อ่าน 28 นาที
ตัวรับที่เชื่อมต่อกับโปรตีนจี
ตัวรับที่เชื่อมต่อกับโปรตีน G ( GPCRs ) หรือที่รู้จักกันในชื่อ ตัวรับโดเมนทรานส์เมมเบรนเจ็ดครั้ง (seven-(pass)-transmembrane domain receptors) , ตัวรับ 7TM , ตัว รับเฮปตา เฮลิคอล...
ตัวรับที่เชื่อมต่อกับโปรตีนจี
| จีพีซีอาร์ | |||||||
|---|---|---|---|---|---|---|---|
 ตัวรับอะดรีเนอร์จิกเบต้า-2ของมนุษย์ที่ซับซ้อนกับ คา ร์ราโซลอ ลซึ่ง เป็นตัวกระตุ้นผกผัน บางส่วน [ 1 ] | |||||||
| ตัวระบุ | |||||||
| เครื่องหมาย | 7tm_1 | ||||||
| พีแฟม | พีเอฟ00001 | ||||||
| ตระกูลพีแฟม | ซีแอล0192 | ||||||
| อีโคด | 5001.1.1 | ||||||
| อินเตอร์โปร | IPR000276 | ||||||
| โปรไซต์ | PDOC00210 | ||||||
| ทีซีดีบี | 9.A.14 | ||||||
| ซูเปอร์แฟมิลี OPM | 6 | ||||||
| โปรตีน OPM | 1 กรัม | ||||||
| ซีดีดี | ซีดี14964 | ||||||
| |||||||

ตัวรับที่เชื่อมต่อกับโปรตีน G ( GPCRs ) หรือที่รู้จักกันในชื่อตัวรับโดเมนทรานส์เมมเบรนเจ็ดครั้ง (seven-(pass)-transmembrane domain receptors) , ตัวรับ 7TM , ตัว รับเฮปตา เฮลิคอล ( heptahelical receptors) , ตัวรับเซอร์เพนไทน์ ( serpentine receptors ) และ ตัวรับที่เชื่อมโยงกับ โปรตีน G ( GPLR ) เป็น กลุ่มโปรตีนที่มีความสัมพันธ์กันทางวิวัฒนาการจำนวนมากซึ่งเป็นตัวรับบนพื้นผิวเซลล์ที่ตรวจจับโมเลกุลภายนอกเซลล์และกระตุ้นการตอบสนองของเซลล์ พวกมันเชื่อมต่อกับ โปรตีน G พวกมันผ่านเยื่อหุ้มเซลล์เจ็ดครั้งในรูปแบบของลูปหกอัน[ 2 ] (ลูปภายนอกเซลล์สามอันที่ทำปฏิกิริยากับ โมเลกุล ลิแกนด์ลูปภายในเซลล์สามอันที่ทำปฏิกิริยากับโปรตีน G บริเวณภายนอกเซลล์ปลาย N และ บริเวณภายในเซลล์ปลาย C [ 2 ] ) ของกรดอะมิโนซึ่งเป็นเหตุผลที่บางครั้งเรียกพวกมันว่าตัวรับทรานส์เมมเบรนเจ็ดครั้ง[ 3 ]ลิแกนด์สามารถจับกับปลาย N-terminus และลูปภายนอกเซลล์ (เช่น ตัวรับกลูตาเมต) หรือจับกับตำแหน่งการจับภายในเฮลิกซ์ทรานส์เมมเบรน ( ตระกูลคล้าย โรดอปซิน ) ลิแกนด์ทั้งหมดจะถูกกระตุ้นด้วยสารกระตุ้นแม้ว่าจะมีการสังเกตการกระตุ้นตัวเองโดยธรรมชาติของตัวรับที่ว่างเปล่าด้วยก็ตาม[ 3 ]
ตัวรับที่เชื่อมต่อกับโปรตีน G พบได้เฉพาะในยูคาริโอตรวมถึงยีสต์และโคอาโนแฟลเจลเลต [ 4 ] ลิแกนด์ที่จับและกระตุ้นตัวรับเหล่านี้ ได้แก่ สารประกอบที่ไวต่อแสงกลิ่น ฟีโรโมนฮอร์โมนและสารสื่อประสาทขนาดของลิแกนด์เหล่านี้แตกต่างกันไปตั้งแต่โมเลกุลขนาดเล็กไปจนถึงเปปไทด์และโปรตีน ขนาดใหญ่ ตัวรับที่เชื่อมต่อกับโปรตีน G มีส่วนเกี่ยวข้องกับโรคต่างๆ มากมาย
มีเส้นทางการส่งสัญญาณหลักสองเส้นทางที่เกี่ยวข้องกับตัวรับที่เชื่อมต่อกับโปรตีน G:
- เส้นทาง การส่งสัญญาณ cAMPและ
- เส้นทางสัญญาณฟอสฟาติดิลอิโนซิทอล[ 5 ]
เมื่อลิแกนด์จับกับ GPCR จะทำให้เกิดการเปลี่ยนแปลงโครงสร้างของ GPCR ซึ่งทำให้สามารถทำหน้าที่เป็นตัวแลกเปลี่ยนนิวคลีโอไทด์กัวนีน (GEF) ได้ จากนั้น GPCR สามารถกระตุ้นโปรตีน G ที่เกี่ยวข้องได้ โดยการแลกเปลี่ยนGDPที่จับกับโปรตีน G กับGTPหน่วยย่อย α ของโปรตีน G พร้อมกับ GTP ที่จับอยู่ สามารถแยกตัวออกจากหน่วยย่อย β และ γ เพื่อส่งผลต่อโปรตีนส่งสัญญาณภายในเซลล์หรือกำหนดเป้าหมายโปรตีนที่ทำหน้าที่โดยตรง ขึ้นอยู่กับชนิดของหน่วยย่อย α ( Gαs , Gαi / o , Gαq /11 , Gα12 /13 ) [ 6 ] : 1160
GPCRs เป็นเป้าหมายยาที่สำคัญ และยาที่ได้รับการอนุมัติจากองค์การอาหารและยา (FDA) ประมาณ 34% [ 7 ] มีเป้าหมายที่สมาชิก 108 ตัวในกลุ่มนี้ ปริมาณการขายทั่วโลกของยาเหล่านี้คาดว่าจะอยู่ที่ 180 พันล้านดอลลาร์สหรัฐในปี 2018 [ 7 ]คาดว่า GPCRs เป็นเป้าหมายของยาประมาณ 50% ที่วางจำหน่ายอยู่ในปัจจุบัน ส่วนใหญ่เป็นเพราะมีส่วนเกี่ยวข้องในเส้นทางการส่งสัญญาณที่เกี่ยวข้องกับโรคต่างๆ มากมาย เช่น โรคทางจิต โรคเมตาบอลิซึมรวมถึงความผิดปกติของต่อมไร้ท่อ โรคภูมิคุ้มกันรวมถึงการติดเชื้อไวรัส โรคหัวใจและหลอดเลือด โรคอักเสบ ความผิดปกติของระบบประสาทรับความรู้สึก และมะเร็ง ความสัมพันธ์ที่ค้นพบมานานแล้วระหว่าง GPCRs กับสารภายในและภายนอกร่างกายหลายชนิด ซึ่งส่งผลให้เกิดการบรรเทาอาการปวด เป็นอีกหนึ่งสาขาการวิจัยทางเภสัชกรรมที่กำลังพัฒนาอย่างรวดเร็ว[ 3 ]
ประวัติและความสำคัญ
ด้วยการค้นพบโครงสร้างแรกของสารประกอบเชิงซ้อนระหว่างตัวรับที่เชื่อมโยงกับโปรตีนจี (GPCR) และไตรเมอร์ของโปรตีนจี (Gαβγ) ในปี 2011 ทำให้เกิดบทใหม่ของการวิจัย GPCR สำหรับการศึกษาโครงสร้างของสวิตช์ระดับโลกที่มีโปรตีนมากกว่าหนึ่งชนิดถูกตรวจสอบ การค้นพบครั้งสำคัญก่อนหน้านี้เกี่ยวข้องกับการค้นพบโครงสร้างผลึกของ GPCR ตัวแรก คือ โรดอปซิน ในปี 2000 และโครงสร้างผลึกของ GPCR ตัวแรกที่มีลิแกนด์ที่แพร่กระจายได้ (β2AR )ในปี 2007 วิธีการจัดเรียงเกลียวทรานส์เมมเบรนทั้งเจ็ดของ GPCR เป็นกลุ่มนั้นคาดเดาได้จากแบบจำลองความละเอียดต่ำของโรดอปซินจากกบจาก การศึกษา ด้วยกล้องจุลทรรศน์อิเล็กตรอน แบบแช่แข็ง ของผลึกสองมิติ โครงสร้างผลึกของโรดอปซินที่ค้นพบในอีกสามปีต่อมานั้นไม่ได้เป็นเรื่องน่าประหลาดใจ ยกเว้นการมีเกลียวไซโตพลาสมิก H8 เพิ่มเติม และตำแหน่งที่แม่นยำของลูปที่ปกคลุมบริเวณที่จับกับเรตินัล อย่างไรก็ตาม มันได้จัดเตรียมโครงสร้างพื้นฐานซึ่งหวังว่าจะเป็นแม่แบบสากลสำหรับการสร้างแบบจำลองโฮโมโลยีและการออกแบบยาสำหรับ GPCR อื่นๆ ซึ่งเป็นแนวคิดที่พิสูจน์แล้วว่ามองโลกในแง่ดีเกินไป[ 8 ]
ผลลัพธ์ที่ได้ในอีก 7 ปีต่อมานั้นน่าประหลาดใจ เพราะการตกผลึกของตัวรับ β 2 -adrenergic (β 2 AR) ที่มีลิแกนด์ที่แพร่กระจายได้เผยให้เห็นรูปร่างของด้านนอกเซลล์ของตัวรับที่แตกต่างจากโรดอปซินอย่างมาก บริเวณนี้มีความสำคัญเพราะเป็นบริเวณที่รับผิดชอบในการจับลิแกนด์และเป็นเป้าหมายของยาหลายชนิด ยิ่งไปกว่านั้น บริเวณที่จับลิแกนด์นั้นกว้างขวางกว่าในโครงสร้างของโรดอปซินมากและเปิดออกสู่ภายนอก ในตัวรับอื่นๆ ที่ตกผลึกในเวลาต่อมาไม่นาน บริเวณที่จับลิแกนด์นั้นเข้าถึงได้ง่ายยิ่งขึ้น โครงสร้างใหม่ที่เสริมด้วยการตรวจสอบทางชีวเคมีได้เปิดเผยกลไกการทำงานของสวิตช์โมเลกุลซึ่งปรับเปลี่ยนโครงสร้างของตัวรับ ทำให้เกิดสถานะการกระตุ้นสำหรับตัวกระตุ้นหรือสถานะการปิดใช้งานอย่างสมบูรณ์หรือบางส่วนสำหรับตัวยับยั้งแบบผกผัน[ 3 ]
รางวัลโนเบลสาขาเคมีประจำปี 2012 มอบให้แก่Brian KobilkaและRobert Lefkowitzสำหรับผลงานของพวกเขาซึ่ง "มีความสำคัญอย่างยิ่งต่อความเข้าใจว่าตัวรับที่เชื่อมโยงกับโปรตีน G ทำงานอย่างไร" [ 9 ]มีรางวัลโนเบลอย่างน้อยเจ็ดรางวัลที่มอบให้แก่แง่มุมต่างๆ ของการส่งสัญญาณผ่านโปรตีน G ณ ปี 2012 ยาที่ขายดีที่สุด 10 อันดับแรกของโลก 2 ชนิด ( Advair DiskusและAbilify ) ออกฤทธิ์โดยการกำหนดเป้าหมายที่ตัวรับที่เชื่อมโยงกับโปรตีน G [ 10 ]
การจำแนกประเภท

ขนาดที่แน่นอนของซูเปอร์แฟมิลี GPCR ยังไม่เป็นที่ทราบแน่ชัด แต่ มีการคาดการณ์ว่า ยีนของมนุษย์ อย่างน้อย 831 ยีน (หรือประมาณ 4% ของจีโนมที่เข้ารหัสโปรตีน ทั้งหมด ) เข้ารหัสสำหรับซูเปอร์แฟมิลีนี้จากการวิเคราะห์ลำดับจีโนม[ 11 ] [ 12 ]แม้ว่าจะ มีการเสนอแผนการจำแนกประเภทมากมาย แต่ซูเปอร์แฟมิลีนี้ถูกแบ่งออกเป็นสามคลาสหลัก (A, B และ C) โดยไม่มี ความคล้ายคลึงกันของลำดับที่ตรวจพบได้ระหว่างคลาส[ 13 ]
กลุ่มที่ใหญ่ที่สุดคือกลุ่ม A ซึ่งคิดเป็นเกือบ 85% ของยีน GPCR ในบรรดา GPCR กลุ่ม A มากกว่าครึ่งหนึ่งคาดว่าจะเข้ารหัสตัวรับกลิ่นในขณะที่ตัวรับที่เหลือจะถูกจับโดยสารประกอบภายในร่างกาย ที่รู้จัก หรือจัดอยู่ใน กลุ่ม ตัวรับกำพร้าแม้ว่าจะไม่มีความคล้ายคลึงกันของลำดับระหว่างกลุ่ม แต่ GPCR ทั้งหมดมีโครงสร้างและกลไกการส่งสัญญาณ ที่เหมือนกัน กลุ่มโรดอปซิน A ขนาดใหญ่มากได้ถูกแบ่งย่อยออกเป็น 19 กลุ่มย่อย ( A1-A19 ) [ 14 ]
ตามระบบ AF แบบคลาสสิก GPCR สามารถจัดกลุ่มได้เป็นหกคลาสตามความเหมือนกันของลำดับและความคล้ายคลึงกันของฟังก์ชัน: [ 15 ] [ 16 ] [ 17 ] [ 18 ]
- คลาส A (หรือ 1) ( คล้ายโรดอปซิน )
- คลาส B (หรือ 2) ( ตระกูลตัวรับซีเครติน )
- คลาส C (หรือ 3) ( กลูตาเมตเมตาโบโทรปิก /ฟีโรโมน)
- คลาส D (หรือ 4) ( ตัวรับฟีโรโมนผสมพันธุ์ของเชื้อรา )
- คลาส E (หรือ 5) ( ตัวรับไซคลิก AMP )
- คลาส F (หรือ 6) ( หยิกฟู / เรียบ )
เมื่อไม่นานมานี้ มีการเสนอระบบการจำแนกประเภททางเลือกที่เรียกว่าGRAFS ( Glutamate , Rhodopsin , Adhesion , Frizzled / Taste2 , Secretin ) สำหรับ GPCR ของสัตว์มีกระดูกสันหลัง [ 11 ] ซึ่งสอดคล้องกับคลาสคลาสสิก C, A, B2, F และ B [ 19 ]
การศึกษาเบื้องต้นที่อิงตามลำดับดีเอ็นเอที่มีอยู่ชี้ให้เห็นว่าจีโนมของมนุษย์เข้ารหัสตัวรับที่เชื่อมโยงกับโปรตีน G ประมาณ 750 ตัว[ 20 ]ซึ่งประมาณ 350 ตัวตรวจจับฮอร์โมน ปัจจัยการเจริญเติบโต และลิแกนด์ภายในร่างกายอื่นๆ ประมาณ 150 ตัวของ GPCR ที่พบในจีโนมของมนุษย์มีหน้าที่ที่ไม่ทราบแน่ชัด
เซิร์ฟเวอร์เว็บบางส่วน[ 21 ]และวิธีการทำนายทางชีวสารสนเทศ[ 22 ] [ 23 ]ได้ถูกนำมาใช้เพื่อทำนายการจำแนกประเภทของ GPCR ตามลำดับกรดอะมิโนเพียงอย่างเดียว โดยใช้แนวทาง องค์ประกอบกรดอะมิโนเทียม
บทบาททางสรีรวิทยา
GPCR มีส่วนเกี่ยวข้องในกระบวนการทางสรีรวิทยาที่หลากหลาย ตัวอย่างบทบาททางสรีรวิทยาของ GPCR ได้แก่:
- ประสาทรับภาพ: ออปซินใช้ ปฏิกิริยา โฟโตไอโซเมอไรเซชันเพื่อแปลงรังสีแม่เหล็กไฟฟ้าเป็นสัญญาณเซลล์ ตัวอย่างเช่น โรดอปซินใช้การแปลง11-cis -retinalเป็นall-trans -retinalเพื่อจุดประสงค์นี้[ 24 ]
- ประสาทรับรส (รสชาติ): ตัวรับ GPCR ในเซลล์รับรสทำหน้าที่ควบคุมการปล่อยสารกัสต์ดูซิน (gustducin)เพื่อตอบสนองต่อสารที่มีรสขม รสอูมามิ และรสหวาน
- ประสาทรับกลิ่น: ตัวรับในเยื่อบุผิวรับกลิ่นจะจับกับสารให้กลิ่น (ตัวรับกลิ่น) และฟีโรโมน (ตัวรับโวเมอโรนาซัล)
- การควบคุมพฤติกรรมและอารมณ์: ตัวรับที่เชื่อมต่อกับโปรตีน G ในสมอง ของสัตว์ เลี้ยงลูกด้วยนม จะจับกับ สารสื่อประสาทหลายชนิดรวมถึงเซโรโทนิน โดปามีนฮิสตามีน นอร์อะดรีนาลีนและGABA ข้อยกเว้นที่สำคัญ ได้แก่ตัวรับGABA A [ 25 ]ตัวรับเซโรโทนิน5-HT 3 [ 26 ]และตัวรับกลูตาเมต( NMDAR , AMPAR และตัวรับไคเนต ) [ 27 ]ซึ่งเป็นช่องไอออน
- การควบคุม กิจกรรม ของระบบภูมิคุ้มกันและการอักเสบ : ตัวรับ เคโมไคน์จะจับกับลิแกนด์ที่ทำหน้าที่เป็นสื่อกลางในการสื่อสารระหว่างเซลล์ของระบบภูมิคุ้มกัน ตัวรับเช่นตัวรับฮิสตามีนจะจับกับสารสื่อกลางการอักเสบและกระตุ้นเซลล์เป้าหมายในการตอบสนองต่อการอักเสบ GPCR ยังมีส่วนเกี่ยวข้องกับการปรับเปลี่ยนภูมิคุ้มกัน เช่น การควบคุมการเหนี่ยวนำของอินเตอร์ลิวคิน[ 28 ]หรือการยับยั้ง การตอบสนองทางภูมิคุ้มกันที่เกิดจาก TLRจากเซลล์ T [ 29 ]
- การส่งสัญญาณ ของระบบประสาทอัตโนมัติ: ทั้ง ระบบประสาทซิ มพาเทติกและพาราซิมพาเทติกถูกควบคุมโดยเส้นทาง GPCR ตัวรับที่เกี่ยวข้อง ได้แก่ตัวรับอะดรีเนอร์จิก ( α1 , α2 , β1 , β2 และ β3ซึ่งβ2 โดดเด่นตรงที่เป็น GPCR ตัวแรกที่มีโครงสร้างผลึกที่ได้รับการกำหนด) สำหรับระบบประสาทซิมพาเทติก และตัวรับอะเซทิลโคลีนมัสคารินิกสำหรับระบบประสาทพาราซิมพาเทติก สารที่จับกับตัวรับอะดรีเนอร์จิกเป็นที่ทราบกันดีว่าสามารถปรับความดันโลหิตและอัตราการเต้นของหัวใจได้ ในขณะที่สารต้านมัสคารินิกทำให้เกิดอาการต่างๆ มากมาย เช่น ม่านตาขยาย ปัสสาวะค้าง ท้องผูก และปากแห้ง ซึ่งเกี่ยวข้องโดยตรงกับการยับยั้งระบบประสาทพาราซิมพาเทติก[ 30 ] [ 31 ]
- การรับรู้ความหนาแน่นของเซลล์: บทบาทใหม่ของ GPCR ในการควบคุมการรับรู้ความหนาแน่นของเซลล์
- การปรับสมดุลของร่างกาย (เช่น สมดุลน้ำ) [ 32 ]
- เกี่ยวข้องกับการเจริญเติบโตและ การ แพร่กระจายของเนื้องอก บางชนิด [ 33 ]
- ใช้ในระบบต่อมไร้ท่อสำหรับฮอร์โมนอนุพันธ์เปปไทด์และกรดอะมิโนที่จับกับ GCPR บนเยื่อหุ้มเซลล์ของเซลล์เป้าหมาย การจับกันนี้จะกระตุ้น cAMP ซึ่งจะไปกระตุ้นไคเนสหลายตัว ทำให้เกิดการตอบสนองของเซลล์ เช่น การถอดรหัสพันธุกรรม
โครงสร้างของตัวรับ
GPCRs เป็นโปรตีนเมมเบรนแบบบูรณาการที่มีโดเมนที่ทะลุผ่านเมมเบรนเจ็ดโดเมนหรือเกลียวทรานส์เมมเบรน [ 34 ] [ 35 ] ส่วนนอกเซลล์ของตัวรับสามารถถูกไกลโคซิเลต ได้ ลูปนอกเซลล์เหล่านี้ยังมี สารตกค้าง ซิสเทอีน ที่อนุรักษ์ไว้อย่างสูงสองตัว ที่สร้างพันธะไดซัลไฟด์เพื่อทำให้โครงสร้างของตัวรับมีเสถียรภาพ โปรตีนเกลียวทรานส์เมมเบรนเจ็ดโดเมนบางชนิด ( แชนเนลโรดอปซิน ) ที่คล้ายกับ GPCRs อาจมีช่องไอออนอยู่ภายในโปรตีน
ในปี 2000 โครงสร้างผลึกแรกของ GPCR ของสัตว์เลี้ยงลูกด้วยนม ซึ่งก็คือโรดอปซิน ของวัว ( 1F88 ) ได้รับการแก้ไข[ 36 ]ในปี 2007 โครงสร้างแรกของ GPCR ของมนุษย์ได้รับการแก้ไข[ 37 ] [ 1 ] [ 38 ] โครงสร้าง GPCR ของตัวรับ β 2 -adrenergicของมนุษย์นี้พิสูจน์แล้วว่ามีความคล้ายคลึงกับโรดอปซินของวัวอย่างมาก โครงสร้างของ GPCR ที่ถูกกระตุ้นหรือจับกับตัวกระตุ้นก็ได้รับการกำหนดแล้วเช่นกัน[ 39 ] [ 40 ] [ 41 ] [ 42 ]โครงสร้างเหล่านี้แสดงให้เห็นว่าการจับของลิแกนด์ที่ด้านนอกเซลล์ของตัวรับนำไปสู่การเปลี่ยนแปลงโครงสร้างที่ด้านไซโตพลาสมิกของตัวรับ การเปลี่ยนแปลงที่ใหญ่ที่สุดคือการเคลื่อนที่ออกไปด้านนอกของส่วนไซโตพลาสมิกของเกลียวทรานส์เมมเบรนที่ 5 และ 6 (TM5 และ TM6) โครงสร้างของตัวรับเบต้า-2 อะดรีเนอร์จิกที่ถูกกระตุ้นในเชิงซ้อนกับ G sยืนยันว่า Gα จับกับโพรงที่สร้างขึ้นโดยการเคลื่อนไหวนี้[ 43 ]
GPCRs มีโครงสร้างคล้ายกับโปรตีนอื่นๆ ที่มีโดเมนทรานส์เมมเบรน 7 โดเมน เช่นโรดอปซินของจุลินทรีย์และตัวรับอะดิโพเนกติน 1 และ 2 ( ADIPOR1และADIPOR2 ) อย่างไรก็ตาม ตัวรับและช่อง 7TMH (เกลียวทรานส์เมมเบรน 7 อัน) เหล่านี้ไม่ได้เชื่อมโยงกับโปรตีน Gนอกจากนี้ ADIPOR1 และ ADIPOR2 ยังมีทิศทางตรงกันข้ามกับ GPCRs ในเยื่อหุ้มเซลล์ (กล่าวคือ GPCRs มักจะมีปลาย N อยู่นอกเซลล์และปลาย C อยู่ในไซโตพลาสซึม ในขณะที่ ADIPORs จะกลับด้าน) [ 44 ]
ความสัมพันธ์ระหว่างโครงสร้างและหน้าที่
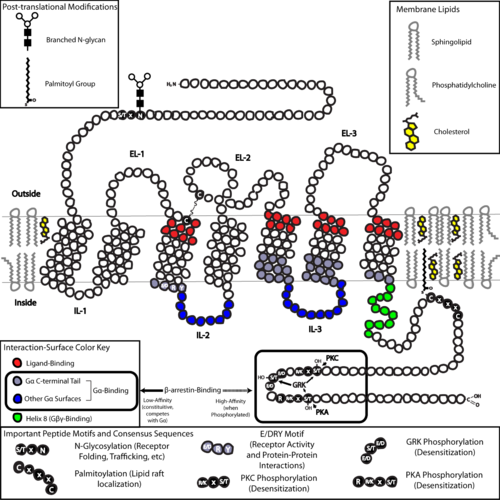
ในแง่ของโครงสร้าง GPCR มีลักษณะเฉพาะคือมีปลาย N-terminus อยู่นอกเซลล์ ตามด้วย เกลียว อัลฟาแบบทรานส์เมมเบ รนเจ็ดเกลียว (7-TM) (TM-1 ถึง TM-7) ที่เชื่อมต่อกันด้วยลูปภายในเซลล์สามลูป (IL-1 ถึง IL-3) และลูปภายนอกเซลล์สามลูป (EL-1 ถึง EL-3) และสุดท้ายคือปลาย C-terminus ภายในเซลล์ GPCR จัดเรียงตัวเองเป็นโครงสร้างสามมิติที่มีลักษณะคล้ายถัง โดยเกลียวทรานส์เมมเบรนทั้งเจ็ดเกลียวจะสร้างโพรงภายในเยื่อหุ้มเซลล์ซึ่งทำหน้าที่เป็น โดเมนจับกับ ลิแกนด์ซึ่งมักถูกปกคลุมด้วย EL-2 อย่างไรก็ตาม ลิแกนด์อาจจับกับส่วนอื่นได้เช่นกัน ดังเช่นในกรณีของลิแกนด์ที่มีขนาดใหญ่กว่า (เช่นโปรตีนหรือเปปไทด์ ขนาดใหญ่) ซึ่งจะโต้ตอบกับลูปภายนอกเซลล์ หรือดังที่แสดงโดย ตัวรับกลูตาเมตแบบเมตาโบโทรปิกคลาส C (mGluRs) จะโต้ตอบกับส่วนหาง N-terminus GPCR คลาส C มีลักษณะเด่นคือหาง N-terminal ขนาดใหญ่ ซึ่งประกอบด้วยโดเมนจับลิแกนด์ด้วย เมื่อกลูตาเมตจับกับ mGluR หาง N-terminal จะเกิดการเปลี่ยนแปลงโครงสร้าง ทำให้เกิดการโต้ตอบกับสารตกค้างของลูปภายนอกเซลล์และโดเมน TM ผลสุดท้ายของ การกระตุ้นด้วย ตัวกระตุ้น ทั้งสามประเภท คือการเปลี่ยนแปลงทิศทางสัมพัทธ์ของเกลียว TM (เปรียบเสมือนการบิดตัว) นำไปสู่พื้นผิวภายในเซลล์ที่กว้างขึ้นและการ "เปิดเผย" สารตกค้างของเกลียวภายในเซลล์และโดเมน TM ที่สำคัญต่อการทำงานของการส่งสัญญาณ (เช่น การจับคู่โปรตีน G) ตัวกระตุ้นผกผันและตัวต้านอาจจับกับตำแหน่งต่างๆ ได้หลายตำแหน่ง แต่ผลสุดท้ายจะต้องเป็นการป้องกันการเปลี่ยนทิศทางของเกลียว TM นี้[ 3 ]
โครงสร้างของปลาย N- และ C-terminal ของ GPCR อาจมีหน้าที่สำคัญนอกเหนือจากการจับกับลิแกนด์ ตัวอย่างเช่น ปลาย C-terminus ของตัวรับมัสคารินิก M3 นั้นเพียงพอแล้ว และโดเมนโพลีเบสิกหกกรดอะมิโน (KKKRRK) ในปลาย C-terminus นั้นจำเป็นสำหรับการประกอบล่วงหน้ากับโปรตีนGq [ 45 ]โดยเฉพาะอย่างยิ่ง ปลาย C-terminus มักมี สารตกค้าง ซีรีน (Ser) หรือทรีโอนีน (Thr) ซึ่งเมื่อถูกฟอสโฟรีเลตจะเพิ่มความสัมพันธ์ของพื้นผิวภายในเซลล์สำหรับการจับกับโปรตีนโครงสร้างที่เรียกว่า β- arrestins (β-arr) [ 46 ]เมื่อจับแล้ว β-arrestins จะป้องกันการจับคู่ ของโปรตีน G ในเชิงสเตอริกและอาจดึงดูดโปรตีนอื่นๆ ทำให้เกิดคอมเพล็กซ์การส่งสัญญาณที่เกี่ยวข้องกับการกระตุ้นวิถีการทำงานของเอนไซม์ extracellular-signal regulated kinase ( ERK ) หรือการนำตัวรับเข้าสู่ เซลล์ (internalization) เนื่องจากการฟอสโฟรีเลชันของสารตกค้าง Ser และ Thr เหล่านี้มักเกิดขึ้นอันเป็นผลมาจากการกระตุ้น GPCR การแยกตัวของโปรตีน G ที่เกิดจาก β-arr และการนำ GPCR เข้าสู่เซลล์จึงเป็นกลไกสำคัญของ การลด ความไว[ 47 ]นอกจากนี้ "เมกะคอมเพล็กซ์" ที่ถูกนำเข้าสู่เซลล์ซึ่งประกอบด้วย GPCR เดี่ยว β-arr (ในรูปแบบหาง) [ 48 ] [ 49 ]และโปรตีน G แบบเฮเทอโรไตรเมอริกก็มีอยู่และอาจอธิบายถึงการส่งสัญญาณโปรตีนจากเอนโดโซมได้[ 50 ] [ 51 ]
โครงสร้างร่วมสุดท้ายใน GPCRs คือการพาลมิโตอิเลชันของตำแหน่งหนึ่งหรือมากกว่าของหางปลาย C หรือลูปภายในเซลล์ พาลมิโตอิเลชันคือการดัดแปลงโควาเลนต์ของ หมู่ ซิสเทอีน (Cys) ผ่านการเพิ่มหมู่แอซิล ที่ไม่ชอบน้ำ และมีผลในการกำหนดเป้าหมายตัวรับไปยังไมโครโดเมนที่อุดมไปด้วยคอเลสเตอรอลและ สฟิง โกลิปิดของเยื่อหุ้มพลาสมาที่เรียกว่าแพลิพิด [ 52 ] เนื่องจากโมเลกุลตัวส่งสัญญาณและตัวกระตุ้นปลายทางจำนวนมากของ GPCRs (รวมถึงโมเลกุลที่เกี่ยวข้องกับ เส้นทาง ป้อนกลับเชิงลบ ) ก็ถูกกำหนดเป้าหมายไปยังแพลิพิดเช่นกัน จึงมีผลในการอำนวยความสะดวกในการส่งสัญญาณของตัวรับอย่างรวดเร็ว[ 52 ]
GPCR ตอบสนองต่อสัญญาณภายนอกเซลล์ที่ส่งผ่านโดยสารกระตุ้นที่หลากหลายมาก ตั้งแต่โปรตีนไปจนถึงสารชีวภาพและโปรตอนแต่ทั้งหมดจะส่งสัญญาณนี้ผ่านกลไกการเชื่อมต่อกับโปรตีน G ซึ่งเป็นไปได้ด้วย โดเมนตัวแลกเปลี่ยนกัวนีน- นิวคลีโอไทด์ ( GEF ) ที่เกิดจากการรวมกันของ IL-2 และ IL-3 ร่วมกับสารตกค้างที่อยู่ติดกันของเกลียว TM ที่เกี่ยวข้อง
กลไก

ตัวรับที่เชื่อมต่อกับโปรตีน G จะถูกกระตุ้นด้วยสัญญาณภายนอกในรูปแบบของลิแกนด์หรือตัวกลางส่งสัญญาณอื่น ๆ ซึ่งจะสร้างการเปลี่ยนแปลงโครงสร้างในตัวรับ ส่งผลให้โปรตีน G ถูกกระตุ้น ผลกระทบเพิ่มเติมขึ้นอยู่กับชนิดของโปรตีน G โปรตีน G จะถูกยับยั้งในภายหลังโดยโปรตีนกระตุ้น GTPase ซึ่งรู้จักกันในชื่อโปรตีน RGS [ 54 ]
การจับตัวของลิแกนด์
GPCR ประกอบด้วยตัวรับอย่างน้อยหนึ่งตัวสำหรับลิแกนด์ต่อไปนี้: ตัวกลางส่งสัญญาณประสาทสัมผัส (เช่นโมเลกุลกระตุ้น แสงและ กลิ่น ); อะดีโนซีน , บอมเบซิน , แบรดิกินิน , เอนโดเทลิน, กรดแกมมา อะมิโนบิวทิริก ( GABA ), ปัจจัยการเจริญเติบโตของเซลล์ตับ ( HGF ), เมลาโนคอร์ทิน , นิว โรเปปไทด์ Y , เปปไทด์โอปิออยด์, ออปซิน, โซมาโตสแตติน , GH , ทาคิคินิน , สมาชิกใน กลุ่ม เปปไทด์ในลำไส้ที่ออกฤทธิ์ต่อหลอดเลือดและวาโซเพรสซิน ; อะมีนชีวภาพ (เช่น โดปามีน , เอ พิเนฟริน , น อร์เอพิเนฟริน , ฮิสตามี น , เซ โรโทนิ นและเมลาโทนิน ); ก ลูตาเมต ( ผล เมตาโบโทรปิก ); กลูคากอน ; อะเซทิลโคลีน ( ผล มัสคารินิก ); เคโมไคน์ ; สารสื่อกลางไขมัน ที่ก่อให้ เกิดการอักเสบ (เช่นโปรสตาแกลน ดิน โปรสตาโนอิดปัจจัยกระตุ้นเกล็ดเลือดและลิวโคไตรอีน ); ฮอร์โมนเปปไทด์ (เช่นแคลซิโทนิน C5a อนาฟิลาทอกซิน ฮอร์โมนกระตุ้นฟอลลิเคิล [FSH] ฮอร์โมนปล่อยโกนาโดโทรปิน [GnRH] นิวโรคินินฮอร์โมนปล่อยไทรอยด์โทร ปิน [TRH] และออกซิโทซิน ); และเอนโดแคนนาบินอยด์
GPCR ที่ทำหน้าที่เป็นตัวรับสำหรับสิ่งเร้าที่ยังไม่ได้รับการระบุเรียกว่าตัวรับกำพร้า[ 55 ]
อย่างไรก็ตาม เมื่อเปรียบเทียบกับตัวรับประเภทอื่น ๆ ที่ได้รับการศึกษา ซึ่งลิแกนด์จะจับกับเยื่อหุ้มเซลล์ภายนอก ลิแกนด์ของ GPCR มักจะจับกับโดเมนทรานส์เมมเบรน อย่างไรก็ตามตัวรับที่ถูกกระตุ้นด้วยโปรตีเอสจะถูกกระตุ้นโดยการตัดส่วนหนึ่งของโดเมนภายนอกเซลล์[ 56 ]
การเปลี่ยนแปลงโครงสร้าง

การส่งสัญญาณผ่านเยื่อหุ้มเซลล์โดยตัวรับยังไม่เป็นที่เข้าใจอย่างสมบูรณ์ เป็นที่ทราบกันว่าในสภาวะที่ไม่ทำงาน GPCR จะจับกับ คอมเพล็กซ์ โปรตีน G แบบเฮเทอโรไตร เม อริก การจับของตัวกระตุ้นกับ GPCR ส่งผลให้เกิดการเปลี่ยนแปลงโครงสร้างในตัวรับ ซึ่งจะถูกส่งต่อไปยังหน่วยย่อย Gα ที่จับอยู่ของโปรตีน G แบบเฮเทอโรไตรเมอ ริกผ่าน พลวัตของโดเมนโปรตีน หน่วยย่อย Gα ที่ถูกกระตุ้นจะแลกเปลี่ยนGTPแทนที่GDPซึ่งจะกระตุ้นให้หน่วยย่อย Gα แยกตัวออกจากไดเมอร์ Gβγ และจากตัวรับ หน่วยย่อย Gα และ Gβγ ที่แยกตัวออกมาจะทำปฏิกิริยากับโปรตีนภายในเซลล์อื่นๆ เพื่อดำเนินการส่งสัญญาณต่อไป ในขณะที่ GPCR ที่เป็นอิสระสามารถจับกับโปรตีน G แบบเฮเทอโรไตรเมอริกอีกตัวเพื่อสร้างคอมเพล็กซ์ใหม่ที่พร้อมจะเริ่มต้นการส่งสัญญาณรอบใหม่[ 57 ]
เชื่อกันว่าโมเลกุลตัวรับมีอยู่ในสมดุล เชิงโครงสร้าง ระหว่างสถานะทางชีวฟิสิกส์ที่ทำงานและไม่ทำงาน[ 58 ]การจับของลิแกนด์กับตัวรับอาจทำให้สมดุลเปลี่ยนไปทางสถานะตัวรับที่ทำงาน ลิแกนด์มีสามประเภท ได้แก่ อะโกนิสต์ ซึ่งเป็นลิแกนด์ที่เปลี่ยนสมดุลไปในทิศทางของสถานะที่ทำงาน อินเวอร์สอะโกนิสต์ซึ่งเป็นลิแกนด์ที่เปลี่ยนสมดุลไปในทิศทางของสถานะที่ไม่ทำงาน และนีโอแอนตาโกนิสต์ ซึ่งเป็นลิแกนด์ที่ไม่ส่งผลต่อสมดุล ยังไม่เป็นที่ทราบแน่ชัดว่าสถานะที่ทำงานและไม่ทำงานแตกต่างกันอย่างไร
วงจรการกระตุ้น/ยับยั้งการทำงานของโปรตีน G

เมื่อตัวรับไม่ทำงาน โดเมนGEFอาจจับกับซับยูนิต α ที่ไม่ทำงานเช่นกันของโปรตีน G แบบเฮเทอโรไตรเมอริกโปรตีน G เหล่านี้เป็นไตรเมอร์ของซับยูนิต α, β และ γ (รู้จักกันในชื่อ Gα, Gβ และ Gγ ตามลำดับ) ซึ่งจะไม่ทำงานเมื่อจับกับกัวโนซีนไดฟอสเฟต (GDP) (หรืออีกทางหนึ่งคือไม่มีนิวคลีโอไทด์กัวนีน) แต่จะทำงานเมื่อจับกับกัวโนซีนไตรฟอสเฟต (GTP) เมื่อตัวรับถูกกระตุ้น โดเมน GEF จะกระตุ้นโปรตีน G แบบอัลโลสเตอริก โดยอำนวย ความสะดวกในการแลกเปลี่ยนโมเลกุล GDP กับ GTP ที่ซับยูนิต α ของโปรตีน G เซลล์รักษาสัดส่วน GTP:GDP ในไซโตพลาสมิกไว้ที่ 10:1 ดังนั้นจึงมั่นใจได้ว่ามีการแลกเปลี่ยนเป็น GTP ในขั้นตอนนี้ หน่วยย่อยของโปรตีน G จะแยกตัวออกจากตัวรับ รวมถึงแยกตัวออกจากกันเองด้วย ทำให้เกิดโมโนเมอร์ Gα-GTP และไดเมอร์ Gβγ ที่จับกันอย่างแน่นหนา ซึ่งขณะนี้พร้อมที่จะปรับเปลี่ยนกิจกรรมของโปรตีนภายในเซลล์อื่นๆ อย่างไรก็ตาม ขอบเขตการแพร่กระจาย ของพวกมันนั้น มีจำกัด เนื่องจากการเติมหมู่ปาล์มิโตอิลให้กับ Gα และการมีอยู่ของ หมู่ ไอโซพรีนอยด์ที่ถูกเติมเข้าไปที่ปลาย C ของ Gγ ด้วยพันธะโควาเลนต์
เนื่องจาก Gα มี ความสามารถ ในการไฮโดรไลซิส GTP→GDP อย่างช้าๆ รูปแบบที่ไม่ทำงานของซับยูนิต α (Gα-GDP) จึงถูกสร้างขึ้นใหม่ในที่สุด ทำให้สามารถรวมตัวกับไดเมอร์ Gβγ เพื่อสร้างโปรตีน G ที่ "พักตัว" ซึ่งสามารถจับกับ GPCR อีกครั้งและรอการกระตุ้น อัตราการไฮโดรไลซิส GTP มักจะเร่งขึ้นเนื่องจากการทำงานของโปรตีนปรับเปลี่ยนแบบอัลโลสเตอริกอีกตระกูลหนึ่งที่เรียกว่าตัวควบคุมการส่งสัญญาณของโปรตีน G หรือโปรตีน RGS ซึ่งเป็น โปรตีนกระตุ้น GTPaseหรือ GAP ชนิดหนึ่ง อันที่จริง โปรตีน ตัวรับผล หลักหลายตัว (เช่นอะดีนิเลตไซเคลส ) ที่ถูกกระตุ้น/ยับยั้งเมื่อมีปฏิสัมพันธ์กับ Gα-GTP ก็มีกิจกรรม GAP ด้วย ดังนั้น แม้ในระยะเริ่มต้นของกระบวนการนี้ การส่งสัญญาณที่เริ่มต้นโดย GPCR ก็มีความสามารถในการยุติตัวเองได้
ครอสทอล์ก

สัญญาณปลายทางของ GPCR แสดงให้เห็นว่าอาจมีปฏิสัมพันธ์กับ สัญญาณ อินทิกรินเช่นFAK [ 59 ] การส่งสัญญาณอินทิกรินจะทำให้ FAK เกิดการฟอสโฟรีเลต ซึ่งสามารถลดกิจกรรม Gαs ของ GPCR ได้
การส่งสัญญาณ

หากตัวรับที่อยู่ในสถานะทำงานพบกับโปรตีน Gมันอาจจะกระตุ้นโปรตีน G ได้ หลักฐานบางอย่างชี้ให้เห็นว่าตัวรับและโปรตีน G นั้นจับคู่กันไว้ล่วงหน้าแล้ว[ 45 ]ตัวอย่างเช่น การจับกันของโปรตีน G กับตัวรับจะส่งผลต่อความสัมพันธ์ของตัวรับกับลิแกนด์ โปรตีน G ที่ถูกกระตุ้นจะจับกับ GTP
การส่งสัญญาณต่อไปนั้นขึ้นอยู่กับชนิดของโปรตีน G เอนไซม์อะดีนิเลตไซเคลสเป็นตัวอย่างของโปรตีนในเซลล์ที่สามารถถูกควบคุมโดยโปรตีน G ในกรณีนี้คือโปรตีน G s การทำงานของอะดีนิเลตไซเคลสจะถูกกระตุ้นเมื่อมันจับกับหน่วยย่อยของโปรตีน G ที่ถูกกระตุ้น การกระตุ้นของอะดีนิเลตไซเคลสจะสิ้นสุดลงเมื่อโปรตีน G กลับสู่สถานะที่จับกับ GDP
อะดีนิเลตไซเคลส (ซึ่งในมนุษย์มี 9 รูปแบบที่ยึดติดกับเยื่อหุ้มเซลล์และ 1 รูปแบบที่อยู่ในไซโตพลาสซึม) อาจถูกกระตุ้นหรือยับยั้งด้วยวิธีอื่น ๆ (เช่น การจับกับ Ca2+/ แคลโมดูลิน ) ซึ่งสามารถปรับเปลี่ยนกิจกรรมของเอนไซม์เหล่านี้ในลักษณะเสริมกันหรือทำงานร่วมกันกับโปรตีน G ได้
เส้นทางการส่งสัญญาณที่ถูกกระตุ้นผ่าน GPCR นั้นถูกจำกัดด้วยลำดับกรดอะมิโนและโครงสร้างสามมิติของ GPCR เอง แต่สุดท้ายแล้วจะถูกกำหนดโดยโครงสร้าง เฉพาะ ที่เสถียรโดยลิแกนด์ เฉพาะ รวมถึงความพร้อมของ โมเลกุล ตัวส่งสัญญาณปัจจุบัน GPCR ถือว่าใช้ตัวส่งสัญญาณหลักสองประเภท ได้แก่โปรตีน Gและβ-arrestinเนื่องจาก β-arrestin มีความสัมพันธ์ สูง เฉพาะกับ รูปแบบ ฟอสโฟรีเลตของ GPCR ส่วนใหญ่ (ดูด้านบนหรือด้านล่าง) การส่งสัญญาณส่วนใหญ่จึงขึ้นอยู่กับการกระตุ้นโปรตีน G อย่างไรก็ตาม ความเป็นไปได้ของการมีปฏิสัมพันธ์ก็อนุญาตให้เกิดการส่งสัญญาณที่ไม่ขึ้นกับโปรตีน G ได้เช่นกัน
การส่งสัญญาณที่ขึ้นอยู่กับโปรตีนจี
มีเส้นทางการส่งสัญญาณหลักสามเส้นทางที่ควบคุมโดยโปรตีน G ซึ่งควบคุมโดย โปรตีน Gα สี่กลุ่มย่อยที่แตกต่างกันโดยความคล้ายคลึงกันของลำดับ ( Gαs , Gαi /o , Gαq /11และGα12 /13 ) โปรตีน G แต่ละกลุ่มย่อยประกอบด้วยโปรตีนหลายชนิด ซึ่งแต่ละชนิดเป็นผลผลิตจากยีน หลายตัว หรือรูปแบบการตัดต่อที่อาจทำให้มีคุณสมบัติการส่งสัญญาณที่แตกต่างกันตั้งแต่เล็กน้อยไปจนถึงชัดเจน แต่โดยทั่วไปแล้วดูเหมือนว่าจะจัดกลุ่มได้อย่างเหมาะสมเป็นสี่กลุ่ม เนื่องจากคุณสมบัติการส่งสัญญาณของชุด βγ ที่เป็นไปได้ต่างๆ ดูเหมือนจะไม่แตกต่างกันอย่างมาก กลุ่มเหล่านี้จึงถูกกำหนดตามไอโซฟอร์มของหน่วยย่อย α [ 6 ] : 1163
แม้ว่า GPCR ส่วนใหญ่จะสามารถกระตุ้น Gα-subtype ได้มากกว่าหนึ่งชนิด แต่ก็ยังแสดงความชอบต่อ subtype หนึ่งมากกว่าอีก subtype หนึ่ง เมื่อ subtype ที่ถูกกระตุ้นขึ้นอยู่กับลิแกนด์ที่จับกับ GPCR นั้น เรียกว่าการเลือกสรรเชิงฟังก์ชัน (หรือที่รู้จักกันในชื่อ การขนส่งที่กำกับโดยตัวกระตุ้น หรือ การกระตุ้นที่จำเพาะต่อโครงสร้าง) อย่างไรก็ตาม การจับของตัวกระตุ้นเฉพาะตัวใดตัวหนึ่งอาจเริ่มต้นการกระตุ้นของ G-protein ที่แตกต่างกันหลายชนิดได้ เนื่องจากตัวกระตุ้นนั้นอาจสามารถทำให้โครงสร้างมากกว่าหนึ่งโครงสร้างของ โดเมน GEF ของ GPCR มีเสถียรภาพได้ แม้กระทั่งในระหว่างการปฏิสัมพันธ์เพียงครั้งเดียว นอกจากนี้ โครงสร้างที่กระตุ้น Gα isoform หนึ่งได้ดีกว่า อาจกระตุ้นอีก isoform หนึ่งได้ หาก isoform ที่ชอบมีน้อยกว่า ยิ่งไปกว่านั้น กลไก ป้อน กลับ อาจส่งผลให้เกิดการดัดแปลงตัวรับ (เช่น การฟอสฟอริเลชั่น) ที่เปลี่ยนแปลงความชอบของ G-protein ได้ โดยไม่คำนึงถึงความแตกต่างปลีกย่อยต่างๆ เหล่านี้ โดยทั่วไปแล้วคู่พันธะที่ GPCR นิยมจับคู่จะถูกกำหนดตามโปรตีน G ที่ถูกกระตุ้นอย่างชัดเจนที่สุดโดย ลิแกนด์ ภายในร่างกายภายใต้สภาวะ ทางสรีรวิทยาหรือการทดลอง ส่วนใหญ่
การส่งสัญญาณ Gα
- ตัวกระตุ้นของทั้งวิถี Gαs และ Gαi /oคือ เอนไซม์ที่สร้างไซ คลิกอะดีโนซีนโมโนฟอสเฟต (cAMP) ซึ่งก็ คืออะ ดีนิเลตไซเคลสหรือ AC แม้ว่าจะมีผลิตภัณฑ์ยีน AC ที่แตกต่างกันถึงสิบชนิดในสัตว์เลี้ยงลูกด้วยนม โดยแต่ละชนิดมีความแตกต่างกันเล็กน้อยในด้าน การกระจายตัว ในเนื้อเยื่อหรือหน้าที่ แต่ทั้งหมด ก็เร่ง ปฏิกิริยาการเปลี่ยนอะดีโนซีนไตรฟอสเฟต (ATP) ในไซโตพลาสมิกไปเป็น cAMP และทั้งหมดก็ถูกกระตุ้นโดยตรงจากโปรตีน G ในกลุ่ม Gαs อย่างไรก็ตามในทางตรงกันข้าม การโต้ตอบกับหน่วยย่อย Gα ของชนิด Gαi /oจะยับยั้งการสร้าง cAMP ของ AC ดังนั้น GPCR ที่เชื่อมต่อกับ Gαs จะต่อต้านการทำงานของ GPCR ที่เชื่อมต่อกับ Gαi /oและในทางกลับกัน ระดับของ cAMP ในไซโตพลาสมิกจึงอาจกำหนดกิจกรรมของช่องไอออน ต่างๆ รวมถึงสมาชิกของ ตระกูล โปรตีนไคเนส A (PKA) ที่จำเพาะต่อซีรีน/ทรีโอนีน ด้วย ดังนั้น cAMP จึงถือเป็นสารสื่อประสาทตัวที่สองและ PKA เป็นตัวกระตุ้น รอง
- ตัวกระตุ้นของวิถี G αq/11คือฟอสโฟลิเปส C-β (PLCβ) ซึ่งเร่งปฏิกิริยาการแตกตัวของฟอสฟาติดิลอิโนซิทอล 4,5-บิสฟอสเฟต (PIP2) ที่ยึดติดกับเยื่อหุ้มเซลล์ ออกเป็นสารส่งสัญญาณรอง ได้แก่ อิโนซิทอล (1,4,5) ไตรฟอสเฟต (IP3) และไดแอซิลกลีเซอรอล (DAG) IP3 ออกฤทธิ์ต่อตัวรับ IP3ที่พบในเยื่อหุ้มของเอน โด พลาสมิกเรติคูลัม (ER) เพื่อกระตุ้น การปลดปล่อย Ca 2+จาก ER ในขณะที่ DAG แพร่กระจายไปตามเยื่อหุ้มพลาสมาซึ่งอาจกระตุ้นรูปแบบใดๆ ของเซริน/ทรีโอนีนไคเนสตัวที่สองที่เรียกว่าโปรตีนไคเนส C (PKC) เนื่องจากไอโซฟอร์มหลายชนิดของ PKC ก็ถูกกระตุ้นโดยการเพิ่มขึ้นของ Ca 2+ภายในเซลล์เช่นกัน ดังนั้นวิถีทั้งสองนี้จึงสามารถมาบรรจบกันเพื่อส่งสัญญาณผ่านตัวกระตุ้นรองตัวเดียวกันได้ แคลเซียมไอออน (Ca 2+) ที่สูงขึ้นภายในเซลล์ จะจับกับและกระตุ้นโปรตีนที่เรียกว่าแคลโมดูลิน (calmodulins) แบบอัลโลสเตอริก (allosteric activation) ซึ่งจะไปกระตุ้นGTPase ขนาดเล็ก ในไซโตพลาสซึมที่เรียกว่า Rhoเมื่อจับกับ GTP แล้ว Rho ก็สามารถไปกระตุ้นโปรตีนต่างๆ ที่รับผิดชอบในการควบคุมโครงสร้างเซลล์เช่นRho-kinase (ROCK) GPCR ส่วนใหญ่ที่เชื่อมต่อกับ G α12/13ก็เชื่อมต่อกับคลาสย่อยอื่นๆ ด้วย โดยส่วนใหญ่จะเป็น G αq/ 11
การส่งสัญญาณ Gβγ
คำอธิบายข้างต้นละเลยผลกระทบของ การส่งสัญญาณ Gβγซึ่งอาจมีความสำคัญเช่นกัน โดยเฉพาะอย่างยิ่งในกรณีของ GPCR ที่เชื่อมต่อกับ Gαi /oที่ถูกกระตุ้น ตัวกระตุ้นหลักของ Gβγ คือช่องไอออนต่างๆ เช่นช่อง K +ที่ควบคุมโดยโปรตีน G (GIRKs), ช่อง Ca2 +ที่ควบคุมด้วยแรงดันไฟฟ้า ชนิด P / QและN รวมถึงไอโซฟอร์มบางชนิดของ AC และ PLC พร้อมกับไอโซฟอร์มบางชนิดของ ฟ อสโฟอิโนซิไทด์-3-ไคเนส (PI3K)
การส่งสัญญาณที่ไม่ขึ้นกับโปรตีนจี
แม้ว่าโดยทั่วไปจะคิดว่า GPCR ทำงานร่วมกันเท่านั้น แต่ GPCR อาจส่งสัญญาณผ่านกลไกที่ไม่ขึ้นกับโปรตีน G และโปรตีน G แบบเฮเทอโรไตรเมอริกอาจมีบทบาทในการทำงานที่ไม่ขึ้นกับ GPCR GPCR อาจส่งสัญญาณอย่างอิสระผ่านโปรตีนหลายชนิดที่กล่าวถึงไปแล้วว่ามีบทบาทในการส่งสัญญาณที่ขึ้นกับโปรตีน G เช่นβ-arrs , GRKsและSrcsการส่งสัญญาณดังกล่าวได้รับการพิสูจน์แล้วว่ามีความเกี่ยวข้องทางสรีรวิทยา ตัวอย่างเช่น การส่งสัญญาณ β-arrestinที่เกิดจากตัวรับเคโมไคน์CXCR3มีความจำเป็นต่อการเคลื่อนที่ของเซลล์ T ที่ถูกกระตุ้นอย่างมีประสิทธิภาพเต็มที่[ 60 ]นอกจากนี้ โปรตีนโครงสร้างเพิ่มเติมที่เกี่ยวข้องกับการกำหนดตำแหน่งย่อยของเซลล์ของ GPCR (เช่น โปรตีนที่มี โดเมน PDZ ) อาจทำหน้าที่เป็นตัวส่งสัญญาณได้เช่นกัน บ่อยครั้งที่ตัวกระตุ้นเป็นสมาชิกของตระกูล MAPK
ตัวอย่าง
ในช่วงปลายทศวรรษ 1990 หลักฐานเริ่มสะสมมากขึ้นเพื่อบ่งชี้ว่า GPCR บางชนิดสามารถส่งสัญญาณได้โดยไม่ต้องมีโปรตีน G โปรตีนไคเนสที่กระตุ้นด้วยไมโท เจน ERK2ซึ่งเป็นตัวกลางการส่งสัญญาณที่สำคัญที่อยู่ปลายทางของการกระตุ้นตัวรับในหลายเส้นทาง ได้รับการแสดงให้เห็นว่าถูกกระตุ้นเพื่อตอบสนองต่อการกระตุ้นตัวรับที่เกิดจาก cAMP ในราเมือกD. discoideumแม้ว่าจะไม่มีหน่วยย่อย α- และ β- ของโปรตีน G ที่เกี่ยวข้องก็ตาม[ 61 ]
ในเซลล์ของสัตว์เลี้ยงลูกด้วยนม ตัวรับ β2-adrenoceptor ที่ได้รับการศึกษาอย่างกว้างขวางนั้นได้รับการพิสูจน์แล้วว่าสามารถกระตุ้นวิถี ERK2 ได้หลังจากที่ arrestin ทำหน้าที่แยกการส่งสัญญาณผ่านโปรตีน G ดังนั้น จึงดูเหมือนว่ากลไกบางอย่างที่เคยเชื่อว่าเกี่ยวข้องกับการลดความไวของตัวรับเพียงอย่างเดียวนั้น แท้จริงแล้วอาจเป็นตัวอย่างของการที่ตัวรับเปลี่ยนวิถีการส่งสัญญาณมากกว่าที่จะถูกปิดใช้งานไปเฉยๆ
ในเซลล์ไตพบว่าตัวรับแบรดิกินิน B2 มีปฏิสัมพันธ์โดยตรงกับโปรตีนไทโรซีนฟอสฟาเทส การมีลำดับ ITIM (immunoreceptor tyrosine-based inhibitory motif) ที่ถูกฟอสโฟรีเลตด้วยไทโรซีนในตัวรับ B2 เป็นสิ่งจำเป็นในการเป็นตัวกลางในปฏิสัมพันธ์นี้ และส่งผลให้เกิดผลต้านการแพร่กระจายของแบรดิกินิน[ 62 ]
การส่งสัญญาณโดยไม่ขึ้นกับ GPCR โดยโปรตีน G แบบเฮเทอโรไตรเมอริก
แม้ว่าจะเป็นสาขาการวิจัยที่ยังค่อนข้างใหม่ แต่ดูเหมือนว่าโปรตีน G แบบเฮเทอโรไตรเมอริกอาจมีส่วนร่วมในการส่งสัญญาณที่ไม่เกี่ยวข้องกับ GPCR ด้วยเช่นกัน มีหลักฐานที่แสดงให้เห็นถึงบทบาทในฐานะตัวส่งสัญญาณในกระบวนการส่งสัญญาณผ่านตัวรับเกือบทุกประเภท รวมถึงอินทิกรินตัวรับไทโรซีนไคเนส (RTK) ตัวรับไซโตไคน์ ( JAK/STAT ) ตลอดจนการปรับเปลี่ยนโปรตีน "เสริม" อื่นๆ เช่นGEF สารยับยั้งการแยกตัวของกัวนีนนิวคลีโอไทด์ (GDI) และโปรตีนฟอสฟาเทสอาจมีโปรตีนเฉพาะในกลุ่มเหล่านี้ที่มีหน้าที่หลักเป็นส่วนหนึ่งของเส้นทางที่ไม่ขึ้นกับ GPCR ซึ่งเรียกว่าตัวกระตุ้นการส่งสัญญาณของโปรตีน G (AGS) ทั้งความแพร่หลายของการปฏิสัมพันธ์เหล่านี้และความสำคัญของหน่วยย่อย Gα เทียบกับ Gβγ ในกระบวนการเหล่านี้ยังคงไม่ชัดเจน
รายละเอียดเกี่ยวกับวิถีการทำงานของ cAMP และ PIP2



มีเส้นทางการส่งสัญญาณหลักสองเส้นทางที่เกี่ยวข้องกับตัวรับที่เชื่อมโยงกับโปรตีน Gได้แก่ เส้นทางการส่งสัญญาณ cAMPและเส้นทางการส่งสัญญาณฟ อสฟา ติดิลอิโนซิทอล[ 5 ]
เส้นทางการส่งสัญญาณ cAMP
กระบวนการส่งสัญญาณ cAMP ประกอบด้วยองค์ประกอบหลัก 5 ส่วน ได้แก่ ตัวรับ ฮอร์โมน กระตุ้น (Rs) หรือตัวรับฮอร์โมน ยับยั้ง (Ri); โปรตีน G ควบคุมกระตุ้น (Gs) หรือโปรตีน G ควบคุมยับยั้ง (Gi); อะเดนิลไซเคลส ; โปรตีนไคเนส A (PKA); และ cAMP ฟอสโฟไดเอสเทอเรส
ตัวรับฮอร์โมนกระตุ้น (Rs) เป็นตัวรับที่สามารถจับกับโมเลกุลสัญญาณกระตุ้น ในขณะที่ตัวรับฮอร์โมนยับยั้ง (Ri) เป็นตัวรับที่สามารถจับกับโมเลกุลสัญญาณยับยั้ง
โปรตีน G ควบคุมแบบกระตุ้น คือโปรตีน G ที่เชื่อมต่อกับตัวรับฮอร์โมนกระตุ้น (Rs) และหน่วยย่อยอัลฟาของมันเมื่อถูกกระตุ้นจะสามารถกระตุ้นการทำงานของเอนไซม์หรือกระบวนการเผาผลาญภายในเซลล์อื่นๆ ในทางตรงกันข้าม โปรตีน G ควบคุมแบบยับยั้ง จะเชื่อมต่อกับตัวรับฮอร์โมนยับยั้ง และหน่วยย่อยอัลฟาของมันเมื่อถูกกระตุ้นจะสามารถยับยั้งการทำงานของเอนไซม์หรือกระบวนการเผาผลาญภายในเซลล์อื่นๆ
อะเดนิลไซเคลสเป็นไกลโคโปรตีนที่มีโครงสร้างแบบทรานส์เมมเบรน 12 ตำแหน่ง ซึ่งทำหน้าที่เร่งปฏิกิริยาการเปลี่ยน ATP ไปเป็น cAMP โดยอาศัยโคแฟคเตอร์ Mg 2+หรือ Mn 2+ cAMP ที่เกิดขึ้นนั้นเป็นสารสื่อสัญญาณรองในกระบวนการเผาผลาญของเซลล์ และเป็นตัวกระตุ้นแบบอัลโลสเตอริกของโปรตีนไคเนส A
โปรตีนไคเนสเอเป็นเอนไซม์สำคัญในกระบวนการเผาผลาญของเซลล์ เนื่องจากความสามารถในการควบคุมกระบวนการเผาผลาญของเซลล์โดยการเติมหมู่ฟอสเฟตให้กับเอนไซม์เฉพาะที่เกี่ยวข้องในวิถีการเผาผลาญ นอกจากนี้ยังสามารถควบคุมการแสดงออกของยีน การหลั่งสารของเซลล์ และการซึมผ่านของเยื่อหุ้มเซลล์ได้อีกด้วย เอนไซม์โปรตีนนี้ประกอบด้วยหน่วยย่อยเร่งปฏิกิริยา 2 หน่วย และหน่วยย่อยควบคุม 2 หน่วย เมื่อไม่มี cAMP คอมเพล็กซ์จะทำงานไม่ได้ เมื่อ cAMP จับกับหน่วยย่อยควบคุม โครงสร้างของหน่วยย่อยควบคุมจะเปลี่ยนไป ทำให้หน่วยย่อยควบคุมแยกตัวออกจากกัน ซึ่งจะกระตุ้นโปรตีนไคเนสเอและทำให้เกิดผลทางชีวภาพต่อไป
จากนั้นสัญญาณเหล่านี้สามารถถูกยุติได้โดย cAMP phosphodiesterase ซึ่งเป็นเอนไซม์ที่ย่อยสลาย cAMP ให้เป็น 5'-AMP และทำให้โปรตีนไคเนส A ไม่ทำงาน
เส้นทางการส่งสัญญาณของฟอสฟาติดิลอิโนซิทอล
ใน วิถีการส่งสัญญาณฟอสฟาติดิลอิโนซิ ทอล โมเลกุลสัญญาณภายนอกเซลล์จะจับกับตัวรับโปรตีนจี (Gq )บนพื้นผิวเซลล์และกระตุ้นเอนไซม์ฟอสโฟลิเปสซีซึ่งอยู่บนเยื่อ หุ้ม เซลล์เอนไซม์ลิเปสจะไฮโดรไลซ์ฟอสฟาติดิลอิโนซิทอล 4,5-บิสฟอสเฟต (PIP2) ให้เป็นสารส่งสัญญาณรองสองชนิด ได้แก่อิโนซิทอล 1,4,5-ไตรฟอสเฟต (IP3)และไดอะซิลกลีเซอรอล (DAG) IP3 จะจับกับตัวรับ IP3ในเยื่อหุ้มของเอนโดพลาสมิกเรติคูลัมเรียบและไมโทคอนเดรียเพื่อเปิดช่องแคลเซียม (Ca2 + ) DAG ช่วยกระตุ้นโปรตีนไคเนสซี (PKC) ซึ่งจะฟอสฟอริเลตโปรตีนอื่นๆ อีกหลายชนิด เปลี่ยนแปลงกิจกรรมเร่งปฏิกิริยาของโปรตีนเหล่านั้น นำไปสู่การตอบสนองของเซลล์
ผลกระทบของ Ca²⁺ ก็มีความน่าทึ่งเช่นกัน: มันทำงานร่วมกับ DAG ในการกระตุ้น PKC และสามารถกระตุ้น วิถีการส่งสัญญาณ ของ CaM kinaseซึ่งโปรตีนcalmodulin (CaM) ที่ถูกควบคุมด้วยแคลเซียมจะจับกับ Ca²⁺ เกิดการเปลี่ยนแปลงโครงสร้าง และกระตุ้น CaM kinase II ซึ่งมีความสามารถพิเศษในการเพิ่มความสามารถในการจับกับ CaM โดยการฟอสโฟรีเลชันตัวเอง ทำให้ CaM ไม่สามารถนำไปใช้ในการกระตุ้นเอนไซม์อื่นๆ ได้ จากนั้นไคเนสจะฟอสโฟรีเลตเอนไซม์เป้าหมาย ควบคุมกิจกรรมของเอนไซม์เหล่านั้น วิถีการส่งสัญญาณทั้งสองเชื่อมต่อกันด้วย Ca²⁺ - CaM ซึ่งเป็นหน่วยย่อยควบคุมของ adenylyl cyclase และ phosphodiesterase ในวิถีการส่งสัญญาณ cAMP ด้วย
การควบคุมตัวรับ
ตัวรับ GPCR จะเกิดการลดความไวเมื่อสัมผัสกับลิแกนด์เป็นเวลานาน การลดความไวมีสองรูปแบบที่ได้รับการยอมรับ ได้แก่ 1) การลดความไว แบบเดียวกัน (homologous desensitization ) ซึ่ง GPCR ที่ถูกกระตุ้นจะถูกลดระดับลง และ 2) การลดความไวแบบต่างชนิด (heterologous desensitization ) ซึ่ง GPCR ที่ถูกกระตุ้นจะทำให้ GPCR อื่นลดระดับลง ปฏิกิริยาสำคัญของการลดระดับนี้คือการฟอสฟอริเลชัน ของ โดเมนตัวรับภายในเซลล์ (หรือไซโตพลาสซึม ) โดย โปรตีนไคเนส
การฟอสฟอริเลชันโดยโปรตีนไคเนสที่ขึ้นอยู่กับ cAMP
โปรตีนไคเนสที่ขึ้นอยู่กับไซคลิก AMP ( โปรตีนไคเนส A ) จะถูกกระตุ้นโดยสายโซ่สัญญาณที่มาจากโปรตีน G (ที่ถูกกระตุ้นโดยตัวรับ) ผ่านทางอะดีนิเลตไซเคลสและไซคลิก AMP (cAMP) ในกลไกป้อนกลับไคเนสที่ถูกกระตุ้นเหล่านี้จะฟอสโฟรีเลตตัวรับ ยิ่งตัวรับยังคงทำงานนานเท่าใด ไคเนสก็จะถูกกระตุ้นมากขึ้น และตัวรับก็จะถูกฟอสโฟรีเลตมากขึ้นเท่านั้น ใน ตัวรับ β2 - adrenoceptor การฟอสโฟรี เลตนี้ส่งผลให้การเชื่อมต่อเปลี่ยนจากคลาส Gs ของโปรตีน G ไปเป็นคลาสGi [ 63 ]การฟอสโฟรีเลตที่เกิดจาก PKA ที่ขึ้นอยู่กับ cAMP สามารถทำให้เกิดการลดความไวแบบต่างชนิดกันในตัวรับอื่นนอกเหนือจากตัวรับที่ถูกกระตุ้น[ 64 ]
การฟอสฟอริเลชันโดย GRK
ไคเนสตัวรับที่เชื่อมต่อกับโปรตีน G (GRKs) เป็นโปรตีนไคเนสที่ฟอสโฟรีเลตเฉพาะ GPCR ที่ทำงานอยู่เท่านั้น[ 65 ]ไคเนสตัวรับที่เชื่อมต่อกับโปรตีน G (GRKs) เป็นตัวปรับสำคัญของการส่งสัญญาณของตัวรับที่เชื่อมต่อกับโปรตีน G (GPCR) พวกมันประกอบด้วยโปรตีนไคเนสเซริน-ทรีโอนีนในสัตว์เลี้ยงลูกด้วยนมเจ็ดชนิดที่ฟอสโฟรีเลตตัวรับที่จับกับตัวกระตุ้น การฟอสโฟรีเลตตัวรับที่เกิดจาก GRKs จะเริ่มต้นอย่างรวดเร็วทำให้การส่งสัญญาณของตัวรับบกพร่องอย่างมากและเกิดการลดความไว กิจกรรมของ GRKs และการกำหนดเป้าหมายในระดับเซลล์ย่อยได้รับการควบคุมอย่างเข้มงวดโดยการโต้ตอบกับโดเมนของตัวรับ หน่วยย่อยของโปรตีน G ไขมัน โปรตีนยึดเกาะ และโปรตีนที่ไวต่อแคลเซียม[ 66 ]
การฟอสฟอริเลชันของตัวรับสามารถส่งผลได้สองประการ:
- การเคลื่อนย้าย : ตัวรับจะถูกนำเข้าไปภายในเซลล์พร้อมกับส่วนของเยื่อหุ้มเซลล์ที่มันฝังอยู่ โดยจะถูกกำจัดฟอสเฟตภายในสภาพแวดล้อมของถุงเวสิเคิลที่เป็นกรด[ 67 ]แล้วจึงนำกลับเข้าไป กลไกนี้ใช้ในการควบคุมการสัมผัสในระยะยาว เช่น ฮอร์โมน โดยอนุญาตให้เกิดการตอบสนองต่อสิ่งเร้าอีกครั้งหลังจากเกิดการลดการตอบสนองต่อสิ่งเร้า หรืออีกทางหนึ่ง ตัวรับอาจถูกย่อยสลายโดยไลโซโซม หรือยังคงอยู่ภายในเซลล์ ซึ่งเชื่อกันว่ามันมีส่วนร่วมในการเริ่มต้นเหตุการณ์การส่งสัญญาณ ซึ่งลักษณะของเหตุการณ์นั้นขึ้นอยู่กับตำแหน่งย่อยของเซลล์ของถุงเวสิเคิลที่อยู่ภายในเซลล์[ 64 ]
- การเชื่อมโยงอาร์เรสติน : ตัวรับที่ถูกฟอสโฟรีเลตสามารถเชื่อมโยงกับ โมเลกุล อาร์เรสตินที่ป้องกันไม่ให้จับ (และกระตุ้น) โปรตีน G ซึ่งมีผลทำให้ปิดการทำงานในช่วงเวลาสั้นๆ กลไกนี้ถูกนำมาใช้ เช่น กับโรดอปซินใน เซลล์ เรตินาเพื่อชดเชยการสัมผัสกับแสงสว่าง ในหลายกรณี การจับของอาร์เรสตินกับตัวรับเป็นสิ่งจำเป็นสำหรับการเคลื่อนย้าย ตัวอย่างเช่น เบต้า-อาร์เรสตินที่จับกับ β2-อะดรีโนรีเซปเตอร์ทำหน้าที่เป็นตัวเชื่อมสำหรับการจับกับคลัทริน และกับหน่วยย่อยเบต้าของ AP2 (โมเลกุลตัวเชื่อมคลัทริน) ดังนั้น อาร์เรสตินในที่นี้จึงทำหน้าที่เป็นโครงสร้างที่ประกอบส่วนประกอบที่จำเป็นสำหรับการเอนโดไซโทซิสที่เกิดจากคลัทรินของ β2-อะดรีโนรีเซปเตอร์ [ 68 ] [ 69 ]
กลไกการยุติสัญญาณของ GPCR
ดังที่กล่าวมาข้างต้น โปรตีน G อาจยุติการทำงานของตัวเองได้เนื่องจาก ความสามารถ ในการไฮโดรไลซิส GTP→GDP โดยธรรมชาติ อย่างไรก็ตาม ปฏิกิริยานี้เกิดขึ้นในอัตรา ที่ช้า (≈0.02 ครั้ง/วินาที) ดังนั้นจึงต้องใช้เวลาประมาณ 50 วินาทีสำหรับโปรตีน G ตัวใดตัวหนึ่งที่จะหยุดการทำงานหากไม่มีปัจจัยอื่นเข้ามาเกี่ยวข้อง อันที่จริง มีโปรตีน RGS ประมาณ 30 ไอโซฟอร์มที่เมื่อจับกับ Gα ผ่านโดเมน GAP ของมัน จะเร่งอัตราการไฮโดรไลซิสให้เร็วขึ้นเป็น ≈30 ครั้ง/วินาที การเพิ่มอัตราขึ้น 1500 เท่านี้ทำให้เซลล์สามารถตอบสนองต่อสัญญาณภายนอกด้วยความเร็วสูง รวมถึงความละเอียด เชิงพื้นที่ เนื่องจากปริมาณของตัวส่งสัญญาณรองที่สามารถสร้างขึ้นได้มีจำกัด และระยะทางที่โปรตีน G สามารถแพร่กระจายได้ใน 0.03 วินาทีมีจำกัด โดยส่วนใหญ่ โปรตีน RGS มีความสามารถในการยับยั้งการทำงานของโปรตีน G ได้หลากหลาย แต่การที่ RGS ตัวใดจะเกี่ยวข้องกับวิถีการส่งสัญญาณใดนั้น ดูเหมือนจะขึ้นอยู่กับเนื้อเยื่อและ GPCR ที่เกี่ยวข้องมากกว่าสิ่งอื่นใด นอกจากนี้ โปรตีน RGS ยังมีหน้าที่เพิ่มเติมในการเพิ่มอัตราการแลกเปลี่ยน GTP-GDP ที่ GPCR (กล่าวคือ เป็นเหมือน co-GEF ชนิดหนึ่ง) ซึ่งช่วยเพิ่มความละเอียดของเวลาในการส่งสัญญาณของ GPCR อีกด้วย
นอกจากนี้ GPCR อาจเกิดการลดความไวลงได้เอง ซึ่งอาจเกิดขึ้นได้ดังนี้:
- เป็นผลโดยตรงจากการจับกันของลิแกนด์ซึ่งการเปลี่ยนแปลงโครงสร้างทำให้เกิดการดึงดูดไคเนสที่ควบคุม GPCR (GRK) ซึ่งจะไปฟอสโฟรี เลตหมู่ เซริน / ทรีโอ นีน ต่างๆของ IL-3 และส่วนปลาย C-terminal เมื่อ GRK ฟอสโฟรีเลตแล้ว ความสัมพันธ์ของ GPCR กับβ-arrestin (β-arrestin-1/2 ในเนื้อเยื่อส่วนใหญ่) จะเพิ่มขึ้น ณ จุดนี้ β-arrestin อาจจับและทำหน้าที่ทั้งขัดขวางการเชื่อมต่อของโปรตีน G และเริ่มต้นกระบวนการนำตัวรับเข้าสู่เซลล์ผ่านเอนโดไซโทซิสที่ อาศัยคลัทริน เนื่องจากมีเพียงตัวรับที่จับกับลิแกนด์เท่านั้นที่ไวต่อการกระตุ้นน้อยลงด้วยกลไกนี้ จึงเรียกว่าการลด ความไวต่อการกระตุ้นแบบเดียวกัน (homologous desensitization)
- ความสัมพันธ์ของ β-arrestin อาจเพิ่มขึ้นในลักษณะการครอบครองลิแกนด์และไม่ขึ้นกับ GRK ผ่านการฟอสโฟรีเลชันของไซต์ ser/thr ที่แตกต่างกัน (แต่ยังรวมถึง IL-3 และหางปลาย C ด้วย) โดย PKC และ PKA การฟอสโฟรีเลชันเหล่านี้มักจะเพียงพอที่จะทำให้การเชื่อมต่อโปรตีน G บกพร่องได้ด้วยตัวเองเช่นกัน[ 70 ]
- ในทางกลับกัน PKC/PKA อาจทำการฟอสฟอริเลต GRK ซึ่งอาจนำไปสู่การฟอสฟอริเลต GPCR และการจับกับ β-arrestin ในลักษณะที่ไม่ขึ้นกับการจับกัน กลไกสองอย่างหลังนี้ทำให้เกิดการลดความไวของ GPCR ตัวหนึ่งเนื่องจากการทำงานของ GPCR ตัวอื่น หรือการลดความไวแบบต่างชนิดกัน GRK อาจมีโดเมน GAP ด้วย ดังนั้นจึงอาจมีส่วนช่วยในการยับยั้งผ่านกลไกที่ไม่ใช่ไคเนส ได้ เช่นกัน นอกจากนี้ยังอาจเกิดการรวมกันของกลไกเหล่านี้ได้ด้วย
เมื่อ β-arrestin จับกับ GPCR แล้ว มันจะเกิดการเปลี่ยนแปลงโครงสร้าง ทำให้มันสามารถทำหน้าที่เป็นโปรตีนโครงสร้างสำหรับคอมเพล็กซ์ตัวเชื่อมต่อที่เรียกว่าAP-2ซึ่งจะดึงดูดโปรตีนอีกตัวหนึ่งที่เรียกว่าclathrin เข้ามา หากมีตัวรับในบริเวณนั้นดึงดูด clathrin เข้ามามากพอในลักษณะนี้ พวกมันจะรวมตัวกันและ เยื่อ หุ้มเซลล์จะโป่งออกด้านในอันเป็นผลมาจากการโต้ตอบระหว่างโมเลกุลของ clathrin ในกระบวนการที่เรียกว่าopsonizationเมื่อหลุมถูกบีบออกจากเยื่อหุ้มเซลล์เนื่องจากการทำงานของโปรตีนอีกสองชนิดที่เรียกว่าamphiphysinและdynaminมันจะกลายเป็นถุงเอนโดไซติก ในขั้นตอนนี้ โมเลกุลตัวเชื่อมต่อและ clathrin จะแยกตัวออกจากกันและตัวรับจะถูกส่งกลับไปยังเยื่อหุ้มเซลล์หรือถูกส่งไปยังไลโซโซมเพื่อย่อยสลาย
ในขั้นตอนใดๆ ของกระบวนการนี้ β-arrestins อาจดึงดูดโปรตีนอื่นๆ เช่นnon-receptor tyrosine kinase (nRTK) หรือc-SRCซึ่งอาจกระตุ้นERK1/2หรือmitogen-activated protein kinase (MAPK) อื่นๆ ผ่านการฟอสฟอริเลชันของGTPase ขนาดเล็กRasหรือดึงดูดโปรตีนในERK cascadeโดยตรง (เช่นRaf-1 , MEK , ERK-1/2) ซึ่งในจุดนี้การส่งสัญญาณจะเริ่มต้นขึ้นเนื่องจากอยู่ใกล้กันมาก เป้าหมายอีกอย่างหนึ่งของ c-SRC คือโมเลกุลไดนามินที่เกี่ยวข้องกับเอนโดไซโทซิส ไดนามินจะรวมตัวกันเป็นพอลิเมอร์รอบคอของเวสิเคิลที่เข้ามา และการฟอสฟอริเลชันโดย c-SRC จะให้พลังงานที่จำเป็นสำหรับการเปลี่ยนแปลงโครงสร้าง ทำให้เกิดการ "แยกตัว" ออกจากเยื่อหุ้มเซลล์ในที่สุด
การควบคุมระดับเซลล์ของ GPCR
การลดความไวของตัวรับเกิดขึ้นผ่านกระบวนการฟอสโฟรีเลชัน การจับกับ β-arr และเอนโดไซโทซิสร่วมกัน ดังที่กล่าวมาข้างต้น การลดระดับเกิดขึ้นเมื่อตัวรับที่ถูกเอนโดไซโทซิสฝังตัวอยู่ในเอนโดโซม ซึ่งจะถูกส่งไปรวมกับออร์แกเนลล์ที่เรียกว่าไลโซโซม เนื่องจากเยื่อหุ้มไลโซโซมอุดมไปด้วยปั๊มโปรตอน ภายในจึงมีค่า pH ต่ำ (≈4.8 เทียบกับ pH≈7.2 ของไซโทซอล) ซึ่งทำหน้าที่ในการทำลายโครงสร้างของ GPCR นอกจากนี้ ไลโซโซมยังมีเอนไซม์ย่อยสลายหลายชนิด รวมถึงโปรตีเอส ซึ่งสามารถทำงานได้เฉพาะในสภาวะ pH ต่ำเช่นนี้ ดังนั้นพันธะเปปไทด์ที่เชื่อมต่อสารตกค้างของ GPCR เข้าด้วยกันอาจถูกตัดขาด การที่ตัวรับจะถูกส่งไปยังไลโซโซม ถูกกักไว้ในเอนโดโซม หรือถูกส่งกลับไปยังเยื่อหุ้มเซลล์นั้นขึ้นอยู่กับปัจจัยหลายอย่าง รวมถึงชนิดของตัวรับและขนาดของสัญญาณ นอกจากนี้ การควบคุม GPCR ยังเกิดขึ้นผ่านปัจจัยการถอดรหัสยีน ปัจจัยเหล่านี้สามารถเพิ่มหรือลดการถอดรหัสยีน และด้วยเหตุนี้จึงเพิ่มหรือลดการสร้างตัวรับใหม่ (การควบคุมขึ้นหรือลง) ที่เคลื่อนที่ไปยังเยื่อหุ้มเซลล์
การรวมตัวของตัวรับ
การรวมตัวกันของตัวรับที่เชื่อมต่อกับโปรตีน G เป็นปรากฏการณ์ที่พบได้ทั่วไป ตัวอย่างหนึ่งที่ได้รับการศึกษาอย่างดีที่สุดคือตัวรับGABA B แบบเมตาโบโทรปิก ตัวรับที่เรียกว่าตัวรับแบบคงที่นี้เกิดขึ้นจากการรวมตัวกันของหน่วยย่อยGABA B R1และGABA B R2 การแสดงออกของ GABA B R1 โดยไม่มี GABA B R2 ในระบบต่างชนิดกันทำให้หน่วยย่อยถูกกักเก็บไว้ในเอนโดพลาสมิกเรติคูลัม ในขณะเดียวกัน การแสดงออกของหน่วยย่อย GABA B R2 เพียงอย่างเดียวทำให้หน่วยย่อยแสดงออกที่พื้นผิว แม้ว่าจะไม่มีกิจกรรมการทำงาน (เช่น ตัวรับไม่จับกับตัวกระตุ้นและไม่สามารถเริ่มต้นการตอบสนองหลังจากการสัมผัสกับตัวกระตุ้น) การแสดงออกของหน่วยย่อยทั้งสองร่วมกันทำให้ตัวรับที่มีฟังก์ชันการทำงานแสดงออกที่เยื่อหุ้มเซลล์ มีการแสดงให้เห็นว่าการจับกันของ GABA B R2 กับ GABA B R1 ทำให้เกิดการปิดบังสัญญาณการกักเก็บ[ 71 ]ของตัวรับที่มีฟังก์ชันการทำงาน[ 72 ]
ที่มาและการกระจายตัวของซูเปอร์แฟมิลี
การส่งสัญญาณผ่านซูเปอร์แฟมิลีของ GPCR มีมาตั้งแต่กำเนิดของสิ่งมีชีวิตหลายเซลล์ GPCR ที่คล้ายกับสัตว์เลี้ยงลูกด้วยนมพบได้ในเชื้อราและได้รับการจัดประเภทตาม ระบบการจำแนกประเภท GRAFSโดยอิงจากลายนิ้วมือของ GPCR [ 19 ]การระบุสมาชิกของซูเปอร์แฟมิลีใน โดเมน ยูคาริโอตและการเปรียบเทียบโมทีฟเฉพาะตระกูล แสดงให้เห็นว่าซูเปอร์แฟมิลีของ GPCR มีต้นกำเนิดร่วมกัน[ 73 ]โมทีฟลักษณะเฉพาะบ่งชี้ว่าสามในห้าตระกูล GRAFS ได้แก่Rhodopsin , AdhesionและFrizzledวิวัฒนาการมาจาก ตัวรับ cAMP ของ Dictyostelium discoideumก่อนการแยกตัวของopisthokontsต่อ มา ตระกูล Secretinวิวัฒนาการมาจาก ตระกูลตัวรับ GPCR Adhesionก่อนการแยกตัวของหนอนตัวกลม[ 19 ] GPCR ของแมลงดูเหมือนจะอยู่ในกลุ่มของตัวเอง และ Taste2 ถูกระบุว่าสืบ เชื้อสายมาจากRhodopsin [ 73 ]โปรดทราบว่า การแบ่ง Secretin / Adhesion นั้น ขึ้นอยู่กับหน้าที่ที่คาดการณ์ไว้มากกว่าลายเซ็น เนื่องจากคลาส B แบบคลาสสิก (7tm_2, Pfam PF00002 ) ใช้ในการระบุทั้งสองอย่างในการศึกษา
ดูเพิ่มเติม
- ฐานข้อมูลตัวรับที่เชื่อมโยงกับโปรตีนจี
- รายการรหัส MeSH (D12.776)
- ตัวรับเมตาโบโทรปิก
- ตัวรับกำพร้า
- เปปดูซินส์เป็นกลุ่มยาที่มุ่งเป้าไปที่ตัวรับ GPCR
- ตัวรับที่ถูกกระตุ้นโดยลิแกนด์สังเคราะห์เพียงอย่างเดียวเทคนิคการควบคุมการส่งสัญญาณของเซลล์ผ่าน GPCR สังเคราะห์
- ซูเปอร์แฟมิลี่ TOG
อ่านเพิ่มเติม
- Vassilatis DK, Hohmann JG, Zeng H, Li F, Ranchalis JE, Mortrud MT และคณะ (เมษายน 2546). "เรพเพอร์ทัวร์ของตัวรับที่เชื่อมโยงกับโปรตีน G ของมนุษย์และหนู" Proceedings of the National Academy of Sciences of the United States of America . 100 (8): 4903– 8. Bibcode : 2003PNAS..100.4903V . doi : 10.1073/ pnas.0230374100 . PMC 153653. PMID 12679517 .
- "ห้องสมุดอ้างอิง GPCR" สืบค้นเมื่อ 11 สิงหาคม 2551
แหล่งอ้างอิงสำหรับแบบจำลองระดับโมเลกุลและทางคณิตศาสตร์สำหรับการตอบสนองของตัวรับในระยะเริ่มต้น
- "รางวัลโนเบลสาขาเคมี ประจำปี 2012" (PDF) . เก็บถาวร(PDF)จากต้นฉบับเมื่อวันที่ 18 ตุลาคม 2012 . เรียกดูเมื่อวันที่ 10 ตุลาคม 2012 .
ลิงก์ภายนอก
- ตัวรับที่เชื่อมต่อกับโปรตีนจี (G-protein-coupled+receptors) ในหัวข้อทางการแพทย์ (Medical Subject Headingsหรือ MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา (US National Library of Medicine )
- เซลล์สายพันธุ์ GPCR ถูกเก็บถาวรเมื่อวันที่ 3 เมษายน 2558 ที่Wayback Machine
- "คู่มือฐานข้อมูลเภสัชวิทยาของ IUPHAR/BPS (GPCRs)"ฐานข้อมูล IUPHARมหาวิทยาลัยเอดินบะระ / สหภาพเภสัชวิทยาพื้นฐานและคลินิกนานาชาติสืบค้นเมื่อ 6 กุมภาพันธ์ 2019
- "GPCRdb"แหล่ง
ข้อมูล แผนภาพ และเครื่องมือบนเว็บสำหรับตัวรับที่เชื่อมต่อกับโปรตีนจี (GPCRs)
; มังค์ ซี, อิสเบิร์ก วี, มอร์ดัลสกี เอส, ฮาร์ปโซอี เค, ราทาจ เค, เฮาเซอร์ AS, และคณะ (กรกฎาคม 2559). GPCRdb: ฐานข้อมูลตัว รับคู่โปรตีน G - บทนำวารสารเภสัชวิทยาอังกฤษ . 173 (14): 2195– 207. ดอย : 10.1111/bph.13509 . PMC 4919580 . PMID27155948 . - "ตัวรับที่เชื่อมโยงกับโปรตีนจีบนเครือข่าย"เก็บถาวรจากต้นฉบับเมื่อวันที่ 23 กรกฎาคม 2554 เรียกดูเมื่อวันที่ 10 พฤศจิกายน 2553
การจำแนกประเภทของ GPCRs
- " ศูนย์เครือข่าย PSI GPCR"เก็บถาวรจากต้นฉบับเมื่อวันที่ 25 กรกฎาคม 2556 เรียกดูเมื่อวันที่ 11 กรกฎาคม 2556
โครงการริเริ่มโครงสร้างโปรตีน: ศูนย์เครือข่ายชีววิทยา มีเป้าหมายเพื่อกำหนดโครงสร้างสามมิติของโปรตีนในกลุ่ม GPCR ที่เป็นตัวแทน
- GPCR-HGmod เก็บถาวรเมื่อวันที่ 1 กุมภาพันธ์ 2016 ที่Wayback Machineฐานข้อมูลแบบจำลองโครงสร้าง 3 มิติของตัวรับ G-protein coupled receptor ของมนุษย์ทั้งหมด สร้างโดยไปป์ไลน์ GPCR -I-TASSER Zhang J, Yang J, Jang R, Zhang Y (สิงหาคม 2015). "GPCR-I-TASSER: แนวทางแบบผสมผสานสำหรับการสร้างแบบจำลองโครงสร้างตัวรับ G Protein-Coupled Receptor และการประยุกต์ใช้กับจีโนมมนุษย์" . Structure . 23 (8): 1538– 1549. doi : 10.1016/j.str.2015.06.007 . PMC 4526412 . PMID 26190572 .
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ตัวรับที่เชื่อมต่อกับโปรตีนจี
ตัวรับที่เชื่อมต่อกับโปรตีน G ( GPCRs ) หรือที่รู้จักกันในชื่อ ตัวรับโดเมนทรานส์เมมเบรนเจ็ดครั้ง (seven-(pass)-transmembrane domain receptors) , ตัวรับ 7TM , ตัว รับเฮปตา เฮลิคอล...
ประวัติและความสำคัญ
ด้วยการค้นพบโครงสร้างแรกของสารประกอบเชิงซ้อนระหว่างตัวรับที่เชื่อมโยงกับโปรตีนจี (GPCR) และไตรเมอร์ของโปรตีนจี (Gαβγ) ในปี 2011 ทำให้เกิดบทใหม่ของการวิจัย GPCR สำหรับการศึกษาโครงสร้างของสวิตช์ระดับโลกที่มีโปรตีนมากกว่าหนึ่งชนิดถูกตรวจสอบ...
การจำแนกประเภท
ขนาดที่แน่นอนของซูเปอร์แฟมิลี GPCR ยังไม่เป็นที่ทราบแน่ชัด แต่ มีการคาดการณ์ว่า ยีน ของมนุษย์ อย่างน้อย 831 ยีน (หรือประมาณ 4% ของ จีโนม ที่เข้ารหัสโปรตีน ทั้งหมด ) เข้ารหัสสำหรับซูเปอร์แฟมิลีนี้จากการวิเคราะห์ลำดับจีโนม[ 11 ] [ 12 ] แม้ว่า จะ...
บทบาททางสรีรวิทยา
GPCR มีส่วนเกี่ยวข้องในกระบวนการทางสรีรวิทยาที่หลากหลาย ตัวอย่างบทบาททางสรีรวิทยาของ GPCR ได้แก่: