อ่าน 5 นาที
ช่องโพแทสเซียมแบบปรับทิศทางเข้า
ช่องโพแทสเซียมแบบปรับทิศทางเข้า ( Kir , IRK ) เป็นกลุ่มย่อยของช่องโพแทสเซียมที่ควบคุมด้วยลิปิดโดย เฉพาะ จนถึงปัจจุบัน...
ช่องโพแทสเซียมแบบปรับทิศทางเข้า
| ช่องโพแทสเซียมแบบปรับทิศทางเข้า | |||||||
|---|---|---|---|---|---|---|---|
 โครงสร้างผลึกของช่องโพแทสเซียมแบบปรับทิศทางเข้า | |||||||
| ตัวระบุ | |||||||
| เครื่องหมาย | รบกวน | ||||||
| พีแฟม | พีเอฟ01007 | ||||||
| ตระกูลพีแฟม | ซีแอล0030 | ||||||
| อินเตอร์โปร | IPR013521 | ||||||
| สโคป2 | 1n9p / SCOPe / SUPFAM | ||||||
| ทีซีดีบี | 1.ก.2 | ||||||
| ซูเปอร์แฟมิลี OPM | 8 | ||||||
| โปรตีน OPM | 3SPG | ||||||
| |||||||
{kind=link}
ช่องโพแทสเซียมแบบปรับทิศทางเข้า ( Kir , IRK ) เป็นกลุ่มย่อยของช่องโพแทสเซียมที่ควบคุมด้วยลิปิดโดย เฉพาะ จนถึงปัจจุบัน มีการระบุกลุ่มย่อยเจ็ดกลุ่มในเซลล์สัตว์เลี้ยงลูกด้วยนมหลายชนิด1 ]พืช[ 2 ]และแบคทีเรีย[ 3 ] ช่อง เหล่านี้ถูกกระตุ้นโดยฟอสฟาติดิลอิโนซิทอล 4,5-บิสฟอสเฟต ( )การทำงานผิดปกติของช่องเหล่านี้เกี่ยวข้องกับโรคหลายชนิด[ 4 ] [ 5 ]ช่อง IRK มีโดเมนรูพรุน ซึ่งคล้ายคลึงกับช่องไอออนที่ควบคุมด้วยแรงดันไฟฟ้าและส่วนของเยื่อหุ้มเซลล์ ที่อยู่ด้านข้าง (TMS) ช่องเหล่านี้อาจมีอยู่ในเยื่อหุ้มเซลล์ในรูปของโฮโมโอ ลิโกเมอร์ หรือเฮเทอโรโอลิโกเมอร์และแต่ละโมโนเมอร์มี TMS ระหว่าง 2 ถึง 4 ในแง่ของหน้าที่ โปรตีนเหล่านี้ขนส่งโพแทสเซียม (K + )โดยมีแนวโน้มที่จะดูดซับ K + มากกว่า ส่งออกK + [ 3 ]กระบวนการปรับทิศทางเข้าด้านในถูกค้นพบโดยBernard Katzในเซลล์กล้ามเนื้อโครงร่างในปี พ.ศ. 2492 [ 6 ]และDenis Nobleในเซลล์กล้ามเนื้อหัวใจในช่วงปี พ.ศ. 2503 [ 7 ]
ภาพรวมของการแก้ไขทิศทางเข้าด้านใน
{kind=link}
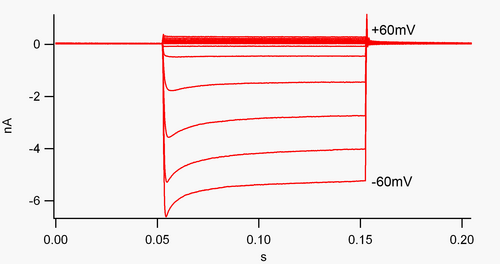
ช่องสัญญาณที่ "ปรับทิศทางการไหลเข้า" คือช่องสัญญาณที่ส่งผ่านกระแสไฟฟ้า (ประจุบวก) ได้ง่ายกว่าในทิศทางเข้าสู่เซลล์มากกว่าในทิศทางออกจากเซลล์ เชื่อกันว่ากระแสไฟฟ้านี้อาจมีบทบาทสำคัญในการควบคุมการทำงานของเซลล์ประสาท โดยช่วยรักษาเสถียรภาพของ ศักย์เยื่อหุ้ม เซลล์ ขณะพัก
ตามหลักการแล้ว กระแสไฟฟ้าไหลเข้า (ประจุบวกเคลื่อนที่เข้าสู่เซลล์) จะแสดงในกราฟการควบคุมแรงดันไฟฟ้าเป็นการเบี่ยงเบนลง ในขณะที่กระแสไฟฟ้าไหลออก (ประจุบวกเคลื่อนที่ออกจากเซลล์) จะแสดงเป็นการเบี่ยงเบนขึ้น ที่ศักย์เยื่อหุ้มเซลล์ที่เป็นลบเมื่อเทียบกับศักย์ผกผัน ของโพแทสเซียม ช่อง K +ที่ปรับกระแสไฟฟ้าเข้า จะช่วยให้ไอออน K +ที่มีประจุบวกไหลเข้าสู่เซลล์ ทำให้ศักย์เยื่อหุ้มเซลล์กลับไปสู่ศักย์พักตัว ดังแสดงในรูปที่ 1: เมื่อศักย์เยื่อหุ้มเซลล์ถูกควบคุมให้เป็นลบเมื่อเทียบกับศักย์พักตัวของช่อง (เช่น -60 mV) กระแสไฟฟ้าไหลเข้า (เช่น ประจุบวกไหลเข้าสู่เซลล์) จะไหลเข้า อย่างไรก็ตาม เมื่อศักย์เยื่อหุ้มเซลล์ถูกตั้งค่าให้เป็นบวกเมื่อเทียบกับศักย์พักตัวของช่อง (เช่น +60 mV) ช่องเหล่านี้จะปล่อยกระแสไฟฟ้าไหลผ่านน้อยมาก กล่าวคือ ช่องนี้ปล่อยกระแสไฟฟ้าไหลเข้ามากกว่าไหลออกในช่วงแรงดันไฟฟ้าที่ใช้งานได้ ช่องเหล่านี้ไม่ใช่ตัวปรับกระแสไฟฟ้าที่สมบูรณ์แบบ เนื่องจากสามารถปล่อยกระแสไฟฟ้าไหลออกได้บ้างในช่วงแรงดันไฟฟ้าสูงถึงประมาณ 30 mV เหนือศักย์พักตัว
ช่องเหล่านี้แตกต่างจากช่องโพแทสเซียมที่โดยทั่วไปมีหน้าที่ในการทำให้เซลล์กลับสู่สภาวะปกติหลังจากเกิดศักย์ไฟฟ้าเช่น ช่อง โพแทสเซียมแบบดีเลย์เรคติไฟเออร์และ ช่องโพแทสเซียม ชนิด Aช่องโพแทสเซียมที่ "ปกติ" เหล่านี้จะนำกระแสโพแทสเซียมออกไปด้านนอก (มากกว่าด้านใน) ที่ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์ลดลง และอาจถือได้ว่าเป็น "การปรับทิศทางออกไปด้านนอก" เมื่อค้นพบครั้งแรก การปรับทิศทางเข้าด้านในถูกตั้งชื่อว่า "การปรับทิศทางที่ผิดปกติ" เพื่อแยกความแตกต่างจากกระแสโพแทสเซียมที่ออกไปด้านนอก[ 8 ]
ตัวปรับกระแสเข้ายังแตกต่างจากช่องโพแทสเซียมโดเมนรูพรุนคู่ซึ่งส่วนใหญ่รับผิดชอบต่อกระแส K + ที่ "รั่วไหล" [ 9 ]ตัวปรับกระแสเข้าบางชนิดที่เรียกว่า "ตัวปรับกระแสเข้าแบบอ่อน" จะนำกระแส K + ที่วัดได้ออกไป ที่แรงดันไฟฟ้าที่เป็นบวกกับศักยภาพการกลับทิศทางของ K + (สอดคล้องกับ แต่มีขนาดใหญ่กว่า กระแสขนาดเล็กเหนือเส้น 0 nA ในรูปที่ 1) พวกมันพร้อมกับช่อง "รั่วไหล" จะสร้างศักยภาพเยื่อหุ้มเซลล์ขณะพัก ช่องปรับกระแสเข้าอื่นๆ ที่เรียกว่า "ตัวปรับกระแสเข้าแบบแรง" จะนำกระแสออกไปน้อยมาก และส่วนใหญ่จะทำงานที่แรงดันไฟฟ้าที่เป็นลบกับศักยภาพการกลับทิศทางของ K +ซึ่งพวกมันจะนำกระแสเข้า (กระแสที่ใหญ่กว่ามากด้านล่างเส้น 0 nA ในรูปที่ 1) [ 10 ]
กลไกการปรับทิศทางเข้าด้านใน
ปรากฏการณ์การแก้ไขกระแสเข้าของช่อง K เป็นผลมาจากการปิดกั้นที่มีความสัมพันธ์สูงโดยโพลีอะมีน ภายในร่างกาย ได้แก่สเปอร์มีนรวมถึง ไอออน แมกนีเซียมซึ่งอุดรูพรุนของช่องที่ศักย์บวก ส่งผลให้กระแสไหลออกลดลง การปิดกั้นที่ขึ้นอยู่กับแรงดันไฟฟ้าโดยโพลีอะมีนนี้ส่งผลให้การนำกระแสมีประสิทธิภาพเฉพาะในทิศทางเข้าเท่านั้น แม้ว่าแนวคิดหลักของการปิดกั้นโดยโพลีอะมีนจะเป็นที่เข้าใจ แต่กลไกเฉพาะยังคงเป็นที่ถกเถียงกันอยู่[ 11 ]
การเปิดใช้งานโดย PIP
ทั้งหมดต้องการฟอสฟาติดิลอิโนซิทอล 4,5-บิสฟอสเฟต (PIP ) เพื่อการกระตุ้น[ 12 ] PIP จับกับและกระตุ้น K 2.2 โดยตรงด้วยคุณสมบัติคล้ายตัวกระตุ้น[ 13 ]ในแง่นี้ ช่อง K เป็นช่องไอออนที่ควบคุมด้วยลิแกนด์ PIP
บทบาท
ช่อง K พบได้ในเซลล์หลายชนิด รวมถึงแมโครฟาจเซลล์หัวใจและไตเม็ดเลือดขาวเซลล์ประสาทและเซลล์บุผนังหลอดเลือด โดยการเป็นตัวกลางในการส่งกระแส K + ที่ทำให้เกิดการลดศักย์ ไฟฟ้า เล็กน้อยที่ศักย์เยื่อหุ้มเซลล์ที่เป็นลบ ช่องเหล่านี้ช่วยสร้างศักย์เยื่อหุ้มเซลล์ขณะพัก และในกรณีของ กลุ่ม K 3ช่องเหล่านี้ช่วยเป็นตัวกลางใน การตอบสนองของ สารสื่อประสาท ที่ยับยั้งการทำงาน แต่บทบาทของช่องเหล่านี้ในสรีรวิทยาของเซลล์จะแตกต่างกันไปตามชนิดของเซลล์:
| ที่ตั้ง | การทำงาน |
|---|---|
| เซลล์กล้ามเนื้อหัวใจ | ช่อง K ระดับศักย์ไฟฟ้าของหัวใจให้ยาวนานขึ้นช่องปรับทิศทางเข้าแบบนี้แตกต่างจากช่องK + แบบปรับทิศทางช้า ซึ่งช่วยคืนขั้วให้กับเซลล์ประสาทและกล้ามเนื้อหลังจากเกิดศักย์ไฟฟ้า และช่องโพแทสเซียมรั่วซึ่งเป็นพื้นฐานสำคัญของ ศักย์ ไฟฟ้า ของเยื่อหุ้มเซลล์ขณะพัก |
| เซลล์เยื่อบุผนังหลอดเลือด | ช่อง K มีส่วนเกี่ยวข้องในการควบคุมเอนไซม์ไนตริกออกไซด์ซินเทส |
| ไต | เอนไซม์ K ทำหน้าที่ส่งออกโพแทสเซียมส่วนเกินไปยังท่อรวมเพื่อขับออกทางปัสสาวะ หรืออาจมีส่วนเกี่ยวข้องกับการดูดซึมโพแทสเซียมกลับเข้าสู่ร่างกาย |
| เซลล์ประสาทและในเซลล์หัวใจ | IRK ที่กระตุ้นด้วยโปรตีน G (K 3)เป็นตัวควบคุมที่สำคัญซึ่งถูกปรับเปลี่ยนโดยสารสื่อประสาท การกลายพันธุ์ใน ช่อง GIRK2นำไปสู่การกลายพันธุ์ของหนูวีเวอร์ หนูที่กลายพันธุ์ "วีเวอร์" มีอาการเดินเซและแสดงอาการเสื่อมของเซลล์ประสาทโดปามีนที่เกิดจากการอักเสบของระบบประสาท[ 14 ]เมื่อเทียบกับหนูควบคุมที่ไม่มีอาการเดินเซ หนูที่กลายพันธุ์วีเวอร์มีความบกพร่องในการประสานงานของการเคลื่อนไหวและการเปลี่ยนแปลงในการเผาผลาญของสมองในระดับภูมิภาค[ 15 ]หนูวีเวอร์ได้รับการศึกษาในห้องปฏิบัติการที่สนใจในการพัฒนาและการเกิดโรคของระบบประสาทมานานกว่า 30 ปี |
| เซลล์เบต้าของตับอ่อน | ช่องK (ประกอบด้วย หน่วยย่อย K 6.2และSUR1 ) ควบคุมการหลั่งอินซูลิน |
ระเบียบข้อบังคับ
การพึ่งพาแรงดันไฟฟ้าอาจถูกควบคุมโดย K + ภายนอก, Mg2 +ภายใน , ATPภายในและ/หรือโปรตีน Gโดเมน P ของช่อง IRK มีความคล้ายคลึงกันในลำดับที่จำกัดกับโดเมนของตระกูล VIC ตัวปรับกระแสไฟฟ้าเข้ามีบทบาทในการกำหนดศักย์เยื่อหุ้มเซลล์ และการปิดช่องเหล่านี้เมื่อเกิดการลดศักย์ไฟฟ้าจะทำให้เกิดศักย์ไฟฟ้าแอคชั่นที่มีระยะเวลานานพร้อมระยะราบเรียบ ตัวปรับกระแสไฟฟ้าเข้าขาดเกลียวรับรู้แรงดันไฟฟ้าภายในที่พบในช่องตระกูล VIC หลายช่อง ในบางกรณี เช่น Kir1.1a, Kir6.1 และ Kir6.2 มีการเสนอว่าการโต้ตอบโดยตรงกับสมาชิกของซูเปอร์แฟมิลี ABC จะทำให้คอมเพล็กซ์เฮเทอโรเมอริกมีคุณสมบัติการทำงานและการควบคุมที่เป็นเอกลักษณ์ รวมถึงความไวต่อ ATP ช่องที่ไวต่อ ATP เหล่านี้พบได้ในเนื้อเยื่อต่างๆ ของร่างกาย พวกมันทำให้กิจกรรมของช่องตอบสนองต่ออัตราส่วน ATP/ADP ในไซโตพลาสซึม (ATP/ADP ที่เพิ่มขึ้นจะปิดช่อง) ตัวรับซัล โฟนิลยูเรีย SUR1 และ SUR2 ของมนุษย์(spQ09428 และ Q15527 ตามลำดับ) เป็นโปรตีน ABC ที่ควบคุมทั้งช่อง Kir6.1 และ Kir6.2 ในการตอบสนองต่อ ATP และ CFTR ( TC #3.A.1.208.4 ) อาจควบคุม Kir1.1a [ 16 ]
โครงสร้าง
โครงสร้างผลึก[ 17 ]และหน้าที่[ 18 ]ของสมาชิกแบคทีเรียในตระกูล IRK-C ได้รับการกำหนดแล้ว KirBac1.1 จากBurkholderia pseudomalleiมีความยาว 333 หน่วยอะมิโนเอซิล (aas) โดยมี TMS สองตัวที่ปลาย N ขนาบข้าง P-loop (หน่วยที่ 1-150) และครึ่งปลาย C ของโปรตีนเป็นแบบชอบน้ำ มันขนส่งแคตไอออนโมโนวาเลนต์ด้วยความเลือกสรร: K ≈ Rb ≈ Cs ≫ Li ≈ Na ≈ NMGM ( N -methyl- D - glucamine ที่ถูกโปรตอน ) กิจกรรมถูกยับยั้งโดย Ba 2+ , Ca 2+และ pH ต่ำ[ 18 ]
การจำแนกประเภท
มีเจ็ดกลุ่มย่อยโดยระบุเป็น K 1 – K 7 [ 1 ]แต่ละกลุ่มย่อยมีสมาชิกหลายตัว (เช่น K 2.1, K 2.2, K 2.3 เป็นต้น) ซึ่งมีลำดับกรดอะมิโนที่เกือบจะเหมือนกันในสัตว์เลี้ยงลูกด้วยนมที่รู้จัก
ช่อง K เกิดจากโปรตีนเมมเบรนแบบโฮโมเตตราเมอ ริก โดยแต่ละหน่วยย่อยของโปรตีนที่เหมือนกันทั้งสี่หน่วยประกอบด้วยอัลฟาเฮลิกซ์ ที่ทะลุผ่านเยื่อหุ้มเซลล์สองอัน (M1 และ M2) เฮเทโรเตตราเมอริกสามารถเกิดขึ้นได้ระหว่างสมาชิกในกลุ่มย่อยเดียวกัน (เช่น K 2.1 และ K 2.3) เมื่อมีการแสดงออกของช่องมากเกินไป
ความหลากหลาย
| ยีน | โปรตีน | ชื่อเรียกอื่น | หน่วยย่อยที่เกี่ยวข้อง |
|---|---|---|---|
| เคซีเอ็นเจ1 | K 1.1 | รอมเค1 | เอ็นเฮอร์ฟ2 |
| เคซีเอ็นเจ2 | K 2.1 | IRK1 | K 2.2, K 4.1, PSD-95 , SAP97 , AKAP79 |
| เคซีเอ็นเจ12 | K 2.2 | อิอาร์เค2 | K 2.1 และ K 2.3 เพื่อสร้างช่องสัญญาณเฮเทอโรเมอริก หน่วยย่อยเสริม: SAP97, Veli-1 , Veli-3 , PSD-95 |
| เคซีเอ็นเจ4 | K 2.3 | ไออาร์เค3 | K 2.1 และ K 2.3 เพื่อสร้างช่องเฮเทอโรเมอริก PSD-95, Chapsyn-110 /PSD-93 |
| เคซีเอ็นเจ14 | K 2.4 | IRK4 | K 2.1 เพื่อสร้างช่องเฮเทอโรเมอริก |
| เคซีเอ็นเจ3 | K 3.1 | GIRK1, KGA | K 3.2, K 3.4, K 3.5, K 3.1 ไม่สามารถใช้งานได้ด้วยตัวเอง |
| เคซีเอ็นเจ6 | K 3.2 | GIRK2 | K 3.1, K 3.3, K 3.4 เพื่อสร้างช่องเฮเทอโรเมอริก |
| เคซีเอ็นเจ9 | K 3.3 | GIRK3 | K 3.1, K 3.2 เพื่อสร้างช่องเฮเทอโรเมอริก |
| เคซีเอ็นเจ5 | K 3.4 | GIRK4 | K 3.1, K 3.2, K 3.3 |
| เคซีเอ็นเจ10 | K 4.1 | K 1.2 | K 4.2, K 5.1 และ K 2.1 เพื่อสร้างช่องเฮเทอโรเมอริก |
| เคซีเอ็นเจ15 | K 4.2 | K 1.3 | |
| เคซีเอ็นเจ16 | K 5.1 | บีอาร์ไอ 9 | |
| เคซีเอ็นเจ8 | K 6.1 | เคเอ | เซอร์ทูบี |
| เคซีเอ็นเจ11 | K 6.2 | เคเอ | SUR1 , SUR2AและSUR2B |
| เคซีเอ็นเจ13 | K 7.1 | K 1.4 |
โรคที่เกี่ยวข้องกับ ช่องK
- ภาวะน้ำตาลในเลือดต่ำจากอินซูลินสูงเรื้อรังในวัยทารกมีความเกี่ยวข้องกับการกลายพันธุ์แบบยีนด้อยในยีน K 6.2 การกลายพันธุ์บางอย่างของยีนนี้ทำให้ความสามารถของช่องสัญญาณในการควบคุมการหลั่งอินซูลินลดลง ส่งผลให้เกิดภาวะน้ำตาลในเลือดต่ำ
- กลุ่มอาการบาร์ตเตอร์อาจเกิดจากการกลายพันธุ์ในช่อง Kภาวะนี้มีลักษณะเฉพาะคือไตไม่สามารถนำโพแทสเซียมกลับมาใช้ใหม่ได้ ทำให้ระดับโพแทสเซียมในร่างกาย
- กลุ่มอาการแอนเดอร์เซนเป็นภาวะหายากที่เกิดจากการกลายพันธุ์หลายตำแหน่งของยีน K 2.1 โดยอาจเป็นลักษณะเด่นหรือลักษณะด้อยขึ้นอยู่กับการกลายพันธุ์นั้น ลักษณะเด่นคืออัมพาตเป็นระยะหัวใจเต้นผิดและ ใบหน้า ผิดรูป (ดูเพิ่มเติมที่ KCNJ2 )
- ภาวะเป็นพิษจากแบเรียมมีแนวโน้มที่จะเกิดจากความสามารถของแบเรียมในการปิดกั้นช่อง
- ภาวะหลอดเลือดแดงแข็ง (โรคหัวใจ)อาจเกี่ยวข้องกับ K+ การสูญเสียกระแสไอออน Kในเซลล์เยื่อบุหลอดเลือดเป็นหนึ่งในตัวบ่งชี้แรกๆ ที่ทราบกันดีของการเกิดภาวะหลอดเลือดแดงแข็ง (จุดเริ่มต้นของโรคหัวใจ)
- ภาวะอัมพาตเป็นระยะจากภาวะไทรอยด์เป็นพิษและโพแทสเซียมต่ำมีความเชื่อมโยงกับการทำงานของ K 2.6 ที่เปลี่ยนแปลงไป [ 19 ]
- กลุ่มอาการ EAST/SeSAMEเกิดจากการกลายพันธุ์ใน KCNJ10 [ 20 ]
ดูเพิ่มเติม
เอกสารอ้างอิง
- ^ a b Kubo Y, Adelman JP, Clapham DE, Jan LY, Karschin A, Kurachi Y และคณะ (ธันวาคม 2548). "สหภาพเภสัชวิทยาระหว่างประเทศ LIV. การตั้งชื่อและความสัมพันธ์ระดับโมเลกุลของช่องโพแทสเซียมที่ปรับทิศทางเข้าด้านใน". บท วิจารณ์ทางเภสัชวิทยา57 (4): 509– 26. doi : 10.1124/pr.57.4.11 . PMID 16382105 . S2CID 11588492 .
- ^ Hedrich R, Moran O, Conti F, Busch H, Becker D, Gambale F และคณะ (1995). "ช่องโพแทสเซียมแบบปรับทิศทางเข้าด้านในในพืชแตกต่างจากช่องโพแทสเซียมในสัตว์เมื่อตอบสนองต่อแรงดันไฟฟ้าและตัวปรับช่อง" European Biophysics Journal . 24 (2): 107– 15. doi : 10.1007/BF00211406 . PMID 8582318 . S2CID 12718513 .
- ^ a b "1.A.2 ตระกูลช่อง K แบบปรับแก้ทิศทางเข้าด้านใน (IRK-C)" . TCDB . สืบค้นเมื่อ2016-04-09 .
- ^ Hansen SB (พฤษภาคม 2015). "การกระตุ้นด้วยลิปิด: รูปแบบ PIP2 ของช่องไอออนที่ควบคุมด้วยลิแกนด์" . Biochimica et Biophysica Acta (BBA) - ชีววิทยาโมเลกุลและเซลล์ของลิปิด . 1851 (5): 620– 8. doi : 10.1016/j.bbalip.2015.01.011 . PMC 4540326 . PMID 25633344 .
- ^ Abraham MR, Jahangir A, Alekseev AE, Terzic A (พฤศจิกายน 1999). "Channelopathies of inwardly rectifying potassium channels" . FASEB Journal . 13 (14): 1901– 10. doi : 10.1096/fasebj.13.14.1901 . PMID 10544173 . S2CID 22205168 .
- ↑แคตซ์, เบอร์นาร์ด (1949) "เลส์ คอนสแตนเตส อิเล็กทริก เดอ ลา เมมเบรน ดู กล้ามเนื้อ" โค้ง. วิทยาศาสตร์ ฟิสิออล . 3 : 285– 299.
- ^ Noble, Denis (ธันวาคม 1965). "คุณสมบัติทางไฟฟ้าของกล้ามเนื้อหัวใจที่เกิดจากการแก้ไขกระแสไฟฟ้าเข้าด้านใน (ผิดปกติ)"วารสารสรีรวิทยาของเซลล์และเปรียบเทียบ66 (S2): 127–135 . doi : 10.1002/jcp.1030660520 . ISSN 0095-9898 .
- ^ Bertil Hille (2001).ช่องไอออนของเยื่อหุ้มเซลล์ที่ไวต่อการกระตุ้นฉบับที่ 3 (Sinauer: Sunderland, MA), หน้า 151. ISBN 0-87893-321-2.
- ^ฮิลเล, หน้า 155.
- ^ฮิลเล, หน้า 153.
- ^ Lopatin AN, Makhina EN, Nichols CG (พฤศจิกายน 1995). "กลไกการปรับทิศทางเข้าของช่องโพแทสเซียม: "การอุดรูพรุนยาว" โดยโพลีอะมีนในไซโตพลาสซึม"วารสารสรีรวิทยาทั่วไป 106 ( 5): 923– 55. doi : 10.1085/jgp.106.5.923 . PMC 2229292 . PMID 8648298 .
- ^ Tucker SJ, Baukrowitz T (พฤษภาคม 2551). "ลิปิดแอนไอออนิกที่มีประจุสูงจับและควบคุมช่องไอออนได้อย่างไร" วารสารสรีรวิทยาทั่วไป 131 ( 5 ) : 431– 8. doi : 10.1085/jgp.200709936 . PMC 2346576 . PMID 18411329 .
- ^ Hansen SB, Tao X, MacKinnon R (สิงหาคม 2011). "โครงสร้างพื้นฐานของการกระตุ้น PIP2 ของช่อง K+ แบบปรับทิศทางเข้าแบบคลาสสิก Kir2.2" Nature . 477 ( 7365 ): 495– 8. Bibcode : 2011Natur.477..495H . doi : 10.1038/nature10370 . PMC 3324908 . PMID 21874019 .
- ^ Peng J, Xie L, Stevenson FF, Melov S, Di Monte DA, Andersen JK (พฤศจิกายน 2549). "การเสื่อมของเซลล์ประสาทโดปามีนในบริเวณนิกโกรสไตรเอตัมในหนูวีเวอร์เกิดจากการอักเสบของระบบประสาทและบรรเทาลงได้ด้วยการให้ยาไมโนไซคลิน"วารสารประสาทวิทยาศาสตร์ 26 ( 45): 11644– 51. doi : 10.1523/JNEUROSCI.3447-06.2006 . PMC 6674792 . PMID 17093086 .
- ^ Strazielle C, Deiss V, Naudon L, Raisman-Vozari R, Lalonde R (ตุลาคม 2549). "ความแปรผันของกิจกรรมไซโตโครมออกซิเดสและการประสานงานของมอเตอร์ในสมองส่วนภูมิภาคในหนู Girk2(Wv) (Weaver) กลายพันธุ์" Neuroscience . 142 (2): 437– 49. doi : 10.1016/j.neuroscience.2006.06.011 . PMID 16844307 . S2CID 33064439 .
- ^ คำขอ WO หมายเลข 0190360 , Wei MH, Chaturvedi K, Guegler K, Webster M, Ketchum KA, Di Francesco V, Beasley E, "โปรตีนขนส่งของมนุษย์ที่แยกได้ โมเลกุลกรดนิวคลีอิกที่เข้ารหัสโปรตีนขนส่งของมนุษย์ และการใช้งาน", เผยแพร่เมื่อวันที่ 29 พฤศจิกายน 2001, มอบสิทธิ์ให้แก่ Apperla Corporation
- ^ Kuo A, Gulbis JM, Antcliff JF, Rahman T, Lowe ED, Zimmer J และคณะ (มิถุนายน 2546). "โครงสร้างผลึกของช่องโพแทสเซียม KirBac1.1 ในสถานะปิด" . Science . 300 (5627): 1922– 6. Bibcode : 2003Sci...300.1922K . doi : 10.1126/science.1085028 . PMID 12738871 . S2CID 2703162 .
- ^ a b Enkvetchakul D, Bhattacharyya J, Jeliazkova I, Groesbeck DK, Cukras CA, Nichols CG (พฤศจิกายน 2547). "การกำหนดลักษณะการทำงานของช่อง Kir ของโปรคาริโอต"วารสารเคมีชีวภาพ 279 ( 45): 47076– 80. doi : 10.1074/jbc.C400417200 . PMC 8629170 . PMID 15448150 .
- ^ Ryan DP, da Silva MR, Soong TW, Fontaine B, Donaldson MR, Kung AW และคณะ (มกราคม 2010). "การกลายพันธุ์ในช่องโพแทสเซียม Kir2.6 ทำให้เกิดความเสี่ยงต่อภาวะอัมพาตเป็นระยะจากภาวะโพแทสเซียมต่ำในต่อมไทรอยด์เป็นพิษ" . Cell . 140 (1): 88– 98. doi : 10.1016/j.cell.2009.12.024 . PMC 2885139 . PMID 20074522 .
- ^ Bockenhauer D, Feather S, Stanescu HC, Bandulik S, Zdebik AA, Reichold M และคณะ (พฤษภาคม 2552). "โรคลมชัก, ภาวะเสียการทรงตัว, หูหนวกจากความเสียหายของเส้นประสาทรับเสียง, โรคท่อน้ำไขสันหลัง และการกลายพันธุ์ของยีน KCNJ10"วารสารการแพทย์นิวอิงแลนด์ 360 ( 19): 1960– 70. doi : 10.1056/NEJMoa0810276 . PMC 3398803 . PMID 19420365 .
อ่านเพิ่มเติม
- Hille B (2001). ช่องไอออนของเยื่อหุ้มเซลล์ที่ไวต่อการกระตุ้น (ฉบับที่ 3). ซันเดอร์แลนด์, แมสซาชูเซตส์: Sinauer. หน้า 149–154 . ISBN 0-87893-321-2.
ลิงก์ภายนอก
- Inward+Rectifier+Potassium+Channelsที่ US National Library of Medicine Medical Subject Headings (MeSH)
- "ช่องโพแทสเซียมที่รับไอออนเข้าภายใน"ฐานข้อมูลตัวรับและช่องไอออนของ IUPHARสหภาพระหว่างประเทศด้านเภสัชวิทยาพื้นฐานและคลินิก
- UMich Orientation of Proteins in Membranes families/family-85 - ตำแหน่งเชิงพื้นที่ของช่องโพแทสเซียมแบบปรับทิศทางเข้าในเยื่อหุ้มเซลล์
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ช่องโพแทสเซียมแบบปรับทิศทางเข้า
ช่องโพแทสเซียมแบบปรับทิศทางเข้า ( Kir , IRK ) เป็นกลุ่มย่อยของช่องโพแทสเซียมที่ควบคุมด้วยลิปิดโดย เฉพาะ จนถึงปัจจุบัน...
ภาพรวมของการแก้ไขทิศทางเข้าด้านใน
รูปที่ 1. บันทึกกระแสไฟฟ้าทั้งเซลล์ของช่องโพแทสเซียม K 2 ที่ปรับทิศทางเข้าด้านในซึ่งแสดงออกในเซลล์ HEK293 (นี่คือกระแสไฟฟ้าที่ปรับทิศทางเข้าด้านในอย่างชัดเจน การเบี่ยงเบนลงด้านล่างคือกระแสไฟฟ้าเข้า การเบี่ยงเบนขึ้นด้านบนคือกระแสไฟฟ้าออก และแกน x...
กลไกการปรับทิศทางเข้าด้านใน
ปรากฏการณ์การแก้ไขกระแสเข้าของช่อง K เป็นผลมาจากการปิดกั้นที่มีความสัมพันธ์สูงโดยโพลีอะมีน ภายในร่างกาย ได้แก่สเปอร์มีนรวมถึง ไอออน แมกนีเซียมซึ่งอุดรูพรุนของช่องที่ศักย์บวก ส่งผลให้กระแสไหลออกลดลง...
การเปิดใช้งานโดย PIP
ทั้งหมดต้องการฟอสฟาติดิลอิโนซิทอล 4,5-บิสฟอสเฟต (PIP ) เพื่อการกระตุ้น[ 12 ] PIP จับกับและกระตุ้น K 2.2 โดยตรงด้วยคุณสมบัติคล้ายตัวกระตุ้น[ 13 ]ในแง่นี้ ช่อง K เป็นช่องไอออนที่ควบคุมด้วยลิแกนด์ PIP