การสังเคราะห์ทางชีวภาพ
การสังเคราะห์ทางชีวภาพคือการสังเคราะห์ทางเคมีที่เกิดขึ้นในบริบททางชีวภาพ โดยส่วนใหญ่มักหมายถึงกระบวนการหลายขั้นตอนที่เร่งปฏิกิริยาโดยเอนไซม์ซึ่งสารเคมีที่ดูดซึมเป็นสารอาหาร (หรือที่ถูกเปลี่ยนรูปผ่านการสังเคราะห์ทางชีวภาพมาก่อน) ทำหน้าที่เป็นสารตั้งต้น ของเอนไซม์ โดยสิ่งมีชีวิตจะเปลี่ยนสารเหล่านั้นให้เป็นผลิตภัณฑ์ ที่เรียบง่ายหรือซับซ้อนมากขึ้น ตัวอย่างของวิถีการสังเคราะห์ทางชีวภาพ ได้แก่ วิถีการผลิต กรด อะมิโน ส่วนประกอบของ เยื่อหุ้มเซลล์ไขมันและนิวคลีโอไทด์แต่ยังรวมถึงการผลิตโมเลกุลขนาดใหญ่ ทางชีวภาพทุกประเภท และ อะ เซทิลโคเอนไซม์เออะดีโนซีนไตรฟอสเฟตนิโคตินาไมด์อะดีนีนไดนิวคลีโอไทด์และโมเลกุลตัวกลางและโมเลกุลธุรกรรมที่สำคัญอื่นๆ ที่จำเป็นสำหรับการเผาผลาญดังนั้น ในการสังเคราะห์ทางชีวภาพสารประกอบ ต่างๆ ตั้งแต่แบบง่ายไปจนถึงแบบซับซ้อน จะถูกเปลี่ยนเป็นสารประกอบอื่นๆ และจึงรวมถึงทั้งการ สร้าง และการสลาย (การสร้างและการทำลาย) ของโมเลกุลที่ซับซ้อน (รวมถึงโมเลกุลขนาด ใหญ่ ) กระบวนการสังเคราะห์ทางชีวภาพมักแสดงด้วยแผนภูมิวิถีการเผาผลาญ วิถีการสังเคราะห์ทางชีวภาพบางอย่างอาจอยู่ภายในออร์แกเนลล์ เดียวของเซลล์ (เช่น วิถี การสังเคราะห์กรดไขมันในไมโทคอนเดรีย ) ในขณะที่บางวิถีเกี่ยวข้องกับเอนไซม์ที่กระจายอยู่ทั่วออร์แกเนลล์และโครงสร้างต่างๆ ของเซลล์ (เช่น การสังเคราะห์โปรตีนบนพื้นผิวเซลล์ที่มีการเติมหมู่ไกลโคซิล)
องค์ประกอบของการสังเคราะห์ทางชีวภาพ
องค์ประกอบของการสังเคราะห์ทางชีวภาพประกอบด้วยสารตั้งต้นพลังงานเคมี (เช่นATP ) และเอนไซม์เร่งปฏิกิริยา ซึ่งอาจต้องการโคเอนไซม์ (เช่นNADH , NADPH ) องค์ประกอบเหล่านี้สร้างโมโนเมอร์ซึ่งเป็นหน่วยพื้นฐานของโมเลกุลขนาดใหญ่ โมเลกุลขนาดใหญ่ทางชีวภาพที่สำคัญบางชนิด ได้แก่โปรตีนซึ่งประกอบด้วย โมโนเมอร์ ของกรดอะมิโนที่เชื่อมต่อกันด้วยพันธะเปปไทด์และ โมเลกุล DNAซึ่งประกอบด้วยนิวคลีโอไทด์ที่เชื่อมต่อกันด้วยพันธะฟอสโฟไดเอสเทอร์
คุณสมบัติของปฏิกิริยาเคมี
การสังเคราะห์ทางชีวภาพเกิดขึ้นเนื่องจากปฏิกิริยาเคมีหลายชุด เพื่อให้ปฏิกิริยาเหล่านี้เกิดขึ้นได้ จำเป็นต้องมีองค์ประกอบดังต่อไปนี้: [ 1 ]
- สารตั้งต้น : สารประกอบเหล่านี้เป็นโมเลกุลเริ่มต้นหรือสารตั้งต้นในปฏิกิริยา อาจมองได้ว่าเป็นสารทำปฏิกิริยาในกระบวนการทางเคมีที่กำหนด
- พลังงานเคมี : พลังงานเคมีสามารถพบได้ในรูปของโมเลกุลที่มีพลังงานสูง โมเลกุลเหล่านี้จำเป็นสำหรับปฏิกิริยาที่ไม่เอื้อต่อพลังงาน นอกจากนี้การไฮโดรไลซิสของสารประกอบเหล่านี้ยังช่วยขับเคลื่อนปฏิกิริยาไปข้างหน้า โมเลกุลที่มีพลังงานสูง เช่นATPมีฟอสเฟต สามหมู่ บ่อยครั้งที่ฟอสเฟตหมู่สุดท้ายจะแยกออกไปในระหว่างการไฮโดรไลซิสและถ่ายโอนไปยังโมเลกุลอื่น
- ตัวเร่งปฏิกิริยา : ตัวอย่างเช่น อาจเป็นไอออนโลหะหรือโคเอนไซม์ซึ่งทำหน้าที่เร่งปฏิกิริยาโดยการเพิ่มอัตราการเกิดปฏิกิริยาและลดพลังงานกระตุ้น
ในความหมายที่ง่ายที่สุด ปฏิกิริยาที่เกิดขึ้นในการสังเคราะห์ทางชีวภาพมีรูปแบบดังต่อไปนี้: [ 2 ]
![{\displaystyle {\mathrm {ตัวทำปฏิกิริยา} {}\mathrel {\xrightarrow[{\mathrm {enzyme} }]{}} {}\mathrm {ผลิตภัณฑ์} }}](https://wikimedia.org/api/rest_v1/media/math/render/svg/d97fbef58f64fb0b7d05d90fa0a73b05f5919fc8)
รูปแบบต่างๆ ของสมการพื้นฐานนี้ซึ่งจะกล่าวถึงในรายละเอียดเพิ่มเติมในภายหลังมีดังนี้: [ 3 ]
- สารประกอบอย่างง่ายที่ถูกเปลี่ยนไปเป็นสารประกอบอื่น โดยปกติแล้วจะเป็นส่วนหนึ่งของกระบวนการปฏิกิริยาหลายขั้นตอน ตัวอย่างของปฏิกิริยาประเภทนี้เกิดขึ้นระหว่างการสร้างกรดนิวคลีอิกและการเติมประจุให้กับtRNAก่อนการแปลรหัสสำหรับบางขั้นตอนเหล่านี้ จำเป็นต้องใช้พลังงานเคมี:
- สารประกอบอย่างง่ายที่ถูกเปลี่ยนไปเป็นสารประกอบอื่นโดยอาศัยโคแฟคเตอร์ ตัวอย่างเช่น การสังเคราะห์ฟอสโฟลิปิดต้องใช้แอซิทิลโคเอ ในขณะที่การสังเคราะห์สฟิงโกลิปิด ซึ่งเป็นส่วนประกอบของเยื่อหุ้มเซลล์อีกชนิดหนึ่งต้องใช้ NADH และ FADH ในการสร้างโครงสร้างหลักของสฟิงโกซีนสมการทั่วไปสำหรับตัวอย่างเหล่านี้คือ:
- สารประกอบอย่างง่ายที่รวมตัวกันเพื่อสร้างโมเลกุลขนาดใหญ่ ตัวอย่างเช่นกรดไขมันรวมตัวกันเพื่อสร้างฟอสโฟลิปิด ในทางกลับกัน ฟอสโฟลิปิดและคอเลสเตอรอล จะ ทำปฏิกิริยากันโดยไม่ใช้พันธะ โควาเลนต์ เพื่อสร้างชั้นไขมันสองชั้นปฏิกิริยานี้สามารถแสดงได้ดังนี้:
![{\displaystyle {{{\text{Precursor}}\ {\text{โมเลกุล}}}{\vphantom {A}}^{+}\mathrm {ATP} {}\mathrel {\longrightleftharpoons } {}{{\text{product}}\ {\text{AMP}}}{}+{}\mathrm {PP} {\vphantom {A}__{\smash[{t}]{\mathrm {ฉัน} }}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/1b8d846dffb93e3d15a4285e813bce8016794122)
![{\displaystyle {{{\text{โมเลกุลตั้งต้น}}}{}+{}{\text{โคแฟคเตอร์}}{}\mathrel {\xrightarrow[{\mathrm {เอนไซม์} }]{}} {}{\text{โมเลกุลขนาดใหญ่}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/851dd7a34e9d2dbddb7852fb502fe9389ff949a2)

ไขมัน

โมเลกุลขนาดใหญ่ที่ซับซ้อนจำนวนมากถูกสังเคราะห์ขึ้นตามรูปแบบของโครงสร้างที่เรียบง่ายและซ้ำกัน[ 4 ]ตัวอย่างเช่น โครงสร้างที่ง่ายที่สุดของลิปิดคือกรดไขมันกรดไขมันเป็น อนุพันธ์ของ ไฮโดรคาร์บอนโดยมีหมู่คาร์บอกซิลเป็น "หัว" และโซ่ไฮโดรคาร์บอนเป็น "หาง" [ 4 ]กรดไขมันเหล่านี้สร้างส่วนประกอบที่ใหญ่ขึ้น ซึ่งจะรวมเอาปฏิสัมพันธ์ที่ไม่ใช่พันธะโควาเลนต์เพื่อสร้างลิปิดไบเลเยอร์[ 4 ] โซ่กรดไขมันพบได้ในส่วนประกอบหลักสองส่วนของลิปิดในเยื่อหุ้มเซลล์ ได้แก่ฟอสโฟลิปิดและสฟิงโกลิปิดส่วนประกอบหลักที่สามของเยื่อหุ้มเซลล์คือคอเลสเตอรอลซึ่งไม่มีหน่วยกรดไขมันเหล่านี้[ 5 ]
ฟอสโฟลิปิดยูคาริโอต
พื้นฐานของเยื่อชีวภาพ ทั้งหมด ประกอบด้วย โครงสร้าง สองชั้นของฟอสโฟลิปิด[ 6 ] โมเลกุลฟอสโฟลิปิดเป็นแอมฟิพาติกคือมี ส่วนหัวที่เป็นขั้ว ชอบน้ำและส่วนหางที่เป็นขั้วไม่ชอบน้ำ[ 4 ] ส่วนหัวของฟอสโฟลิปิดจะทำปฏิกิริยากันเองและกับตัวกลางที่เป็นน้ำ ในขณะที่ส่วนหางที่เป็นไฮโดรคาร์บอนจะวางตัวอยู่ตรงกลาง ห่างจากน้ำ[ 7 ]ปฏิกิริยาเหล่านี้เป็นแรงผลักดันให้เกิดโครงสร้างสองชั้นที่ทำหน้าที่เป็นกำแพงกั้นไอออนและโมเลกุล[ 8 ]
ฟอสโฟลิปิดมีหลายประเภท ดังนั้นเส้นทางการสังเคราะห์จึงแตกต่างกัน อย่างไรก็ตาม ขั้นตอนแรกในการสังเคราะห์ฟอสโฟลิปิดเกี่ยวข้องกับการสร้างฟอสฟาติเดตหรือ ไดอะซิลกลีเซอรอ ล3-ฟอสเฟตที่เอนโดพลาสมิกเรติคูลัม และเยื่อหุ้มไมโทคอนเดรียชั้นนอก[ 7 ] เส้นทางการสังเคราะห์มีดังต่อไปนี้:

เส้นทางเริ่มต้นด้วยกลีเซอรอล 3-ฟอสเฟต ซึ่งจะถูกแปลงเป็นไลโซฟอสฟาติเดตโดยการเพิ่มสายกรดไขมันที่ได้จากอะซิลโคเอนไซม์เอ [ 9 ] จากนั้น ไลโซฟอสฟาติเดตจะถูกแปลงเป็นฟอสฟาติเดตโดยการเพิ่มสายกรดไขมันอีกสายหนึ่งที่ได้จากอะซิลโคเอนไซม์เอตัวที่สอง ขั้นตอนทั้งหมดนี้ถูกเร่งปฏิกิริยาโดยเอนไซม์ กลีเซอรอลฟอสเฟตอะซิ ลทรานสเฟอเรส[ 9 ]การสังเคราะห์ฟอสโฟลิปิดยังคงดำเนินต่อไปในเอนโดพลาสมิกเรติคูลัม และเส้นทางการสังเคราะห์ทางชีวภาพจะแตกต่างกันไปขึ้นอยู่กับส่วนประกอบของฟอสโฟลิปิดแต่ละชนิด[ 9 ]
สฟิงโกลิปิด
เช่นเดียวกับฟอสโฟลิปิด อนุพันธ์ของกรดไขมันเหล่านี้มีส่วนหัวที่เป็นขั้วและส่วนหางที่ไม่เป็นขั้ว[ 5 ] ต่างจากฟอสโฟลิปิด สฟิงโกลิปิดมีโครงสร้าง หลักเป็นสฟิ งโกซีน[ 10 ]สฟิงโกลิปิดมีอยู่ใน เซลล์ ยูคาริโอตและมีปริมาณมากเป็นพิเศษในระบบประสาทส่วนกลาง [ 7 ] ตัวอย่าง เช่นสฟิ งโกไมอีลิน เป็นส่วนหนึ่งของปลอกไมอีลินของเส้นใยประสาท[ 11 ]
สฟิงโกลิปิดเกิดจากเซราไมด์ซึ่งประกอบด้วยโซ่กรดไขมันที่ติดอยู่กับหมู่เอมีนของโครงสร้างสฟิงโกซีน เซราไมด์เหล่านี้ถูกสังเคราะห์ขึ้นจากการอะซิเลชันของสฟิงโกซีน[ 11 ] เส้นทางการสังเคราะห์ทางชีวภาพของสฟิงโกซีนมีดังต่อไปนี้:

ดังที่ภาพแสดงให้เห็น ในระหว่างการสังเคราะห์สฟิงโกซีนพาลมิโทอิล CoAและเซอรีนจะเกิดปฏิกิริยาควบแน่นซึ่งส่งผลให้เกิดการสร้าง 3-ดีไฮโดรสฟิงกานีน[ 7 ]จากนั้นผลิตภัณฑ์นี้จะถูกรีดิวซ์เพื่อสร้างไดไฮโดรสฟิงโกซีน ซึ่งจะถูกแปลงเป็นสฟิงโกซีนผ่านปฏิกิริยาออกซิเดชันโดยFAD [ 7 ]
คอเลสเตอรอล
ลิปิดชนิดนี้เป็นของโมเลกุลประเภทหนึ่งที่เรียกว่าสเตอรอล [ 5 ] สเตอรอลมีวงแหวนที่เชื่อมต่อกันสี่วงและหมู่ไฮดรอกซิล [ 5 ] คอเลสเตอรอลเป็นโมเลกุลที่สำคัญอย่างยิ่ง ไม่เพียงแต่ทำหน้าที่เป็นส่วนประกอบของเยื่อหุ้มไขมันเท่านั้น แต่ยังเป็นสารตั้งต้นของ ฮอร์โมน สเตียรอยด์ หลายชนิด รวมถึงคอร์ติซอล เทสโทสเตอโรนและเอสโทรเจน[ 12 ]
คอเลสเตอรอลถูกสังเคราะห์จาก อะ เซทิล CoA [ 12 ] เส้นทางแสดงไว้ด้านล่าง:

โดยทั่วไปแล้ว การสังเคราะห์นี้เกิดขึ้นในสามขั้นตอน โดยขั้นตอนแรกเกิดขึ้นในไซโตพลาสซึมและขั้นตอนที่สองและสามเกิดขึ้นในเอนโดพลาสมิกเรติคูลัม[ 9 ]ขั้นตอนต่างๆ มีดังนี้: [ 12 ]
- 1. การสังเคราะห์ไอโซเพนเทนิลไพโรฟอสเฟตซึ่งเป็น "หน่วยพื้นฐาน" ของคอเลสเตอรอล
- 2. การก่อตัวของสควาเลนผ่านการควบแน่นของโมเลกุลไอโซเพนเทนิลฟอสเฟตจำนวนหกโมเลกุล
- 3. การเปลี่ยนสควาเลนเป็นคอเลสเตอรอลผ่านปฏิกิริยาทางเอนไซม์หลายขั้นตอน
นิวคลีโอไทด์
การสังเคราะห์ทางชีวภาพของนิวคลีโอไทด์เกี่ยวข้องกับ ปฏิกิริยา ที่เร่งด้วย เอนไซม์ ซึ่งเปลี่ยนสารตั้งต้นให้เป็นผลิตภัณฑ์ที่ซับซ้อนมากขึ้น[ 1 ]นิวคลีโอไทด์เป็นหน่วยพื้นฐานของDNAและRNA นิวคลีโอ ไทด์ประกอบด้วยวงแหวนห้าสมาชิกที่เกิดจาก น้ำตาล ไรโบสใน RNA และ น้ำตาล ดีออกซีไรโบสใน DNA น้ำตาลเหล่านี้เชื่อมต่อกับ เบส พิวรีนหรือไพริมิดีนด้วยพันธะไกลโคไซด์และ หมู่ ฟอสเฟตที่ตำแหน่ง 5'ของน้ำตาล[ 13 ]
นิวคลีโอไทด์พิวรีน

นิวคลีโอไทด์ DNA อะดีโนซีนและกัวโนซีนประกอบด้วยเบสพิวรีนที่เชื่อมต่อกับน้ำตาลไรโบสด้วยพันธะไกลโคไซด์ ในกรณีของนิวคลีโอไทด์ RNA ดีออกซีอะดีโนซีนและดีออกซีกัวโนซีน เบสพิวรีนจะเชื่อมต่อกับน้ำตาลดีออกซีไรโบสด้วยพันธะไกลโคไซด์ เบสพิวรีนบนนิวคลีโอไทด์ DNA และ RNA ถูกสังเคราะห์ขึ้นในกลไกปฏิกิริยา 12 ขั้นตอนที่มีอยู่ในสิ่งมีชีวิตเซลล์เดียวส่วนใหญ่ยูคาริโอต ชั้นสูงใช้ กลไกปฏิกิริยาที่คล้ายกันใน 10 ขั้นตอน เบสพิวรีนถูกสังเคราะห์โดยการเปลี่ยนฟอสโฟไรโบซิลไพโรฟอสเฟต (PRPP) เป็นอิโนซีนโมโนฟอสเฟต (IMP) ซึ่งเป็นตัวกลางสำคัญตัวแรกในการสังเคราะห์เบสพิวรีน[ 14 ] การดัดแปลงเอนไซม์เพิ่มเติมของIMPจะสร้างเบสอะดีโนซีนและกัวโนซีนของนิวคลีโอไทด์
- ขั้นตอนแรกในการสังเคราะห์พิวรีนคือปฏิกิริยาควบแน่นซึ่งดำเนินการโดยเอนไซม์กลูตามีน-PRPP อะมิโดทรานสเฟอ เรส เอนไซม์นี้จะถ่ายโอนหมู่เอมีนจากกลูตามีนไปยัง PRPP ทำให้เกิด5-ฟอสโฟริโบซิลอะมี น ขั้นตอนต่อไปต้อง อาศัยการกระตุ้นไกลซีนโดยการเติม หมู่ ฟอสเฟตจากATP
- GAR synthetase [ 15 ] ทำการควบแน่นของไกลซีนที่ถูกกระตุ้นบน PRPP ทำให้เกิด ไกลซีนอะไมด์ ไรโบนิวคลีโอไทด์ (GAR)
- เอนไซม์ GAR transformylaseจะเพิ่มหมู่ฟอร์มิลเข้าไปที่หมู่เอมีนของ GAR ทำให้เกิดเป็นฟอร์มิลไกลซินาไมด์ไรโบนิวคลีโอไทด์ (FGAR)
- FGAR amidotransferase [ 16 ] เร่งปฏิกิริยาการเพิ่มกลุ่มไนโตรเจนให้กับ FGAR ทำให้เกิดฟอร์มิลไกลซินามิดีนไรโบนิวคลีโอไทด์ (FGAM)
- เอนไซม์ FGAM cyclaseทำหน้าที่เร่งปฏิกิริยาการปิดวงแหวน ซึ่งเกี่ยวข้องกับการกำจัดโมเลกุลน้ำ ทำให้เกิดวงแหวนอิมิดาโซล 5 วง หรือ 5-อะมิโนอิมิดาโซลไรโบนิวคลีโอไทด์ (AIR)
- N5-CAIR synthetase ถ่ายโอน หมู่ คาร์บอกซิลทำให้เกิด N5-carboxyaminoimidazole ribonucleotide (N5-CAIR) ซึ่งเป็นสารตัวกลาง[ 17 ]
- N5-CAIR mutaseจัดเรียงกลุ่มฟังก์ชันคาร์บอกซิลใหม่และถ่ายโอนไปยังวงแหวนอิมิดาโซล ทำให้เกิดคาร์บอก ซีอะ มิโน-อิมิดาโซลไรโบนิวคลีโอไทด์ (CAIR) กลไกสองขั้นตอนของการสร้าง CAIR จาก AIR ส่วนใหญ่พบในสิ่งมีชีวิตเซลล์เดียว ยูคาริโอตชั้นสูงมีเอนไซม์ AIR คาร์บอกซิเลส[ 18 ] ซึ่งถ่ายโอนกลุ่มคาร์บอกซิลโดยตรงไปยังวงแหวนอิมิดาโซลของ AIR ทำให้เกิด CAIR
- เอนไซม์ SAICAR synthetaseจะสร้างพันธะเปปไทด์ระหว่างแอสปาร์เทตและหมู่คาร์บอกซิลที่เพิ่มเข้ามาของวงแหวนอิมิดาโซล ทำให้เกิดN-succinyl-5-aminoimidazole-4-carboxamide ribonucleotide (SAICAR)
- เอนไซม์ SAICAR lyaseจะกำจัดโครงสร้างคาร์บอนของแอสปาร์เทตที่เติมเข้าไป เหลือไว้เพียงหมู่เอมีโน และเกิดเป็น5-อะมิโนอิมิดาโซล-4-คาร์บอกซาไมด์ไรโบนิวคลีโอไทด์ (AICAR)
- เอนไซม์ AICAR transformylaseจะถ่ายโอนหมู่คาร์บอนิลไปยัง AICAR ทำให้เกิดN-formylaminoimidazole-4-carboxamide ribonucleotide (FAICAR)
- ขั้นตอนสุดท้ายเกี่ยวข้องกับเอนไซม์IMP synthaseซึ่งทำหน้าที่ปิดวงแหวนพิวรีนและสร้างสารตัวกลางอินโนซีนโมโนฟอสเฟต[ 5 ]
นิวคลีโอไทด์ไพริมิดีน
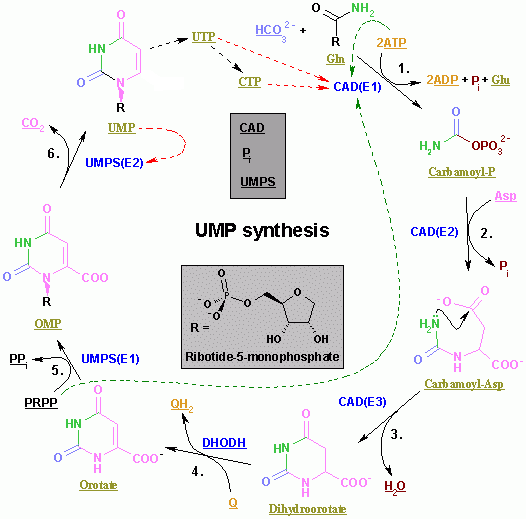
นิว คลีโอไทด์เบสอื่นๆ ของ DNA และ RNA ที่เชื่อมต่อกับน้ำตาลไรโบสผ่านพันธะไกลโคไซด์ ได้แก่ไทมีนไซโตซีนและยูราซิล (ซึ่งพบเฉพาะใน RNA เท่านั้น) การสังเคราะห์ ยูริดีนโมโนฟอสเฟตเกี่ยวข้องกับเอนไซม์ที่อยู่ในเยื่อหุ้มชั้นในของไมโทคอนเดรียและเอนไซม์อเนกประสงค์ที่อยู่ในไซโทซอล[ 19 ]
- ขั้นตอนแรกเกี่ยวข้องกับเอนไซม์คาร์บาโมอิลฟอสเฟตซินเทสที่รวมกลูตามีนกับCO2ในปฏิกิริยาที่ต้องอาศัย ATP เพื่อสร้างร์บาโมอิลฟอสเฟต
- แอสปาร์เทตคาร์บาโมอิลทรานสเฟอเรสจะรวมคาร์บาโมอิลฟอสเฟตกับแอสปาร์เทตเพื่อสร้างยูริโดซัคซิเนต
- ไดไฮโดรออโรเทสทำหน้าที่ปิดวงแหวนซึ่งเป็นปฏิกิริยาที่สูญเสียน้ำไป เพื่อสร้างไดไฮโดรออโรเทต
- ไดไฮโดรออโรเทตดีไฮโดรจีเนสซึ่งตั้งอยู่ในเยื่อหุ้มชั้นในของไมโตคอนเดรีย[ 19 ]ออกซิไดซ์ไดไฮโดรออโรเทตเป็นออโรเทต
- เอนไซม์โอโรเทตฟอสโฟริโบซิลไฮโดรเลส (OMP ไพโรฟอสโฟริเลส) จะควบแน่นโอโรเทตกับPRPPเพื่อสร้างโอโรทิดีน-5'-ฟอสเฟต
- OMP decarboxylaseเร่งปฏิกิริยาการเปลี่ยน orotidine-5'-phosphate เป็นUMP [ 20 ]
หลังจากสังเคราะห์นิวคลีโอไทด์เบสยูริดีนแล้ว เบสอื่นๆ เช่น ไซโตซีนและไทมีนก็จะถูกสังเคราะห์ขึ้น การสังเคราะห์ไซโตซีนเป็นปฏิกิริยาสองขั้นตอนซึ่งเกี่ยวข้องกับการแปลง UMP เป็นUTP การเติม ฟอสเฟต ลงใน UMP จะถูกเร่งปฏิกิริยาโดยเอนไซม์ไคเนส เอนไซม์ CTP ซินเทสจะเร่งปฏิกิริยาขั้นตอนต่อไป คือ การแปลง UTP เป็นCTPโดยการถ่ายโอนหมู่เอมีนจากกลูตามีนไปยังยูริดีน ซึ่งจะสร้างเบสไซโตซีนของ CTP [ 21 ]กลไกซึ่งแสดงปฏิกิริยา UTP + ATP + กลูตามีน ⇔ CTP + ADP + กลูตาเมต มีดังต่อไปนี้:


ไซโตซีนเป็นนิวคลีโอไทด์ที่มีอยู่ในทั้ง DNA และ RNA อย่างไรก็ตาม ยูราซิลพบได้เฉพาะใน RNA เท่านั้น ดังนั้น หลังจากที่ UTP ถูกสังเคราะห์แล้ว จะต้องถูกแปลงเป็น รูปแบบ ดีออกซีเพื่อนำไปรวมเข้ากับ DNA การแปลงนี้เกี่ยวข้องกับเอนไซม์ไรโบนิวคลีโอไซด์ไตรฟอสเฟตเรดักเทสปฏิกิริยานี้ที่กำจัด 2'-OH ของน้ำตาลไรโบสเพื่อสร้างดีออกซีไรโบสจะไม่ได้รับผลกระทบจากเบสที่ติดอยู่กับน้ำตาล ความไม่จำเพาะนี้ทำให้ไรโบนิวคลีโอไซด์ไตรฟอสเฟตเรดักเทสสามารถแปลงนิวคลีโอไทด์ไตรฟอสเฟต ทั้งหมด เป็น ดีออก ซีไรโบนิวคลีโอไทด์ด้วยกลไกที่คล้ายกัน[ 21 ]
ตรงกันข้ามกับยูราซิล เบสไทมีนส่วนใหญ่พบใน DNA ไม่ใช่ RNA โดยปกติเซลล์จะไม่มีเบสไทมีนที่เชื่อมต่อกับน้ำตาลไรโบสใน RNA ซึ่งบ่งชี้ว่าเซลล์จะสังเคราะห์ไทมีนที่เชื่อมต่อกับดีออกซีไรโบสเท่านั้น เอนไซม์ไทมิดิเลตซินเทสมีหน้าที่ในการสังเคราะห์สารตกค้างไทมีนจากdUMPเป็นdTMPปฏิกิริยานี้จะถ่ายโอน หมู่ เมทิลไปยังเบสยูราซิลของ dUMP เพื่อสร้าง dTMP [ 21 ]ปฏิกิริยาของไทมิดิเลตซินเทส dUMP + 5,10-เมทิลีนเตตระไฮโดรโฟเลต ⇔ dTMP + ไดไฮโดรโฟเลต แสดงไว้ทางด้านขวา
ดีเอ็นเอ

แม้ว่าจะมีข้อแตกต่างระหว่าง การสังเคราะห์ DNA ในสิ่งมีชีวิตยูคาริโอตและโปรคาริโอตแต่ส่วนต่อไปนี้จะกล่าวถึงลักษณะสำคัญของการจำลอง DNA ที่สิ่งมีชีวิตทั้งสองชนิดมีร่วมกัน
ดีเอ็นเอประกอบด้วยนิวคลีโอไทด์ที่เชื่อมต่อกันด้วยพันธะฟอสโฟไดเอสเทอร์ [ 4 ] การสังเคราะห์ดีเอ็นเอซึ่งเกิดขึ้นในนิวเคลียสเป็น กระบวนการ แบบกึ่งอนุรักษ์ซึ่งหมายความว่าโมเลกุลดีเอ็นเอที่ได้จะมีสายเดิมจากโครงสร้างแม่และสายใหม่[ 22 ] การสังเคราะห์ดีเอ็นเอถูกเร่งปฏิกิริยาโดยตระกูลของดีเอ็นเอพอลิเมอเรสซึ่งต้องการดีออกซีนิวคลีโอไซด์ไตรฟอสเฟตสี่ตัว สายแม่แบบและไพรเมอร์ที่มี 3'OH อิสระเพื่อรวมนิวคลีโอไทด์[ 23 ]
เพื่อให้การจำลองดีเอ็นเอเกิดขึ้น เอนไซม์ที่เรียกว่า เฮลิเคสจะสร้างจุดแยกการจำลองโดยคลายเกลียวดีเอ็นเอ[ 23 ]โทโปไอโซเมอเรสที่จุดแยกการจำลองจะกำจัดซูเปอร์คอยล์ที่เกิดจากการคลายเกลียวดีเอ็นเอ และโปรตีนที่จับกับดีเอ็นเอสายเดี่ยวจะรักษาแม่แบบดีเอ็นเอสายเดี่ยวสองอันให้คงที่ก่อนการจำลอง[ 13 ]
การสังเคราะห์ DNA เริ่มต้นโดยRNA polymerase primaseซึ่งสร้าง RNA primer ที่มี 3'OH อิสระ[ 23 ] primer นี้จะถูกยึดติดกับแม่แบบ DNA สายเดี่ยว และ DNA polymerase จะยืดสายโซ่โดยการรวมนิวคลีโอไทด์เข้าไป นอกจากนี้ DNA polymerase ยังตรวจสอบความถูกต้องของสาย DNA ที่สังเคราะห์ขึ้นใหม่ด้วย[ 23 ]
ในระหว่างปฏิกิริยาพอลิเมอไรเซชันที่เร่งปฏิกิริยาโดย DNA พอลิเมอเรสการโจมตีแบบนิวคลีโอฟิลิกเกิดขึ้นโดย 3'OH ของสายโซ่ที่กำลังเติบโตบนอะตอมฟอสฟอรัสในสุดของดีออกซีนิวคลีโอไซด์ไตรฟอสเฟต ซึ่งส่งผลให้เกิดการสร้างสะพานฟอสโฟไดเอสเทอร์ที่ยึดนิวคลีโอไทด์ใหม่และปล่อยไพโรฟอสเฟต ออก มา[ 9 ]
ในระหว่างการจำลองแบบ จะมีการสร้างสายดีเอ็นเอสองประเภทพร้อมกัน ได้แก่สายนำ (leading strand ) ซึ่งถูกสังเคราะห์อย่างต่อเนื่องและเติบโตไปทางจุดแยกการจำลองแบบ และสายตาม (lagging strand ) ซึ่งถูกสร้างขึ้นอย่างไม่ต่อเนื่องในชิ้นส่วนโอคาซากิ (Okazaki fragments)และเติบโตออกไปจากจุดแยกการจำลองแบบ[ 22 ] ชิ้นส่วนโอคาซากิจะ ถูกเชื่อม ต่อกัน ด้วยพันธะโควาเลนต์ โดยเอนไซม์ดีเอ็นเอไลเกส (DNA ligase ) เพื่อสร้างสายที่ต่อเนื่อง[ 22 ] จากนั้น เพื่อให้การจำลองแบบดีเอ็นเอเสร็จสมบูรณ์ ไพรเมอร์อาร์เอ็นเอ (RNA primers) จะถูกกำจัดออกไป และช่องว่างที่เกิดขึ้นจะถูกแทนที่ด้วยดีเอ็นเอและเชื่อมต่อกันโดยใช้เอนไซม์ดีเอ็นเอไลเกส[ 22 ]
กรดอะมิโน
โปรตีนเป็นพอลิเมอร์ที่ประกอบขึ้นจากกรดอะมิโนที่เชื่อมต่อกันด้วยพันธะเปปไทด์มีกรดอะมิโนมากกว่า 300 ชนิดที่พบในธรรมชาติ ซึ่งมีเพียง 22 ชนิดเท่านั้นที่เรียกว่ากรดอะมิโนโปรตีนโนเจนิกซึ่งเป็นหน่วยพื้นฐานของโปรตีน[ 24 ] มีเพียงพืชสีเขียวและจุลินทรีย์ ส่วนใหญ่เท่านั้น ที่สามารถสังเคราะห์กรดอะมิโนมาตรฐานทั้ง 20 ชนิดที่จำเป็นสำหรับสิ่งมีชีวิตทุกชนิดได้ สัตว์ เลี้ยงลูกด้วยนมสามารถสังเคราะห์กรดอะมิโนมาตรฐานได้เพียง 10 ชนิดจาก 20 ชนิดเท่านั้น กรดอะมิโนอื่นๆ ได้แก่วาลีน เมไท โอนีนลิวซีนไอ โซลิวซีน ฟี นิล อะลานีนไล ซีน ทรี โอนีน และ ท ริปโตเฟนสำหรับผู้ใหญ่ และฮิสติดีนและอาร์จินีน สำหรับทารก นั้นได้มาจากอาหาร[ 25 ]
โครงสร้างพื้นฐานของกรดอะมิโน

โครงสร้างทั่วไปของกรดอะมิโนมาตรฐานประกอบด้วยหมู่เอมีนหลักหมู่คาร์บอกซิลและหมู่ฟังก์ชันที่ติดอยู่กับคาร์บอนอัลฟากรดอะมิโนที่แตกต่างกันจะถูกระบุโดยหมู่ฟังก์ชัน เนื่องจากมีหมู่ที่แตกต่างกันสามหมู่ที่ติดอยู่กับคาร์บอนอัลฟา กรดอะมิโนจึงเป็นโมเลกุลที่ไม่สมมาตรสำหรับกรดอะมิโนมาตรฐานทั้งหมด ยกเว้นไกลซีนคาร์บอนอัลฟาเป็นศูนย์กลางไครัลในกรณีของไกลซีน คาร์บอนอัลฟามีอะตอมไฮโดรเจนสองอะตอม จึงเพิ่มความสมมาตรให้กับโมเลกุลนี้ ยกเว้นโพรลีนกรดอะมิโนทั้งหมดที่พบในสิ่งมีชีวิตมี โครงสร้างแบบ L-isoformโพรลีนมีหมู่ฟังก์ชันบนคาร์บอนอัลฟาที่สร้างวงแหวนกับหมู่เอมีน[ 24 ]

แหล่งไนโตรเจน
ขั้นตอนสำคัญอย่างหนึ่งในการสังเคราะห์กรดอะมิโนคือการเติมหมู่ไนโตรเจนลงบนคาร์บอนอัลฟา ในเซลล์มีกระบวนการหลักสองกระบวนการในการเติมหมู่ไนโตรเจน กระบวนการหนึ่งเกี่ยวข้องกับเอนไซม์กลูตามีนออกโซกลูตาเรตอะมิโนทรานสเฟอเรส (GOGAT) ซึ่งจะกำจัดหมู่เอไมด์อะมิโนของกลูตามีนและถ่ายโอนไปยัง2-ออกโซกลูตาเรต ทำให้เกิดโมเลกุล กลูตา เมต สองโมเลกุล ในปฏิกิริยาเร่งปฏิกิริยานี้ กลูตามีนทำหน้าที่เป็นแหล่งไนโตรเจน ภาพประกอบแสดงปฏิกิริยานี้แสดงอยู่ทางด้านขวา
เส้นทางอื่นในการรวมไนโตรเจนเข้ากับคาร์บอนอัลฟาของกรดอะมิโนเกี่ยวข้องกับเอนไซม์กลูตาเมตดีไฮโดรจีเนส (GDH) GDH สามารถถ่ายโอนแอมโมเนียไปยัง 2-ออกโซกลูตาเรตและสร้างกลูตาเมตได้ นอกจากนี้ เอนไซม์กลูตามีนซินเทส (GS) ยังสามารถถ่ายโอนแอมโมเนียไปยังกลูตาเมตและสังเคราะห์กลูตามีน ซึ่งเป็นการเติมเต็มกลูตามีน[ 26 ]
กรดอะมิโนในกลุ่มกลูตาเมต
ตระกูลกลูตาเมตของกรดอะมิโนประกอบด้วยกรดอะมิโนที่ได้มาจากกรดอะมิโนกลูตาเมต ตระกูลนี้ได้แก่ กลูตาเมต กลูตามีนโพรลีนและอาร์จินีนนอกจากนี้ ตระกูลนี้ยังรวมถึงกรดอะมิโนไลซีนซึ่งได้มาจากα-คีโตกลูตาเรต[ 27 ]
การสังเคราะห์กลูตาเมตและกลูตามีนเป็นขั้นตอนสำคัญในการดูดซึมไนโตรเจนที่กล่าวถึงข้างต้น เอนไซม์GOGATและGDHทำหน้าที่เร่งปฏิกิริยา การดูดซึมไนโตรเจน
ในแบคทีเรีย เอนไซม์กลูตาเมต 5-ไคเนสจะเริ่มต้นการสังเคราะห์โพรลีนโดยการถ่ายโอนหมู่ฟอสเฟตจาก ATP ไปยังกลูตาเมต ปฏิกิริยาถัดไปจะถูกเร่งปฏิกิริยาโดยเอนไซม์ไพร์โรลีน-5-คาร์บอกซิเลตซินเทส (P5CS) ซึ่งเร่งปฏิกิริยาการรีดิวซ์ หมู่ ϒ-คาร์บอกซิลของ L-กลูตาเมต 5-ฟอสเฟต ส่งผลให้เกิดกลูตาเมตเซมิอัลดีไฮด์ ซึ่งจะเกิดการสร้างวงแหวนโดยธรรมชาติเป็นไพร์โรลีน-5-คาร์บอกซิเลต ไพร์โรลีน-5-คาร์บอกซิเลตจะถูกรีดิวซ์เพิ่มเติมโดยเอนไซม์ไพร์โรลีน-5-คาร์บอกซิเลตรีดักเทส (P5CR) เพื่อให้ได้กรดอะมิโนโพรลีน[ 28 ]
ในขั้นตอนแรกของการสังเคราะห์อาร์จินีนในแบคทีเรีย กลูตาเมตจะถูกอะเซทิเลตโดยการถ่ายโอนหมู่แอเซทิลจากแอเซทิล-โคเอที่ตำแหน่ง N-α ซึ่งจะป้องกันการเกิดวงแหวนโดยธรรมชาติ เอนไซม์N-acetylglutamate synthase (glutamate N-acetyltransferase) ทำหน้าที่เร่งปฏิกิริยาในขั้นตอนการอะเซทิเลต ขั้นตอนต่อมาจะถูกเร่งปฏิกิริยาโดยเอนไซม์N-acetylglutamate kinase , N-acetyl-gamma-glutamyl-phosphate reductaseและacetylornithine/succinyldiamino pimelate aminotransferaseและได้ผลลัพธ์เป็น N-acetyl-L-ornithine หมู่แอเซทิลของ acetylornithine จะถูกกำจัดออกโดยเอนไซม์acetylornithinase (AO) หรือornithine acetyltransferase (OAT) และได้ผลลัพธ์เป็นornithineจากนั้นเอนไซม์ซิทรูลีนและอาร์จินิโนซัคซิเนตจะเปลี่ยนออร์นิทีนเป็นอาร์จินีน[ 29 ]

มีเส้นทางการสังเคราะห์ไลซีนที่แตกต่างกันสองเส้นทาง ได้แก่ เส้นทางกรดไดอะมิโนพิเมลิก และเส้นทางอัลฟา-อะมิโนอะดิเพต เส้นทางการสังเคราะห์ที่พบได้บ่อยที่สุดในสองเส้นทางนี้คือเส้นทางกรดไดอะมิโนพิเมลิก ซึ่งประกอบด้วยปฏิกิริยาเอนไซม์หลายอย่างที่เพิ่มกลุ่มคาร์บอนให้กับแอสปาร์เทตเพื่อให้ได้ไลซีน: [ 30 ]
- แอสปาร์เทตไคเนสเริ่มต้นกระบวนการสังเคราะห์กรดไดอะมิโนพิเมลิกโดยการฟอสฟอริเลตแอสปาร์เทตและสร้างแอสปาร์ทิลฟอสเฟต
- เอนไซม์แอสปาร์เทตเซมิอัลดีไฮด์ดีไฮโดรจีเนสทำหน้าที่ เร่งปฏิกิริยารีดักชันของแอสปาร์ทิลฟอสเฟตโดยอาศัย NADPHเพื่อให้ได้แอสปาร์เทตเซมิอัลดีไฮด์
- เอนไซม์ 4-hydroxy-tetrahydrodipicolinate synthaseจะเพิ่ม หมู่ ไพรูเวตให้กับ β-aspartyl-4-semialdehyde และกำจัดโมเลกุลน้ำออกไป ทำให้เกิดการสร้างวงแหวนและได้เป็น (2S,4S)-4-hydroxy-2,3,4,5-tetrahydrodipicolinate
- 4-ไฮดรอกซี-เตตระไฮโดรไดพิโคลิเนต รีดักเทส เร่งปฏิกิริยารีดักชันของ (2S,4S)-4-ไฮดรอกซี-2,3,4,5-เตตระไฮโดรไดพิโคลิเนต โดย NADPH เพื่อให้ได้ Δ'-พิเพอริดีน-2,6-ไดคาร์บอกซิเลต (2,3,4,5-เตตระไฮโดรไดพิโคลิเนต) และ
- เอนไซม์ Tetrahydrodipicolinate acyltransferaseทำหน้าที่เร่งปฏิกิริยาอะซิติเลชัน ซึ่งส่งผลให้เกิดการเปิดวงแหวนและให้ผลผลิตเป็น N-acetyl α-amino-ε-ketopimelate
- N-succinyl-α-amino-ε-ketopimelate-glutamate aminotransaminaseเร่งปฏิกิริยาการถ่ายโอนหมู่เอมีนซึ่งกำจัดหมู่คีโตของ N-acetyl α-amino-ε-ketopimelate และแทนที่ด้วยหมู่เอมีนเพื่อให้ได้ N-succinyl-L-diaminopimelate [ 31 ]
- N-acyldiaminopimelate deacylaseเร่งปฏิกิริยาการดีอะซิเลชันของ N-succinyl-L-diaminopimelate เพื่อให้ได้ L,L-diaminopimelate [ 32 ]
- DAP epimeraseเร่งปฏิกิริยาการเปลี่ยน L,L-diaminopimelate ไปเป็นL,L-diaminopimelate รูปแบบเมโซ[ 33 ]
- เอนไซม์ DAP decarboxylaseทำหน้าที่เร่งปฏิกิริยาการกำจัดหมู่คาร์บอกซิล ทำให้ได้แอล-ไลซีน
กรดอะมิโนในกลุ่มเซริน
กรด อะมิโนในกลุ่มเซ รีนประกอบด้วย เซรีนซิสเทอีนและไกลซีนจุลินทรีย์และพืชส่วนใหญ่ได้รับกำมะถันเพื่อสังเคราะห์เมไทโอนีนจากกรดอะมิโนซิสเทอีน นอกจากนี้ การเปลี่ยนเซรีนเป็นไกลซีนยังให้คาร์บอนที่จำเป็นสำหรับการสังเคราะห์เมไทโอนีนและฮิสติดีน อีก ด้วย[ 27 ]
ในระหว่างการสังเคราะห์เซรีน[ 34 ]เอนไซม์ฟอสโฟกลีเซอเรตดีไฮโดรจี เนส จะเร่งปฏิกิริยาเริ่มต้นที่ออกซิไดซ์3-ฟอสโฟ-D-กลีเซอเรตเพื่อให้ได้3-ฟอสโฟโนออกซีไพรูเวต [ 35 ] ปฏิกิริยา ต่อไปนี้ถูกเร่งโดยเอนไซม์ฟอสโฟเซรีนอะมิโนทรานสเฟอเรสซึ่งถ่ายโอนหมู่เอมีนจากกลูตาเมตไปยัง 3-ฟอสโฟโนออกซีไพรูเวตเพื่อให้ได้L-ฟอสโฟเซรีน [ 36 ] ขั้น ตอนสุดท้ายถูกเร่งโดยเอนไซม์ฟอสโฟเซรีนฟอสฟาเทสซึ่งกำจัดหมู่ฟอสเฟตออกจาก L-ฟอสโฟเซรีนเพื่อให้ได้L-เซรีน[ 37 ]
มีเส้นทางการสังเคราะห์ไกลซีนที่รู้จักกันสองเส้นทาง สิ่งมีชีวิตที่ใช้เอทานอลและอะซิเตตเป็นแหล่งคาร์บอนหลักจะใช้ เส้นทาง ไกลโคเนโอเจนิคในการสังเคราะห์ไกลซีน เส้นทางการสังเคราะห์ไกลซีนอีกเส้นทางหนึ่งเรียกว่า เส้นทาง ไกลโคไลซิสเส้นทางนี้จะเปลี่ยนซีรีนที่สังเคราะห์จากสารตัวกลางของไกลโคไลซิสให้เป็นไกลซีน ในเส้นทางไกลโคไลซิส เอนไซม์ซีรีนไฮดรอกซีเมทิลทรานสเฟอเร ส จะเร่งปฏิกิริยาการแตกตัวของซีรีนเพื่อให้ได้ไกลซีน และถ่ายโอนหมู่คาร์บอนที่แตกตัวของซีรีนไปยังเตตระไฮโดรโฟเลตทำให้เกิด5,10-เมทิลีนเตตระไฮโดรโฟเลต[ 38 ]
การสังเคราะห์ซิสเตอีนเป็นปฏิกิริยาสองขั้นตอนที่เกี่ยวข้องกับการรวมกำมะถันอนินทรี ย์ ในจุลินทรีย์และพืช เอนไซม์เซอรีนอะเซทิลทรานสเฟอเรสจะเร่งปฏิกิริยาการถ่ายโอนหมู่แอเซทิลจากแอเซทิล-CoAไปยัง L-เซอรีนเพื่อให้ได้O-แอเซทิล-L-เซอรีน [ 39 ] ขั้น ตอนปฏิกิริยาถัดไปซึ่งเร่งปฏิกิริยาโดยเอนไซม์O-แอเซทิลเซอรีน (ไทออล) ไลเอสจะแทนที่หมู่แอเซทิลของ O-แอเซทิล-L-เซอรีนด้วยซัลไฟด์เพื่อให้ได้ซิสเตอีน[ 40 ]
กรดอะมิโนในกลุ่มแอสปาร์เทต
กรด อะมิโนในกลุ่มแอสปา ร์เทตประกอบด้วย: ทรีโอนีนไลซีนเมไทโอนีนไอ โซ ลิวซีน และแอสปาร์เทต ไลซีนและไอโซลิวซีนถือเป็นส่วนหนึ่งของกลุ่มแอสปาร์เทต แม้ว่าโครงสร้างคาร์บอนบางส่วนจะมาจากไพรูเวตก็ตาม ในกรณีของเมไทโอนีน คาร์บอนเมทิลได้มาจากซีรีนและหมู่ซัลเฟอร์ แต่ในสิ่งมีชีวิตส่วนใหญ่ ได้มาจากซิสเทอีน[ 27 ]
การสังเคราะห์แอสปาร์เทตเป็นปฏิกิริยาขั้นตอนเดียวที่เร่งปฏิกิริยาโดยเอนไซม์ตัวเดียว เอนไซม์แอสปาร์ เทตอะมิโนทรานสเฟอเรส เร่งปฏิกิริยาการถ่ายโอนหมู่เอมีนจากแอสปาร์เทตไปยังอัลฟา-คีโตกลูตาเรตเพื่อให้ได้กลูตาเมตและออกซาโลอะซิเตต [ 41 ] แอ สปาราจีนถูกสังเคราะห์โดยการเติมหมู่เอมีนลงบนแอสปาร์เทตโดยอาศัย ATP เอนไซม์แอสปาราจีนซินเทสเร่งปฏิกิริยาการเติมไนโตรเจนจากกลูตามีนหรือแอมโมเนียที่ละลายได้ไปยังแอสปาร์เทตเพื่อให้ได้แอสปาราจีน[ 42 ]
เส้นทางการสังเคราะห์กรดไดอะมิโนพิเมลิกของไลซีนเป็นของตระกูลกรดอะมิโนแอสปาร์เทต เส้นทางนี้เกี่ยวข้องกับปฏิกิริยาเร่งปฏิกิริยาโดยเอนไซม์เก้าปฏิกิริยาที่เปลี่ยนแอสปาร์เทตเป็นไลซีน[ 43 ]
- แอสปาร์เทตไคเนสเร่งปฏิกิริยาขั้นตอนเริ่มต้นในเส้นทางไดอะมิโนพิเมลิกแอซิดโดยการถ่ายโอนฟอสฟอริลจาก ATP ไปยังกลุ่มคาร์บอกซิเลตของแอสปาร์เทต ซึ่งให้ผลลัพธ์เป็นแอสปาร์ทิล-β-ฟอสเฟต[ 44 ]
- แอสปาร์เทต-เซมิอัลดีไฮด์ดีไฮโดรจีเนสเร่งปฏิกิริยารีดักชันโดยการกำจัดฟอสเฟตของแอสปาร์ทิล-β-ฟอสเฟตเพื่อให้ได้แอสปาร์เทต-β-เซมิอัลดีไฮด์[ 45 ]
- ไดไฮโดรไดพิโคลิเนตซินเทสเร่งปฏิกิริยา การ ควบแน่นของแอสปาร์เทต-β-เซมิอัลดีไฮด์กับไพรูเวตเพื่อให้ได้กรดไดไฮโดรไดพิโคลิก[ 46 ]
- 4-ไฮดรอกซี-เตตระไฮโดรไดพิโคลิเนต รีดักเทสเร่งปฏิกิริยารีดักชันของกรดไดไฮโดรไดพิโคลินิกเพื่อให้ได้กรดเตตระไฮโดรไดพิโคลินิก[ 47 ]
- Tetrahydrodipicolinate N-succinyltransferaseเร่งปฏิกิริยาการถ่ายโอนกลุ่มซัคซินิลจากซัคซินิล-CoA ไปยังกรดเตตระไฮโดรไดพิโคลิกเพื่อให้ได้ N-succinyl-L-2,6-diaminoheptanedioate [ 48 ]
- N-succinyldiaminopimelate aminotransferase เร่งปฏิกิริยาการถ่ายโอนหมู่เอมีนจากกลูตาเมตไปยัง N-succinyl-L-2,6-diaminoheptanedioate เพื่อให้ได้ N-succinyl-L,L-diaminopimelic acid [ 49 ]
- ซัคซินิล-ไดอะมิโนพิเมเลต เดซัคซินิเลสเร่งปฏิกิริยาการกำจัดหมู่เอซิลจากกรด N-ซัคซินิล-L,L-ไดอะมิโนพิเมลิก เพื่อให้ได้กรด L,L-ไดอะมิโนพิเมลิก[ 50 ]
- ไดอะมิโนพิเมเลตอีพิเมอเรสเร่งปฏิกิริยาการผกผันของคาร์บอนอัลฟาของกรด L,L-ไดอะมิโนพิเมลิกเพื่อให้ได้กรดเมโซ-ไดอะมิโนพิเมลิก[ 51 ]
- ไซอะมิโนพิเมเลตดีคาร์บอกซิเลสเร่งปฏิกิริยาขั้นตอนสุดท้ายในการสังเคราะห์ไลซีนโดยการกำจัดหมู่คาร์บอนไดออกไซด์ออกจากเมโซ-ไดอะมิโนพิเมลิกแอซิดเพื่อให้ได้แอล-ไลซีน[ 52 ]
โปรตีน

การสังเคราะห์โปรตีนเกิดขึ้นผ่านกระบวนการที่เรียกว่าการแปล [ 53 ] ใน ระหว่างการแปล สารพันธุกรรมที่เรียกว่าmRNA จะถูกอ่านโดยไรโบโซมเพื่อสร้างสายโพลีเปปไทด์ ของโปรตีน [ 53 ] กระบวนการนี้ต้องการRNA ถ่ายโอน (tRNA) ซึ่งทำหน้าที่เป็นตัวเชื่อมโดยการจับ กับ กรดอะมิโนที่ปลายด้านหนึ่งและโต้ตอบกับ mRNA ที่ปลายอีกด้านหนึ่ง การจับคู่ระหว่าง tRNA และ mRNA ในภายหลังทำให้มั่นใจได้ว่ากรดอะมิโนที่ถูกต้องจะถูกเพิ่มเข้าไปในสาย[ 53 ] การสังเคราะห์โปรตีนเกิดขึ้นในสามขั้นตอน ได้แก่ การเริ่มต้น การยืดตัว และการสิ้นสุด[ 13 ] การแปล ของโปรคาริโอต ( อาร์เคียและแบคทีเรีย ) แตกต่างจากการแปลของยูคาริโอตอย่างไรก็ตาม ส่วนนี้จะเน้นไปที่ความเหมือนกันระหว่างสิ่งมีชีวิตทั้งสองเป็นส่วนใหญ่
ข้อมูลพื้นฐานเพิ่มเติม
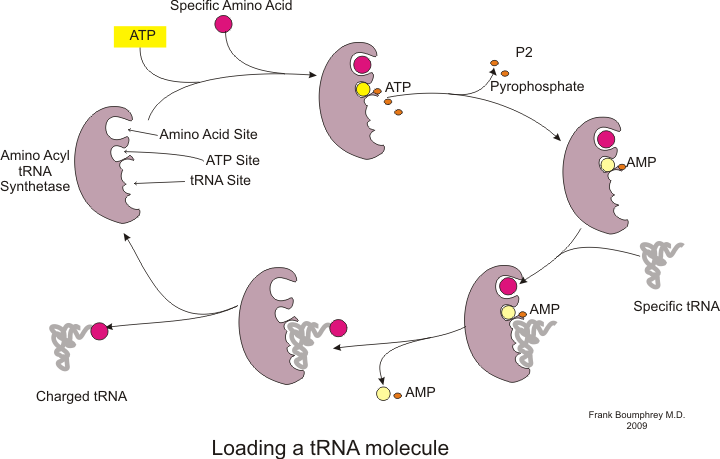
ก่อนที่การแปลจะเริ่มต้นได้ กระบวนการจับกรดอะมิโนเฉพาะกับ tRNA ที่สอดคล้องกันจะต้องเกิดขึ้น ปฏิกิริยานี้เรียกว่าการเติม tRNA ซึ่งถูกเร่งปฏิกิริยาโดยอะมิโนเอซิล tRNA ซินเทส [ 54 ] tRNAซินเทสเฉพาะมีหน้าที่ในการจดจำและเติมกรดอะมิโนเฉพาะ[ 54 ]นอกจากนี้ เอนไซม์นี้ยังมีบริเวณตัวแยกแยะพิเศษเพื่อให้แน่ใจว่าการจับกันระหว่าง tRNA และกรดอะมิโนที่เกี่ยวข้องนั้นถูกต้อง[ 54 ]ขั้นตอนแรกสำหรับการเชื่อมต่อกรดอะมิโนกับ tRNA ที่สอดคล้องกันคือการสร้างอะมิโนเอซิล-AMP: [ 54 ]
![{\displaystyle {{{\text{Amino}}\ {\text{acid}}}{}+{}\mathrm {ATP} {}\mathrel {\longrightleftharpoons } {}{\text{aminoacyl-AMP}}{}+{}\mathrm {PP} {\vphantom {A}__{\smash[{t}]{\mathrm {i} }}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/430f5fad4c9b8a572dbded184d45ec2a9b36693a)
ตามด้วยการถ่ายโอนกลุ่มอะมิโนเอซิลจากอะมิโนเอซิล-AMP ไปยังโมเลกุล tRNA โมเลกุลที่ได้คืออะมิโนเอซิล-tRNA : [ 54 ]

การรวมกันของสองขั้นตอนดังกล่าว ซึ่งทั้งสองขั้นตอนถูกเร่งปฏิกิริยาโดยเอนไซม์อะมิโนเอซิลทีอาร์เอ็นเอซินเทส จะสร้างทีอาร์เอ็นเอที่มีประจุพร้อมที่จะเพิ่มกรดอะมิโนเข้าไปในสายโพลีเปปไทด์ที่กำลังเติบโต
นอกจากจะจับกับกรดอะมิโนแล้ว tRNA ยังมีหน่วยนิวคลีโอไทด์สามตัวที่เรียกว่าแอนติโคดอนซึ่งจับคู่กับนิวคลีโอไทด์สามตัวที่เฉพาะเจาะจงบน mRNA ที่เรียกว่าโคดอนโคดอนเข้ารหัสกรดอะมิโนที่เฉพาะเจาะจง[ 55 ] ปฏิสัมพันธ์นี้เป็นไปได้ด้วยไรโบโซม ซึ่งทำหน้าที่เป็นแหล่งสังเคราะห์โปรตีน ไรโบโซมมีตำแหน่งการจับ tRNA สามตำแหน่ง ได้แก่ ตำแหน่งอะมิโนเอซิล (ตำแหน่ง A) ตำแหน่งเปปทิดิล (ตำแหน่ง P) และตำแหน่งทางออก (ตำแหน่ง E) [ 56 ]
มีโคดอนจำนวนมากภายในทรานสคริปต์ mRNA และเป็นเรื่องปกติมากที่กรดอะมิโนจะถูกกำหนดโดยโคดอนมากกว่าหนึ่งตัว ปรากฏการณ์นี้เรียกว่าความเสื่อม[ 57 ]โดยรวมแล้วมีโคดอน 64 ตัว โคดอน 61 ตัวแต่ละรหัสสำหรับกรดอะมิโน 1 ใน 20 ตัว ในขณะที่โคดอนที่เหลือระบุการสิ้นสุดของสายโซ่[ 57 ]
การแปลเป็นขั้นตอน
ดังที่กล่าวไว้ก่อนหน้านี้ การแปลรหัสพันธุกรรมเกิดขึ้นในสามขั้นตอน ได้แก่ การเริ่มต้น การยืดออก และการสิ้นสุด

ขั้นตอนที่ 1: การเริ่มต้น
การเสร็จสิ้นของขั้นตอนเริ่มต้นขึ้นอยู่กับเหตุการณ์สามประการต่อไปนี้: [ 13 ]
1. การดึงดูดไรโบโซมเข้าสู่ mRNA
2. การจับตัวของ tRNA ตัวเริ่มต้นที่มีประจุเข้ากับตำแหน่ง P ของไรโบโซม
3. การจัดเรียงตัวที่ถูกต้องของไรโบโซมให้ตรงกับรหัสเริ่มต้นของ mRNA
ขั้นตอนที่ 2: การยืด
หลังจากการเริ่มต้น สายโซ่โพลีเปปไทด์จะขยายออกไปโดยผ่านปฏิกิริยาแอนติโคดอน:โคดอน โดยไรโบโซมจะเพิ่มกรดอะมิโนเข้าไปในสายโซ่โพลีเปปไทด์ทีละตัว ขั้นตอนต่อไปนี้จะต้องเกิดขึ้นเพื่อให้แน่ใจว่ามีการเพิ่มกรดอะมิโนอย่างถูกต้อง: [ 58 ]
1. การจับกันของ tRNA ที่ถูกต้องเข้ากับตำแหน่ง A ของไรโบโซม
2. การเกิดพันธะเปปไทด์ระหว่าง tRNA ในตำแหน่ง A กับสายโพลีเปปไทด์ที่ติดอยู่กับ tRNA ในตำแหน่ง P
3. การเคลื่อนย้ายหรือการเคลื่อนที่ไปข้างหน้าของคอมเพล็กซ์ tRNA-mRNA โดยนิวคลีโอไทด์สามตัว
กระบวนการเคลื่อนย้าย "เริ่มต้น" การเคลื่อนที่ของ tRNA ที่ตำแหน่ง E และย้าย tRNA จากตำแหน่ง A ไปยังตำแหน่ง P ทำให้ตำแหน่ง A ว่างลงเพื่อให้ tRNA ตัวใหม่เข้ามาเพิ่มกรดอะมิโนอีกตัวหนึ่ง
ขั้นตอนที่ 3: การยุติ
ขั้นตอนสุดท้ายของการแปลเกิดขึ้นเมื่อรหัสหยุดเข้าสู่ไซต์ A [ 1 ]จากนั้นขั้นตอนต่อไปนี้จะเกิดขึ้น:
1. การรับรู้โคดอนโดยปัจจัยปลดปล่อยซึ่งทำให้เกิดการไฮโดรไลซิสของสายโซ่โพลีเปปไทด์จาก tRNA ที่อยู่ในไซต์ P [ 1 ]
2. การปล่อยโซ่โพลีเปปไทด์[ 57 ]
3. การแยกตัวและการ "รีไซเคิล" ของไรโบโซมสำหรับกระบวนการแปลในอนาคต[ 57 ]
ตารางสรุปผู้มีบทบาทสำคัญในวงการแปลมีดังต่อไปนี้:
| ผู้เล่นหลักในวงการแปล | ขั้นตอนการแปล | วัตถุประสงค์ |
|---|---|---|
| tRNA synthetase | ก่อนการเริ่มต้น | รับผิดชอบในการชาร์จ tRNA |
| เอ็มอาร์เอ็นเอ | การเริ่มต้น การยืดออก การสิ้นสุด | แม่แบบสำหรับการสังเคราะห์โปรตีน ประกอบด้วยบริเวณที่เรียกว่าโคดอน ซึ่งเข้ารหัสกรดอะมิโน |
| ทีอาร์เอ็นเอ | การเริ่มต้น การยืดออก การสิ้นสุด | จับกับตำแหน่ง A, P, E ของไรโบโซม; แอนติโคดอนจับคู่กับโคดอนของ mRNA เพื่อให้แน่ใจว่ากรดอะมิโนที่ถูกต้องจะถูกรวมเข้ากับสายโพลีเปปไทด์ที่กำลังเติบโต |
| ไรโบโซม | การเริ่มต้น การยืดออก การสิ้นสุด | ควบคุมการสังเคราะห์โปรตีนและเร่งปฏิกิริยาการสร้างพันธะเปปไทด์ |
โรคที่เกี่ยวข้องกับการขาดสารโมเลกุลขนาดใหญ่

ความผิดพลาดในกระบวนการสังเคราะห์ทางชีวภาพอาจส่งผลเสียร้ายแรง รวมถึงการผิดรูปของโมเลกุลขนาดใหญ่ หรือการผลิตโมเลกุลที่ทำหน้าที่ได้ไม่เพียงพอ ตัวอย่างด้านล่างนี้แสดงให้เห็นถึงความเสียหายที่เกิดขึ้นเนื่องจากความไม่มีประสิทธิภาพเหล่านี้
- ภาวะคอเลสเตอรอลสูงในครอบครัว : ความผิดปกตินี้มีลักษณะเฉพาะคือการขาดตัวรับ LDL ที่ ใช้งานได้ [ 59 ]การขาดการสร้างตัวรับ LDL อาจทำให้เกิดตัวรับที่ผิดพลาดซึ่งขัดขวางเส้นทางเอนโดไซโทซิส ยับยั้งการเข้าของ LDL เข้าสู่ตับและเซลล์อื่นๆ[ 59 ] สิ่งนี้ทำให้เกิดการสะสมของ LDL ในพลาสมาในเลือด ซึ่งส่งผลให้เกิดคราบพลัคหลอดเลือดแดงที่ทำให้หลอดเลือดตีบและเพิ่มความเสี่ยงต่อการเกิดโรคหัวใจวาย[ 59 ]
- กลุ่มอาการเลช-นัยฮาน : โรคทางพันธุกรรมนี้มีลักษณะเฉพาะคือการทำร้ายตัวเองความบกพร่องทางสติปัญญา และโรคเกาต์[ 60 ] เกิดจากการขาด เอนไซม์ ไฮโปแซนทีน-กัวนีนฟอสโฟริโบซิลทรานสเฟอเรสซึ่งเป็นเอนไซม์ที่จำเป็นสำหรับการสร้างนิวคลีโอไทด์พิวรีน[ 60 ] การขาดเอนไซม์ทำให้ระดับของนิวคลีโอไทด์ที่จำเป็นลดลงและทำให้เกิดการสะสมของสารตัวกลาง ในการสังเคราะห์ทางชีวภาพ ซึ่งส่งผลให้เกิดพฤติกรรมผิดปกติดังกล่าว[ 60 ]
- ภาวะภูมิคุ้มกันบกพร่องร่วมรุนแรง (SCID) : SCID มีลักษณะเฉพาะคือการสูญเสียเซลล์T [ 61 ]การขาดแคลนส่วนประกอบของระบบภูมิคุ้มกันเหล่านี้ทำให้มีความเสี่ยงต่อเชื้อโรคมากขึ้น เนื่องจากผู้ป่วยไม่สามารถสร้างความจำทางภูมิคุ้มกันได้[ 61 ]ความผิดปกติทางภูมิคุ้มกันนี้เกิดจากการขาดกิจกรรมของอะดีโนซีนดีอะมี เนส ซึ่งทำให้เกิดการสะสมของdATPโมเลกุล dATP เหล่านี้จะยับยั้งไรโบนิวคลีโอไทด์รีดักเทส ซึ่งป้องกันการสังเคราะห์ DNA [ 61 ]
- โรคฮันติงตัน : โรค ทางระบบประสาท นี้ เกิดจากข้อผิดพลาดที่เกิดขึ้นระหว่างการสังเคราะห์ DNA [ 62 ]ข้อผิดพลาดหรือการกลายพันธุ์เหล่านี้ทำให้เกิดการแสดงออกของ โปรตีน ฮันติงติน กลายพันธุ์ ซึ่งมี สารตกค้าง ของกลูตามี นซ้ำๆ ที่ถูกเข้ารหัสโดยการขยายการทำซ้ำของไตรนิวคลีโอไทด์ CAGในยีน[ 62 ] โรคฮันติงตันมีลักษณะเฉพาะคือการสูญเสียเซลล์ประสาทและกลิโอซิสอาการของโรคนี้ได้แก่ ความผิดปกติของการเคลื่อนไหว ความเสื่อมถอย ทางสติปัญญาและความผิดปกติทางพฤติกรรม[ 63 ]