อ่าน 43 นาที
ประวัติวิวัฒนาการของพืช
วิวัฒนาการของพืชส่งผลให้เกิดความซับซ้อนที่หลากหลาย ตั้งแต่แผ่นสาหร่าย ยุคแรกสุด ของอาร์คีพลาส ติดเซลล์เดียวที่ วิวัฒนาการผ่านเอนโดซิมไบโอซิสไปจนถึงสาหร่ายสีเขียวในทะเลและน้ำจืด...
ประวัติวิวัฒนาการของพืช

วิวัฒนาการของพืชส่งผลให้เกิดความซับซ้อนที่หลากหลาย ตั้งแต่แผ่นสาหร่าย ยุคแรกสุด ของอาร์คีพลาส ติดเซลล์เดียวที่ วิวัฒนาการผ่านเอนโดซิมไบโอซิสไปจนถึงสาหร่ายสีเขียวในทะเลและน้ำจืด แบบหลายเซลล์ ไปจนถึงไบรโอไฟต์บนบกที่มีส ปอร์ ไลโคพอดและเฟิร์นและในที่สุดก็มาถึงพืชเมล็ดเปลือยและพืชดอก ( พืช มีดอก ) ที่ซับซ้อน ซึ่งมีเมล็ดในปัจจุบัน ในขณะที่กลุ่มแรกๆ หลายกลุ่มยังคงเจริญเติบโตได้ดี ดังเช่น สาหร่าย สีแดงและสีเขียวในสภาพแวดล้อมทางทะเล กลุ่มที่เกิดขึ้นใหม่ในภายหลังได้เข้ามาแทนที่กลุ่มที่เคยมีบทบาทเด่นทางนิเวศวิทยามาก่อน ตัวอย่างเช่น การขึ้นมามีบทบาทของพืชดอกเหนือพืชเมล็ดเปลือยในสภาพแวดล้อมบนบก[ 1 ] : 498
มีหลักฐานว่าไซยาโนแบคทีเรียและ ยูคาริโอต แบบทัลลอย ด์หลายเซลล์ อาศัยอยู่ในชุมชนน้ำจืดบนบกตั้งแต่เมื่อ 1 พันล้านปีก่อน[ 2 ]และชุมชนของสิ่งมีชีวิตหลายเซลล์ที่ซับซ้อนซึ่งสังเคราะห์แสงได้มีอยู่บนบกในช่วงปลายยุคพรีแคมเบรียนประมาณ850 ล้านปีก่อน[ 3 ]
หลักฐานการกำเนิดของ พืชบก เอ็มบริโอไฟต์ปรากฏขึ้นครั้งแรกในช่วงกลางยุคออร์โดวิเชียน (ประมาณ470 ล้านปีก่อน ) ในช่วงกลางยุคเดวอนเนียน (ประมาณ390 ล้านปีก่อน ) หลักฐานฟอสซิลแสดงให้เห็นว่าลักษณะหลายอย่างที่พบในพืชบกในปัจจุบันมีอยู่แล้ว รวมถึงรากและใบเมื่อไม่นานมานี้ หลักฐานทางธรณีเคมีชี้ให้เห็นว่าในช่วงเวลานี้ อาณาจักรบนบกส่วนใหญ่ถูกยึดครอง ซึ่งเปลี่ยนแปลงสภาพแวดล้อมการผุกร่อนบนบกทั่วโลก[ 4 ]ในช่วงปลายยุคเดวอนเนียน (ประมาณ370 ล้านปีก่อน ) พืชที่สร้างสปอร์อิสระบางชนิด เช่นอาร์คีออปเทอริสมีเนื้อเยื่อหลอดเลือดทุติยภูมิที่สร้างเนื้อไม้และก่อตัวเป็นป่าต้นไม้ สูง นอกจากนี้ ในช่วงปลายยุคเดวอนเนียนเอลกินเซีย ซึ่งเป็น เฟิร์นเมล็ดแรกได้วิวัฒนาการเมล็ด[ 5 ] นวัตกรรมทางวิวัฒนาการยังคงดำเนินต่อไปตลอดช่วงที่เหลือของ ยุค ฟาเนโรโซอิกและยังคงดำเนินต่อไปจนถึงปัจจุบัน กลุ่มพืชส่วนใหญ่ไม่ได้รับผลกระทบมากนักจากเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในยุคเพอร์เมียน-ไทร แอสสิก แม้ว่าโครงสร้างของชุมชนจะเปลี่ยนแปลงไปก็ตาม นี่อาจเป็นพื้นฐานสำหรับการปรากฏตัวของพืชดอกในยุคไทรแอสสิก (ประมาณ200 ล้านปีก่อน ) และการกระจายตัวของพืชดอกในยุคครีเทเชียสและพาลีโอจีนในเวลาต่อมากลุ่มพืชที่สำคัญกลุ่มล่าสุดที่วิวัฒนาการขึ้นมาคือหญ้าซึ่งมีความสำคัญในยุคพาลีโอจีนตอนกลาง ตั้งแต่ประมาณ40 ล้านปีก่อนหญ้า รวมถึงกลุ่มพืชอื่นๆ อีกมากมาย ได้วิวัฒนาการกลไกการเผาผลาญแบบใหม่เพื่อความอยู่รอดในสภาพที่มี CO2 ต่ำและสภาพอากาศอบอุ่นแห้งแล้งของเขต ร้อน ในช่วง10ล้านปี ที่ผ่าน มา
การล่าอาณานิคมของที่ดิน
ความแตกต่าง
พืชบกวิวัฒนาการมาจากกลุ่มสาหร่ายสีเขียว น้ำจืด อาจจะเร็วที่สุดเมื่อ 850 ล้านปีก่อน[ 3 ]แต่พืชที่มีลักษณะคล้ายสาหร่ายอาจวิวัฒนาการมาตั้งแต่ 1 พันล้านปีก่อน[ 2 ]ญาติที่ใกล้ชิดที่สุดของพืชบกในปัจจุบันคือชาโรไฟต์โดยเฉพาะอย่างยิ่งCharalesหาก Charales ในปัจจุบันมีความคล้ายคลึงกับบรรพบุรุษที่ห่างไกลที่พวกมันมีร่วมกับพืชบก นั่นหมายความว่าพืชบกวิวัฒนาการมาจากสาหร่ายที่มีกิ่งก้านสาขาและเป็นเส้นใยที่อาศัยอยู่ในน้ำจืดตื้น[ 6 ]อาจจะอยู่ที่ขอบของสระน้ำที่แห้งเหือดตามฤดูกาล[ 7 ]อย่างไรก็ตาม หลักฐานล่าสุดบางอย่างชี้ให้เห็นว่าพืชบกอาจมีต้นกำเนิดมาจากชาโรไฟต์ บนบกเซลล์เดียว ที่คล้ายกับKlebsormidiophyceae ที่มีอยู่ในปัจจุบัน [ 8 ]สาหร่ายจะมีวงจรชีวิตแบบแฮพลอนติก ในช่วงเวลาสั้นๆ โครโมโซม คู่ ( สภาวะ ดิพลอยด์ ) จะเกิดขึ้นเมื่อไข่และอสุจิรวมตัวกันเพื่อสร้างไซโกตซึ่งจะแบ่งตัวทันทีด้วยไมโอซิสเพื่อสร้างเซลล์ที่มีโครโมโซมที่ไม่จับคู่กันครึ่งหนึ่ง ( สภาวะ แฮพลอยด์ ) ปฏิสัมพันธ์แบบร่วมมือกับเชื้อราอาจช่วยให้พืชยุคแรกปรับตัวเข้ากับความเครียดของสภาพแวดล้อมบนบกได้[ 9 ]

ความท้าทายต่อการตั้งถิ่นฐานบนที่ดิน
พืชไม่ใช่สิ่งมีชีวิตสังเคราะห์แสงกลุ่มแรกบนบก อัตราการผุกร่อนบ่งชี้ว่าสิ่งมีชีวิตที่สามารถสังเคราะห์แสงได้นั้นอาศัยอยู่บนบกมาตั้งแต่1,200 ล้านปีก่อน [ 7 ]และพบฟอสซิลจุลินทรีย์ในตะกอนทะเลสาบน้ำจืดตั้งแต่1,000 ล้านปีก่อน [ 10 ]แต่บันทึกไอโซโทปคาร์บอนบ่งชี้ว่าพวกมันมีจำนวนน้อยเกินไปที่จะส่งผลกระทบต่อองค์ประกอบของบรรยากาศจนกระทั่งประมาณ850 ล้านปีก่อน [ 3 ] สิ่งมีชีวิตเหล่านี้ แม้ว่าจะมีความหลากหลายทางสายพันธุ์[ 11 ]ก็อาจมีขนาดเล็กและเรียบง่าย ก่อตัวเป็นเพียงคราบสาหร่ายเท่านั้น[ 7 ]
เนื่องจากไลเคนเป็นขั้นตอนแรกในการสืบทอดทางนิเวศวิทยา ขั้นต้น ในบริบทปัจจุบัน สมมติฐานหนึ่งก็คือไลเคนขึ้นมาบนบกก่อนและอำนวยความสะดวกในการตั้งถิ่นฐานของพืช อย่างไรก็ตาม ทั้งวิวัฒนาการระดับโมเลกุลและบันทึกฟอสซิลดูเหมือนจะขัดแย้งกับสมมติฐานนี้[ 12 ]
มีเหตุผลหลายประการที่เป็นไปได้ว่าทำไมพืชบกจึงใช้เวลานานมากในการเกิดขึ้น อาจเป็นเพราะ 'พิษ' ในบรรยากาศทำให้ยูคาริโอตไม่สามารถเข้ามาอาศัยบนบกได้ก่อนที่พืชบกจะเกิดขึ้น[ 13 ]หรืออาจเป็นเพราะต้องใช้เวลานานมากในการวิวัฒนาการให้มีความซับซ้อนที่จำเป็น[ 14 ]ความท้าทายสำคัญในการปรับตัวบนบกคือการขาดแคลนดิน ที่ เหมาะสม[ 15 ]ตลอดบันทึกฟอสซิล ดินได้รับการอนุรักษ์ไว้ ทำให้ได้รับข้อมูลเกี่ยวกับลักษณะของดินในยุคแรก ก่อนที่พืชบกจะเกิดขึ้น ดินบนบกมีทรัพยากรที่จำเป็นต่อชีวิตน้อย เช่นไนโตรเจนและฟอสฟอรัสและมีความสามารถในการกักเก็บน้ำน้อย
การปรับตัวเพื่อการตั้งถิ่นฐานบนบก
หลักฐานของพืชบกยุคแรกสุดปรากฏขึ้นเมื่อประมาณ470 ล้านปีก่อนในหินยุคออร์โดวิเชียนตอนกลางตอนล่างจากซาอุดีอาระเบีย[ 16 ]และกอนด์วานา[ 17 ]ในรูปแบบของสปอร์ที่เรียกว่าคริปโตสปอร์สปอร์เหล่านี้มีผนังที่ทำจากสปอโรพอลเลนินซึ่งเป็นวัสดุที่ทนต่อการผุกร่อนอย่างมาก ทำให้พวกมันได้รับการอนุรักษ์ไว้อย่างดีในบันทึกฟอสซิล สปอร์เหล่านี้ถูกผลิตขึ้นเป็นเดี่ยว (โมนาด) เป็นคู่ (ไดแอด) หรือเป็นกลุ่มสี่ (เตตระด) และโครงสร้างจุลภาคของพวกมันคล้ายกับสปอร์ของลิเวอร์เวิร์ต ในปัจจุบัน ซึ่งบ่งชี้ว่าพวกมันมีระดับการจัดระเบียบที่เทียบเท่ากัน[ 18 ]ผนังของพวกมันมีสปอโรพอลเลนิน ซึ่งเป็นหลักฐานเพิ่มเติมของความสัมพันธ์แบบเอ็มบริโอไฟต์[ 19 ]
สปอร์แบบไตรเลทที่คล้ายกับของพืชมีท่อลำเลียงปรากฏขึ้นในเวลาไม่นานหลังจากนั้น ในหินยุคออร์โดวิเชียนตอนบนเมื่อประมาณ 455 ล้านปีก่อน[ 20 ] [ 21 ]ขึ้นอยู่กับเวลาที่เททราดแยกออก สปอร์แต่ละอันจากสี่อันอาจมี "เครื่องหมายไตรเลท" ซึ่งเป็นรูปตัว Y สะท้อนถึงจุดที่แต่ละเซลล์ถูกบีบอัดเข้ากับเซลล์ข้างเคียง[ 22 ]อย่างไรก็ตาม สิ่งนี้ต้องการให้ผนังสปอร์มีความแข็งแรงและทนทานในระยะแรก ความทนทานนี้เกี่ยวข้องอย่างใกล้ชิดกับการมีผนังด้านนอกที่ทนต่อการแห้ง ซึ่งเป็นลักษณะที่ใช้ได้เฉพาะเมื่อสปอร์ต้องอยู่รอดนอกน้ำ อันที่จริง แม้แต่เอ็มบริโอไฟต์ที่กลับไปอยู่ในน้ำก็ไม่มีผนังที่ทนทาน ดังนั้นจึงไม่มีเครื่องหมายไตรเลท[ 22 ]การตรวจสอบสปอร์ของสาหร่ายอย่างละเอียดแสดงให้เห็นว่าไม่มีสปอร์ใดที่มีไตรเลท ไม่ว่าจะเป็นเพราะผนังของสปอร์ไม่แข็งแรงพอ หรือในกรณีที่หายากที่สปอร์แข็งแรงพอ สปอร์ก็จะกระจายตัวออกไปก่อนที่จะถูกบีบอัดจนเพียงพอที่จะเกิดเครื่องหมาย หรือไม่ก็ไม่พอดีกับเตตระเฮดรัลเตตระด[ 22 ]
ซากดึกดำบรรพ์ขนาดใหญ่ที่เก่าแก่ที่สุดของพืชบกคือ สิ่งมีชีวิต แบบทัลลอยด์ซึ่งอาศัยอยู่ในพื้นที่ชุ่มน้ำริมแม่น้ำและพบว่าปกคลุมพื้นที่ราบน้ำท่วมถึงในยุคไซลูเรียนตอนต้น ส่วนใหญ่ พวกมันสามารถอยู่รอดได้ก็ต่อเมื่อพื้นดินมีน้ำขัง[ 23 ]นอกจากนี้ยังมีแผ่นจุลินทรีย์อีกด้วย[ 24 ]
เมื่อพืชขึ้นมาอยู่บนบกแล้ว มีสองแนวทางในการรับมือกับการขาดน้ำ พืชกลุ่มไบรโอไฟต์ในปัจจุบันอาจหลีกเลี่ยงหรือยอมจำนนต่อการขาดน้ำ โดยจำกัดถิ่นที่อยู่ให้อยู่ในสภาพแวดล้อมที่ชื้น หรืออาจแห้งเหี่ยวและหยุดกระบวนการเผาผลาญไว้ชั่วคราว จนกว่าจะมีน้ำมาเพิ่มเติม เช่น ในสกุลTargionia ซึ่งเป็นพืชกลุ่มลิเวอร์เวิ ร์ ต ส่วนพืชกลุ่มแทรคีโอไฟต์จะต้านทานการขาดน้ำโดยการควบคุมอัตราการสูญเสียน้ำ พวกมันมี ชั้น คิวติเคิล นอกที่กันน้ำได้ ในบริเวณที่สัมผัสกับอากาศ (เช่นเดียวกับไบรโอไฟต์บางชนิด) เพื่อลดการสูญเสียน้ำ แต่เนื่องจากการปกคลุมทั้งหมดจะทำให้พวกมันขาดคาร์บอนไดออกไซด์ในบรรยากาศ แทรคีโอไฟต์จึงใช้ช่องเปิดที่หลากหลาย คือปากใบเพื่อควบคุมอัตราการแลกเปลี่ยนก๊าซ นอกจากนี้ แทรคีโอไฟต์ยังพัฒนาเนื้อเยื่อท่อลำเลียงเพื่อช่วยในการเคลื่อนย้ายน้ำภายในสิ่งมีชีวิต (ดูด้านล่าง ) และเปลี่ยนจากวงจรชีวิตที่โดมินันต์ด้วยแกมีโทไฟต์ (ดูด้านล่าง ) เนื้อเยื่อท่อลำเลียงยังช่วยให้พืชเจริญเติบโตในแนวตั้งได้โดยไม่ต้องอาศัยน้ำ และปูทางไปสู่การวิวัฒนาการของพืชขนาดใหญ่บนบก
ผลที่ตามมา
เหตุการณ์การเกิดธารน้ำแข็งทั่วโลกที่เรียกว่าSnowball Earthซึ่งเกิดขึ้นในช่วงประมาณ 720-635 ล้านปีก่อนใน ยุค Cryogenianเชื่อกันว่าเกิดจากสิ่งมีชีวิตสังเคราะห์แสงในยุคแรกๆ อย่างน้อยบางส่วน ซึ่งช่วยลดความเข้มข้นของคาร์บอนไดออกไซด์และลดปรากฏการณ์เรือนกระจกในบรรยากาศ[ 25 ]ส่งผลให้เกิด สภาพภูมิ อากาศแบบยุคน้ำแข็งจาก การศึกษาโดยใช้ นาฬิกาโมเลกุลในช่วงทศวรรษที่ผ่านมา การศึกษาในปี 2022 พบว่าเวลาโดยประมาณสำหรับการกำเนิดของสเตรปโตไฟต์ แบบหลายเซลล์ (ทั้งหมด ยกเว้นกลุ่มพื้นฐานเซลล์เดียวMesostigmatophyceae ) ตกอยู่ในยุค Cryogenian ที่เย็น ในขณะที่การแยกตัวของสเตรปโตไฟต์ในภายหลังตกอยู่ใน ยุค Ediacaran ที่อบอุ่น ซึ่งพวกเขาตีความว่าเป็นข้อบ่งชี้ถึงแรงกดดันในการคัดเลือกโดยยุคน้ำแข็งต่อสิ่งมีชีวิตที่สังเคราะห์แสง ซึ่งกลุ่มหนึ่งประสบความสำเร็จในการอยู่รอดในสภาพแวดล้อมที่ค่อนข้างอบอุ่นซึ่งยังคงสามารถอาศัยอยู่ได้ และต่อมาเจริญเติบโตในยุค Ediacaran ตอนปลายและ ยุค Phanerozoicบนบกในฐานะเอ็มบริโอไฟต์ การศึกษายังตั้งทฤษฎีว่าสัณฐานวิทยาเซลล์เดียวและคุณลักษณะเฉพาะอื่นๆ ของZygnematophyceaeอาจสะท้อนถึงการปรับตัวเพิ่มเติมให้เข้ากับวิถีชีวิตที่ชอบความเย็น[ 26 ]การก่อตั้งพืชบนบกเพิ่มอัตราการสะสมของออกซิเจนในบรรยากาศ เนื่องจากพืชบนบกผลิตออกซิเจนเป็นของเสีย เมื่อความเข้มข้นนี้เพิ่มขึ้นเกิน 13% เมื่อราว 0.45 พันล้านปีก่อน[ 27 ]ไฟป่าจึงเกิดขึ้นได้ ซึ่งเห็นได้จากถ่านในบันทึกฟอสซิล[ 28 ]นอกเหนือจากช่องว่างที่เป็นข้อถกเถียงในช่วงปลายยุคดีโวเนียนแล้ว ถ่านก็มีอยู่เรื่อยมาตั้งแต่นั้นเป็นต้นมา
การกลายเป็นถ่านเป็นรูปแบบ การเกิดซากดึกดำบรรพ์ที่สำคัญอย่างหนึ่งไฟป่าหรือการฝังกลบในเถ้าภูเขาไฟร้อนจะขับไล่สารประกอบระเหยออกไป เหลือเพียงสารตกค้างที่เป็นคาร์บอนบริสุทธิ์ ซึ่งไม่สามารถเป็นแหล่งอาหารที่เหมาะสมสำหรับเชื้อรา สัตว์กินพืช หรือสัตว์กินซาก ดังนั้นจึงมีแนวโน้มที่จะได้รับการอนุรักษ์ไว้ นอกจากนี้ยังมีความแข็งแรงและสามารถทนต่อแรงกดดันได้ แสดงรายละเอียดที่ประณีต บางครั้งถึงระดับเซลล์ย่อยในซากดึกดำบรรพ์
นอกจากการปรากฏของถ่านในบันทึกหินแล้ว การปรับตัวของพืชบนบกยังมีส่วนสำคัญต่อการเปลี่ยนแปลงทางธรณีวิทยาและภูมิทัศน์ ยุคออร์โดวิเชียนและไซลูเรียนแสดงให้เห็นสัดส่วนของหินโคลนในบันทึกทางธรณีวิทยามากกว่า 1.4 เท่าเมื่อเทียบกับ 90% ก่อนหน้าของประวัติศาสตร์โลก และการเพิ่มขึ้นของหินโคลนนี้ถือเป็นผลมาจากการที่พืชบนบกกักเก็บโคลนไว้ในสภาพแวดล้อมบนบก[ 29 ]
วิวัฒนาการของวงจรชีวิต

พืชหลายเซลล์ทุกชนิดมีวงจรชีวิตประกอบด้วยสองรุ่นหรือสองระยะ ระยะแก มีโทไฟ ต์ เป็น ระยะแฮพลอยด์ที่มีโครโมโซมชุดเดียว (ใช้สัญลักษณ์ 1n ) และสร้างแกมีต (อสุจิและไข่) ระยะสปอโรไฟต์ เป็น ระยะดิ พลอยด์ ที่มีโครโมโซมเป็นคู่ (ใช้สัญลักษณ์ 2n ) และสร้างสปอร์ ระยะแกมีโทไฟต์และสปอโรไฟต์อาจมีรูปร่างเหมือนกัน คือมีลักษณะเหมือนกันในสาหร่ายบางชนิด เช่นUlva lactucaแต่จะแตกต่างกันมากในพืชบกสมัยใหม่ทั้งหมด ซึ่งเป็นภาวะที่เรียกว่าเฮเทอโรโมฟี
รูปแบบในวิวัฒนาการของพืชคือการเปลี่ยนจากโฮโมมอร์ฟีไปเป็นเฮเทอโรมอร์ฟี บรรพบุรุษของพืชบกที่เป็นสาหร่ายเกือบจะแน่นอนว่าเป็นแฮพลอยด์คือเป็นแฮพลอยด์ตลอดวงจรชีวิต โดยมีไซโกตเซลล์เดียวที่ให้ระยะ 2N พืชบกทั้งหมด (เช่นเอ็มบริโอไฟต์ ) เป็นดิพลอยด์ นั่นคือทั้งระยะแฮพลอยด์และดิพลอยด์เป็นหลายเซลล์[ 1 ]มีแนวโน้มสองประการที่เห็นได้ชัด: ไบรโอไฟต์ ( ลิเวอร์เวิร์ตมอสและฮอร์นเวิร์ต ) ได้พัฒนาแกมีโทไฟต์ให้เป็นระยะเด่นของวงจรชีวิต โดยสปอโรไฟต์แทบจะขึ้นอยู่กับแกมีโทไฟต์ทั้งหมดพืชมีท่อลำเลียงได้พัฒนาสปอโรไฟต์ให้เป็นระยะเด่น โดยแกมีโทไฟต์ลดลงอย่างมากในพืช มีเมล็ด
มีการเสนอว่าพื้นฐานสำหรับการเกิดขึ้นของระยะดิพลอยด์ของวงจรชีวิตเป็นระยะเด่นคือ ดิพลอยด์ช่วยให้สามารถปกปิดการแสดงออกของการกลายพันธุ์ที่เป็นอันตรายผ่านการเสริมยีน[ 30 ] [ 31 ]ดังนั้น หากจีโนมของพ่อแม่ตัวใดตัวหนึ่งในเซลล์ดิพลอยด์มีการกลายพันธุ์ที่นำไปสู่ข้อบกพร่องในผลิตภัณฑ์ยีน หนึ่งตัวหรือมากกว่านั้น ข้อบกพร่องเหล่านี้สามารถชดเชยได้ด้วยจีโนมของพ่อแม่อีกตัวหนึ่ง (ซึ่งอย่างไรก็ตามอาจมีข้อบกพร่องในยีนอื่นๆ ของตัวเอง) เมื่อระยะดิพลอยด์กลายเป็นระยะเด่น ผลกระทบของการปกปิดน่าจะทำให้ขนาดของจีโนมและด้วยเหตุนี้ปริมาณข้อมูล จึงเพิ่มขึ้นโดยไม่มีข้อจำกัดในการปรับปรุงความแม่นยำของการจำลองแบบ โอกาสในการเพิ่มปริมาณข้อมูลด้วยต้นทุนต่ำเป็นสิ่งที่เป็นประโยชน์เพราะช่วยให้สามารถเข้ารหัสการปรับตัวใหม่ๆ ได้ มุมมองนี้ถูกท้าทายด้วยหลักฐานที่แสดงให้เห็นว่าการคัดเลือกไม่มีประสิทธิภาพในระยะแฮพลอยด์มากกว่าในระยะดิพลอยด์ของวงจรชีวิตของมอสและพืชดอก[ 32 ]
มีทฤษฎีที่แข่งขันกันอยู่สองทฤษฎีเพื่ออธิบายการปรากฏของวงจรชีวิตแบบไดพลอบิออนติก
ทฤษฎีการแทรกสอด (หรือที่รู้จักกันในชื่อทฤษฎีแบบตรงข้ามหรือแบบแทรก) [ 33 ]ถือว่าการแทรกสอดของระยะสปอโรไฟต์แบบหลายเซลล์ระหว่างรุ่นแกมีโตไฟต์สองรุ่นที่ต่อเนื่องกันนั้นเป็นนวัตกรรมที่เกิดจากการแบ่งไมโอซิสก่อนหน้าในไซโกตที่งอกใหม่ซึ่งมีการแบ่งไมโทซิสหนึ่งรอบหรือมากกว่านั้น ทำให้เกิดเนื้อเยื่อหลายเซลล์แบบดิพลอยด์บางส่วนก่อนที่การแบ่งไมโอซิสจะสร้างสปอร์ในที่สุด ทฤษฎีนี้บ่งชี้ว่าสปอโรไฟต์แรกมีสัณฐานวิทยาที่แตกต่างและเรียบง่ายกว่าแกมีโตไฟต์ที่มันพึ่งพา[ 33 ]สิ่งนี้ดูเหมือนจะสอดคล้องกับสิ่งที่เรารู้เกี่ยวกับไบรโอไฟต์ ซึ่งแกมีโตไฟต์แบบทัลลอยด์ที่เจริญเติบโตจะหล่อเลี้ยงสปอโรไฟต์ที่เรียบง่าย ซึ่งประกอบด้วยเพียงแค่สปอแรนเจียมที่ไม่มีกิ่งก้านบนก้านเท่านั้น ความซับซ้อนที่เพิ่มขึ้นของสปอโรไฟต์ที่เรียบง่ายแต่ดั้งเดิม รวมถึงการได้รับเซลล์สังเคราะห์แสงในที่สุด จะทำให้มันเป็นอิสระจากการพึ่งพาแกมีโทไฟต์ ดังที่พบในฮอร์นเวิร์ตบางชนิด ( Anthoceros ) และในที่สุดจะส่งผลให้สปอโรไฟต์พัฒนาอวัยวะและเนื้อเยื่อหลอดเลือด และกลายเป็นระยะที่เด่นกว่า ดังเช่นในทราคีโอไฟต์ (พืชมีหลอดเลือด) [ 1 ]ทฤษฎีนี้อาจได้รับการสนับสนุนจากการสังเกตว่าCooksonia ที่มีขนาดเล็กกว่า จะต้องได้รับการสนับสนุนจากรุ่นแกมีโทไฟต์ การปรากฏของขนาดแกนที่ใหญ่ขึ้นที่สังเกตได้ ซึ่งมีพื้นที่สำหรับเนื้อเยื่อสังเคราะห์แสงและทำให้สามารถดำรงอยู่ได้ด้วยตนเอง จึงเป็นเส้นทางที่เป็นไปได้สำหรับการพัฒนาระยะสปอโรไฟต์ที่พึ่งพาตนเองได้[ 33 ]
สมมติฐานทางเลือกที่เรียกว่าทฤษฎีการเปลี่ยนแปลง (หรือทฤษฎีโฮโมโลกัส) ระบุว่าสปอโรไฟต์อาจปรากฏขึ้นอย่างฉับพลันโดยการชะลอการเกิดไมโอซิสจนกระทั่งสปอโรไฟต์หลายเซลล์ที่พัฒนาเต็มที่ได้ก่อตัวขึ้น เนื่องจากวัสดุพันธุกรรมเดียวกันจะถูกใช้โดยทั้งระยะแฮพลอยด์และดิพลอยด์ ดังนั้นพวกมันจึงดูเหมือนกัน นี่อธิบายพฤติกรรมของสาหร่ายบางชนิด เช่นUlva lactucaซึ่งสร้างระยะสลับกันของสปอโรไฟต์และแกมีโตไฟต์ที่เหมือนกัน การปรับตัวในภายหลังต่อสภาพแวดล้อมบนบกที่แห้งแล้งซึ่งทำให้การสืบพันธุ์แบบอาศัยเพศเป็นไปได้ยาก อาจส่งผลให้แกมีโตไฟต์ที่พร้อมสืบพันธุ์แบบอาศัยเพศมีความเรียบง่ายขึ้น และระยะสปอโรไฟต์มีความซับซ้อนมากขึ้นเพื่อกระจายสปอร์ที่กันน้ำได้ดีขึ้น[ 1 ]เนื้อเยื่อของสปอโรไฟต์และแกมีโตไฟต์ของพืชมีท่อลำเลียง เช่นRhyniaที่ได้รับการอนุรักษ์ไว้ในหินเชิร์ต Rhynieมีความซับซ้อนคล้ายกัน ซึ่งถือเป็นการสนับสนุนสมมติฐานนี้[ 33 ] [ 34 ] [ 35 ]ในทางตรงกันข้าม พืชหลอดเลือดสมัยใหม่ ยกเว้นPsilotumมีสปอโรไฟต์และแกมีโตไฟต์ที่มีรูปร่างต่างกัน โดยที่แกมีโตไฟต์แทบจะไม่มีเนื้อเยื่อหลอดเลือดเลย[ 36 ]
วิวัฒนาการของกายวิภาคของพืช
ความสัมพันธ์แบบพึ่งพาอาศัยกันของไมคอร์ไรซาแบบอาร์บัสคูลาร์
ไม่มีหลักฐานว่าพืชบกยุคแรกในยุคไซลูเรียนและเดโวเนียนตอนต้นมีราก แม้ว่าจะมีหลักฐานฟอสซิลของไรโซอยด์ในหลายชนิด เช่นHorneophyton ก็ตาม พืชบกยุคแรกสุดไม่มีระบบหลอดเลือดสำหรับการขนส่งน้ำและสารอาหารเช่นกันAglaophytonซึ่งเป็นพืชมีหลอดเลือดที่ไม่มีรากที่รู้จักจากฟอสซิลเดโวเนียนในหินเชิร์ต Rhynie [ 37 ]เป็นพืชบกชนิดแรกที่ถูกค้นพบว่ามี ความสัมพันธ์ แบบพึ่งพาอาศัยกันกับเชื้อรา[ 38 ]ซึ่งก่อตัวเป็นไมคอร์ไรซาแบบอาร์บัสคูลาร์ หรือ "รากเชื้อราคล้ายต้นไม้" ในเซลล์ทรงกระบอกที่กำหนดไว้อย่างชัดเจน (วงแหวนในภาคตัดขวาง) ในคอร์เทกซ์ของลำต้น เชื้อรากินน้ำตาลของพืช แลกกับสารอาหารที่สร้างขึ้นหรือสกัดจากดิน (โดยเฉพาะฟอสเฟต ) ซึ่งพืชจะไม่สามารถเข้าถึงได้หากไม่มีเชื้อรา เช่นเดียวกับพืชบกไร้รากอื่นๆ ในยุคไซลูเรียนและยุคดีโวเนียนตอนต้นAglaophytonอาจอาศัยเชื้อราไมคอร์ไรซาแบบอาร์บัสคูลาร์ในการดูดซับน้ำและสารอาหารจากดิน[ 39 ]
เชื้อราเหล่านี้อยู่ในไฟลัมGlomeromycota [ 40 ] ซึ่งเป็นกลุ่มที่น่าจะปรากฏตัวครั้งแรก เมื่อ 1 พันล้านปีก่อน และยังคงสร้างความสัมพันธ์แบบไมคอร์ไรซาแบบอาร์บัสคูลาร์กับกลุ่มพืชบกหลักๆ ทุกกลุ่มตั้งแต่ไบรโอไฟต์ไปจนถึงเทอริโดไฟต์ จิมโนสเปิร์ม และแองจิโอสเปิร์ม และกับพืชมีท่อลำเลียงมากกว่า 80% [ 41 ]
หลักฐานจากการวิเคราะห์ลำดับดีเอ็นเอแสดงให้เห็นว่าความสัมพันธ์แบบพึ่งพาอาศัยกันของไมคอร์ไรซาแบบอาร์บัสคูลาร์เกิดขึ้นในบรรพบุรุษร่วมของกลุ่มพืชบกเหล่านี้ในช่วงการเปลี่ยนผ่านไปสู่บนบก[ 42 ]และอาจเป็นขั้นตอนสำคัญที่ทำให้พวกมันสามารถตั้งรกรากบนบกได้[ 43 ]เชื้อราไมคอร์ไรซาที่ปรากฏขึ้นก่อนที่พืชเหล่านี้จะวิวัฒนาการราก จะช่วยให้พืชได้รับน้ำและแร่ธาตุ เช่นฟอสฟอรัสแลกกับสารประกอบอินทรีย์ที่พวกมันไม่สามารถสังเคราะห์เองได้[ 41 ]เชื้อราดังกล่าวช่วยเพิ่มผลผลิตแม้แต่พืชที่เรียบง่าย เช่น ลิเวอร์เวิร์ต[ 44 ] [ 45 ]
คิวติเคิล ปากใบ และช่องว่างระหว่างเซลล์
ในการสังเคราะห์แสง พืชต้องดูดซับ CO2 จากบรรยากาศ อย่างไรก็ตาม การทำให้เนื้อเยื่อพร้อมสำหรับการดูดซับ CO2 จะทำให้น้ำระเหย ซึ่งต้องแลกมาด้วยต้นทุน[ 46 ]น้ำจะสูญเสียไปเร็วกว่าการดูดซับ CO2 มากดังนั้นพืชจึงจำเป็นต้องเติมน้ำเข้าไปใหม่ พืชบกในยุคแรกๆ ขนส่งน้ำผ่านช่องว่างภายในผนังเซลล์ที่มีรูพรุน ต่อมาพวกมันได้พัฒนาลักษณะทางกายวิภาคสามประการที่ช่วยให้สามารถควบคุมการสูญเสียน้ำที่หลีกเลี่ยงไม่ได้ซึ่งเกิดขึ้นพร้อมกับการดูดซับ CO2 ประการแรก เปลือกนอกที่กันน้ำหรือคิวติเคิลได้พัฒนาขึ้นเพื่อลดการสูญเสียน้ำ ประการที่สอง ช่องเปิดที่เปลี่ยนแปลงได้ หรือ ปากใบที่สามารถเปิดและปิดเพื่อควบคุมปริมาณน้ำที่สูญเสียไปจากการระเหยในระหว่างการดูดซับ CO2 และประการที่สาม ช่องว่างระหว่างเซลล์พาเรนไคมาที่สังเคราะห์แสงได้ ซึ่งช่วยให้การกระจาย CO2 ภายในไปยังคลอโรพลาสต์ดีขึ้นระบบสามส่วนนี้ช่วยให้เกิดโฮโมไฮดรีที่ดีขึ้น ซึ่งเป็นการควบคุมปริมาณน้ำในเนื้อเยื่อ โดยเฉพาะอย่างยิ่งเมื่อปริมาณน้ำไม่คงที่[ 47 ] ความเข้มข้นของ CO2ที่สูงในยุคไซลูเรียนและเดโวเนียนตอนต้น ซึ่งเป็นช่วงที่พืชเริ่มเข้ามาอาศัยบนบกเป็นครั้งแรก หมายความว่าพืชใช้น้ำได้อย่างมีประสิทธิภาพมากขึ้น เมื่อพืชดูดซับ CO2 จากชั้นบรรยากาศ น้ำก็จะสูญเสียไปมากขึ้นในกระบวนการดูดซับ และกลไกการดูดซับและขนส่งน้ำที่ซับซ้อนยิ่งขึ้นจึงวิวัฒนาการขึ้น[ 46 ]พืชที่เติบโตขึ้นไปในอากาศจำเป็นต้องมีระบบในการขนส่งน้ำจากดินไปยังส่วนต่างๆ ของพืชที่อยู่เหนือดิน โดยเฉพาะอย่างยิ่งส่วนที่สังเคราะห์แสง เมื่อสิ้นสุดยุคคาร์บอนิเฟอรัสเมื่อความเข้มข้นของ CO2 ลดลงจนใกล้เคียงกับปัจจุบัน น้ำจะสูญเสียไปประมาณ 17 เท่าต่อหน่วยการดูดซับCO2 [ 46 ]อย่างไรก็ตาม แม้ในยุคแรกๆ ที่ "ง่าย" น้ำก็เป็นสิ่งที่มีค่าและต้องถูกขนส่งไปยังส่วนต่างๆ ของพืชจากดินที่เปียกชื้นเพื่อหลีกเลี่ยงการแห้งเหี่ยว[ 47 ]
น้ำสามารถซึมผ่านเนื้อผ้าที่มีช่องว่างเล็กๆ ได้ด้วยแรงดึงดูดของเส้นเลือดฝอยในคอลัมน์น้ำแคบๆ เช่น ภายในผนังเซลล์พืชหรือในเทรคีด เมื่อโมเลกุลระเหยจากปลายด้านหนึ่ง โมเลกุลเหล่านั้นจะดึงโมเลกุลที่อยู่ด้านหลังไปตามช่องทาง ดังนั้น การระเหยเพียงอย่างเดียวจึงเป็นแรงขับเคลื่อนสำหรับการขนส่งน้ำในพืช[ 46 ] อย่างไรก็ตาม หากไม่มีท่อลำเลียงเฉพาะ กลไก แรงดึงและแรงยึดเหนี่ยว นี้ อาจทำให้เกิดแรงดันลบมากพอที่จะทำให้เซลล์นำน้ำยุบตัวลง จำกัดการขนส่งน้ำได้ไม่เกินสองสามเซนติเมตร และด้วยเหตุนี้จึงจำกัดขนาดของพืชในยุคแรกๆ[ 46 ]

ไซเล็ม
เพื่อให้หลุดพ้นจากข้อจำกัดของขนาดเล็กและความชื้นคงที่ที่ระบบการขนส่งพาเรนไคมาติก สร้างขึ้น พืชจึงต้องการระบบการขนส่งน้ำที่มีประสิทธิภาพมากขึ้น เมื่อพืชเจริญเติบโตขึ้น เนื้อเยื่อหลอดเลือด สำหรับการขนส่งน้ำที่เฉพาะเจาะจง จึงวิวัฒนาการขึ้น โดยเริ่มแรกในรูปแบบของไฮดรอยด์แบบง่ายๆ ที่พบในขนของสปอโรไฟต์มอส เซลล์ยาวแบบง่ายๆ เหล่านี้จะตายและเต็มไปด้วยน้ำเมื่อเจริญเต็มที่ ทำให้เกิดช่องทางสำหรับการขนส่งน้ำ แต่ผนังที่บางและไม่เสริมแรงจะยุบตัวลงภายใต้แรงตึงของน้ำเพียงเล็กน้อย ซึ่งจำกัดความสูงของพืชเซลล์ไซเล็มเทรคีดซึ่งเป็นเซลล์ที่กว้างกว่าและมี ผนังเซลล์เสริมแรงด้วย ลิกนินซึ่งทนต่อการยุบตัวภายใต้แรงตึงที่เกิดจากความเครียดจากน้ำได้ดีกว่า พบในกลุ่มพืชมากกว่าหนึ่งกลุ่มในช่วงกลางยุคไซลูเรียน และอาจมีต้นกำเนิดทางวิวัฒนาการเดียว อาจอยู่ในกลุ่มฮอร์นเวิร์ต[ 48 ]ซึ่งรวมเทรคีโอไฟต์ทั้งหมดเข้าด้วยกัน หรืออีกทางหนึ่ง พวกมันอาจวิวัฒนาการขึ้นมากกว่าหนึ่งครั้ง[ 46 ] ต่อมาในยุคครีเทเชียส เทรคีดส์ถูกพัฒนาเป็นเวสิเคิลในพืชดอก [ 46 ] เมื่อกลไกการลำเลียงน้ำและคิวติเคิลกันน้ำพัฒนาขึ้น พืชจึงสามารถอยู่รอดได้โดยไม่ต้องมีฟิล์มน้ำปกคลุมอยู่ตลอดเวลา การเปลี่ยนแปลงจากpoikilohydryไปเป็นhomoiohydry นี้ เปิดโอกาสใหม่สำหรับการตั้งถิ่นฐาน[ 46 ] [ 47 ]
AglaophytonและHorneophytonซึ่งเป็นพืชก่อนมีท่อลำเลียงน้ำในยุคดีโวเนียนตอนต้นมีท่อลำเลียงน้ำที่ไม่เสริมแรง โดยมีโครงสร้างผนังที่คล้ายกับไฮดรอยด์ของมอสมาก แต่พวกมันเติบโตควบคู่ไปกับพืช มีท่อ ลำเลียงน้ำ หลายชนิด เช่นRhynia gwynne-vaughaniiซึ่งมีท่อลำเลียงน้ำไซเล็มที่เสริมแรงด้วยแถบลิกนินอย่างดี ฟอสซิลขนาดใหญ่ที่เก่าแก่ที่สุดที่ทราบว่ามีท่อลำเลียงน้ำไซเล็มคือพืชขนาดเล็กในยุคกลางไซลูเรียนของสกุลCooksonia [ 49 ] อย่างไรก็ตามแถบที่หนาขึ้นบนผนังของชิ้นส่วนท่อที่แยกออกมานั้นปรากฏให้เห็นตั้งแต่ยุคไซลูเรียนตอนต้นเป็นต้นไป[ 50 ]
พืชยังคงคิดค้นวิธีการลดความต้านทานต่อการไหลภายในเซลล์อย่างต่อเนื่อง เพิ่มประสิทธิภาพในการขนส่งน้ำ และเพิ่มความต้านทานของทราคีดต่อการยุบตัวภายใต้แรงดึง[ 51 ] [ 52 ]ในช่วงต้นยุคดีโวเนียน เส้นผ่านศูนย์กลางทราคีดสูงสุดเพิ่มขึ้นตามเวลา แต่อาจถึงจุดคงที่ในโซสเตอโรฟิลล์ในช่วงกลางยุคดี โวเนียน [ 51 ] อัตราการขนส่งโดยรวมยังขึ้นอยู่กับพื้นที่หน้าตัดโดยรวมของกลุ่มไซเล็มเอง และพืชบางชนิดในช่วงกลางยุคดีโวเนียน เช่น ไตรเมอโรไฟต์ มีสเตลที่ใหญ่กว่าบรรพบุรุษในยุคแรกมาก[ 51 ]แม้ว่าทราคีดที่กว้างขึ้นจะให้อัตราการขนส่งน้ำที่สูงขึ้น แต่ก็เพิ่มความเสี่ยงต่อการเกิดโพรงอากาศ ซึ่งเป็นการก่อตัวของฟองอากาศที่เกิดจากการแตกของคอลัมน์น้ำภายใต้แรงดึง[ 46 ]รูเล็กๆ ในผนังทราคีดช่วยให้น้ำไหลผ่านทราคีดที่บกพร่องได้ ในขณะเดียวกันก็ป้องกันไม่ให้ฟองอากาศผ่านเข้าไปได้[ 46 ]แต่ต้องแลกมาด้วยอัตราการไหลที่จำกัด ในยุคคาร์บอนิเฟอรัส พืชเมล็ดเปลือยได้พัฒนารูที่มีขอบ [ 53 ] [ 54 ] โครงสร้างคล้ายวาล์วที่ช่วยให้รูที่มีการนำไฟฟ้าสูงสามารถปิดผนึก ได้เมื่อด้านหนึ่งของทราคีดมีความดันลดลง
ทราคีดมีผนังปลายที่ไม่เป็นรูพรุนและมีหลุม ซึ่งทำให้เกิดแรงต้านทานต่อการไหลของน้ำอย่างมาก[ 51 ]แต่ก็อาจมีข้อดีในการแยกฟองอากาศที่เกิดจากการเกิดโพรงอากาศหรือการแข็งตัว ท่อลำเลียงน้ำวิวัฒนาการขึ้นครั้งแรกในช่วงยุคเพอร์เมียนตอนปลายที่แห้งแล้งและมี CO2 ต่ำในหญ้าหางม้า เฟิร์น และเซลาจิเนลลาเลสอย่างอิสระ และต่อมาปรากฏขึ้นในยุคครีเทเชียสตอนกลางในพืชกลุ่มเนโทไฟต์และพืชดอก[ 46 ] สมาชิกของท่อลำเลียงน้ำเป็นท่อเปิดที่ไม่มีผนังปลาย และเรียงต่อกันเป็นท่อเดียวต่อเนื่องกันเพื่อทำหน้าที่เสมือนเป็นท่อลำเลียงน้ำเดียว[ 51 ]ท่อลำเลียงน้ำช่วยให้ไม้ที่มีพื้นที่หน้าตัดเท่ากันสามารถลำเลียงน้ำได้มากกว่าทราคีดมาก[ 46 ]สิ่งนี้ทำให้พืชสามารถเติมเส้นใย โครงสร้างลงในลำต้นได้มากขึ้น และยังเปิดช่องทางใหม่ให้กับไม้เลื้อย ซึ่งสามารถลำเลียงน้ำได้โดยไม่ต้องหนาเท่ากับต้นไม้ที่มันเติบโตอยู่[ 46 ]แม้จะมีข้อดีเหล่านี้ แต่เนื้อไม้ที่สร้างจากทราคีดนั้นเบากว่ามาก จึงผลิตได้ถูกกว่า เนื่องจากท่อลำเลียงน้ำต้องได้รับการเสริมแรงมากขึ้นเพื่อหลีกเลี่ยงการเกิดโพรงอากาศ[ 46 ]เมื่อพืชวิวัฒนาการจนสามารถควบคุมการระเหยของน้ำและการลำเลียงน้ำได้ในระดับนี้ พวกมันจึง กลายเป็นพืชที่อาศัยน้ำ ได้ เองอย่างแท้จริง สามารถดึงน้ำจากสิ่งแวดล้อมผ่านอวัยวะคล้ายรากแทนที่จะพึ่งพาความชื้นบนผิว ทำให้พวกมันสามารถเติบโตได้ใหญ่ขึ้นมาก[ 47 ] [ 46 ]แต่เนื่องจากความเป็นอิสระจากสิ่งแวดล้อมมากขึ้น พืชมีท่อลำเลียงส่วนใหญ่จึงสูญเสียความสามารถในการอยู่รอดจากการขาดน้ำ ซึ่งเป็นลักษณะที่สูญเสียไปอย่างน่าเสียดาย[ 46 ]ในพืชบกยุคแรก การรองรับส่วนใหญ่เกิดจากแรงดันเต่ง โดยเฉพาะอย่างยิ่งชั้นนอกของเซลล์ที่เรียกว่าสเตอโรมทราคีด และไม่ใช่จากไซเล็ม ซึ่งมีขนาดเล็กเกินไป อ่อนแอเกินไป และอยู่ในตำแหน่งตรงกลางมากเกินไปที่จะให้การสนับสนุนโครงสร้างได้มากนัก[ 46 ] พืชที่มีไซเล็มรองซึ่งปรากฏขึ้นในช่วงกลางยุคดีโวเนียน เช่นไตรเมอโรไฟต์และโปรจิมโนสเปิร์มมีหน้าตัดของหลอดเลือดที่ใหญ่กว่ามาก ทำให้เกิดเนื้อเยื่อไม้ที่แข็งแรง
เอนโดเดอร์มิส
เอนโดเดอร์มิสอาจวิวัฒนาการในรากพืชยุคแรกในช่วงยุคดีโวเนียน แต่หลักฐานฟอสซิลแรกสุดสำหรับโครงสร้างดังกล่าวคือยุคคาร์บอนิเฟอรัส[ 46 ]เอนโดเดอร์มิสในรากจะล้อมรอบเนื้อเยื่อขนส่งน้ำและควบคุมการแลกเปลี่ยนไอออนระหว่างน้ำใต้ดินกับเนื้อเยื่อ และป้องกันเชื้อโรคที่ไม่พึงประสงค์ ฯลฯ ไม่ให้เข้าสู่ระบบขนส่งน้ำ เอนโดเดอร์มิสยังสามารถสร้างแรงดันขึ้นด้านบน บังคับให้น้ำไหลออกจากรากเมื่อการคายน้ำไม่เพียงพอที่จะเป็นตัวขับเคลื่อน
วิวัฒนาการของสัณฐานวิทยาของพืช
ออกจาก
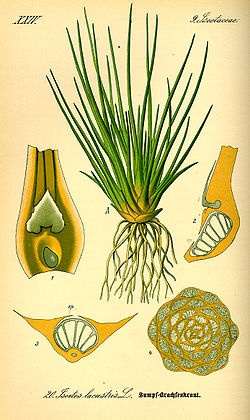


ใบเป็นอวัยวะ สังเคราะห์แสงหลักของพืชสมัยใหม่ การกำเนิดของใบเกือบจะแน่นอนว่าเกิดขึ้นจากการลดลงของความเข้มข้นของ CO2 ในบรรยากาศในช่วงยุคดีโวเนียน ซึ่งเพิ่มประสิทธิภาพในการดักจับคาร์บอนไดออกไซด์เพื่อการสังเคราะห์แสง[ 55 ] [ 56 ]
ใบไม้มีการวิวัฒนาการมากกว่าหนึ่งครั้ง โดยพิจารณาจากโครงสร้าง ใบไม้ถูกจัดประเภทเป็นสองประเภท ได้แก่ไมโครฟิลล์ซึ่งไม่มีเส้นใบที่ซับซ้อนและอาจมีต้นกำเนิดมาจากส่วนที่ยื่นออกมาคล้ายหนามที่เรียกว่าเอเนชัน และเม กาฟิลล์ ซึ่งมีขนาดใหญ่และมีเส้นใบที่ ซับซ้อน ซึ่งอาจเกิดขึ้นจากการดัดแปลงกลุ่มกิ่งก้าน มีการเสนอว่าโครงสร้างเหล่านี้เกิดขึ้นอย่างอิสระ[ 57 ]ตามทฤษฎีเทโลมของวอลเตอร์ ซิมเมอร์แมน[ 58 ] เมกาฟิลล์ ได้วิวัฒนาการมาจากพืชที่แสดงสถาปัตยกรรมแบบกิ่งก้านสามมิติ ผ่านการเปลี่ยนแปลงสาม ขั้นตอน ได้แก่ การทับซ้อนซึ่งนำไปสู่ตำแหน่งด้านข้างที่เป็นลักษณะเฉพาะของใบไม้การแบนราบซึ่งเกี่ยวข้องกับการสร้างสถาปัตยกรรมแบบแบนราบการเชื่อมต่อหรือการหลอมรวมซึ่งรวมกิ่งก้านแบบแบนราบเข้าด้วยกัน จึงนำไปสู่การสร้าง แผ่น ใบ ที่เหมาะสม ขั้นตอนทั้งสามนี้เกิดขึ้นหลายครั้งในการวิวัฒนาการของใบไม้ในปัจจุบัน[ 59 ]
It is widely believed that the telome theory is well supported by fossil evidence. However, Wolfgang Hagemann questioned it for morphological and ecological reasons and proposed an alternative theory.[60][61] Whereas according to the telome theory the most primitive land plants have a three-dimensional branching system of radially symmetrical axes (telomes), according to Hagemann's alternative the opposite is proposed: the most primitive land plants that gave rise to vascular plants were flat, thalloid, leaf-like, without axes, somewhat like a liverwort or fern prothallus. Axes such as stems and roots evolved later as new organs. Rolf Sattler proposed an overarching process-oriented view that leaves some limited room for both the telome theory and Hagemann's alternative and in addition takes into consideration the whole continuum between dorsiventral (flat) and radial (cylindrical) structures that can be found in fossil and living land plants.[62][63] This view is supported by research in molecular genetics. Thus, James (2009)[64] concluded that "it is now widely accepted that... radiality [characteristic of axes such as stems] and dorsiventrality [characteristic of leaves] are but extremes of a continuous spectrum. In fact, it is simply the timing of the KNOX gene expression".
Before the evolution of leaves, plants had the photosynthetic apparatus on the stems, which they retain albeit leaves have largely assumed that job. Today's megaphyll leaves probably became commonplace some 360mya, about 40my after the simple leafless plants had colonized the land in the Early Devonian. This spread has been linked to the fall in the atmospheric carbon dioxide concentrations in the Late Paleozoic era associated with a rise in density of stomata on leaf surface.[55] This would have resulted in greater transpiration rates and gas exchange, but especially at high CO2 concentrations, large leaves with fewer stomata would have heated to lethal temperatures in full sunlight. Increasing the stomatal density allowed for a better-cooled leaf, thus making its spread feasible, but increased CO2 uptake at the expense of decreased water use efficiency.[56][65]
ไรนิโอไฟต์ของหินเชิร์ตไรนีประกอบด้วยแกนที่เรียวบางและไม่มีลวดลายเท่านั้นไตรเมอโรไฟต์ ในยุคดีโวเนียนตอนต้นถึงตอนกลาง อาจถือได้ว่ามีใบ พืชมีท่อลำเลียงกลุ่มนี้สามารถจำแนกได้จากกลุ่มของสปอแรนเจียที่ปลาย ซึ่งประดับอยู่ที่ปลายแกนที่อาจแตกออกเป็นสองหรือสามแฉก[ 1 ]สิ่งมีชีวิตบางชนิด เช่นPsilophyton มีส่วนยื่น ที่เรียกว่าenationsซึ่งเป็นส่วนที่ยื่นออกมาจากลำต้นขนาดเล็กและมีหนาม ไม่มีท่อลำเลียงของตัวเอง
โซสเตอโรฟิลล์มีความสำคัญอยู่แล้วในช่วงปลายยุคไซลูเรียน ซึ่งเร็วกว่าไรนิโอไฟต์ที่มีความซับซ้อนเทียบเท่ากันมาก[ 66 ]กลุ่มนี้สามารถจำแนกได้จากสปอแรนเจียรูปไตที่เติบโตบนกิ่งด้านข้างสั้นๆ ใกล้กับแกนหลัก บางครั้งแตกแขนงเป็นรูปตัว H ที่โดดเด่น[ 1 ]โซสเตอโรฟิลล์จำนวนมากมีเอ็นเนชัน (เนื้อเยื่อที่งอกออกมาเล็กๆ บนพื้นผิวที่มีรูปร่างหลากหลาย) บนแกน แต่ไม่มีเอ็นเนชันใดที่มีร่องรอยหลอดเลือด หลักฐานแรกของเอ็นเนชันที่มีหลอดเลือดเกิดขึ้นในฟอสซิลคลับมอสที่รู้จักกันในชื่อBaragwanathiaซึ่งปรากฏในบันทึกฟอสซิลแล้วในช่วงปลายยุคไซลูเรียน[ 67 ]ในสิ่งมีชีวิตนี้ ร่องรอยใบเหล่านี้ต่อเนื่องเข้าไปในใบเพื่อสร้างเส้นกลางใบ[ 68 ]ทฤษฎีหนึ่งที่เรียกว่า "ทฤษฎีเอเนชั่น" ระบุว่าใบไมโครฟิลล์ของคลับมอสส์พัฒนาโดยการเจริญเติบโตของโปรโตสเตลที่เชื่อมต่อกับเอเนชั่นที่มีอยู่[ 1 ]ใบของสกุลAsteroxylon จาก Rhynie ซึ่งถูกเก็บรักษาไว้ในหินเชิร์ต Rhynie เกือบ 20 ล้านปีต่อมาหลังจากBaragwanathiaมีระบบหลอดเลือดแบบดั้งเดิม – ในรูปแบบของร่องรอยใบที่แยกออกจากโปรโตสเตลกลางไปยัง "ใบ" แต่ละใบ[ 69 ] AsteroxylonและBaragwanathiaถือกันอย่างกว้างขวางว่าเป็นไลโคพอดดั้งเดิม[ 1 ]ซึ่งเป็นกลุ่มที่ยังคงมีอยู่จนถึงปัจจุบัน โดยมีตัวแทนคือควิลล์เวิร์ต สไปค์มอสส์และคลับมอสส์ไลโคพอดมีไมโครฟิลล์ ที่โดดเด่น ซึ่งกำหนดไว้ว่าเป็นใบที่มีร่องรอยหลอดเลือดเพียงเส้นเดียว ใบขนาดเล็กอาจเจริญเติบโตได้จนมีขนาดพอสมควร โดยใบของพืชในอันดับ Lepidodendralesอาจยาวได้มากกว่าหนึ่งเมตร แต่เกือบทั้งหมดจะมีเพียงมัดท่อลำเลียงเพียงมัดเดียว ยกเว้นการแตกแขนงที่พบได้น้อยในพืช สกุล Selaginella บางชนิด
ใบขนาดใหญ่ ที่คุ้นเคยกันดี นั้น เชื่อกันว่าเกิดขึ้นอย่างอิสระถึงสี่ครั้ง ได้แก่ ในเฟิร์น หญ้าหางม้า โปรจิมโนสเปิร์ม และพืชมีเมล็ด[ 70 ]ดูเหมือนว่าใบขนาดใหญ่เหล่านี้จะเกิดขึ้นจากการดัดแปลง กิ่ง ที่แตกออกเป็นสองแฉก ซึ่งในตอนแรกจะซ้อนทับกัน (หรือ "ทับซ้อน") กลายเป็นแบนราบหรือเรียบ และในที่สุดก็พัฒนาเป็น "ใย" และวิวัฒนาการไปทีละน้อยจนกลายเป็นโครงสร้างที่คล้ายใบมากขึ้น[ 68 ]ตามทฤษฎีเทโลมของซิมเมอร์แมน ใบขนาดใหญ่เหล่านี้ ประกอบด้วยกลุ่มของกิ่งที่เชื่อมต่อกันเป็นใย[ 68 ]ดังนั้น "ช่องว่างของใบ" ที่เหลืออยู่ตรงที่มัดหลอดเลือดของใบแยกออกจากมัดหลอดเลือดของกิ่งหลักจึงดูเหมือนแกนสองแกนที่แยกออกจากกัน[ 68 ]ในแต่ละกลุ่มทั้งสี่กลุ่มที่วิวัฒนาการใบขนาดใหญ่ ใบของพวกมันวิวัฒนาการขึ้นครั้งแรกในช่วงปลายยุคดีโวเนียนถึงต้นยุคคาร์บอนิเฟอรัส โดยมีความหลากหลายอย่างรวดเร็วจนกระทั่งรูปแบบต่างๆ คงที่ในช่วงกลางยุคคาร์บอนิเฟอรัส[ 70 ]
การหยุดชะงักของการกระจายตัวเพิ่มเติมอาจเกิดจากข้อจำกัดในการพัฒนา[ 70 ]ซึ่งทำให้เกิดคำถามว่าทำไมใบไม้จึงใช้เวลานานมากในการวิวัฒนาการตั้งแต่แรก พืชอยู่บนบกมาอย่างน้อย 50 ล้านปีก่อนที่เมกะฟิลล์จะมีความสำคัญ อย่างไรก็ตาม มีเมโซฟิลล์ขนาดเล็กและหายากที่พบในสกุลEophyllophyton ในยุคดีโวเนียนตอนต้น ดังนั้นการพัฒนาจึงไม่น่าจะเป็นอุปสรรคต่อการปรากฏตัวของพวกมัน[ 71 ]คำอธิบายที่ดีที่สุดในขณะนี้คือ CO2 ในบรรยากาศลดลงอย่างรวดเร็วในช่วงเวลานี้ โดยลดลงประมาณ 90% ในช่วงยุคดีโวเนียน[ 72 ]ซึ่งจำเป็นต้องเพิ่มความหนาแน่นของปากใบขึ้น 100 เท่าเพื่อรักษาระดับการสังเคราะห์แสง เมื่อปากใบเปิดเพื่อให้น้ำระเหยออกจากใบ จะมีผลทำให้เกิดความเย็น ซึ่งเป็นผลมาจากการสูญเสียความร้อนแฝงของการระเหย ดูเหมือนว่าความหนาแน่นของปากใบที่ต่ำในช่วงต้นยุคดีโวเนียนหมายความว่าการระเหยและการระบายความร้อนด้วยการระเหยมีจำกัด และใบไม้จะร้อนเกินไปหากเจริญเติบโตจนมีขนาดใหญ่ ความหนาแน่นของปากใบไม่สามารถเพิ่มขึ้นได้ เนื่องจากเนื้อเยื่อท่อลำเลียงน้ำแบบดั้งเดิมและระบบรากที่จำกัดจะไม่สามารถส่งน้ำได้เร็วพอที่จะตรงกับอัตราการคายน้ำ[ 56 ] เห็นได้ชัดว่าใบไม้ไม่ได้มีประโยชน์เสมอไป ดังที่แสดง ให้ เห็นโดยการสูญเสียใบรองที่เกิดขึ้นบ่อยครั้ง ตัวอย่างเช่นต้นกระบองเพชรและเฟิร์นแส้Psilotum
วิวัฒนาการขั้นที่สองสามารถปกปิดต้นกำเนิดวิวัฒนาการที่แท้จริงของใบบางชนิดได้ เฟิร์นบางสกุลมีใบที่ซับซ้อนซึ่งติดอยู่กับแกนเทียมโดยการเจริญเติบโตของมัดหลอดเลือด ทำให้ไม่มีช่องว่างระหว่างใบ[ 68 ]
ต้นไม้ ผลัดใบต้องเผชิญกับข้อเสียอีกประการหนึ่งของการมีใบ ความเชื่อที่แพร่หลายว่าพืชจะผลัดใบเมื่อวันสั้นลงนั้นไม่ถูกต้อง เพราะต้นไม้ไม่ผลัดใบเจริญเติบโตได้ดีในวงกลมอาร์กติกในช่วง ยุค เรือนกระจกครั้งล่าสุด[ 73 ]เหตุผลที่ยอมรับกันโดยทั่วไปสำหรับการผลัดใบในช่วงฤดูหนาวคือเพื่อรับมือกับสภาพอากาศ – แรงลมและน้ำหนักของหิมะจะรับมือได้ดีกว่ามากหากไม่มีใบเพื่อเพิ่มพื้นที่ผิว การผลัดใบตามฤดูกาลได้วิวัฒนาการขึ้นอย่างอิสระหลายครั้งและพบได้ในกิงโกเลสพินอไฟตาบางชนิดและพืชดอกบางชนิด[ 74 ]การผลัดใบอาจเกิดขึ้นเพื่อตอบสนองต่อแรงกดดันจากแมลง อาจมีต้นทุนน้อยกว่าที่จะสูญเสียใบทั้งหมดในช่วงฤดูหนาวหรือฤดูแล้งมากกว่าที่จะลงทุนทรัพยากรในการซ่อมแซมต่อไป[ 75 ]
ราก


วิวัฒนาการของรากมีผลกระทบในระดับโลก โดยการรบกวนดินและส่งเสริมความเป็นกรดของดิน (โดยการดูดซับสารอาหาร เช่น ไนเตรตและฟอสเฟต[ 76 ] ) ทำให้ดินผุพังได้ลึกขึ้น ส่งผลให้สารประกอบคาร์บอนแทรกซึมลงไปในดินลึกขึ้น[ 77 ]ซึ่งส่งผลกระทบอย่างมากต่อสภาพภูมิอากาศ[ 78 ]ผลกระทบเหล่านี้อาจรุนแรงมากจนนำไปสู่การสูญพันธุ์ครั้งใหญ่[ 79 ]
แม้ว่าจะมีร่องรอยคล้ายรากในดินฟอสซิลในช่วงปลายยุคไซลูเรียน[ 80 ]แต่ฟอสซิลร่างกายแสดงให้เห็นว่าพืชยุคแรกสุดไม่มีราก หลายชนิดมีกิ่งก้านแผ่ราบไปตามพื้นดิน โดยมีแกนหรือทัลลัส ตั้งตรง กระจายอยู่บ้าง และบางชนิดยังมีกิ่งใต้ดินที่ไม่สังเคราะห์แสงซึ่งไม่มีปากใบ รากมีหมวกราก ซึ่ง แตกต่างจากกิ่งก้านที่เฉพาะเจาะจง[ 7 ]ดังนั้นในขณะที่พืชยุคไซลูเรียน-เดโวเนียน เช่นRhyniaและHorneophytonมีสิ่งที่เทียบเท่ากับรากในทางสรีรวิทยา[ 81 ] [ 82 ]ราก – ซึ่งนิยามว่าเป็นอวัยวะที่แยกออกจากลำต้น – ก็ไม่ได้ปรากฏขึ้นจนกระทั่งภายหลัง[ 7 ]น่าเสียดายที่รากมักไม่ได้รับการเก็บรักษาไว้ในบันทึกฟอสซิล[ 7 ]
ไรซอยด์ – โครงสร้างขนาดเล็กที่ทำหน้าที่เหมือนราก โดยปกติมีเส้นผ่านศูนย์กลางเท่ากับเซลล์ – น่าจะวิวัฒนาการมาตั้งแต่ยุคแรกๆ อาจจะก่อนที่พืชจะเข้ามาอาศัยอยู่บนบกด้วยซ้ำ พวกมันถูกพบในCharaceaeซึ่งเป็นกลุ่มสาหร่ายที่เป็นญาติใกล้ชิดกับพืชบก[ 7 ]อย่างไรก็ตาม ไรซอยด์น่าจะวิวัฒนาการมากกว่าหนึ่งครั้ง ไรซีนของไลเคนตัวอย่างเช่น ทำหน้าที่คล้ายกัน แม้แต่สัตว์บางชนิด ( Lamellibrachia ) ก็มีโครงสร้างคล้ายราก[ 7 ]ไรซอยด์สามารถมองเห็นได้อย่างชัดเจนใน ฟอสซิลหิน เชิร์ต Rhynieและมีอยู่ในพืชมีท่อลำเลียงยุคแรกๆ ส่วนใหญ่ และจากพื้นฐานนี้ดูเหมือนว่าจะเป็นลางบอกเหตุของรากพืชที่แท้จริง[ 83 ]
โครงสร้างที่ซับซ้อนกว่านั้นพบได้ทั่วไปในหินเชิร์ต Rhynie และฟอสซิลอื่นๆ อีกมากมายที่มีอายุในยุคดีโวเนียนตอนต้นที่เทียบเคียงกันได้มีโครงสร้างที่ดูเหมือนและทำหน้าที่เหมือนราก[ 7 ]ไรนิโอไฟต์มีไรโซอยด์ขนาดเล็ก และไตรเมอโรไฟต์และไลโคพอดที่เป็นพืชล้มลุกในหินเชิร์ตมีโครงสร้างคล้ายรากที่แทรกซึมลงไปในดินลึกไม่กี่เซนติเมตร[ 84 ]อย่างไรก็ตาม ฟอสซิลเหล่านี้ไม่มีชิ้นใดแสดงคุณสมบัติทั้งหมดของรากสมัยใหม่[ 7 ]ยกเว้นAsteroxylonซึ่งเพิ่งได้รับการยอมรับว่ามีรากที่วิวัฒนาการอย่างอิสระจากรากของพืชมีท่อลำเลียงในปัจจุบัน[ 85 ]รากและโครงสร้างคล้ายรากกลายเป็นเรื่องธรรมดามากขึ้นและแทรกซึมลึกขึ้นในช่วงยุคดีโวเนียนโดยต้นไลโคพอดสร้างรากยาวประมาณ 20 เซนติเมตรในช่วงยุคอีเฟเลียนและกิเวเทียน ต่อมาก็มีโปรจิมโนสเปิร์มซึ่งมีรากลึกถึงประมาณหนึ่งเมตรในช่วงยุคฟราสเนียน[ 84 ]พืชเมล็ดเปลือยแท้และเฟิร์นไซโกปเทอริดยังสร้างระบบรากตื้นในช่วงฟาเมนเนียนด้วย[ 84 ]
ไรโซฟอร์ของไลโคพอดให้วิธีการสร้างรากที่แตกต่างออกไปเล็กน้อย พวกมันเทียบเท่ากับลำต้น โดยมีอวัยวะที่เทียบเท่ากับใบทำหน้าที่เป็นรากฝอย[ 7 ]โครงสร้างที่คล้ายกันนี้พบได้ในไลโคพอดIsoetes ที่มีอยู่ในปัจจุบัน และสิ่งนี้ดูเหมือนจะเป็นหลักฐานว่ารากวิวัฒนาการอย่างอิสระอย่างน้อยสองครั้งในไลโคไฟต์และพืชชนิดอื่น[ 7 ]ข้อเสนอนี้ได้รับการสนับสนุนจากการศึกษาที่แสดงให้เห็นว่ารากเริ่มต้นและการเจริญเติบโตของรากได้รับการส่งเสริมโดยกลไกที่แตกต่างกันในไลโคไฟต์และยูฟิลโลไฟต์[ 86 ]
พืชที่มีรากในยุคแรกนั้นก้าวหน้าไปเพียงเล็กน้อยเมื่อเทียบกับบรรพบุรุษในยุคไซลูเรียน โดยไม่มีระบบรากที่เฉพาะเจาะจง อย่างไรก็ตาม สามารถมองเห็นแกนที่แผ่ราบได้อย่างชัดเจนว่ามีการเจริญเติบโตที่คล้ายกับไรโซอยด์ของไบรโอไฟต์ในปัจจุบัน[ 87 ]
เมื่อถึงช่วงปลายยุคดีโวเนียน กลุ่มพืชส่วนใหญ่ได้พัฒนาระบบรากขึ้นมาเองโดยอิสระ[ 87 ]เมื่อรากมีขนาดใหญ่ขึ้น ก็สามารถรองรับต้นไม้ขนาดใหญ่ขึ้นได้ และดินก็ผุพังลงไปลึกกว่าเดิม[ 79 ]การผุพังที่ลึกขึ้นนี้ไม่เพียงแต่ส่งผลต่อการลดลงของ CO 2 ดังที่กล่าวมาแล้วเท่านั้น แต่ยังเปิดแหล่งที่อยู่อาศัยใหม่สำหรับการตั้งรกรากของเชื้อราและสัตว์อีกด้วย[ 84 ]
รากที่แคบที่สุดของพืชสมัยใหม่มีเส้นผ่านศูนย์กลางเพียง 40 ไมโครเมตร และไม่สามารถลำเลียงน้ำได้หากแคบกว่านี้[ 7 ]ในทางตรงกันข้าม รากฟอสซิลที่เก่าแก่ที่สุดที่ค้นพบนั้นแคบลงจาก 3 มิลลิเมตรเหลือต่ำกว่า 700 ไมโครเมตรในเส้นผ่านศูนย์กลาง แน่นอนว่ากระบวนการเกิดฟอสซิลเป็นตัวควบคุมขั้นสุดท้ายของความหนาที่มองเห็นได้[ 7 ]
รูปทรงต้นไม้


ภูมิทัศน์ในยุคดีโวเนียนตอนต้นนั้นปราศจากพืชพรรณที่สูงเกินระดับเอว ความสูงที่มากกว่านั้นให้ความได้เปรียบในการแข่งขันในการเก็บเกี่ยวแสงแดดเพื่อการสังเคราะห์แสง การบังเงาคู่แข่ง และการกระจายสปอร์ เนื่องจากสปอร์ (และต่อมาคือเมล็ด) สามารถถูกลมพัดไปได้ไกลกว่าหากเริ่มต้นจากที่สูงกว่า ระบบท่อลำเลียงที่มีประสิทธิภาพจึงจำเป็นต่อการเจริญเติบโตให้สูงขึ้น เพื่อให้พืชมีลักษณะเป็นต้นไม้พืชต้องพัฒนา เนื้อเยื่อ ไม้ที่ให้ทั้งการค้ำจุนและการลำเลียงน้ำ ดังนั้นจึงจำเป็นต้องพัฒนาความสามารถในการเจริญเติบโตทุติยภูมิ เนื้อเยื่อแกนกลางของพืชที่กำลังเจริญเติบโตทุติยภูมิจะถูกล้อมรอบด้วยแคมเบียมหลอดเลือดซึ่งเป็นวงแหวนของเซลล์เมริสเตมที่สร้างไซเล็มเพิ่มเติมด้านในและโฟลเอมด้านนอก เนื่องจากเซลล์ไซเล็มประกอบด้วยเนื้อเยื่อที่ตายแล้วและมีลิกนิน วงแหวนไซเล็มที่เกิดขึ้นใหม่จะถูกเพิ่มเข้าไปในวงแหวนที่มีอยู่แล้ว ทำให้เกิดเป็นเนื้อไม้ ฟอสซิลของพืชจากยุคดีโวเนียนตอนต้นแสดงให้เห็นว่ารูปแบบของเนื้อไม้ที่เรียบง่ายปรากฏขึ้นครั้งแรกอย่างน้อย 400 ล้านปีก่อน ในช่วงเวลาที่พืชบกทั้งหมดมีขนาดเล็กและเป็นพืชล้มลุก[ 88 ]เนื่องจากไม้มีวิวัฒนาการมานานก่อนพุ่มไม้และต้นไม้ จึงเป็นไปได้ว่าจุดประสงค์ดั้งเดิมของมันคือการขนส่งน้ำ และต่อมาจึงนำมาใช้เพื่อการรองรับทางกล[ 89 ]
พืชชนิดแรกที่พัฒนาการเจริญเติบโตทุติยภูมิและมีลักษณะเป็นเนื้อไม้ เห็นได้ชัดว่าเป็นเฟิร์น และในช่วงต้นยุคดีโวเนียนตอนกลาง เฟิร์นสายพันธุ์Wattiezaก็มีความสูงถึง 8 เมตรและมีลักษณะคล้ายต้นไม้แล้ว[ 90 ]

กลุ่มอื่นๆ ใช้เวลาไม่นานในการพัฒนารูปร่างคล้ายต้นไม้Archaeopteris ในช่วงปลายยุคดีโวเนียน ซึ่ง เป็นบรรพบุรุษของพืชเมล็ดเปลือยที่วิวัฒนาการมาจากไตรเมอโรไฟต์[ 91 ]มีความสูงถึง 30 เมตร พืชเมล็ดเปลือยเป็นพืชกลุ่มแรกที่พัฒนาเนื้อไม้แท้ ซึ่งเติบโตจากแคมเบียมสองด้าน การปรากฏตัวครั้งแรกของพืชกลุ่มนี้ คือRellimiaเกิดขึ้นในช่วงกลางยุคดีโวเนียน[ 92 ]เชื่อกันว่าเนื้อไม้แท้มีการวิวัฒนาการเพียงครั้งเดียว ทำให้เกิดแนวคิดของกลุ่ม "ลิกโนไฟต์" ขึ้นมา
ป่า Archaeopterisได้รับการเสริมด้วยไลโคพอดที่มีลักษณะเป็นต้นไม้ในรูปแบบของLepidodendralesซึ่งมีความสูงเกิน 50 เมตรและกว้าง 2 เมตรที่โคนต้น ไลโคพอดที่มีลักษณะเป็นต้นไม้เหล่านี้กลายเป็นพืชเด่นในป่าช่วงปลายยุคดีโวเนียนและยุคคาร์บอนิเฟอรัส ซึ่งเป็นแหล่งกำเนิดของแหล่งถ่านหิน[ 93 ] Lepidodendrales แตกต่างจากต้นไม้สมัยใหม่ตรงที่มีการเจริญเติบโตแบบจำกัด: หลังจากสะสมสารอาหารสำรองไว้ที่ความสูงระดับต่ำ พืชจะ "แตกกิ่ง" เป็นลำต้นเดียวไปจนถึงความสูงที่กำหนดโดยพันธุกรรม แตกกิ่งที่ระดับนั้น แพร่สปอร์ และตาย[ 94 ]พวกมันประกอบด้วยเนื้อไม้ "ราคาถูก" เพื่อให้สามารถเจริญเติบโตได้อย่างรวดเร็ว โดยอย่างน้อยครึ่งหนึ่งของลำต้นประกอบด้วยโพรงที่เต็มไปด้วยเนื้อเยื่อแกนกลาง[ 1 ]เนื้อไม้ของพวกมันยังถูกสร้างขึ้นโดยแคมเบียมหลอดเลือดแบบหน้าเดียว ซึ่งไม่ได้สร้างโฟลเอ็มใหม่ หมายความว่าลำต้นไม่สามารถเติบโตให้กว้างขึ้นได้เมื่อเวลาผ่านไป
หญ้าหางม้าสกุลCalamitesปรากฏขึ้นในยุคคาร์บอนิเฟอรัส แตกต่างจากหญ้าหางม้า สกุลEquisetumในปัจจุบันCalamitesมีแคมเบียมหลอดเลือดแบบหน้าเดียว ทำให้สามารถสร้างเนื้อไม้และเติบโตได้สูงเกิน 10 เมตร และแตกกิ่งก้านสาขาได้ซ้ำแล้วซ้ำเล่า
แม้ว่ารูปร่างของต้นไม้ในยุคแรกจะคล้ายกับต้นไม้ในปัจจุบัน แต่ พืช มีเมล็ดหรือพืชสเปิร์ม ซึ่งเป็นกลุ่มที่ประกอบด้วยต้นไม้สมัยใหม่ทั้งหมด ยังไม่วิวัฒนาการ กลุ่มต้นไม้ที่โดดเด่นในปัจจุบันล้วนเป็นพืชมีเมล็ด ได้แก่ พืชเมล็ดเปลือย ซึ่งรวมถึงต้นสน และพืชดอก ซึ่งรวมถึงต้นไม้ที่มีผลและดอกทั้งหมด ไม่มีต้นไม้ที่ออกสปอร์อิสระเช่นArchaeopterisอยู่ในพืชที่ยังมีชีวิตอยู่ เป็นที่เชื่อกันมานานแล้วว่าพืชดอกเกิดขึ้นจากภายในพืชเมล็ดเปลือย แต่หลักฐานทางโมเลกุลล่าสุดชี้ให้เห็นว่าตัวแทนที่ยังมีชีวิตอยู่ของพวกมันก่อตัวเป็นสองกลุ่มที่แตกต่างกัน[ 95 ] [ 96 ] [ 97 ]ข้อมูลทางโมเลกุลยังไม่สอดคล้องกับข้อมูลทางสัณฐานวิทยาอย่างสมบูรณ์[ 98 ] [ 99 ] [ 100 ]แต่กำลังเป็นที่ยอมรับว่าการสนับสนุนทางสัณฐานวิทยาสำหรับพาราไฟลีนั้นไม่แข็งแกร่งนัก[ 101 ] สิ่งนี้จะนำไปสู่ข้อสรุปว่าทั้งสองกลุ่มเกิดขึ้นจากภายในพืชกลุ่มเทอริโดสเปิร์มซึ่งน่าจะเกิดขึ้นตั้งแต่ยุคเพอร์เมียน[ 101 ]
พืชดอกและบรรพบุรุษของพวกมันมีบทบาทเพียงเล็กน้อยจนกระทั่งมีการแตกแขนงออกไปในช่วงยุคครีเทเชียส พวกมันเริ่มต้นจากการเป็นสิ่งมีชีวิตขนาดเล็กที่ชอบความชื้นในชั้นล่างของป่า และมีการแตกแขนงออกไปเรื่อยๆ นับตั้งแต่ยุคครีเทเชียส[ 102 ]จนกลายเป็นสมาชิกที่โดดเด่นของป่าที่ไม่ใช่เขตหนาวในปัจจุบัน
เมล็ดพันธุ์


พืชบกในยุคแรกสืบพันธุ์ในลักษณะเดียวกับเฟิร์น: สปอร์งอกเป็นแกมีโทไฟต์ขนาดเล็ก ซึ่งผลิตไข่และ/หรืออสุจิ อสุจิเหล่านี้จะว่ายน้ำข้ามดินชื้นไปหาอวัยวะเพศเมีย (อาร์คีโกเนีย) บนแกมีโทไฟต์เดียวกันหรือแกมีโทไฟต์อื่น ซึ่งพวกมันจะรวมกับไข่เพื่อสร้างเอ็มบริโอ ซึ่งจะงอกเป็นสปอโรไฟต์[ 84 ]
พืช เฮเทอโรสปอริก (Heterosporic plants) ตามชื่อที่บ่งบอก มีสปอร์สองขนาด คือไมโครสปอร์และเมกาสปอร์ ซึ่งจะงอกเป็นไมโครกามีโทไฟต์และเมกากามีโทไฟต์ตามลำดับ ระบบนี้ปูทางไปสู่ไข่และเมล็ด: ในกรณีที่รุนแรงที่สุด เมกาสปอแรนเจียมสามารถบรรจุเมกาสปอร์เททราดได้เพียงกลุ่มเดียว และเพื่อให้การเปลี่ยนไปเป็นไข่ที่แท้จริงสมบูรณ์ เมกาสปอร์สามในเททราดดั้งเดิมอาจถูกทำลาย เหลือเมกาสปอร์หนึ่งอันต่อเมกาสปอแรนเจียม
การเปลี่ยนผ่านไปสู่ไข่ยังคงดำเนินต่อไปโดยเมกะสปอร์นี้ถูก "กักขัง" ไว้ในสปอแรนเจียมขณะที่มันงอก จากนั้นเมกะกาเมโทไฟต์จะถูกบรรจุอยู่ภายในเยื่อหุ้มที่กันน้ำซึ่งห่อหุ้มเมล็ดไว้ ละอองเรณูซึ่งมีไมโครกาเมโทไฟต์ที่งอกมาจากไมโครสปอร์ ถูกนำมาใช้ในการกระจายตัวของแกมีตเพศผู้ โดยจะปล่อยสเปิร์มแฟลเจลเลตที่ไวต่อการแห้งเมื่อไปถึงเมกะกาเมโทไฟต์ที่พร้อมรับเท่านั้น[ 1 ]
ไลโคพอดและสเฟโนปซิดได้ก้าวไปไกลพอสมควรในเส้นทางสู่การมีเมล็ดโดยไม่เคยข้ามขีดจำกัด มีการค้นพบเมกะสปอร์ของไลโคพอดที่เป็นฟอสซิลที่มีเส้นผ่านศูนย์กลางถึง 1 ซม. และล้อมรอบด้วยเนื้อเยื่อพืช (Lepidocarpon, Achlamydocarpon) ซึ่งเมกะสปอร์เหล่านี้สามารถงอกเป็นเมกะกาเมโทไฟต์ได้ในแหล่งกำเนิดอย่างไรก็ตาม พวกมันยังไม่ถือว่าเป็นไข่ เนื่องจากนิวเคลลัส ซึ่งเป็นชั้นหุ้มสปอร์ด้านใน ไม่ได้ห่อหุ้มสปอร์ไว้อย่างสมบูรณ์ ยังมีช่องเล็กๆ (ไมโครไพล์) เหลืออยู่ ซึ่งหมายความว่าเมกะสปอแรนเจียมยังคงสัมผัสกับบรรยากาศ สิ่งนี้มีผลสองประการ ประการแรก คือ มันไม่ทนต่อการแห้งอย่างสมบูรณ์ และประการที่สอง อสุจิไม่จำเป็นต้อง "เจาะ" เพื่อเข้าถึงอาร์คีโกเนียของเมกะสปอร์[ 1 ]
มีการระบุพืชมีเมล็ดในยุคดีโวเนียนตอนกลาง จากเบลเยียม ซึ่งมีอายุเก่าแก่กว่า พืชมีเมล็ดที่ เก่าแก่ที่สุด ประมาณ 20 ล้านปีรันคาเรีย (Runcaria)มีขนาดเล็กและสมมาตรตามแนวรัศมี เป็นเมกาสปอแรน เจียมที่มีเยื่อหุ้มล้อม รอบด้วย คัพพูล เมกาสปอแรนเจียมมีส่วนยื่นปลายที่ยังไม่เปิดยื่นออกมาเหนือเยื่อหุ้มที่มีหลายแฉกคาดว่าส่วนยื่นนี้มีส่วนเกี่ยวข้องกับการผสมเกสรโดยลม รันคา เรี ยให้ความกระจ่างใหม่เกี่ยวกับลำดับการได้มาซึ่งลักษณะที่นำไปสู่เมล็ดรันคาเรียมีคุณสมบัติทั้งหมดของพืชมีเมล็ด ยกเว้นเปลือกเมล็ด ที่แข็ง และระบบนำทางละอองเรณูไปยังไข่[ 103 ]
พืชมีเมล็ดชนิดแรก (แปลตรงตัวว่า "พืชมีเมล็ด") – กล่าวคือ พืชชนิดแรกที่สร้างเมล็ดที่แท้จริง – เรียกว่า เท อริโดสเปิ ร์ม (pteridosperms ) ซึ่งแปลตรงตัวว่า "เฟิร์นมีเมล็ด" เนื่องจากใบของพวกมันประกอบด้วยใบคล้ายเฟิร์น แม้ว่าพวกมันจะไม่ได้มีความสัมพันธ์ใกล้ชิดกับเฟิร์นก็ตาม หลักฐานฟอสซิลที่เก่าแก่ที่สุดของพืชมีเมล็ดมีอายุอยู่ในช่วงปลายยุคดีโวเนียน และดูเหมือนว่าพวกมันจะวิวัฒนาการมาจากกลุ่มก่อนหน้าที่เรียกว่า โปรจิมโนส เปิร์ม (progymnosperms ) พืชมีเมล็ดในยุคแรกเหล่านี้มีตั้งแต่ต้นไม้ไปจนถึงไม้พุ่มเลื้อยขนาดเล็ก เช่นเดียวกับโปรจิมโนสเปิร์มในยุคแรกส่วนใหญ่ พวกมันเป็นพืชเนื้อไม้ที่มีใบคล้ายเฟิร์น พวกมันทั้งหมดมีไข่ แต่ไม่มีกรวย ผลไม้ หรือสิ่งอื่นที่คล้ายกัน แม้ว่าจะเป็นเรื่องยากที่จะติดตามวิวัฒนาการของเมล็ดในยุคแรก แต่สายพันธุ์ของเฟิร์นมีเมล็ดอาจสืบย้อนไปได้จากไตรเมอโรไฟต์ (trimerophytes) ที่เรียบง่ายไปจนถึงอะนิวโรไฟต์ ( Aneurophytes ) ที่มีสปอร์ชนิดเดียวกัน [ 1 ] พืชมีเมล็ดมีการวิวัฒนาการครั้งใหญ่ครั้งแรกใน ยุค ฟาเมนเนียน (Famennian era) [ 104 ]
รูปแบบเมล็ดแบบนี้พบได้ในพืชเมล็ดเปลือยเกือบทั้งหมด (พืชเมล็ดเปลือย) ซึ่งส่วนใหญ่จะห่อหุ้มเมล็ดด้วยกรวยไม้หรือเนื้อเยื่อหุ้มเมล็ด ( เช่น ต้นยิว ) แต่ไม่มีพืชชนิดใดที่ห่อหุ้มเมล็ดอย่างสมบูรณ์ พืชดอก (พืชเมล็ดมีท่อลำเลียง) เป็นกลุ่มเดียวที่ห่อหุ้มเมล็ดอย่างสมบูรณ์ด้วยเนื้อเยื่อหุ้มเมล็ด (carpel)
เมล็ดที่ห่อหุ้มอย่างสมบูรณ์ได้เปิดเส้นทางใหม่ให้พืชได้ดำเนินตาม นั่นคือการพักตัวของเมล็ดตัวอ่อนซึ่งแยกตัวออกจากบรรยากาศภายนอกอย่างสมบูรณ์และได้รับการปกป้องจากการแห้ง สามารถอยู่รอดได้ในสภาวะแห้งแล้งหลายปีก่อนที่จะงอก เมล็ดพืชเมล็ดเปลือยจากยุคคาร์บอนิเฟอรัสตอนปลายพบว่ามีตัวอ่อน ซึ่งบ่งชี้ว่ามีช่วงเวลาที่ยาวนานระหว่างการปฏิสนธิและการงอก[ 105 ]ช่วงเวลานี้เกี่ยวข้องกับการเข้าสู่ ยุค เรือนกระจกของโลกซึ่งส่งผลให้ความแห้งแล้งเพิ่มขึ้น สิ่งนี้ชี้ให้เห็นว่าการพักตัวเกิดขึ้นเพื่อตอบสนองต่อสภาพภูมิอากาศที่แห้งแล้งมากขึ้น ซึ่งการรอช่วงเวลาที่มีความชื้นก่อนการงอกกลายเป็นสิ่งที่เป็นประโยชน์[ 105 ]ความก้าวหน้าทางวิวัฒนาการนี้ดูเหมือนจะเปิดประตูสู่โอกาส พื้นที่ที่ไม่เอื้ออำนวยก่อนหน้านี้ เช่น เนินเขาที่แห้งแล้ง สามารถทนทานได้ และในไม่ช้าก็ถูกปกคลุมด้วยต้นไม้[ 105 ]
เมล็ดพืชมีข้อดีเพิ่มเติมสำหรับผู้ถือครอง ได้แก่ การเพิ่มอัตราความสำเร็จของแกมีโทไฟต์ที่ได้รับการผสมพันธุ์ และเนื่องจากสารอาหารสามารถ "บรรจุ" ไว้ในเอ็มบริโอได้ เมล็ดจึงสามารถงอกได้อย่างรวดเร็วในสภาพแวดล้อมที่ไม่เอื้ออำนวย และเติบโตจนมีขนาดที่สามารถดูแลตัวเองได้เร็วขึ้น[ 84 ]ตัวอย่างเช่น หากไม่มีเอนโดสเปิร์ม ต้นกล้าที่เติบโตในสภาพแวดล้อมที่แห้งแล้งจะไม่มีแหล่งสำรองพลังงานเพียงพอที่จะสร้างรากให้ลึกพอที่จะไปถึงระดับน้ำใต้ดินก่อนที่จะตายเพราะขาดน้ำ[ 84 ]ในทำนองเดียวกัน เมล็ดที่งอกในที่ร่มเงาของต้นไม้ต้องการแหล่งสำรองพลังงานเพิ่มเติมเพื่อเติบโตให้สูงพอที่จะรับแสงได้เพียงพอสำหรับการดำรงชีวิต[ 84 ] การรวมกันของข้อดีเหล่านี้ทำให้พืชมีเมล็ดได้เปรียบทางนิเวศวิทยาเหนือสกุลArchaeopteris ที่เคยครองอำนาจมาก่อน จึงเพิ่มความหลากหลายทางชีวภาพของป่าในยุคแรก[ 84 ]
ถึงแม้จะมีข้อดีเหล่านี้ แต่โดยทั่วไปแล้วไข่ที่ได้รับการผสมพันธุ์แล้วมักจะไม่เจริญเติบโตเป็นเมล็ด[ 106 ]นอกจากนี้ ในระหว่างการพักตัวของเมล็ด (ซึ่งมักเกี่ยวข้องกับสภาวะที่ไม่สามารถคาดเดาได้และก่อให้เกิดความเครียด) ความเสียหายของ DNA ก็จะสะสมมากขึ้น[ 107 ] [ 108 ] [ 109 ]ดังนั้น ความเสียหายของ DNA จึงดูเหมือนจะเป็นปัญหาพื้นฐานต่อการอยู่รอดของพืชมีเมล็ด เช่นเดียวกับที่ความเสียหายของ DNA เป็นปัญหาสำคัญต่อสิ่งมีชีวิตโดยทั่วไป[ 110 ]
ดอกไม้

ดอกไม้เป็นใบที่ดัดแปลงแล้วซึ่งพบได้เฉพาะในพืชดอกซึ่งปรากฏในบันทึกฟอสซิลค่อนข้างช้า กลุ่มนี้กำเนิดและแตกแขนงออกไปในช่วงต้นยุคครีเทเชียส และมีความสำคัญทางนิเวศวิทยาหลังจากนั้น[ 111 ]โครงสร้างคล้ายดอกไม้ปรากฏครั้งแรกใน บันทึก ฟอสซิลเมื่อประมาณ 130 ล้านปีก่อน ในยุคครีเทเชียส [ 112 ] อย่างไรก็ตามในปี 2018 นักวิทยาศาสตร์รายงานการค้นพบดอกไม้ ฟอสซิล จากเมื่อประมาณ 180 ล้านปีก่อน ซึ่งเร็วกว่าที่เคยคิดไว้ถึง 50 ล้านปี[ 113 ]อย่างไรก็ตาม การตีความนี้ถูกโต้แย้งอย่างมาก[ 114 ]
โครงสร้างที่มีสีสันและ/หรือมีกลิ่นฉุนล้อมรอบโคนของพืช เช่นไซแคดและGnetalesทำให้การกำหนดนิยามที่ชัดเจนของคำว่า "ดอกไม้" เป็นเรื่องยาก[ 100 ]
หน้าที่หลักของดอกไม้คือการสืบพันธุ์ซึ่งก่อนการวิวัฒนาการของดอกไม้และพืชดอกนั้น เป็นหน้าที่ของไมโครสปอโรฟิลล์และเมกาสปอโรฟิลล์ ดอกไม้สามารถถือได้ว่าเป็นนวัตกรรม ทางวิวัฒนาการที่ทรงพลัง เพราะการมีอยู่ของดอกไม้ทำให้โลกของพืชสามารถเข้าถึงวิธีการและกลไกใหม่ๆ ในการสืบพันธุ์ได้
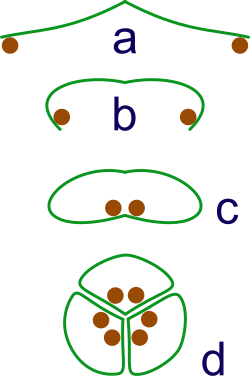
เชื่อกันมานานแล้วว่าพืชดอกมีวิวัฒนาการมาจากกลุ่มจิมโนสเปิร์มตามมุมมองทางสัณฐานวิทยาแบบดั้งเดิม พวกมันมีความสัมพันธ์ใกล้ชิดกับกลุ่มGnetalesอย่างไรก็ตาม ดังที่กล่าวไว้ข้างต้น หลักฐานทางโมเลกุลล่าสุดขัดแย้งกับสมมติฐานนี้[ 96 ] [ 97 ]และยังชี้ให้เห็นเพิ่มเติมว่า Gnetales มีความสัมพันธ์ใกล้ชิดกับกลุ่มจิมโนสเปิร์มบางกลุ่มมากกว่าแองจิโอสเปิร์ม[ 95 ]และจิมโนสเปิร์มที่ยังมีชีวิตอยู่ก่อตัวเป็นกลุ่มที่แตกต่างจากแองจิโอสเปิร์ม[ 95 ] [ 96 ] [ 97 ]โดยทั้งสองกลุ่มแยกออกจากกันเมื่อประมาณ300 ล้านปีก่อน[ 115 ]
ความสัมพันธ์ของกลุ่มลำต้นกับพืชดอกมีความสำคัญในการกำหนดวิวัฒนาการของดอกไม้ กลุ่มลำต้นให้ข้อมูลเชิงลึกเกี่ยวกับสถานะของ "ทางแยก" ก่อนหน้านี้บนเส้นทางสู่สถานะปัจจุบัน การบรรจบกันเพิ่มความเสี่ยงในการระบุกลุ่มลำต้นผิดพลาด เนื่องจากการปกป้องเมกาแกมีโทไฟต์เป็นสิ่งที่พึงปรารถนาในเชิงวิวัฒนาการ จึงเป็นไปได้ว่าหลายกลุ่มได้วิวัฒนาการเปลือกหุ้มป้องกันแยกจากกัน ในดอกไม้ การป้องกันนี้อยู่ในรูปของคาร์เพล ซึ่งวิวัฒนาการมาจากใบและถูกนำมาใช้ในบทบาทการป้องกัน โดยปกป้องไข่ ไข่เหล่านี้ได้รับการปกป้องเพิ่มเติมด้วย เยื่อหุ้มสองชั้น
การเจาะทะลุชั้นป้องกันเหล่านี้จำเป็นต้องมีอะไรมากกว่าไมโครแกมีโทไฟต์ที่ลอยอยู่พืชดอกมีละอองเรณูที่ประกอบด้วยเซลล์เพียงสามเซลล์ เซลล์หนึ่งทำหน้าที่เจาะทะลุเปลือกหุ้มและสร้างท่อให้เซลล์สเปิร์มสองเซลล์ไหลลงไป เมกะแกมีโทไฟต์มีเพียงเจ็ดเซลล์ ในจำนวนนี้ หนึ่งเซลล์จะรวมกับเซลล์สเปิร์มเพื่อสร้างนิวเคลียสของไข่ และอีกเซลล์หนึ่งจะรวมกับสเปิร์มอีกเซลล์หนึ่งและทำหน้าที่สร้างเอนโดสเปิร์ม ที่อุดมไปด้วยสารอาหาร ส่วนเซลล์อื่นๆ ทำหน้าที่เสริม กระบวนการ " การปฏิสนธิสองครั้ง " นี้เป็นเอกลักษณ์และพบได้ทั่วไปในพืชดอกทุกชนิด

ในบันทึกฟอสซิล มีกลุ่มพืชที่น่าสนใจสามกลุ่มที่มีโครงสร้างคล้ายดอกไม้ กลุ่มแรกคือGlossopterisซึ่งเป็นพืชกลุ่ม เฟิร์นในยุคเพ อร์เมียน ที่มีใบโค้งงอคล้ายกลีบดอก กลุ่มที่สองคือ Caytoniaในยุค มีโซโซอิก มีลักษณะคล้ายดอกไม้มากยิ่งขึ้น โดยมีไข่ที่อยู่ภายใน แต่มีเยื่อหุ้มเพียงชั้นเดียว นอกจากนี้ รายละเอียดของละอองเรณูและเกสรตัวผู้ยังทำให้พวกมันแตกต่างจากพืชดอกแท้จริงอีกด้วย
พืช ในอันดับ Bennettitalesมีอวัยวะคล้ายดอกไม้ที่โดดเด่น ซึ่งได้รับการปกป้องด้วยกลีบเลี้ยงที่อาจมีบทบาทคล้ายกับกลีบดอกและกลีบเลี้ยงของดอกไม้จริง อย่างไรก็ตาม โครงสร้างคล้ายดอกไม้เหล่านี้วิวัฒนาการขึ้นอย่างอิสระ เนื่องจากพืชในอันดับ Bennettitales มีความสัมพันธ์ใกล้ชิดกับไซแคดและแปะก๊วยมากกว่าพืชดอก[ 116 ]
อย่างไรก็ตาม ไม่พบดอกไม้ที่แท้จริงในกลุ่มใดๆ ยกเว้นกลุ่มที่ยังมีชีวิตอยู่ในปัจจุบัน การวิเคราะห์ทางสัณฐานวิทยาและโมเลกุลส่วนใหญ่จัดให้Amborella , nymphaealesและAustrobaileyaceaeอยู่ในกลุ่มพื้นฐานที่เรียกว่า "ANA" กลุ่มนี้ดูเหมือนจะแยกตัวออกมาในช่วงต้นยุคครีเทเชียส ประมาณ130 ล้านปีก่อน ซึ่งเป็นช่วงเวลาเดียวกับฟอสซิลพืชดอกที่เก่าแก่ที่สุด [ 117 ] [ 118 ] และหลังจากละอองเรณูที่คล้ายพืชดอกครั้งแรก เมื่อ 136 ล้านปีก่อน[ 101 ]แมกโนลิดแยกตัวออกมาหลังจากนั้นไม่นาน และการแพร่กระจายอย่างรวดเร็วทำให้เกิดยูไดคอตและโมโนคอตเมื่อ125 ล้านปีก่อน [ 101 ] เมื่อสิ้นสุดยุคครีเทเชียส เมื่อ 66 ล้านปีก่อนพืชดอกมากกว่า 50% ของอันดับในปัจจุบันได้วิวัฒนาการขึ้น และกลุ่มนี้คิดเป็น 70% ของสายพันธุ์ทั่วโลก[ 119 ]ในช่วงเวลานี้เองที่ต้นไม้ดอกกลายเป็นพืชเด่นเหนือต้นสน[ 1 ] : 498
ลักษณะของกลุ่ม "ANA" พื้นฐานบ่งชี้ว่าพืชดอกมีต้นกำเนิดในพื้นที่มืดชื้นและถูกรบกวนบ่อยครั้ง[ 120 ]ดูเหมือนว่าพืชดอกจะยังคงถูกจำกัดอยู่ในถิ่นที่อยู่ดังกล่าวตลอดช่วงยุคครีเทเชียส โดยครอบครองช่องว่างของพืชล้มลุกขนาดเล็กในช่วงต้นของลำดับการเปลี่ยนแปลง[ 119 ]สิ่งนี้อาจจำกัดความสำคัญในเบื้องต้นของพวกมัน แต่ให้ความยืดหยุ่นแก่พวกมันซึ่งเป็นสาเหตุของความรวดเร็วในการกระจายพันธุ์ในภายหลังในถิ่นที่อยู่อื่นๆ[ 120 ]
| มุมมองแบบดั้งเดิม | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
| มุมมองสมัยใหม่ | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
บางคนเสนอว่าพืชดอกเกิดขึ้นจากเฟิร์นเมล็ดที่ไม่รู้จัก Pteridophyte และมองว่าไซแคดเป็นเฟิร์นเมล็ดที่มีชีวิตที่มีทั้งใบที่มีเมล็ดและใบที่ไม่มีเมล็ด ( Cycas revoluta ) [ 99 ]
ในเดือนสิงหาคม พ.ศ. 2560 นักวิทยาศาสตร์ได้นำเสนอคำอธิบายโดยละเอียดและการสร้างแบบจำลอง 3 มิติของดอกไม้ชนิดแรกที่มีชีวิตอยู่เมื่อประมาณ 140 ล้านปีก่อน[ 122 ] [ 123 ]
ที่มาของดอกไม้
วงศ์Amborellaceaeถือเป็นกลุ่มพี่น้องของพืชดอกที่ยังมีชีวิตอยู่ทั้งหมด จีโนมฉบับร่างของAmborella trichopodaได้รับการเผยแพร่ในเดือนธันวาคม พ.ศ. 2556 [ 124 ]โดยการเปรียบเทียบจีโนมของมันกับจีโนมของพืชดอกที่ยังมีชีวิตอยู่ทั้งหมด จะทำให้สามารถระบุลักษณะที่น่าจะเป็นไปได้มากที่สุดของบรรพบุรุษของA. trichopodaและพืชดอกอื่นๆ ทั้งหมด นั่นคือพืชดอกบรรพบุรุษ[ 125 ]
ดูเหมือนว่าในระดับของอวัยวะใบอาจเป็นบรรพบุรุษของดอกไม้ หรืออย่างน้อยก็อวัยวะบางส่วนของดอกไม้ เมื่อยีนสำคัญบางตัวที่เกี่ยวข้องกับการพัฒนาของดอกไม้เกิดการกลายพันธุ์ กลุ่มของโครงสร้างคล้ายใบก็จะเกิดขึ้นแทนที่ดอกไม้ ดังนั้น ในช่วงเวลาใดเวลาหนึ่งในประวัติศาสตร์ โปรแกรมการพัฒนาที่นำไปสู่การสร้างใบจะต้องถูกเปลี่ยนแปลงไปเพื่อสร้างดอกไม้ นอกจากนี้ อาจมีกรอบการทำงานที่แข็งแกร่งโดยรวมซึ่งเป็นพื้นฐานในการสร้างความหลากหลายของดอกไม้ ตัวอย่างเช่น ยีนที่เรียกว่าLEAFY (LFY)ซึ่งเกี่ยวข้องกับการพัฒนาของดอกไม้ในArabidopsis thaliana ยีน ที่คล้ายคลึงกันนี้พบได้ในพืชดอกหลากหลายชนิด เช่นมะเขือเทศ ดอกสแนปดรากอน ถั่วลันเตาข้าวโพดและแม้แต่พืชเมล็ด เปลือย การแสดงออกของArabidopsis thaliana LFY ในพืชที่อยู่ห่างไกล เช่นต้นป็อปลาร์และส้มก็ส่งผลให้เกิดการสร้างดอกไม้ในพืชเหล่านี้ เช่นกัน ยีน LFYควบคุมการแสดงออกของยีนบางตัวที่อยู่ในตระกูลMADS-boxยีนเหล่านี้ทำหน้าที่ควบคุมการเจริญเติบโตของดอกไม้โดยตรง
หน้าที่การปรับตัวของดอกไม้
ดอกไม้น่าจะเกิดขึ้นระหว่างวิวัฒนาการของพืชในฐานะการปรับตัวเพื่ออำนวยความสะดวกในการผสมข้ามพันธุ์(การผสมข้าม ) ซึ่งเป็นกระบวนการที่นำไปสู่การปกปิดการกลายพันธุ์ แบบด้อยที่เป็นอันตราย ในจีโนม ของลูกหลาน ผลการปกปิดการแสดงออกของการกลายพันธุ์ที่เป็นอันตรายนี้เรียกว่าการเสริมกันทางพันธุกรรม[ 126 ] ผลการปกปิดที่เป็นประโยชน์ของการผสมข้ามพันธุ์นี้ยังถือเป็นพื้นฐานของ ความแข็งแรงของลูกผสมหรือเฮเทอโรซิสในลูกหลาน เมื่อดอกไม้ได้รับการสถาปนาขึ้นในสายพันธุ์โดยอาศัยหน้าที่ในการปรับตัวเพื่อส่งเสริมการผสมข้ามพันธุ์ การเปลี่ยนไปใช้การผสมพันธุ์ในสายเลือดเดียวกันในภายหลังมักจะกลายเป็นข้อเสีย โดยส่วนใหญ่เป็นเพราะมันอนุญาตให้มีการแสดงออกของการกลายพันธุ์แบบด้อยที่เป็นอันตรายที่ถูกปกปิดไว้ก่อนหน้านี้ นั่นคือภาวะซึมเศร้าจากการผสมพันธุ์ในสายเลือดเดียวกันนอกจากนี้ไมโอซิสซึ่งเป็นกระบวนการที่สร้างลูกหลานจากเมล็ดในพืชดอก ยังเป็นกลไกโดยตรงในการซ่อมแซม DNAผ่านการรวมตัวทางพันธุกรรม[ 127 ] ดังนั้น ในพืชดอก กระบวนการสืบพันธุ์แบบอาศัยเพศพื้นฐานสองประการคือ การผสมข้ามพันธุ์ (การผสมข้าม) และไมโอซิส และกระบวนการทั้งสองนี้ดูเหมือนจะได้รับการรักษาไว้โดยอาศัยข้อดีของการเสริมกันทางพันธุกรรมและการซ่อมแซม DNA แบบรีคอมบิเนชันตามลำดับ[ 126 ]
วิวัฒนาการของตระกูล MADS-box
สมาชิกใน กลุ่มปัจจัยถอดรหัส MADS-boxมีบทบาทสำคัญและได้รับการอนุรักษ์ไว้ในเชิงวิวัฒนาการในการพัฒนาของดอกไม้ ตามแบบจำลอง ABC ของการพัฒนาของดอกไม้จะมีการสร้างโซนสามโซน ได้แก่ A, B และ C ภายในดอกตูมที่กำลังพัฒนา โดยการทำงานของปัจจัยถอดรหัส บางชนิด ที่เป็นสมาชิกใน กลุ่ม MADS-boxในบรรดาปัจจัยเหล่านี้ หน้าที่ของยีนในโดเมน B และ C ได้รับการอนุรักษ์ไว้ในเชิงวิวัฒนาการมากกว่ายีนในโดเมน A ยีนเหล่านี้จำนวนมากเกิดขึ้นจากการจำลองยีนของบรรพบุรุษในกลุ่มนี้ และหลายยีนก็มีหน้าที่ซ้ำซ้อนกัน
วิวัฒนาการของ ตระกูล MADS-boxได้รับการศึกษาอย่างกว้างขวาง ยีนเหล่านี้มีอยู่ในเฟิร์น ด้วย แต่การแพร่กระจายและความหลากหลายนั้นสูงกว่ามากในพืชดอก [ 128 ] ดูเหมือนว่าจะมีรูปแบบค่อนข้างมากเกี่ยวกับวิวัฒนาการของตระกูลนี้ ลองพิจารณาวิวัฒนาการของยีนAGAMOUS (AG) ในบริเวณ C ยีนนี้ แสดงออกในดอกไม้ในปัจจุบันในเกสรตัวผู้และเกสรตัวเมียซึ่งเป็นอวัยวะสืบพันธุ์ บรรพบุรุษของมันในพืชเมล็ดเปลือยก็มีรูปแบบการแสดงออกเช่นเดียวกัน โดยแสดงออกในสโตรบิลีซึ่งเป็นอวัยวะที่ผลิตละอองเรณูหรือไข่[ 129 ]ในทำนองเดียวกัน บรรพบุรุษของยีน B (AP3 และ PI)แสดงออกเฉพาะในอวัยวะสืบพันธุ์เพศผู้ในพืชเมล็ดเปลือย ลูกหลานของพวกมันในพืชดอกสมัยใหม่ก็แสดงออกเฉพาะในเกสรตัวผู้ซึ่งเป็นอวัยวะสืบพันธุ์เพศผู้เช่นกัน ดังนั้น พืชจึงนำส่วนประกอบที่มีอยู่เดิมมาใช้ในรูปแบบใหม่เพื่อสร้างดอกไม้ดอกแรก นี่เป็นรูปแบบที่เกิดขึ้นซ้ำๆใน วิวัฒนาการ
ปัจจัยที่มีอิทธิพลต่อความหลากหลายของพืชพรรณ
โครงสร้างดอกของพืชมีความหลากหลายอย่างมาก โดยทั่วไปเกิดจากการเปลี่ยนแปลงใน ยีน MADS-boxและรูปแบบการแสดงออกของยีนเหล่านั้น ตัวอย่างเช่น หญ้ามีโครงสร้างดอกที่เป็นเอกลักษณ์ เกสรตัวเมียและเกสรตัวผู้ถูกล้อมรอบด้วยลอดิคูลที่มีลักษณะคล้ายเกล็ดและใบประดับสองใบ คือ เลมมาและพาเลีย แต่หลักฐานทางพันธุกรรมและสัณฐานวิทยาบ่งชี้ว่าลอดิคูลเป็นโฮโมล็อกกับกลีบดอกของพืชใบเลี้ยงคู่[ 130 ]พาเลียและเลมมาอาจเป็นโฮโมล็อกกับกลีบเลี้ยงในกลุ่มอื่นๆ หรืออาจเป็นโครงสร้างเฉพาะของหญ้า อีกตัวอย่างหนึ่งคือLinaria vulgarisซึ่งมีสมมาตรของดอกสองแบบ คือ สมมาตรแบบรัศมีและสมมาตรแบบทวิภาคสมมาตรเหล่านี้เกิดจาก การเปลี่ยนแปลง ทางเอพิเจเนติกในยีนเพียงยีนเดียวที่เรียกว่าCYCLOIDEA [ 112 ]

Arabidopsis thalianaมียีนชื่อAGAMOUSซึ่งมีบทบาทสำคัญในการกำหนดจำนวนกลีบดอกกลีบเลี้ยงและอวัยวะอื่นๆ ที่เกิดขึ้น การกลายพันธุ์ในยีนนี้ทำให้เนื้อเยื่อเจริญ ของดอก มีชะตากรรมที่ไม่แน่นอน และทำให้เกิดการเพิ่มจำนวนของอวัยวะดอกในกุหลาบ ดอกคาร์เนชั่นและดอกมอร์นิ่งกลอ รี่ แบบดอกซ้อนลักษณะเหล่านี้ได้รับการคัดเลือกโดยนักจัดสวนเนื่องจากมีจำนวนกลีบดอก เพิ่ม ขึ้น[ 131 ]การศึกษาหลายชิ้นในพืชหลากหลายชนิด เช่นเพทูเนียมะเขือเทศอิมพาเทียนส์ข้าวโพดฯลฯชี้ให้เห็นว่าความหลากหลายของดอกไม้จำนวนมากเป็นผลมาจากการเปลี่ยนแปลงเล็กน้อยในยีนที่ควบคุมการพัฒนาของดอกไม้[ 132 ]
โครงการ Floral Genome Projectยืนยันว่าแบบจำลอง ABC ของการพัฒนาของดอกไม้ไม่ได้ถูกอนุรักษ์ไว้ในพืชดอก ทั้งหมด บางครั้งโดเมนการแสดงออกก็เปลี่ยนแปลงไป เช่นในกรณีของพืชใบเลี้ยงเดี่ยว หลายชนิด และในพืชดอกพื้นฐานบางชนิด เช่นAmborella แบบ จำลองการพัฒนาของดอกไม้ที่แตกต่างกัน เช่นแบบจำลองขอบเขตที่จางหายไปหรือแบบจำลองขอบเขตที่ทับซ้อนกันซึ่งเสนอโดเมนการแสดงออกที่ไม่ตายตัว อาจอธิบายสถาปัตยกรรมเหล่านี้ได้[ 133 ]มีความเป็นไปได้ว่าตั้งแต่พืชดอกพื้นฐานไปจนถึงพืชดอกสมัยใหม่ โดเมนของสถาปัตยกรรมดอกไม้มีความคงที่มากขึ้นเรื่อยๆ ผ่านวิวัฒนาการ
ช่วงเวลาออกดอก
ลักษณะทางดอกไม้ประการหนึ่งที่เป็นหัวข้อของการคัดเลือกโดยธรรมชาติคือเวลาออกดอก พืชบางชนิดออกดอกเร็วในวงจรชีวิต ในขณะที่บางชนิดต้องการระยะเวลาของการกระตุ้นด้วยความเย็นก่อนออกดอก ผลลัพธ์นี้ขึ้นอยู่กับปัจจัยต่างๆ เช่นอุณหภูมิความเข้มของแสงการมีอยู่ของแมลงผสมเกสรและสัญญาณสิ่งแวดล้อมอื่นๆ ยีนเช่นCONSTANS (CO) Flowering Locus C ( FLC ) และFRIGIDAควบคุมการรวมสัญญาณสิ่งแวดล้อมเข้าสู่เส้นทางการพัฒนาของดอกไม้ ความแปรผันในตำแหน่งเหล่านี้มีความเกี่ยวข้องกับความแปรผันของเวลาออกดอกระหว่างพืช ตัวอย่างเช่น พันธุ์ Arabidopsis thalianaที่เติบโตในเขตหนาวและ เขต อบอุ่นต้องการการกระตุ้นด้วยความเย็นเป็นเวลานานก่อนที่จะออกดอก ในขณะที่ พันธุ์ เขตร้อนและสายพันธุ์ในห้องปฏิบัติการทั่วไปไม่ต้องการ ความแปรผันนี้เกิดจากการกลายพันธุ์ใน ยีน FLCและFRIGIDAทำให้ยีนเหล่านี้ไม่ทำงาน[ 134 ]
ยีนจำนวนมากที่เกี่ยวข้องกับกระบวนการนี้ได้รับการอนุรักษ์ไว้ในพืชทุกชนิดที่ศึกษา อย่างไรก็ตาม บางครั้งแม้จะมีการอนุรักษ์ทางพันธุกรรม กลไกการทำงานกลับแตกต่างกัน ตัวอย่างเช่นข้าวเป็นพืชวันสั้น ในขณะที่Arabidopsis thalianaเป็นพืชวันยาว พืชทั้งสองชนิดมีโปรตีนCOและFLOWERING LOCUS T (FT)แต่ในArabidopsis thalianaนั้นCOช่วยเพิ่ม การผลิต FTในขณะที่ในข้าว โฮโมล็อก ของ COจะยับยั้ง การผลิต FTส่งผลให้เกิดผลกระทบที่ตรงกันข้ามอย่างสิ้นเชิง[ 135 ]
ทฤษฎีวิวัฒนาการของดอกไม้
ทฤษฎีแอนโธไฟต์นั้นอิงจากการสังเกตว่ากลุ่มพืชเมล็ดเปลือยGnetalesมีรังไข่ ที่มีลักษณะคล้ายดอกไม้ มีเส้นเลือด ที่พัฒนาบางส่วน เหมือนที่พบในพืชดอกและเมกะสปอ แรนเจียม ถูกหุ้มด้วยเยื่อหุ้มสามชั้น คล้ายกับ โครงสร้าง รังไข่ของดอกไม้พืชดอก อย่างไรก็ตาม หลักฐานอื่นๆ อีกมากมายแสดงให้เห็นว่า Gnetales ไม่มีความเกี่ยวข้องกับพืชดอก[ 136 ]
ทฤษฎี " ส่วนใหญ่เป็นเพศผู้"มีพื้นฐานทางพันธุกรรมมากกว่า ผู้สนับสนุนทฤษฎีนี้ชี้ให้เห็นว่าพืชเมล็ดเปลือยมีสำเนาของยีนLFY ที่คล้ายคลึงกันมากถึงสองชุด ในขณะที่พืชดอกมีเพียงชุดเดียว การวิเคราะห์ นาฬิกาโมเลกุล แสดงให้เห็นว่าพาราโลก LFYอีกชุดหนึ่งสูญหายไปในพืชดอกในช่วงเวลาเดียวกันกับที่ฟอสซิลดอกไม้มีจำนวนมาก ซึ่งบ่งชี้ว่าเหตุการณ์นี้อาจนำไปสู่การวิวัฒนาการของดอกไม้[ 137 ]ตามทฤษฎีนี้ การสูญเสียพาราโลกLFY ชุดหนึ่ง ทำให้ดอกไม้มีลักษณะเป็นเพศผู้มากขึ้น โดยที่ไข่ถูกแสดงออกในตำแหน่งที่ไม่เหมาะสม ไข่เหล่านี้ในตอนแรกทำหน้าที่ดึงดูดแมลงผสมเกสรแต่ในเวลาต่อมาอาจถูกรวมเข้ากับแกนกลางของดอกไม้
กลไกและตัวแปรในการวิวัฒนาการของรูปร่างพืช

แม้ว่าปัจจัยด้านสิ่งแวดล้อมจะมีส่วนสำคัญต่อการเปลี่ยนแปลงทางวิวัฒนาการ แต่ก็เป็นเพียงตัวกระตุ้นให้เกิดการคัดเลือกโดยธรรมชาติ เท่านั้น การเปลี่ยนแปลงเกิดขึ้นจากปรากฏการณ์ในระดับพันธุกรรม ได้แก่การกลายพันธุ์การจัดเรียงโครโมโซมใหม่ และ การเปลี่ยนแปลง ทางอีพีเจเนติกส์ แม้ว่า การกลายพันธุ์ประเภททั่วไปจะพบได้ในสิ่งมีชีวิตทั่วโลก แต่ในพืชนั้น มีกลไกอื่นๆ ที่ได้รับการระบุว่ามีความสำคัญอย่างยิ่ง
การเพิ่มจำนวนจีโนมเป็นปรากฏการณ์ที่ค่อนข้างพบได้บ่อยในวิวัฒนาการของพืช และส่งผลให้เกิดภาวะโพลีพลอยดีซึ่งเป็นลักษณะทั่วไปในพืช มีการประมาณการว่าอย่างน้อยครึ่งหนึ่ง (และอาจจะทั้งหมด) ของพืชเคยมีการเพิ่มจำนวนจีโนมในประวัติศาสตร์ของพวกมันการเพิ่มจำนวนจีโนม เกี่ยวข้องกับ การจำลองยีน ทำให้เกิดความซ้ำซ้อนทางหน้าที่ในยีนส่วนใหญ่ ยีนที่จำลองขึ้นมาอาจได้รับหน้าที่ใหม่ ไม่ว่าจะโดยการเปลี่ยนแปลงรูปแบบการแสดงออกหรือการเปลี่ยนแปลงกิจกรรม เชื่อกันว่าภาวะโพลีพลอยดีและการจำลองยีนเป็นหนึ่งในแรงผลักดันที่ทรงพลังที่สุดในวิวัฒนาการของรูปร่างพืช แม้ว่าจะไม่ทราบสาเหตุที่แน่ชัดว่าทำไม การเพิ่มจำนวน จีโนม จึงเป็นกระบวนการที่เกิดขึ้นบ่อยในพืช เหตุผลหนึ่งที่เป็นไปได้คือการผลิต สารเมตาบอไลต์ทุติยภูมิจำนวนมากในเซลล์พืช สารเหล่านี้บางส่วนอาจรบกวนกระบวนการแยกโครโมโซมตามปกติ ทำให้เกิดการจำลองจีโนม

เมื่อไม่นานมานี้ พบว่าพืชมี ตระกูล ไมโครอาร์เอ็นเอ จำนวนมาก ซึ่งได้รับการอนุรักษ์ไว้ในสายพันธุ์พืชหลายสายพันธุ์ เมื่อเปรียบเทียบกับสัตว์แม้ว่าจำนวนตระกูลไมโครอาร์เอ็นเอของพืชจะน้อยกว่าในสัตว์ แต่ขนาดของแต่ละตระกูลนั้นใหญ่กว่ามาก ยีน ไมโครอาร์เอ็นเอยังกระจายตัวอยู่ในจีโนมมากกว่าในสัตว์ ซึ่งยีนไมโครอาร์เอ็นเอจะกระจุกตัวอยู่มากกว่า มีการเสนอว่าตระกูลไมโครอาร์เอ็นเอเหล่านี้ขยายตัวโดยการจำลองแบบของบริเวณโครโมโซม[ 138 ] พบว่า ยีนไมโครอาร์เอ็นเอจำนวนมากที่เกี่ยวข้องกับการควบคุมการพัฒนาของพืช นั้น ได้รับการอนุรักษ์ไว้ค่อนข้างดีระหว่างพืชที่ศึกษา
การนำพืชมาปลูกเลี้ยง เช่นข้าวโพดข้าวข้าวบาร์เลย์ข้าวสาลีฯลฯ ถือเป็นแรงผลักดันสำคัญในการวิวัฒนาการของพืชเหล่านี้ งานวิจัยเกี่ยวกับต้นกำเนิดของข้าวโพดพบว่า ข้าวโพดเป็นพืชที่ได้รับการดัดแปลงมาจากพืชป่าในเม็กซิโกที่เรียกว่าเทโอซินเต (teosinte ) เทโอซินเตอยู่ในสกุลZea เช่นเดียวกับข้าวโพด แต่มี ช่อดอกขนาดเล็กมากมีฝักแข็ง 5-10 ฝัก และลำต้นแตกกิ่งก้านสาขามากและแผ่กว้าง

การผสมข้ามพันธุ์ระหว่างทีโอซินเต้สายพันธุ์หนึ่งกับข้าวโพดจะให้ลูกหลานที่เจริญพันธุ์ได้ซึ่งมีลักษณะฟีโนไทป์อยู่ระหว่างข้าวโพดและทีโอซินเต้ การวิเคราะห์ QTLยังเผยให้เห็นตำแหน่งยีนบางตำแหน่งที่เมื่อกลายพันธุ์ในข้าวโพดแล้วจะให้ลำต้นหรือฝักที่มีลักษณะคล้ายทีโอซินเต้ การวิเคราะห์ นาฬิกาโมเลกุลของยีนเหล่านี้ประมาณการว่าต้นกำเนิดของยีนเหล่านี้มีอายุราว 9,000 ปีที่แล้ว ซึ่งสอดคล้องกับบันทึกอื่นๆ เกี่ยวกับการปลูกข้าวโพด เชื่อกันว่ากลุ่มเกษตรกรกลุ่มเล็กๆ กลุ่มหนึ่งได้คัดเลือกทีโอซินเต้กลายพันธุ์ตามธรรมชาติที่มีลักษณะคล้ายข้าวโพดเมื่อประมาณ 9,000 ปีที่แล้วในเม็กซิโก และทำการคัดเลือกอย่างต่อเนื่องจนได้ต้นข้าวโพดที่เราคุ้นเคยในปัจจุบัน[ 139 ]
ดอกกะหล่ำที่กินได้เป็นพืชที่ได้รับการพัฒนาสายพันธุ์มาจากBrassica oleracea ซึ่งเป็นพืชป่า โดย ดอกกะหล่ำไม่มีช่อดอกที่หนาแน่นและไม่แตกต่างกันเรียกว่า "เคิร์ด" เหมือนกับดอกกะหล่ำ ดอกกะหล่ำมีการกลายพันธุ์เพียงจุดเดียวในยีนที่เรียกว่าCALซึ่งควบคุม การแบ่งตัว ของเนื้อเยื่อเจริญไปเป็น ช่อดอก การ กลายพันธุ์นี้ทำให้เซลล์ที่เนื้อเยื่อเจริญของดอกมีลักษณะที่ไม่แตกต่างกัน และแทนที่จะเจริญเติบโตเป็นดอกไม้เซลล์เหล่านั้นกลับเจริญเติบโตเป็นกลุ่มเซลล์เนื้อเยื่อเจริญของช่อดอกที่หนาแน่นและหยุดการเจริญเติบโต[ 140 ]การกลายพันธุ์นี้ได้รับการคัดเลือกผ่านการพัฒนาสายพันธุ์มาตั้งแต่สมัยจักรวรรดิ กรีก เป็นอย่างน้อย
วิวัฒนาการของวิถีการสังเคราะห์แสง

เส้นทางการเผาผลาญC4เป็นนวัตกรรมวิวัฒนาการล่าสุดที่มีค่าในพืช ซึ่งเกี่ยวข้องกับการเปลี่ยนแปลงการปรับตัวที่ซับซ้อนต่อสรีรวิทยาและรูปแบบ การ แสดงออกของยีน[ 141 ]
การสังเคราะห์แสง เป็นกระบวนการทางเคมีที่ซับซ้อนซึ่งอาศัย เอนไซม์และโคเอนไซม์หลายชนิดเอนไซม์RuBisCOมีหน้าที่ "ตรึง" CO2 กล่าวคือ มันจะนำ CO2 มาเชื่อมต่อกับโมเลกุลที่มีคาร์บอนเป็นองค์ประกอบเพื่อสร้างน้ำตาล ซึ่งพืชสามารถนำไปใช้ได้ และปล่อยโมเลกุลออกซิเจนออกมา อย่างไรก็ตาม เอนไซม์นี้มีประสิทธิภาพต่ำมาก และเมื่ออุณหภูมิแวดล้อมสูงขึ้น มันจะตรึงออกซิเจนแทน CO2 มากขึ้นเรื่อยๆในกระบวนการที่เรียกว่าการหายใจระดับแสง (photorespiration ) กระบวนการนี้สิ้นเปลืองพลังงาน เนื่องจากพืชต้องใช้พลังงานในการเปลี่ยนผลิตภัณฑ์จากการหายใจระดับแสงกลับไปเป็นรูปแบบที่สามารถทำปฏิกิริยากับCO2ได้
การรวมคาร์บอน
โดยทั่วไปแล้ว วิธีหลักสองวิธีในการเพิ่มความเข้มข้นของคาร์บอนไดออกไซด์ในพืช ได้แก่ 1) กลไกการเพิ่มความเข้มข้นของ CO2 ทางชีวเคมี (CCM) และ 2) กลไกการเพิ่มความเข้มข้นของ CO2 ทางชีวฟิสิกส์กลไก CCM ทางชีวเคมี เช่น การสังเคราะห์แสงแบบ C4 และ CAM จะเพิ่มความเข้มข้นของ CO2 โดยใช้เอนไซม์ฟอสโฟอีโนลไพรูเวตคาร์บอกซิเลสในการจับคาร์บอนอนินทรีย์เข้ากับน้ำตาลสี่คาร์บอนระดับกลาง ซึ่งสามารถเปลี่ยนกลับเป็น RuBP และ CO2 เพื่อการตรึงคาร์บอนในภายหลังโดย Rubisco ส่วนกลไก CCM ทางชีวฟิสิกส์ เช่นคาร์บอกซีโซมและ ไพรีนอยด์ จะเพิ่มความเข้มข้นของ CO2 ในตำแหน่งเฉพาะผ่านการทำงานร่วมกันของคาร์บอนิกแอน ไฮ เดรสและช่องแอนไอออน
พืช C4 พัฒนากลไกการรวมคาร์บอนโดยการเพิ่มความเข้มข้นของ CO2 รอบๆ RuBisCO และกีดกันออกซิเจนออกไป ซึ่งจะช่วยเพิ่มประสิทธิภาพของการสังเคราะห์แสงโดยลดการหายใจระดับเซลล์ (photorespiration) กระบวนการรวม CO2 รอบๆ RuBisCO นั้นต้องการพลังงานมากกว่าการปล่อยให้ก๊าซแพร่กระจายแต่ภายใต้เงื่อนไขบางอย่าง เช่น อุณหภูมิสูง (>25 °C) ความเข้มข้นของ CO2 ต่ำหรือความเข้มข้นของออกซิเจนสูง จะคุ้มค่าในแง่ของการลดการสูญเสียน้ำตาลจากการหายใจระดับเซลล์
กระบวนการเมตาบอลิซึมแบบ C4ชนิดหนึ่งใช้โครงสร้างที่เรียกว่าKranz anatomyซึ่งจะลำเลียง CO2 ผ่านชั้นมีโซฟิลล์ด้านนอก โดยอาศัยโมเลกุลอินทรีย์หลายชนิด ไปยังเซลล์หุ้มมัดท่อลำเลียงส่วนกลาง ซึ่ง CO2 จะถูกปล่อยออกมา ด้วยวิธีนี้ CO2 จะมีความเข้มข้นอยู่ใกล้บริเวณที่ RuBisCO ทำงาน เนื่องจาก RuBisCO ทำงานในสภาพแวดล้อมที่มี CO2 มากกว่าปกติจึงทำงานได้อย่างมีประสิทธิภาพมากขึ้น
กลไกที่สองคือการสังเคราะห์แสงแบบ CAM ซึ่งแยกการสังเคราะห์แสงออกจากการทำงานของ RuBisCO ในเชิงเวลา RuBisCO จะทำงานเฉพาะในเวลากลางวันเท่านั้น เมื่อปากใบปิดสนิทและมี CO2 เกิดขึ้นจากการสลายตัวของสารเคมี มา เลต จากนั้นจะมีการดึง CO2เพิ่มเติมจากบรรยากาศเมื่อปากใบเปิดออกในเวลากลางคืนที่อากาศเย็นและชื้น ซึ่งช่วยลดการสูญเสียน้ำ
กลไกที่สามที่มีอยู่ในพืช คือ CCM ที่ใช้ไพรีนอยด์ พบได้เฉพาะในสายพันธุ์ฮอร์นเวิร์ต เท่านั้น [ 142 ]ในกลไกนี้ RuBisCO จะมีความเข้มข้นในไพรีนอยด์ ซึ่งเป็นช่องที่ไม่มีเยื่อหุ้ม โดยการนำเข้าคาร์บอนอนินทรีย์ในรูปของไบคาร์บอเนต การนำเข้านี้เชื่อว่าขึ้นอยู่กับการประสานงานของคาร์บอนิกแอนไฮเดรสและช่องแอนไอออน และใช้ประโยชน์จากความแตกต่างของ pH ตามธรรมชาติระหว่างไซโตซอล สโตรมา ของคลอโรพลาสต์ และลูเมน ของไทลาคอย ด์
บันทึกวิวัฒนาการ
เส้นทางทั้งสองนี้ ซึ่งมีผลเช่นเดียวกันต่อ RuBisCO ได้วิวัฒนาการขึ้นหลายครั้งโดยอิสระ – อันที่จริง C4 เพียงอย่างเดียวเกิดขึ้นถึง 62 ครั้งในพืช 18 วงศ์ ที่แตกต่างกัน ดูเหมือนว่า 'การปรับตัวเบื้องต้น' หลายอย่างได้ปูทางให้กับ C4 ทำให้เกิดการรวมกลุ่มกันในกลุ่มย่อยบางกลุ่ม: โดยส่วนใหญ่แล้ว C4 มักเกิดขึ้นในพืชที่มีลักษณะเฉพาะอยู่แล้ว เช่น เนื้อเยื่อหุ้มมัดหลอดเลือดที่กว้างขวาง[ 143 ]เส้นทางวิวัฒนาการที่เป็นไปได้หลายเส้นทางซึ่งส่งผลให้เกิดฟีโนไทป์C4 นั้นสามารถระบุลักษณะได้โดยใช้การอนุมานแบบเบย์เซียน[ 141 ] ซึ่งยืนยันว่าการปรับตัวที่ไม่เกี่ยวข้องกับการสังเคราะห์แสงมักเป็นก้าวสำคัญทางวิวัฒนาการสำหรับการพัฒนา C4 ต่อไป
โครงสร้าง C4 ถูกใช้โดยหญ้าบางชนิด ในขณะที่ CAM ถูกใช้โดยพืชอวบน้ำและกระบองเพชร หลายชนิด ลักษณะ C4 ดูเหมือนจะเกิดขึ้นในช่วงโอลิโกซีนประมาณ25 ถึง 32ล้านปีก่อน[ 144 ] อย่างไรก็ตาม พวกมัน ไม่ได้มีความสำคัญทางนิเวศวิทยาจนกระทั่งถึงไมโอซีน 6 ถึง 7ล้านปีก่อน [ 145 ] ที่ น่าทึ่งคือ ฟอสซิลที่ กลาย เป็นถ่านบางส่วน ยังคงรักษาเนื้อเยื่อที่จัดเรียงเป็นโครงสร้าง Kranz โดยมีเซลล์ปลอกมัดที่สมบูรณ์[ 146 ]ทำให้สามารถระบุการมีอยู่ของการเผาผลาญ C4 ได้ เครื่องหมายไอโซโทปถูกใช้เพื่ออนุมานการกระจายและความสำคัญของพวกมัน พืช C3 เลือกใช้ไอโซโทป คาร์บอน ที่เบากว่าในบรรยากาศ คือ12Cซึ่งมีส่วนร่วมในเส้นทางเคมีที่เกี่ยวข้องกับการตรึงได้ง่ายกว่า เนื่องจากกระบวนการเผาผลาญ C4 เกี่ยวข้องกับขั้นตอนทางเคมีเพิ่มเติม ผลกระทบนี้จึงเด่นชัดขึ้น สามารถวิเคราะห์ วัสดุจากพืช เพื่อหาอัตราส่วนของคาร์บอน-13 ที่หนักกว่า ต่อคาร์บอน-12ได้ อัตราส่วนนี้เรียกว่าδ 13 Cพืช C3 มี น้ำหนักเบากว่าอัตราส่วนในบรรยากาศโดยเฉลี่ยประมาณ 14‰ (ส่วนต่อพัน) ในขณะที่พืช C4 มีน้ำหนักเบากว่าประมาณ 28‰ ค่าδ 13 Cพืช CAM ขึ้นอยู่กับเปอร์เซ็นต์ของคาร์บอนที่ตรึงในเวลากลางคืนเมื่อเทียบกับคาร์บอนที่ตรึงในเวลากลางวัน โดยจะใกล้เคียงกับพืช C3 หากตรึงคาร์บอนส่วนใหญ่ในเวลากลางวัน และจะใกล้เคียงกับพืช C4 หากตรึงคาร์บอนทั้งหมดในเวลากลางคืน[ 147 ]
ซากดึกดำบรรพ์ดั้งเดิมที่มีปริมาณเพียงพอสำหรับการวิเคราะห์หญ้าโดยตรงนั้นหายาก แต่ซากม้าเป็นตัวแทนที่ดี พวกมันแพร่หลายไปทั่วโลกในช่วงเวลาที่สนใจ และกินหญ้าเป็นอาหารเกือบทั้งหมด มีวลีเก่าแก่ในบรรพชีวินวิทยาไอโซโทปว่า "คุณคือสิ่งที่คุณกิน (บวกอีกเล็กน้อย)" – ซึ่งหมายถึงข้อเท็จจริงที่ว่าสิ่งมีชีวิตสะท้อนองค์ประกอบไอโซโทปของสิ่งที่พวกมันกิน บวกกับปัจจัยการปรับเล็กน้อย มีบันทึกที่ดีเกี่ยวกับฟันม้าทั่วโลก และค่า δ 13 C ของพวกมันบันทึกแสดงให้เห็นการเปลี่ยนแปลงเชิงลบอย่างรวดเร็วเมื่อประมาณ6 ถึง 7ล้านปีก่อนในช่วงยุคเมสซิเนียน ซึ่งตีความได้ว่าเป็นผลมาจากการเพิ่มขึ้นของพืช C4 ในระดับโลก[ 145 ]
ข้อดีของ C 4
แม้ว่า C4 จะช่วยเพิ่มประสิทธิภาพของ RuBisCO แต่ความเข้มข้นของคาร์บอนนั้นต้องการพลังงานสูงมาก ซึ่งหมายความว่าพืช C4 จะได้เปรียบสิ่งมีชีวิต C3 ในบางสภาวะเท่านั้นกล่าวคือ อุณหภูมิสูงและปริมาณน้ำฝนต่ำ พืช C4 ยังต้องการแสงแดดในระดับสูงเพื่อเจริญเติบโต[ 148 ]แบบจำลองชี้ให้เห็นว่า หากไม่มีไฟป่าที่กำจัดต้นไม้และพุ่มไม้ที่ให้ร่มเงา จะไม่มีพื้นที่สำหรับพืชC4 [ 149 ]แต่ไฟป่าเกิดขึ้นมาแล้ว 400 ล้านปี ยุคคาร์บอนิเฟอรัส (ประมาณ300 ล้านปีก่อน ) มีระดับออกซิเจนสูงมาก เกือบจะเพียงพอที่จะทำให้เกิดการเผาไหม้เองได้[ 150 ] และมี CO2 ต่ำมากแต่ไม่พบร่องรอยไอโซโทปของ C4 นอกจากนี้ดูเหมือนว่าจะไม่มีตัวกระตุ้นที่ฉับพลันสำหรับการเพิ่มขึ้นของไมโอ ซีน
ในช่วงยุคไมโอซีน บรรยากาศและสภาพภูมิอากาศค่อนข้างคงที่ หากจะมีอะไรเปลี่ยนแปลง CO2 ก็เพิ่มขึ้นอย่างค่อยเป็นค่อยไปตั้งแต่14 ถึง 9ล้านปีก่อนก่อนที่จะคงที่ในระดับความเข้มข้นที่คล้ายกับยุคโฮโลซีน[ 151 ]ซึ่งแสดงให้เห็นว่ามันไม่ได้มีบทบาทสำคัญในการกระตุ้นวิวัฒนาการ ของ C4 [ 144 ]หญ้าเอง (กลุ่มที่ก่อให้เกิด C4 มากที่สุด)น่าจะมีอยู่มานานกว่า 60 ล้านปีแล้ว ดังนั้นจึงมีเวลามากพอที่จะวิวัฒนาการ C4 [ 152 ] [ 153 ] ซึ่ง ในกรณีใดๆ ก็ตาม C4มีอยู่ในกลุ่มที่หลากหลายและวิวัฒนาการขึ้นอย่างอิสระ มีสัญญาณที่ชัดเจนของการเปลี่ยนแปลงสภาพภูมิอากาศในเอเชียใต้[ 144 ]ความแห้งแล้งที่เพิ่มขึ้น – ดังนั้นความถี่และความรุนแรงของไฟป่าที่เพิ่มขึ้น – อาจนำไปสู่ความสำคัญของทุ่งหญ้าที่เพิ่มขึ้น[ 154 ]อย่างไรก็ตาม สิ่งนี้ยากที่จะสอดคล้องกับบันทึกของอเมริกาเหนือ[ 144 ]เป็นไปได้ว่าสัญญาณดังกล่าวเป็นผลทางชีวภาพทั้งหมด ซึ่งเกิดจากไฟ[ 155 ]ที่เร่งการวิวัฒนาการของหญ้า ซึ่งทั้งเพิ่มการผุกร่อนและรวมคาร์บอนเข้าไปในตะกอนมากขึ้น ทำให้ระดับ CO 2 ในบรรยากาศลดลง [ 155 ]สุดท้าย มีหลักฐานว่าการเริ่มต้นของ C 4ตั้งแต่9 ถึง 7ล้านปีก่อนเป็นสัญญาณที่มีอคติ ซึ่งเป็นจริงเฉพาะในทวีปอเมริกาเหนือเท่านั้น เนื่องจากตัวอย่างส่วนใหญ่มาจากที่นั่น หลักฐานที่เกิดขึ้นใหม่ชี้ให้เห็นว่าทุ่งหญ้าวิวัฒนาการไปสู่สถานะที่โดดเด่นอย่างน้อย 15 ล้านปีก่อนในทวีปอเมริกาใต้
วิวัฒนาการของการควบคุมการถอดรหัสทางพันธุกรรม
ปัจจัยการถอดรหัสและเครือข่ายควบคุมการถอดรหัสมีบทบาทสำคัญในการพัฒนาและการตอบสนองต่อความเครียดของพืช รวมถึงวิวัฒนาการของพืชด้วย ในระหว่างการลงจอดของพืช ตระกูลปัจจัยการถอดรหัสใหม่ ๆ จำนวนมากได้เกิดขึ้นและถูกเชื่อมโยงเข้ากับเครือข่ายการพัฒนาแบบหลายเซลล์ การสืบพันธุ์ และการพัฒนาอวัยวะ ซึ่งมีส่วนช่วยให้พืชบกมีรูปร่างที่ซับซ้อนมากขึ้น[ 156 ]
วิวัฒนาการของเมตาบอลิซึมรอง

เมตาโบไลต์รอง เป็นสารประกอบที่ มีน้ำหนักโมเลกุลต่ำบางครั้งมีโครงสร้างที่ซับซ้อน ซึ่งไม่จำเป็นต่อกระบวนการเจริญเติบโต การพัฒนาหรือการสืบพันธุ์ ตามปกติ พวกมันทำหน้าที่ในกระบวนการที่หลากหลาย เช่นภูมิคุ้มกันการป้องกันการถูกสัตว์ กินพืชกัดกิน การดึงดูดแมลงผสมเกสรการสื่อสารระหว่างพืช การรักษา ความสัมพันธ์ แบบพึ่งพาอาศัยกันกับจุลินทรีย์ในดิน หรือการเพิ่มอัตราการปฏิสนธิดังนั้นจึงมีความสำคัญจากมุมมองด้านวิวัฒนาการและการพัฒนา เมตาโบไลต์รองมีความหลากหลายทั้งในด้านโครงสร้างและหน้าที่ และคาดว่าเอนไซม์หลายแสนชนิดอาจเกี่ยวข้องกับกระบวนการผลิตพวกมัน โดยประมาณ 15–25% ของจีโนมเข้ารหัสเอนไซม์เหล่านี้ และแต่ละสปีชีส์มีเมตาโบไลต์รองที่เป็นเอกลักษณ์เฉพาะตัว[ 157 ]เมตาโบไลต์เหล่านี้หลายชนิด เช่นกรดซาลิไซลิกมีความสำคัญทางการแพทย์ต่อมนุษย์
จุดประสงค์ของการผลิตเมตาโบไลต์รองจำนวนมาก โดยมีสัดส่วนที่สำคัญของเมตาโบโลมที่อุทิศให้กับกิจกรรมนี้ยังไม่ชัดเจน มีการตั้งสมมติฐานว่าสารเคมีเหล่านี้ส่วนใหญ่ช่วยในการสร้างภูมิคุ้มกัน และผลที่ตามมาคือความหลากหลายของเมตาโบไลต์เหล่านี้เป็นผลมาจากการแข่งขัน อย่างต่อเนื่อง ระหว่างพืชและปรสิตของพวกมัน มีหลักฐานบางอย่างที่สนับสนุนกรณีนี้ คำถามสำคัญเกี่ยวข้องกับต้นทุนการสืบพันธุ์ในการรักษาคลังยีนจำนวนมากที่อุทิศให้กับการผลิตเมตาโบไลต์รอง มีการเสนอแบบจำลองต่างๆ ที่ตรวจสอบแง่มุมนี้ของคำถาม แต่ยังไม่มีข้อสรุปเกี่ยวกับขอบเขตของต้นทุน[ 158 ]เนื่องจากยังคงยากที่จะคาดการณ์ว่าพืชที่มีเมตาโบไลต์รองมากขึ้นจะเพิ่มการอยู่รอดหรือความสำเร็จในการสืบพันธุ์เมื่อเทียบกับพืชอื่นๆ ในบริเวณใกล้เคียงหรือไม่
การผลิตเมตาโบไลต์รองดูเหมือนจะเกิดขึ้นค่อนข้างเร็วในช่วงวิวัฒนาการ ในพืช ดูเหมือนว่าเมตาโบไลต์รองจะแพร่กระจายออกไปโดยใช้กลไกต่างๆ รวมถึงการจำลองยีนหรือวิวัฒนาการของยีนใหม่ นอกจากนี้ การวิจัยยังแสดงให้เห็นว่าความหลากหลายในสารประกอบบางชนิดเหล่านี้อาจได้รับการคัดเลือกในเชิงบวก แม้ว่าบทบาทของวิวัฒนาการของยีนใหม่ในวิวัฒนาการของเมตาโบลิซึมรองจะชัดเจน แต่ก็มีตัวอย่างหลายอย่างที่เมตาโบไลต์ใหม่เกิดขึ้นจากการเปลี่ยนแปลงเล็กน้อยในปฏิกิริยา ตัวอย่างเช่น มีการเสนอว่า ไซยาโนเจนไกลโคไซด์มีวิวัฒนาการหลายครั้งในสายพันธุ์พืชที่แตกต่างกัน มีกรณีวิวัฒนาการแบบลู่เข้า หลายกรณี ตัวอย่างเช่น เอนไซม์สำหรับการสังเคราะห์ลิโมนีน ซึ่งเป็นเทอร์พีน มีความคล้ายคลึงกันระหว่างพืชดอกและพืชเมล็ดเปลือยมากกว่าเอนไซม์สังเคราะห์เทอร์พีนของพวกมันเอง สิ่งนี้ชี้ให้เห็นถึงวิวัฒนาการที่เป็นอิสระของเส้นทางการสังเคราะห์ลิโมนีนในสองสายพันธุ์นี้[ 159 ]
วิวัฒนาการของปฏิสัมพันธ์ระหว่างพืชและจุลินทรีย์
กำเนิดของจุลินทรีย์บนโลก ย้อนกลับไปถึงจุดเริ่มต้นของสิ่งมีชีวิตเมื่อกว่า 3.5 พันล้านปีก่อน บ่งชี้ว่าปฏิสัมพันธ์ระหว่างจุลินทรีย์ได้มีการวิวัฒนาการและมีความหลากหลายอย่างต่อเนื่องตลอดเวลา ก่อนที่พืชจะเริ่มเข้ามาอาศัยบนบกเมื่อ 450 ล้านปีก่อน ดังนั้น จึงเป็นไปได้ว่าปฏิสัมพันธ์ระหว่างจุลินทรีย์ทั้งภายในและระหว่างอาณาจักรเป็นตัวขับเคลื่อนที่สำคัญของการก่อตั้งกลุ่มจุลินทรีย์ ที่เกี่ยวข้องกับพืช ที่บริเวณรอยต่อระหว่างดินและราก อย่างไรก็ตาม ยังไม่ชัดเจนว่าปฏิสัมพันธ์เหล่านี้ในไรโซสเฟียร์/ฟิลโลสเฟียร์และในส่วนของพืชที่อยู่ภายใน (เช่น ภายในโฮสต์) มีส่วนในการกำหนดกลุ่มจุลินทรีย์ในธรรมชาติมากน้อยเพียงใด และการปรับตัวของจุลินทรีย์ให้เข้ากับถิ่นที่อยู่ของพืชเป็นตัวขับเคลื่อนกลยุทธ์ปฏิสัมพันธ์ระหว่างจุลินทรีย์ที่เฉพาะเจาะจงกับถิ่นที่อยู่ซึ่งส่งผลต่อความแข็งแรงของพืชหรือไม่ นอกจากนี้ การมีส่วนร่วมของปฏิสัมพันธ์ระหว่างจุลินทรีย์แบบแข่งขันและแบบร่วมมือกันต่อโครงสร้างชุมชนโดยรวมยังคงประเมินได้ยากในธรรมชาติเนื่องจากมีสัญญาณรบกวนในสิ่งแวดล้อมสูง[ 160 ]
ดูเพิ่มเติม
- วิวัฒนาการของการกินพืช
- ประวัติวิวัฒนาการของสิ่งมีชีวิต
- บรรพพฤกษศาสตร์
- ชีววิทยาการพัฒนาเชิงวิวัฒนาการของพืช
- ลำดับเหตุการณ์วิวัฒนาการของพืช
ลิงก์ภายนอก
- วิวัฒนาการและพฤกษศาสตร์ โบราณ ที่บริแทนนิกา
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ประวัติวิวัฒนาการของพืช
วิวัฒนาการของพืชส่งผลให้เกิดความซับซ้อนที่หลากหลาย ตั้งแต่แผ่นสาหร่าย ยุคแรกสุด ของอาร์คีพลาส ติดเซลล์เดียวที่ วิวัฒนาการผ่านเอนโดซิมไบโอซิสไปจนถึงสาหร่ายสีเขียวในทะเลและน้ำจืด...
ความแตกต่าง
พืชบก วิวัฒนาการมาจากกลุ่ม สาหร่ายสีเขียว น้ำจืด อาจจะเร็วที่สุดเมื่อ 850 ล้านปีก่อน [ 3 ] แต่พืชที่มีลักษณะคล้ายสาหร่ายอาจวิวัฒนาการมาตั้งแต่ 1 พันล้านปีก่อน [ 2 ] ญาติที่ใกล้ชิดที่สุดของพืชบกในปัจจุบันคือ ชาโรไฟต์ โดยเฉพาะอย่างยิ่ง Charales หาก Charales...
ความท้าทายต่อการตั้งถิ่นฐานบนที่ดิน
พืชไม่ใช่สิ่งมีชีวิตสังเคราะห์แสงกลุ่มแรกบนบก อัตราการผุกร่อนบ่งชี้ว่าสิ่งมีชีวิตที่สามารถสังเคราะห์แสงได้นั้นอาศัยอยู่บนบกมา ตั้งแต่ 1,200 ล้านปีก่อน [ 7 ] และ พบ ฟอสซิล จุลินทรีย์ในตะกอนทะเลสาบน้ำจืดตั้งแต่ 1,000 ล้านปีก่อน [ 10 ] แต่ บันทึกไอโซโทปคาร์บอน...
การปรับตัวเพื่อการตั้งถิ่นฐานบนบก
หลักฐานของพืชบกยุคแรกสุดปรากฏขึ้นเมื่อประมาณ 470 ล้านปีก่อน ในหินยุคออร์โดวิเชียนตอนกลางตอนล่างจาก ซาอุดีอาระเบีย [ 16 ] และ กอนด์วานา [ 17 ] ในรูปแบบของสปอร์ที่เรียกว่า คริปโตสปอร์ สปอร์เหล่านี้มีผนังที่ทำจาก สปอโรพอลเลนิน...