อ่าน 29 นาที
เอนไซม์
เอนไซม์เป็น โมเลกุลชีวภาพขนาดใหญ่ ซึ่ง โดยทั่วไปเป็น โปรตีน ทำหน้าที่เป็น ตัวเร่งปฏิกิริยา ทางชีวภาพ เร่ง ปฏิกิริยาเคมี โดยไม่ถูกใช้ไปในกระบวนการ โมเลกุล...
เอนไซม์

| ส่วนหนึ่งของชุดบทความเกี่ยวกับ |
| ชีวเคมี |
|---|
 |
|
เอนไซม์เป็นโมเลกุลชีวภาพขนาดใหญ่ซึ่งโดยทั่วไปเป็นโปรตีนทำหน้าที่เป็นตัวเร่งปฏิกิริยา ทางชีวภาพ เร่งปฏิกิริยาเคมีโดยไม่ถูกใช้ไปในกระบวนการโมเลกุลที่เอนไซม์ทำงานด้วยเรียกว่าสารตั้งต้นซึ่งจะถูกเปลี่ยนเป็นผลิตภัณฑ์กระบวนการเผาผลาญเกือบทั้งหมดภายในเซลล์ขึ้นอยู่กับการเร่งปฏิกิริยาของเอนไซม์เพื่อให้เกิดขึ้นในอัตราที่เกี่ยวข้องทางชีวภาพ[ 1 ] : 8.1 วิถีการเผาผลาญโดยทั่วไปประกอบด้วยขั้นตอนที่เร่งปฏิกิริยาโดยเอนไซม์หลายขั้นตอน การศึกษาเกี่ยวกับเอนไซม์เรียกว่าเอนไซม์วิทยาและสาขาที่เกี่ยวข้องมุ่งเน้นไปที่ เอนไซม์ เทียมซึ่งเป็นโปรตีนที่สูญเสียกิจกรรมการเร่งปฏิกิริยา แต่อาจยังคงรักษาหน้าที่ในการควบคุมหรือเป็นโครงสร้าง ซึ่งมักบ่งชี้โดยการเปลี่ยนแปลงใน ลำดับ กรดอะมิโนหรือพฤติกรรม 'เร่งปฏิกิริยาเทียม' ที่ผิดปกติ[ 2 ] [ 3 ]
เอนไซม์เป็นที่รู้จักกันดีว่าสามารถเร่งปฏิกิริยาทางชีวเคมีได้มากกว่า 5,000 ชนิด[ 4 ]ตัวเร่งปฏิกิริยาทางชีวภาพอื่นๆ ได้แก่โมเลกุล RNA ที่เป็นตัวเร่งปฏิกิริยาหรือไรโบไซม์ ซึ่งบางครั้งถูกจัดประเภทเป็นเอนไซม์แม้ว่าจะประกอบด้วย RNA แทนที่จะเป็นโปรตีน เมื่อไม่นานมานี้คอนเดนเซตของโมเลกุลชีวภาพได้รับการยอมรับว่าเป็นตัวเร่งปฏิกิริยาทางชีวภาพประเภทที่สาม ซึ่งสามารถเร่งปฏิกิริยาได้โดยการสร้างส่วนต่อประสานและเกรเดียนต์ เช่น เกรเดียนต์ไอออนิก ที่ขับเคลื่อนกระบวนการทางชีวเคมี แม้ว่าโปรตีนที่เป็นส่วนประกอบจะไม่ใช่ตัวเร่งปฏิกิริยาโดยเนื้อแท้ก็ตาม[ 5 ]
เอนไซม์ช่วยเพิ่มอัตราการเกิดปฏิกิริยา โดยการลด พลังงานกระตุ้นของปฏิกิริยาซึ่งมักจะลดลงหลายล้านเท่า ตัวอย่างที่โดดเด่นคือโอโรทิดีน 5′-ฟอสเฟต ดีคาร์บอกซิเลส ซึ่งเร่งปฏิกิริยาที่ปกติแล้วต้องใช้เวลาหลายล้านปีจึงจะเกิดขึ้นได้ภายในไม่กี่มิลลิวินาที[ 6 ] [ 7 ]เช่นเดียวกับตัวเร่งปฏิกิริยาทั้งหมด เอนไซม์ไม่มีผลต่อสมดุล โดยรวม ของปฏิกิริยาและจะถูกสร้างขึ้นใหม่เมื่อสิ้นสุดแต่ละรอบ สิ่งที่ทำให้เอนไซม์แตกต่างออกไปคือความจำเพาะสูง ซึ่งกำหนดโดยโครงสร้างสามมิติ ที่เป็นเอกลักษณ์ และความไวต่อปัจจัยต่างๆ เช่นอุณหภูมิและpHกิจกรรมของเอนไซม์สามารถเพิ่มขึ้นได้ด้วยตัวกระตุ้นหรือลดลงได้ด้วยตัวยับยั้งซึ่งหลายชนิดทำหน้าที่เป็นยาหรือสารพิษนอกเหนือจากสภาวะที่เหมาะสม เอนไซม์อาจสูญเสียโครงสร้างผ่านการเสียสภาพทำให้สูญเสียการทำงาน
เอนไซม์มีการใช้งานจริงอย่างกว้างขวาง ในอุตสาหกรรม เอนไซม์ถูกใช้เพื่อเร่งปฏิกิริยาการผลิตยาปฏิชีวนะและโมเลกุลที่ซับซ้อนอื่นๆ ในชีวิตประจำวัน เอนไซม์ในผงซักฟอกชีวภาพช่วยย่อยสลายโปรตีน แป้ง และคราบไขมัน ทำให้ประสิทธิภาพในการทำความสะอาดดีขึ้น ปาเปนและเอนไซม์ย่อยโปรตีนอื่นๆ ถูกใช้ในสารทำให้เนื้อนุ่มเพื่อย่อยสลายโปรตีน ปรับปรุงเนื้อสัมผัสและความสามารถในการย่อย ความจำเพาะและประสิทธิภาพของเอนไซม์ทำให้เอนไซม์มีความสำคัญอย่างยิ่งทั้งในระบบชีวภาพและกระบวนการเชิงพาณิชย์

ที่มาและประวัติความเป็นมา
ในช่วงปลายศตวรรษที่ 17 และต้นศตวรรษที่ 18 การย่อยเนื้อสัตว์โดยการหลั่งของกระเพาะอาหาร[ 8 ]และการเปลี่ยนแป้งเป็นน้ำตาลโดยสารสกัดจากพืชและน้ำลายเป็นที่ทราบกันดีอยู่แล้ว แต่กลไกที่ทำให้เกิดสิ่งเหล่านี้ยังไม่ได้รับการระบุ[ 9 ]
นักเคมีชาวฝรั่งเศสAnselme Payenเป็นคนแรกที่ค้นพบเอนไซม์ไดแอสเตสในปี พ.ศ. 2476 [ 10 ]ไม่กี่ทศวรรษต่อมา เมื่อศึกษาการหมักน้ำตาลเป็นแอลกอฮอล์โดยยีสต์Louis Pasteurสรุปว่าการหมักนี้เกิดจากพลังชีวิตที่บรรจุอยู่ภายในเซลล์ยีสต์ที่เรียกว่า "เฟอร์เมนต์" ซึ่งเชื่อกันว่าทำงานได้เฉพาะในสิ่งมีชีวิตเท่านั้น เขาเขียนว่า "การหมักแอลกอฮอล์เป็นกระบวนการที่สัมพันธ์กับชีวิตและการจัดระเบียบของเซลล์ยีสต์ ไม่ใช่กับการตายหรือการเน่าเปื่อยของเซลล์" [ 11 ]
ในปี พ.ศ. 2420 วิลเฮล์ม คูห์เน (ค.ศ. 2480–2443) นักสรีรวิทยาชาวเยอรมัน ได้ใช้คำว่าเอนไซม์ เป็นครั้งแรก ซึ่งมาจากภาษากรีกโบราณἔνζυμον (énzymon) ' ทำให้ขึ้นฟูในยีสต์'เพื่ออธิบายกระบวนการนี้[ 12 ] ต่อมา คำว่าเอนไซม์ถูกนำมาใช้เพื่ออ้างถึงสารที่ไม่มีชีวิต เช่นเปปซินและคำว่าการหมักถูกนำมาใช้เพื่ออ้างถึงกิจกรรมทางเคมีที่ผลิตโดยสิ่งมีชีวิต[ 13 ]

เอดูอาร์ด บูชเนอร์ส่งผลงานวิจัยชิ้นแรกเกี่ยวกับการศึกษาสารสกัดจากยีสต์ในปี 1897 จากการทดลองหลายครั้งที่มหาวิทยาลัยเบอร์ลินเขาพบว่าน้ำตาลถูกหมักโดยสารสกัดจากยีสต์แม้ว่าจะไม่มีเซลล์ยีสต์ที่ยังมีชีวิตอยู่ในส่วนผสมก็ตาม[ 14 ]เขาตั้งชื่อเอนไซม์ที่ทำให้เกิดการหมักซูโครสว่า " ไซเมส " [ 15 ]ในปี 1907 เขาได้รับรางวัลโนเบลสาขาเคมีจาก "การค้นพบการหมักแบบปราศจากเซลล์" ตามแบบอย่างของบูชเนอร์ เอนไซม์มักจะถูกตั้งชื่อตามปฏิกิริยาที่พวกมันทำ: คำต่อท้าย-aseจะรวมกับชื่อของสารตั้งต้น (เช่นแลคเตสคือเอนไซม์ที่ย่อยสลายแลคโตส ) หรือกับประเภทของปฏิกิริยา (เช่นดีเอ็นเอพอลิเมอเรสสร้างพอลิเมอร์ดีเอ็นเอ) [ 16 ]
เอกลักษณ์ทางชีวเคมีของเอนไซม์ยังไม่เป็นที่รู้จักในช่วงต้นทศวรรษ 1900 นักวิทยาศาสตร์หลายคนสังเกตว่ากิจกรรมของเอนไซม์เกี่ยวข้องกับโปรตีน แต่คนอื่นๆ (เช่นริชาร์ด วิลสแตตเตอร์ ผู้ได้รับรางวัลโนเบล ) โต้แย้งว่าโปรตีนเป็นเพียงตัวพาของเอนไซม์ที่แท้จริง และโปรตีนนั้นไม่สามารถเร่งปฏิกิริยาได้ด้วยตัวเอง[ 17 ]ในปี 1926 เจมส์ บี. ซัมเนอร์แสดงให้เห็นว่าเอนไซม์ยูรีเอสเป็นโปรตีนบริสุทธิ์และตกผลึกมันได้ เขาทำเช่นเดียวกันกับเอนไซม์คาตาเลสในปี 1937 ข้อสรุปที่ว่าโปรตีนบริสุทธิ์สามารถเป็นเอนไซม์ได้นั้นได้รับการพิสูจน์อย่างชัดเจนโดยจอห์น ฮาวเวิร์ด นอร์ธรอปและเวนเดลล์ เมเรดิธ สแตนลีย์ผู้ซึ่งทำงานเกี่ยวกับเอนไซม์ย่อยอาหารเปปซิน (1930) ทริปซินและไคโมทริปซินนักวิทยาศาสตร์ทั้งสามคนนี้ได้รับรางวัลโนเบลสาขาเคมีในปี 1946 [ 18 ]
การค้นพบว่าเอนไซม์สามารถตกผลึกได้ในที่สุดก็ทำให้สามารถไขโครงสร้างของเอนไซม์ได้โดยใช้การตกผลึกด้วยรังสีเอกซ์โดยเริ่มแรกทำกับไลโซไซม์ซึ่งเป็นเอนไซม์ที่พบในน้ำตา น้ำลาย และไข่ขาวที่ย่อยสลายเปลือกของแบคทีเรียบางชนิด โครงสร้างของไลโซไซม์ได้รับการไขโดยกลุ่มที่นำโดยเดวิด ชิลตัน ฟิลลิปส์และตีพิมพ์ในปี 1965 [ 19 ]โครงสร้างที่มีความละเอียดสูงของไลโซไซม์นี้ถือเป็นจุดเริ่มต้นของสาขาชีววิทยาเชิงโครงสร้างและความพยายามที่จะเข้าใจว่าเอนไซม์ทำงานอย่างไรในระดับอะตอม[ 20 ]
การจำแนกและการตั้งชื่อ
เอนไซม์สามารถจำแนกได้ตามเกณฑ์หลักสองประการ ได้แก่ กิจกรรมของเอนไซม์ หรือ ความคล้ายคลึงกัน ของลำดับกรดอะมิโน (และด้วยเหตุนี้จึงแสดงถึงความสัมพันธ์ทางวิวัฒนาการ)
กิจกรรมของเอนไซม์
ชื่อของเอนไซม์มักได้มาจากสารตั้งต้นหรือปฏิกิริยาเคมีที่เอนไซม์เร่งปฏิกิริยา โดยคำจะลงท้ายด้วย-ase [ 1 ] : 8.1.3 ตัวอย่างเช่นแลคเตสแอลกอฮอล์ดีไฮโดรจีเนสและดีเอ็นเอพอลิเมอเรสเอนไซม์ที่แตกต่างกันแต่เร่งปฏิกิริยาเคมีเดียวกันเรียกว่าไอโซเอนไซม์[ 1 ] : 10.3
สหภาพระหว่างประเทศด้านชีวเคมีและชีววิทยาโมเลกุลได้พัฒนาระบบการตั้งชื่อสำหรับเอนไซม์ โดยใช้หมายเลข EC (ย่อมาจาก "Enzyme Commission")เอนไซม์แต่ละชนิดจะถูกอธิบายด้วย "EC" ตามด้วยลำดับตัวเลขสี่หลัก ซึ่งแสดงถึงลำดับชั้นของกิจกรรมของเอนไซม์ (จากทั่วไปมากไปจนถึงเฉพาะเจาะจงมาก) กล่าวคือ ตัวเลขแรกจะจัดประเภทเอนไซม์อย่างกว้างๆ ตามกลไกการทำงาน ในขณะที่ตัวเลขอื่นๆ จะเพิ่มความเฉพาะเจาะจงมากขึ้นเรื่อยๆ[ 21 ]
การจัดระดับชั้นสูงสุดคือ:
- EC 1, ออกซิโดรีดักเทส : ทำ หน้าที่เร่งปฏิกิริยา ออกซิเดชัน /รีดักชัน
- EC 2, ทรานสเฟอเรส : ทำหน้าที่ถ่ายโอนหมู่ฟังก์ชัน ( เช่นหมู่เมทิลหรือหมู่ฟอสเฟต)
- EC 3, ไฮโดรเลส : ทำหน้าที่เร่งปฏิกิริยาไฮโดรไลซิสของพันธะต่างๆ
- EC 4, ไลเอส : ทำหน้าที่ตัดพันธะต่างๆ ด้วยวิธีการอื่นนอกเหนือจากไฮโดรไลซิสและออกซิเดชัน
- EC 5, ไอโซเมอเรส : เร่งปฏิกิริยา การเปลี่ยนแปลง ไอโซเมอไรเซชันภายในโมเลกุลเดียว
- EC 6, ไลเกส : ทำหน้าที่เชื่อมโมเลกุลสองโมเลกุลเข้าด้วยกันด้วยพันธะโควาเลนต์
- EC 7, ทรานสโลเคส : ทำหน้าที่เร่งปฏิกิริยาการเคลื่อนย้ายไอออนหรือโมเลกุลผ่านเยื่อหุ้มเซลล์ หรือการแยกไอออนหรือโมเลกุลเหล่านั้นภายในเยื่อหุ้มเซลล์
ส่วนเหล่านี้แบ่งย่อยตามคุณลักษณะอื่นๆ เช่น สารตั้งต้น ผลิตภัณฑ์ และกลไกทางเคมีเอนไซม์จะถูกระบุอย่างครบถ้วนด้วยการกำหนดตัวเลขสี่ตัว ตัวอย่างเช่นเฮกโซไคเนส (EC 2.7.1.1) เป็นทรานสเฟอเรส (EC 2) ที่เพิ่มหมู่ฟอสเฟต (EC 2.7) ให้กับน้ำตาลเฮกโซส ซึ่งเป็นโมเลกุลที่มีหมู่แอลกอฮอล์ (EC 2.7.1) [ 22 ]
ความคล้ายคลึงของลำดับ
หมวดหมู่ EC ไม่ได้สะท้อนถึงความคล้ายคลึงของลำดับ ตัวอย่างเช่น ไลเกสสองตัวที่มีหมายเลข EC เดียวกันซึ่งเร่งปฏิกิริยาเดียวกันเป๊ะ อาจมีลำดับที่แตกต่างกันโดยสิ้นเชิง เอนไซม์ เช่นเดียวกับโปรตีนอื่นๆ ได้ถูกจัดประเภทตามความคล้ายคลึงของลำดับเป็นตระกูลต่างๆ มากมาย โดยไม่ขึ้นอยู่กับหน้าที่ของมัน ตระกูลเหล่านี้ได้รับการบันทึกไว้ในฐานข้อมูลโปรตีนและตระกูลโปรตีนต่างๆ มากมายเช่น Pfam [ 23 ]
เอนไซม์ไอโซฟังก์ชันที่ไม่เหมือนกัน
เอนไซม์ที่ไม่เกี่ยวข้องกันแต่มีกิจกรรมเอนไซม์เดียวกันเรียกว่าเอนไซม์ไอโซฟังก์ชันที่ไม่เหมือนกัน[ 24 ]การถ่ายโอนยีนในแนวนอนอาจแพร่กระจายยีนเหล่านี้ไปยังสายพันธุ์ที่ไม่เกี่ยวข้องกัน โดยเฉพาะแบคทีเรีย ซึ่งยีนเหล่านี้สามารถแทนที่ยีนภายในที่มีฟังก์ชันเดียวกัน ทำให้เกิดการแทนที่ยีนที่ไม่เหมือนกัน
โครงสร้าง

โดยทั่วไปเอนไซม์เป็นโปรตีนทรงกลมทำงานเดี่ยวๆ หรือในสารประกอบ ที่ใหญ่กว่า ลำดับของกรดอะมิโนจะระบุโครงสร้าง ซึ่งจะกำหนดกิจกรรมเร่งปฏิกิริยาของเอนไซม์[ 25 ]แม้ว่าโครงสร้างจะกำหนดหน้าที่ แต่ยังไม่สามารถทำนายกิจกรรมของเอนไซม์ใหม่ได้จากโครงสร้างเพียงอย่างเดียว[ 26 ]โครงสร้างของเอนไซม์จะคลายตัว ( เสียสภาพ ) เมื่อได้รับความร้อนหรือสัมผัสกับสารเคมีที่ทำให้เสียสภาพ และการรบกวนโครงสร้างนี้มักทำให้สูญเสียกิจกรรม[ 27 ]การเสียสภาพของเอนไซม์มักเกี่ยวข้องกับอุณหภูมิที่สูงกว่าระดับปกติของสายพันธุ์นั้นๆ ดังนั้น เอนไซม์จากแบคทีเรียที่อาศัยอยู่ในสภาพแวดล้อมภูเขาไฟ เช่นน้ำพุร้อนจึงเป็นที่ต้องการของผู้ใช้ในอุตสาหกรรมเนื่องจากความสามารถในการทำงานที่อุณหภูมิสูง ทำให้ปฏิกิริยาที่เร่งด้วยเอนไซม์สามารถดำเนินการได้ในอัตราที่สูงมาก
โดยทั่วไปเอนไซม์จะมีขนาดใหญ่กว่าสารตั้งต้นมาก ขนาดมีตั้งแต่เพียง 62 หน่วยกรดอะมิโน สำหรับโมโนเมอร์ของ4-oxalocrotonate tautomerase [ 28 ] ไปจนถึงมากกว่า 2,500 หน่วยในเอนไซม์สังเคราะห์กรดไขมันของสัตว์[ 29 ]มีเพียงส่วนเล็ก ๆ ของโครงสร้าง (ประมาณ 2–4 กรดอะมิโน) เท่านั้นที่เกี่ยวข้องโดยตรงกับการเร่งปฏิกิริยา: ตำแหน่งเร่งปฏิกิริยา[ 30 ]ตำแหน่งเร่งปฏิกิริยานี้ตั้งอยู่ถัดจากตำแหน่งการจับ หนึ่งตำแหน่งหรือมากกว่า ซึ่งกรดอะมิโนจะจัดเรียงสารตั้งต้น ตำแหน่งเร่งปฏิกิริยาและตำแหน่งการจับรวมกันเป็น ตำแหน่งออกฤทธิ์ของเอนไซม์โครงสร้างเอนไซม์ส่วนใหญ่ที่เหลือทำหน้าที่รักษาทิศทางที่แม่นยำและพลวัตของตำแหน่งออกฤทธิ์[ 31 ]
ในเอนไซม์บางชนิด กรดอะมิโนไม่ได้มีส่วนเกี่ยวข้องโดยตรงกับการเร่งปฏิกิริยา แต่เอนไซม์จะมีตำแหน่งสำหรับจับและจัดวางโคแฟคเตอร์เร่ง ปฏิกิริยา [ 31 ]โครงสร้างของเอนไซม์อาจมีตำแหน่งอัลโลสเตอริกซึ่งการจับของโมเลกุลขนาดเล็กทำให้เกิดการเปลี่ยนแปลงโครงสร้างที่เพิ่มหรือลดกิจกรรม[ 32 ]
มี ตัวเร่งปฏิกิริยาทางชีวภาพที่ใช้RNAจำนวนเล็กน้อย ที่เรียกว่า ไรโบไซม์ซึ่งสามารถทำงานได้ด้วยตัวเองหรือร่วมกับโปรตีนก็ได้ ไรโบโซม เป็นส่วนประกอบที่พบได้บ่อยที่สุด ซึ่งเป็นส่วนประกอบของโปรตีนและ RNA ที่ทำหน้าที่เร่งปฏิกิริยา[ 1 ] : 2.2
กลไก

การยึดเกาะของสารตั้งต้น
เอนไซม์ต้องจับกับสารตั้งต้นก่อนจึงจะสามารถเร่งปฏิกิริยาเคมีได้ เอนไซม์มักมีความจำเพาะสูงมากต่อสารตั้งต้นที่มันจับและปฏิกิริยาเคมีที่เร่งความจำเพาะนี้เกิดขึ้นได้จากการจับกับช่องว่างที่มีรูปร่าง ประจุ และ ลักษณะ ไฮโดรฟิลิก / ไฮโดรโฟบิก ที่เข้ากันได้ กับสารตั้งต้น ดังนั้นเอนไซม์จึงสามารถแยกแยะโมเลกุลของสารตั้งต้นที่คล้ายคลึงกันมากได้ เพื่อให้เกิดความจำเพาะทางเคมีความจำเพาะทางตำแหน่งและ ความ จำเพาะทางสเตอริโอ[ 33 ]
เอนไซม์บางชนิดที่แสดงความจำเพาะและความแม่นยำสูงสุดเกี่ยวข้องกับการคัดลอกและการแสดงออกของจีโนมเอนไซม์เหล่านี้บางชนิดมีกลไก " การตรวจสอบความถูกต้อง " โดยเอนไซม์เช่นDNA polymeraseจะเร่งปฏิกิริยาในขั้นตอนแรก จากนั้นตรวจสอบว่าผลิตภัณฑ์ถูกต้องในขั้นตอนที่สอง[ 34 ]กระบวนการสองขั้นตอนนี้ส่งผลให้อัตราข้อผิดพลาดเฉลี่ยน้อยกว่า 1 ข้อผิดพลาดใน 100 ล้านปฏิกิริยาใน polymerase ของสัตว์เลี้ยงลูกด้วยนมที่มีความแม่นยำสูง[ 1 ] : 5.3.1 กลไกการตรวจสอบความ ถูกต้องที่คล้ายกันนี้ยังพบในRNA polymerase [ 35 ] aminoacyl tRNA synthetases [ 36 ]และไรโบโซม[ 37 ]
ในทางกลับกัน เอนไซม์บางชนิดแสดงคุณสมบัติการทำงานที่หลากหลาย โดยมีความจำเพาะกว้างและทำงานบนสารตั้งต้นที่เกี่ยวข้องทางสรีรวิทยาที่แตกต่างกันหลายชนิด เอนไซม์หลายชนิดมีกิจกรรมข้างเคียงเล็กน้อยซึ่งเกิดขึ้นโดยบังเอิญ (เช่นเป็นกลาง ) ซึ่งอาจเป็นจุดเริ่มต้นสำหรับการคัดเลือกเชิงวิวัฒนาการของฟังก์ชันใหม่[ 38 ] [ 39 ]

แบบจำลอง "ล็อคและกุญแจ"
เพื่ออธิบายความจำเพาะของเอนไซม์ที่สังเกตได้ ในปี พ.ศ. 2437 Emil Fischerเสนอว่าทั้งเอนไซม์และสารตั้งต้นมีรูปร่างทางเรขาคณิตที่เสริมกันอย่างเฉพาะเจาะจงซึ่งเข้ากันได้พอดี[ 40 ] [ 41 ]ซึ่งมักเรียกกันว่าแบบจำลอง "กุญแจและแม่กุญแจ" [ 1 ] : 8.3.2 แบบจำลองในยุคแรกนี้อธิบายความจำเพาะของเอนไซม์ได้ แต่ไม่สามารถอธิบายการทำให้สถานะการเปลี่ยนผ่านมีเสถียรภาพซึ่งเอนไซม์บรรลุได้[ 42 ]
แบบจำลองความพอดีที่เหนี่ยวนำ
ในปี พ.ศ. 2491 แดเนียล คอชแลนด์เสนอการปรับเปลี่ยนแบบจำลองกุญแจและแม่กุญแจ: เนื่องจากเอนไซม์เป็นโครงสร้างที่ค่อนข้างยืดหยุ่น บริเวณออกฤทธิ์จึงถูกปรับเปลี่ยนรูปร่างอย่างต่อเนื่องโดยปฏิสัมพันธ์กับสารตั้งต้นในขณะที่สารตั้งต้นมีปฏิสัมพันธ์กับเอนไซม์[ 43 ]ผลก็คือ สารตั้งต้นไม่ได้เพียงแค่จับกับบริเวณออกฤทธิ์ที่แข็งตัวเท่านั้น แต่โซ่ข้าง ของกรดอะมิโน ที่ประกอบเป็นบริเวณออกฤทธิ์จะถูกหล่อหลอมให้อยู่ในตำแหน่งที่แม่นยำซึ่งช่วยให้เอนไซม์สามารถทำหน้าที่เร่งปฏิกิริยาได้ ในบางกรณี เช่นไกลโคซิเดสโมเลกุลของสารตั้งต้นจะเปลี่ยนรูปร่างเล็กน้อยเมื่อเข้าสู่บริเวณออกฤทธิ์[ 44 ]บริเวณออกฤทธิ์จะเปลี่ยนแปลงต่อไปจนกว่าสารตั้งต้นจะถูกจับอย่างสมบูรณ์ ณ จุดนั้น รูปร่างสุดท้ายและการกระจายประจุจะถูกกำหนด[ 45 ] การปรับตัวแบบเหนี่ยวนำอาจช่วยเพิ่มความแม่นยำของการจดจำโมเลกุลในสภาวะที่มีการแข่งขันและสัญญาณรบกวนผ่านกลไกการตรวจสอบความถูกต้องของโครงสร้าง[ 46 ]
การเร่งปฏิกิริยา
เอนไซม์สามารถเร่งปฏิกิริยาได้หลายวิธี ซึ่งทั้งหมดนี้ช่วยลดพลังงานกระตุ้น (ΔG ‡ พลังงานอิสระของกิบส์ ) [ 47 ]
- โดยการทำให้สถานะการเปลี่ยนผ่านมีเสถียรภาพ:
- สร้างสภาพแวดล้อมที่มีการกระจายประจุที่เสริมกับสถานะการเปลี่ยนผ่านเพื่อลดพลังงาน[ 48 ]
- โดยการเสนอเส้นทางปฏิกิริยาทางเลือก:
- ทำปฏิกิริยาชั่วคราวกับสารตั้งต้น ก่อให้เกิดตัวกลางโคเวเลนต์เพื่อให้สถานะการเปลี่ยนผ่านพลังงานต่ำลง[ 49 ]
- โดยการทำให้สถานะพื้นฐานของสารตั้งต้นไม่เสถียร:
- บิดเบือนสารตั้งต้นที่ผูกติดให้เป็นรูปแบบสถานะการเปลี่ยนผ่านเพื่อลดพลังงานที่จำเป็นในการเข้าถึงสถานะการเปลี่ยนผ่าน[ 50 ]
- โดยการจัดเรียงสารตั้งต้นให้เป็นรูปแบบที่มีประสิทธิภาพเพื่อลดการเปลี่ยนแปลงเอนโทรปี ของปฏิกิริยา [ 51 ] (การมีส่วนร่วมของกลไกนี้ต่อการเร่งปฏิกิริยาค่อนข้างน้อย) [ 52 ]
เอนไซม์อาจใช้กลไกเหล่านี้หลายอย่างพร้อมกัน ตัวอย่างเช่นโปรตีเอ ส เช่นทริปซินทำการเร่งปฏิกิริยาแบบโควาเลนต์โดยใช้ไตรแอดเร่งปฏิกิริยา ทำให้การสะสมประจุบนสถานะเปลี่ยนผ่านมีเสถียรภาพโดยใช้ช่องออกซิแอนไอออน และทำการ ไฮโดรไลซิสให้สมบูรณ์โดยใช้สารตั้งต้นน้ำที่มีทิศทาง[ 53 ]
พลวัต
เอนไซม์ไม่ใช่โครงสร้างที่แข็งและคงที่ แต่มีกลไกการเคลื่อนไหวภายในที่ซับซ้อน นั่นคือ การเคลื่อนไหวของส่วนต่างๆ ของโครงสร้างเอนไซม์ เช่น กรดอะมิโนแต่ละตัว กลุ่มของกรดอะมิโนที่ก่อตัวเป็นลูปโปรตีนหรือหน่วยโครงสร้างทุติย ภูมิ หรือแม้แต่ โดเมนโปรตีนทั้งหมดการเคลื่อนไหวเหล่านี้ก่อให้เกิดกลุ่มโครงสร้างที่แตกต่างกันเล็กน้อยซึ่งสามารถเปลี่ยนไปมาระหว่างกันได้ที่สมดุลสถานะต่างๆ ภายในกลุ่มนี้อาจเกี่ยวข้องกับแง่มุมต่างๆ ของการทำงานของเอนไซม์ ตัวอย่างเช่น โครงสร้างที่แตกต่างกันของเอนไซม์ไดไฮโดรโฟเลตเรดักเท ส เกี่ยวข้องกับการจับกับสารตั้งต้น การเร่งปฏิกิริยา การปลดปล่อยโคแฟคเตอร์ และการปลดปล่อยผลิตภัณฑ์ของวัฏจักรเร่งปฏิกิริยา[ 54 ]ซึ่งสอดคล้องกับทฤษฎีเรโซแนนซ์เร่งปฏิกิริยาการเปลี่ยนผ่านระหว่างโครงสร้างที่แตกต่างกันในระหว่างวัฏจักรเร่งปฏิกิริยาเกี่ยวข้องกับ การเคลื่อนไหว แบบยืดหยุ่นหนืด ภายใน ซึ่งอำนวยความสะดวกโดย บริเวณที่ มีความเครียด สูง ซึ่งกรดอะมิโนมีการจัดเรียงใหม่[ 55 ]
การนำเสนอพื้นผิว
การนำเสนอซับสเตรตเป็นกระบวนการที่เอนไซม์ถูกแยกออกจากซับสเตรต เอนไซม์สามารถถูกแยกไปยังเยื่อหุ้มพลาสมาโดยแยกจากซับสเตรตในนิวเคลียสหรือไซโตซอล[ 56 ]หรือภายในเยื่อหุ้มเซลล์ เอนไซม์สามารถถูกแยกเข้าไปในลิพิดแรฟต์โดยแยกจากซับสเตรตในบริเวณที่ไม่เป็นระเบียบ เมื่อเอนไซม์ถูกปล่อยออกมา มันจะผสมกับซับสเตรต หรืออีกทางหนึ่ง เอนไซม์สามารถถูกแยกไว้ใกล้กับซับสเตรตเพื่อกระตุ้นเอนไซม์ ตัวอย่างเช่น เอนไซม์สามารถละลายได้ และเมื่อถูกกระตุ้นจะจับกับลิพิดในเยื่อหุ้มพลาสมา จากนั้นจึงออกฤทธิ์ต่อโมเลกุลในเยื่อหุ้มพลาสมา[ 57 ]
การปรับเปลี่ยนแบบอัลโลสเตอริก
ตำแหน่งอัลโลสเตอริกเป็นช่องว่างบนเอนไซม์ ซึ่งแตกต่างจากตำแหน่งออกฤทธิ์ ที่จับกับโมเลกุลในสภาพแวดล้อมของเซลล์ โมเลกุลเหล่านี้จะทำให้เกิดการเปลี่ยนแปลงในโครงสร้างหรือพลวัตของเอนไซม์ ซึ่งจะถูกส่งต่อไปยังตำแหน่งออกฤทธิ์และส่งผลต่ออัตราการเกิดปฏิกิริยาของเอนไซม์[ 58 ]ด้วยวิธีนี้ ปฏิสัมพันธ์แบบอัลโลสเตอริกสามารถยับยั้งหรือกระตุ้นเอนไซม์ได้ ปฏิสัมพันธ์แบบอัลโลสเตอริกกับเมตาบอไลต์ต้นน้ำหรือปลายน้ำในวิถีเมตาบอลิซึมของเอนไซม์ทำให้เกิด การควบคุม แบบป้อนกลับเปลี่ยนแปลงกิจกรรมของเอนไซม์ตามการไหลผ่านส่วนที่เหลือของวิถี[ 59 ]
โคแฟกเตอร์

เอนไซม์บางชนิดไม่ต้องการส่วนประกอบเพิ่มเติมเพื่อแสดงกิจกรรมเต็มที่ ส่วนเอนไซม์อื่นๆ ต้องการโมเลกุลที่ไม่ใช่โปรตีนที่เรียกว่าโคแฟคเตอร์เพื่อจับกับเอนไซม์เพื่อให้เกิดกิจกรรม[ 60 ]โคแฟคเตอร์อาจเป็น สารประกอบ อนินทรีย์ (เช่นไอออน โลหะ และคลัสเตอร์เหล็ก-กำมะถัน ) หรือสารประกอบอินทรีย์ (เช่นฟลาวินและฮีม ) โคแฟคเตอร์เหล่านี้มีหน้าที่หลายอย่าง ตัวอย่างเช่น ไอออนโลหะสามารถช่วยในการทำให้สปีชีส์นิวคลีโอฟิลิกมีเสถียรภาพภายในบริเวณออกฤทธิ์[ 61 ]โคแฟคเตอร์อินทรีย์อาจเป็นโคเอนไซม์ซึ่งถูกปล่อยออกมาจากบริเวณออกฤทธิ์ของเอนไซม์ระหว่างปฏิกิริยา หรือกลุ่มโปรสเตติกซึ่งจับกับเอนไซม์อย่างแน่นหนา กลุ่มโปรสเตติกอินทรีย์สามารถจับกันด้วยพันธะโควาเลนต์ได้ (เช่นไบโอตินในเอนไซม์เช่นไพรูเวตคาร์บอก ซิเลส ) [ 62 ]
ตัวอย่างของเอนไซม์ที่มีโคแฟคเตอร์คือคาร์บอนิกแอนไฮดราสซึ่งใช้โคแฟคเตอร์สังกะสีที่จับเป็นส่วนหนึ่งของตำแหน่งออกฤทธิ์[ 63 ]ไอออนหรือโมเลกุลที่จับแน่นเหล่านี้มักพบในตำแหน่งออกฤทธิ์และเกี่ยวข้องกับการเร่งปฏิกิริยา[ 1 ] : 8.1.1 ตัวอย่างเช่น โคแฟคเตอร์ฟลาวินและฮีมมักเกี่ยวข้องกับปฏิกิริยารีดอกซ์[ 1 ] : 17
เอนไซม์ที่ต้องการโคแฟคเตอร์แต่ไม่มีโคแฟคเตอร์จับอยู่เรียกว่าอะโปเอนไซม์หรืออะโปโปรตีนเอนไซม์ที่รวมกับโคแฟคเตอร์ที่จำเป็นสำหรับการทำงานเรียกว่าโฮโลเอนไซม์ (หรือฮาโลเอนไซม์) คำว่าโฮโลเอนไซม์ยังสามารถใช้กับเอนไซม์ที่มีโปรตีนย่อยหลายหน่วย เช่น ดีเอ็นเอพอลิเมอเรสซึ่งในกรณีนี้ โฮโลเอนไซม์คือสารประกอบที่สมบูรณ์ซึ่งประกอบด้วยหน่วยย่อยทั้งหมดที่จำเป็นสำหรับการทำงาน[ 1 ] : 8.1.1
โคเอนไซม์
โคเอนไซม์เป็นโมเลกุลอินทรีย์ขนาดเล็กที่สามารถจับกับเอนไซม์ได้อย่างหลวมๆ หรือแน่นหนา โคเอนไซม์ทำหน้าที่ขนส่งกลุ่มเคมีจากเอนไซม์หนึ่งไปยังอีกเอนไซม์หนึ่ง[ 64 ]ตัวอย่างเช่นNADH , NADPHและอะดีโนซีนไตรฟอสเฟต (ATP) โคเอนไซม์บางชนิด เช่นฟลาวินโมโนนิวคลี โอไทด์ (FMN), ฟลาวินอะดีนีนไดนิวคลีโอไทด์ (FAD), ไทอามีนไพ โรฟอสเฟต ( TPP) และเตตระไฮโดรโฟเลต( THF) ได้มาจากวิตามินโคเอนไซม์เหล่านี้ไม่สามารถสังเคราะห์ขึ้นใหม่ได้ในร่างกายและสารประกอบที่เกี่ยวข้องอย่างใกล้ชิด (วิตามิน) ต้องได้รับจากอาหาร กลุ่มเคมีที่ขนส่ง ได้แก่:
- ไอออนไฮไดรด์ (H − ) ซึ่งถูกขนส่งโดยNAD หรือ NADP +
- หมู่ฟอสเฟตที่อยู่บนอะดีโนซีนไตรฟอสเฟต
- หมู่แอเซทิลที่อยู่บนโคเอนไซม์เอ
- หมู่ฟอร์มิล เมทิล หรือเมทิล ที่นำโดยกรดโฟลิกและ
- หมู่เมทิลที่นำโดยS-adenosylmethionine [ 64 ]
เนื่องจากโคเอนไซม์มีการเปลี่ยนแปลงทางเคมีอันเป็นผลมาจากการทำงานของเอนไซม์ จึงเป็นประโยชน์ที่จะพิจารณาว่าโคเอนไซม์เป็นสารตั้งต้นประเภทพิเศษ หรือสารตั้งต้นตัวที่สอง ซึ่งพบได้ในเอนไซม์หลายชนิด ตัวอย่างเช่น เป็นที่ทราบกันว่ามีเอนไซม์ประมาณ 1,000 ชนิดที่ใช้โคเอนไซม์ NADH [ 65 ]
โดยปกติแล้วโคเอนไซม์จะถูกสร้างขึ้นใหม่อย่างต่อเนื่องและรักษาระดับความเข้มข้นให้คงที่ภายในเซลล์ ตัวอย่างเช่น NADPH จะถูกสร้างขึ้นใหม่ผ่านทางวิถีเพนโทสฟอสเฟตและS -adenosylmethionine โดยเมไทโอนีนอะดีโนซิลทรานสเฟอเรส การสร้างใหม่อย่างต่อเนื่องนี้หมายความว่าโคเอนไซม์จำนวนเล็กน้อยสามารถถูกนำไปใช้อย่างเข้มข้นได้ ตัวอย่างเช่น ร่างกายมนุษย์สร้าง ATP ได้ถึงน้ำหนักตัวของตัวเองในแต่ละวัน[ 66 ]
อุณหพลศาสตร์
เช่นเดียวกับตัวเร่งปฏิกิริยาทั้งหมด เอนไซม์ไม่เปลี่ยนแปลงตำแหน่งของสมดุลทางเคมีของปฏิกิริยา ในการมีอยู่ของเอนไซม์ ปฏิกิริยาจะดำเนินไปในทิศทางเดียวกับที่เกิดขึ้นโดยไม่มีเอนไซม์ เพียงแต่เร็วกว่า[ 1 ] : 8.2.3 ตัวอย่างเช่นคาร์บอนิกแอนไฮดราสเร่งปฏิกิริยาในทิศทางใดก็ได้ขึ้นอยู่กับความเข้มข้นของสารตั้งต้น: [ 67 ]
| (ในเนื้อเยื่อ ; ความเข้มข้นของ CO2 สูง ) | 1 |
![{\displaystyle {\mathrm {CO} {\vphantom {A}__{\smash[{t}]{2}}{}+{}\mathrm {H} {\vphantom {A}}_{\smash[{t}]{2}}\mathrm {O} {}\mathrel {\xrightarrow {\text{คาร์บอนิกแอนไฮไดเรส}} } {}\mathrm {H} {\vphantom {A}__{\smash[{t}]{2}}\mathrm {CO} {\vphantom {A}__{\smash[{t}]{3}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/cb4c8837b26e96fe552c17d863f93e0618cd998b)
| (ในปอด ; ความเข้มข้นของ CO2 ต่ำ ) | 2 |
![{\displaystyle {\mathrm {CO} {\vphantom {A}__{\smash[{t}]{2}}{}+{}\mathrm {H} {\vphantom {A}}_{\smash[{t}]{2}}\mathrm {O} {}\mathrel {\xleftarrow {\text{คาร์บอนิกแอนไฮไดเรส}} } {}\mathrm {H} {\vphantom {A}__{\smash[{t}]{2}}\mathrm {CO} {\vphantom {A}__{\smash[{t}]{3}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/618e95485aa1c3c44a29c557ac448ae5b544ff07)
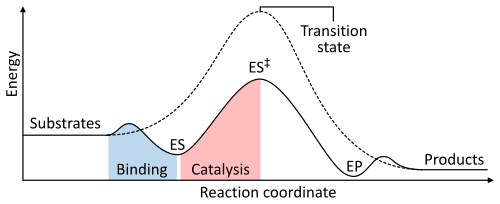
อัตราการเกิดปฏิกิริยาขึ้นอยู่กับพลังงานกระตุ้นที่จำเป็นในการสร้างสถานะเปลี่ยนผ่านซึ่งจะสลายตัวเป็นผลิตภัณฑ์ เอนไซม์ช่วยเพิ่มอัตราการเกิดปฏิกิริยาโดยการลดพลังงานของสถานะเปลี่ยนผ่าน ขั้นแรก การจับกันจะก่อให้เกิดสารเชิงซ้อนเอนไซม์-ซับสเตรต (ES) ที่มีพลังงานต่ำ ขั้นที่สอง เอนไซม์จะทำให้สถานะเปลี่ยนผ่านมีเสถียรภาพมากขึ้น ทำให้ต้องใช้พลังงานน้อยลงในการเกิดขึ้นเมื่อเทียบกับปฏิกิริยาที่ไม่มีตัวเร่งปฏิกิริยา (ES ‡ ) สุดท้าย สารเชิงซ้อนเอนไซม์-ผลิตภัณฑ์ (EP) จะแยกตัวออกเพื่อปลดปล่อยผลิตภัณฑ์[ 1 ] : 8.3
เอนไซม์สามารถเชื่อมโยงปฏิกิริยาสองปฏิกิริยาขึ้นไปได้ เพื่อให้ปฏิกิริยาที่เอื้อต่ออุณหพลศาสตร์สามารถใช้ "ขับเคลื่อน" ปฏิกิริยาที่ไม่เอื้อต่ออุณหพลศาสตร์ได้ เพื่อให้พลังงานรวมของผลิตภัณฑ์ต่ำกว่าสารตั้งต้น ตัวอย่างเช่น การไฮโดรไลซิสของATPมักถูกใช้เพื่อขับเคลื่อนปฏิกิริยาเคมีอื่นๆ[ 68 ]
จลนศาสตร์


จลนพลศาสตร์ของเอนไซม์คือการศึกษาว่าเอนไซม์จับกับสารตั้งต้นและเปลี่ยนให้เป็นผลิตภัณฑ์ได้อย่างไร[ 69 ]ข้อมูลอัตราที่ใช้ในการวิเคราะห์จลนพลศาสตร์มักได้มาจากการทดสอบเอนไซม์ในปี 1913 Leonor MichaelisและMaud Leonora Mentenได้เสนอทฤษฎีเชิงปริมาณของจลนพลศาสตร์ของเอนไซม์ ซึ่งเรียกว่าจลนพลศาสตร์ของ Michaelis–Menten [ 70 ] ผลงานสำคัญของ Michaelis และ Menten คือการคิดถึงปฏิกิริยาของเอนไซม์ในสองขั้นตอน ในขั้นตอนแรก สารตั้งต้นจะจับกับเอนไซม์แบบย้อนกลับได้ ก่อให้เกิดสารเชิงซ้อนเอนไซม์-สารตั้งต้น ซึ่งบางครั้งเรียกว่าสารเชิงซ้อน Michaelis–Menten เพื่อเป็นเกียรติแก่พวกเขา จากนั้นเอนไซม์จะเร่งปฏิกิริยาทางเคมีและปล่อยผลิตภัณฑ์ออกมา งานนี้ได้รับการพัฒนาเพิ่มเติมโดยG. E. BriggsและJ. B. S. Haldaneซึ่งได้สมการจลนพลศาสตร์ที่ยังคงใช้กันอย่างแพร่หลายในปัจจุบัน[ 71 ]
อัตราการทำงานของเอนไซม์ขึ้นอยู่กับ สภาวะ ของสารละลายและความเข้มข้น ของสารตั้งต้น ในการหาความเร็วสูงสุดของปฏิกิริยาเอนไซม์ จะต้องเพิ่มความเข้มข้นของสารตั้งต้นจนกว่าจะเห็นอัตราการเกิดผลิตภัณฑ์คงที่ ซึ่งแสดงไว้ในเส้นโค้งความอิ่มตัวทางด้านขวา ความอิ่มตัวเกิดขึ้นเนื่องจากเมื่อความเข้มข้นของสารตั้งต้นเพิ่มขึ้น เอนไซม์อิสระจะถูกเปลี่ยนเป็นสารเชิงซ้อน ES ที่จับกับสารตั้งต้นมากขึ้นเรื่อยๆ ที่อัตราการเกิดปฏิกิริยาสูงสุด ( Vmax )ของเอนไซม์ ตำแหน่งที่ใช้งานของเอนไซม์ทั้งหมดจะถูกจับกับสารตั้งต้น และปริมาณของสารเชิงซ้อน ES จะเท่ากับปริมาณเอนไซม์ทั้งหมด[ 1 ] : 8.4
Vmaxเป็นเพียงพารามิเตอร์จลนศาสตร์ที่สำคัญตัวหนึ่งเท่านั้น ปริมาณของสารตั้งต้นที่จำเป็นเพื่อให้ได้อัตราการเกิดปฏิกิริยาที่กำหนดก็มีความสำคัญเช่นกัน ซึ่งกำหนดโดยค่าคงที่ Michaelis–Menten ( Km ) ซึ่งเป็นความเข้มข้นของสารตั้งต้นที่จำเป็นสำหรับเอนไซม์ในการเข้าถึงอัตราการเกิดปฏิกิริยาครึ่งหนึ่งของอัตราสูงสุด โดยทั่วไป เอนไซม์แต่ละชนิดจะมีค่าKm ที่เป็นลักษณะ เฉพาะสำหรับสารตั้งต้นที่กำหนด อีกค่าคงที่ที่มีประโยชน์คือkcat หรือที่เรียกว่าเลขการหมุนเวียนซึ่งเป็นจำนวนโมเลกุลของสารตั้งต้นที่ถูกจัดการโดยไซต์ที่ใช้งานหนึ่งไซต์ต่อวินาที[ 1 ] : 8.4
The efficiency of an enzyme can be expressed in terms of kcat/Km. This is also called the specificity constant and incorporates the rate constants for all steps in the reaction up to and including the first irreversible step. Because the specificity constant reflects both affinity and catalytic ability, it is useful for comparing different enzymes against each other, or the same enzyme with different substrates. The theoretical maximum for the specificity constant is called the diffusion limit and is about 108 to 109 (M−1 s−1). At this point every collision of the enzyme with its substrate will result in catalysis, and the rate of product formation is not limited by the reaction rate but by the diffusion rate. Enzymes with this property are called catalytically perfect or kinetically perfect. Example of such enzymes are triose-phosphate isomerase, carbonic anhydrase, acetylcholinesterase, catalase, fumarase, β-lactamase, and superoxide dismutase.[1]: 8.4.2 The turnover of such enzymes can reach several million reactions per second.[1]: 9.2 But most enzymes are far from perfect: the average values of and are about and , respectively.[72]




Michaelis–Menten kinetics relies on the law of mass action, which is derived from the assumptions of free diffusion and thermodynamically driven random collision. Many biochemical or cellular processes deviate significantly from these conditions, because of macromolecular crowding and constrained molecular movement.[73] More recent, complex extensions of the model attempt to correct for these effects.[74]
Inhibition


Enzyme reaction rates can be decreased by various types of enzyme inhibitors.[75]: 73–74
Types of inhibition
Competitive
สารยับยั้งการแข่งขันและสารตั้งต้นไม่สามารถจับกับเอนไซม์ได้ในเวลาเดียวกัน[ 76 ]บ่อยครั้งที่สารยับยั้งการแข่งขันมีลักษณะคล้ายกับสารตั้งต้นที่แท้จริงของเอนไซม์อย่างมาก ตัวอย่างเช่น ยาเมโทเทรกเซตเป็นสารยับยั้งการแข่งขันของเอนไซม์ไดไฮโดรโฟเลตเรดักเทสซึ่งเร่งปฏิกิริยารีดักชันของไดไฮโดรโฟเลตเป็นเตตระไฮโดรโฟเลต[ 77 ]ความคล้ายคลึงกันระหว่างโครงสร้างของไดไฮโดรโฟเลตและยานี้แสดงอยู่ในรูปที่แนบมาด้วย การยับยั้งประเภทนี้สามารถเอาชนะได้ด้วยความเข้มข้นของสารตั้งต้นที่สูง ในบางกรณี สารยับยั้งสามารถจับกับตำแหน่งอื่นที่ไม่ใช่ตำแหน่งการจับของสารตั้งต้นปกติ และออกฤทธิ์แบบอัลโลสเตอริกเพื่อเปลี่ยนรูปร่างของตำแหน่งการจับปกติ[ 78 ]
ไม่แข่งขัน
สารยับยั้งแบบไม่แข่งขันจะจับกับตำแหน่งอื่นที่ไม่ใช่ตำแหน่งที่สารตั้งต้นจับ สารตั้งต้นยังคงจับด้วยความสัมพันธ์ตามปกติ ดังนั้น K mจึงยังคงเท่าเดิม อย่างไรก็ตาม สารยับยั้งจะลดประสิทธิภาพการเร่งปฏิกิริยาของเอนไซม์ ทำให้ V maxลดลง ในทางตรงกันข้ามกับการยับยั้งแบบแข่งขัน การยับยั้งแบบไม่แข่งขันไม่สามารถเอาชนะได้ด้วยความเข้มข้นของสารตั้งต้นสูง[ 75 ] : 76–78
ไม่สามารถแข่งขันได้
สารยับยั้งที่ไม่แข่งขันไม่สามารถจับกับเอนไซม์อิสระได้ แต่จะจับกับเอนไซม์-ซับสเตรตคอมเพล็กซ์เท่านั้น ดังนั้น สารยับยั้งประเภทนี้จึงมีประสิทธิภาพมากที่สุดที่ความเข้มข้นของซับสเตรตสูง ในกรณีที่มีสารยับยั้ง เอนไซม์-ซับสเตรตคอมเพล็กซ์จะไม่ทำงาน[ 75 ] : 78 การยับยั้งประเภทนี้เกิดขึ้นได้ยาก[ 79 ]
ผสม
สารยับยั้งแบบผสมจะจับกับไซต์อัลโลสเตอริก และการจับกันของสารตั้งต้นและสารยับยั้งจะส่งผลกระทบต่อกันและกัน การทำงานของเอนไซม์จะลดลงแต่ไม่หมดไปเมื่อจับกับสารยับยั้ง สารยับยั้งประเภทนี้ไม่เป็นไปตามสมการ Michaelis–Menten [ 75 ] : 76–78
ไม่สามารถย้อนกลับได้
สารยับยั้งที่ไม่สามารถย้อนกลับได้จะทำให้เอนไซม์ไม่ทำงานอย่างถาวร โดยปกติจะสร้างพันธะโควาเลนต์กับโปรตีน[ 80 ]เพนิซิลลิน[ 81 ]และแอสไพริน[ 82 ]เป็นยาที่ใช้กันทั่วไปซึ่งออกฤทธิ์ในลักษณะนี้
หน้าที่ของสารยับยั้ง
ในสิ่งมีชีวิตหลายชนิด สารยับยั้งอาจทำหน้าที่เป็นส่วนหนึ่งของ กลไก ป้อนกลับหากเอนไซม์ผลิตสารหนึ่งมากเกินไปในสิ่งมีชีวิต สารนั้นอาจทำหน้าที่เป็นสารยับยั้งเอนไซม์ที่จุดเริ่มต้นของเส้นทางที่ผลิตสารนั้น ทำให้การผลิตสารนั้นช้าลงหรือหยุดลงเมื่อมีปริมาณเพียงพอ นี่เป็นรูปแบบหนึ่งของการป้อนกลับเชิงลบเส้นทางการเผาผลาญหลัก เช่นวัฏจักรกรดซิตริกใช้กลไกนี้[ 1 ] : 17.2.2
เนื่องจากสารยับยั้งจะปรับเปลี่ยนการทำงานของเอนไซม์ จึงมักถูกนำมาใช้เป็นยา ยาหลายชนิดเป็นสารยับยั้งแบบแข่งขันที่ผันกลับได้ ซึ่งมีลักษณะคล้ายกับสารตั้งต้นดั้งเดิมของเอนไซม์ เช่นเมโทเทรกเซตข้างต้น ตัวอย่างที่รู้จักกันดีอื่นๆ ได้แก่สแตตินที่ใช้รักษาคอเลสเตอรอล สูง [ 83 ]และสารยับยั้งโปรตีเอส ที่ ใช้รักษาการ ติดเชื้อไวรัส เรโทรเช่นเอชไอวี[ 84 ]ตัวอย่างทั่วไปของสารยับยั้งที่ไม่สามารถผันกลับได้ซึ่งใช้เป็นยาคือแอสไพรินซึ่งยับยั้ง เอนไซม์ COX-1และ COX-2 ที่ผลิตสารสื่อการอักเสบอย่างโปรสตาแกลนดิน[ 82 ]สารยับยั้งเอนไซม์อื่นๆ เป็นสารพิษ ตัวอย่างเช่น สารพิษไซยาไนด์เป็นสารยับยั้งเอนไซม์ที่ไม่สามารถผันกลับได้ ซึ่งรวมตัวกับทองแดงและเหล็กในบริเวณออกฤทธิ์ของเอนไซม์ไซโตโครมซีออกซิเดสและปิดกั้น การหายใจ ระดับเซลล์ [ 85 ]
ปัจจัยที่มีผลต่อการทำงานของเอนไซม์
เนื่องจากเอนไซม์ประกอบด้วยโปรตีน การทำงานของเอนไซม์จึงไวต่อการเปลี่ยนแปลงของปัจจัยทางกายภาพและเคมีหลายอย่าง เช่น ค่า pH อุณหภูมิ ความเข้มข้นของสารตั้งต้น เป็นต้น
ตารางต่อไปนี้แสดงค่า pH ที่เหมาะสมที่สุดสำหรับเอนไซม์ต่างๆ[ 86 ]
| เอนไซม์ | ค่า pH ที่เหมาะสม | คำอธิบายค่า pH |
|---|---|---|
| เปปซิน | 1.5–1.6 | มีความเป็นกรดสูงมาก |
| อินเวอร์เทส | 4.5 | กรด |
| ไลเปส (กระเพาะอาหาร) | 4.0–5.0 | กรด |
| ไลเปส (น้ำมันละหุ่ง) | 4.7 | กรด |
| ไลเปส (ตับอ่อน) | 8.0 | ด่าง |
| อะไมเลส (มอลต์) | 4.6–5.2 | กรด |
| อะไมเลส (ตับอ่อน) | 6.7–7.0 | ความเป็นกรด-ความเป็นกลาง |
| เซลโลไบเอส | 5.0 | กรด |
| มอลเทส | 6.1–6.8 | กรด |
| ซูเครส | 6.2 | กรด |
| เอนไซม์คาตาเลส | 7.0 | เป็นกลาง |
| ยูรีเอส | 7.0 | เป็นกลาง |
| โคลีนเอสเทอเรส | 7.0 | เป็นกลาง |
| ไรโบเอนไซม์ | 7.0–7.5 | เป็นกลาง |
| ฟูมาราเซ่ | 7.8 | ด่าง |
| ทริปซิน | 7.8–8.7 | ด่าง |
| อะดีโนซีนไตรฟอสเฟต | 9.0 | ด่าง |
| อาร์จิเนส | 10.0 | มีฤทธิ์เป็นด่างสูง |
หน้าที่ทางชีวภาพ
เอนไซม์ทำ หน้าที่หลากหลายภายในสิ่งมีชีวิต เอนไซม์มีความสำคัญอย่างยิ่งต่อการส่งสัญญาณและการควบคุมเซลล์ โดยมักจะผ่านทางไคเนสและฟอสฟาเทส [ 87 ] นอกจากนี้ยังสร้างการเคลื่อนไหว โดยไมโอซินจะไฮโดรไลซ์อะดีโนซีนไตรฟอสเฟต (ATP) เพื่อสร้างการหดตัวของกล้ามเนื้อและยังขนส่งสิ่งของไปรอบๆ เซลล์ในฐานะส่วนหนึ่งของโครงร่างเซลล์ [ 88 ] ATPaseอื่นๆในเยื่อหุ้มเซลล์เป็นปั๊มไอออนที่เกี่ยวข้องกับการขนส่งแบบแอ ค ทีฟ เอนไซม์ยังมีส่วนเกี่ยวข้องกับหน้าที่ที่แปลกใหม่กว่า เช่นลูซิเฟอเรสที่สร้างแสงในหิ่งห้อย[ 89 ]ไวรัสยังสามารถมีเอนไซม์สำหรับการติดเชื้อเซลล์ เช่น อินทิเกรส และรีเวิร์สทรานสคริปเทสของ HIVหรือสำหรับการปล่อยไวรัสออกจากเซลล์ เช่นนิวรามินิเดสของไวรัสไข้หวัดใหญ่[ 90 ]
หน้าที่สำคัญของเอนไซม์คือในระบบย่อยอาหารของสัตว์ เอนไซม์ เช่นอะไมเลสและโปรตีเอสจะย่อยโมเลกุลขนาดใหญ่ ( แป้งหรือโปรตีนตามลำดับ) ให้เป็นโมเลกุลขนาดเล็ก เพื่อให้สามารถดูดซึมได้โดยลำไส้ ตัวอย่างเช่น โมเลกุลของแป้งมีขนาดใหญ่เกินกว่าจะดูดซึมได้จากลำไส้ แต่เอนไซม์จะไฮโดรไลซ์โซ่ของแป้งให้เป็นโมเลกุลขนาดเล็ก เช่นมอลโทสและในที่สุดก็เป็นกลูโคสซึ่งสามารถดูดซึมได้ เอนไซม์แต่ละชนิดย่อยสารอาหารที่แตกต่างกัน ในสัตว์เคี้ยวเอื้องซึ่งกินพืชเป็นอาหาร จุลินทรีย์ในลำไส้จะผลิตเอนไซม์อีกชนิดหนึ่งคือเซลลูเลสเพื่อย่อยผนังเซลล์เซลลูโลสของเส้นใยพืช[ 91 ]
การเผาผลาญ

เอนไซม์หลายชนิดสามารถทำงานร่วมกันตามลำดับที่เฉพาะเจาะจง ทำให้เกิดวิถีเมตาบอลิซึม [ 1 ] : 30.1 ในวิถีเมตาบอลิซึม เอนไซม์หนึ่งตัวจะใช้ผลิตภัณฑ์ของเอนไซม์อีกตัวเป็นสารตั้งต้น หลังจากปฏิกิริยาเร่งปฏิกิริยา ผลิตภัณฑ์นั้นจะถูกส่งต่อไปยังเอนไซม์อีกตัว บางครั้งเอนไซม์มากกว่าหนึ่งตัวสามารถเร่งปฏิกิริยาเดียวกันได้พร้อมกัน ซึ่งอาจทำให้เกิดการควบคุมที่ซับซ้อนมากขึ้น เช่น เอนไซม์ตัวหนึ่งมีกิจกรรมคงที่ต่ำ แต่เอนไซม์ตัวที่สองมีกิจกรรมสูงที่สามารถเหนี่ยวนำได้[ 92 ]
เอนไซม์เป็นตัวกำหนดขั้นตอนที่เกิดขึ้นในเส้นทางเหล่านี้ หากไม่มีเอนไซม์ การเผาผลาญจะไม่สามารถดำเนินไปตามขั้นตอนเดียวกันและไม่สามารถควบคุมให้ตอบสนองความต้องการของเซลล์ได้ เส้นทางการเผาผลาญหลักส่วนใหญ่จะถูกควบคุมในไม่กี่ขั้นตอน โดยทั่วไปผ่านเอนไซม์ที่มีกิจกรรมเกี่ยวข้องกับการฟอสโฟรีเลชันโดย ATP เนื่องจากปฏิกิริยานี้ปลดปล่อยพลังงานจำนวนมาก ปฏิกิริยาอื่นๆ ที่ไม่เอื้อต่ออุณหพลศาสตร์จึงสามารถเชื่อมโยงกับการไฮโดรไลซิสของ ATP ทำให้เกิดปฏิกิริยาการเผาผลาญที่เชื่อมโยงกันโดยรวม[ 1 ] : 30.1
การควบคุมกิจกรรม
มีห้าวิธีหลักที่ควบคุมการทำงานของเอนไซม์ในเซลล์[ 1 ] : 30.1.1
ระเบียบข้อบังคับ
เอนไซม์สามารถถูกกระตุ้นหรือยับยั้งโดยโมเลกุลอื่นๆ ได้ ตัวอย่างเช่น ผลิตภัณฑ์สุดท้ายของวิถีเมตาบอลิซึมมักจะเป็นตัวยับยั้งเอนไซม์ตัวแรกๆ ของวิถีนั้น (โดยปกติจะเป็นขั้นตอนที่ไม่สามารถย้อนกลับได้ขั้นตอนแรก เรียกว่าขั้นตอนที่ผูกพัน) ดังนั้นจึงควบคุมปริมาณผลิตภัณฑ์สุดท้ายที่สร้างขึ้นโดยวิถีนั้น กลไกการควบคุมดังกล่าวเรียกว่ากลไกป้อนกลับเชิงลบเนื่องจากปริมาณผลิตภัณฑ์สุดท้ายที่ผลิตได้ถูกควบคุมโดยความเข้มข้นของตัวมันเอง[ 93 ] : 141–48 กลไกป้อนกลับเชิงลบสามารถปรับอัตราการสังเคราะห์ของเมตาบอไลต์ระดับกลางได้อย่างมีประสิทธิภาพตามความต้องการของเซลล์ ซึ่งช่วยในการจัดสรรวัสดุและพลังงานอย่างมีประสิทธิภาพ และป้องกันการผลิตผลิตภัณฑ์สุดท้ายมากเกินไป เช่นเดียวกับอุปกรณ์รักษาสมดุล อื่นๆ การควบคุมการทำงานของเอนไซม์ช่วยรักษาสภาพแวดล้อมภายในที่คงที่ในสิ่งมีชีวิต[ 93 ] : 141
การดัดแปลงหลังการแปล
ตัวอย่างของการดัดแปลงหลังการแปลได้แก่ ฟอ สโฟรีเลชันไมริสโตอิเลชันและไกลโคซิเลชัน [ 93 ] : 149–69 ตัวอย่างเช่น ในการตอบสนองต่ออินซูลินฟอสโฟรีเลชันของเอนไซม์หลายตัว รวมถึงไกลโคเจน ซินเทส ช่วยควบคุมการสังเคราะห์หรือการสลายไกลโคเจนและช่วยให้เซลล์ตอบสนองต่อการเปลี่ยนแปลงของน้ำตาลในเลือด [ 94 ] อีกตัวอย่างหนึ่งของการดัดแปลงหลังการแปลคือการตัดสายโซ่โพลีเปปไทด์ ไคโมทริ ปซิน ซึ่งเป็นโปรตีเอสย่อยอาหาร ถูกผลิตในรูปที่ไม่ทำงานเป็นไคโมทริปซินโนเจนในตับอ่อนและถูกขนส่งในรูปทรงนี้ไปยังกระเพาะอาหารซึ่งจะถูกกระตุ้นให้ทำงาน ซึ่งจะหยุดเอนไซม์จากการย่อยตับอ่อนหรือเนื้อเยื่ออื่นๆ ก่อนที่จะเข้าสู่ลำไส้ สารตั้งต้นที่ไม่ทำงานของเอนไซม์ชนิดนี้เรียกว่าไซโมเจน[ 93 ] : 149–53 หรือโปรเอนไซม์
ปริมาณ
การผลิตเอนไซม์ ( การถอดรหัสและการแปลยีนเอนไซม์) สามารถเพิ่มขึ้นหรือลดลงได้โดยเซลล์เพื่อตอบสนองต่อการเปลี่ยนแปลงในสภาพแวดล้อมของเซลล์ รูปแบบการควบคุมยีน นี้ เรียกว่าการเหนี่ยวนำเอนไซม์ตัวอย่างเช่น แบคทีเรียอาจดื้อต่อยาปฏิชีวนะเช่นเพนิซิลลินเนื่องจากเอนไซม์ที่เรียกว่าเบต้า-แลคตาเมสถูกเหนี่ยวนำให้ไฮโดรไลซ์วงแหวนเบต้า-แลคแทม ที่สำคัญ ภายในโมเลกุลของเพนิซิลลิน[ 95 ]อีกตัวอย่างหนึ่งมาจากเอนไซม์ในตับที่เรียกว่าไซโตโครม P450 ออกซิเดสซึ่งมีความสำคัญในการเผาผลาญยาการเหนี่ยวนำหรือการยับยั้งเอนไซม์เหล่านี้สามารถทำให้เกิดปฏิกิริยาระหว่างยาได้ [ 96 ] ระดับเอนไซม์ยังสามารถควบคุมได้โดยการเปลี่ยนอัตราการย่อยสลาย ของ เอนไซม์[ 1 ] : 30.1.1 สิ่งที่ตรงข้ามกับการเหนี่ยว นำ เอนไซม์คือการยับยั้งเอนไซม์
การกระจายตัวในระดับเซลล์ย่อย
เอนไซม์สามารถแบ่งส่วนได้ โดยมีวิถีการเผาผลาญที่แตกต่างกันเกิดขึ้นในส่วนต่างๆ ของเซลล์ตัวอย่างเช่นกรดไขมันถูกสังเคราะห์โดยเอนไซม์ชุดหนึ่งในไซโตซอล เอน โดพลาสมิกเรติคูลัมและกอลจิและถูกใช้โดยเอนไซม์อีกชุดหนึ่งเป็นแหล่งพลังงานในไมโทคอนเดรียผ่านกระบวนการβ-ออกซิเดชัน [ 97 ] นอกจากนี้การเคลื่อนย้ายของเอนไซม์ไปยังส่วนต่างๆ อาจเปลี่ยนแปลงระดับการโปรตอน (เช่น ไซโตพลาส ม์ที่เป็นกลาง และไลโซโซมที่เป็นกรด) หรือสถานะออกซิเดชัน (เช่นเพ ริพลาสม์ที่เป็นออกซิไดซ์หรือไซ โตพลาสม์ที่เป็นรีดิวซ์) ซึ่งส่งผลต่อกิจกรรมของเอนไซม์[ 98 ]ในทางตรงกันข้ามกับการแบ่งส่วนเข้าไปในออร์แกเนลล์ที่มีเยื่อหุ้ม การกำหนดตำแหน่งย่อยของเซลล์ของเอนไซม์อาจเปลี่ยนแปลงได้ผ่านการสร้างพอลิเมอร์ของเอนไซม์เป็นเส้นใยไซโตพลาสมิกขนาดใหญ่[ 99 ] [ 100 ]
ความเชี่ยวชาญเฉพาะด้านขององค์กร
ในยูคาริโอตหลายเซลล์ เซลล์ในอวัยวะและเนื้อเยื่อ ต่างๆ มีรูปแบบการแสดงออกของยีน ที่แตกต่างกัน ดังนั้นจึงมีชุดเอนไซม์ที่แตกต่างกัน (เรียกว่าไอโซเอนไซม์ ) ที่พร้อมสำหรับปฏิกิริยาเมตาบอลิซึม ซึ่งเป็นกลไกในการควบคุมเมตาบอลิซึมโดยรวมของสิ่งมีชีวิต ตัวอย่างเช่นเฮกโซไคเนสซึ่งเป็นเอนไซม์ตัวแรกใน วิถีไกล โคไลซิสมีรูปแบบเฉพาะที่เรียกว่ากลูโคไคเนสที่แสดงออกในตับและตับอ่อนซึ่งมีความสัมพันธ์กับกลูโคสน้อยกว่า แต่มีความไวต่อความเข้มข้นของกลูโคสมากกว่า[ 101 ]เอนไซม์นี้มีส่วนเกี่ยวข้องในการตรวจจับน้ำตาลในเลือดและควบคุมการผลิตอินซูลิน[ 102 ]
การมีส่วนร่วมในโรค


เนื่องจากการควบคุมกิจกรรมของเอนไซม์อย่างเข้มงวดเป็นสิ่งจำเป็นสำหรับภาวะสมดุลการทำงานผิดปกติใดๆ (การกลายพันธุ์ การผลิตมากเกินไป การผลิตน้อยเกินไป หรือการขาดหายไป) ของเอนไซม์ที่สำคัญเพียงตัวเดียวอาจนำไปสู่โรคทางพันธุกรรมได้ การทำงานผิดปกติของเอนไซม์เพียงชนิดเดียวจากหลายพันชนิดที่มีอยู่ในร่างกายมนุษย์อาจถึงแก่ชีวิตได้ ตัวอย่างของโรคทางพันธุกรรมที่ร้ายแรงเนื่องจากเอนไซม์ไม่เพียงพอคือโรคเทย์-แซคส์ซึ่งผู้ป่วยขาดเอนไซม์เฮกโซซามิเนส[ 103 ] [ 104 ]
ตัวอย่างหนึ่งของการขาดเอนไซม์คือโรค ฟีนิลคีโตนูเรียชนิดที่พบได้บ่อยที่สุดการกลายพันธุ์ของกรดอะมิโนเดี่ยวหลายชนิดในเอนไซม์ฟีนิลอะลานีนไฮดรอก ซิเล ส ซึ่งเป็นตัวเร่งปฏิกิริยาในขั้นตอนแรกของการย่อยสลายฟีนิลอะลานีนส่งผลให้เกิดการสะสมของฟีนิลอะลานีนและผลิตภัณฑ์ที่เกี่ยวข้อง การกลายพันธุ์บางส่วนเกิดขึ้นในบริเวณออกฤทธิ์ ซึ่งขัดขวางการจับและการเร่งปฏิกิริยาโดยตรง แต่หลายการกลายพันธุ์อยู่ห่างจากบริเวณออกฤทธิ์และลดกิจกรรมโดยการทำให้โครงสร้างโปรตีนไม่เสถียร หรือส่งผลต่อการเกิดโอลิโกเมอไรเซชันที่ถูกต้อง[ 105 ] [ 106 ]ซึ่งอาจนำไปสู่ความบกพร่องทางสติปัญญาหากไม่ได้รับการรักษา[ 107 ]อีกตัวอย่างหนึ่งคือภาวะขาดเอนไซม์ซูโดโคลีนเอสเทอเรสซึ่งความสามารถของร่างกายในการย่อยสลายยาโคลีนเอสเทอร์บกพร่อง[ 108 ] การให้เอนไซม์ทางปากสามารถใช้รักษาภาวะขาดเอนไซม์บางชนิดได้ เช่นภาวะตับอ่อนทำงานบกพร่อง[ 109 ]และภาวะไม่ทนต่อแลคโตส[ 110 ]
อีกวิธีหนึ่งที่ความผิดปกติของเอนไซม์สามารถก่อให้เกิดโรคได้มาจากการกลายพันธุ์ในเซลล์สืบพันธุ์ของยีนที่เข้ารหัสเอนไซม์ซ่อมแซม DNA ความบกพร่องในเอนไซม์เหล่านี้ทำให้เกิดมะเร็งเนื่องจากเซลล์มีความสามารถในการซ่อมแซมการกลายพันธุ์ใน จีโนม น้อยลง ทำให้เกิดการสะสมของการกลายพันธุ์อย่างช้าๆ และส่งผลให้เกิดมะเร็งตัวอย่างของกลุ่มอาการมะเร็ง ทางพันธุกรรมดังกล่าว คือxeroderma pigmentosumซึ่งทำให้เกิดมะเร็งผิวหนังแม้เพียงการสัมผัสกับรังสีอัลตราไวโอเลตเพียง เล็กน้อย [ 111 ] [ 112 ]
วิวัฒนาการ
เช่นเดียวกับโปรตีนอื่นๆ เอนไซม์ก็เปลี่ยนแปลงไปตามกาลเวลาผ่านการกลายพันธุ์และการเปลี่ยนแปลงลำดับกรดอะมิโน เนื่องจากเอนไซม์มีบทบาทสำคัญในกระบวนการ เผาผลาญ การวิวัฒนาการของเอนไซม์จึงมีบทบาทสำคัญในการปรับตัวดังนั้นคำถามสำคัญคือ เอนไซม์สามารถเปลี่ยนแปลงกิจกรรมทางเอนไซม์ได้หรือไม่และอย่างไร โดยทั่วไปเป็นที่ยอมรับกันว่ากิจกรรมของเอนไซม์ใหม่ๆ จำนวนมากได้วิวัฒนาการมาจากการเพิ่มจำนวนยีนและการกลายพันธุ์ของสำเนาที่เพิ่มจำนวนขึ้นมา แม้ว่าวิวัฒนาการอาจเกิดขึ้นได้โดยไม่ต้องมีการเพิ่มจำนวนยีนก็ตาม ตัวอย่างหนึ่งของเอนไซม์ที่เปลี่ยนแปลงกิจกรรมคือบรรพบุรุษของเมไธโอนิลอะมิโนเปปติเดส (MAP) และครีเอทีนอะมิดิโนไฮโดรเลส ( ครีเอทีเน ส ) ซึ่งมีความคล้ายคลึงกันอย่างชัดเจน แต่เร่งปฏิกิริยาที่แตกต่างกันมาก (MAP กำจัดเมไธโอนีนที่ปลายอะมิโนในโปรตีนใหม่ ในขณะที่ครีเอทีเนสไฮโดรไลซ์ครีเอทีนเป็นซาร์โคซีนและยูเรีย ) นอกจากนี้ MAP ยังขึ้นอยู่กับไอออนโลหะ ในขณะที่ครีเอทีเนสไม่ขึ้นอยู่กับไอออนโลหะ ดังนั้นคุณสมบัตินี้จึงหายไปตามกาลเวลาด้วย[ 113 ]การเปลี่ยนแปลงเล็กน้อยของกิจกรรมเอนไซม์เป็นเรื่องปกติอย่างยิ่งในหมู่เอนไซม์ โดยเฉพาะอย่างยิ่ง ความจำเพาะในการจับกับสารตั้งต้น (ดูข้างต้น) สามารถเปลี่ยนแปลงได้อย่างง่ายดายและรวดเร็วด้วยการเปลี่ยนแปลงกรดอะมิโนเพียงตัวเดียวในช่องจับสารตั้งต้น ซึ่งมักพบเห็นได้ในกลุ่มเอนไซม์หลัก เช่นไคเนส[ 114 ]
ปัจจุบัน การวิวัฒนาการเทียม (ในหลอดทดลอง) ถูกนำมาใช้กันอย่างแพร่หลายเพื่อปรับเปลี่ยนกิจกรรมหรือความจำเพาะของเอนไซม์สำหรับการใช้งานในอุตสาหกรรม (ดูด้านล่าง)
การใช้งานในอุตสาหกรรม
เอนไซม์ถูกนำมาใช้ในอุตสาหกรรมเคมีและการใช้งานทางอุตสาหกรรมอื่นๆ เมื่อต้องการตัวเร่งปฏิกิริยาที่มีความจำเพาะสูงมาก โดยทั่วไปแล้ว เอนไซม์มีข้อจำกัดในจำนวนปฏิกิริยาที่พวกมันพัฒนาขึ้นมาเพื่อเร่งปฏิกิริยา และยังมีข้อจำกัดในเรื่องความไม่เสถียรในตัวทำละลายอินทรีย์และที่อุณหภูมิสูง ด้วยเหตุนี้วิศวกรรมโปรตีนจึงเป็นสาขาการวิจัยที่กำลังดำเนินอยู่ และเกี่ยวข้องกับความพยายามในการสร้างเอนไซม์ใหม่ที่มีคุณสมบัติแปลกใหม่ ไม่ว่าจะผ่านการออกแบบอย่างมีเหตุผลหรือวิวัฒนาการในหลอดทดลอง[ 115 ] [ 116 ]ความพยายามเหล่านี้เริ่มประสบความสำเร็จ และขณะนี้มีเอนไซม์บางชนิดที่ได้รับการออกแบบ "ตั้งแต่เริ่มต้น" เพื่อเร่งปฏิกิริยาที่ไม่เกิดขึ้นในธรรมชาติ[ 117 ]
ดูเพิ่มเติม
ฐานข้อมูลเอนไซม์
อ่านเพิ่มเติม
|
|
ลิงก์ภายนอก
 สื่อที่เกี่ยวข้องกับเอนไซม์ในวิกิมีเดียคอมมอนส์
สื่อที่เกี่ยวข้องกับเอนไซม์ในวิกิมีเดียคอมมอนส์
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เอนไซม์
เอนไซม์เป็น โมเลกุลชีวภาพขนาดใหญ่ ซึ่ง โดยทั่วไปเป็น โปรตีน ทำหน้าที่เป็น ตัวเร่งปฏิกิริยา ทางชีวภาพ เร่ง ปฏิกิริยาเคมี โดยไม่ถูกใช้ไปในกระบวนการ โมเลกุล...
ที่มาและประวัติความเป็นมา
ในช่วงปลายศตวรรษที่ 17 และต้นศตวรรษที่ 18 การย่อย เนื้อสัตว์ โดยการหลั่งของกระเพาะอาหาร [ 8 ] และการเปลี่ยน แป้ง เป็น น้ำตาล โดยสารสกัดจากพืชและ น้ำลาย เป็นที่ทราบกันดีอยู่แล้ว แต่กลไกที่ทำให้เกิดสิ่งเหล่านี้ยังไม่ได้รับการระบุ [ 9 ]
การจำแนกและการตั้งชื่อ
เอนไซม์สามารถจำแนกได้ตามเกณฑ์หลักสองประการ ได้แก่ กิจกรรมของเอนไซม์ หรือ ความคล้ายคลึงกัน ของลำดับกรดอะมิโน (และด้วยเหตุนี้จึงแสดงถึงความสัมพันธ์ทางวิวัฒนาการ)
กิจกรรมของเอนไซม์
ชื่อของเอนไซม์มักได้มาจากสารตั้งต้นหรือปฏิกิริยาเคมีที่เอนไซม์เร่งปฏิกิริยา โดยคำจะลงท้ายด้วย-ase [ 1 ] : 8.1.3 ตัวอย่างเช่น แลคเตส แอลกอฮอล์ ดีไฮโดรจีเนส และ ดีเอ็นเอพอลิเมอเรส เอนไซม์ที่แตกต่างกันแต่เร่งปฏิกิริยาเคมีเดียวกันเรียกว่าไอ โซเอนไซม์ [ 1 ] : 10.3