อ่าน 88 นาที
ไทแรนโนซอรัส
ไทแรนโนซอรัส ( / t ɪ ˌ r æ n ə ˈ s ɔː r ə s , t aɪ - / ) [ a ] เป็น สกุล ของ ไดโนเสาร์ เทอ โรพอด ขนาดใหญ่ ไทแรนโนซอรัสเร็ก ซ์ ( เร็กซ์ แปลว่า 'ราชา' ใน ภาษาละติน )...
ไทแรนโนซอรัส
| ไทแรนโนซอรัส | |
|---|---|
 | |
| การจำลองแบบตัวอย่างต้นแบบ ของ ทีเร็กซ์ ที่พิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติคาร์เนกี | |
| การจำแนกทางวิทยาศาสตร์ | |
| อาณาจักร: | แอนิมอลเลีย |
| ไฟลัม: | คอร์ดาต้า |
| ระดับ: | สัตว์เลื้อยคลาน |
| กลุ่มสายพันธุ์ : | ไดโนเสาร์ |
| กลุ่มสายพันธุ์ : | ซอริสเชีย |
| กลุ่มสายพันธุ์ : | เทโรโปดา |
| ซูเปอร์แฟมิลี่: | † ไทแรนโนซอรอยเดีย |
| ตระกูล: | † ไทแรนโนซอริเด |
| เผ่า: | † ไทแรนโนซอรินี |
| ประเภท: | † ไทแรนโนซอรัสออสบอร์น , 1905 |
| ชนิดต้นแบบ | |
| † ไทแรนโนซอรัส เร็กซ์ ออสบอร์น, 1905 | |
| สายพันธุ์อื่นๆ | |
| คำพ้องความหมาย | |
คำพ้องความหมายของสกุล
ความเหมือนกันของสายพันธุ์
| |
ไทแรนโนซอรัส ( / t ɪ ˌ r æ n ə ˈ s ɔː r ə s , t aɪ - / ) [ a ] เป็นสกุลของ ไดโนเสาร์ เทอโรพอด ขนาดใหญ่ ไทแรนโนซอรัสเร็กซ์ (เร็กซ์แปลว่า 'ราชา' ในภาษาละติน ) ซึ่งมักย่อเป็น T. rexหรือเรียกกันทั่วไปว่า T-Rexเป็นหนึ่งในเทอโรพอดที่มีการค้นพบฟอสซิลมากที่สุด มันอาศัยอยู่ทั่วบริเวณที่ปัจจุบันคืออเมริกาเหนือ ตะวันตก ซึ่งในสมัยนั้นเป็นทวีปเกาะที่รู้จักกันในชื่อลารามิ เดีย ไทแรนโนซอรัสมีถิ่นที่อยู่กว้างกว่าไทแรนโนซอริเด ชนิดอื่นๆ ฟอสซิล ของมันพบได้ใน ชั้นหินทางธรณีวิทยาที่หลากหลายซึ่งมีอายุย้อนไปถึง ยุค มาสทริชเชียน ตอนปลาย ของ ยุค ครีเทเชียสประมาณ 69 ถึง 66 ล้านปีก่อนโดยมีตัวอย่างที่แยกออกมาซึ่งอาจบ่งชี้ถึงต้นกำเนิดที่เก่ากว่าในยุคแคมพาเนียน ตอนกลาง มันเป็นหนึ่งในสมาชิกกลุ่มไทแรนโนซอริเดียกลุ่มสุดท้ายที่รู้จัก และเป็นหนึ่งในไดโนเสาร์ที่ไม่ใช่ นก กลุ่ม สุดท้าย ที่ยังมีชีวิตอยู่ก่อนเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในยุคครีเทเชียส-พาลีโอจีน
เช่นเดียวกับไดโนเสาร์วงศ์ไทแรนโน ซอรัสชนิดอื่นๆ ไทแรนโนซอรัส เป็น สัตว์กิน เนื้อ สองขาที่มีกะโหลก ขนาดใหญ่สมดุลด้วยหางที่ยาวและหนัก เมื่อเทียบกับขาหลังที่ใหญ่และแข็งแรง ขาหน้าของไทแรนโนซอรัสกลับสั้นแต่แข็งแรงผิดปกติเมื่อเทียบกับขนาด และมีนิ้วที่มีเล็บสองนิ้ว ตัวอย่างที่สมบูรณ์ที่สุดมีความยาว 12.3–12.4 เมตร (40–41 ฟุต) แต่จากการประมาณการในปัจจุบันส่วนใหญ่ไทแรนโนซอรัสอาจมีขนาดใหญ่กว่า 13 เมตร (43 ฟุต) ความสูงที่สะโพก 3.7–4 เมตร (12–13 ฟุต) และน้ำหนัก 8.8 ตัน (8.7 ตันยาว; 9.7 ตันสั้น) แม้ว่าไดโนเสาร์เทอโรพอดชนิดอื่นๆ อาจมีขนาดใกล้เคียงหรือใหญ่กว่าไทแรนโนซอ รัส แต่ไทแรนโนซอรัส ก็ยังคงเป็นหนึ่งในสัตว์นักล่าบนบกที่ใหญ่ที่สุดเท่าที่รู้จัก โดยมีแรงกัดที่คาดการณ์ไว้ว่ามากที่สุดในบรรดาสัตว์บกทั้งหมดไทแรนโนซอรัส เร็กซ์เป็นสัตว์กินเนื้อที่ใหญ่ที่สุดในสภาพแวดล้อมของมัน และ น่าจะเป็น ผู้ล่าสูงสุดโดย ล่าไดโนเสาร์กลุ่ม ฮาโดร ซอร์ สัตว์กินพืชหุ้มเกราะวัยอ่อนอย่างเซราทอปเซียนและแอนคิโลซอร์และอาจรวมถึงซอโรพอดด้วย อย่างไรก็ตาม ผู้เชี่ยวชาญบางคนเสนอว่าไดโนเสาร์ชนิดนี้เป็นสัตว์กินซาก เป็นหลัก คำถามที่ว่าไทแรนโนซอรัสเป็นผู้ล่าสูงสุดหรือเป็นเพียงสัตว์กินซากนั้น เป็นหนึ่งในข้อถกเถียงที่ยาวนานที่สุดในวงการบรรพชีวินวิทยาปัจจุบันนักบรรพชีวินวิทยาส่วนใหญ่ยอมรับว่าไทแรนโนซอรัสเป็นทั้งผู้ล่าและสัตว์กินซาก
ซากดึกดำบรรพ์ของไทแรนโนซอรัส เร็กซ์บางชิ้นมีโครงกระดูกที่เกือบสมบูรณ์มีการพบเนื้อเยื่ออ่อนและโปรตีน ในซากดึกดำบรรพ์อย่างน้อยหนึ่งชิ้น ความอุดมสมบูรณ์ของซากดึกดำบรรพ์ทำให้สามารถทำการวิจัยอย่างมีนัยสำคัญในหลายแง่มุมทางชีววิทยาของสัตว์ชนิดนี้ รวมถึงวงจรชีวิตและ กลไกทางชีวภาพพฤติกรรมการกินสรีรวิทยาและความเร็วที่เป็นไปได้ของไทแรนโนซอรัส เร็กซ์เป็นหัวข้อที่ถกเถียง กันอยู่ การจำแนกทางอนุกรมวิธาน ของมันก็ยังเป็นที่ถกเถียงเช่นกัน ทาร์โบซอ รัส บาตาร์จากเอเชียมีความสัมพันธ์ใกล้ชิดกับไทแรนโนซอรัส มาก และบางครั้งก็ถูกมองว่าเป็นสายพันธุ์หนึ่งในสกุลนี้ ไทแรนโนซอริเดหลายชนิดในอเมริกาเหนือถูกจัดให้เป็นชื่อพ้องกับไทแรนโนซอรัสในขณะที่ ซากดึกดำบรรพ์ของ ไทแรนโนซอรัส บางชิ้นถูกเสนอให้เป็นสายพันธุ์ที่แตกต่างกัน ความถูกต้องของสายพันธุ์เหล่านี้ เช่น T. mcraeensisที่เพิ่งค้นพบเมื่อไม่นานมานี้ยังคงเป็นที่ถกเถียงกันอยู่
ไทแรนโนซอรัสเป็นหนึ่งในไดโนเสาร์ที่รู้จักกันดีที่สุดนับตั้งแต่ต้นศตวรรษที่ 20 ไรลีย์ แบล็ก นักเขียนด้านวิทยาศาสตร์ เรียกมันว่า "สุดยอดไดโนเสาร์" ฟอสซิลของมันเป็นที่ดึงดูดใจในพิพิธภัณฑ์ต่างๆ และมันยังปรากฏในสื่อต่างๆ เช่นจูราสสิก พาร์ค
ประวัติการวิจัย
การค้นพบที่เก่าแก่ที่สุด

ฟันของสิ่งที่ปัจจุบันได้รับการบันทึกว่าเป็นไทแรนโนซอรัส เร็กซ์ถูกค้นพบในเดือนกรกฎาคม ค.ศ. 1874 บนภูเขาเซาท์เทเบิล (โคโลราโด)โดยปีเตอร์ ที. ดอตสัน นักศึกษาจากวิทยาลัยจาร์วิสฮอลล์ (โคโลราโด)ภายใต้การดูแลของอาร์เธอร์ เลคส์ใกล้เมืองโกลเดน รัฐ โคโลราโดในตะกอนที่เป็นของกลุ่มหินเดนเวอร์[ 1 ] [ 2 ]ในช่วงต้นทศวรรษ ค.ศ. 1890 จอห์ น เบลล์ แฮทเชอร์ได้รวบรวมชิ้นส่วนกระดูกส่วนลำตัวในไวโอมิง ตะวันออก ฟอสซิลเหล่านั้นเชื่อกันว่าเป็นของสายพันธุ์ขนาดใหญ่Ornithomimus grandis (ปัจจุบันคือ Deinodon ) แต่ปัจจุบันถือว่าเป็นซากของที. เร็ก ซ์ [ 3 ]
ในปี ค.ศ. 1892 เอ็ดเวิร์ด ดริงเกอร์ โคปพบชิ้นส่วนกระดูกสันหลังสองชิ้นของไดโนเสาร์ขนาดใหญ่ โคปเชื่อว่าชิ้นส่วนเหล่านั้นเป็นของไดโนเสาร์ "อะกาธาอุมิด" ( เซราทอปซิด ) และตั้งชื่อว่ามาโนสปอนดิลัส กิกัสซึ่งหมายถึง "กระดูกสันหลังที่มีรูพรุนขนาดยักษ์" โดยอ้างอิงถึงช่องเปิดจำนวนมากสำหรับหลอดเลือดที่เขาพบในกระดูก[ 3 ] ซาก ของM. gigasถูกระบุโดยแฮทเชอร์ในปี ค.ศ. 1907 ว่าเป็นของเทโรพอด ไม่ใช่เซราทอปซิด[ 4 ]
เฮนรี แฟร์ฟิลด์ ออสบอร์นตระหนักถึงความคล้ายคลึงกันระหว่างManospondylus gigasและT. rexตั้งแต่ปี 1917 ซึ่งในเวลานั้นกระดูกสันหลังชิ้นที่สองได้หายไปแล้ว เนื่องจากกระดูกสันหลังของManospondylus มีลักษณะเป็นชิ้นส่วน ออสบอร์นจึงไม่ได้จัดให้ทั้งสองสกุลเป็นชื่อพ้อง แต่กลับพิจารณาว่าสกุลที่เก่ากว่านั้นยังไม่สามารถระบุได้[ 5 ]ในเดือนมิถุนายน ปี 2000 สถาบันแบล็กฮิลส์ ได้ค้นพบโครงกระดูกของ ไทแรนโนซอรัสประมาณ 10% ( BHI 6248) ในสถานที่ซึ่งอาจเป็นแหล่งที่ พบ M. gigas ดั้งเดิม [ 6 ]ในขณะที่ลาร์สันในตอนแรกพิจารณาว่าตัวอย่างนี้เป็นสัตว์ตัวเดียวกันกับ ตัวอย่างต้นแบบของ M. gigasและแนะนำว่าควรให้ความสำคัญมากกว่าT. rexนักวิจัยคนอื่นๆ รวมถึงคริสโตเฟอร์ โบรชู ยังคงไม่แน่ใจว่าชื่อT. rexจะถูกละทิ้งหรือไม่[ 7 ]ในปี 2546 Brochu ได้โต้แย้งเพิ่มเติมว่า แม้ว่าแท็กซาทั้งสองน่าจะแสดงถึงสปีชีส์เดียวกัน แต่เป็นไปไม่ได้ที่จะพิสูจน์ว่าตัวอย่างที่ค้นพบโดยสถาบัน Black Hills แสดงถึงสัตว์ตัวเดียวกันกับM. gigasเนื่องจากการเก็บรักษาตัวอย่างต้นแบบของM. gigas ที่ไม่ดี นอกจากนี้ ตาม การแก้ไขประมวล กฎการตั้งชื่อสัตว์สากล ปี 2543 ชื่อที่ใช้กันทั่วไป เช่นT. rexมีลำดับความสำคัญเหนือกว่าชื่อที่มีลำดับความสำคัญทางเทคนิค แต่ไม่ได้ใช้เป็นชื่อที่ถูกต้องในเอกสารทางการหลังจากปี 1899 เป็นเวลากว่าห้าสิบปี เช่นM. gigas [ 8 ] [ 9 ]
การค้นพบและการตั้งชื่อโครงกระดูก

บาร์นัม บราวน์ผู้ช่วยภัณฑารักษ์ของพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติอเมริกันพบโครงกระดูกบางส่วนของทีเร็กซ์ เป็นครั้งแรก ในไวโอมิงตะวันออกในปี 1900 บราวน์พบโครงกระดูกบางส่วนอีกชิ้นหนึ่งในแหล่งหินเฮลล์ครีกในมอนแทนาในปี 1902 ซึ่งประกอบด้วยกระดูกฟอสซิลประมาณ 34 ชิ้น[ 10 ] บราวน์เขียนไว้ในขณะนั้นว่า "เหมืองหมายเลข 1 ประกอบด้วยกระดูกต้นขา กระดูกเชิงกราน กระดูกต้นแขน กระดูกสันหลัง 3 ชิ้น และกระดูกอีก 2 ชิ้นที่ไม่สามารถระบุได้ของไดโนเสาร์กินเนื้อขนาดใหญ่ที่ มาร์ชไม่ได้บรรยายไว้... ผมไม่เคยเห็นอะไรแบบนี้มาก่อนจากยุคครีเทเชียส " [ 11 ]เฮนรี แฟร์ฟิลด์ ออสบอร์น ประธานพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติอเมริกันได้ตั้งชื่อโครงกระดูกที่สองว่าT. rexในปี 1905 ชื่อสกุลมาจากคำภาษากรีกτύραννος ( tyrannosแปลว่า "ทรราช") และσαῦρος ( saurosแปลว่า "กิ้งก่า") ออสบอร์นใช้คำภาษาละตินrexซึ่งแปลว่า "ราชา" สำหรับชื่อเฉพาะ ดังนั้น ชื่อวิทยาศาสตร์ แบบเต็ม จึงแปลว่า "กิ้งก่าทรราชราชา" หรือ "กิ้งก่าทรราชราชา" ซึ่งเน้นขนาดของสัตว์และอำนาจที่สันนิษฐานว่าเหนือกว่าสายพันธุ์อื่นในสมัยนั้น[ 10 ]

Osborn ตั้งชื่อตัวอย่างอีกชิ้นหนึ่งว่า Dynamosaurus imperiosusในบทความเมื่อปี พ.ศ. 2448 [ 10 ]ในปี พ.ศ. 2449 Osborn ตระหนักว่าโครงกระดูกทั้งสองมาจากสายพันธุ์เดียวกัน และเลือกTyrannosaurusเป็นชื่อที่ต้องการ[ 12 ]ในปี พ.ศ. 2484 ตัวอย่างต้นแบบของ T. rexถูกขายให้กับพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติคาร์เนกีในเมืองพิตต์สเบิร์ก รัฐเพนซิลเวเนีย ในราคา 7,000 ดอลลาร์ (เทียบเท่ากับ 153,224 ดอลลาร์ในปี พ.ศ. 2568) [ 11 ] ปัจจุบันวัสดุ Dynamosaurusดั้งเดิมอยู่ในคอลเลกชันของพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติแห่งลอนดอน[ 13 ] ต่อมา Dynamosaurusได้รับการยกย่องจากการบรรยายลักษณะของไดโนเสาร์วงศ์ Tyrannosauridae อีกสายพันธุ์หนึ่งในปี พ.ศ. 2561 โดย Andrew McDonald และเพื่อนร่วมงาน ในชื่อ Dynamoterror dynastesซึ่งชื่อนี้ถูกเลือกโดยอ้างอิงจากชื่อในปี พ.ศ. 2448 เนื่องจากเป็น "ชื่อโปรดในวัยเด็ก" ของ McDonald [ 14 ]
ตั้งแต่ช่วงทศวรรษ 1910 จนถึงปลายทศวรรษ 1950 การค้นพบของบาร์นัมยังคงเป็นตัวอย่างเดียวของไทแรนโนซอรัสเนื่องจากภาวะเศรษฐกิจตกต่ำครั้งใหญ่และสงครามทำให้บรรดานักบรรพชีวินวิทยาจำนวนมากไม่สามารถออกไปสำรวจภาคสนามได้[ 6 ]
การศึกษาต่อเนื่อง

ตั้งแต่ช่วงทศวรรษ 1960 เป็นต้นมา มีความสนใจในไทแรนโนซอรัส เพิ่มขึ้นอีกครั้ง ส่งผลให้มีการค้นพบโครงกระดูก 42 โครง (สมบูรณ์ 5–80% ตามจำนวนกระดูก) จากทางตะวันตกของทวีปอเมริกาเหนือ[ 6 ]ในปี 1967 ดร. วิลเลียม แมคแมนนิส ได้ค้นพบและกู้คืนโครงกระดูกที่ชื่อว่า "MOR 008" ซึ่งสมบูรณ์ 15% ตามจำนวนกระดูก และมีกะโหลกศีรษะที่สร้างขึ้นใหม่จัดแสดงอยู่ที่พิพิธภัณฑ์แห่งเทือกเขาร็ อกกี้ ทศวรรษ 1990 มีการค้นพบมากมาย โดยมีจำนวนการค้นพบเกือบสองเท่าของทุกปีที่ผ่านมา รวมถึงโครงกระดูกที่สมบูรณ์ที่สุดสองโครงที่พบจนถึงปัจจุบัน ได้แก่ซูและสแตน[ 6 ]
ซู เฮนดริกสัน นัก บรรพชีวินวิทยา สมัครเล่น ค้นพบโครงกระดูก ไทแรนโนซอรัสที่สมบูรณ์ที่สุด (ประมาณ 85%) และใหญ่ที่สุดในแหล่งหินเฮลล์ครีกเมื่อวันที่ 12 สิงหาคม พ.ศ. 2533 โครงกระดูกนี้ ซึ่งตั้งชื่อตามผู้ค้นพบว่า ซู เป็นประเด็นของการต่อสู้ทางกฎหมายเกี่ยวกับกรรมสิทธิ์ ในปี พ.ศ. 2540 คดีความได้ยุติลงโดยตัดสินให้มอริส วิลเลียมส์ เจ้าของที่ดินเดิมเป็นฝ่ายชนะ คอลเลกชันฟอสซิลนี้ถูกซื้อโดยพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติฟิลด์ในการประมูลในราคา 7.6 ล้านดอลลาร์ ทำให้เป็นโครงกระดูกไดโนเสาร์ที่แพงที่สุดจนกระทั่งมีการขายสแตนในราคา 31.8 ล้านดอลลาร์ในปี พ.ศ. 2563 [ 15 ]ตั้งแต่ปี พ.ศ. 2541 ถึง พ.ศ. 2542 เจ้าหน้าที่ของพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติฟิลด์ใช้เวลากว่า 25,000 ชั่วโมงในการนำหินออกจากกระดูก[ 16 ]จากนั้นกระดูกถูกส่งไปยังรัฐนิวเจอร์ซีย์เพื่อสร้างฐานตั้ง แล้วส่งกลับไปยังชิคาโกเพื่อประกอบขั้นสุดท้าย โครงกระดูกที่จัดแสดงเปิดให้ประชาชนเข้าชมเมื่อวันที่ 17 พฤษภาคม พ.ศ. 2543 ที่พิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติฟิลด์ การศึกษาเกี่ยวกับกระดูกฟอสซิลของตัวอย่างนี้แสดงให้เห็นว่าซูโตเต็มที่เมื่ออายุ 19 ปีและเสียชีวิตเมื่ออายุ 28 ปี ซึ่งเป็นอายุขัยโดยประมาณที่ยาวนานที่สุดของไทแรนโนซอรัสที่รู้จัก[ 17 ]

ไทแรนโนซอรัสอีกตัวหนึ่งซึ่งมีชื่อเล่นว่า สแตน (NHMAD 2020.00001 เดิมคือ BHI 3033 [ 18 ] ) เพื่อเป็นเกียรติแก่นักบรรพชีวินวิทยาสมัครเล่น สแตน แซคริสัน ถูกค้นพบจากชั้นหินเฮลล์ครีกในปี 1992 สแตนเป็นโครงกระดูกที่สมบูรณ์ที่สุดเป็นอันดับสองที่พบ โดยมีกระดูก 199 ชิ้น คิดเป็น 70% ของทั้งหมด[ 19 ]ไทแรนโนซอรัสตัวนี้ยังมีพยาธิสภาพของกระดูกหลายอย่าง รวมถึงซี่โครงที่หักและสมานตัวแล้ว คอที่หัก (และสมานตัวแล้ว) และรูขนาดใหญ่ที่ด้านหลังศีรษะ ซึ่งมีขนาดประมาณฟัน ของ ไทแรนโนซอรัส[ 20 ]
ในปี พ.ศ. 2541 บัคกี้ เดอร์ฟลิงเกอร์ วัย 20 ปี สังเกตเห็นนิ้วเท้าของทีเร็กซ์ โผล่ขึ้นมาเหนือพื้นดิน ทำให้เขากลายเป็นบุคคลที่อายุน้อยที่สุดที่ค้นพบ ไทแรนโนซอรัสตัวอย่างดังกล่าวได้รับการตั้งชื่อว่าบัคกี้เพื่อเป็นเกียรติแก่ผู้ค้นพบ เป็นไทแรนโนซอรัสวัยหนุ่ม สูง 3.0 เมตร (10 ฟุต) และยาว 11 เมตร (35 ฟุต) บัคกี้เป็นไทแรนโนซอรัสตัว แรก ที่ถูกค้นพบซึ่งยังคงมีกระดูกอก (กระดูกรูปตัววี) บัคกี้จัดแสดงถาวรอยู่ที่พิพิธภัณฑ์เด็กแห่งอินเดียนาโพลิส[ 21 ]

ในช่วงฤดูร้อนปี 2000 ทีมงานที่จัดตั้งโดยJack Horner ได้ค้นพบโครงกระดูก Tyrannosaurusจำนวน 5 โครงใกล้กับอ่างเก็บน้ำ Fort Peck [ 22 ] ในปี 2001 โครงกระดูกของTyrannosaurus วัยเยาว์ที่สมบูรณ์ 50% ถูกค้นพบในชั้นหิน Hell Creek โดยทีมงานจากพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติ Burpeeโครงกระดูกนี้มีชื่อว่า Jane (BMRP 2002.4.1) เดิมทีคิดว่าเป็นโครงกระดูกแรกที่รู้จักของTyrannosaurus แคระNanotyrannusแต่ผู้วิจัยคนอื่นๆ ถือว่ามันเป็น โครงกระดูก T. rexวัย เยาว์ที่สมบูรณ์ที่สุด [ 23 ]ปัจจุบัน Jane ซึ่งจัดแสดงอยู่ที่พิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติ Burpee ได้รับการอธิบายใหม่ว่าเป็นโฮโลไทป์ของN. lethaeus [ 24 ]ในปี 2545 โครงกระดูกที่ได้รับฉายาว่า "Wyrex" ซึ่งค้นพบโดยนักสะสมสมัครเล่น Dan Wells และ Don Wyrick มีกระดูก 114 ชิ้นและสมบูรณ์ 38% การขุดค้นเสร็จสิ้นภายใน 3 สัปดาห์ในปี 2547 โดยสถาบัน Black Hillsด้วย การขุดค้น Tyrannosaurus แบบสด ทางออนไลน์ ครั้งแรก ซึ่งมีการรายงานรายวัน รูปภาพ และวิดีโอ[ 6 ]
ในปี พ.ศ. 2549 มหาวิทยาลัยแห่งรัฐมอนแทนาได้เปิดเผยว่ามี กะโหลก ไทแรนโนซอรัส ที่ใหญ่ที่สุด เท่าที่เคยค้นพบ (จากตัวอย่างชื่อ MOR 008) ซึ่งมีความยาว 5 ฟุต (152 ซม.) [ 25 ]การเปรียบเทียบในภายหลังระบุว่าหัวที่ยาวที่สุดคือ 136.5 เซนติเมตร (53.7 นิ้ว) (จากตัวอย่าง LACM 23844) และหัวที่กว้างที่สุดคือ 90.2 เซนติเมตร (35.5 นิ้ว) (จาก Sue) [ 26 ]
คำอธิบาย
ขนาด

ทีเร็กซ์เป็นหนึ่งในสัตว์กินเนื้อบนบกที่ใหญ่ที่สุดตลอดกาล ตัวอย่างที่ใหญ่ที่สุดและสมบูรณ์ที่สุดชิ้นหนึ่ง ซึ่งมีชื่อเล่นว่าซู (FMNH PR2081) ตั้งอยู่ที่พิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติฟิลด์ในชิคาโก ซูมีความยาว 12.3–12.8 เมตร (40–42 ฟุต) [ 27 ] [ 28 ] [ 29 ]และสูงที่สะโพก 3.66–3.96 เมตร (12.0–13.0 ฟุต) [ 27 ] [ 30 ] [ 31 ]และจากการศึกษาล่าสุดโดยใช้เทคนิคที่หลากหลาย มวลร่างกายสูงสุดได้รับการประมาณไว้ที่ 8.4–9.22 ตัน (8.27–9.07 ตันยาว; 9.26–10.16 ตันสั้น) [ 32 ] [ 33 ] [ 34 ]ตัวอย่างที่มีชื่อเล่นว่าScotty (RSM P2523.8) ซึ่งตั้งอยู่ที่พิพิธภัณฑ์ Royal Saskatchewan Museumมีรายงานว่ามีความยาว 13 เมตร (43 ฟุต) โดยใช้วิธีการประมาณมวลที่ขยายจากเส้นรอบวงของกระดูกต้นขา Scotty ถูกประมาณว่าเป็นตัวอย่างที่ใหญ่ที่สุดเท่าที่รู้จัก โดยมีมวลร่างกาย 8.87 ตัน (8.73 ตันยาว; 9.78 ตันสั้น) [ 32 ] [ 35 ]
ไม่ใช่ว่า ตัวอย่าง ไทแรนโนซอรัสที่ โตเต็มวัยทุกตัว ที่ถูกค้นพบจะมีขนาดใหญ่เท่ากันเสมอไป ในอดีต การประมาณมวลเฉลี่ยของตัวเต็มวัยมีความแตกต่างกันอย่างมากในช่วงหลายปีที่ผ่านมา ตั้งแต่ต่ำสุดที่ 4.5 ตัน (4.4 ตันยาว; 5.0 ตันสั้น) [ 36 ] [ 37 ]ไปจนถึงมากกว่า 7.2 ตัน (7.1 ตันยาว; 7.9 ตันสั้น) [ 38 ]โดยการประมาณค่าในปัจจุบันส่วนใหญ่จะอยู่ระหว่าง 5.4 ถึง 8.0 ตัน (5.3 ถึง 7.9 ตันยาว; 6.0 ถึง 8.8 ตันสั้น) [ 28 ] [ 39 ] [ 40 ] [ 41 ] [ 42 ]
การศึกษาในปี 2024 พบว่ามีหลักฐานเพียงเล็กน้อยเกี่ยวกับความแตกต่างทางเพศตามขนาดในทีเร็กซ์[ 43 ]
กะโหลก

กะโหลก T. rexที่ใหญ่ที่สุดเท่าที่รู้จักมีความยาวถึง 1.52 เมตร (5 ฟุต 0 นิ้ว) [ 25 ] [ 27 ]ช่อง เปิด ขนาดใหญ่ในกะโหลกช่วยลดน้ำหนัก เช่นเดียวกับเทโรพอดกินเนื้อทั้งหมด ในด้านอื่นๆกะโหลกของTyrannosaurus แตกต่างอย่างมากจากกะโหลกของเทโรพอดขนาดใหญ่ที่ไม่ใช่ ไทแรนโนซอริเดมันกว้างมากที่ด้านหลัง แต่มีจมูกแคบ ทำให้มองเห็นแบบสองตาได้ดีเป็นพิเศษ[ 44 ] [ 45 ]กระดูกกะโหลกมีขนาดใหญ่ และกระดูกจมูกและกระดูกอื่นๆ บางส่วนเชื่อมติดกัน ป้องกันการเคลื่อนไหวระหว่างกัน แต่หลายชิ้นมีโพรงอากาศ (มี "รังผึ้ง" ของช่องอากาศเล็กๆ) จึงทำให้เบากว่า คุณสมบัติเหล่านี้และคุณสมบัติอื่นๆ ที่เสริมความแข็งแรงของกะโหลกเป็นส่วนหนึ่งของ แนวโน้ม ของไทแรนโนซอริเดที่มุ่งไปสู่การกัดที่ทรงพลังมากขึ้น ซึ่งเหนือกว่าการกัดของเทโรพอดที่ไม่ใช่ไทแรนโนซอริเดทั้งหมดอย่างง่ายดาย[ 46 ] [ 47 ] [ 48 ]ปลายขากรรไกรบนมีรูปทรงตัว U (สัตว์กินเนื้อที่ไม่ใช่ไทแรนโนซอรัสส่วนใหญ่มีขากรรไกรบนรูปทรงตัว V) ซึ่งช่วยเพิ่มปริมาณเนื้อเยื่อและกระดูกที่ไทแรนโนซอรัสสามารถฉีกออกได้ด้วยการกัดเพียงครั้งเดียว แม้ว่ามันจะเพิ่มแรงกดดันต่อฟันหน้าด้วยก็ตาม[ 49 ]
ฟันของทีเร็กซ์แสดงให้เห็นถึงความแตกต่างของรูปร่างอย่างเห็นได้ชัด[ 8 ] [ 50 ]ฟันพรีแม็กซิลลารีซึ่งมีข้างละสี่ซี่ที่ด้านหน้าของขากรรไกรบน เรียงตัวกันอย่างหนาแน่น มี รูปทรงตัว Dในส่วนตัดขวาง มีสันเสริมความแข็งแรงที่พื้นผิวด้านหลัง มีลักษณะเป็นฟันตัด (ปลายฟันมีลักษณะคล้ายใบมีดสิ่ว) และโค้งไปด้านหลัง รูปทรงตัว Dในส่วนตัดขวาง สันเสริมความแข็งแรง และการโค้งไปด้านหลัง ช่วยลดความเสี่ยงที่ฟันจะหักเมื่อไทแรนโนซอรัสกัดและดึง ฟันที่เหลือมีความแข็งแรงเหมือน "กล้วยพิษ" มากกว่ามีดสั้น มีระยะห่างกันมากขึ้น และยังมีสันเสริมความแข็งแรงด้วย[ 51 ]ฟันในขากรรไกรบน ซึ่งมีข้างละสิบสองซี่ในตัวที่โตเต็มวัย[ 8 ]มีขนาดใหญ่กว่าฟันในขากรรไกรล่าง ยกเว้นที่ด้านหลัง ฟันที่ใหญ่ที่สุดที่พบจนถึงขณะนี้คาดว่ามีความยาว 30.5 ซม. (12.0 นิ้ว) รวมรากเมื่อสัตว์ยังมีชีวิตอยู่ ทำให้เป็นฟันที่ใหญ่ที่สุดของไดโนเสาร์กินเนื้อที่เคยพบมา[ 52 ]ขากรรไกรล่างแข็งแรงกระดูกขากรรไกร ด้านหน้า มีฟัน 13 ซี่ ด้านหลังแถวฟัน ขากรรไกรล่างจะสูงขึ้นอย่างเห็นได้ชัด[ 8 ]ขากรรไกรบนและล่างของไทแรนโนซอรัสเช่นเดียวกับไดโนเสาร์หลายชนิด มีรูเล็กๆ จำนวนมากในกระดูก มีการเสนอหน้าที่ต่างๆ ของรูเหล่านี้ เช่น ระบบรับความรู้สึกคล้ายจระเข้[ 53 ]หรือหลักฐานของโครงสร้างภายนอกช่องปากเช่น เกล็ดหรือริมฝีปาก[ 54 ] [ 55 ] [ 56 ]โดยการวิจัยในภายหลังเกี่ยวกับรูปแบบการสึกหรอของฟันเทโรพอดสนับสนุนข้อเสนอดังกล่าว[ 57 ]
โครงกระดูก


กระดูกสันหลังของไทแรนโนซอรัสประกอบด้วยกระดูกสันหลังส่วนคอ 10 ชิ้น กระดูกสันหลังส่วนหลัง 13 ชิ้น และกระดูกสันหลังส่วนกระเบนเหน็บ 5 ชิ้น จำนวนกระดูกสันหลังส่วนหางไม่เป็นที่ทราบแน่ชัดและอาจแตกต่างกันไปในแต่ละตัว แต่คาดว่ามีอย่างน้อย 40 ชิ้น ซูมีกระดูกสันหลังส่วนหาง 47 ชิ้น[ 8 ] คอของทีเร็กซ์มีลักษณะโค้งเป็นรูปตัว S ตามธรรมชาติเช่นเดียวกับเทอโรพอดชนิดอื่นๆ เมื่อเทียบกับชนิดอื่นๆ คอของทีเร็กซ์นั้นสั้น ลึก และมีกล้ามเนื้อมากเป็นพิเศษเพื่อรองรับหัวขนาดใหญ่ กระดูกสันหลังชิ้นที่สองหรือแกนกลางนั้นสั้นเป็นพิเศษ กระดูกสันหลังส่วนคอที่เหลือมีลักษณะโอพิสโทโคอีลัสเล็กน้อย กล่าวคือ ด้านหน้าของกระดูกสันหลังนูนและด้านหลังเว้า กระดูกสันหลังมีเพลอโรโคอีลเดี่ยว ซึ่งเป็นช่องลมที่เกิดจากถุงลมอยู่ด้านข้าง[ 8 ]กระดูกสันหลังส่วนลำตัวแข็งแรงแต่มีเอวแคบ ด้านล่างของลำตัวมีลักษณะเป็นสันนูน ด้านหน้าเว้าและมีร่องลึกในแนวตั้ง มีช่องอกขนาดใหญ่ กระดูกสันหลังส่วนประสาทมีด้านหน้าและด้านหลังที่ขรุขระมากเพื่อยึดเอ็นที่แข็งแรง กระดูกสันหลังส่วนกระเบนเหน็บเชื่อมติดกันทั้งในส่วนของตัวกระดูกสันหลังและกระดูกสันหลังส่วนประสาท มีโพรงอากาศอยู่ภายใน เชื่อมต่อกับกระดูกเชิงกรานด้วยกระบวนการตามขวางและซี่โครงส่วนกระเบนเหน็บ หางมีขนาดใหญ่และยาวปานกลาง เพื่อรักษาสมดุลของหัวและลำตัวขนาดใหญ่ และเพื่อให้มีพื้นที่สำหรับกล้ามเนื้อเคลื่อนที่ ขนาดใหญ่ ที่ยึดติดกับกระดูกต้นขา กระดูกสันหลังส่วนหางชิ้นที่สิบสามเป็นจุดเปลี่ยนระหว่างฐานหางที่ลึกและหางส่วนกลางที่แข็งตัวด้วยกระบวนการข้อต่อด้านหน้าที่ค่อนข้างยาว ด้านล่างของลำตัวปกคลุมด้วยซี่โครงท้องแบบแบ่งเป็นปล้องจำนวนสิบแปดหรือสิบเก้าคู่[ 8 ]

กระดูกหัวไหล่ยาวกว่าแขนขาหน้าทั้งหมด กระดูกสะบักมีแกนแคบแต่ขยายออกเป็นพิเศษที่ปลายด้านบน เชื่อมต่อกับกระดูกโคราคอยด์ซึ่งมีลักษณะกลมโดยส่วนที่ยื่นออกมาข้างหน้ายาว กระดูกสะบักทั้งสองข้างเชื่อมต่อกันด้วยกระดูกเฟอร์คู ล่าขนาดเล็ก กระดูกอกคู่น่าจะทำจากกระดูกอ่อนเท่านั้น[ 8 ]
แขนส่วนหน้าหรือแขนนั้นสั้นมาก กระดูกต้นแขนส่วนบนหรือกระดูกฮิวเมอรัสสั้นแต่แข็งแรง มีปลายบนที่แคบและมีหัวกลมเป็นพิเศษ กระดูกปลายแขนส่วนล่างหรือกระดูกอัลนาและเรเดียสเป็นกระดูกตรง สั้นกว่ากระดูกฮิวเมอรัสมาก กระดูกฝ่ามือชิ้น ที่สอง ยาวและกว้างกว่าชิ้นแรก ในขณะที่ปกติในเทอโรพอดจะเป็นตรงกันข้าม แขนส่วนหน้ามีเพียงสองนิ้วที่มีเล็บ[ 8 ] พร้อมกับ กระดูกฝ่ามือชิ้นที่สามขนาดเล็กคล้ายเฝือกซึ่งเป็นส่วนที่เหลือของนิ้วที่สาม[ 58 ]

กระดูกเชิงกรานเป็นโครงสร้างขนาดใหญ่ กระดูกส่วนบนคือกระดูกอิเลียม นั้น ยาวและสูงมาก ทำให้มีพื้นที่ยึดเกาะที่กว้างขวางสำหรับกล้ามเนื้อขาหลัง กระดูกหัวหน่าวส่วนหน้าสิ้นสุด ลงที่ส่วนปลายกระดูกหัวหน่าวขนาดใหญ่ ซึ่งยาวกว่าแกนทั้งหมดของกระดูกนั้น กระดูกอิสเคียมส่วนหลังนั้นเรียวและตรง ชี้เฉียงไปด้านหลังและลงล่าง[ 8 ]
เมื่อเปรียบเทียบกับแขน ขาหลังของเทโรพอดนั้นยาวที่สุดเมื่อเทียบกับขนาดตัว ในเท้า กระดูกฝ่าเท้ามีลักษณะ "arctometatarsalian" ซึ่งหมายความว่าส่วนของกระดูกฝ่าเท้าที่สามใกล้ข้อเท้ามีลักษณะแคบ กระดูกฝ่าเท้าที่สามยังมีลักษณะโค้งงอเป็นพิเศษอีกด้วย[ 8 ]เพื่อชดเชยขนาดตัวที่ใหญ่โตของสัตว์ กระดูกหลายชิ้นทั่วทั้งโครงกระดูกจึงมีลักษณะกลวง ช่วยลดน้ำหนักโดยไม่สูญเสียความแข็งแรงอย่างมีนัยสำคัญ[ 8 ]
การจำแนกประเภท

ไทแรนโนซอรัสเป็น สกุล ต้นแบบของวงศ์ใหญ่ไทแรนโนซอโรอิเดีย วงศ์ ไทแรนโนซอริเด และวงศ์ย่อยไทแรนโนซอรีเน กล่าวอีกนัยหนึ่งคือเป็นมาตรฐานที่นักบรรพชีวินวิทยาใช้ในการตัดสินใจว่าจะรวมสายพันธุ์อื่น ๆ ไว้ในกลุ่มเดียวกันหรือ ไม่ สมาชิกอื่น ๆ ของวงศ์ย่อยไทแรนโนซอรีเน ได้แก่ ดาสเปลโตซอรัสในอเมริกาเหนือและทาร์โบซอรัส ในเอเชีย [ 23 ] [ 59 ]ซึ่งบางครั้งถูกจัดให้เป็นชื่อพ้องกับไทแรนโนซอรัส[ 60 ]
เดิมทีเชื่อกันว่าไทแรนโนซอริเดสเป็นลูกหลานของเทโรพอดขนาดใหญ่ในยุคก่อนหน้า เช่นเมกาโลซอรัสและคาร์โนซอรัสแม้ว่าเมื่อไม่นานมานี้พวกมันจะถูกจัดกลุ่มใหม่ร่วมกับโคเอลูโรซอรัสซึ่ง โดยทั่วไปมีขนาดเล็กกว่า [ 49 ]กลุ่มไทแรนโนซอรัสที่เก่าแก่ที่สุดคือโปรเซราโตซอริเดสที่มีหงอน ในขณะที่สมาชิกที่พัฒนาในภายหลังและวิวัฒนาการมากขึ้นอยู่ในกลุ่ม แพนไทแรนโนซอเรีย ไทแรนโนซอรัสเริ่มต้นจากการเป็นเทโร พอดขนาดเล็ก อย่างไรก็ตามอย่างน้อยบางชนิดก็มีขนาดใหญ่ขึ้นใน ช่วงต้นยุคครี เทเชียสเป็นไปได้มากว่าไทแรนโนซอรอยด์จะมีบทบาทสำคัญมากขึ้นหลังจากความหลากหลายของอัลโลซอรอยด์และเมกาโลซอรอยด์ลดลงในช่วงต้นของยุคครีเทเชียสตอนปลาย[ 61 ]
ไทแรนโนซอรัสมีลักษณะเด่นคือกระดูกจมูกที่เชื่อมติดกันและการจัดเรียงฟัน แพนทีแรนโนซอรัสมีลักษณะเฉพาะที่สะโพก รวมถึงรูที่ขยายใหญ่ขึ้นในกระดูกควอดเรต กระดูกโพสต์ออร์บิทัลที่กว้าง และกระดูกจมูกรูปทรงนาฬิกาทราย แพนทีแรนโนซอรัสที่วิวัฒนาการมามากกว่าบางชนิดไม่มีโพรงอากาศในจมูก และมีอัตราส่วนกระดูกต้นแขนต่อกระดูกต้นขาที่ต่ำกว่า โดยแขนเริ่มลดขนาดลง แพนทีแรนโนซอรัสบางชนิดเริ่มพัฒนากระดูกอาร์คโตเมตาตาร์ซัส ยูทีแรนโนซอรัสมีพื้นผิวที่หยาบบนกระดูกจมูก และช่องเปิดขากรรไกรล่างลดขนาดลงภายนอก ไทแรนโนซอริเดไม่มีกะโหลกที่เคลื่อนไหวได้หรือสันพิเศษบนกระดูกจมูก และมีกระดูกน้ำตาที่มีกระบวนการที่โดดเด่น ไทแรนโนซอริเดยังมีคานระหว่างช่องเปิดที่มีขนาดเล็กกว่าครึ่งหนึ่งของช่องเปิดขากรรไกรบน[ 62 ]
การวิเคราะห์ ทางวิวัฒนาการหลายครั้งพบว่าTarbosaurus bataarเป็นกลุ่มพี่น้องของT. rex [ 59 ] การค้นพบLythronax ซึ่ง เป็น ไทแรนโนซอริเด อีกตัวหนึ่งบ่งชี้ว่าTarbosaurusและTyrannosaurusมีความสัมพันธ์ใกล้ชิดกัน โดยก่อตัวเป็นกลุ่มเดียวกันกับZhuchengtyrannusซึ่งเป็นไทแรนโนซอริเดจากเอเชียเช่นกัน โดยLythronax เป็นกลุ่มพี่น้องของพวก มัน[ 63 ] [ 64 ]การศึกษาเพิ่มเติมในปี 2016 โดย Steve Brusatte, Thomas Carr และเพื่อนร่วมงาน ยังบ่งชี้ว่าTyrannosaurusอาจเป็นผู้อพยพมาจากเอเชีย และอาจเป็นลูกหลานของTarbosaurus ด้วย [ 65 ]
ด้านล่างนี้คือแผนภูมิวิวัฒนาการของ Tyrannosauridae โดยอิงจากการวิเคราะห์ทางวิวัฒนาการที่ดำเนินการโดย Loewen และเพื่อนร่วมงานในปี 2013 [ 63 ]
ในการบรรยายลักษณะของTyrannosaurus mcraeensis ในปี 2024 Dalman และคณะได้ค้นพบผลลัพธ์ที่คล้ายคลึงกับการวิเคราะห์ก่อนหน้านี้ โดยTyrannosaurusเป็นกลุ่มพี่น้องกับกลุ่มที่ประกอบด้วยTarbosaurusและZhuchengtyrannusซึ่งเรียกว่า Tyrannosaurini นอกจากนี้พวกเขายังพบหลักฐานสนับสนุน กลุ่ม โมโนฟิเลติกที่ประกอบด้วยDaspletosaurusและThanatotheristesซึ่งโดยทั่วไปเรียกว่าDaspletosaurini [ 66 ] [ 67 ]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
สายพันธุ์เพิ่มเติม

ในปี พ.ศ. 2498 นักบรรพชีวินวิทยา ชาวโซเวียต Evgeny Maleevได้ตั้งชื่อสายพันธุ์ใหม่ว่าTyrannosaurus bataarจากมองโกเลีย [ 68 ]ในปี พ.ศ. 2508 สายพันธุ์นี้ได้รับการเปลี่ยนชื่อเป็นสกุลที่แยกต่างหาก คือTarbosaurus bataar [ 69 ]ใน ขณะที่นักบรรพชีวินวิทยาส่วนใหญ่ยังคงถือว่าทั้งสองเป็น สกุลที่แยกต่างหาก แต่ผู้เขียนบางคน เช่นThomas Holtz , Kenneth CarpenterและThomas Carrโต้แย้งว่าทั้งสองสายพันธุ์มีความคล้ายคลึงกันมากพอที่จะถือว่าเป็นสมาชิกของสกุลเดียวกัน โดยคืนชื่อวิทยาศาสตร์ดั้งเดิมของสายพันธุ์ มองโกเลีย [ 49 ] [ 70 ] [ 53 ]
ตัวอย่างบางส่วนจากแหล่งสะสมยุคครีเทเชียสตอนปลายของจีนได้รับการอธิบายว่าเป็นสายพันธุ์ใหม่ของTyrannosaurusได้แก่T. "lanpingensis" โดยอิงจาก ฟัน ด้านข้าง ที่แยกออกมา จากชั้นหินสีแดงของยูนนานในปี 1975; T. " turpanensis " จากชั้นหิน Subashiในแอ่ง Turpan มณฑลซินเจียงในปี 1978; และ T. luanchuanensis จากชั้นหิน Quibaในแอ่ง Tantou มณฑลเหอหนานในปี 1979–1980 [ 71 ] [ 72 ] [ 73 ]สองกลุ่มแรกได้รับการตีพิมพ์โดยไม่มีคำอธิบายโดยละเอียด ดังนั้นจึงเป็น nomina nuda T. "turpanensis" และT. luanchuanensisได้รับการระบุเบื้องต้นว่าเป็นชื่อพ้องรองของTarbosaurus bataarโดย Holtz ในปี 2004; Holtz ยังระบุT. "lanpingensis" แต่เป็น nomen dubium [ 59 ] [ 72 ] [ 73 ]
VGI หมายเลข 231/3 กระดูกนิ้ว ขนาดใหญ่ ซึ่ง Yarkov กำหนดให้เป็นTyrannosaurus sp. ในปี 2000 ถูกค้นพบในยุค Maastrichtian ตอนล่างของBereslavka ประเทศรัสเซียในปี 2004 Averianov และ Yarkov ได้ตีความใหม่ว่าเป็นกระดูกฝ่ามือชิ้นที่ 1 หรือกระดูกฝ่าเท้าชิ้น ที่ 1 ซึ่งอาจเป็นของเซราโท ซอ ร์[ 74 ]ในภาพรวมปี 2023 Averianov และ Lopatin กล่าวถึงตัวอย่างนี้รวมถึงฟันซี่เดียวจากแหล่งเดียวกันว่าเป็น Theropoda ที่ระบุไม่ได้[ 75 ]
ในปี 2001 ฟันและกระดูกฝ่าเท้าของไดโนเสาร์วงศ์ Tyrannosauridae หลายชิ้นที่ขุดพบในเหมืองหินใกล้เมือง Zhuchengประเทศจีน ถูกจัดโดยนักบรรพชีวินวิทยาชาวจีนHu Chengzhiให้เป็นสายพันธุ์ใหม่ที่ตั้งขึ้นคือTyrannosaurus zhuchengensisอย่างไรก็ตาม ในปี 2011 ในสถานที่ใกล้เคียงกัน กระดูกขากรรไกรบนด้านขวาและกระดูกขากรรไกรล่างด้านซ้ายถูกจัดให้เป็นสกุลใหม่ที่ตั้งขึ้นคือZhuchengtyrannusเป็นไปได้ว่าT. zhuchengensisอาจเป็นชื่อพ้องกับZhuchengtyrannusไม่ว่าในกรณีใดT. zhuchengensisถือเป็นชื่อที่ไม่แน่ชัดเนื่องจากตัวอย่างต้นแบบขาดลักษณะเฉพาะที่ต่ำกว่าระดับ Tyrannosaurinae [ 76 ]
ในปี 2006 กระดูกน้ำตาชิ้นเล็กๆ ของไดโนเสาร์วงศ์ Tyrannosauridae (CM 9401) จากชั้นหิน Judith River Formationในเขต Fergus County รัฐ Montana ถูกระบุว่าเป็น ? Tyrannosaurus sp. กระดูกน้ำตาข้างขวาชิ้นนี้ถูกเก็บรวบรวมมาพร้อมกับตัวอย่างต้นแบบของDeinosuchus rugosusซึ่งเป็นจระเข้ยักษ์ และยังไม่ได้รับการอธิบายจนกระทั่งนักบรรพชีวินวิทยาDale Russell ระบุใหม่ว่าเป็นของไดโนเสาร์วงศ์ Tyrannosauridae ในช่วงทศวรรษ 1980 กระดูกน้ำตาชิ้นนี้มีลักษณะคล้ายคลึงกับของTyrannosaurus rexทั้งขนาดและรูปร่าง ที่น่าสังเกตคือ มันไม่มี "เขากระดูกน้ำตา" ที่มักพบในไดโนเสาร์วงศ์ Tyrannosauridae รุ่นก่อนๆ เช่นAlbertosaurusและGorgosaurusแต่กลับมีลักษณะเป็นร่องขรุขระตามพื้นผิวด้านบน ซึ่งสอดคล้องกับT. rexและTarbosaurus ซึ่งเป็นญาติใน เอเชีย ขนาดที่ใหญ่โตของตัวอย่างนี้ทำให้มันอยู่ในช่วงของตัวอย่างT. rex ที่รู้จักกัน ซึ่งบ่งชี้ว่ามีไดโนเสาร์วงศ์ Tyrannosauridae ขนาดใหญ่ในช่วงยุค Campanian (~75 ล้านปีก่อน) ซึ่งเป็นช่วงเวลาที่เร็วกว่ายุค Maastrichtian ที่กำหนดไว้ (~68–66 ล้านปีก่อน) สำหรับTyrannosaurus rexอย่างไรก็ตาม อายุและแหล่งที่มาที่แน่นอนของ CM 9401 ยังคงไม่แน่นอนเนื่องจากขาดเอกสารภาคสนามโดยละเอียด[ 77 ]
ในปี 2018 บทความที่อธิบายฟันของไทแรนโนซอริเดจากชั้นหินทูเมดิซีนระบุว่าฟันพรีแม็กซิลลารี (YPM VPPU 023469) มีความคล้ายคลึงกับฟันของซู อย่างมาก โดยไม่รวมไทแรนโนซอริเดในยุคแคมพาเนียน นอกจากนี้ ผู้เขียนบทความนี้ยังแนะนำว่า CM 9401 ก็มาจากชั้นหินทูเมดิซีนเช่นกัน เนื่องจากมีความคล้ายคลึงกันในการเก็บรักษาระหว่างแหล่งที่พบและแอนติไคลน์วิลโลว์ครีก ซึ่งเป็นที่ที่พบฟัน[ 78 ]ที่น่าสังเกตคือ สิ่งนี้จะทำให้ตัวอย่างทั้งสองอยู่ในแฟลกบัตต์เมมเบอร์ของชั้นหินทูเมดิซีน ซึ่งมีอายุตั้งแต่ 77 ถึง 76.3 ล้านปี เก่าแก่กว่า ตัวอย่าง ไทแรนโนซอรัส อื่นๆ มาก และร่วมสมัยกับ ดาสเปลโตซอรัสโดยตรง ในปี 2025 ตัวอย่างเหล่านี้ที่มีอายุทางธรณีวิทยาเก่าแก่ถูกใช้เป็นหลักฐานโดยชาร์ลี เชเรอร์ เพื่อ เสนอแนะว่าไทแรนโนซอรินีไม่ได้วิวัฒนาการโดยตรงจาก ดาสเปล โตซอรัส[ 79 ]
ไทแรนโนซอรัส อิมพีเรียเลเตอร์และไทแรนโนซอรัส เรจินา
ในการศึกษาปี 2022 เกรกอรี เอส. พอลและเพื่อนร่วมงานได้โต้แย้งว่าไทแรนโนซอรัส เร็กซ์ตามความเข้าใจดั้งเดิมนั้น แท้จริงแล้วประกอบด้วยสามสายพันธุ์ ได้แก่ สายพันธุ์ต้นแบบไทแรนโนซอรัส เร็กซ์ และอีกสองสายพันธุ์ที่ไม่เคยได้รับการยอมรับมาก่อน คือที. อิ มพี เรีย เลเตอร์ ( T. imperator หมาย ถึง "จักรพรรดิกิ้งก่าทรราช") และ ที. เรจินา ( T. regina หมายถึง "ราชินีกิ้งก่าทรราช" ) ตัวอย่างต้นแบบของสายพันธุ์แรก (ที. อิมพีเรียเลเตอร์) คือ ตัวอย่างจาก ถ้ำซูและตัวอย่างต้นแบบของสายพันธุ์หลัง ( ที. เรจินา ) คือแวนเคล เร็กซ์การแบ่งออกเป็นหลายสายพันธุ์นั้นขึ้นอยู่กับความแปรผันอย่างมากในสัดส่วนและความแข็งแรงของกระดูกต้นขา (และส่วนประกอบโครงกระดูกอื่นๆ) ใน ตัวอย่าง ที. เร็กซ์นอกจากนี้ยังใช้ลักษณะรูปร่างที่แข็งแรงและบอบบาง รวมถึงจำนวนฟันหน้าขนาดเล็กในขากรรไกรล่างเป็นหลักฐานประกอบด้วย พอลและเพื่อนร่วมงานได้จัดประเภทสายพันธุ์ดังนี้: T. rexแสดงให้เห็นกายวิภาคที่แข็งแรง อัตราส่วนความยาวต่อเส้นรอบวงของกระดูกต้นขาอยู่ในระดับปานกลาง และมีฟันกรามล่างรูปฟันตัดที่เรียวเล็กเพียงซี่เดียวT. imperatorมีโครงสร้างแข็งแรง มีอัตราส่วนความยาวต่อเส้นรอบวงของกระดูกต้นขาต่ำ และมีฟันที่เรียวเล็กสองซี่ และT. reginaมีโครงสร้างที่บอบบางกว่า มีอัตราส่วนกระดูกต้นขาสูง และมีฟันที่เรียวเล็กหนึ่งซี่[ 80 ]
อย่างไรก็ตาม นักบรรพชีวินวิทยาชั้นนำหลายคน รวมถึงStephen Brusatte , Thomas Carr , Thomas Holtz , David Hone, Jingmai O'ConnorและLindsay Zannoได้วิพากษ์วิจารณ์การศึกษาหรือแสดงความสงสัยต่อข้อสรุปเมื่อได้รับการติดต่อจากสื่อต่างๆ เพื่อขอความคิดเห็น[ 81 ] [ 82 ] [ 83 ]คำวิจารณ์ของพวกเขาได้รับการตีพิมพ์ในเอกสารทางเทคนิคในภายหลัง[ 84 ] ทั้ง Holtz และ Zanno ต่างกล่าวว่ามีความเป็นไปได้ที่ Tyrannosaurusมากกว่าหนึ่งสายพันธุ์จะมีอยู่ แต่รู้สึกว่าการศึกษาใหม่นี้ไม่เพียงพอที่จะสนับสนุนสายพันธุ์ที่เสนอ Holtz กล่าวว่า แม้ว่าTyrannosaurus imperatorจะเป็นสายพันธุ์ที่แตกต่างจากTyrannosaurus rexแต่ก็อาจเป็นสายพันธุ์เดียวกันกับNanotyrannus lancensisและจะต้องเรียกว่าTyrannosaurus lancensis โอคอนเนอร์ ภัณฑารักษ์ที่พิพิธภัณฑ์ฟิลด์ ซึ่ง เป็นที่จัดแสดงตัวอย่างต้นแบบ T. imperatorชื่อซู ถือว่าสายพันธุ์ใหม่นี้ได้รับการสนับสนุนน้อยเกินไปจนไม่คุ้มค่าที่จะแก้ไขป้ายนิทรรศการ บรูซัตต์ คาร์ และโอคอนเนอร์ มองว่าลักษณะเด่นที่เสนอระหว่างสายพันธุ์ต่างๆ นั้นสะท้อนถึงความแปรผันตามธรรมชาติภายในสายพันธุ์เดียวกัน ทั้งคาร์และโอคอนเนอร์ต่างแสดงความกังวลเกี่ยวกับความไม่สามารถของงานวิจัยที่จะระบุได้ว่าตัวอย่างที่ได้รับการอนุรักษ์ไว้อย่างดีหลายชิ้นนั้นเป็นของสายพันธุ์ใดในบรรดาสายพันธุ์ที่เสนอ นักบรรพชีวินวิทยาอีกคนหนึ่งฟิลิป เจ. เคอร์รีเดิมทีเป็นผู้ร่วมเขียนงานวิจัยนี้ แต่ได้ถอนตัวออกไปเนื่องจากไม่ต้องการมีส่วนร่วมในการตั้งชื่อสายพันธุ์ใหม่[ 81 ]
ในบทความที่ตีพิมพ์ในภายหลังในปี 2025 Paul ยังคงยืนยันความถูกต้องของT. imperatorและT. reginaโดยโต้แย้งว่าสายพันธุ์ที่เสนอเหล่านี้สามารถแยกแยะได้เพิ่มเติมโดยรูปร่างของปุ่มหลังเบ้าตา (ส่วนที่เป็นกระดูกขรุขระด้านหลังดวงตา) [ 85 ]
ไทแรนโนซอรัส แมคเรเอนซิส
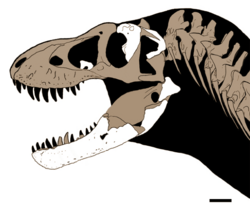

ในปี 2024 Dalman และเพื่อนร่วมงานได้อธิบายซากดึกดำบรรพ์ของไทแรนโนซอร์ที่ค้นพบในปี 1983 ในชั้นหิน Hall Lake Formation ยุค Campanian-ต้น Maastrichtian ในรัฐนิวเม็กซิโกซากดึกดำบรรพ์ (NMMNH P-3698) ซึ่งเก็บรักษาไว้ที่ พิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติและวิทยาศาสตร์แห่งรัฐนิวเม็กซิโก ประกอบด้วย กระดูก postorbital ขวา, กระดูก squamosalขวา, กระดูก palatine ซ้าย และกระดูกmaxilla ที่ไม่สมบูรณ์จากกะโหลกศีรษะ กระดูก dentaryซ้าย, กระดูก splenial ขวา , กระดูก prearticular ขวา, กระดูก angular ขวา และกระดูก articular ขวา จากขากรรไกรล่าง ฟันที่แยกออกมา และกระดูกรูปตัววี [ 66 ] กระดูก บางส่วนถูก กล่าวถึงสั้นๆ ในปี 1984 ว่าเป็นของT. rex [ 86 ]และอธิบายไว้ในปี 1986 [ 87 ]
Lehman และ Carpenter (1990) เสนอว่า NMMNH P-3698 เป็นของสกุลไทแรนโนซอริเดใหม่[ 88 ]ในขณะที่ Carr และ Williamson (2000) ไม่เห็นด้วยกับข้ออ้างของพวกเขา[ 89 ] Sullivan และ Lucas (2015) โต้แย้งว่ามีหลักฐานน้อยมากที่จะสนับสนุนว่า NMMNH P-3698 เป็นตัวอย่างของTyrannosaurus rexดังนั้นพวกเขาจึงจัดประเภทเบื้องต้นเป็น cf. Tyrannosaurus sp.; พวกเขายังพิจารณาว่าไทแรนโนซอรัส McRae มีชีวิตอยู่ก่อนยุคLancian (ก่อน 67 ล้านปีก่อน) โดยพิจารณาจากความสัมพันธ์ที่เป็นไปได้กับฟอสซิลAlamosaurus [ 90 ]

Dalman et al. (2024) เสนอชื่อใหม่ว่าTyrannosaurus mcraeensisสำหรับตัวอย่างต้นแบบ (NMMNH P-3698) โดยอ้างอิงถึงกลุ่มหิน McRae ซึ่งเป็นชั้นหินที่ Hall Lake Formation เป็นส่วนหนึ่ง ตัวอย่างต้นแบบของT. mcraeensisพบในชั้นหินที่มีอายุเก่ากว่าช่วงเวลาที่ยอมรับกันของT. rex ประมาณสองสามล้านปี ซึ่ง T. rex มีชีวิตอยู่ในช่วงปลายยุค Maastrichtian ชั้นหินเหล่านี้ได้รับการประเมินอายุเบื้องต้นว่าอยู่ระหว่าง 72.7 ถึง 70.9 ล้านปี ซึ่งสอดคล้องกับช่วงปลายยุค Campanian หรือต้นยุค Maastrichtian [ 66 ]อย่างไรก็ตาม ในบทคัดย่อ การประชุมปี 2024 ชั้นหินทางธรณีวิทยาเฉพาะที่พบT. mcraeensis นั้น คาดว่ามีอายุน้อยกว่า 69.0 ± 0.4 ล้านปี และเก่ากว่า 66.0 ล้านปี โดยอิงจากหินทรายจากแหล่งฟอสซิลเดียวกัน[ 91 ]ซึ่งแสดงให้เห็นว่าอายุของT. mcraeensisนั้นใกล้เคียงกับT. rexมากกว่าที่เคยคิดไว้[ 79 ] T. mcraeensisมีความยาวประมาณ 12 เมตร (39 ฟุต) ซึ่งมีขนาดใกล้เคียงกับT. rex ตัวเต็มวัย ทั้งสองชนิดแตกต่างกันที่ลักษณะของกะโหลกศีรษะ ในบรรดาลักษณะเหล่านี้ ขากรรไกรล่างของT. mcraeensisนั้นยาวกว่าและมีคางที่ไม่เด่นชัด และขากรรไกรล่างตื้นกว่าของT. rexซึ่งบ่งชี้ว่ามีแรงกัดที่อ่อนกว่า ฟันก็ทู่กว่าและแบนกว่าด้านข้าง ในขณะที่สันหลังเบ้าตาไม่เด่นชัด ในทำนองเดียวกัน กายวิภาคของโครงกระดูกแสดงให้เห็นลักษณะร่วมกันกับTarbosaurusและZhuchengtyrannus [ 66 ] [ 92 ]
ความถูกต้องของT. mcraeensisถูกตั้งคำถามโดยนักวิจัยคนอื่นๆ ในปี 2025 มอร์ริสันและคณะได้กล่าวถึงความไม่แน่นอนในการประมาณอายุของT. mcraeensis ในครั้งก่อน ผู้เขียนโต้แย้งว่า การรวมฟอสซิลไททาโนซอรัส (เช่นAlamosaurus ) เป็นตัวบ่งชี้ทางชีวธรณีวิทยาของสัตว์ในชั้นหิน Hall Lake Formation ทำให้การกำหนดอายุเป็นปัญหา เนื่องจากฟอสซิลดังกล่าวมีความแปรปรวนอย่างมากในอัตราการตกตะกอนที่สันนิษฐานไว้ การศึกษายังชี้ให้เห็นถึงความไม่แน่นอนในรูปร่างของ ตัวอย่างต้นแบบของ T. mcraeensisซึ่งอยู่นอกเหนือความแปรปรวนที่รู้จักของT. rex อย่างน่าเชื่อถือ ไม่ว่าอายุและความถูกต้องของT. mcraeensis จะเป็นอย่างไร ผู้เขียนสรุปว่าสกุลTyrannosaurusน่าจะมีต้นกำเนิดมาจาก Laramidia และบรรพบุรุษของสกุลนี้อาจอพยพมาจากเอเชีย[ 93 ]ต่อมาในปีนั้น Voris และเพื่อนร่วมงานถือว่าT. mcraeensisเป็นชื่อพ้องรองของT. rexโดยเห็นด้วยกับข้อมูลทางธรณีวิทยาที่มีปัญหา และพบว่าลักษณะการวินิจฉัยที่เสนอทั้งหมดของT. mcraeensisอยู่ในช่วงความแปรผันของตัวอย่างT. rex ที่รู้จัก [ 94 ]ในการศึกษากะโหลกต้นแบบของT. rexนั้น Carr ก็ไม่ถือว่าT. mcraeensisเป็นสายพันธุ์ที่ถูกต้องเช่นกัน โดยเห็นด้วยกับลักษณะที่เสนอของ กะโหลกต้นแบบ T. mcraeensisที่มองเห็นได้ทั่วทั้งกะโหลกย่อยของT. rex [ 95 ]
นาโนไทแรนนัส

ฟอสซิลไทแรนโนซอร์อื่นๆ ที่พบในชั้นหินเดียวกันกับT. rexเดิมทีถูกจัดประเภทเป็น taxa ที่แยกจากกัน รวมถึงAublysodonและAlbertosaurus megagracilis [ 60 ] โดย ฟอสซิล หลังนี้ได้รับการตั้งชื่อว่าDinotyrannus megagracilisในปี 1995 [ 96 ]ปัจจุบันฟอสซิลเหล่านี้ถือกันโดยทั่วไปว่าเป็นของ T. rex วัยเยาว์[ 97 ]ยกเว้นกะโหลกขนาดเล็กแต่เกือบสมบูรณ์จากมอนแทนา ซึ่งมีความยาว 57.2 เซนติเมตร (1.88 ฟุต) กะโหลกศีรษะชิ้นนี้CMNH 7541 เดิมทีถูกจัดประเภทเป็นสายพันธุ์Gorgosaurus ( G. lancensis ) โดยCharles W. Gilmoreในปี 1946 [ 98 ]ในปี 1988 ตัวอย่างนี้ได้รับการอธิบายใหม่โดยRobert T. Bakker , Michael Williams ซึ่งในขณะนั้นเป็นภัณฑารักษ์ด้านบรรพชีวินวิทยาที่พิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติคลีฟแลนด์ และPhil Currieซึ่งเป็นสถานที่เก็บรักษาตัวอย่างดั้งเดิมและจัดแสดงอยู่ในปัจจุบัน การวิจัยเบื้องต้นของพวกเขาบ่งชี้ว่ากระดูกกะโหลกศีรษะเชื่อมติดกัน และดังนั้นจึงแสดงถึงตัวอย่างที่โตเต็มวัย ด้วยเหตุนี้ Bakker และเพื่อนร่วมงานจึงกำหนดกะโหลกศีรษะนี้ให้กับสกุลใหม่ที่ชื่อว่าNanotyrannus (หมายถึง "ทรราชแคระ" เนื่องจากขนาดตัวที่โตเต็มวัยที่ดูเหมือนจะเล็ก) [ 99 ]อย่างไรก็ตาม ในปี 1999 การวิเคราะห์อย่างละเอียดโดยThomas Carr เปิดเผยว่าตัวอย่างนี้เป็นของวัยเยาว์ ทำให้ Carr และนักบรรพชีวินวิทยา คนอื่นๆ อีกหลายคนพิจารณาว่าเป็นT. rex วัยเยาว์ [ 100 ] [ 101 ]

ในปี 2001 ซากดึกดำบรรพ์ของทีเร็กซ์วัยเยาว์ที่สมบูรณ์กว่า (ได้รับฉายาว่า "เจน" หมายเลขทะเบียน BMRP 2002.4.1) ซึ่งคาดว่าน่าจะเป็นสายพันธุ์เดียวกับ ซากดึกดำบรรพ์ Nanotyrannus ดั้งเดิม ได้ถูกค้นพบ การค้นพบนี้กระตุ้นให้มีการประชุมเกี่ยวกับทีเร็กซ์ โดยมุ่งเน้นไปที่ประเด็นความถูกต้องของNanotyrannusซึ่งจัดขึ้นที่พิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติเบอร์พีในปี 2005 นักบรรพชีวินวิทยาหลายคนที่เคยตีพิมพ์ความเห็นว่าN. lancensisเป็นสายพันธุ์ที่ถูกต้อง รวมถึงเคอร์รีและวิลเลียมส์ มองว่าการค้นพบ "เจน" เป็นการยืนยันว่าNanotyrannus นั้นเป็น ทีเร็กซ์ วัย เยาว์อย่างแท้จริง[ 102 ] [ 103 ] [ 104 ]ปีเตอร์ ลาร์สันยังคงสนับสนุนสมมติฐานที่ว่าNanotyrannus lancensisเป็นสายพันธุ์ที่แยกจากกันแต่มีความสัมพันธ์ใกล้ชิดกัน และยังโต้แย้งว่าStygivenator (LACM 28471) ซึ่งโดยทั่วไปถือว่าเป็นลูกของTyrannosaurus rexอาจเป็นNanotyrannusที่ มีอายุน้อยกว่า [ 105 ] [ 106 ]
ในช่วงปลายปี 2011 มีรายงานข่าวเกี่ยวกับการค้นพบซากดึกดำบรรพ์ของเทโรพอดที่สมบูรณ์เกือบทั้งหมดชิ้นใหม่ในปี 2006 ซึ่งพบอยู่เคียงข้างซากดึกดำบรรพ์ ของเซราทอ ปซิด โรเบิร์ต บักเกอร์และปีเตอร์ ลาร์สันได้ทำการศึกษาซากดึกดำบรรพ์ดังกล่าวในสถานที่จริง และระบุว่าเซราทอป เซียนคือไทรเซราทอปส์ และเทโรพอดคือนาโนไทแรนนัส[ 107 ]เป็นไปไม่ได้ที่จะระบุว่าซากดึกดำบรรพ์ของเทโรพอดซึ่งมีชื่อเล่นว่า "บลัดดี แมรี" นั้นแตกต่างจากที.เร็กซ์หรือ ไม่ เนื่องจากซากดึกดำบรรพ์ยังคงอยู่ในมือของเอกชนจนถึงปี 2020 เมื่อศาลฎีกาแห่งรัฐมอนแทนา ตัดสิน ให้กรรมสิทธิ์ในซากดึกดำบรรพ์ตกเป็นของแมรี แอนน์และไลจ์ เมอร์เรย์ เจ้าของที่ดิน ซึ่งตกลงที่จะขายซากดึกดำบรรพ์คู่ให้กับพิพิธภัณฑ์ในสหรัฐอเมริกา[ 108 ] ซากดึกดำบรรพ์ดังกล่าว ถูกซื้อโดยพิพิธภัณฑ์วิทยาศาสตร์ธรรมชาติแห่งนอร์ทแคโรไลนาในปี 2020 หลังจากนั้นพิพิธภัณฑ์ได้สร้างห้องปฏิบัติการวิจัยเพิ่มเติมเพื่อเตรียมศึกษา และจัดแสดงซากดึกดำบรรพ์[ 109 ] [ 110 ]ในปี 2025 นักบรรพชีวินวิทยาLindsay Zannoและ James Napoli ได้ตีพิมพ์คำอธิบายเบื้องต้นของตัวอย่าง "Bloody Mary" ซึ่งเป็นส่วนหนึ่งของการแก้ไขสกุลNanotyrannus อย่างกว้างขวาง พวกเขาระบุว่าตัวอย่างนี้เป็นNanotyrannus ที่มีโครงกระดูกโตเต็มวัย และได้ให้หลักฐานหลายประการที่สนับสนุนความถูกต้องของสกุลนี้ พวกเขายังได้อธิบายชนิดที่สองN. lethaeusโดยอิงจากตัวอย่าง Jane เนื่องจากมีความแตกต่างที่เห็นได้ชัดระหว่างมันกับN. lancensis [ 24 ]
บรรพชีววิทยา
ประวัติชีวิต

การระบุตัวอย่างหลายชิ้นว่าเป็นทีเร็กซ์ วัยเยาว์ ทำให้นักวิทยาศาสตร์สามารถบันทึก การเปลี่ยนแปลง ทางพัฒนาการของสายพันธุ์ ประเมินอายุขัย และกำหนดอัตราการเติบโตของสัตว์ได้ ตัวอย่างต้นแบบ ของ Stygivenator ( LACM 28471 หรือ "เทโรพอดแห่งจอร์แดน") ซึ่งอาจเป็นทีเร็กซ์ วัยเยาว์ คาดว่ามีน้ำหนักเพียง 30 กก. (66 ปอนด์) ในขณะที่ตัวเต็มวัยที่ใหญ่ที่สุด เช่นFMNH PR2081 (ซู) น่าจะมีน้ำหนักประมาณ 5,650 กก. (12,460 ปอนด์) การวิเคราะห์ ทางเนื้อเยื่อวิทยาของกระดูกทีเร็กซ์แสดงให้เห็นว่า LACM 28471 มีอายุเพียง 2 ปีเมื่อตาย ซูได้รับการประเมินอายุเบื้องต้นที่ 28 ปี ซึ่งในขณะนั้นถือว่าใกล้เคียงกับอายุสูงสุดของสายพันธุ์[ 39 ]แต่การศึกษาล่าสุดประเมินอายุของซูไว้ที่ 27–33 ปี[ 111 ]ในทำนองเดียวกันTrix (RGM 792.000)คาดว่ามีอายุอย่างน้อย 30 ปี ณ เวลาที่เสียชีวิต[ 112 ]
การศึกษาเนื้อเยื่อวิทยาช่วยให้สามารถกำหนดอายุของตัวอย่างอื่นๆ ได้ด้วย สามารถสร้างกราฟแสดงการเจริญเติบโตได้เมื่อนำอายุของตัวอย่างต่างๆ มาพล็อตลงบนกราฟพร้อมกับมวลของพวกมัน กราฟการเจริญเติบโต ของทีเร็กซ์มีลักษณะเป็นรูปตัว S โดยลูกทีเร็กซ์จะมีน้ำหนักต่ำกว่า 1,800 กิโลกรัม (4,000 ปอนด์) จนกระทั่งอายุประมาณ 14 ปี เมื่อขนาดตัวเริ่มเพิ่มขึ้นอย่างรวดเร็ว ในช่วงการเจริญเติบโตอย่างรวดเร็วนี้ทีเร็กซ์ วัยเยาว์ จะมีน้ำหนักเพิ่มขึ้นเฉลี่ย 600 กิโลกรัม (1,300 ปอนด์) ต่อปีเป็นเวลาสี่ปีถัดไป เมื่ออายุ 18 ปี กราฟจะทรงตัวอีกครั้ง ซึ่งบ่งชี้ว่าการเจริญเติบโตชะลอตัวลงอย่างมาก ตัวอย่างเช่น น้ำหนักของซู อายุ 28 ปี ต่างจากทีเร็กซ์จากแคนาดา อายุ 22 ปี เพียง 600 กิโลกรัม (1,300 ปอนด์) เท่านั้น ( RTMP 81.12.1) [ 39 ]การศึกษาทางเนื้อเยื่อวิทยาในปี 2004 ที่ดำเนินการโดยนักวิจัยหลายคนยืนยันผลลัพธ์เหล่านี้ โดยพบว่าการเจริญเติบโตอย่างรวดเร็วเริ่มชะลอตัวลงเมื่ออายุประมาณ 16 ปี[ 113 ]
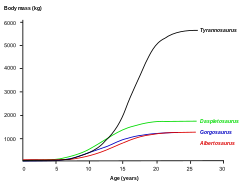
การศึกษาโดยฮัทชินสันและเพื่อนร่วมงานในปี 2011 ยืนยันวิธีการประมาณค่าก่อนหน้านี้โดยทั่วไป แต่การประมาณอัตราการเติบโตสูงสุดของพวกเขาสูงกว่าอย่างมีนัยสำคัญ พบว่า "อัตราการเติบโตสูงสุดของ T. rex ในช่วงระยะทวีคูณคือ 1790 กก./ปี" [ 28 ]แม้ว่าผลลัพธ์เหล่านี้จะสูงกว่าการประมาณค่าก่อนหน้านี้มาก ผู้เขียนตั้งข้อสังเกตว่าผลลัพธ์เหล่านี้ลดความแตกต่างอย่างมากระหว่างอัตราการเติบโตจริงกับอัตราที่คาดหวังจากสัตว์ที่มีขนาดเท่านี้[ 28 ]การเปลี่ยนแปลงอย่างกะทันหันของอัตราการเติบโตในช่วงท้ายของการเติบโตอย่างรวดเร็วอาจบ่งชี้ถึงวุฒิภาวะทางกายภาพ ซึ่งเป็นสมมติฐานที่ได้รับการสนับสนุนจากการค้นพบเนื้อเยื่อไขกระดูกในกระดูกต้นขา ของ T. rexอายุ 16 ถึง 20 ปีจากมอนแทนา ( MOR 1125 หรือที่รู้จักกันในชื่อB-rex ) เนื้อเยื่อไขกระดูกพบได้เฉพาะในนกตัวเมียในช่วงตกไข่ ซึ่งบ่งชี้ว่า B-rex อยู่ในวัยเจริญพันธุ์[ 114 ]การศึกษาเพิ่มเติมบ่งชี้ว่าตัวอย่างนี้มีอายุ 18 ปี[ 115 ]ในปี 2016 แมรี ฮิกบี ชไวเซอร์ และลินด์เซย์ แซนโน และเพื่อนร่วมงานได้ยืนยันในที่สุดว่าเนื้อเยื่ออ่อนภายในกระดูกต้นขาของ MOR 1125 คือเนื้อเยื่อไขกระดูก ซึ่งยืนยันตัวตนของตัวอย่างว่าเป็นเพศเมีย การค้นพบ เนื้อเยื่อ ไขกระดูกภายในไทแรนโนซอรัสอาจมีคุณค่าในการกำหนดเพศของไดโนเสาร์สายพันธุ์อื่นในการตรวจสอบในอนาคต เนื่องจากองค์ประกอบทางเคมีของเนื้อเยื่อไขกระดูกนั้นชัดเจน[ 116 ]ไทแรนโนซอริเดชนิดอื่น ๆ แสดงเส้นโค้งการเจริญเติบโตที่คล้ายคลึงกันมาก แม้ว่าจะมีอัตราการเจริญเติบโตที่ต่ำกว่าซึ่งสอดคล้องกับขนาดตัวเต็มวัยที่ต่ำกว่า[ 117 ]
งานวิจัยเพิ่มเติมที่ตีพิมพ์ในปี 2020 โดยวูดเวิร์ดและคณะ ในวารสารScience Advancesระบุว่า ในระหว่างการเจริญเติบโตจากวัยเยาว์ไปสู่วัยผู้ใหญ่ไทแรนโนซอรัสสามารถชะลอการเจริญเติบโตเพื่อรับมือกับปัจจัยด้านสิ่งแวดล้อม เช่น การขาดแคลนอาหาร โดยอ้างอิงจากตัวอย่าง BMRP 2002.4.1 และ BMRP 2006.4.4 ซึ่งมีอายุระหว่าง 13 ถึง 15 ปี และเก็บรักษาไว้ที่พิพิธภัณฑ์เบอร์พีในรัฐอิลลินอยส์ ซึ่งในขณะนั้นเรียกว่า ตัวอย่าง ไทแรนโน ซอรัสวัยเยาว์ ผู้เขียนเสนอว่าอัตราการเจริญเติบโตของไทแรนโนซอรัสขึ้นอยู่กับความอุดมสมบูรณ์ของทรัพยากร งานวิจัยนี้ยังแสดงให้เห็นว่าในสภาพแวดล้อมที่เปลี่ยนแปลงไปไทแรนโนซอรัสมีความเหมาะสมเป็นพิเศษกับสภาพแวดล้อมที่เปลี่ยนแปลงไปในแต่ละปีในแง่ของความอุดมสมบูรณ์ของทรัพยากร ซึ่งบ่งชี้ว่าสัตว์นักล่าขนาดกลางชนิดอื่นอาจมีปัญหาในการเอาชีวิตรอดในสภาพที่โหดร้ายเช่นนี้ และอธิบายถึงการแบ่งส่วนของนิเวศวิทยา (niche partitioning) ระหว่างไทแรนโนซอรัสวัยเยาว์และวัยผู้ใหญ่ การศึกษายังชี้ให้เห็นเพิ่มเติมว่าTyrannosaurusและNanotyrannusเป็นชื่อเดียวกัน เนื่องจากการวิเคราะห์วงแหวนการเจริญเติบโตในกระดูกของตัวอย่างทั้งสองที่ศึกษา[ 118 ] [ 119 ]
ในปี 2026 Woodward, Myhrvold และ Horner ได้ทำการวิเคราะห์ทางเนื้อเยื่อวิทยาอย่างครอบคลุมของตัวอย่างไทแรนโนซอรัส 17 ตัวอย่าง และได้ให้เหตุผลว่าไทแรนโนซอรัสอาจมีอัตราการเติบโตประจำปีที่ค่อยเป็นค่อยไปมากกว่าที่ระบุไว้ในการศึกษาและหลักฐานก่อนหน้านี้เกี่ยวกับระยะวัยรุ่นที่ยาวนาน โดยถึงขนาดสูงสุดเมื่ออายุประมาณ 35-40 ปี ขีดจำกัดบนของการประมาณอัตราการเติบโตประจำปีอยู่ที่ประมาณ 43-53 ปี พวกเขายังพบว่าเส้นทางการเติบโตของ BMRP 2002.4.1 และ BMRP 2006.4.4 ไม่สอดคล้องกับตัวอย่างไทแรนโนซอรัส อื่นๆ ในแบบจำลองเส้นโค้งการเติบโตของพวกเขา แม้ว่าพวกเขาจะยอมรับความเป็นไปได้ที่ตัวอย่างที่ยังไม่โตเต็มวัยเหล่านี้จะเป็น Nanotyrannusตามที่ Zanno และ Napoli (2025) เสนอไว้ แต่พวกเขาก็ตั้งข้อสังเกตว่าความไม่สอดคล้องกันของตัวอย่างเหล่านี้ที่สังเกตได้ในเส้นโค้งการเติบโตไม่จำเป็นต้องมีน้ำหนักต่อข้อเสนอที่ว่าNanotyrannusเป็นอนุกรมวิธานที่แตกต่างกัน[ 120 ]
จากการศึกษาพบ ว่า กว่าครึ่งหนึ่งของ ตัวอย่าง ทีเร็กซ์ ที่รู้จักกัน นั้นเสียชีวิตภายในหกปีหลังจากถึงวัยเจริญพันธุ์ ซึ่งเป็นรูปแบบที่พบในไดโนเสาร์วงศ์ไทแรนโนซอรัสชนิดอื่นๆ และในนกและสัตว์เลี้ยงลูกด้วยนมขนาดใหญ่ที่มีอายุยืนยาวบางชนิดในปัจจุบัน สัตว์เหล่านี้มีลักษณะเด่นคืออัตราการตายของลูกอ่อนสูง ตามด้วยอัตราการตายที่ค่อนข้างต่ำในวัยรุ่น อัตราการตายจะเพิ่มขึ้นอีกครั้งหลังจากถึงวัยเจริญพันธุ์ ส่วนหนึ่งเนื่องจากความเครียดจากการสืบพันธุ์ การศึกษาหนึ่งชี้ให้เห็นว่าความหายากของ ฟอสซิล ทีเร็กซ์ วัยเยาว์ นั้นส่วนหนึ่งเกิดจากอัตราการตายของวัยรุ่นที่ต่ำ สัตว์เหล่านี้ไม่ได้ตายเป็นจำนวนมากในช่วงวัยดังกล่าว จึงไม่ค่อยพบฟอสซิล ความหายากนี้อาจเกิดจากความไม่สมบูรณ์ของบันทึกฟอสซิลหรืออคติของนักสะสมฟอสซิลที่เลือกเฉพาะตัวอย่างที่ใหญ่กว่าและน่าตื่นตาตื่นใจกว่า[ 117 ]ในการบรรยายเมื่อปี 2556 โทมัส โฮลท์ซ จูเนียร์ เสนอว่าไดโนเสาร์ "มีชีวิตอยู่อย่างรวดเร็วและตายเร็ว" เพราะพวกมันสืบพันธุ์ได้อย่างรวดเร็ว ในขณะที่สัตว์เลี้ยงลูกด้วยนมมีอายุขัยยาวนานเพราะใช้เวลานานกว่าในการสืบพันธุ์[ 121 ]เกรกอรี เอส. พอล ก็เขียนเช่นกันว่าไทแรนโนซอรัสสืบพันธุ์ได้อย่างรวดเร็วและตายเร็ว แต่ให้เหตุผลว่าอายุขัยที่สั้นของพวกมันเกิดจากชีวิตที่อันตรายที่พวกมันต้องเผชิญ[ 122 ]
ผิวหนังและอาจมีลักษณะเป็นขนคล้ายเส้นใย

การค้นพบไดโนเสาร์มีขน นำไปสู่การถกเถียงว่า ไทแรนโนซอรัสอาจมีขนหรือไม่ และมีมากน้อยเพียงใด[ 123 ] [ 124 ]โครงสร้างที่เป็นเส้นใย ซึ่งโดยทั่วไปถือว่าเป็นสารตั้งต้นของขน ได้รับการรายงานใน ไดลอง พาราโดซัส ไทแรนโนซอรัสขนาดเล็ก ซึ่งเป็นไทแรนโนซอรัสพื้นฐานจากยุคครีเทเชียสตอนต้นของแหล่ง หินอี้เซียน ในประเทศจีนในปี 2547 [ 125 ]เนื่องจาก ร่องรอย ของผิวหนังของไทแรนโนซอรัสขนาดใหญ่ที่รู้จักในเวลานั้นแสดงหลักฐานของเกล็ดนักวิจัยที่ศึกษาไดลองจึงคาดการณ์ว่าขนที่เป็นฉนวนอาจหายไปในสายพันธุ์ขนาดใหญ่เนื่องจากอัตราส่วนพื้นผิวต่อปริมาตรที่เล็กกว่า[ 125 ]การค้นพบสายพันธุ์ยักษ์Yutyrannus huali ในเวลาต่อมา ซึ่งมาจาก Yixian เช่นกัน แสดงให้เห็นว่าแม้แต่ไดโนเสาร์วงศ์ Tyrannosauria ขนาดใหญ่บางตัวก็มีขนปกคลุมร่างกายส่วนใหญ่ ทำให้เกิดข้อสงสัยเกี่ยวกับสมมติฐานที่ว่าขนเป็นลักษณะที่เกี่ยวข้องกับขนาด[ 126 ]การศึกษาในปี 2017 ได้ทบทวนรอยประทับผิวหนังที่รู้จักของไดโนเสาร์วงศ์ Tyrannosauria รวมถึงตัวอย่างของTyrannosaurusที่มีชื่อเล่นว่า "Wyrex" (HMNS 2006.1743.01 เดิมชื่อ BHI 6230) ซึ่งเก็บรักษาเกล็ดโมเสกเป็นหย่อมๆ บนหาง สะโพก และคอ[ 123 ]การศึกษาสรุปว่าการปกคลุมด้วยขนของไดโนเสาร์วงศ์ Tyrannosauria ขนาดใหญ่ เช่นTyrannosaurusนั้น หากมีอยู่ ก็จะจำกัดอยู่เฉพาะด้านบนของลำตัวเท่านั้น[ 123 ]
บทคัดย่อการประชุมที่ตีพิมพ์ในปี 2016 เสนอว่าเทโรพอด เช่นไทแรนโนซอรัสมีฟันบนปกคลุมด้วยริมฝีปาก แทนที่จะเป็นฟันเปลือยอย่างที่พบในจระเข้โดยอ้างอิงจากการมีอยู่ของเคลือบฟันซึ่งตามการศึกษาพบว่าจำเป็นต้องคงความชุ่มชื้น ซึ่งเป็นปัญหาที่สัตว์น้ำอย่างจระเข้ไม่ได้เผชิญ[ 55 ]อย่างไรก็ตาม มีการวิพากษ์วิจารณ์ที่สนับสนุนแนวคิดเรื่องริมฝีปาก โดยการศึกษาเชิงวิเคราะห์ในปี 2017 เสนอว่าไทแรนโนซอริเดมีเกล็ดขนาดใหญ่และแบนบนจมูกแทนที่จะเป็นริมฝีปากเหมือนจระเข้ในปัจจุบัน[ 53 ] [ 127 ]แต่จระเข้มีผิวหนังที่แตกเป็นรอยและมีเคราติน ไม่ใช่เกล็ดแบน จากการสังเกตความขรุขระเป็นเนินของไทแรนโนซอริเด และเปรียบเทียบกับกิ้งก่าที่ยังมีชีวิตอยู่ นักวิจัยพบว่าไทแรนโนซอริเดมีเกล็ดแบบเกล็ดงูมากกว่าผิวหนังแบบจระเข้[ 56 ] [ 128 ]
ในปี 2023 คัลเลนและเพื่อนร่วมงานสนับสนุนแนวคิดที่ว่าไดโนเสาร์กินเนื้อจำพวกเทโรพอด เช่น ไทแรนโนซอริเด มีริมฝีปากโดยพิจารณาจากรูปแบบทางกายวิภาค เช่น รูบนใบหน้าและขากรรไกร ซึ่งคล้ายคลึงกับสัตว์เลื้อยคลาน ในปัจจุบัน เช่นจิ้งจกมอนิเตอร์หรืออีกัวน่าทะเล มากกว่า จระเข้ในปัจจุบันเช่นอัลลิเกเตอร์การเปรียบเทียบฟันของดาสเปลโตซอรัสและอัลลิเกเตอร์อเมริกันแสดงให้เห็นว่าเคลือบฟันของไทแรนโนซอริเดไม่มีการสึกหรออย่างมีนัยสำคัญ ในขณะที่ฟันของจระเข้ในปัจจุบันมีการสึกกร่อนที่ด้านริมฝีปากและสึกหรออย่างมาก นี่แสดงให้เห็นว่าฟันของเทโรพอดน่าจะถูกทำให้ชุ่มชื้นด้วยริมฝีปาก จากความสัมพันธ์ระหว่างความชุ่มชื้นและความต้านทานต่อการสึกหรอ ผู้เขียนจึงโต้แย้งว่าไม่น่าเป็นไปได้ที่ฟันของเทโรพอด รวมถึงไทแรนโนซอริเด จะยังคงไม่สึกหรอเมื่อสัมผัสกับอากาศเป็นเวลานาน เพราะการรักษาความชุ่มชื้นนั้นทำได้ยาก ผู้เขียนยังทำการวิเคราะห์การถดถอยเพื่อแสดงความสัมพันธ์ระหว่างความสูงของฟันและความยาวของกะโหลก และพบว่าวารานิดเช่น จิ้งจกมอนิเตอร์จระเข้มีอัตราส่วนความสูงของฟันต่อความยาวของกะโหลกมากกว่าไทแรนโนซอรัส อย่างมาก ซึ่งบ่งชี้ว่าฟันของเทโรพอดไม่ได้ใหญ่เกินไปจนถูกปกคลุมด้วยเนื้อเยื่อภายนอกช่องปากเมื่อปิดปาก[ 57 ]
ความแตกต่างทางเพศ

เมื่อจำนวนตัวอย่างที่รู้จักเพิ่มขึ้น นักวิทยาศาสตร์เริ่มวิเคราะห์ความแปรผันระหว่างแต่ละตัวและค้นพบสิ่งที่ดูเหมือนจะเป็นรูปร่างสองแบบที่แตกต่างกัน หรือมอร์ฟคล้ายกับไดโนเสาร์เทอโรพอดบางชนิด เนื่องจากมอร์ฟหนึ่งมีโครงสร้างที่แข็งแรงกว่า จึงถูกเรียกว่า มอร์ฟ 'แข็งแรง' ในขณะที่อีกมอร์ฟหนึ่งถูกเรียกว่า ' บอบบาง ' ความแตกต่าง ทางสัณฐานวิทยาหลายประการที่เกี่ยวข้องกับมอร์ฟทั้งสองถูกนำมาใช้ในการวิเคราะห์ความแตกต่างทางเพศในทีเร็กซ์โดยปกติแล้วมอร์ฟ 'แข็งแรง' มักถูกสันนิษฐานว่าเป็นเพศเมีย ตัวอย่างเช่นกระดูกเชิงกรานของตัวอย่าง 'แข็งแรง' หลายตัวดูเหมือนจะกว้างกว่า อาจเพื่อให้ไข่ ผ่าน ได้[ 129 ]นอกจากนี้ยังคิดว่าสัณฐานวิทยา 'แข็งแรง' สัมพันธ์กับกระดูกสันหลังส่วนหางข้อแรกที่ลดลง ซึ่งเห็นได้ชัดว่าเพื่อให้ไข่ผ่านออกจากระบบสืบพันธุ์ได้ดังที่เคยมีรายงานผิดพลาดสำหรับจระเข้ [ 130 ]
ในช่วงไม่กี่ปีที่ผ่านมา หลักฐานเกี่ยวกับความแตกต่างทางเพศได้อ่อนลง การศึกษาในปี 2548 รายงานว่าข้ออ้างก่อนหน้านี้เกี่ยวกับความแตกต่างทางเพศในกายวิภาคของกระดูกหางจระเข้นั้นผิดพลาด ทำให้เกิดข้อสงสัยเกี่ยวกับการมีอยู่ของความแตกต่างทางเพศที่คล้ายคลึงกันระหว่างเพศของT. rex [ 131 ]พบกระดูกหางขนาดเต็มบนกระดูกสันหลังส่วนหางชิ้นแรกของซู ซึ่งเป็นตัวอย่างที่แข็งแรงมาก แสดงให้เห็นว่าลักษณะนี้ไม่สามารถใช้เพื่อแยกความแตกต่างระหว่างสองรูปแบบได้อยู่แล้ว เนื่องจากพบตัวอย่างT. rex ตั้งแต่ ซัสแคตเชวันไปจนถึงนิวเม็กซิโกความแตกต่างระหว่างแต่ละตัวอาจบ่งชี้ถึงความแปรผันทางภูมิศาสตร์มากกว่าความแตกต่างทางเพศ ความแตกต่างอาจเกี่ยวข้องกับอายุด้วย โดยตัวอย่างที่ 'แข็งแรง' อาจเป็นสัตว์ที่มีอายุมากกว่า[ 8 ]
มีเพียงตัวอย่าง ไทแรนโนซอรัสเพียงชิ้นเดียวเท่านั้นที่ได้รับการพิสูจน์อย่างแน่ชัดว่าเป็นเพศใด การตรวจสอบ B-rex แสดงให้เห็นถึงการรักษาเนื้อเยื่ออ่อนภายในกระดูกหลายชิ้น เนื้อเยื่อบางส่วนนี้ได้รับการระบุว่าเป็นเนื้อเยื่อไขกระดูก ซึ่งเป็นเนื้อเยื่อพิเศษที่เจริญเติบโตเฉพาะในนกสมัยใหม่เท่านั้น โดยเป็นแหล่งแคลเซียมสำหรับการสร้างเปลือกไข่ในระหว่างการตกไข่เนื่องจากมีเพียงนกตัวเมียเท่านั้นที่วางไข่ เนื้อเยื่อไขกระดูกจึงพบได้ตามธรรมชาติเฉพาะในตัวเมียเท่านั้น แม้ว่าตัวผู้จะสามารถผลิตได้เมื่อได้รับฮอร์โมน สืบพันธุ์ของตัวเมีย เช่นเอสโตรเจนสิ่งนี้ชี้ให้เห็นอย่างชัดเจนว่า B-rex เป็นตัวเมียและเสียชีวิตในระหว่างการตกไข่[ 114 ]การวิจัยล่าสุดแสดงให้เห็นว่าเนื้อเยื่อไขกระดูกไม่เคยพบในจระเข้ซึ่งเชื่อกันว่าเป็นญาติที่ใกล้ชิดที่สุดของไดโนเสาร์ การมีอยู่ร่วมกันของเนื้อเยื่อไขกระดูกในนกและไดโนเสาร์เทอโรพอดอื่นๆ เป็นหลักฐานเพิ่มเติมของ ความสัมพันธ์ ทางวิวัฒนาการ ที่ใกล้ชิด ระหว่างทั้งสอง[ 132 ]
ท่าทาง

เช่นเดียวกับไดโนเสาร์สองขา หลายชนิด ทีเร็กซ์ถูกวาดภาพในอดีตว่าเป็น 'สัตว์สามขาที่มีชีวิต' โดยลำตัวทำมุม 45 องศาหรือน้อยกว่าจากแนวตั้ง และหางลากไปกับพื้น คล้ายกับจิงโจ้แนวคิดนี้มีมาตั้งแต่การสร้างภาพฮาโดรซอรัส ขึ้นใหม่โดย โจเซฟ เลดี้ ในปี 1865 ซึ่งเป็นภาพแรกที่แสดงให้เห็นไดโนเสาร์ในท่าสองขา[ 133 ] ในปี 1915 เฮนรี แฟร์ฟิลด์ ออสบอร์นอดีตประธานพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติอเมริกันเชื่อมั่นว่าสิ่งมีชีวิตนี้ยืนตัวตรง จึงได้เปิดเผยโครงกระดูก ทีเร็กซ์ ที่สมบูรณ์เป็นครั้งแรก ซึ่งจัดเรียงในลักษณะนี้ โครงกระดูกนี้อยู่ในท่าตั้งตรงเป็นเวลา 77 ปี จนกระทั่งถูกรื้อถอนในปี 1992 [ 134 ]
ในปี 1970 นักวิทยาศาสตร์ตระหนักว่าท่าทางนี้ไม่ถูกต้องและสัตว์ที่มีชีวิตไม่สามารถรักษาไว้ได้ เนื่องจากจะส่งผลให้ข้อต่อ หลายข้อ หลุดหรืออ่อนแอลงรวมถึงข้อสะโพกและข้อต่อระหว่างศีรษะกับกระดูกสันหลัง[ 135 ] หุ่น จำลองที่ไม่ถูกต้องของ AMNH เป็นแรงบันดาลใจให้เกิดภาพที่คล้ายคลึงกันในภาพยนตร์และภาพวาดหลายเรื่อง (เช่นภาพจิตรกรรมฝาผนังที่มีชื่อเสียงของRudolph Zallinger เรื่อง The Age of Reptilesในพิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติ Peabodyของมหาวิทยาลัยเยล ) [ 136 ]จนกระทั่งถึงทศวรรษ 1990 เมื่อภาพยนตร์เช่นJurassic Parkได้นำเสนอท่าทางที่ถูกต้องมากขึ้นแก่สาธารณชน[ 137 ]ภาพที่แสดงในพิพิธภัณฑ์ ศิลปะ และภาพยนตร์ในปัจจุบันแสดงให้เห็นT. rexโดยมีลำตัวขนานกับพื้นโดยประมาณและหางยื่นไปด้านหลังลำตัวเพื่อรักษาสมดุลของศีรษะ[ 138 ]
ในการนั่งลงไทแรนโนซอรัสอาจจะถ่ายน้ำหนักไปด้านหลังและวางน้ำหนักลงบนกระดูกหัวหน่าว ซึ่งเป็นส่วนขยายกว้างที่ปลายกระดูกหัวหน่าวในไดโนเสาร์บางชนิด เมื่อน้ำหนักวางอยู่บนกระดูกเชิงกราน มันอาจสามารถขยับขาหลังได้อย่างอิสระ การลุกขึ้นยืนอีกครั้งอาจต้องอาศัยการทรงตัวจากขาหน้าที่เล็กจิ๋ว[ 139 ] [ 135 ]ทฤษฎีการวิดพื้นของนิวแมนซึ่งเป็นที่รู้จักกันในชื่อนี้ยังคงเป็นที่ถกเถียงกันอยู่ อย่างไรก็ตามไทแรนโนซอรัสน่าจะสามารถลุกขึ้นยืนได้หากมันล้ม ซึ่งจะต้องวางแขนขาไว้ต่ำกว่าจุดศูนย์ถ่วง โดยใช้หางเป็นตัวถ่วงดุลที่มีประสิทธิภาพ รอยแตกร้าวที่หายแล้วในขาหน้าถูกนำเสนอเป็นทั้งหลักฐานว่าแขนไม่น่าจะมีประโยชน์มากนัก[ 140 ] [ 141 ]และเป็นหลักฐานว่าแขนนั้นถูกใช้งานและได้รับบาดเจ็บ[ 142 ]เช่นเดียวกับส่วนอื่นๆ ของร่างกาย
อาวุธ

เมื่อ มีการค้นพบ ทีเร็กซ์เป็นครั้งแรกกระดูกต้นแขนเป็นส่วนประกอบเพียงส่วนเดียวของแขนขาหน้าที่รู้จัก[ 10 ]สำหรับโครงกระดูกที่จัดแสดงครั้งแรกซึ่งสาธารณชนได้เห็นในปี 1915 ออสบอร์นได้เปลี่ยนแขนขาหน้าที่ยาวกว่าและมีสามนิ้วเหมือนกับของอัลโลซอรัส [ 5 ] หนึ่งปีก่อนหน้า นั้น ลอว์เรนซ์ แลมบ์ได้อธิบายถึงแขนขาหน้าที่สั้นและมีสองนิ้วของกอร์โกซอรัสซึ่งเป็น ญาติใกล้ชิด [ 143 ]สิ่งนี้ชี้ให้เห็นอย่างชัดเจนว่าทีเร็กซ์มีแขนขาหน้าที่คล้ายคลึงกัน แต่สมมติฐาน นี้ ยังไม่ได้รับการยืนยันจนกระทั่ง มีการระบุแขนขาหน้า ของทีเร็กซ์ ที่สมบูรณ์เป็นครั้งแรก ในปี 1989 ซึ่งเป็นของ MOR 555 ("แวนเคลเร็กซ์") [ 144 ] [ 145 ]ซากของซูยังรวมถึงแขนขาหน้าที่สมบูรณ์ด้วย[ 8 ] แขน ของทีเร็กซ์มีขนาดเล็กมากเมื่อเทียบกับขนาดตัวโดยรวม โดยมีความยาวเพียง 1 เมตร (3.3 ฟุต) และนักวิชาการบางคนเรียกมันว่า อวัยวะ ที่เหลืออยู่อย่างไรก็ตาม กระดูกแสดงให้เห็นพื้นที่ขนาดใหญ่สำหรับ การยึดเกาะ ของกล้ามเนื้อซึ่งบ่งชี้ถึงความแข็งแรงอย่างมาก สิ่งนี้ได้รับการยอมรับตั้งแต่ปี 1906 โดยออสบอร์น ซึ่งคาดการณ์ว่าแขนขาหน้าอาจถูกใช้เพื่อจับคู่ระหว่างการผสมพันธุ์[ 12 ]นิวแมน (1970) เสนอว่าแขนขาหน้าถูกใช้เพื่อช่วยให้ไทแรนโนซอรัสลุกขึ้นจากท่านอนคว่ำ[ 135 ]ตั้งแต่นั้นมา มีการเสนอหน้าที่อื่นๆ แม้ว่านักวิชาการบางคนจะพบว่าไม่น่าเป็นไปได้[ 141 ] Padian (2022) โต้แย้งว่าการลดขนาดของแขนในไทแรนโนซอริเดไม่ได้มีหน้าที่เฉพาะเจาะจง แต่เป็นการปรับตัวรอง โดยระบุว่าเมื่อไทแรนโนซอริเดพัฒนากะโหลกและขากรรไกรให้ใหญ่ขึ้นและแข็งแรงขึ้น แขนก็จะเล็กลงเพื่อหลีกเลี่ยงการถูกกัดหรือฉีกขาดโดยตัวอื่น โดยเฉพาะอย่างยิ่งในระหว่างการหาอาหารเป็นกลุ่ม[ 141 ]

อีกความเป็นไปได้หนึ่งคือ แขนขาหน้าอาจใช้จับเหยื่อที่ดิ้นรนขณะที่มันถูกฆ่าโดยขากรรไกรขนาดมหึมาของทีเร็กซ์ สมมติฐานนี้อาจได้รับการสนับสนุนจาก การวิเคราะห์ทางชีวกลศาสตร์ กระดูกแขนขาหน้า ของ ทีเร็กซ์ มีกระดูกชั้น นอกที่หนามาก ซึ่งตีความได้ว่าเป็นหลักฐานว่ามันถูกพัฒนาขึ้นมาเพื่อรับน้ำหนักมาก กล้ามเนื้อ ไบเซปส์ ของ ทีเร็กซ์ ที่โตเต็มวัย สามารถยกน้ำหนักได้ถึง 199 กิโลกรัม (439 ปอนด์) ด้วยตัวเอง กล้ามเนื้ออื่นๆ เช่น กล้ามเนื้อเบรเคียลิสจะทำงานร่วมกับกล้ามเนื้อไบเซปส์เพื่อให้การงอข้อศอกมีประสิทธิภาพมากยิ่งขึ้น กล้ามเนื้อ ไบเซปส์ของทีเร็กซ์ มีกำลังมากกว่ากล้ามเนื้อไบเซป ส์ของมนุษย์ถึง 3.5 เท่า แขนท่อนล่างของ ทีเร็กซ์มีการเคลื่อนไหวที่จำกัด โดยข้อต่อไหล่และข้อศอกอนุญาตให้เคลื่อนไหวได้เพียง 40 และ 45 องศาตามลำดับ ในทางตรงกันข้าม ข้อต่อสองข้อเดียวกันในไดโนนิคัสช่วยให้เคลื่อนไหวได้ถึง 88 และ 130 องศา ตามลำดับ ในขณะที่แขนของมนุษย์สามารถหมุนได้ 360 องศาที่ไหล่และเคลื่อนไหวได้ 165 องศาที่ข้อศอก โครงสร้างที่แข็งแรงของกระดูกแขน ความแข็งแรงของกล้ามเนื้อ และช่วงการเคลื่อนไหวที่จำกัด อาจบ่งชี้ถึงระบบที่วิวัฒนาการมาเพื่อยึดเกาะให้แน่นแม้จะเผชิญกับแรงกดดันจากสัตว์เหยื่อที่ดิ้นรน ในคำอธิบายทางวิทยาศาสตร์โดยละเอียดครั้งแรกเกี่ยวกับแขนขาหน้าของไทแรนโนซอรัส นักบรรพชีวินวิทยา เคนเนธ คาร์เพนเตอร์ และแมตต์ สมิธ ได้ปฏิเสธความคิดที่ว่าแขนขาหน้าไม่มีประโยชน์หรือว่าไทแรนโนซอรัสเป็นสัตว์กินซากโดยสมบูรณ์[ 146 ]
แนวคิดที่ว่าแขนทำหน้าที่เป็นอาวุธเมื่อล่าเหยื่อได้รับการเสนอโดยSteven M. Stanleyซึ่งแนะนำว่าแขนใช้สำหรับฟันเหยื่อ โดยเฉพาะอย่างยิ่งการใช้กรงเล็บเพื่อสร้างบาดแผลยาวและลึกให้กับเหยื่ออย่างรวดเร็ว[ 147 ] Padian ปฏิเสธแนวคิดนี้ โดยโต้แย้งว่า Stanley สรุปโดยอิงจากขนาดและช่วงการเคลื่อนไหวของแขนขาหน้าที่ประเมินไม่ถูกต้อง[ 141 ]
การควบคุมอุณหภูมิร่างกาย

ไทแรนโนซอรัสเช่นเดียวกับไดโนเสาร์ส่วนใหญ่ เคยถูกคิดว่ามีการเผา ผลาญแบบเอคโทเท อร์มิก ("สัตว์เลือดเย็น") เหมือนสัตว์เลื้อยคลานแนวคิดเรื่องเอคโทเทอร์มิกของไดโนเสาร์ถูกท้าทายโดยนักวิทยาศาสตร์อย่างRobert T. BakkerและJohn Ostromในช่วงต้นของ " ยุคฟื้นฟูไดโนเสาร์ " ซึ่งเริ่มต้นในช่วงปลายทศวรรษ 1960 [ 148 ] [ 149 ] มีการอ้างว่า ทีเร็กซ์เองเป็นเอนโดเทอร์มิก ("สัตว์เลือดอุ่น") ซึ่งหมายถึงวิถีชีวิตที่กระฉับกระเฉงมาก[ 37 ]ตั้งแต่นั้นมา นักบรรพชีวินวิทยาหลายคนได้พยายามกำหนดความสามารถของไทแรนโนซอรัสในการควบคุมอุณหภูมิร่างกาย หลักฐานทางเนื้อเยื่อวิทยาของอัตราการเจริญเติบโตที่สูงในทีเร็กซ์ วัยเยาว์ ซึ่งเทียบได้กับสัตว์เลี้ยงลูกด้วยนมและนก อาจสนับสนุนสมมติฐานของการเผาผลาญที่สูง เส้นกราฟการเจริญเติบโตบ่งชี้ว่า เช่นเดียวกับในสัตว์เลี้ยงลูกด้วยนมและนก การเจริญเติบโต ของทีเร็กซ์ส่วนใหญ่จำกัดอยู่ที่สัตว์ที่ยังไม่โตเต็มที่ มากกว่าการเจริญเติบโตที่ไม่แน่นอนที่พบในสัตว์มีกระดูกสันหลัง อื่นๆ ส่วน ใหญ่[ 113 ]
อัตราส่วน ไอโซโทปออกซิเจนในกระดูกฟอสซิลบางครั้งใช้เพื่อกำหนดอุณหภูมิที่กระดูกถูกสะสม เนื่องจากอัตราส่วนระหว่างไอโซโทปบางชนิดมีความสัมพันธ์กับอุณหภูมิ ในตัวอย่างหนึ่ง อัตราส่วนไอโซโทปในกระดูกจากส่วนต่างๆ ของร่างกายบ่งชี้ถึงความแตกต่างของอุณหภูมิไม่เกิน 4 ถึง 5 °C (7 ถึง 9 °F) ระหว่างกระดูกสันหลังของลำตัวและกระดูกหน้าแข้งของขาด้านล่าง ช่วงอุณหภูมิที่แคบระหว่างแกนกลางของร่างกายและส่วนปลายนี้ถูกอ้างโดยนักบรรพชีวินวิทยา Reese Barrick และนักธรณีเคมี William Showers ว่าบ่งชี้ว่าT. rexรักษาอุณหภูมิภายในร่างกายให้คงที่ ( ภาวะอุณหภูมิคงที่ ) และมีกระบวนการเผาผลาญอยู่ระหว่างสัตว์เลื้อยคลานที่มีอุณหภูมิภายนอกและสัตว์เลี้ยงลูกด้วยนมที่มีอุณหภูมิภายใน[ 150 ]นักวิทยาศาสตร์คนอื่นๆ ชี้ให้เห็นว่าอัตราส่วนของไอโซโทปออกซิเจนในฟอสซิลในปัจจุบันไม่จำเป็นต้องแสดงถึงอัตราส่วนเดียวกันในอดีตอันไกลโพ้น และอาจมีการเปลี่ยนแปลงระหว่างหรือหลังการเกิดฟอสซิล ( ไดอะเจเนซิส ) [ 151 ] Barrick และ Showers ได้ปกป้องข้อสรุปของพวกเขาในเอกสารฉบับต่อมา โดยพบผลลัพธ์ที่คล้ายกันในไดโนเสาร์เทอโรพอดอีกชนิดหนึ่งจากทวีปอื่นและเมื่อหลายสิบล้านปีก่อน ( Giganotosaurus ) [ 152 ] ไดโนเสาร์กลุ่ม Ornithischianยังแสดงหลักฐานของภาวะอุณหภูมิคงที่ ในขณะที่ กิ้งก่ากลุ่ม Varanidจากยุคเดียวกันไม่ได้ แสดงหลักฐานดังกล่าว [ 153 ]ในปี 2022 Wiemann และเพื่อนร่วมงานใช้วิธีการที่แตกต่างออกไป นั่นคือการวิเคราะห์สเปกตรัมของสัญญาณลิโปออกซิเดชัน ซึ่งเป็นผลพลอยได้จากการฟอสโฟรีเลชันแบบออกซิเดชันและมีความสัมพันธ์กับอัตราการเผาผลาญ เพื่อแสดงให้เห็นว่าไดโนเสาร์หลายสกุลรวมถึงTyrannosaurusมีการเผาผลาญแบบเอนโดเทอร์มิก เทียบเท่ากับนกในปัจจุบันและสูงกว่าสัตว์เลี้ยงลูกด้วยนม พวกเขายังเสนอแนะว่าการเผาผลาญดังกล่าวเป็นลักษณะดั้งเดิมของไดโนเสาร์ทุกชนิด[ 154 ]
แม้ว่าT. rexจะแสดงหลักฐานของภาวะอุณหภูมิคงที่ แต่ก็ไม่ได้หมายความว่ามันเป็นสัตว์เลือดอุ่นเสมอไป การควบคุมอุณหภูมิดังกล่าวอาจอธิบายได้ด้วยภาวะอุณหภูมิร่างกายสูงเกินปกติ เช่นเดียวกับ เต่าทะเลบางชนิดที่ยังมีชีวิตอยู่[ 155 ] [ 156 ] [ 157 ] คล้ายกับจระเข้ในปัจจุบัน ช่องเปิด (dorsotemporal fenestrae) ในส่วนบนของกะโหลกของTyrannosaurusอาจช่วยในการควบคุมอุณหภูมิได้[ 158 ]
เนื้อเยื่ออ่อน

ในวารสารScience ฉบับเดือนมีนาคม พ.ศ. 2548 แมรี ฮิกบี ชไวเซอร์จากมหาวิทยาลัยแห่งรัฐนอร์ทแคโรไลนาและเพื่อนร่วมงานได้ประกาศการค้นพบเนื้อเยื่ออ่อนจากโพรงไขกระดูกของกระดูกขาที่กลายเป็นฟอสซิลของทีเร็กซ์กระดูกชิ้นนี้ถูกทำให้แตกโดยเจตนา แม้ว่าจะไม่เต็มใจก็ตาม เพื่อการขนส่ง และไม่ได้ถูกเก็บรักษาไว้ตามปกติ โดยเฉพาะอย่างยิ่งเพราะชไวเซอร์หวังที่จะทดสอบหาเนื้อเยื่ออ่อน[ 159 ]ไดโนเสาร์ตัวนี้ได้รับการกำหนดให้เป็นตัวอย่างหมายเลข 1125 ของพิพิธภัณฑ์แห่งเทือกเขาร็อกกี หรือ MOR 1125 ซึ่งก่อนหน้านี้ถูกขุดพบจากชั้นหินเฮลล์ครีก พบ หลอดเลือดที่ยืดหยุ่นและแตก แขนง และเนื้อเยื่อเมทริกซ์ กระดูกที่เป็นเส้นใยแต่ยืดหยุ่นนอกจากนี้ยังพบ โครงสร้างขนาดเล็กที่คล้ายกับ เซลล์เม็ดเลือด อยู่ภายในเมทริกซ์และหลอดเลือด โครงสร้างเหล่านี้มีความคล้ายคลึงกับเซลล์เม็ดเลือดและหลอดเลือด ของนกกระจอกเทศนักวิจัยไม่ทราบว่ากระบวนการที่ไม่รู้จักซึ่งแตกต่างจากการเกิดฟอสซิลตามปกติได้รักษาวัสดุนี้ไว้หรือไม่ หรือวัสดุนี้เป็นของเดิม และพวกเขาระมัดระวังที่จะไม่กล่าวอ้างใดๆ เกี่ยวกับการเก็บรักษา[ 160 ]หากพบว่าเป็นวัสดุดั้งเดิม โปรตีนที่หลงเหลืออยู่อาจใช้เป็นวิธีการคาดเดาเนื้อหา DNA ของไดโนเสาร์ที่เกี่ยวข้องได้โดยอ้อม เนื่องจากโปรตีนแต่ละชนิดมักถูกสร้างขึ้นโดยยีนเฉพาะ การไม่พบหลักฐานก่อนหน้านี้อาจเป็นผลมาจากการที่ผู้คนสันนิษฐานว่าเนื้อเยื่อที่ได้รับการอนุรักษ์นั้นเป็นไปไม่ได้ จึงไม่ได้ค้นหา นับตั้งแต่การค้นพบครั้งแรก มีไทแรนโนซอรัสอีก 2 ตัวและฮาโดรซอรัสอีก 1 ตัวที่ถูกพบว่ามีโครงสร้างคล้ายเนื้อเยื่อดังกล่าว[ 159 ]การวิจัยเกี่ยวกับเนื้อเยื่อบางส่วนที่เกี่ยวข้องได้ชี้ให้เห็นว่านกเป็นญาติใกล้ชิดกับไทแรนโนซอรัสมากกว่าสัตว์สมัยใหม่ชนิดอื่นๆ[ 161 ]นอกจากนี้ยังพบเคมีดั้งเดิมใน MOR 1125 โดยพิจารณาจากการอนุรักษ์องค์ประกอบที่เกี่ยวข้องกับการปรับโครงสร้างและการสะสมใหม่ของกระดูก (กำมะถัน แคลเซียม สังกะสี) ซึ่งแสดงให้เห็นว่าเปลือกกระดูกนั้นคล้ายกับของนกที่ยังมีชีวิตอยู่[ 162 ]
ในการศึกษาที่รายงานในวารสาร Scienceในเดือนเมษายน พ.ศ. 2550 Asara และเพื่อนร่วมงานสรุปว่าร่องรอยของ โปรตีน คอลลาเจน 7 ร่องรอย ที่ตรวจพบใน กระดูก T. rex ที่บริสุทธิ์ นั้นตรงกับที่รายงานในไก่ มากที่สุด รองลงมาคือกบและนิวท์ การค้นพบโปรตีนจากสิ่งมีชีวิตที่มีอายุหลายสิบล้านปี พร้อมกับร่องรอยที่คล้ายกันที่ทีมพบในกระดูกมาสโตดอนที่มีอายุอย่างน้อย 160,000 ปี พลิกมุมมองดั้งเดิมเกี่ยวกับฟอสซิลและอาจทำให้นักบรรพชีวินวิทยาเปลี่ยนจุดสนใจจากการล่ากระดูกไปสู่ชีวเคมี ก่อนการค้นพบเหล่านี้ นักวิทยาศาสตร์ส่วนใหญ่สันนิษฐานว่าการกลายเป็นฟอสซิลได้แทนที่เนื้อเยื่อที่มีชีวิตทั้งหมดด้วยแร่ธาตุที่ไม่ทำปฏิกิริยา นักบรรพชีวินวิทยา Hans Larsson จากมหาวิทยาลัย McGill ในมอนทรีออล ซึ่งไม่ได้เป็นส่วนหนึ่งของการศึกษา เรียกการค้นพบนี้ว่า "เหตุการณ์สำคัญ" และแนะนำว่าไดโนเสาร์อาจ "เข้าสู่สาขาชีววิทยาโมเลกุลและผลักดันบรรพชีวินวิทยาเข้าสู่โลกสมัยใหม่ได้อย่างแท้จริง" [ 163 ]
เนื้อเยื่ออ่อนที่สันนิษฐานไว้นั้นถูกตั้งคำถามโดยโทมัส เคย์ จากมหาวิทยาลัยวอชิงตันและผู้ร่วมเขียนของเขาในปี 2008 พวกเขาโต้แย้งว่าสิ่งที่อยู่ภายในกระดูกไทแรนโนซอร์จริงๆ แล้วคือไบโอฟิล์ม เหนียวๆ ที่สร้างขึ้นโดยแบคทีเรียซึ่งเคลือบช่องว่างที่ครั้งหนึ่งเคยมีหลอดเลือดและเซลล์อยู่[ 164 ]นักวิจัยพบว่าสิ่งที่ก่อนหน้านี้ระบุว่าเป็นเศษเซลล์เม็ดเลือด เนื่องจากมีธาตุเหล็กอยู่ แท้จริงแล้วคือฟราม บอย ด์ ซึ่งเป็นทรงกลมแร่ขนาดเล็กที่มีธาตุเหล็ก พวกเขาพบทรงกลมที่คล้ายกันในฟอสซิลอื่นๆ อีกหลายชนิดจากหลายยุคสมัย รวมถึงแอมโมไนต์ในแอมโมไนต์ พวกเขาพบทรงกลมในตำแหน่งที่ธาตุเหล็กที่บรรจุอยู่ไม่น่าจะมีความสัมพันธ์กับการมีอยู่ของเลือด[ 165 ]ชไวเซอร์ได้วิพากษ์วิจารณ์ข้ออ้างของเคย์อย่างรุนแรงและโต้แย้งว่าไม่มีรายงานหลักฐานว่าไบโอฟิล์มสามารถสร้างท่อกลวงที่แตกแขนงได้เหมือนที่พบในงานวิจัยของเธอ[ 166 ] San Antonio, Schweitzer และเพื่อนร่วมงานได้ตีพิมพ์การวิเคราะห์ในปี 2011 เกี่ยวกับส่วนของคอลลาเจนที่ได้รับการกู้คืน โดยพบว่าเป็นส่วนด้านในของขดคอลลาเจนที่ได้รับการอนุรักษ์ไว้ ซึ่งเป็นไปตามที่คาดไว้จากระยะเวลาการย่อยสลายโปรตีนที่ยาวนาน[ 167 ]งานวิจัยอื่นท้าทายการระบุเนื้อเยื่ออ่อนว่าเป็นไบโอฟิล์ม และยืนยันการค้นพบ "โครงสร้างแตกแขนงคล้ายหลอดเลือด" จากภายในกระดูกฟอสซิล[ 168 ]
ความเร็ว

นักวิทยาศาสตร์ได้กำหนดช่วงความเร็วสูงสุดที่เป็นไปได้สำหรับการวิ่งของ ไทแรนโนซอรัสไว้หลากหลายช่วงโดยส่วนใหญ่จะอยู่ที่ประมาณ 9 เมตรต่อวินาที (32 กม./ชม.; 20 ไมล์/ชม.) แต่ต่ำสุดที่ 4.5–6.8 เมตรต่อวินาที (16–24 กม./ชม.; 10–15 ไมล์/ชม.) และสูงสุดที่ 20 เมตรต่อวินาที (72 กม./ชม.; 45 ไมล์/ชม.) แม้ว่าการวิ่งด้วยความเร็วระดับนี้จะเป็นไปได้ยากมากก็ตามไทแรนโนซอรัสเป็นสัตว์กินเนื้อที่มีขนาดใหญ่และหนัก ดังนั้นจึงไม่น่าจะวิ่งได้เร็วมากนักเมื่อเทียบกับเทโรพอดตัวอื่นๆ เช่นคาร์โนทอรัสหรือไจแกนโนทอซอรัส [ 169 ] นักวิจัยได้อาศัยเทคนิคการประมาณค่าต่างๆ เนื่องจากในขณะที่มีร่องรอยการเดินของเทโรพอดขนาดใหญ่จำนวนมาก แต่ไม่มีร่องรอยใดแสดงหลักฐานการวิ่ง[ 170 ]
รายงานปี 2002 ใช้แบบจำลองทางคณิตศาสตร์ (ตรวจสอบความถูกต้องโดยการนำไปใช้กับสัตว์มีชีวิต 3 ชนิด ได้แก่จระเข้ ไก่และมนุษย์และอีก 8 ชนิด รวมถึงนกอีมูและนกกระจอกเทศ[ 170 ] ) เพื่อวัดมวลกล้ามเนื้อขาที่จำเป็นสำหรับการวิ่งเร็ว (มากกว่า 40 กม./ชม. หรือ 25 ไมล์ต่อชั่วโมง) [ 169 ]นักวิทยาศาสตร์ที่คิดว่าไทแรนโนซอรัสสามารถวิ่งได้ชี้ให้เห็นว่ากระดูกกลวงและลักษณะอื่นๆ ที่จะทำให้ร่างกายเบาลงอาจทำให้น้ำหนักตัวของผู้ใหญ่อยู่ที่ประมาณ 4.5 เมตริกตัน (5.0 ตันสั้น) หรือสัตว์อื่นๆ เช่นนกกระจอกเทศและม้าที่มีขาที่ยาวและยืดหยุ่นได้ สามารถทำความเร็วได้สูงด้วยก้าวที่ช้ากว่าแต่ยาวกว่า[ 170 ] ความเร็วสูงสุดที่เสนอสำหรับ ไทแรนโนซอรัสเกิน 40 กิโลเมตรต่อชั่วโมง (25 ไมล์ต่อชั่วโมง) แต่ถือว่าเป็นไปไม่ได้เพราะจะต้องใช้กล้ามเนื้อขาที่แข็งแรงเป็นพิเศษประมาณ 40–86% ของมวลร่างกายทั้งหมด แม้แต่ความเร็วปานกลางก็ยังต้องใช้กล้ามเนื้อขาขนาดใหญ่ หากมวลกล้ามเนื้อน้อยกว่านี้ การเดินหรือวิ่งเหยาะๆ ก็จะทำได้เพียง 18 กิโลเมตรต่อชั่วโมง (11 ไมล์ต่อชั่วโมง) เท่านั้น[ 169 ]โฮลทซ์ตั้งข้อสังเกตว่าไทแรนโนซอริเดและกลุ่มที่ใกล้เคียงกันบางกลุ่มมี ส่วนประกอบของขาหลัง ส่วนปลาย (หน้าแข้ง บวกเท้า บวกนิ้วเท้า) ที่ยาวกว่าเมื่อเทียบกับความยาวของกระดูกต้นขามากกว่าเทโรพอดอื่นๆ ส่วนใหญ่ และไทแรนโนซอริเดและญาติใกล้เคียงมีกระดูกฝ่าเท้าที่ เชื่อมต่อกันอย่างแน่นหนา [ 171 ]กระดูกฝ่าเท้าชิ้นที่สามถูกบีบอยู่ระหว่างกระดูกฝ่าเท้าชิ้นที่สองและชิ้นที่สี่เพื่อสร้างเป็นหน่วยเดียวที่เรียกว่าอาร์คโตเมตาตาร์ซัสคุณลักษณะของข้อเท้านี้อาจช่วยให้สัตว์วิ่งได้อย่างมีประสิทธิภาพมากขึ้น[ 172 ]คุณลักษณะของขาเหล่านี้ร่วมกันทำให้ไทแรนโนซอรัสสามารถส่งแรงเคลื่อนที่จากเท้าไปยังขาด้านล่างได้อย่างมีประสิทธิภาพมากกว่าในเทโรพอดรุ่นก่อนๆ[ 171 ]

นอกจากนี้ การศึกษาในปี 2020 ชี้ให้เห็นว่าไทแรนโนซอรัสและไดโนเสาร์ในวงศ์ไทแรนโนซอรัสอื่นๆ เดินได้อย่างมีประสิทธิภาพเป็นพิเศษ การศึกษาโดย Dececchi และคณะ ได้เปรียบเทียบสัดส่วนขา มวลร่างกาย และลักษณะการเดินของไดโนเสาร์เทอโรพอดมากกว่า 70 สายพันธุ์ รวมถึงไทแรนโนซอรัสและญาติของมัน จากนั้นทีมวิจัยได้ใช้วิธีการต่างๆ เพื่อประเมินความเร็วสูงสุดของไดโนเสาร์แต่ละตัวขณะวิ่ง รวมถึงปริมาณพลังงานที่ไดโนเสาร์แต่ละตัวใช้ไปขณะเคลื่อนที่ด้วยความเร็วที่ผ่อนคลายกว่า เช่น การเดิน ในบรรดาสายพันธุ์ขนาดเล็กถึงขนาดกลาง เช่น ไดโนเสาร์ในวงศ์ดรอมิโอซอรัส ขาที่ยาวกว่าดูเหมือนจะเป็นการปรับตัวเพื่อการวิ่งที่เร็วขึ้น ซึ่งสอดคล้องกับผลการวิจัยก่อนหน้านี้ของนักวิจัยคนอื่นๆ แต่สำหรับเทอโรพอดที่มีน้ำหนักมากกว่า 1,000 กิโลกรัม (2,200 ปอนด์) ความเร็วในการวิ่งสูงสุดถูกจำกัดด้วยขนาดร่างกาย ดังนั้นจึงพบว่าขาที่ยาวกว่ามีความสัมพันธ์กับการเดินที่ใช้พลังงานต่ำแทน ผลการวิจัยยังชี้ให้เห็นว่า ไดโนเสาร์เทอโรพอดขนาดเล็กมีวิวัฒนาการขาที่ยาวเพื่อช่วยในการล่าเหยื่อและหลบหนีจากผู้ล่าขนาดใหญ่ ในขณะที่ไดโนเสาร์เทอโรพอดขนาดใหญ่ที่มีวิวัฒนาการขาที่ยาวนั้นทำไปเพื่อลดต้นทุนด้านพลังงานและเพิ่มประสิทธิภาพในการหาอาหาร เนื่องจากพวกมันไม่ต้องเผชิญกับแรงกดดันจากการถูกล่าในฐานะผู้ล่าสูงสุด เมื่อเปรียบเทียบกับกลุ่มเทอโรพอดดั้งเดิมในงานวิจัย ไดโนเสาร์ตระกูลไทแรนโนซอรัส เช่นไทแรนโนซอรัสเอง แสดงให้เห็นถึงประสิทธิภาพในการหาอาหารที่เพิ่มขึ้นอย่างเห็นได้ชัดเนื่องจากการใช้พลังงานลดลงในระหว่างการล่าหรือการกินซาก ซึ่งส่งผลให้ไทแรนโนซอรัสไม่จำเป็นต้องออกล่าบ่อยและต้องการอาหารน้อยลงในการดำรงชีวิต นอกจากนี้ งานวิจัยนี้ร่วมกับงานวิจัยที่แสดงให้เห็นว่าไทแรนโนซอรัสมีความคล่องแคล่วว่องไวกว่าเทอโรพอดขนาดใหญ่อื่นๆ บ่งชี้ว่าพวกมันปรับตัวได้ดีมากในการซุ่มโจมตีจากระยะไกลแล้วเร่งความเร็วเพื่อเข้าโจมตีและฆ่าเหยื่อ สามารถสังเกตเห็นความคล้ายคลึงกันระหว่างไทแรนโนซอริเดและหมาป่าในปัจจุบันได้ โดยได้รับการสนับสนุนจากหลักฐานที่ว่าไทแรนโนซอริเดอย่างน้อยบางชนิดล่าเหยื่อเป็นกลุ่ม[ 173 ] [ 174 ]
งานวิจัยที่ตีพิมพ์ในปี 2021 โดย Pasha van Bijlert และคณะ ได้คำนวณความเร็วในการเดินที่เหมาะสมของไทแรนโนซอรัสโดยรายงานความเร็วที่ 1.28 เมตรต่อวินาที (4.6 กิโลเมตรต่อชั่วโมง; 2.9 ไมล์ต่อชั่วโมง) ในขณะเดิน สัตว์จะลดการใช้พลังงานโดยการเลือกจังหวะการก้าวเดินที่เหมาะสม ซึ่งทำให้ส่วนต่างๆ ของร่างกายสั่นสะเทือนไดโนเสาร์ก็เช่นกันแต่การศึกษาในอดีตไม่ได้คำนึงถึงผลกระทบของหางที่มีต่อความเร็วในการเดินอย่างเต็มที่ ตามที่ผู้เขียนกล่าวไว้ เมื่อไดโนเสาร์เดิน หางของมันจะแกว่งขึ้นลงเล็กน้อยในแต่ละก้าว อันเป็นผลมาจากเอ็นยึดระหว่างกระดูกสันหลังที่ค้ำยันหาง เช่นเดียวกับยางยืด เอ็นเหล่านี้จะเก็บพลังงานไว้เมื่อถูกยืดออกเนื่องจากการแกว่งของหาง โดยใช้แบบจำลอง 3 มิติของไทแรนโนซอรัสตัวอย่างTrixได้มีการสร้างกล้ามเนื้อและเอ็นขึ้นใหม่เพื่อจำลองการเคลื่อนไหวของหาง ส่งผลให้ไทแรนโนซอรัส มีจังหวะการเดินที่ประหยัดพลังงาน คล้ายกับที่พบในสัตว์มีชีวิต เช่น มนุษย์ นกกระจอกเทศ และยีราฟ[ 175 ]
การศึกษาในปี 2017 ประเมินความเร็วสูงสุดในการวิ่งของไทแรนโนซอรัสไว้ที่ 17 ไมล์ต่อชั่วโมง (27 กิโลเมตรต่อชั่วโมง) โดยคาดการณ์ว่าไทแรนโนซอรัสใช้พลังงานสำรองหมดไปนานก่อนที่จะถึงความเร็วสูงสุด ส่งผลให้มีความสัมพันธ์แบบพาราโบลาระหว่างขนาดและความเร็ว[ 176 ] [ 177 ]การศึกษาอีกฉบับในปี 2017 ตั้งสมมติฐานว่าไทแรนโนซอรัส ที่โตเต็มวัย ไม่สามารถวิ่งได้เนื่องจากน้ำหนักโครงกระดูกสูง โดยใช้การประมาณน้ำหนักที่คำนวณได้ 7 ตัน แบบจำลองแสดงให้เห็นว่าความเร็วที่สูงกว่า 11 ไมล์ต่อชั่วโมง (18 กิโลเมตรต่อชั่วโมง) อาจทำให้กระดูกขาของไทแรนโนซอรัส แตกหัก ได้ การค้นพบนี้อาจหมายความว่าการวิ่งเป็นไปไม่ได้สำหรับไดโนเสาร์เทอโรพอดขนาดยักษ์อื่นๆ เช่นไจแกนโนโตซอรัส มาปูซอรัสและ อะโครแคน โทซอรัส[ 178 ] อย่างไรก็ตาม การศึกษาของ Eric Snively และเพื่อนร่วมงานที่ตีพิมพ์ในปี 2019 ระบุว่าTyrannosaurusและไดโนเสาร์วงศ์ Tyrannosauridae อื่นๆ มีความคล่องตัวมากกว่าไดโนเสาร์วงศ์ Allosauroididae และไดโนเสาร์วงศ์ Theropodidae อื่นๆ ที่มีขนาดใกล้เคียงกัน เนื่องจากมีแรงเฉื่อยในการหมุนต่ำเมื่อเทียบกับมวลร่างกาย ประกอบกับกล้ามเนื้อขาขนาดใหญ่ ส่งผลให้มีการตั้งสมมติฐานว่าTyrannosaurusสามารถเลี้ยวได้อย่างรวดเร็ว และน่าจะสามารถหมุนตัวได้เร็วขึ้นเมื่ออยู่ใกล้เหยื่อ หรือในขณะที่เลี้ยว ไดโนเสาร์วงศ์ Theropodidae อาจ "หมุนตัว" บนเท้าข้างเดียวที่วางอยู่บนพื้น ในขณะที่ขาอีกข้างหนึ่งถูกยกขึ้นแกว่งไปมาในระหว่างการไล่ล่า ผลการศึกษานี้อาจช่วยให้เข้าใจว่าความคล่องแคล่วว่องไวมีส่วนช่วยให้วิวัฒนาการของไดโนเสาร์วงศ์ Tyrannosauridae ประสบความสำเร็จได้อย่างไร[ 179 ]ในปี 2026 Boeye และเพื่อนร่วมงานได้วิเคราะห์กลไกทางชีวภาพของเท้าของT. rexและพบว่าเท้าของมันน่าจะทำงานคล้ายกับนกในปัจจุบัน สิ่งนี้อาจส่งผลให้ระยะก้าวสั้นลง ความถี่ก้าวสูงขึ้น และความเร็วสูงสุดเพิ่มขึ้นเล็กน้อย[ 18 ]
รอยเท้าที่เป็นไปได้

รอยเท้าฟอสซิลที่แยกจากกันสองรอยได้รับการระบุเบื้องต้นว่าเป็นของทีเร็กซ์ รอยแรกถูกค้นพบที่ฟิลมอนต์ สเกาต์ แรนช์ รัฐนิวเม็กซิโก ในปี 1983 โดยนักธรณีวิทยาชาวอเมริกัน ชาร์ลส์ พิลล์มอร์ เดิมทีคิดว่าเป็นของ ไดโนเสาร์ฮา โดรซอริเดียการตรวจสอบรอยเท้าเผยให้เห็น "ส้นเท้า" ขนาดใหญ่ที่ไม่เคยพบใน รอยเท้า ไดโนเสาร์ออร์นิโทพอดและร่องรอยของสิ่งที่อาจเป็นนิ้วหัวแม่เท้า ซึ่งเป็นนิ้วที่สี่คล้ายเล็บเท้าของไทแรนโนซอร์ รอยเท้าดังกล่าวได้รับการตีพิมพ์เป็นสกุลรอยเท้าTyrannosauripus pillmoreiในปี 1994 โดยมาร์ติน ล็อกลีย์และเอเดรียน ฮันต์ ล็อกลีย์และฮันต์แนะนำว่ามีความเป็นไปได้สูงที่รอยเท้าดังกล่าวเป็นของทีเร็กซ์ซึ่งจะทำให้เป็นรอยเท้าแรกที่รู้จักของสายพันธุ์นี้ รอยเท้าดังกล่าวเกิดขึ้นในพื้นที่ชุ่มน้ำที่เป็นโคลนซึ่งเคยมีพืชปกคลุม มีความยาว 83 เซนติเมตร (33 นิ้ว) และกว้าง 71 เซนติเมตร (28 นิ้ว) [ 180 ]
รอยเท้าที่สองที่อาจเป็นของไทแรนโนซอรัสได้รับการรายงานครั้งแรกในปี 2007 โดยฟิล แมนนิง นักบรรพชีวินวิทยาชาวอังกฤษ จากแหล่งหินเฮลล์ครีก ในรัฐมอนแทนา รอยเท้าที่สองนี้มีความยาว 72 เซนติเมตร (28 นิ้ว) สั้นกว่ารอยเท้าที่ล็อคลีย์และฮันท์อธิบายไว้ ยังไม่ชัดเจนว่ารอยเท้านี้เป็นของไทแรนโนซอรัส หรือไม่ แม้ว่า ไทแรนโนซอรัสจะเป็นเทโรพอดขนาดใหญ่เพียงชนิดเดียวที่ทราบว่าเคยอาศัยอยู่ในแหล่งหินเฮลล์ครีก[ 181 ] [ 182 ]
รอยเท้าชุดหนึ่งในเกลนร็อก รัฐไวโอมิง ซึ่งมีอายุย้อนไปถึง ยุค มาสทริชเชียนของยุคครีเทเชียสตอนปลาย และมาจากชั้นหินแลนซ์ได้รับการอธิบายโดยสก็อตต์ เพอร์สันส์ ฟิล เคอร์รี และเพื่อนร่วมงานในปี 2016 และเชื่อว่าเป็นของทีเร็กซ์ วัยเยาว์ หรือนาโนไทแรนนัส แลนเซนซิส จากการวัดและพิจารณาจากตำแหน่งของรอยเท้า เชื่อว่าสัตว์ตัวนี้กำลังเดินทางด้วยความเร็วในการเดินประมาณ 2.8 ถึง 5 ไมล์ต่อชั่วโมง และคาดว่ามีความสูงที่สะโพก 1.56 ถึง 2.06 เมตร (5.1 ถึง 6.8 ฟุต) [ 183 ] [ 184 ] [ 185 ]บทความติดตามผลปรากฏขึ้นในปี 2017 โดยเพิ่มการประมาณความเร็วขึ้น 50–80% [ 186 ]
รอยเท้าและร่องรอยฟอสซิลหายากที่พบในนิวเม็กซิโกและไวโอมิง ซึ่งจัดอยู่ในสกุลรอยเท้าTyrannosauripusได้รับการสันนิษฐานว่าเป็นรอยเท้าของTyrannosaurusโดยพิจารณาจากอายุทางธรณีวิทยาของหินที่พบ ตัวอย่างแรกที่พบในปี 1994 ได้รับการอธิบายโดย Lockley และ Hunt และประกอบด้วยรอยเท้าขนาดใหญ่เพียงรอยเดียว ฟอสซิลรอยเท้าอีกคู่หนึ่งที่ได้รับการอธิบายในปี 2021 แสดงให้เห็นไดโนเสาร์วงศ์ Tyrannosauridae ขนาดใหญ่กำลังลุกขึ้นจากท่านอนคว่ำโดยใช้ข้อศอกร่วมกับแผ่นรองฝ่าเท้าเพื่อยืน ฟอสซิลสองชุดที่ไม่เหมือนใครนี้พบใน Ludlow รัฐโคโลราโด และ Cimarron รัฐนิวเม็กซิโก[ 187 ]ฟอสซิลรอยเท้าอีกชุดหนึ่งที่ได้รับการอธิบายในปี 2018 ซึ่งอาจเป็นของTyrannosaurusหรือNanotyrannus วัยเยาว์ ถูกค้นพบในชั้นหิน Lance Formation ของไวโอมิง ร่องรอยดังกล่าวเป็นหลักฐานที่หาได้ยากเกี่ยวกับความเร็วในการเดินของไดโนเสาร์วงศ์ไทแรนโนซอรัส และคาดว่าไดโนเสาร์ที่ทิ้งร่องรอยนั้นเคลื่อนที่ด้วยความเร็ว 4.5–8.0 กิโลเมตรต่อชั่วโมง (2.8–5.0 ไมล์ต่อชั่วโมง) ซึ่งเร็วกว่าที่เคยสันนิษฐานไว้ก่อนหน้านี้สำหรับการประเมินความเร็วในการเดินของไดโนเสาร์วงศ์ไทแรนโนซอรัส[ 188 ]
สมองและประสาทสัมผัส

การศึกษาที่ดำเนินการโดยLawrence Witmerและ Ryan Ridgely จากมหาวิทยาลัยโอไฮโอพบว่าTyrannosaurusมีความสามารถทางประสาทสัมผัสที่สูงขึ้นเช่นเดียวกับcoelurosaurs อื่นๆ โดยเน้นการเคลื่อนไหวของดวงตาและศีรษะที่ค่อนข้างรวดเร็วและประสานกัน ความสามารถในการรับรู้เสียงความถี่ต่ำที่เพิ่มขึ้น ซึ่งจะช่วยให้ tyrannosaurus สามารถติดตามการเคลื่อนไหวของเหยื่อจากระยะไกล และประสาทสัมผัสในการดมกลิ่นที่เพิ่มขึ้น[ 189 ]การศึกษาที่ตีพิมพ์โดย Kent Stevens สรุปว่าTyrannosaurusมีสายตาที่เฉียบคม โดยการใช้การวัดขอบเขตการ มองเห็นที่ดัดแปลง กับการสร้างใบหน้าของไดโนเสาร์หลายชนิดรวมถึงTyrannosaurusการศึกษาพบว่าTyrannosaurusมีขอบเขตการมองเห็นแบบสองตาที่ 55 องศา ซึ่งเหนือกว่าเหยี่ยวในปัจจุบัน Stevens ประมาณการว่าTyrannosaurusมีความคมชัดในการมองเห็นมากกว่ามนุษย์ถึง 13 เท่า และเหนือกว่าความคมชัดในการมองเห็นของนกอินทรี ซึ่งมีความคมชัดมากกว่าคนถึง 3.6 เท่า สตีเวนส์ประเมินจุดไกลสุดที่จำกัด (นั่นคือ ระยะทางที่สามารถมองเห็นวัตถุแยกจากเส้นขอบฟ้าได้) ไว้ที่ 6 กิโลเมตร (3.7 ไมล์) ซึ่งไกลกว่า 1.6 กิโลเมตร (1 ไมล์) ที่มนุษย์สามารถมองเห็นได้[ 44 ] [ 45 ] [ 190 ]
Thomas Holtz Jr. ตั้งข้อสังเกตว่าการรับรู้เชิงลึกที่สูงของTyrannosaurusอาจเป็นผลมาจากเหยื่อที่มันต้องล่า โดยสังเกตว่ามันต้องล่าเซราทอปเซียน เช่นไทรเซราทอปส์ แอ นคิโลซอรัส เช่นแอนคิโลซอรัสและฮาโดรซอรัส เขาเสนอว่าสิ่งนี้ทำให้ความแม่นยำมีความสำคัญมากขึ้นสำหรับTyrannosaurusทำให้มันสามารถ "เข้าไป โจมตี และล้มมันลง" ในทางตรงกันข้ามAcrocanthosaurus มีการรับรู้เชิงลึกที่จำกัดเนื่องจากพวกมันล่าซอโรพอด ขนาดใหญ่ ซึ่งค่อนข้างหายากในช่วงเวลาของTyrannosaurus [ 121 ]
แม้ว่าจะไม่พบวงแหวนสเคลราของไทแรนโนซอรัส แต่ เคนเนธ คาร์เพนเตอร์ได้ประมาณขนาดของมันโดยอิงจากของกอร์โกซอรัสวงแหวนสเคลราที่คาดการณ์ไว้สำหรับตัวอย่างสแตนมีเส้นผ่านศูนย์กลางประมาณ 7 ซม. (2.8 นิ้ว) โดยมีเส้นผ่านศูนย์กลางช่องเปิดภายในประมาณ 3.5 ซม. (1.4 นิ้ว) จากสัดส่วนของดวงตาในสัตว์เลื้อยคลานที่ยังมีชีวิตอยู่ สิ่งนี้บ่งชี้ว่าเส้นผ่านศูนย์กลางรูม่านตาประมาณ 2.5 ซม. (0.98 นิ้ว) เส้นผ่านศูนย์กลางม่านตาประมาณเท่ากับวงแหวนสเคลรา และเส้นผ่านศูนย์กลางลูกตา 11–12 ซม. (4.3–4.7 นิ้ว) คาร์เพนเตอร์ยังประมาณความลึกของลูกตาไว้ที่ประมาณ 7.7–9.6 ซม. (3.0–3.8 นิ้ว) จากการคำนวณเหล่านี้ค่า f-numberสำหรับดวงตาของสแตนคือ 3–3.8 เนื่องจาก สัตว์ ที่หากิน ในเวลากลางวัน มีค่า f-number 2.1 หรือสูงกว่า สิ่งนี้จะบ่งชี้ว่าไทแรนโนซอรัสมีการมองเห็นในที่แสงน้อยไม่ดีและล่าเหยื่อในเวลากลางวัน[ 191 ]
ไทแรนโนซอรัสมีปุ่มรับกลิ่นและเส้นประสาทรับกลิ่น ขนาดใหญ่มาก เมื่อเทียบกับขนาดสมอง ซึ่งเป็นอวัยวะที่รับผิดชอบต่อประสาทสัมผัสในการดมกลิ่นที่เฉียบคม สิ่งนี้บ่งชี้ว่าประสาทสัมผัสในการดมกลิ่นได้รับการพัฒนาอย่างมาก และแสดงให้เห็นว่าไทแรนโนซอรัสสามารถตรวจจับซากสัตว์ได้ด้วยกลิ่นเพียงอย่างเดียวในระยะไกล ประสาทสัมผัสในการดมกลิ่นของไทแรนโนซอรัสอาจเทียบได้กับแร้ง ในปัจจุบัน ซึ่งใช้กลิ่นในการติดตามซากสัตว์เพื่อกินซาก การวิจัยเกี่ยวกับปุ่มรับกลิ่นแสดงให้เห็นว่าทีเร็กซ์มีประสาทสัมผัสในการดมกลิ่นที่พัฒนามากที่สุดในบรรดาไดโนเสาร์ที่ไม่ใช่นก 21 ชนิดที่นำมาศึกษา[ 192 ]

T. rexมีหูชั้นในที่ ยาวมาก ซึ่งค่อนข้างผิดปกติในกลุ่มเทอโรพอดความยาวของหูชั้นในมักเกี่ยวข้องกับความสามารถในการได้ยิน หรืออย่างน้อยก็ความสำคัญของการได้ยินในพฤติกรรม ซึ่งบ่งชี้ว่าการได้ยินเป็นประสาทสัมผัสที่สำคัญเป็นพิเศษสำหรับไทแรนโนซอรัส โดยเฉพาะอย่างยิ่ง ข้อมูลชี้ให้เห็นว่าT. rexได้ยินดีที่สุดในช่วงความถี่ต่ำ และเสียงความถี่ต่ำเป็นส่วนสำคัญของพฤติกรรมของไทแรนโนซอรัส[ 189 ]การศึกษาในปี 2017 โดย Thomas Carr และเพื่อนร่วมงานพบว่าจมูกของไทแรนโนซอรัสมีความไวสูง โดยพิจารณาจากจำนวนช่องเปิดเล็กๆ จำนวนมากในกระดูกใบหน้าของDaspletosaurus ที่เกี่ยวข้อง ซึ่งมีเซลล์ประสาทรับความรู้สึกการศึกษาดังกล่าวคาดการณ์ว่าไทแรนโนซอรัสอาจใช้จมูกที่ไวต่อความรู้สึกเพื่อวัดอุณหภูมิของรังและเพื่อหยิบไข่และลูกอ่อนอย่างเบามือ ดังเช่นที่พบในจระเข้ในปัจจุบัน[ 53 ] การศึกษาอีกชิ้นหนึ่งที่ตีพิมพ์ในปี 2021 ชี้ให้เห็นเพิ่มเติมว่าไทแรนโนซอรัสมีประสาทสัมผัสที่ไวมาก โดยอาศัยท่อประสาทและหลอดเลือดที่ด้านหน้าของขากรรไกร ซึ่งมันสามารถใช้เพื่อตรวจจับและกินเหยื่อได้ดียิ่งขึ้น การศึกษาที่ตีพิมพ์โดย Kawabe และ Hittori และคณะ ชี้ให้เห็นว่าไทแรนโนซอรัสยังสามารถรับรู้ความแตกต่างเล็กน้อยของวัสดุและการเคลื่อนไหวได้อย่างแม่นยำ ทำให้มันสามารถใช้กลยุทธ์การกินที่แตกต่างกันในส่วนต่างๆ ของซากเหยื่อได้ ขึ้นอยู่กับสถานการณ์ ท่อประสาทและหลอดเลือดที่ไวต่อความรู้สึกในไทแรนโนซอรัสยังอาจปรับตัวให้เข้ากับการเคลื่อนไหวและพฤติกรรมที่ละเอียดอ่อน เช่น การสร้างรัง การดูแลลูก และพฤติกรรมทางสังคมอื่นๆ เช่น การสื่อสารภายในสายพันธุ์เดียวกัน ผลการศึกษาชิ้นนี้ยังสอดคล้องกับผลการศึกษาในไทแรนโนซอริเดที่เกี่ยวข้องอย่างDaspletosaurus horneriและอัลโลซอรอยด์Neovenatorซึ่งมีการปรับตัวของระบบประสาทและหลอดเลือดที่คล้ายคลึงกัน แสดงให้เห็นว่าใบหน้าของเทโรพอดมีความไวต่อแรงกดและการสัมผัสสูง[ 193 ] [ 194 ]อย่างไรก็ตาม การศึกษาล่าสุดที่ทบทวนวิวัฒนาการของท่อไตรเจมินัลในกลุ่มซอรอปซิดส์ระบุว่า เครือข่ายท่อประสาทและหลอดเลือดที่หนาแน่นกว่ามากในจมูกและขากรรไกรล่างนั้นพบได้บ่อยในกลุ่มที่อาศัยอยู่ในน้ำหรือกึ่งน้ำ (เช่นสไปโนซอรัส , ฮาลส์คารัปเตอร์ , เพลซิโอ ซอรัส ) และกลุ่มที่พัฒนาแรมโฟเทกา (เช่น ซีนานา ธาเซีย ) ในขณะที่เครือข่ายท่อในไทแรนโนซอรัสดูเรียบง่ายกว่า แต่ก็ยังมีการพัฒนามากกว่าในกลุ่มออร์นิธิสเชียนส่วนใหญ่ และกลุ่มที่อาศัยอยู่บนบกโดยรวม เช่น ไทแรนโนซอริเดและนีโอเวเนเตอร์อาจมีความไวต่อการสัมผัสใบหน้าโดยเฉลี่ยสำหรับเทอโรพอดบนบกที่ไม่มีฟัน แม้ว่าจะต้องมีการวิจัยเพิ่มเติมก็ตาม ท่อประสาทและหลอดเลือดในไทแรนโนซอรัสอาจรองรับโครงสร้างเนื้อเยื่ออ่อนเพื่อการควบคุมอุณหภูมิหรือการส่งสัญญาณทางสังคม ซึ่งข้อหลังนี้สามารถยืนยันได้จากข้อเท็จจริงที่ว่าเครือข่ายท่อประสาทและหลอดเลือดอาจมีการเปลี่ยนแปลงในระหว่างการเจริญเติบโต[ 195 ]
งานวิจัยโดย Grant R. Hurlburt, Ryan C. Ridgely และ Lawrence Witmer ได้ทำการประมาณค่า อัตราส่วนการพัฒนาสมอง ( Encephalization Quotientsหรือ EQs) โดยอ้างอิงจากสัตว์เลื้อยคลานและนก รวมถึงประมาณค่าอัตราส่วนของมวลสมองส่วนซีรีบรัมต่อมวลสมองทั้งหมด งานวิจัยสรุปว่าไทแรนโนซอรัสมีสมองที่ใหญ่ที่สุดเมื่อเทียบกับไดโนเสาร์ที่ไม่ใช่นกที่โตเต็มวัยทั้งหมด ยกเว้นไดโนเสาร์ในกลุ่มมานิแรปทอริฟอร์มขนาดเล็กบางชนิด (เช่นแบมบิแรปเตอร์ , โทรดอนและออร์นิโทมิมัส ) งานวิจัยพบว่า ขนาดสมองสัมพัทธ์ของ ไทแรนโนซอรัสยังคงอยู่ในช่วงของสัตว์เลื้อยคลานในปัจจุบัน โดยสูงกว่าค่าเฉลี่ยของ EQs ของสัตว์เลื้อยคลานที่ไม่ใช่นกไม่เกิน 2 ส่วนเบี่ยงเบนมาตรฐานค่าประมาณอัตราส่วนของมวลสมองส่วนซีรีบรัมต่อมวลสมองทั้งหมดจะอยู่ในช่วง 47.5 ถึง 49.53 เปอร์เซ็นต์ จากการศึกษาพบว่า ค่านี้สูงกว่าค่าประมาณต่ำสุดของนกที่ยังมีชีวิตอยู่ (44.6 เปอร์เซ็นต์) แต่ยังคงใกล้เคียงกับอัตราส่วนทั่วไปของจระเข้ที่โตเต็มวัยทางเพศที่เล็กที่สุด ซึ่งมีช่วงตั้งแต่ 45.9–47.9 เปอร์เซ็นต์[ 196 ]การศึกษาอื่นๆ เช่น การศึกษาของ Steve Brusatte ระบุว่าค่าสัมประสิทธิ์การพัฒนาสมองของTyrannosaurusมีช่วงใกล้เคียงกัน (2.0–2.4) กับลิงชิมแปนซี (2.2–2.5) แม้ว่าเรื่องนี้อาจเป็นที่ถกเถียงกันได้ เนื่องจากค่าสัมประสิทธิ์การพัฒนาสมองของสัตว์เลื้อยคลานและสัตว์เลี้ยงลูกด้วยนมไม่เท่ากัน[ 197 ]
พฤติกรรมทางสังคม

ฟิลิป เจ. เคอร์รีเสนอว่าไทแรนโนซอรัสอาจเป็นนักล่าเป็นฝูงโดยเปรียบเทียบทีเร็กซ์กับสายพันธุ์ที่เกี่ยวข้องอย่างทาร์โบซอรัส บาตาร์และอัลเบอร์โตซอรัส ซาร์โคฟากั ส โดยอ้างหลักฐานฟอสซิลที่อาจบ่งชี้ถึง พฤติกรรม รวมกลุ่ม (ซึ่งหมายถึงสัตว์ที่เดินทางเป็นฝูง) [ 198 ]การค้นพบ โครงกระดูก ทีเร็กซ์สามโครงที่อยู่ใกล้กัน ใน เซาท์ดาโคตา อาจบ่งชี้ถึงการรวมกลุ่มเป็นฝูง [ 199 ] [ 200 ]การล่าเป็นฝูงแบบร่วมมือกันอาจเป็นกลยุทธ์ที่มีประสิทธิภาพในการปราบเหยื่อที่มีการปรับตัวต่อต้านผู้ล่า ขั้นสูง ซึ่งอาจเป็นอันตรายถึงชีวิตได้ เช่นไทรเซราทอปส์และแอนคิโลซอรัส[ 198 ]
สมมติฐานการ ล่าเป็นฝูงของทีเร็กซ์ ของเคอร์รี ถูกวิพากษ์วิจารณ์ว่าไม่ได้ผ่านการตรวจสอบโดยผู้เชี่ยวชาญแต่กลับถูกกล่าวถึงในการสัมภาษณ์ทางโทรทัศน์และหนังสือชื่อDino Gangs [ 201 ] ทฤษฎีการล่าเป็นฝูงของทีเร็กซ์ของเคอร์รีนั้นอิงตามการเปรียบเทียบกับสายพันธุ์อื่นเป็นหลัก คือTarbosaurus bataarหลักฐานเกี่ยวกับการรวมกลุ่มกันของT. bataarเองนั้นยังไม่ได้รับการตรวจสอบโดยผู้เชี่ยวชาญ และเคอร์รีเองก็ยอมรับว่าสามารถตีความได้โดยอ้างอิงจากหลักฐานในสายพันธุ์อื่นที่ใกล้เคียงกันเท่านั้น ตามที่เคอร์รีกล่าว การรวมกลุ่มกันของAlbertosaurus sarcophagusได้รับการสนับสนุนจากการค้นพบซากดึกดำบรรพ์ 26 ตัวที่มีอายุแตกต่างกันในแหล่งซากดึกดำบรรพ์ Dry Island เขาตัดความเป็นไปได้ของกับดักผู้ล่าออกไปเนื่องจากสภาพการเก็บรักษาของซากแต่ละตัวคล้ายคลึงกันและแทบไม่มีสัตว์กินพืชเลย[ 201 ] [ 202 ]
หลักฐานเพิ่มเติมที่สนับสนุนพฤติกรรมการรวมกลุ่มของไดโนเสาร์วงศ์ Tyrannosauridae สามารถพบได้จากร่องรอย ฟอสซิล จากชั้นหินWapiti Formation ยุคครีเทเชียสตอนบน ทางตะวันออกเฉียงเหนือของบริติชโคลัมเบียประเทศแคนาดา ซึ่งเกิดจากไดโนเสาร์วงศ์ Tyrannosauridae สามตัวที่เดินทางไปในทิศทางเดียวกัน[ 203 ] [ 204 ]ตามที่นักวิทยาศาสตร์ที่ประเมินโครงการ Dino Gangs ระบุ หลักฐานการล่าเป็นฝูงในTarbosaurusและAlbertosaurusนั้นอ่อนแอและอิงจากซากโครงกระดูกที่เป็นกลุ่ม ซึ่งอาจมีคำอธิบายอื่นที่ใช้ได้ (เช่น ภัยแล้งหรือน้ำท่วมทำให้ไดโนเสาร์ต้องตายพร้อมกันในที่เดียว) [ 201 ]นักวิจัยคนอื่นๆ ได้ตั้งข้อสันนิษฐานว่า แทนที่จะเป็นกลุ่มสังคมขนาดใหญ่ของเทโรพอด การค้นพบเหล่านี้บางส่วนแสดงถึงพฤติกรรมที่คล้ายกับ การรุมกินซากศพแบบ มังกรโคโมโดถึงขั้นกล่าวว่าพฤติกรรมการล่าเป็นฝูงที่แท้จริงอาจไม่มีอยู่ในไดโนเสาร์ที่ไม่ใช่นก เนื่องจากเป็นพฤติกรรมที่หายากในสัตว์นักล่าในปัจจุบัน[ 205 ]พยาธิสภาพของ ตัวอย่าง ไทแรนโนซอรัสได้รับการเสนอแนะว่าเป็นหลักฐานของการโจมตีโดยสายพันธุ์เดียวกัน รวมถึง "ไวเร็กซ์" ที่มีรูทะลุผ่านกระดูกหางและบาดแผลรุนแรงที่หางซึ่งแสดงให้เห็นสัญญาณของการปรับโครงสร้างกระดูก (ไม่ใช่การงอกใหม่) [ 206 ] [ 207 ]
อาหารและพฤติกรรมการกินอาหาร


นักบรรพชีวินวิทยาส่วนใหญ่ยอมรับว่าไทแรนโนซอรัสเป็นนักล่า ที่กระตือรือร้น และเช่นเดียวกับสัตว์กินเนื้อ ขนาดใหญ่ส่วนใหญ่ มันจะฉวย โอกาสกินซาก สัตว์ ด้วย [ 208 ]ทีเร็กซ์เป็น สัตว์ กินเนื้อที่ใหญ่ที่สุดในสภาพแวดล้อมของมัน และ น่าจะ เป็นนักล่าสูงสุดโดยล่าฮาโดรซอร์สัตว์กินพืชที่มีเกราะ เช่นเซราทอปเซียนและแอนคิโลซอร์และอาจรวมถึงซอโรพอด (เช่นอลาโมซอรัส ) ด้วย [ 209 ] ค่า δ 44/42 Ca ของเคลือบฟันยังชี้ให้เห็นถึงความเป็นไปได้ที่ทีเร็กซ์จะกินซากสัตว์เลื้อยคลานและปลาทะเลที่ถูกคลื่นซัดขึ้นฝั่งในทะเลสาบภายในตะวันตกเป็นครั้งคราว[ 210 ]การศึกษาในปี 2012 โดย Karl Bates และ Peter Falkingham พบว่าTyrannosaurusมีแรงกัดที่ทรงพลังที่สุดในบรรดาสัตว์บกที่เคยมีชีวิตอยู่ โดยพบว่าTyrannosaurus ตัวเต็มวัยสามารถออกแรง กัดที่ฟันหลังได้ถึง 35,000 ถึง 57,000 N (7,868 ถึง 12,814 lbf ) [ 211 ] [ 212 ] [ 213 ] Mason B. Meers ได้ทำการประมาณค่าที่สูงกว่านี้ในปี 2003 [ 47 ]ซึ่งทำให้มันสามารถบดกระดูกได้ในระหว่างการกัดซ้ำๆ และกินซากไดโนเสาร์ขนาดใหญ่ได้อย่างสมบูรณ์[ 26 ] Stephan Lautenschlager และเพื่อนร่วมงานคำนวณว่าTyrannosaurusสามารถอ้าปากได้สูงสุดประมาณ 80 องศา ซึ่งเป็นการปรับตัวที่จำเป็นสำหรับมุมขากรรไกรที่หลากหลายเพื่อเสริมพลังให้กับแรงกัดที่แข็งแกร่งของสิ่งมีชีวิต[ 214 ] [ 215 ]
อย่างไรก็ตาม มีการถกเถียงกันว่าไทแรนโนซอรัสเป็นสัตว์นักล่า เป็นหลัก หรือเป็นสัตว์กิน ซาก โดยแท้จริง การถกเถียงนี้มีต้นกำเนิดมาจากการศึกษาในปี 1917 โดยแลมบ์ ซึ่งโต้แย้งว่าเทโรพอดขนาดใหญ่เป็นสัตว์กินซากโดยแท้จริงเพราะ ฟันของ กอร์โกซอรัสแทบไม่มีร่องรอยการสึกหรอเลย[ 216 ]ข้อโต้แย้งนี้ละเลยข้อเท็จจริงที่ว่าเทโรพอดเปลี่ยนฟันของพวกมันค่อนข้างเร็ว นับตั้งแต่การค้นพบไทแรนโนซอรัส ครั้งแรก นักวิทยาศาสตร์ส่วนใหญ่ต่างคาดเดาว่ามันเป็นสัตว์นักล่า เช่นเดียวกับสัตว์นักล่าขนาดใหญ่ในปัจจุบัน มันจะกินซากหรือขโมยเหยื่อของสัตว์นักล่าตัวอื่นได้อย่างง่ายดายหากมีโอกาส[ 217 ]
นักบรรพชีวินวิทยาJack Hornerเป็นผู้สนับสนุนหลักของมุมมองที่ว่าTyrannosaurusไม่ใช่นักล่าเลย แต่เป็นสัตว์กินซากโดยเฉพาะ[ 144 ] [ 218 ] [ 219 ]เขาได้นำเสนอข้อโต้แย้งในวรรณกรรมที่เป็นที่นิยมเพื่อสนับสนุนสมมติฐานสัตว์กินซากโดยแท้จริง:

- แขนของไทแรนโนซอรัสสั้นเมื่อเทียบกับสัตว์นักล่าชนิดอื่นที่รู้จัก ฮอร์เนอร์แย้งว่าแขนนั้นสั้นเกินไปที่จะสร้างแรงยึดจับที่จำเป็นเพื่อจับเหยื่อไว้ได้[ 220 ]นักบรรพชีวินวิทยาคนอื่นๆ เช่นโทมัส โฮลท์ซ จูเนียร์แย้งว่ามีสัตว์นักล่าในปัจจุบันจำนวนมากที่ไม่ใช้แขนขาหน้าในการล่า เช่นหมาป่าไฮยีน่าและนกเลขารวมถึงสัตว์ที่สูญพันธุ์ไปแล้วอื่นๆ ที่คิดว่าเป็นสัตว์นักล่าที่ไม่ใช้แขนขาหน้า เช่นฟอรัสราซิด[ 221 ] [ 222 ]
- ไทแรนโนซอรัสมีปุ่มรับกลิ่นและเส้นประสาทรับกลิ่น ขนาดใหญ่ (เมื่อเทียบกับขนาดสมอง) ซึ่งบ่งชี้ว่ามีประสาทรับกลิ่นที่พัฒนาอย่างมาก ซึ่งสามารถดมกลิ่นซากสัตว์ได้ในระยะไกล เช่นเดียวกับนกแร้ง ในปัจจุบัน การวิจัยเกี่ยวกับปุ่มรับกลิ่นของไดโนเสาร์แสดงให้เห็นว่าไทแรนโนซอรัสมีประสาทรับกลิ่นที่พัฒนามากที่สุดในบรรดาไดโนเสาร์ 21 ชนิดที่นำมาศึกษา[ 192 ]
- ฟันของไทแรนโนซอรัสสามารถบดกระดูกได้ และด้วยเหตุนี้จึงสามารถสกัดอาหาร ( ไขกระดูก ) ได้มากที่สุดเท่าที่จะเป็นไปได้จากซากสัตว์ ซึ่งโดยปกติแล้วจะเป็นส่วนที่มีคุณค่าทางโภชนาการน้อยที่สุด Karen Chin และเพื่อนร่วมงานได้พบเศษกระดูกในโคโปรไลต์ (อุจจาระที่กลายเป็นฟอสซิล) ซึ่งพวกเขาเชื่อว่าเป็นของไทแรนโนซอรัส แต่ชี้ให้เห็นว่าฟันของไทแรนโนซอรัสไม่ได้ปรับตัวให้เหมาะสมกับการเคี้ยวกระดูกอย่างเป็นระบบเหมือนที่ไฮยีน่าทำเพื่อสกัดไขกระดูก[ 223 ]
- เนื่องจากเหยื่อที่เป็นไปได้ของ ไทแรนโนซอรัสอย่างน้อยบางส่วนสามารถเคลื่อนที่ได้อย่างรวดเร็ว หลักฐานที่ว่ามันเดินแทนที่จะวิ่งอาจบ่งชี้ว่ามันเป็นสัตว์กินซาก[ 218 ]ในทางกลับกัน การวิเคราะห์ล่าสุดชี้ให้เห็นว่าไทแรนโนซอรัสแม้จะช้ากว่านักล่าบนบกขนาดใหญ่ในปัจจุบัน แต่ก็อาจจะเร็วพอที่จะล่าฮาโดรซอร์และเซราทอปเซียนขนาด ใหญ่ได้ [ 169 ] [ 182 ]
หลักฐานอื่นๆ ชี้ให้เห็นถึงพฤติกรรมการล่าเหยื่อในไทแรนโนซอรัสเบ้าตาของไทแรนโนซอรัสอยู่ในตำแหน่งที่ดวงตาจะชี้ไปข้างหน้า ทำให้พวกมันมีวิสัยทัศน์แบบสองตาที่ดีกว่าเหยี่ยว ในปัจจุบันเล็กน้อย ไม่ชัดเจนว่าเหตุใดการคัดเลือกโดยธรรมชาติจึงสนับสนุนแนวโน้มระยะยาวนี้ หากไทแรนโนซอรัสเป็นสัตว์กินซากอย่างเดียว ซึ่งไม่จำเป็นต้องมีการรับรู้เชิงลึกขั้น สูง ที่การมองเห็นแบบสามมิติให้มา[ 44 ] [ 45 ]ในสัตว์สมัยใหม่ การมองเห็นแบบสองตาพบได้ส่วนใหญ่ในสัตว์นักล่า

โครงกระดูกของไดโนเสาร์ฮาโดรซอริเดียEdmontosaurus annectensจากรัฐมอนแทนาได้รับการอธิบายโดยพบร่องรอยความเสียหายที่กระดูกสันหลัง ส่วนหางซึ่งเกิดจากการโจมตีของไทแรนโนซอรัสที่หายดีแล้ว ข้อเท็จจริงที่ว่าความเสียหายดูเหมือนจะหายดีแล้วนั้นบ่งชี้ว่าEdmontosaurusรอดชีวิตจากการโจมตีของไทแรนโนซอรัสต่อเป้าหมายที่มีชีวิต กล่าวคือ ไทแรนโนซอรัสพยายามล่าเหยื่ออย่าง กระตือรือร้น [ 224 ]แม้ว่าจะมีความเห็นพ้องกันว่ารอยกัดที่หางเกิดจากไทแรน โนซอรัส แต่ก็มีหลักฐานบางอย่างที่แสดงให้เห็นว่าอาจเกิดจากปัจจัยอื่น ตัวอย่างเช่น การศึกษาในปี 2014 ชี้ให้เห็นว่าบาดแผลที่หางอาจเกิดจากการที่Edmontosaurusเหยียบกัน[ 225 ]ในขณะที่การศึกษาอีกฉบับในปี 2020 สนับสนุนสมมติฐานที่ว่าความเครียดทางชีวกลศาสตร์เป็นสาเหตุของบาดแผลที่หาง[ 226 ]นอกจากนี้ยังมีหลักฐานของการโต้ตอบที่ก้าวร้าวระหว่างไทรเซราทอปส์และไทแรนโนซอรัสในรูปแบบของรอยฟันของไทแรนโนซอรัสที่หายดีบางส่วนบนเขาคิ้วและกระดูกสควอโมซัล (กระดูกของแผ่นคอ ) ของ ไทร เซราทอ ปส์ เขาที่ถูกกัดก็หักเช่นกัน โดยมีกระดูกงอกใหม่หลังจากหัก ไม่ทราบแน่ชัดว่าลักษณะการโต้ตอบเป็นอย่างไร สัตว์ตัวใดตัวหนึ่งอาจเป็นฝ่ายรุกก็ได้[ 227 ]เนื่องจาก บาดแผล ของไทรเซราทอปส์หายดีแล้ว จึงเป็นไปได้มากที่สุดว่าไทรเซราทอปส์รอดชีวิตจากการเผชิญหน้าและสามารถเอาชนะไทแรนโนซอรัส ได้ ในการต่อสู้กับไทรเซราทอปส์ตัวผู้ไทรเซราทอปส์น่าจะป้องกันตัวเองโดยการสร้างบาดแผลร้ายแรงให้กับไทแรนโนซอรัสโดยใช้เขาแหลมคมของมัน[ 228 ]การศึกษาของซูพบกระดูกน่องและกระดูกสันหลังส่วนหางที่หักและสมานตัว กระดูกใบหน้าที่มีรอยแผลเป็น และฟันของไทแรนโนซอรัส ตัวอื่น ฝังอยู่ในกระดูกสันหลังส่วนคอ ซึ่งเป็นหลักฐานแสดงถึงพฤติกรรมก้าวร้าว[ 229 ]การศึกษาเกี่ยวกับกระดูกสันหลังของฮาโดรซอร์จากแหล่งหินเฮลล์ครีกที่ถูกเจาะด้วยฟันของไทแรนโนซอรัส วัยเยาว์ในระยะสุดท้าย แสดงให้เห็นว่าแม้จะขาดการปรับตัวในการบดกระดูกเหมือนตัวเต็มวัย แต่ตัวอ่อนก็ยังสามารถใช้เทคนิคการกินโดยการเจาะกระดูกแบบเดียวกับตัวเต็มวัยได้[ 230 ]
In 1992, William Abler suggested that Tyrannosaurus may have had infectious saliva used to kill its prey. Abler observed that the serrations (tiny protuberances) on the cutting edges of the teeth are closely spaced, enclosing little chambers, which he argued trapped pieces of carcass with bacteria, giving Tyrannosaurus a deadly, infectious bite, as the Komodo dragon was mistakenly thought to have.[231] Jack Horner and Don Lessem, in a 1993 popular book, questioned Abler's hypothesis, arguing that Tyrannosaurus's tooth serrations as more like cubes in shape than the serrations on a Komodo monitor's teeth, which are rounded.[144]: 214–215 The basis for an infectious bite in Tyrannosaurus or any other theropod was further undermined when it was found Komodo dragons do not actually have an infectious bite nor hunt by biting prey and tracking it as it weakens.[232]
Tyrannosaurus probably primarily processed carcasses with lateral shakes of the head, like crocodilians. The head was not as maneuverable as the skulls of allosauroids, due to flat joints of the neck vertebrae.[233]
Evidence also strongly suggests that tyrannosaurs were at least occasionally cannibalistic. Tyrannosaurus itself has strong evidence pointing towards it having been cannibalistic in at least a scavenging capacity based on tooth marks on the foot bones, humerus, and metatarsals of one specimen.[234]
Parenting
While there is no direct evidence of Tyrannosaurus raising their young (the rarity of juvenile and nest Tyrannosaur fossils has left researchers guessing), it has been suggested that like its closest living relatives, birds and crocodiles (modern archosaurs), Tyrannosaurus may have protected and fed its young. Crocodilians and birds are often suggested by some paleontologists to be modern analogues for dinosaur parenting.[235] Direct evidence of parental behavior exists in other dinosaurs such as Maiasaura peeblesorum, the first dinosaur to have been discovered to raise its young, as well as more closely related Oviraptorids, the latter suggesting parental behavior in theropods.[236][237][238][239][240]
Pathology


{kind=link}
In 2001, Bruce Rothschild and others published a study examining evidence for stress fractures and tendon avulsions in theropod dinosaurs and the implications for their behavior. Since stress fractures are caused by repeated trauma rather than singular events they are more likely to be caused by regular behavior than other types of injuries. Of the 81 Tyrannosaurus foot bones examined in the study, one was found to have a stress fracture, while none of the 10 hand bones were found to have stress fractures. The researchers found tendon avulsions only among Tyrannosaurus and Allosaurus. An avulsion injury left a divot on the humerus of Sue the T. rex, apparently located at the origin of the deltoid or teres major muscles. The presence of avulsion injuries being limited to the forelimb and shoulder in both Tyrannosaurus and Allosaurus suggests that theropods may have had a musculature more complex than and functionally different from those of birds. The researchers concluded that Sue's tendon avulsion was probably obtained from struggling prey. The presence of stress fractures and tendon avulsions, in general, provides evidence for a "very active" predation-based diet rather than obligate scavenging.[241]
A 2009 study showed that smooth-edged holes in the skulls of several specimens might have been caused by Trichomonas-like parasites that commonly infect birds. According to the study, seriously infected individuals, including "Sue" and MOR 980 ("Peck's Rex"), might therefore have died from starvation after feeding became increasingly difficult. Previously, these holes had been explained by the bacterious bone infection Actinomycosis or by intraspecific attacks.[242] A subsequent study found that while trichomoniasis has many attributes of the model proposed (osteolytic, intra oral) several features make the assumption that it was the cause of death less supportable by evidence. For example, the observed sharp margins with little reactive bone shown by the radiographs of Trichomonas-infected birds are dissimilar to the reactive bone seen in the affected T. rex specimens. Also, trichomoniasis can be very rapidly fatal in birds (14 days or less) albeit in its milder form, and this suggests that if a Trichomonas-like protozoan is the culprit, trichomoniasis was less acute in its non-avian dinosaur form during the Late Cretaceous. Finally, the relative size of this type of lesions is much larger in small bird throats, and may not have been enough to choke a T. rex.[243] A more recent study examining the pathologies concluded that the osseous alteration observed most closely resembles those around healing human cranial trepanations and healing fractures in the Triassic reptile Stagonolepis, in the absence of infection. The possible cause may instead have been intraspecific combat.[244]
One study of Tyrannosaurus specimens with tooth marks in the bones attributable to the same genus was presented as evidence of cannibalism.[234] Tooth marks in the humerus, foot bones and metatarsals, may indicate opportunistic scavenging, rather than wounds caused by combat with another T. rex.[234][245] Other tyrannosaurids may also have practiced cannibalism.[234]
Paleoecology

Tyrannosaurus lived during what is referred to as the Lancian faunal stage (Maastrichtian age) at the end of the Late Cretaceous. Tyrannosaurus ranged from Canada in the north to at least New Mexico in the south of Laramidia.[6] Several notable Tyrannosaurus remains have been found in the Hell Creek Formation. During the Maastrichtian this area was maritime to subtropical, with a warm and humid climate.[246] The flora consisted mostly of angiosperms, but also included trees like dawn redwood (Metasequoia).[247] Archosaurs discovered from this formation include the ceratopsiansLeptoceratops, Torosaurus, and Triceratops,[248] the hadrosaurid Edmontosaurus annectens,[249] the parksosauridThescelosaurus,[250] the ankylosaursAnkylosaurus and Denversaurus,[251] the pachycephalosaursPachycephalosaurus, Platytholus and Sphaerotholus,[252][253] the paravian theropods Acheroraptor and Pectinodon,[254] other maniraptoran theropods Anzu, Eoneophron, Trierarchuncus and indeterminate ornithomimids,[255][256] the tyrannosauroidNanotyrannus,[24] and the azhdarchid pterosaur Infernodrakon.[257]

Another formation with Tyrannosaurus remains is the Lance Formation of Wyoming. This has been interpreted as a bayou environment similar to today's Gulf Coast. The fauna was very similar to Hell Creek, but with Struthiomimus replacing its relative Ornithomimus. The small ceratopsian Leptoceratops also lived in the area.[258]
In its southern range, specifically based on remains discovered from the North Horn Formation of Utah, Tyrannosaurus rex lived alongside the titanosaurUtetitan, the ceratopsidTorosaurus and the indeterminate troodontids and hadrosaurids.[259][260][261]Tyrannosaurus mcraeensis from the McRae Group of New Mexico coexisted with the ceratopsidSierraceratops and possibly the titanosaur Alamosaurus.[66] Potential remains identified as cf. Tyrannosaurus have also been discovered from the Javelina Formation of Texas,[66] where the remains of the titanosaur (either Alamosaurus or Utetitan),[261] the ceratopsid Bravoceratops, the pterosaursQuetzalcoatlus and Wellnhopterus, and possible species of troodontids and hadrosaurids are found.[262][263][264] Its southern range is thought to have been dominated by semi-arid inland plains, following the probable retreat of the Western Interior Seaway as global sea levels fell.[265]
Tyrannosaurus may have also inhabited Mexico's Lomas Coloradas Formation in Sonora. Though skeletal evidence is lacking, six shed and broken teeth from the fossil bed have been thoroughly compared with other theropod genera and appear to be identical to those of Tyrannosaurus. If true, the evidence indicates the range of Tyrannosaurus was possibly more extensive than previously believed.[266] It is possible that tyrannosaurs were originally Asian species, migrating to North America before the end of the Cretaceous period.[267]
Population estimates

According to studies published in 2021 by Charles Marshall et al., the total population of adult Tyrannosaurus at any given time was perhaps 20,000 individuals, with computer estimations also suggesting a total population no lower than 1,300 and no higher than 328,000. The authors themselves suggest that the estimate of 20,000 individuals is probably lower than what should be expected, especially when factoring in that disease pandemics could easily wipe out such a small population. Over the span of the genus' existence, it is estimated that there were about 127,000 generations and that this added up to a total of roughly 2.5 billion animals until their extinction.[268][269]
In the same paper, it is suggested that in a population of Tyrannosaurus adults numbering 20,000, the number of individuals living in an area the size of California could be as high as 3,800 animals, while an area the size of Washington D.C. could support a population of only two adult Tyrannosaurus. The study does not take into account the number of juvenile animals in the genus present in this population estimate due to their occupation of a different niche than the adults, and thus it is likely the total population was much higher when accounting for this factor. Simultaneously, studies of living carnivores suggest that some predator populations are higher in density than others of similar weight (such as jaguars and hyenas, which are similar in weight but have vastly differing population densities). Lastly, the study suggests that in most cases, only one in 80 million Tyrannosaurus would become fossilized, while the chances were likely as high as one in every 16,000 of an individual becoming fossilized in areas that had more dense populations.[268][269]
Meiri (2022) questioned the reliability of the estimates, citing uncertainty in metabolic rate, body size, sex and age-specific survival rates, habitat requirements and range size variability as shortcomings Marshall et al. did not take into account.[270] The authors of the original publication replied that while they agree that their reported uncertainties were probably too small, their framework is flexible enough to accommodate uncerainty in physiology, and that their calculations do not depend on short-term changes in population density and geographic range, but rather on their long-term averages. Finally, they remark that they did estimate the range of reasonable survivorship curves and that they did include uncertainty in the time of onset of sexual maturity and in the growth curve by incorporating the uncertainty in the maximum body mass.[271]
Cultural significance

Tyrannosaurus is the most widely known dinosaur in popular culture. Science writer Riley Black states, "In all of prehistory, there is no animal that commands our attention quite like Tyrannosaurus rex, the king of the tyrant lizards. Since the time this dinosaur was officially named in 1905, the enormous carnivore has stood as the ultimate dinosaur."[272] Paleontologist David Hone notes the popularity of Tyrannosaurus models and skeletal displays in museums and writes that films like Jurassic Park and King Kong "would not have been the same without it."[273]T. rex is the only dinosaur that is commonly known to the general public by its full scientific name (binomial name) and the scientific abbreviation T. rex has also come into wide usage.[8]Robert T. Bakker notes this in The Dinosaur Heresies and explains that, "a name like 'T. rex' is just irresistible to the tongue."[37]
See also
Notes
- ^lit.'tyrant lizard'; from Ancient Greekτύραννος (túrannos)'tyrant' and σαῦρος (saûros)'lizard'
Further reading
- Farlow, J. O.; Gatesy, S. M.; Holtz, T. R. Jr.; Hutchinson, J. R.; Robinson, J. M. (2000). "Theropod Locomotion". American Zoologist. 40 (4): 640–663. doi:10.1093/icb/40.4.640. JSTOR 3884284.
External links
 Media from Commons
Media from Commons Quotations from Wikiquote
Quotations from Wikiquote Texts from Wikisource
Texts from Wikisource Textbooks from Wikibooks
Textbooks from Wikibooks Taxa from Wikispecies
Taxa from Wikispecies
- The University of Edinburgh Lecture Dr Stephen Brusatte – Tyrannosaur Discoveries Feb 20, 2015
- 28 species in the tyrannosaur family tree, when and where they livedStephen BrusatteThomas Carr 2016
- Australia's answer to T-Rex, State Library of Queensland
Exhibits
- American Museum of Natural History
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ไทแรนโนซอรัส
ไทแรนโนซอรัส ( / t ɪ ˌ r æ n ə ˈ s ɔː r ə s , t aɪ - / ) [ a ] เป็น สกุล ของ ไดโนเสาร์ เทอ โรพอด ขนาดใหญ่ ไทแรนโนซอรัสเร็ก ซ์ ( เร็กซ์ แปลว่า 'ราชา' ใน ภาษาละติน )...
การค้นพบที่เก่าแก่ที่สุด
ฟันของสิ่งที่ปัจจุบันได้รับการบันทึกว่าเป็น ไทแรนโนซอรัส เร็กซ์ ถูกค้นพบในเดือนกรกฎาคม ค.ศ. 1874 บน ภูเขาเซาท์เทเบิล (โคโลราโด) โดย ปีเตอร์ ที.
การค้นพบและการตั้งชื่อโครงกระดูก
บาร์นัม บราวน์ ผู้ช่วยภัณฑารักษ์ของ พิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติอเมริกัน พบโครงกระดูกบางส่วนของ ทีเร็กซ์ เป็นครั้งแรก ในไวโอมิงตะวันออกในปี 1900 บราวน์พบโครงกระดูกบางส่วนอีกชิ้นหนึ่งใน แหล่งหินเฮลล์ครีก ในมอนแทนาในปี 1902 ซึ่งประกอบด้วยกระดูกฟอสซิลประมาณ...
การศึกษาต่อเนื่อง
ตั้งแต่ช่วงทศวรรษ 1960 เป็นต้นมา มีความสนใจใน ไทแรนโนซอรัส เพิ่มขึ้นอีกครั้ง ส่งผลให้มีการค้นพบโครงกระดูก 42 โครง (สมบูรณ์ 5–80% ตามจำนวนกระดูก) จากทางตะวันตกของทวีปอเมริกาเหนือ [ 6 ] ในปี 1967 ดร.