อ่าน 20 นาที
การตายของเซลล์ตามโปรแกรม
การตายของเซลล์ตามโปรแกรม ( PCD ) บางครั้งเรียกว่าการฆ่าตัวตายของเซลล์หรือการฆ่าตัวตายของเซลล์ คือการตายของเซลล์อันเป็นผลมาจากเหตุการณ์ภายในเซลล์ เช่นอะพอพโทซิสหรือออโตฟาจี
การตายของเซลล์ตามโปรแกรม
การตายของเซลล์ตามโปรแกรม ( PCD ) บางครั้งเรียกว่าการฆ่าตัวตายของเซลล์หรือการฆ่าตัวตายของเซลล์[ 1 ] [ 2 ] [ 3 ]คือการตายของเซลล์อันเป็นผลมาจากเหตุการณ์ภายในเซลล์ เช่นอะพอพโทซิสหรือออโตฟาจี [ 4 ] [ 5 ] PCDเกิดขึ้นในกระบวนการทางชีวภาพซึ่งโดยปกติแล้วจะให้ประโยชน์ในระหว่างวงจรชีวิต ของสิ่งมีชีวิต ตัวอย่างเช่นการแยกนิ้วมือและนิ้วเท้าในตัวอ่อนมนุษย์ที่กำลังพัฒนาเกิดขึ้นเนื่องจากเซลล์ระหว่างนิ้วมือเกิดอะพอพโทซิส ผลที่ได้คือนิ้วมือแยกออกจากกัน PCD ทำหน้าที่พื้นฐานในระหว่างการพัฒนาเนื้อเยื่อ ของพืชและสัตว์
อะพอพโทซิสและออโตฟาจีต่างก็เป็นรูปแบบของการตายของเซลล์แบบมีโปรแกรม[ 6 ]เนโครซิสคือการตายของเซลล์ที่เกิดจากปัจจัยภายนอก เช่น การบาดเจ็บหรือการติดเชื้อ และเกิดขึ้นได้หลายรูปแบบ เนโครซิสถูกมองว่าเป็นกระบวนการที่ไม่เป็นไปตามสรีรวิทยาซึ่งเกิดขึ้นจากการติดเชื้อหรือการบาดเจ็บมานานแล้ว[ 6 ]แต่ในช่วงปี 2000 รูปแบบของเนโครซิสแบบมีโปรแกรมที่เรียกว่าเนโครพโทซิส [ 7 ] ได้รับการยอมรับว่าเป็นอีกรูปแบบหนึ่งของการตายของเซลล์แบบมีโปรแกรม มีการตั้งสมมติฐานว่าเนโครพโทซิสสามารถทำหน้าที่เป็นกลไกสำรองในการตายของเซลล์แทนอะพอพโทซิสเมื่อการส่งสัญญาณอะพอพโทซิสถูกปิดกั้นโดยปัจจัยภายในหรือภายนอก เช่น ไวรัสหรือการกลายพันธุ์ เมื่อไม่นานมานี้ ยังมีการค้นพบเนโครซิสแบบมีการควบคุมประเภทอื่นๆ อีกด้วย ซึ่งมีเหตุการณ์การส่งสัญญาณหลายอย่างร่วมกับเนโครพโทซิสและอะพอพโทซิส[ 8 ]
ประวัติศาสตร์
แนวคิดเรื่อง "การตายของเซลล์ตามโปรแกรม" ถูกนำมาใช้โดยLockshin & Williams [ 9 ]ในปี พ.ศ. 2507 โดยเกี่ยวข้องกับ การพัฒนาเนื้อเยื่อ ของแมลงประมาณแปดปีก่อนที่คำว่า "อะพอพโทซิส" จะถูกบัญญัติขึ้น อย่างไรก็ตาม คำว่า PCD เป็นแหล่งที่มาของความสับสน และ Durand และ Ramsey [ 10 ]ได้พัฒนาแนวคิดนี้โดยให้คำจำกัดความเชิงกลไกและวิวัฒนาการ PCD ได้กลายเป็นคำทั่วไปที่อ้างถึงการตายของเซลล์ทุกประเภทที่มีองค์ประกอบทางพันธุกรรม[ 11 ]
ความเข้าใจเบื้องต้นเกี่ยวกับกลไกนี้มาจากการศึกษาBCL2ซึ่งเป็นผลิตภัณฑ์ของยีนก่อมะเร็งที่ถูกกระตุ้นโดยการย้ายตำแหน่ง ของ โครโมโซม ที่มักพบในมะเร็งต่อมน้ำ เหลืองชนิดฟอลลิคูลาร์ แตกต่างจากยีนมะเร็งอื่นๆ ที่ส่งเสริมมะเร็งโดยการกระตุ้นการแพร่กระจายของเซลล์ BCL2 ส่งเสริมมะเร็งโดยการหยุดยั้งเซลล์มะเร็งต่อมน้ำเหลืองไม่ให้สามารถฆ่าตัวเองได้[ 12 ]
PCD ได้รับความสนใจและการวิจัยเพิ่มมากขึ้น แนวโน้มนี้ได้รับการเน้นย้ำด้วยการมอบรางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ประจำ ปี 2002 ให้แก่ซิดนีย์ เบรนเนอร์ ( สหราชอาณาจักร ) เอช. โรเบิร์ต ฮอร์วิตซ์ (สหรัฐอเมริกา) และจอห์น อี. ซัลสตัน (สหราชอาณาจักร) [ 13 ]
ประเภท
- อะพอพโทซิสหรือ การตายของเซลล์ประเภทที่ 1
- การตายของเซลล์แบบออโตฟาจิกหรือการตายของเซลล์ประเภท II ( ไซโตพลาสมิก : มีลักษณะเฉพาะคือการก่อตัวของแวคิวโอล ขนาดใหญ่ ที่กัดกินออร์แกเนลล์ตามลำดับที่เฉพาะเจาะจงก่อนที่จะทำลายนิวเคลียส ) [ 14 ]
อะพอพโทซิส
อะพอพโทซิสเป็นกระบวนการตายของเซลล์ตามโปรแกรม (PCD) ที่อาจเกิดขึ้นในสิ่งมีชีวิตหลายเซลล์ [ 15 ] เหตุการณ์ทางชีวเคมีนำไปสู่การเปลี่ยนแปลงของเซลล์ ( สัณฐานวิทยา ) และการตายของเซลล์ การเปลี่ยนแปลงเหล่านี้รวมถึงการเกิดฟองการหดตัวของเซลล์การแตกตัวของนิวเคลียสการควบแน่นของโครมาติน และ การแตกตัวของ ดีเอ็นเอในโครโมโซม ปัจจุบันเชื่อกันว่าในบริบทของการพัฒนา เซลล์จะถูกกระตุ้นให้ฆ่าตัวตายในขณะที่อยู่ในสภาวะสมดุล การขาดปัจจัยการอยู่รอดบางอย่างอาจเป็นแรงผลักดันให้เกิดการฆ่าตัวตาย ดูเหมือนว่าจะมีความแปรผันในสัณฐานวิทยาและชีวเคมีของเส้นทางการฆ่าตัวตายเหล่านี้ บางเส้นทางเดินตาม "อะพอพโทซิส" บางเส้นทางเดินตามเส้นทางการกำจัดที่ทั่วไปกว่า แต่ทั้งสองเส้นทางมักมีแรงจูงใจทางพันธุกรรมและสังเคราะห์ มีหลักฐานบางอย่างที่บ่งชี้ว่าอาการบางอย่างของ "อะพอพโทซิส" เช่น การกระตุ้นเอนโดนิวคลีเอส สามารถเกิดขึ้นได้โดยไม่จำเป็นต้องเกี่ยวข้องกับกระบวนการทางพันธุกรรม อย่างไรก็ตาม คาดว่าอะพอพโทซิสที่แท้จริงและการตายของเซลล์ตามโปรแกรมจะต้องเกิดขึ้นโดยอาศัยการควบคุมทางพันธุกรรม นอกจากนี้ยังเริ่มชัดเจนขึ้นว่าไมโทซิสและอะพอพโทซิสมีการสลับหรือเชื่อมโยงกันในบางลักษณะ และความสมดุลที่เกิดขึ้นนั้นขึ้นอยู่กับสัญญาณที่ได้รับจากปัจจัยการเจริญเติบโตหรือการอยู่รอดที่เหมาะสม[ 16 ]
เส้นทางภายนอกเทียบกับเส้นทางภายใน
มีเส้นทางที่เป็นไปได้สองเส้นทางที่แตกต่างกันที่อาจเกิดขึ้นเมื่อจำเป็นต้องมีอะพอพโทซิส ได้แก่ เส้นทางภายนอกและเส้นทางภายใน ทั้งสองเส้นทางเกี่ยวข้องกับการใช้แคสเปส ซึ่งมีความสำคัญต่อการตายของเซลล์[ 17 ]
วิถีภายนอก
เส้นทางภายนอกเกี่ยวข้องกับการโต้ตอบระหว่างตัวรับและลิแกนด์ที่เฉพาะเจาะจง ลิแกนด์ FAS จะจับกับตัวรับ FAS หรือลิแกนด์ TNF-alpha สามารถจับกับตัวรับ TNF ได้ ในทั้งสองกรณีนี้จะมีการกระตุ้นแคสเปสเริ่มต้น เส้นทางภายนอกสามารถถูกกระตุ้นได้สองวิธี วิธีแรกคือผ่านการจับลิแกนด์ TNF-alpha อย่างรวดเร็วหรือผ่านเซลล์ T ที่เป็นพิษต่อเซลล์ เซลล์ T ที่เป็นพิษต่อเซลล์สามารถเกาะติดกับเยื่อหุ้มเซลล์ ทำให้มีการปล่อยแกรนไซม์ B ออกมา แกรนไซม์ B จะเจาะเยื่อหุ้มเซลล์เป้าหมายและทำให้มีการปล่อยเพอร์ฟอรินออกมา ในที่สุดเพอร์ฟอรินจะสร้างรูพรุนในเยื่อหุ้มเซลล์และปล่อยแคสเปสออกมา ซึ่งนำไปสู่การกระตุ้นแคสเปส 3 แคสเปสเริ่มต้นนี้อาจทำให้เกิดการแตกตัวของแคสเปส 3 ที่ไม่ทำงาน ทำให้กลายเป็นแคสเปส 3 ที่แตกตัวแล้ว นี่คือโมเลกุลสุดท้ายที่จำเป็นในการกระตุ้นการตายของเซลล์[ 18 ]
วิถีภายใน
กลไกการตายของเซลล์แบบอะพอพโทซิสเกิดขึ้นจากความเสียหายของเซลล์ เช่น ความเสียหายของดีเอ็นเอ หรือการสัมผัสกับรังสียูวี กลไกนี้เกิดขึ้นในไมโทคอนเดรียและถูกควบคุมโดยเซนเซอร์ที่เรียกว่าเซนเซอร์ Bcl และโปรตีนสองชนิดที่เรียกว่า BAX และ BAK โปรตีนเหล่านี้พบได้ในสัตว์เลี้ยงลูกด้วยนมชั้นสูงส่วนใหญ่ เนื่องจากพวกมันสามารถเจาะเยื่อหุ้มไมโทคอนเดรียชั้นนอกได้ ทำให้พวกมันเป็นส่วนสำคัญในการควบคุมการตายของเซลล์โดยอะพอพโทซิส พวกมันทำเช่นนั้นโดยการควบคุมการสร้างรูพรุนภายในเยื่อหุ้ม ซึ่งจำเป็นต่อการปล่อยไซโตโครมซี อย่างไรก็ตาม ไซโตโครมซีจะถูกปล่อยออกมาก็ต่อเมื่อเยื่อหุ้มไมโทคอนเดรียเสียหายเท่านั้น เมื่อตรวจพบไซโตโครมซีแล้ว คอมเพล็กซ์อะพอพโทโซมจะถูกสร้างขึ้น คอมเพล็กซ์นี้จะกระตุ้นเอนไซม์แคสเปสซึ่งเป็นเอนไซม์ที่ทำให้เซลล์ตาย การฆ่าเซลล์นี้อาจมีความสำคัญเนื่องจากป้องกันการเจริญเติบโตของเซลล์มากเกินไปซึ่งอาจนำไปสู่โรคต่างๆ เช่น มะเร็ง นอกจากนี้ยังมีโปรตีนอีกสองชนิดที่ควรกล่าวถึงซึ่งยับยั้งการปล่อยไซโตโครมซีในไมโทคอนเดรีย Bcl-2 และ Bcl-xl เป็นสารต้านอะพอพโทซิส ดังนั้นจึงป้องกันการตายของเซลล์ มีการกลายพันธุ์ที่อาจเกิดขึ้นได้ซึ่งทำให้เกิดการทำงานมากเกินไปของ Bcl-2 ซึ่งก็คือการย้ายตำแหน่งระหว่างโครโมโซม 14 และ 18 การทำงานที่มากเกินไปนี้อาจส่งผลให้เกิดมะเร็งต่อมน้ำเหลืองชนิดฟอลลิคูลาร์ได้[ 19 ]
ออโตฟาจี
มาโครออโตฟาจีซึ่งมักเรียกว่าออโตฟาจีเป็น กระบวนการ สลายที่ส่งผลให้เกิด การย่อยสลายของสาร ในไซโตพลา สซึม จำนวนมากโปรตีนที่ผิดปกติ และออร์แกเนลล์ ส่วนเกินหรือที่เสียหายโดยออโตฟาโก โซมและ ไลโซ โซม[ 6 ]
โดยทั่วไปแล้ว กระบวนการ ออโตฟาจีจะถูกกระตุ้นเมื่อ ขาด สารอาหารแต่ก็ยังเกี่ยวข้องกับ กระบวนการ ทางสรีรวิทยาและพยาธิวิทยาเช่น การเจริญเติบโต การเปลี่ยนแปลงรูปร่าง โรค ความ เสื่อมของระบบประสาท ความเครียดการติดเชื้อและมะเร็ง
กลไก
ตัวควบคุมที่สำคัญของการเหนี่ยวนำออโตฟาจีคือไคเนสmTORซึ่งเมื่อถูกกระตุ้นจะยับยั้งออโตฟาจีและเมื่อไม่ถูกกระตุ้นจะส่งเสริม ออโตฟาจี ไคเน สเซริน / ทรี โอนีนที่เกี่ยวข้องสามตัว ได้แก่ UNC-51-like kinase -1, -2 และ -3 (ULK1, ULK2, UKL3) ซึ่งมีบทบาทคล้ายกับAtg1 ในยีสต์ ทำงานอยู่ภายใต้การควบคุมของคอมเพล็กซ์mTOR ULK1และULK2สร้างคอมเพล็กซ์ขนาดใหญ่ร่วมกับโฮโมล็อกของผลิตภัณฑ์ยีนที่เกี่ยวข้องกับออโตฟาจี (Atg) ในสัตว์เลี้ยงลูกด้วยนม (mAtg13) และโปรตีนโครงสร้าง FIP200 คอมเพล็กซ์ PI3K คลาส III ซึ่งประกอบด้วย hVps34, Beclin-1 , p150 และโปรตีนที่คล้ายกับ Atg14 หรือยีนที่เกี่ยวข้องกับความต้านทานต่อรังสีอัลตราไวโอเลต (UVRAG) จำเป็นสำหรับการเหนี่ยวนำออโตฟาจี
ยีนATG ควบคุม การสร้าง ออโตฟาโกโซมผ่านคอมเพล็กซ์ATG12 - ATG5และ LC3-II ( ATG8 - II ) ATG12จะถูกเชื่อมต่อกับATG5ใน ปฏิกิริยาคล้าย ยูบิควิตินซึ่งต้องใช้ATG7และATG10คอมเพล็กซ์ Atg12–Atg5 จะทำปฏิกิริยาแบบไม่เกิดพันธะโควาเลนต์กับ ATG16 เพื่อสร้างคอมเพล็กซ์ขนาดใหญ่ LC3/ ATG8จะถูกตัดที่ปลาย C โดยโปรตี เอส ATG4 เพื่อสร้าง LC3-I ในไซโตพลาสซึม LC3-I จะถูกเชื่อมต่อกับฟอสฟาติดิลเอทานอลามีน (PE) ในปฏิกิริยาคล้ายยูบิควิตินซึ่งต้องใช้ Atg7 และ Atg3 รูปแบบของ LC3 ที่มีไขมันเกาะอยู่ ซึ่งรู้จักกันในชื่อ LC3-II จะติดอยู่กับเยื่อหุ้มออโตฟาโกโซม[ 20 ]
ออโตฟาจีและอะพอพโทซิสมีความเชื่อมโยงกันทั้งในเชิงบวกและเชิงลบ และมีการสื่อสารข้ามกันอย่างกว้างขวางระหว่างทั้งสอง ในช่วงที่ขาดสารอาหารออโตฟาจีทำหน้าที่เป็นกลไกการอยู่รอด อย่างไรก็ตาม ออโตฟาจีที่มากเกินไปอาจนำไปสู่การตายของเซลล์ซึ่งเป็นกระบวนการ ที่มี ลักษณะ แตกต่างจากอะพอพโทซิส สัญญาณ กระตุ้น อะพอพโทซิสหลายอย่างเช่นTNF , TRAILและFADDยังกระตุ้นออโตฟาจีอีกด้วย นอกจากนี้Bcl-2ยังยับยั้ง ออโตฟาจีที่ขึ้นอยู่กับ Beclin-1ดังนั้นจึงทำหน้าที่ทั้งเป็นตัวควบคุมการอยู่รอดและตัวควบคุมต่อต้านออโตฟาจี[ 20 ]
ประเภทอื่นๆ
นอกจาก PCD สองประเภทข้างต้นแล้ว ยังมีการค้นพบเส้นทางอื่นๆ อีกด้วย[ 21 ] เส้นทางการตายทางเลือกเหล่านี้ เรียกว่า "การตายของเซลล์แบบโปรแกรมที่ไม่ใช่อะพอพโทซิส" (หรือ " การตายของเซลล์แบบโปรแกรมที่ไม่ขึ้นกับแคส เปส " หรือ "เนโครพโทซิส") ซึ่งมีประสิทธิภาพเท่ากับอะพอพโทซิส และสามารถทำหน้าที่เป็นกลไกสำรองหรือเป็นประเภทหลักของ PCD ก็ได้[ 22 ]
รูปแบบอื่น ๆ ของการตายของเซลล์ตามโปรแกรม ได้แก่อะโนอิคิสซึ่งเกือบจะเหมือนกับอะพอพโทซิส ยกเว้นการเหนี่ยวนำ คอร์นิฟิเคชันซึ่งเป็นรูปแบบการตายของเซลล์เฉพาะในชั้นหนังกำพร้า เอ็กซิโทท็อกซิซิตี้ เฟอร์โรพโทซิส ซึ่งเป็นรูปแบบการตายของเซลล์ที่ขึ้นอยู่กับธาตุเหล็ก[ 23 ]และการเสื่อมสภาพของวอลเลเรียน
เนโครพโทซิสเป็นรูปแบบหนึ่งของเนโครซิสหรือการตายของเซลล์ที่เกิดจากการอักเสบตามโปรแกรม โดยทั่วไป เนโครซิสเกี่ยวข้องกับการตายของเซลล์ที่ไม่ได้กำหนดโปรแกรมไว้ ซึ่งเกิดจากความเสียหายของเซลล์หรือการแทรกซึมของเชื้อโรค ตรงกันข้ามกับการตายของเซลล์ที่เป็นระเบียบตามโปรแกรมผ่านอะพอพโทซิสเนโมซิสเป็นอีกรูปแบบหนึ่งของเนโครซิสตามโปรแกรมที่เกิดขึ้นในไฟโบรบ ลาสต์ [ 24 ]
อีริปโทซิสเป็นรูปแบบหนึ่งของการตายของเม็ดเลือดแดง แบบฆ่าตัวตาย [ 25 ]
อะโพเนโครซิสเป็นการผสมผสานระหว่างอะพอพโทซิสและเนโครซิส และหมายถึงกระบวนการอะพอพโทซิสที่ไม่สมบูรณ์ซึ่งเสร็จสมบูรณ์ด้วยเนโครซิส[ 26 ]
NETosisคือกระบวนการตายของเซลล์ที่เกิดจากนิวโทรฟิล ส่งผลให้เกิด NETs [ 27 ]
พาราปโทซิสเป็นการตายของเซลล์อีกประเภทหนึ่งที่ไม่ใช่อะพอปโทซิส ซึ่งเกิดขึ้นโดยMAPKผ่านการกระตุ้นของIGF-1โดยมีลักษณะเฉพาะคือการก่อตัวของแวคิวโอลภายในเซลล์และการบวมของไมโตคอนเดรีย[ 28 ]
ไพรอพโทซิสซึ่งเป็นการตายของเซลล์ประเภทหนึ่งที่ทำให้เกิดการอักเสบ เกิดขึ้นโดยอาศัยแคสเปส 1ซึ่งเป็นเอนไซม์ที่ไม่เกี่ยวข้องกับอะพอพโทซิส ในการตอบสนองต่อการติดเชื้อจากจุลินทรีย์บางชนิด[ 28 ]
เซลล์พืชมีกระบวนการ PCD เฉพาะที่คล้ายกับการตายของเซลล์แบบออโตฟาจี อย่างไรก็ตาม ลักษณะทั่วไปบางประการของ PCD นั้นมีการอนุรักษ์ไว้อย่างดีทั้งในพืชและสัตว์[ 22 ]
ปัจจัยที่ทำให้เกิดการฝ่อ
ปัจจัยที่ทำให้เซลล์ฝ่อคือแรงที่ทำให้เซลล์ตาย เฉพาะแรงตามธรรมชาติที่กระทำต่อเซลล์เท่านั้นที่ถือว่าเป็นปัจจัยที่ทำให้ เซลล์ฝ่อ ในขณะที่ตัวอย่างเช่น ตัวแทนของการทำร้ายทางกลหรือทางเคมี หรือการแตกตัวของเซลล์นั้นไม่ถือว่าเป็นปัจจัยที่ทำให้เซลล์ฝ่อ ประเภทของปัจจัยที่ทำให้เซลล์ฝ่อที่พบได้ทั่วไป ได้แก่: [ 29 ]
- ภาระงานลดลง
- การสูญเสียการทำงานของเส้นประสาท
- ปริมาณเลือดที่ไหลเวียนลดลง
- โภชนาการไม่เพียงพอ
- การสูญเสียการกระตุ้นจากต่อมไร้ท่อ
- ความชรา
- การบีบอัด
บทบาทในการพัฒนาของระบบประสาท

การขยายตัวเริ่มต้นของระบบประสาท ที่กำลังพัฒนา จะถูกชดเชยด้วยการกำจัดเซลล์ประสาทและกระบวนการของเซลล์ประสาท[ 30 ]ในระหว่างการพัฒนาของระบบประสาท เซลล์ประสาทที่กำลังพัฒนาเกือบ 50% จะถูกกำจัดออกไปตามธรรมชาติด้วยการตายของเซลล์ตามโปรแกรม (PCD) [ 31 ] การตายของเซลล์ตามโปรแกรม (PCD) ในระบบประสาทได้รับการค้นพบครั้งแรกในปี พ.ศ. 2449 โดย John Beard [ 32 ]ตั้งแต่นั้นมาก็มีทฤษฎีหลายทฤษฎีที่ถูกเสนอขึ้นเพื่อทำความเข้าใจความสำคัญทางชีววิทยาของการตายของเซลล์ตามโปรแกรม (PCD) ในระหว่าง การพัฒนาของ ระบบประสาท[ 33 ]
บทบาทในการพัฒนาของระบบประสาท
มีการสังเกตพบ PCD ในระบบประสาทที่กำลังพัฒนาทั้งในเซลล์ที่กำลังแบ่งตัวและเซลล์หลังการแบ่งตัว[ 30 ]ทฤษฎีหนึ่งเสนอว่า PCD เป็นกลไกการปรับตัวเพื่อควบคุมจำนวนเซลล์ต้นกำเนิด ในมนุษย์ PCD ในเซลล์ต้นกำเนิดเริ่มต้นในสัปดาห์ ที่7 ของการตั้งครรภ์และคงอยู่จนถึงไตรมาสแรก[ 34 ]กระบวนการตายของเซลล์นี้ได้รับการระบุในบริเวณต้นกำเนิดของเปลือกสมอง สมองน้อยทาลามัส ก้านสมองและไขสันหลังรวมถึงบริเวณอื่นๆ[ 33 ]ในสัปดาห์ที่ 19-23 ของการตั้งครรภ์ พบ PCD ในเซลล์หลังการแบ่งตัว[ 35 ]ทฤษฎีที่แพร่หลายที่อธิบายการสังเกตนี้คือทฤษฎี neurotrophic ซึ่งระบุว่า PCD จำเป็นต่อการปรับการเชื่อมต่อระหว่างเซลล์ประสาทและอินพุตขาเข้าและเป้าหมายขาออกให้เหมาะสมที่สุด[ 33 ]ทฤษฎีอีกทฤษฎีหนึ่งเสนอว่า PCD ที่เกิดขึ้นในระหว่างการพัฒนาของระบบประสาทเกิดขึ้นเพื่อแก้ไขข้อผิดพลาดในเซลล์ประสาทที่อพยพไปยังตำแหน่งที่ไม่ถูกต้อง เชื่อมต่อกับเป้าหมายที่ไม่ถูกต้อง หรือมีแอกซอนที่ผิดทางในระหว่างการค้นหาเส้นทาง[ 36 ]เป็นไปได้ว่า PCD ในระหว่างการพัฒนาของระบบประสาททำหน้าที่ต่างๆ ที่กำหนดโดยระยะการพัฒนา ประเภทของเซลล์ และแม้แต่สายพันธุ์[ 33 ]
ทฤษฎีนิวโรโทรฟิก
ทฤษฎีนิวโรโทรฟิกเป็นสมมติฐานหลักที่ใช้ในการอธิบายบทบาทของการตายของเซลล์ตามโปรแกรมในระบบประสาทที่กำลังพัฒนา[ 37 ]ทฤษฎีนี้ตั้งสมมติฐานว่า เพื่อให้แน่ใจว่ามีการเชื่อมต่อเส้นประสาทไปยังเป้าหมายอย่างเหมาะสม เซลล์ประสาทจำนวนมากเกินไปจะถูกผลิตขึ้นก่อน จากนั้นเซลล์ประสาทเหล่านั้นจะแข่งขันกันเพื่อแย่งชิงปัจจัยนิวโรโทรฟิก ที่ปกป้องในปริมาณจำกัด และมีเพียงส่วนน้อยเท่านั้นที่รอดชีวิต ในขณะที่เซลล์ประสาทอื่นๆ จะตายไปตามโปรแกรมการตายของเซลล์[ 34 ]นอกจากนี้ ทฤษฎียังระบุว่าปัจจัยที่กำหนดไว้ล่วงหน้าจะควบคุมปริมาณเซลล์ประสาทที่รอดชีวิต และขนาดของประชากรเซลล์ประสาทที่เชื่อมต่อเส้นประสาทนั้นมีความสัมพันธ์โดยตรงกับอิทธิพลของบริเวณเป้าหมาย[ 38 ]
แนวคิดพื้นฐานที่ว่าเซลล์เป้าหมายหลั่งสารดึงดูดหรือสารกระตุ้น และปลายประสาทที่กำลังเจริญเติบโตมี ความไวต่อ สารเคมี ดึงดูดนั้น ถูกนำเสนอครั้งแรกโดยSantiago Ramon y Cajalในปี 1892 [ 39 ] Cajal นำเสนอแนวคิดนี้เพื่ออธิบาย "แรงอันชาญฉลาด" ที่แอกซอนดูเหมือนจะใช้เมื่อค้นหาเป้าหมาย แต่ยอมรับว่าเขาไม่มีข้อมูลเชิงประจักษ์[ 39 ]ทฤษฎีนี้ได้รับความสนใจมากขึ้นเมื่อการทดลองจัดการเป้าหมายของแอกซอนส่งผลให้เซลล์ประสาทที่เชื่อมต่อทั้งหมดตายลง ซึ่งพัฒนาแนวคิดของการควบคุมที่ได้มาจากเป้าหมาย ซึ่งกลายเป็นหลักการสำคัญในทฤษฎีนิวโรโทรฟิก[ 40 ] [ 41 ]การทดลองที่สนับสนุนทฤษฎีนี้เพิ่มเติมนำไปสู่การระบุปัจจัยนิวโรโทรฟิกตัวแรก คือปัจจัยการเจริญเติบโตของเส้นประสาท (NGF) [ 42 ]
ระบบประสาทส่วนปลายเทียบกับระบบประสาทส่วนกลาง

กลไกที่แตกต่างกันควบคุม PCD ในระบบประสาทส่วนปลาย (PNS) เมื่อเทียบกับระบบประสาทส่วนกลาง (CNS) ใน PNS การทำงานของเส้นประสาทที่เป้าหมายจะเป็นสัดส่วนกับปริมาณของปัจจัยนิวโรโทรฟิก NGF และ NT3 ที่ปล่อยออกมาจากเป้าหมาย[ 43 ] [ 44 ]การแสดงออกของตัวรับนิวโรโทรฟินTrkAและTrkCเพียงพอที่จะกระตุ้นให้เกิดอะพอพโทซิสได้แม้ไม่มีลิแกนด์[ 31 ]ดังนั้น จึงคาดการณ์ได้ว่า PCD ใน PNS ขึ้นอยู่กับการปล่อยปัจจัยนิวโรโทรฟิก และเป็นไปตามแนวคิดของทฤษฎีนิวโรโทรฟิก[ 45 ]
การตายของเซลล์ตามโปรแกรมในระบบประสาทส่วนกลางไม่ได้ขึ้นอยู่กับปัจจัยการเจริญเติบโต ภายนอก แต่อาศัยสัญญาณที่มาจากภายใน ในเปลือกสมองส่วนหน้าอัตราส่วน 4:1 ของเซลล์ประสาท ตัวกลางที่กระตุ้นและยับยั้ง จะถูกรักษาไว้โดยกลไกการตายของเซลล์แบบอะพอพโทซิส ซึ่งดูเหมือนว่าจะไม่ขึ้นอยู่กับสภาพแวดล้อม[ 44 ]หลักฐานสนับสนุนมาจากการทดลองที่เซลล์ต้นกำเนิดของเซลล์ประสาทตัวกลางถูกปลูกถ่ายเข้าไปในเปลือกสมองส่วนหน้าของหนูหรือเพาะเลี้ยงในหลอดทดลอง [ 46 ] เซลล์ที่ปลูกถ่ายตายเมื่ออายุได้สองสัปดาห์ ซึ่งเป็นอายุเดียวกับที่เซลล์ประสาทตัวกลางภายในร่างกายเกิดอะพอพโทซิส ไม่ว่าขนาดของการปลูกถ่ายจะเป็นเท่าใด สัดส่วนของเซลล์ที่เกิดอะพอพโทซิสก็ยังคงคงที่ นอกจากนี้ การรบกวนTrkBซึ่งเป็นตัวรับสำหรับปัจจัยการเจริญเติบโตของเซลล์ประสาทที่ได้จากสมอง (Bdnf) ก็ไม่มีผลต่อการตายของเซลล์ นอกจากนี้ยังแสดงให้เห็นว่าในหนูที่ไม่มีปัจจัยโปรอะพอพโทซิสBax (โปรตีน X ที่เกี่ยวข้องกับ Bcl-2) เปอร์เซ็นต์ของเซลล์ประสาทตัวกลางที่รอดชีวิตมีมากกว่าเมื่อเทียบกับหนูป่า[ 46 ]ผลการค้นพบเหล่านี้บ่งชี้ว่าการตายของเซลล์ตามโปรแกรมใน CNS ใช้ประโยชน์จากการส่งสัญญาณผ่าน Bax บางส่วน และเป็นอิสระจาก BDNF และสิ่งแวดล้อม กลไกการเกิดอะพอพโทซิสใน CNS ยังไม่เป็นที่เข้าใจดีนัก แต่เชื่อกันว่าการเกิดอะพอพโทซิสของอินเตอร์นิวรอนเป็นกระบวนการที่เกิดขึ้นเองโดยอิสระ[ 45 ]
การพัฒนาของระบบประสาทในกรณีที่ขาดสิ่งนี้
การตายของเซลล์ตามโปรแกรมสามารถลดลงหรือกำจัดได้ในระบบประสาทที่กำลังพัฒนาโดยการลบยีนที่กระตุ้นการตายของเซลล์หรือโดยการแสดงออกมากเกินไปของยีนที่ต่อต้านการตายของเซลล์ การไม่มีหรือการลดลงของ PCD อาจทำให้เกิดความผิดปกติทางกายวิภาคที่ร้ายแรง แต่ก็อาจส่งผลให้เกิดผลกระทบน้อยที่สุดได้เช่นกัน ขึ้นอยู่กับยีนเป้าหมาย ประชากรเซลล์ประสาท และระยะของการพัฒนา[ 33 ]การแพร่กระจายของเซลล์ต้นกำเนิดมากเกินไปที่นำไปสู่ความผิดปกติของสมองอย่างรุนแรงมักเป็นอันตรายถึงชีวิต ดังที่เห็นในหนูที่ขาดcaspase-3หรือcaspase-9 ซึ่งพัฒนาเป็น exencephalyในสมองส่วนหน้า[ 47 ] [ 48 ]อย่างไรก็ตาม ก้านสมอง ไขสันหลัง และปมประสาทส่วนปลายของหนูเหล่านี้พัฒนาตามปกติ ซึ่งบ่งชี้ว่าการมีส่วนร่วมของcaspaseใน PCD ในระหว่างการพัฒนาขึ้นอยู่กับบริเวณสมองและชนิดของเซลล์[ 49 ]การกำจัดหรือการยับยั้งปัจจัยกระตุ้นโปรตีเอสอะพอพโทซิส 1 ( APAF1 ) ยังส่งผลให้เกิดความผิดปกติและการตายของตัวอ่อนเพิ่มขึ้น[ 50 ] [ 51 ] [ 52 ]การจัดการโปรตีนควบคุมอะพอพโทซิสBcl-2และ Bax (การแสดงออกมากเกินไปของ Bcl-2 หรือการลบ Bax) ทำให้จำนวนเซลล์ประสาทเพิ่มขึ้นในบางบริเวณของระบบประสาท เช่นเรตินานิวเคลียสไตรเจมินัล ซีรีเบลลัม และไขสันหลัง[ 53 ] [ 54 ] [ 55 ] [ 56 ] [ 57 ] [ 58 ] [ 59 ]อย่างไรก็ตาม PCD ของเซลล์ประสาทเนื่องจากการลบ Bax หรือการแสดงออกมากเกินไปของ Bcl-2 ไม่ได้ส่งผลให้เกิดความผิดปกติทางสัณฐานวิทยาหรือพฤติกรรมที่เด่นชัดในหนู ตัวอย่างเช่น หนูที่แสดงออก Bcl-2 มากเกินไปโดยทั่วไปจะมีทักษะการเคลื่อนไหวและการมองเห็นปกติ และแสดงอาการบกพร่องเฉพาะในพฤติกรรมที่ซับซ้อน เช่น การเรียนรู้และความวิตกกังวล[ 60 ] [ 61 ] [ 62 ]ลักษณะพฤติกรรมปกติของหนูเหล่านี้บ่งชี้ว่าอาจมีกลไกการปรับตัวเข้ามาเกี่ยวข้องเพื่อชดเชยเซลล์ประสาทส่วนเกิน[ 33 ]
สัตว์ไม่มีกระดูกสันหลังและสัตว์มีกระดูกสันหลัง

การเรียนรู้เกี่ยวกับ PCD ในสิ่งมีชีวิตชนิดต่างๆ เป็นสิ่งจำเป็นในการทำความเข้าใจพื้นฐานทางวิวัฒนาการและเหตุผลของการเกิดอะพอพโทซิสในการพัฒนาของระบบประสาท ในระหว่างการพัฒนาของระบบ ประสาท ของสัตว์ไม่มีกระดูกสันหลัง PCD มีบทบาทที่แตกต่างกันในแต่ละชนิด[ 63 ]ความคล้ายคลึงกันของกลไกการตายของเซลล์แบบไม่สมมาตรในหนอนตัวกลมและปลิงบ่งชี้ว่า PCD อาจมีความสำคัญทางวิวัฒนาการในการพัฒนาของระบบประสาท[ 64 ]ในหนอนตัวกลม PCD เกิดขึ้นในชั่วโมงแรกของการพัฒนา ส่งผลให้เซลล์ที่ไม่ใช่เซลล์สืบพันธุ์ 12% รวมถึงเซลล์ประสาทถูกกำจัด[ 65 ]การตายของเซลล์ในสัตว์ขาปล้องเกิดขึ้นครั้งแรกในระบบประสาทเมื่อ เซลล์ เอกโตเดิร์มเกิดการเปลี่ยนแปลง และเซลล์ลูกหนึ่งเซลล์กลายเป็นนิวโรบลาสต์ ส่วนอีกเซลล์หนึ่งจะเกิดอะพอพโทซิส[ 66 ]นอกจากนี้ การตายของเซลล์ที่กำหนดเป้าหมายตามเพศนำไปสู่การสร้างเส้นประสาทที่แตกต่างกันของอวัยวะเฉพาะในเพศผู้และเพศเมีย[ 67 ]ในแมลงหวี่ PCD เป็นสิ่งจำเป็นในการแบ่งส่วนและการกำหนดลักษณะเฉพาะในระหว่างการพัฒนา
ตรงกันข้ามกับสัตว์ไม่มีกระดูกสันหลัง กลไกการตายของเซลล์แบบโปรแกรมพบว่ามีการอนุรักษ์ไว้มากกว่าในสัตว์มีกระดูกสันหลังการศึกษาอย่างกว้างขวางที่ดำเนินการกับสัตว์มีกระดูกสันหลังหลายชนิดแสดงให้เห็นว่า PCD ของเซลล์ประสาทและเซลล์เกลียเกิดขึ้นในส่วนใหญ่ของระบบประสาทระหว่างการพัฒนา มีการสังเกตพบก่อนและระหว่างการสร้างไซแนปส์ในระบบประสาทส่วนกลางและระบบประสาทส่วนปลาย[ 33 ]อย่างไรก็ตาม มีความแตกต่างเล็กน้อยระหว่างสายพันธุ์สัตว์มีกระดูกสันหลัง ตัวอย่างเช่นสัตว์เลี้ยงลูกด้วยนมแสดงให้เห็นการแตกแขนงอย่างกว้างขวางตามด้วย PCD ในเรตินา ในขณะที่นกไม่เป็นเช่นนั้น[ 68 ]แม้ว่าการปรับแต่งไซแนปส์ในระบบสัตว์มีกระดูกสันหลังส่วนใหญ่จะขึ้นอยู่กับ PCD แต่กลไกวิวัฒนาการอื่นๆ ก็มีบทบาทเช่นกัน[ 33 ]
ในเนื้อเยื่อพืช
การตายของเซลล์ตามโปรแกรมในพืชมีลักษณะทางโมเลกุลคล้ายคลึงกับ การตายของเซลล์ แบบอะพอพโทซิส ในสัตว์ แต่ก็มีความแตกต่างกันด้วย ความแตกต่างที่เห็นได้ชัดที่สุดคือการมีผนังเซลล์และไม่มีระบบภูมิคุ้มกันที่กำจัดชิ้นส่วนของเซลล์ที่ตายแล้ว แทนที่จะเป็นการตอบสนองทางภูมิคุ้มกัน เซลล์ที่กำลังจะตายจะสังเคราะห์สารเพื่อสลายตัวเองและนำไปไว้ในแวคิวโอลซึ่งจะแตกออกเมื่อเซลล์ตาย[ 69 ]
ใน "APL ควบคุมเอกลักษณ์ของเนื้อเยื่อหลอดเลือดในArabidopsis " [ 70 ] Martin Bonke และเพื่อนร่วมงานของเขาได้ระบุว่าหนึ่งในสองระบบการขนส่งระยะไกลในพืชมีหลอดเลือดคือไซเล็มประกอบด้วยเซลล์หลายประเภท "ซึ่งการแบ่งแยกนั้นเกี่ยวข้องกับการสะสมของผนังเซลล์ ที่หนาขึ้นอย่างซับซ้อน และการตายของเซลล์ตามโปรแกรม" ผู้เขียนเน้นย้ำว่าผลิตภัณฑ์ของ PCD ในพืชมีบทบาทโครงสร้างที่สำคัญ
ลักษณะทางสัณฐานวิทยาและชีวเคมีพื้นฐานของ PCD ได้รับการอนุรักษ์ไว้ทั้งในอาณาจักร พืชและ สัตว์[ 71 ]เซลล์พืชบางประเภทดำเนินโปรแกรมการตายของเซลล์ที่ไม่เหมือนใคร ซึ่งมีลักษณะร่วมกันกับอะพอพโทซิสในสัตว์ เช่น การย่อยสลาย DNA ในนิวเคลียสแต่ก็มีลักษณะเฉพาะของตัวเองด้วย เช่น การย่อยสลายของ นิวเคลียสที่เกิดจากการยุบตัวของแวคิวโอลใน องค์ประกอบ ท่อลำเลียงของไซเล็ม[ 72 ]
Janneke Balk และ Christopher J. Leaver จากภาควิชาวิทยาศาสตร์พืชมหาวิทยาลัยออกซ์ฟอร์ดได้ทำการวิจัยเกี่ยวกับการกลายพันธุ์ในจีโนมไมโทคอนเดรียของ เซลล์ ดอกทานตะวันผลการวิจัยนี้ชี้ให้เห็นว่าไมโทคอนเดรีย มีบทบาทสำคัญเช่นเดียวกันใน PCD ของพืชมีท่อลำเลียงเช่นเดียวกับใน เซลล์ยูคาริโอตอื่นๆ[ 73 ]
PCD ในละอองเกสรช่วยป้องกันการผสมพันธุ์ในสายเลือดเดียวกัน
ระหว่างการผสมเกสรพืชจะบังคับใช้ความไม่เข้ากันเอง ( SI ) ซึ่งเป็นวิธีการสำคัญในการป้องกันการผสมตัวเองการวิจัยเกี่ยวกับต้นป๊อปปี้ข้าวโพด ( Papaver rhoeas ) พบว่าโปรตีนในเกสรตัวเมียที่ละอองเรณูตกลงมานั้น จะมีปฏิสัมพันธ์กับละอองเรณูและกระตุ้นให้เกิด PCD ในละอองเรณูที่ไม่เข้ากัน (เช่น ละอองเรณูที่ผสม ตัวเอง ) นักวิจัย Steven G. Thomas และVernonica E. Franklin-Tongยังพบว่าการตอบสนองดังกล่าวเกี่ยวข้องกับการยับยั้งการเจริญเติบโตของท่อละอองเรณู อย่างรวดเร็ว ตามด้วย PCD [ 74 ]
ในราเมือก
ราเมือก สังคมDictyostelium discoideumมีลักษณะพิเศษคือสามารถปรับตัวให้มี พฤติกรรมคล้าย อะมีบา ที่เป็นผู้ล่า ใน รูปแบบ เซลล์เดียวหรือรวมตัวกันเป็น รูปแบบคล้าย ทากที่ เคลื่อนที่ได้ เมื่อกระจายสปอร์ที่จะให้กำเนิดรุ่น ต่อ ไป[ 75 ]
ก้านประกอบด้วยเซลล์ที่ตายแล้วซึ่งผ่านกระบวนการ PCD ชนิดหนึ่งที่มีลักษณะร่วมกันหลายประการกับการตายของเซลล์แบบออโตฟาจิก ได้แก่ การเกิดแวคิวโอลขนาดใหญ่ภายในเซลล์ การควบแน่นของโครมาติน ในระดับหนึ่ง แต่ไม่มีการแตกตัวของ DNA [ 76 ]บทบาทเชิงโครงสร้างของสารตกค้างที่เหลือจากเซลล์ที่ตายแล้วนั้นชวนให้นึกถึงผลิตภัณฑ์ของ PCD ในเนื้อเยื่อพืช
D. discoideumเป็นราเมือกชนิดหนึ่ง ซึ่งเป็นส่วนหนึ่งของสาขาที่อาจกำเนิดมาจากบรรพบุรุษยูคาริโอต เมื่อประมาณหนึ่ง พันล้านปีก่อนปัจจุบัน ดูเหมือนว่าพวกมันจะกำเนิดขึ้นหลังจากบรรพบุรุษของพืชสีเขียวและบรรพบุรุษของเชื้อราและสัตว์ได้แยกตัวออกจากกันแล้ว แต่ นอกเหนือจากตำแหน่งของพวกมันในแผนภูมิ วิวัฒนาการ แล้ว ข้อเท็จจริงที่ว่ามีการสังเกต PCD ในD. discoideum ที่เรียบง่ายและมี โครโมโซม หกชุด นั้น มีความสำคัญเพิ่มเติมอีกด้วย นั่นคือ ช่วยให้สามารถศึกษาเส้นทางการพัฒนาของ PCD ที่ไม่ขึ้นอยู่กับแคสเปสซึ่งเป็นลักษณะเฉพาะของอะพอพโทซิส[ 77 ]
ต้นกำเนิดเชิงวิวัฒนาการของอะพอพโทซิสในไมโทคอนเดรีย
การเกิดการตายของเซลล์ตามโปรแกรมในโปรติสต์เป็นไปได้[ 78 ] [ 79 ]แต่ยังคงเป็นที่ถกเถียงกันอยู่ บางคนจัดประเภทการตายในสิ่งมีชีวิตเหล่านั้นเป็นการตายของเซลล์แบบอะพอพโทซิสที่ไม่ได้รับการควบคุม[ 80 ] [ 79 ]
นักชีววิทยาสงสัยมานานแล้วว่าไมโตคอนเดรียมีต้นกำเนิดมาจากแบคทีเรียที่ถูกรวมเข้าเป็นเอนโดซิมไบออนต์ ("อาศัยอยู่ร่วมกันภายใน") ของเซลล์ยูคาริโอติกขนาดใหญ่ลินน์ มาร์กูลิส เป็นผู้ที่สนับสนุน ทฤษฎีนี้มาตั้งแต่ปี 1967 ซึ่งต่อมาได้รับการยอมรับอย่างกว้างขวาง[ 81 ]หลักฐานที่น่าเชื่อถือที่สุดสำหรับทฤษฎีนี้คือข้อเท็จจริงที่ว่าไมโตคอนเดรียมีดีเอ็นเอ ของตัวเอง และมียีนและอุปกรณ์ การจำลองแบบ
ขั้นตอน วิวัฒนาการนี้ถือเป็นความเสี่ยงสำหรับเซลล์ยูคาริโอตดั้งเดิม ซึ่งเริ่มกลืนกิน แบคทีเรีย ที่ผลิตพลังงานเช่นเดียวกับเป็นขั้นตอนที่อันตรายสำหรับบรรพบุรุษของไมโทคอนเดรีย ซึ่งเริ่มบุกรุกโฮสต์ โปรโตยูคาริโอตของพวกมัน กระบวนการนี้ยังคงปรากฏให้เห็นในปัจจุบัน ระหว่างเซลล์เม็ดเลือดขาวของมนุษย์กับแบคทีเรีย ส่วนใหญ่แล้ว แบคทีเรียที่บุกรุกจะถูกทำลายโดยเซลล์เม็ดเลือดขาว อย่างไรก็ตาม ไม่ใช่เรื่องแปลกที่สงครามเคมีที่กระทำโดยโปรคาริโอตจะประสบความสำเร็จ โดยมีผลที่ตามมาคือการติดเชื้อจากความเสียหายที่เกิดขึ้น
หนึ่งในเหตุการณ์วิวัฒนาการที่หายากเหล่านี้ ซึ่งเกิดขึ้นเมื่อประมาณสองพันล้านปีก่อน ทำให้ยูคาริโอตบางชนิดและโปรคาริโอตที่ผลิตพลังงานสามารถอยู่ร่วมกันและได้รับประโยชน์ร่วมกันจากภาวะพึ่งพาอาศัย กัน ได้[ 82 ]
เซลล์ยูคาริโอตที่มีไมโตคอนเดรียดำรงชีวิตอยู่ระหว่างความเป็นและความตาย เนื่องจากไมโตคอนเดรียยังคงรักษาสารโมเลกุลที่สามารถกระตุ้นการฆ่าตัวตายของเซลล์ได้[ 83 ]ยังไม่เป็นที่แน่ชัดว่าเหตุใดกลไกอะพอพโทซิสจึงยังคงอยู่ในสิ่งมีชีวิตเซลล์เดียวในปัจจุบัน กระบวนการนี้ได้วิวัฒนาการให้เกิดขึ้นเฉพาะเมื่อมีการกำหนด[ 84 ]ให้กับเซลล์ (เช่น การตอบสนองจากเซลล์ข้างเคียง ความเครียด หรือความเสียหายของ DNA ) ไมโตคอนเดรียจะปล่อยตัว กระตุ้น แคสเปส ที่กระตุ้นกระบวนการ ทางชีวเคมีที่เหนี่ยวนำให้เกิดการตายของเซลล์ด้วยเหตุนี้กลไก การฆ่าตัวตายของเซลล์ จึงมีความสำคัญต่อชีวิตของเราทุกคนในปัจจุบัน
ความเสียหายของดีเอ็นเอและอะพอพโทซิส
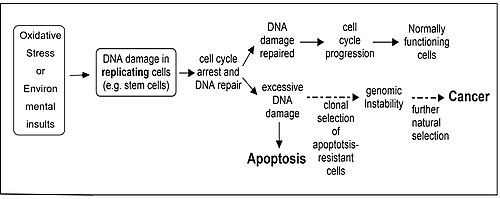
การซ่อมแซมความเสียหายของ DNAและอะพอพโทซิสเป็นกระบวนการทางเอนไซม์สองอย่างที่จำเป็นสำหรับการรักษา ความสมบูรณ์ของ จีโนมในมนุษย์ เซลล์ที่ขาดความสามารถในการซ่อมแซม DNA มีแนวโน้มที่จะสะสมความเสียหายของ DNAและเมื่อเซลล์ดังกล่าวมีข้อบกพร่องในอะพอพโทซิสด้วย พวกมันก็มีแนวโน้มที่จะอยู่รอดได้แม้จะมีความเสียหายของ DNA มากเกินไป[ 85 ] การจำลองแบบของ DNA ในเซลล์ดังกล่าวทำให้เกิดการกลายพันธุ์และการกลายพันธุ์เหล่านี้อาจทำให้เกิดมะเร็ง (ดูรูป) เส้นทางเอนไซม์หลายเส้นทางได้วิวัฒนาการขึ้นเพื่อซ่อมแซมความเสียหายของ DNA ประเภทต่างๆ และพบว่าในเส้นทางการซ่อมแซม DNA ที่ได้รับการศึกษาอย่างดีห้าเส้นทาง เอนไซม์บางชนิดมีบทบาทสองอย่าง โดยบทบาทหนึ่งคือการมีส่วนร่วมในการซ่อมแซมความเสียหายเฉพาะประเภท และบทบาทที่สองคือการกระตุ้นให้เกิดอะพอพโทซิสหากระดับความเสียหายของ DNA ดังกล่าวเกินความสามารถในการซ่อมแซมของเซลล์[ 85 ] โปรตีน ที่ มีบทบาทสองอย่างนี้มีแนวโน้มที่จะป้องกันการเกิดมะเร็ง โปรตีนที่ทำหน้าที่ในบทบาทสองอย่างนี้สำหรับแต่ละกระบวนการซ่อมแซม ได้แก่: (1) การซ่อมแซม DNA ที่ไม่ตรงกันMSH2 , MSH6 , MLH1และPMS2 ; (2) การซ่อมแซมการตัดฐาน , APEX1 (REF1/APE), โพลี(ADP-ริโบส) โพลีเมอเรส (PARP); (3) การซ่อมแซมการตัดนิวคลีโอไทด์ , XPB , XPD ( ERCC2 ), p53 , p33 ( ING1b ); (4) การเชื่อมต่อปลายที่ไม่เหมือนกัน , หน่วยย่อยเร่งปฏิกิริยาของDNA-PK ; (5) การ ซ่อมแซม การรวมตัวใหม่ที่เหมือนกัน , BRCA1 , ATM , ATR , WRN , BLM , Tip60 , p53
การตายตามโปรแกรมของสิ่งมีชีวิตทั้งตัว
ความสำคัญทางคลินิก
เอบีแอล
พบว่าออนโคยีน BCR-ABL มีส่วนเกี่ยวข้องกับการเกิดมะเร็งในมนุษย์[ 86 ]
ซี-ไมค
c-Mycมีส่วนเกี่ยวข้องในการควบคุมอะพอพโทซิสผ่านบทบาทในการลดการแสดงออกของ ยีน Bcl-2บทบาทของมันคือการเจริญเติบโตที่ผิดปกติของเนื้อเยื่อ[ 86 ]
การแพร่กระจาย
ลักษณะทางโมเลกุลของเซลล์ที่แพร่กระจายคือการแสดงออกของยีนอะพอพโทซิสหลายตัวที่เปลี่ยนแปลงไป[ 86 ]
ดูเพิ่มเติม
- อะโนอิกิส
- ปัจจัยกระตุ้นอะพอพโทซิส
- อะพอพโทซิสเทียบกับ ซูโดอะพอ พโทซิส
- อะพอปโทโซม
- การแตกตัวของดีเอ็นเอในกระบวนการอะพอพโทซิส
- การสลายตัวด้วยตนเอง (ชีววิทยา)
- ออโตฟาจี
- ออโตสคิซิส
- บีซีแอล-2
- สารกระตุ้นการตายของโดเมนที่ทำปฏิกิริยากับ BH3 (BID)
- คาลเพนส์
- แคสเปส
- ความเสียหายของเซลล์
- การเกิดเคราติน
- ไซโตโครม ซี
- ความเป็นพิษต่อเซลล์
- ไดอาโบล โฮโมล็อก
- เอนโทซิส
- ความเป็นพิษต่อเซลล์ประสาท
- เฟอร์โรพโทซิส
- อินฟลามาโซม
- รูพรุนการเปลี่ยนแปลงสภาพการซึมผ่านของไมโตคอนเดรีย
- หายนะของการแบ่งเซลล์แบบไมโทซิส
- เนโครไบโอโลยี
- เนโครพโทซิส
- เนื้อเยื่อตาย
- p53 upregulated modulator of apoptosis (PUMA)
- พาราปโทซิส
- พาร์ธาเนโทส
- ไพรอพโทซิส
- ไรพีไอ ไคเนส
- การเสื่อมสภาพแบบวอลเลเรียน
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การตายของเซลล์ตามโปรแกรม
การตายของเซลล์ตามโปรแกรม ( PCD ) บางครั้งเรียกว่าการฆ่าตัวตายของเซลล์หรือการฆ่าตัวตายของเซลล์ คือการตายของเซลล์อันเป็นผลมาจากเหตุการณ์ภายในเซลล์ เช่นอะพอพโทซิสหรือออโตฟาจี
ประวัติศาสตร์
แนวคิดเรื่อง "การตายของเซลล์ตามโปรแกรม" ถูกนำมาใช้โดย Lockshin & Williams [ 9 ] ในปี พ.ศ.
ประเภท
ภาพรวมของเส้นทางการส่งสัญญาณที่เกี่ยวข้องกับกระบวนการ อะพอพโทซิส อะพอพโทซิส หรือ การตายของเซลล์ประเภทที่ 1 การตายของเซลล์แบบออโตฟาจิก หรือการตายของเซลล์ประเภท II ( ไซโตพลาสมิก : มีลักษณะเฉพาะคือการก่อตัวของ แวคิวโอล ขนาดใหญ่ ที่กัดกิน ออร์แกเนลล์...
อะพอพโทซิส
อะพอพโทซิส เป็นกระบวนการตายของเซลล์ตามโปรแกรม (PCD) ที่อาจเกิดขึ้นใน สิ่งมีชีวิตหลายเซลล์ [ 15 ] เหตุการณ์ ทางชีวเคมี นำไปสู่การเปลี่ยนแปลงของเซลล์ ( สัณฐานวิทยา ) และการตายของเซลล์ การเปลี่ยนแปลงเหล่านี้รวมถึง การเกิดฟอง การหดตัวของเซลล์การแตกตัว...