และยังพบได้จาก แหล่งโบราณคดี ของ H. sapiens ยุคแรก ในยูเรเซียเมื่อประมาณ 50,000 ปีก่อน
รายชื่อสายพันธุ์
รายชื่อนี้เรียง ลำดับ ตามลำดับเวลา ตาม สกุลในตารางชื่อชนิด/ชนิดย่อยบางชนิดเป็นที่รู้จักกันดี ในขณะที่บางชนิดยังไม่เป็นที่รู้จักมากนัก โดยเฉพาะในสกุลHomoโปรดดูบทความเพื่อดูข้อมูลเพิ่มเติม
ดูเพิ่มเติม
- วิวัฒนาการแบบปรับตัวในจีโนมมนุษย์
- ความสัมพันธ์แบบมิตร-ศัตรู
- มานุษยวิทยา
- อาร์คีโอเจเนติกส์
- มานุษยวิทยาชีวภาพ
- มานุษยวิทยาทันตกรรม
- ทฤษฎีการถ่ายทอดทางพันธุกรรมแบบคู่
- วิวัฒนาการของศีลธรรม
- เวชศาสตร์วิวัฒนาการ
- ประสาทวิทยาเชิงวิวัฒนาการ
- กำเนิดเชิงวิวัฒนาการของศาสนา
- จิตวิทยาเชิงวิวัฒนาการ
- นิเวศวิทยาพฤติกรรมมนุษย์
- ต้นกำเนิดของมนุษย์
- ความหลงเหลือของมนุษย์
- รายชื่อฟอสซิลวิวัฒนาการของมนุษย์
- บรรพชีวินวิทยาโมเลกุล
- ภาวะกลืนไม่เข้าคายไม่ออกทางสูติกรรม
- ที่มาของภาษา
- ที่มาของคำพูด
- ยุคก่อนประวัติศาสตร์ของการเปลือยกายและการสวมเสื้อผ้า
- การคัดเลือกทางเพศในมนุษย์
- บาดแผลทางใจที่ส่งต่อกันมาหลายรุ่น
- ลำดับเหตุการณ์วิวัฒนาการของมนุษย์
หมายเหตุ
- ^ a b cการประมาณอายุตามปกติของH. habilisอยู่ที่ประมาณ 2.1 ถึง 2.3 ล้านปี[ 42 ] [ 180 ]ข้อเสนอแนะสำหรับการเลื่อนอายุกลับไปเป็น 2.8 ล้านปีเกิดขึ้นในปี 2015 โดยอิงจากการค้นพบกระดูกขากรรไกร[ 181 ]
- ^อย่าสับสนกับ วงศ์ Pongidae ซึ่ง เป็นวงศ์ที่เลิกใช้แล้ว ที่ใช้จัดกลุ่มอุรังอุตังกอริลลาและชิมแปนซี เข้าด้วยกัน เพื่อแยกพวกมันออกจากมนุษย์
- ^ยังไม่มีข้อตกลงทั่วไปเกี่ยวกับสาย การสืบเชื้อสาย พิเศษของ H. sapiensจาก H. erectusบางชนิดที่ปรากฏในภาพอาจไม่ได้เป็นบรรพบุรุษทางวิวัฒนาการโดยตรงของ H. sapiensและอาจไม่ได้สืบเชื้อสายมาจากกันโดยตรง ได้แก่:
- H. heidelbergensisอาจไม่ได้สืบเชื้อสายมาจากH. antecessor [ 152 ]
- H. heidelbergensisไม่น่าจะเป็นบรรพบุรุษของH. sapiens และ H. antecessorก็ไม่ใช่เช่นกัน[ 152 ]
- H. ergasterมักถูกพิจารณาว่าเป็นบรรพบุรุษทางวิวัฒนาการถัดไปของH. sapiensต่อจากH. erectusอย่างไรก็ตาม ยังมีความไม่แน่นอนอย่างมากเกี่ยวกับความถูกต้องของการจัดประเภทให้เป็นสปีชีส์ที่แยกต่างหาก จาก H. erectus [ 153 ]
- ^ H. erectusในความหมายแคบ (สายพันธุ์เอเชีย) สูญพันธุ์ไปแล้วเมื่อ 140,000 ปีที่แล้ว Homo erectus soloensisที่พบในชวาถือเป็น H. erectus ที่รอดชีวิตมาได้ล่าสุดเท่าที่ทราบ เดิมทีมีการกำหนดอายุไว้ว่าประมาณ 50,000 ถึง 40,000 ปีที่แล้ว แต่การศึกษาในปี 2011 ได้เลื่อนวันที่สูญพันธุ์ของ H. erectus soloensis กลับไป เป็น 143,000 ปีที่แล้วเป็นอย่างช้าที่สุด ซึ่งน่าจะเป็นไปได้ก่อน 550,000 ปีที่แล้ว [ 183 ]
- ^คำภาษาละตินที่หมายถึงเพศชายที่เป็นผู้ใหญ่เท่านั้นคือ vir
- ^โปรดดู บทความเกี่ยว กับการตั้งชื่อแบบทวิภาคและระบบธรรมชาติ (Systema Naturae )
อ่านเพิ่มเติม
- อเล็กซานเดอร์, ริชาร์ด ดี. (1990). มนุษย์วิวัฒนาการมาได้อย่างไร? ข้อคิดเกี่ยวกับสายพันธุ์ที่มีเอกลักษณ์เฉพาะตัว (PDF)สิ่งพิมพ์พิเศษ แอนน์ อาร์เบอร์: พิพิธภัณฑ์สัตว์วิทยา มหาวิทยาลัยมิชิแกนหน้า 1–38 . LCCN 90623893 . OCLC 22860997 . เก็บถาวรจากต้นฉบับ(PDF)เมื่อวันที่ 4 มีนาคม 2016 . สืบค้นเมื่อ6 พฤษภาคม 2015 .
- บาร์ตัน, นิโคลัส เอช. ; บริกส์, เดเร็ก อีจี ; ไอเซน, โจนาธาน เอ. ; และคณะ (2007). วิวัฒนาการ . สำนักพิมพ์โคลด์สปริงฮาร์เบอร์แล็บพลาคิวเลชัน . ISBN 978-0-87969-684-9LCCN 2007010767 OCLC 86090399
- เบลล์วูด, ปีเตอร์ (2022). การเดินทางห้าล้านปี: การเดินทางของมนุษย์จากลิงสู่เกษตรกรรม . พรินซ์ตัน: สำนักพิมพ์มหาวิทยาลัยพรินซ์ตัน. ISBN 978-0-691-19757-9.
- Enard, Wolfgang; Przeworski, Molly; Fisher, Simon E. ; และคณะ (22 สิงหาคม 2545). "วิวัฒนาการระดับโมเลกุลของFOXP2ยีนที่เกี่ยวข้องกับการพูดและภาษา" Nature . 418 (6900): 869– 872. Bibcode : 2002Natur.418..869E . doi : 10.1038/nature01025 . hdl : 11858/00-001M-0000-0012-CB89-A . ISSN 0028-0836 . PMID 12192408 . S2CID 4416233 .
- Flinn, Mark V.; Geary, David C. ; Ward, Carol V. (2005). "การครอบงำทางนิเวศวิทยา การแข่งขันทางสังคม และการแข่งขันด้านอาวุธแบบพันธมิตร: เหตุใดมนุษย์จึงวิวัฒนาการสติปัญญาที่เหนือกว่า" (PDF)วิวัฒนาการและพฤติกรรมมนุษย์ 26 ( 1): 10– 46. Bibcode : 2005EHumB..26...10F . doi : 10.1016/j.evolhumbehav.2004.08.005 . ISSN 1090-5138 . เก็บถาวร(PDF)จากต้นฉบับเมื่อวันที่ 4 มิถุนายน 2007 . สืบค้นเมื่อ5 พฤษภาคม 2015 .
- Galinon-Melenec, Béatrice (2015). "จาก "ร่องรอย" และ "ร่องรอยของมนุษย์" สู่ "กระบวนทัศน์ร่องรอยของมนุษย์"ใน Parrend, Pierre; Bourgine, Paul; Collet, Pierre (บรรณาธิการ). การประชุมออนไลน์ระดับโลกครั้งแรกเกี่ยวกับระบบที่ซับซ้อนในวิทยาเขตดิจิทัล . เทมเป, แอริโซนา: สปริงเกอร์ .
- กิบบอนส์, แอนน์ (2006). มนุษย์คนแรก: การแข่งขันเพื่อค้นหาบรรพบุรุษยุคแรกสุดของเรา . นิวยอร์ก: ดับเบิลเดย์ . ISBN 978-0-385-51226-8LCCN 2005053780 OCLC 61652817
- Hartwig, Walter C. , บรรณาธิการ (2002). บันทึกฟอสซิลของไพรเมต . การศึกษาเคมบริดจ์ด้านมานุษยวิทยาชีวภาพและวิวัฒนาการ. เล่มที่ 33. สำนักพิมพ์มหาวิทยาลัยเคมบริดจ์ . ISBN 978-0-521-66315-1LCCN 2001037847 OCLC 47254191
- ไฮซ์มันน์, เอลมาร์ พีเจ; เริ่ม, David R. (พฤศจิกายน 2544). "โฮมินอยด์ยูเรเชียนที่เก่าแก่ที่สุด" (PDF ) วารสารวิวัฒนาการของมนุษย์ . 41 (5): 463– 481. Bibcode : 2001JHumE..41..463H . ดอย : 10.1006/jhev.2001.0495 . ISSN 0047-2484 . PMID11681862 . S2CID 21230992 .
- ฮิลล์, แอนดรูว์; วอร์ด, สตีเวน (1988). "ต้นกำเนิดของโฮมินิดี: บันทึกวิวัฒนาการของโฮมินอยด์ขนาดใหญ่ในแอฟริกา ระหว่าง 14 ล้านปีก่อนและ 4 ล้านปีก่อน"วารสารมานุษยวิทยากายภาพอเมริกัน 31 ( 59): 49– 83. รหัสบรรณานุกรม : 1988AJPA...31S..49H . doi : 10.1002/ajpa.1330310505 . ISSN 0002-9483 .
- Hoagland, Hudson (1964). "วิทยาศาสตร์และมนุษยนิยมใหม่". Science . 143 (3602): 111– 114. Bibcode : 1964Sci...143..111H . doi : 10.1126/science.143.3602.111 . PMID 17781189 .
- Ijdo, Jacob W.; Baldini, Antonio; Ward, David C.; และคณะ (15 ตุลาคม 1991). "ต้นกำเนิดของโครโมโซม 2 ของมนุษย์: การหลอมรวมเทโลเมียร์-เทโลเมียร์ในบรรพบุรุษ" . Proceedings of the National Academy of Sciences of the United States of America . 88 (20): 9051– 9055. Bibcode : 1991PNAS...88.9051I . doi : 10.1073/pnas.88.20.9051 . ISSN 0027-8424 . PMC 52649 . PMID 1924367 .– โครโมโซมของลิงบรรพบุรุษสองตัวรวมกัน ก่อให้เกิดโครโมโซมคู่ที่ 2 ของมนุษย์
- โจฮันสัน, โดนัลด์ ; หว่อง, เคท (2010). มรดกของลูซี่: การค้นหาต้นกำเนิดของมนุษย์ . นิวยอร์ก: สำนักพิมพ์ทรีริเวอร์ส. ISBN 978-0-307-39640-2LCCN 2010483830 OCLC 419801728
- Jones, Steve; Martin, Robert D.; Pilbeam , David, บรรณาธิการ (1994) [1992]. สารานุกรมวิวัฒนาการมนุษย์แห่งเคมบริดจ์คำนำโดย Richard Dawkins สำนักพิมพ์มหาวิทยาลัยเคมบริดจ์ ISBN 978-0-521-32370-3LCCN 92018037 OCLC 444512451 (หนังสือเล่มนี้มีบทต่างๆ ที่มีประโยชน์และอัดแน่นไปด้วยข้อมูลเกี่ยวกับวิวัฒนาการของไพรเมตโดยทั่วไป และวิวัฒนาการของมนุษย์โดยเฉพาะ รวมถึงประวัติศาสตร์ของฟอสซิล)
- คาวาบาตะ, ฮิโรชิ; ไคฟุ, โยสุเกะ (2020). หลงทางในวิวัฒนาการ: สำรวจเส้นทางของมนุษยชาติในเอเชีย . โตเกียว: มูลนิธิอุตสาหกรรมสิ่งพิมพ์เพื่อวัฒนธรรมแห่งญี่ปุ่น. ISBN 978-4-86658-133-0เก็บถาวรจากต้นฉบับเมื่อวันที่ 20 ตุลาคม 2022 เรียกดูเมื่อวันที่ 20 ตุลาคม 2022
- ลีคีย์, ริชาร์ด อี.; ลูวิน, โรเจอร์ (1992). การพิจารณาต้นกำเนิดใหม่: ในการค้นหาสิ่งที่ทำให้เราเป็นมนุษย์ . นิวยอร์ก: ดับเบิลเดย์ . ISBN 978-0-385-41264-3LCCN 92006661 OCLC 25373161
- เลวิน, โรเจอร์ (1997). กระดูกแห่งข้อขัดแย้ง: ข้อถกเถียงในการค้นหาต้นกำเนิดของมนุษย์ (ฉบับที่ 2). สำนักพิมพ์มหาวิทยาลัยชิคาโก . ISBN 978-0-226-47651-3LCCN 97000972 OCLC 36181117
- มอร์วูด, ไมค์; แวน อูสเตอร์ซี, เพนนี (2007). มนุษย์คนใหม่: การค้นพบที่น่าตกใจและเรื่องราวแปลกประหลาดของ 'ฮอบบิท' แห่งฟลอเรส ประเทศอินโดนีเซีย . นิวยอร์ก: สำนักพิมพ์ สมิธโซเนียน / ฮาร์เปอร์คอลลินส์ . ISBN 978-0-06-089908-0LCCN 2006052267 OCLC 76481584
- โอปเพนไฮเมอร์, สตีเฟน (2003). จากสวนเอเดน: การตั้งถิ่นฐานของมนุษย์ทั่วโลก . ลอนดอน: คอนสเตเบิล แอนด์ โรบินสัน . ISBN 978-1-84119-697-8LCCN 2005482222 OCLC 52195607
- Ovchinnikov, Igor V.; Götherström, Anders; Romanova, Galina P.; และคณะ (30 มีนาคม 2000). "การวิเคราะห์โมเลกุลของดีเอ็นเอของมนุษย์นีแอนเดอร์ทัลจากเทือกเขาคอเคซัสตอนเหนือ" Nature . 404 ( 6777): 490– 493. Bibcode : 2000Natur.404..490O . doi : 10.1038/35006625 . ISSN 0028-0836 . PMID 10761915. S2CID 3101375 .
- โรเบิร์ตส์, อลิซ เอ็ม. (2009). การเดินทางอันน่าทึ่งของมนุษย์: เรื่องราวของการที่เราเข้ามาตั้งอาณานิคมบนโลก . ลอนดอน: สำนักพิมพ์บลูมส์เบอรี . ISBN 978-0-7475-9839-8. OCLC 310156315 .
- Shreeve, James (1995). ปริศนาของมนุษย์นีแอนเดอร์ทัล: ไขปริศนาต้นกำเนิดของมนุษย์ยุคใหม่ . นิวยอร์ก: William Morrow & Co. ISBN 978-0-688-09407-2LCCN 95006337 OCLC 32088673
- สตริงเกอร์, คริส บี. (2011). กำเนิดของเผ่าพันธุ์มนุษย์ . ลอนดอน: อัลเลน เลน . ISBN 978-1-84614-140-9LCCN 2011489742 OCLC 689522193
- สตริงเกอร์, คริส บี.; แอนดรูว์ส, ปีเตอร์ (2005). โลกแห่งวิวัฒนาการของมนุษย์ฉบับสมบูรณ์ . ลอนดอน/นิวยอร์ก: เทมส์ แอนด์ ฮัดสัน . ISBN 978-0-500-05132-0LCCN 2004110563 OCLC 224377190
- สตริงเกอร์, คริสโตเฟอร์; แม็คกี, โรบิน (1997). การอพยพจากแอฟริกา: ต้นกำเนิดของมนุษยชาติยุคใหม่ . นิวยอร์ก: เฮนรี โฮลต์ แอนด์ โค. ISBN 978-0-8050-2759-4LCCN 96037718 OCLC 36001167
- ไซค์ส, รีเบคก้า แร็กก์ (2020). ญาติสนิท: ชีวิต ความรัก ความตาย และศิลปะของมนุษย์นีแอนเดอร์ทาล . ลอนดอน: บลูมส์เบอรี ซิกมา . ISBN 978-1-4729-3749-0.
- แทตเตอร์ซอลล์, เอียน (2008). เส้นทางฟอสซิล: เรารู้ได้อย่างไรว่าสิ่งที่เราคิดว่าเรารู้เกี่ยวกับวิวัฒนาการของมนุษย์ (ฉบับที่ 2). สำนักพิมพ์มหาวิทยาลัยออกซ์ฟอร์ด . ISBN 978-0-19-536766-9LCCN 2008013654 OCLC 218188644
- แวน อูสเตอร์ซี, เพนนี (1999). กระดูกมังกร: เรื่องราวของมนุษย์ปักกิ่ง . เซนต์ลีโอนาร์ดส์, นิวเซาท์เวลส์: อัลเลน แอนด์ อันวินออสเตรเลีย. ISBN 978-1-86508-123-6LCCN 00300421 OCLC 45853997
- เวด, นิโคลัส (2006). ก่อนรุ่งอรุณ: การกู้คืนประวัติศาสตร์ที่สูญหายของบรรพบุรุษของเรา . นิวยอร์ก: เพนกวิน . ISBN 978-1-59420-079-3LCCN 2005055293 OCLC 62282400
- วอล์คเกอร์, อลัน; ชิปแมน, แพท (1996). ภูมิปัญญาแห่งกระดูก: การค้นหาต้นกำเนิดของมนุษย์ . ลอนดอน: ไวเดนเฟลด์ แอนด์ นิโคลสัน . ISBN 978-0-297-81670-6. OCLC 35202130 .
- ไวส์, มาร์ค แอล.; แมนน์, อลัน อี. (1985). ชีววิทยาและพฤติกรรมของมนุษย์: มุมมองทางมานุษยวิทยา (ฉบับที่ 4). บอสตัน: ลิตเติล บราวน์ แอนด์ โค. ISBN 978-0-316-92894-6LCCN 85000158 OCLC 11726796 (หนังสือเล่มนี้มีคำอธิบายที่เข้าใจง่ายเกี่ยวกับมนุษย์และไพรเมตที่ไม่ใช่มนุษย์ วิวัฒนาการของพวกมัน และประวัติฟอสซิล)
- เวลส์, สเปนเซอร์ (2003) [ตีพิมพ์ครั้งแรกในปี 2002; พรินซ์ตัน, นิวเจอร์ซีย์: สำนักพิมพ์มหาวิทยาลัยพรินซ์ตัน] การเดินทางของมนุษย์: การผจญภัยทางพันธุกรรม (ฉบับปกอ่อนของสำนักพิมพ์แรนดอมเฮาส์) นิวยอร์ก: แรนดอมเฮาส์ISBN 978-0-8129-7146-0LCCN 2003066679 OCLC 53287806
ลิงก์ภายนอก
- " เชื้อชาติ วิวัฒนาการ และวิทยาศาสตร์เกี่ยวกับต้นกำเนิดของมนุษย์ " โดย อลิสัน ฮอปเปอร์, Scientific American (5 กรกฎาคม 2021)
- "วิวัฒนาการของมนุษย์" บีบีซี ไซแอนซ์ แอนด์เนเจอร์สืบค้นเมื่อ6 พฤษภาคม 2015
- " การกำเนิดของมนุษย์" สถาบันต้นกำเนิดมนุษย์มหาวิทยาลัยรัฐแอริโซนาสืบค้นเมื่อ 6 พฤษภาคม 2558
- "กระดูก หิน และยีน: ต้นกำเนิดของมนุษย์ยุคใหม่" (ชุดบรรยายวิดีโอ) สถาบันการแพทย์โฮเวิร์ด ฮิวส์เก็บถาวรจากต้นฉบับเมื่อวันที่ 24 เมษายน 2558 เรียกดูเมื่อวันที่ 6 พฤษภาคม 2558
- " ภาพประกอบ วิวัฒนาการ : บทที่ 25" สำนักพิมพ์ห้องปฏิบัติการโคล ด์สปริงฮาร์เบอร์สืบค้นเมื่อ6 พฤษภาคม 2558– ภาพประกอบจากหนังสือวิวัฒนาการ (2007)
- "วิวัฒนาการของมนุษย์"โครงการต้นกำเนิดมนุษย์ สถาบันส มิธโซเนียนสืบค้นเมื่อ24 มิถุนายน 2013
- "ลำดับเหตุการณ์วิวัฒนาการของมนุษย์" . ArchaeologyInfo.com. เก็บถาวรจากต้นฉบับเมื่อวันที่ 18 มิถุนายน 2013 . เรียกดูเมื่อวันที่ 24 มิถุนายน 2013 .
- วิดีโอ "ร่องรอยมนุษย์" (2015) มหาวิทยาลัยนอร์มังดี UNIHAVRE, CNRS , IDEES, ห้องปฏิบัติการด้านร่องรอยมนุษย์ หน่วยระบบที่ซับซ้อน Unitwin วิทยาเขตดิจิทัล UNESCO
- แลมเบิร์ต, ทิม (ผู้อำนวยการสร้าง) (24 มิถุนายน 2015). ชนพื้นเมืองดั้งเดิม . ลอนดอน: วอลล์ทูวอลล์เทเลวิชั่น . OCLC 910115743. สืบค้นเมื่อ18 กรกฎาคม 2015 .
- วิดีโอ "การหล่อหลอมมนุษยชาติ"ปี 2013 มหาวิทยาลัยเยล
- เส้นเวลาของมนุษย์ (แบบอินเทอร์แอ็กทีฟ) – สถาบัน สมิธโซเนียนพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติแห่งชาติ (สิงหาคม 2559)
- วิวัฒนาการของมนุษย์การสนทนาทางวิทยุ BBC Radio 4 กับ สตีฟ โจนส์, เฟร็ด สปอร์ และ มาร์กาเร็ต เคล็ก ( ในยุคของเรา , 16 กุมภาพันธ์ 2549)
- ลำดับเหตุการณ์วิวัฒนาการของมนุษย์ (Home Sapiens ) − สถาบันสมิธโซเนียน (กุมภาพันธ์ 2021)
- ประวัติศาสตร์วิวัฒนาการของมนุษย์ในสหรัฐอเมริกา – Salon (24 สิงหาคม 2021)
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ วิวัฒนาการของมนุษย์
โฮโมเซเปียนส์เป็นสายพันธุ์ที่แตกต่างกันของ วงศ์ โฮมินิดในกลุ่มไพรเมตซึ่งรวมถึงลิงใหญ่ทั้งหมดตลอดประวัติศาสตร์วิวัฒนาการมนุษย์ค่อยๆ พัฒนาลักษณะต่างๆ...
Early evolution of primates
The evolutionary history of primates can be traced back 65 million years. [ 13 ] [ 14 ] [ 15 ] [ 16 ] [ 17 ] The details of the origins and early evolution of primates are however still unknown to a large extent due to scarcity of fossil evidence.
การแยกสายวิวัฒนาการของมนุษย์จากลิงใหญ่ชนิดอื่นๆ
ฟอสซิลของ Nakalipithecus ที่พบในเคนยาอาจเป็นตัวอย่างของสายพันธุ์ที่ใกล้เคียงกับบรรพบุรุษร่วมสุดท้ายของกอริลลา ชิมแปนซี และมนุษย์ หลักฐานทางโมเลกุลชี้ให้เห็นว่าระหว่าง 8 ถึง 4 ล้านปีก่อน กอริลลาและชิมแปนซี (สกุล Pan ) ได้แยกตัวออกจากสายพันธุ์ที่นำไปสู่มนุษย์...
สกุลออ สตราโลพิเทคัส
สกุล Australopithecus วิวัฒนาการขึ้นในแอฟริกาตะวันออกเมื่อประมาณ 4 ล้านปีก่อน ก่อนที่จะแพร่กระจายไปทั่วทวีปและสูญพันธุ์ไปในที่สุดเมื่อ 2 ล้านปีก่อน ในช่วงเวลานั้นมีออสตราโลพิเทคัสหลายรูปแบบอาศัยอยู่ รวมถึง Australopithecus anamensis , A. afarensis , A.